双特异性T细胞活化性抗原结合分子的制作方法
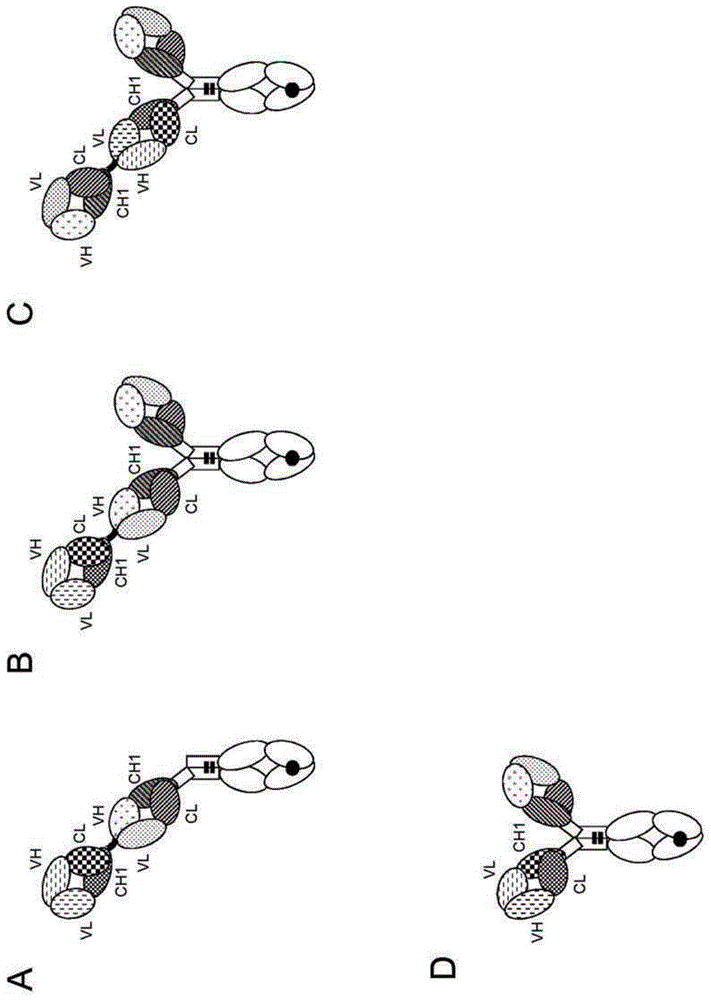
发明领域本发明一般涉及用于活化t细胞的双特异性抗原结合分子。另外,本发明涉及编码此类双特异性抗原结合分子的多核苷酸,以及包含此类多核苷酸的载体和宿主细胞。本发明进一步涉及用于生成本发明双特异性抗原结合分子的方法,以及在疾病治疗中使用这些双特异性抗原结合分子的方法。发明背景在多种临床背景中常常期望选择性破坏个别细胞或特定细胞类型。例如,特异性破坏肿瘤细胞而使健康细胞和组织保持完整且不受损是癌症疗法的首要目的。实现这点的一种有吸引力的方式是通过诱导针对肿瘤的免疫应答,使得免疫效应细胞诸如天然杀伤(nk)细胞或细胞毒性t淋巴细胞(ctl)攻击并破坏肿瘤细胞。ctl构成免疫系统最有力的效应细胞,然而它们不能通过由常规治疗性抗体的fc域介导的效应器机制来激活。在此点上,最近数年对双特异性抗体变得感兴趣,其设计为用一个“臂”结合靶细胞上的表面抗原,而用第二个“臂”结合t细胞受体(tcr)复合物的活化性的、不变的组分。此类抗体对其两种靶物的同时结合会迫使靶细胞和t细胞之间的暂时相互作用,引起任何细胞毒性t细胞活化和随后的靶细胞裂解。因此,免疫应答重定向于靶细胞,而且不依赖于靶细胞的肽抗原呈递或t细胞的特异性,其对于ctl的正常mhc限制性活化会是相关的。在此背景中,至关重要的是ctl仅在靶细胞向其呈递双特异性抗体时活化,即模拟免疫突触。特别期望的是不需要淋巴细胞预条件化或共刺激来引发靶细胞有效裂解的双特异性抗体。已开发出数种双特异性抗体形式并研究了它们对调查中的t细胞介导的免疫疗法的适宜性。其中,所谓的bite(双特异性t细胞衔接物(engager))分子已得到非常好的表征,而且在临床中已显示出一些前景(综述见nagorsen和expcellres317,1255-1260(2011))。bite是串联scfv分子,其中两个scfv分子通过柔性接头融合。针对t细胞衔接评估的其它双特异性形式包括双抗体(holliger等,proteng9,299-305(1996))及其衍生物,诸如串联双抗体(kipriyanov等,jmolbiol293,41-66(1999))。一项最近的进展是所谓的dart(双重亲和力重靶向)分子,它们基于双抗体形式但特征在于实现额外稳定化的c端二硫桥(moore等,blood117,4542-51(2011))。所谓的triomab(它们是完整杂合小鼠/大鼠igg分子,而且目前亦在临床试验中进行评估)代表了尺寸更大的形式(综述于seimetz等,cancertreatrev36,458-467(2010))。正在开发的多种形式显示免疫疗法中归因于t细胞重定向和活化的极大潜力。然而,生成对此合适的双特异性抗体的任务绝不是微不足道的,而是牵涉到许多必须满足的与抗体功效、毒性、适用性和生产能力有关的挑战。小构建体诸如例如bite分子(尽管能够有效交联效应器和靶细胞)具有非常短的血清半衰期,从而需要通过连续输注对患者施用它们。另一方面,igg样形式(尽管具有长半衰期的极大益处)受制于与igg分子固有的天然效应器功能有关的毒性。它们的免疫原性潜力构成了成功治疗性开发的igg样双特异性抗体(尤其是非人形式)的另一个不利特征。最后,双特异性抗体的一般开发中的一项主要挑战是以临床充足的数量和纯度生产双特异性抗体构建体,原因自傲与具有不同特异性的抗体重和轻链在共表达后的错配,这降低了正确装配的构建体的产率且导致许多无功能的副产物,而期望的双特异性抗体可能难以与之分开。鉴于与目前可获得用于t细胞介导的免疫疗法的双特异性抗体有关的难点和缺点,仍然存在对此类分子的新的改进形式的需要。本发明提供设计用于t细胞活化和重定向的双特异性抗原结合分子,其组合了优良的功效和生产能力与较低的毒性和有利的药动学特性。发明概述在第一个方面,本发明提供一种t细胞活化性双特异性抗原结合分子,其包含(i)第一抗原结合模块,它是能够特异性结合cd3的fab分子,包含至少一个选自seqidno:4,seqidno:5和seqidno:6的重链互补决定区(cdr)和至少一个选自seqidno:8,seqidno:9和seqidno:10的轻链cdr;(ii)第二抗原结合模块,它是能够特异性结合靶细胞抗原的fab分子。在一个实施方案中,所述第一抗原结合模块(它是能够特异性结合cd3的fab分子)包含包含与选自seqidno:3,seqidno:32和seqidno:33的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与选自seqidno:7和seqidno:31的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个实施方案中,所述第一抗原结合模块(它是能够特异性结合cd3的fab分子)包含包含与seqidno:3的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:7的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个特定的实施方案中,所述第二抗原结合模块能够特异性结合癌胚抗原(cea,ceacam5)且包含至少一个选自seqidno:24,seqidno:25和seqidno:26的重链互补决定区(cdr)和至少一个选自seqidno:28,seqidno:29和seqidno:30的轻链cdr。在另一个特定的实施方案中,所述第二抗原结合模块能够特异性结合cea且包含包含与seqidno:23的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:27的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在另一个特定的实施方案中,所述第二抗原结合模块能够特异性结合mcsp(cspg4)且包含至少一个选自seqidno:14,seqidno:15,seqidno:16,seqidno:35,seqidno:37,seqidno:38和seqidno:40的重链互补决定区(cdr)和至少一个选自seqidno:18,seqidno:19,seqidno:20,seqidno:44,seqidno:45,seqidno:48,seqidno:49和seqidno:50的轻链cdr。在另一个特定的实施方案中,所述第二抗原结合模块能够特异性结合黑素瘤相关硫酸软骨素蛋白聚糖(mcsp,cspg4)且包含至少一个选自seqidno:14,seqidno:15和seqidno:16的重链互补决定区(cdr)和至少一个选自seqidno:18,seqidno:19和seqidno:20的轻链cdr。在另一个特定的实施方案中,所述第二抗原结合模块能够特异性结合mcsp且包含包含与选自seqidno:13,seqidno:34,seqidno:36,seqidno:39和seqidno:41的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与选自seqidno:17,seqidno:43,seqidno:46,seqidno:47和seqidno:51的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在另一个特定的实施方案中,所述第二抗原结合模块能够特异性结合mcsp且包含包含与seqidno:13的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:17的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个具体的实施方案中,所述第一抗原结合模块是交换fab分子,其中fab轻链和fab重链的可变或恒定区任一是交换的。在一个甚至更加具体的实施方案中,所述第一抗原结合模块是交换fab分子,其中fab轻链和fab重链的恒定区是交换的。在一个实施方案中,所述第二抗原结合模块是常规fab分子。在又一个具体的实施方案中,所述t细胞活化性双特异性抗原结合分子中存在不超过一个能够特异性结合cd3的抗原结合模块(即所述t细胞活化性双特异性抗原结合分子提供对cd3的单价结合)。在又一个实施方案中,所述t细胞活化性双特异性抗原结合分子进一步包含第三抗原结合模块,它是能够特异性结合靶细胞抗原的fab分子。在一个实施方案中,所述第三抗原结合模块是常规fab分子。在一个实施方案中,所述第三抗原结合模块与所述第二抗原结合模块相同。在一个具体的实施方案中,所述t细胞活化性双特异性抗原结合分子进一步包含第三抗原结合模块,它是能够特异性结合cea的fab分子,且包含至少一个选自seqidno:24,seqidno:25和seqidno:26的重链互补决定区(cdr)和至少一个选自seqidno:28,seqidno:29和seqidno:30的轻链cdr。在一个具体的实施方案中,所述t细胞活化性双特异性抗原结合分子进一步包含第三抗原结合模块,它是能够特异性结合cea的fab分子,且包含包含与seqidno:23的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:27的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个实施方案中,所述t细胞活化性双特异性抗原结合分子进一步包含第三抗原结合模块,它是能够特异性结合mcsp的fab分子,且包含至少一个选自seqidno:14,seqidno:15,seqidno:16,seqidno:35,seqidno:37,seqidno:38和seqidno:40的重链互补决定区(cdr)和至少一个选自seqidno:18,seqidno:19,seqidno:20,seqidno:44,seqidno:45,seqidno:48,seqidno:49和seqidno:50的轻链cdr。在一个具体的实施方案中,所述t细胞活化性双特异性抗原结合分子进一步包含第三抗原结合模块,它是能够特异性结合mcsp的fab分子,且包含至少一个选自seqidno:14,seqidno:15和seqidno:16的重链互补决定区(cdr)和至少一个选自seqidno:18,seqidno:19和seqidno:20的轻链cdr。在一个实施方案中,所述t细胞活化性双特异性抗原结合分子进一步包含第三抗原结合模块,它是能够特异性结合mcsp的fab分子,且包含包含与选自seqidno:13,seqidno:34,seqidno:36,seqidno:39和seqidno:41的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与选自seqidno:17,seqidno:43,seqidno:46,seqidno:47和seqidno:51的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个具体的实施方案中,所述t细胞活化性双特异性抗原结合分子进一步包含第三抗原结合模块,它是能够特异性结合mcsp的fab分子,且包含包含与seqidno:13的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:17的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一些实施方案中,所述t细胞活化性双特异性抗原结合分子的第一和第二抗原结合模块彼此融合,任选经由肽接头。在一个此类实施方案中,所述第二抗原结合模块在fab重链的c端融合至所述第一抗原结合模块的fab重链的n端。在另一个此类实施方案中,所述第一抗原结合模块在fab重链的c端融合至所述第二抗原结合模块的fab重链的n端。在其中(i)所述第二抗原结合模块在fab重链的c端融合至所述第一抗原结合模块的fab重链的n端或者(ii)所述第一抗原结合模块在fab重链的c端融合至所述第二抗原结合模块的fab重链的n端的实施方案中,所述第一抗原结合模块的fab轻链和所述第二抗原结合模块的fab轻链另外可以彼此融合,任选经由肽接头。在一个实施方案中,所述t细胞活化性双特异性抗原结合分子进一步包含(iii)由能够稳定联合的第一和第二亚基构成的fc域。在一个实施方案中,所述t细胞活化性双特异性抗原结合分子的第二抗原结合模块在fab重链的c端融合至所述fc域的第一或第二亚基的n端。在另一个实施方案中,所述第一抗原结合模块在fab重链的c端融合至所述fc域的第一或第二亚基的n端。在一个实施方案中,所述t细胞活化性双特异性抗原结合分子的第一和第二抗原结合模块各自在fab重链的c端融合至所述fc域的亚基之一的n端。在一个实施方案中,所述第三抗原结合模块在fab重链的c端融合至所述fc域的第一或第二亚基的n端。在一个具体的实施方案中,所述t细胞活化性抗原结合分子的第二和第三抗原结合模块各自在fab重链的c端融合至所述fc域的亚基之一的n端,且所述第一抗原结合模块在fab重链的c端融合至所述第二抗原结合模块的fab重链的n端。在另一个具体的实施方案中,所述t细胞活化性抗原结合分子的第一和第三抗原结合模块各自在fab重链的c端融合至所述fc域的亚基之一的n端,且所述第二抗原结合模块在fab重链的c端融合至所述第一抗原结合模块的fab重链的n端。所述t细胞活化性双特异性抗原结合分子的各构件可以直接或经由合适的肽接头融合。在一个实施方案中,所述第一和第三抗原结合模块和fc域是免疫球蛋白分子的部分。在一个具体的实施方案中,所述免疫球蛋白分子是igg类免疫球蛋白。在一个甚至更加具体的实施方案中,所述免疫球蛋白是igg1亚类免疫球蛋白。在另一个实施方案中,所述免疫球蛋白是igg4亚类免疫球蛋白。在一个具体的实施方案中,所述fc域是iggfc域。在一个特定的实施方案中,所述fc域是igg1fc域。在另一个特定的实施方案中,所述fc域是igg4fc域。在一个甚至更加特定的实施方案中,所述fc域是包含氨基酸替代s228p(依照kabat的eu编号方式)的igg4fc域。在具体的实施方案中,所述fc域是人fc域。在具体的实施方案中,所述fc域包含促进第一和第二fc域亚基联合的修饰。在一个特定的此类实施方案中,所述fc域的第一亚基的ch3域中的一个氨基酸残基用具有更大侧链体积的氨基酸残基替换,由此在所述第一亚基的ch3域内生成隆起,其可安置于所述第二亚基的ch3域内的空腔中,而且所述fc域的第二亚基的ch3域中的一个氨基酸残基用具有更小侧链体积的氨基酸残基替换,由此在所述第二亚基的ch3域内生成空腔,其内可安置所述第一亚基的ch3域内的隆起。在一个具体的实施方案中,相比于天然igg1fc域,所述fc域展现出降低的对fc受体的结合亲和力和/或降低的效应器功能。在某些实施方案中,相比于非工程化fc域,所述fc域被工程化改造为具有降低的对fc受体的结合亲和力和/或降低的效应器功能。在一个实施方案中,所述fc域包含一处或多处降低对fc受体的结合和/或效应器功能的氨基酸替代。在一个实施方案中,所述fc域中的一处或多处降低对fc受体的结合和/或效应器功能的氨基酸替代在选自下组的一个或多个位置处:l234,l235和p329(依照kabat的eu编号方式)。在具体的实施方案中,所述fc域的每个亚基包含三处降低对fc受体的结合和/或效应器功能的氨基酸替代,其中所述氨基酸替代是l234a,l235a和p329g。在一个此类实施方案中,所述fc域是igg1fc域,具体是人igg1fc域。在其它实施方案中,所述fc域的每个亚基包含两处降低对fc受体的结合和/或效应器功能的氨基酸替代,其中所述氨基酸替代是l235e和p329g。在一个此类实施方案中,所述fc域是igg4fc域,具体是人igg4fc域。在一个实施方案中,所述t细胞活化性双特异性抗原结合分子的fc域是igg4fc域且包含氨基酸替代l235e和s228p(sple)。在一个实施方案中,所述fc受体是fcγ受体。在一个实施方案中,所述fc受体是人fc受体。在一个实施方案中,所述fc受体是活化性fc受体。在一个特定的实施方案中,所述fc受体是人fcγriia,fcγri和/或fcγriiia。在一个实施方案中,所述效应器功能是抗体依赖性细胞介导的细胞毒性(adcc)。依照本发明的另一个方面,提供一种分离的多核苷酸,其编码本发明的t细胞活化性双特异性抗原结合分子或其片段。本发明还涵盖由本发明多核苷酸编码的多肽。本发明还提供包含本发明的分离的多核苷酸的表达载体,以及包含本发明的分离的多核苷酸或表达载体的宿主细胞。在一些实施方案中,所述宿主细胞是真核细胞,具体是哺乳动物细胞。在另一个方面,提供一种生成本发明的t细胞活化性双特异性抗原结合分子的方法,其包括下述步骤:a)在适合于表达所述t细胞活化性双特异性抗原结合分子的条件下培养本发明的宿主细胞,并b)回收所述t细胞活化性双特异性抗原结合分子。本发明还涵盖通过本发明的方法生成的t细胞活化性双特异性抗原结合分子。本发明进一步提供一种药物组合物,其包含本发明的t细胞活化性双特异性抗原结合分子和药学可接受载剂。本发明还涵盖使用本发明的t细胞活化性双特异性抗原结合分子和药物组合物的方法。在一个方面,本发明提供本发明的t细胞活化性双特异性抗原结合分子或药物组合物,其用作药物。在一个方面,提供依照本发明的t细胞活化性双特异性抗原结合分子或药物组合物,其用于治疗有所需要的个体中的疾病。在一个特定的实施方案中,所述疾病是癌症。还提供本发明的t细胞活化性双特异性抗原结合分子在制备用于治疗有所需要的个体中的疾病的药物中的用途;以及一种治疗个体中的疾病的方法,其包括对所述个体施用治疗有效量的组合物,所述组合物包含药学可接受形式的依照本发明的t细胞活化性双特异性抗原结合分子。在一个特定的实施方案中,所述疾病是癌症。在任一个上文实施方案中,所述个体优选是哺乳动物,具体是人。本发明还提供用于诱导靶细胞,具体是肿瘤细胞裂解的方法,其包括在t细胞(具体是细胞毒性t细胞)存在下使靶细胞与本发明的t细胞活化性双特异性抗原结合分子接触。附图简述图1。本发明的t细胞活化性双特异性抗原结合分子(tcb)的例示性构造。(a)“1+1iggcrossfab”分子的示图。(b)“2+1iggcrossfab”分子的示图。(c)具有备选次序的crossfab和fab构件(“倒转的”)的“2+1iggcrossfab”分子的示图。(d)“1+1crossmab”分子的示图。(e)“2+1iggcrossfab,连接的轻链”分子的示图。(f)“1+1iggcrossfab,连接的轻链”分子的示图。(g)“2+1iggcrossfab,倒转的,连接的轻链”分子的示图。(h)“1+1iggcrossfab,倒转的,连接的轻链”分子的示图。黑点:fc域中促进异二聚化的任选修饰。图2。亲和力成熟的抗mcsp克隆与非成熟的亲本克隆(m4-3ml2)相比的比对。图3。mcsptcb(2+1crossfab-iggp329glala倒转的)分子的示意图。图4。mcsptcb(2+1crossfab-iggp329glala倒转的,seqidno:12,53,54和55)的ce-sds分析。显示的电泳图是mcsptcb的sds-page:a)非还原性,b)还原性。图5。ceatcb(2+1crossfab-iggp329glala倒转的)分子的示意图。图6。ceatcb(2+1crossfab-iggp329glala倒转的,seqidno:22,56,57和58)分子的ce-sds分析。显示的电泳图是ceatcb的sds-page:a)非还原性,b)还原性。图7。mcsptcb(seqidno:12,53,54和55)对a375细胞(mcsp+)(a)和jurkat细胞(cd3+)(b)的结合。“非靶向tcb”:针对cd3但无第二抗原的双特异性抗体(seqidno:59,60,61和62)。图8。由mcsptcb抗体(seqidno:12,53,54和55)诱导的对a375(高mcsp)(a),mv-3(中等mcsp)(b),hct-116(低mcsp)(c)和ls180(mcsp阴性)(d)靶细胞的t细胞杀伤(e:t=10:1,效应人pbmc,温育时间24h)。“非靶向tcb”:针对cd3但无第二抗原的双特异性抗体(seqidno:59,60,61和62)。图9。由mcsptcb抗体(seqidno:12,53,54和55)诱导的t细胞介导的mv3黑素瘤细胞杀伤(e:t=10:1,24h温育)之后,人cd8+(a,b)和cd4+(c,d)t细胞上cd25和cd69的上调。“非靶向tcb”:针对cd3但无第二抗原的双特异性抗体(seqidno:59,60,61和62)。图10。由mcsptcb抗体(seqidno:12,53,54和55)诱导的t细胞介导的mv3黑素瘤细胞杀伤(e:t=10:1,24h温育)之后,人pbmc的il-2(a),ifn-γ(b),tnfα(c),il-4(d),il-10(e)和粒酶b(f)分泌。“非靶向tcb”:针对cd3但无第二抗原的双特异性抗体(seqidno:59,60,61和62)。图11。ceatcb(seqidno:22,56,57和58)对表达cea的a549肺腺癌细胞(a)和表达cd3的永生化人和猕猴t淋巴细胞系(分别为jurkat(b)和hsc-f(c))的结合。图12。由ceatcb(seqidno:22,56,57和58)诱导的对hpafii(高cea)(a,e),bxpc-3(中等cea)(b,f),aspc-1(低cea)(c,g)和hct-116(cea阴性)(d,h)细胞的t细胞杀伤。e:t=10:1,效应人pbmc,温育时间24h(a-d)或48h(e-h)。“非靶向tcb”:针对cd3但无第二抗原的双特异性抗体(seqidno:59,60,61和62)。图13。由ceatcb(seqidno:22,56,57和58)诱导的t细胞介导的hpafii(高cea)(a,e),bxpc-3(中等cea)(b,f),aspc-1(低cea)(c,g)和hct-116(cea阴性)(d,h)细胞杀伤之后5天,人cd8+和cd4+t细胞增殖(a-d)和人cd8+和cd4t细胞上的cd25上调(e-h)。“dp47tcb”:针对cd3但无第二抗原的双特异性抗体(seqidno:59,60,61和62)。图14。由ceatcb(seqidno:22,56,57和58)诱导的t细胞介导的mkn45肿瘤细胞杀伤(e:t=10:1,48h温育)之后的ifn-γ(a),tnfα(b),粒酶b(c),il-2(d),il-6(e)和il-10(f)分泌。“非靶向tcb”:针对cd3但无第二抗原的双特异性抗体(seqidno:59,60,61和62)。图15。在渐增浓度的脱落cea(scea)存在下,由ceatcb(seqidno:22,56,57和58)诱导的t细胞介导的对表达cea的ls180肿瘤靶细胞的杀伤,与ceatcb和scea一起温育后24h(a)或48h(b)检测。图16。t细胞介导的对过表达人cea的a549(肺腺癌)细胞(a549-hcea)的杀伤,与ceatcb(seqidno:22,56,57和58)和作为效应细胞的人pbmc(a,c)或猕猴pbmc(b,d)一起温育后21h(a,b)和40h(c,d)评估。图17。在0.8nm(a),4nm(b)和20nm(c),由ceatcb(seqidno:22,56,57和58)诱导的t细胞介导的对表达cea的人结肠直肠癌细胞系的杀伤。(d)cea表达和20nmceatcb的%比裂解之间的相关性,(e)cea表达和ceatcb的ec50之间的相关性。图18。ceatcb(seqidno:22,56,57和58)在与人pbmc共移植的ls174t-fluc2人结肠癌中的体内抗肿瘤功效(e:t比5:1)。结果显示不同研究组中来自12只小鼠的肿瘤体积的平均值和sem,通过测径器(a和c)和通过生物发光(总通量,b和d)测量。(a,b)第1天开始的早期处理,(c,d)第7天开始的延迟处理。使用mcsptcb(seqidno:12,53,54和55)作为阴性对照。图19。ceatcb(seqidno:22,56,57和58)在与人pbmc共移植的ls174t-fluc2人结肠癌中的体内抗肿瘤功效(e:t比1:1)。结果显示不同研究组中来自10只小鼠的肿瘤体积的平均值和sem,通过测径器(a)和通过生物发光(总通量,b)测量。使用mcsptcb(seqidno:12,53,54和55)作为阴性对照。图20。鼠源化ceatcb在免疫活性hucd3ε/hucea转基因小鼠中的panco2-hucea同位肿瘤模型中的体内功效。图21。ceatcb的热稳定性。以0.05℃/min在温度斜坡25-75℃中测量的动态光散射。以灰色显示副本。图22。mcsptcb的热稳定性。以0.05℃/min在温度斜坡25-75℃中测量的动态光散射。以灰色线显示副本。图23。由mcsptcb(seqidno:12,53,54和55)和mcsp1+1crossmabtcb抗体诱导的t细胞介导的(a)a375(高mcsp),(b)mv-3(中等mcsp)和(c)hct-116(低mcsp)肿瘤靶细胞杀伤。(d)使用ls180(mcsp阴性肿瘤细胞系)作为阴性对照。在靶细胞与抗体和效应细胞(人pbmc)一起温育后24h(a-d)和48h(e-h)评估肿瘤细胞杀伤。图24。由mcsptcb(seqidno:12,53,54和55)和mcsp1+1crossmabtcb抗体介导的对表达mcsp的肿瘤细胞(a375,a-d和mv-3,e-h)的t细胞杀伤后cd8+和cd4+t细胞上的cd25和cd69上调。图25。dp47gstcb(2+1crossfab-iggp329glala倒转的=“非靶向tcb”seqidno:59,60,61和62)的ce-sds分析,其含有dp47gs作为非结合抗体且含有人源化ch2527作为抗cd3抗体。显示的电泳图是dp47gstcb的sds-page:a)非还原性,b)还原性。发明详述定义除非在下文另外定义,术语在本文中如本领域中一般使用的那样使用。如本文中使用的,术语“抗原结合分子”在其最广义上指特异性结合抗原性决定簇的分子。抗原结合分子的例子是免疫球蛋白及其衍生物,例如片段。术语“双特异性”意指抗原结合分子能够特异性结合至少两种不同的抗原性决定簇。通常,双特异性抗原结合分子包含两种抗原结合位点,其中每种特异于不同的抗原性决定簇。在某些实施方案中,所述双特异性抗原结合分子能够同时结合两种抗原性决定簇,特别是在两种不同的细胞上表达的两种抗原性决定簇。如本文中使用的,术语“价”指抗原结合分子中规定数目的抗原结合位点的存在。因而,术语“对抗原的单价结合”指抗原结合分子中一个(且不超过一个)特异于抗原的抗原结合位点的存在。“抗原结合位点”指抗原结合分子上提供与抗原相互作用的位点,即一个或多个氨基酸残基。例如,抗体的抗原结合位点包含来自互补性决定区(cdr)的氨基酸残基。天然的免疫球蛋白分子通常具有两个抗原结合位点,fab分子通常具有单个抗原结合位点。如本文中使用的,术语“抗原结合模块”指特异性结合抗原性决定簇的多肽分子。在一个实施方案中,抗原结合模块能够将与其附接的实体(例如第二抗原结合模块)引导至靶部位,例如至特定类型的肿瘤细胞或携有抗原性决定簇的肿瘤基质。在另一个实施方案中,抗原结合模块能够经由其靶抗原例如t细胞受体复合物抗原来激活信号传导。抗原结合模块包括如本文中另外定义的抗体及其片段。具体的抗原结合模块包括抗体的抗原结合域,其包含抗体重链可变区和抗体轻链可变区。在某些实施方案中,抗原结合模块可以包含抗体恒定区,如本文中另外定义和本领域中已知的。可用的重链恒定区包括以下5种同种型中的任一种:α、δ、ε、γ或μ。可用的轻链恒定区包括以下2种同种型中的任一种:κ和λ。如本文中使用的,术语“抗原性决定簇”与“抗原”和“表位”同义,并且指多肽大分子上与抗原结合模块结合,从而形成抗原结合模块-抗原复合物的位点(例如氨基酸的连续区段或由不连续氨基酸的不同区构成的构象性构造)。可用的抗原性决定簇可以在例如肿瘤细胞表面上、病毒感染的细胞的表面上、其它患病细胞的表面上、免疫细胞的表面上、游离在血液血清中和/或在胞外基质(ecm)中找到。除非另外指示,本文中称为抗原的蛋白质(例如mcsp、cea、cd3)可以是来自任何脊椎动物来源,包括哺乳动物如灵长类(例如人)和啮齿类(例如小鼠和大鼠)的任何天然形式蛋白质。在一个具体的实施方案中,抗原是人蛋白。在对本文中的特定蛋白质进行提述的情况下,该术语涵盖“全长”、未加工的蛋白质以及起因于细胞中加工的蛋白质的任何形式。该术语还涵盖蛋白质的天然存在变体,例如剪接变体或等位变体。可用作抗原的例示性人蛋白包括但不限于:黑素瘤关联硫酸软骨素蛋白聚糖(mcsp),亦称为硫酸软骨素蛋白聚糖4(cspg4,uniprotno.q6uvk1(版本70),ncbirefseqno.np_001888.2);癌胚抗原(cea),亦称为癌胚抗原相关细胞粘着分子5(ceacam5,uniprotno.p06731(版本119),ncbirefseqno.np_004354.2);和cd3,特别是cd3的ε亚基(对于人序列,参见uniprotno.p07766(版本130)、ncbirefseqno.np_000724.1、seqidno:103;或对于猕猴[食蟹猴(macacafascicularis)]序列,参见uniprotno.q95li5(版本49)、ncbigenbankno.bab71849.1、seqidno:104)。在某些实施方案中,本发明的t细胞活化性双特异性抗原结合分子结合在来自不同物种的cd3或靶抗原间保守的cd3或靶细胞抗原的表位。在某些实施方案中,本发明的t细胞活化性双特异性抗原结合分子结合cd3和ceacam5,但是不结合ceacam1或ceacam6。“特异性结合”意指结合对于抗原是选择性的,并且能与不想要的或非特异性的相互作用区别开来。抗原结合模块结合特定抗原性决定簇的能力能经由酶联免疫吸附测定法(elisa)或本领域技术人员熟知的其它技术,例如表面等离振子共振技术(在biacore仪上分析)(liljeblad等,glycoj17,323-329(2000)),以及传统的结合测定法(heeley,endocrres28,217-229(2002))来测量。在一个实施方案中,抗原结合模块对无关蛋白的结合程度是该抗原结合模块对抗原结合的小于约10%,如例如通过spr测量的。在某些实施方案中,结合抗原的抗原结合模块,或包含该抗原结合模块的抗原结合分子具有≤1μm、≤100nm、≤10nm、≤1nm、≤0.1nm、≤0.01nm、或≤0.001nm(例如10-8m或更少,例如10-8m至10-13m,例如10-9m至10-13m)的解离常数(kd)。“亲和力”指分子(例如受体)的单一结合位点与其结合配偶体(例如配体)之间非共价相互作用总和的强度。除非另外指示,如本文中使用的,“结合亲和力”指反映结合对的成员(例如抗原结合模块和抗原,或受体及其配体)之间1:1相互作用的固有结合亲和力。分子x对其配偶体y的亲和力通常可以以解离常数(kd)来表述,其为解离与结合速率常数(分别为k解离和k结合)的比率。如此,相等的亲和力可能包含不同的速率常数,只要速率常数的比率保持相同。亲和力可以通过本领域知道的确立方法来测量,包括本文中描述的那些方法。用于测量亲和力的一种具体方法是表面等离振子共振(spr)。“降低的结合”,例如降低的对fc受体的结合,指相应相互作用的亲和力降低,如例如通过spr测量的。为了清楚,该术语还包括亲和力降低至0(或低于分析方法的检测限),即完全消除相互作用。相反,“升高的结合”指相应相互作用的亲和力升高。如本文中使用的,“t细胞活化”指t淋巴细胞,特别是细胞毒性t淋巴细胞的一种或多种细胞应答,其选自:增殖、分化、细胞因子分泌、细胞毒性效应分子释放、细胞毒性活性和活化标志物的表达。本发明的t细胞活化性双特异性抗原结合分子能够诱导t细胞活化。合适的测量t细胞活化的测定法是本文中所述
技术领域:
中已知的。如本文中使用的,“靶细胞抗原”指靶细胞表面上呈现的抗原性决定簇,所述靶细胞例如肿瘤中的细胞如癌细胞或肿瘤基质的细胞。如本文中使用的,术语“第一”和“第二”就抗原结合模块等而言为了在有超过一个每类模块时便于区分而使用。除非明确如此陈述,这些术语的使用不意图赋予t细胞活化性双特异性抗原结合分子的特定次序或取向。“fab分子”指由免疫球蛋白的重链(“fab重链”)的vh和ch1域以及轻链(“fab轻链”)的vl和cl域组成的蛋白质。“融合”意指组分(例如fab分子和fc域亚基)直接地或经由一种或多种肽接头通过肽键连接。如本文中使用的,术语“单链”指包含通过肽键线性连接的氨基酸单体的分子。在某些实施方案中,抗原结合模块之一是单链fab分子,即其中通过肽接头连接fab轻链和fab重链以形成单一肽链的fab分子。在一个具体的此类实施方案中,在单链fab分子中fab轻链的c端连接于fab重链的n端。“交换”fab分子(亦称为“crossfab”)意指其中交换fab重链和轻链的可变区或恒定区的fab分子,即交换fab分子包含由轻链可变区和重链恒定区构成的肽链,和由重链可变区和轻链恒定区构成的肽链。为了清楚,在其中交换fab轻链和fab重链的可变区的交换fab分子中,包含重链恒定区的肽链在本文中称为交换fab分子的“重链”。相反,在其中交换fab轻链和fab重链的恒定区的交换fab分子中,包含重链可变区的肽链在本文中称为交换fab分子的“重链”。与之对比,“常规”fab分子意指处于其天然型式的fab分子,即包含由重链可变和恒定区构成的重链(vh-ch1)和由轻链可变和恒定区构成的轻链(vl-cl)。术语“免疫球蛋白分子”指具有天然存在抗体的结构的蛋白质。例如,igg类的免疫球蛋白是约150,000道尔顿的异四聚体糖蛋白,其由二硫键连接的两条轻链和两条重链构成。从n端至c端,每条重链具有可变区(vh),也称为可变重域或重链可变域,接着是3个恒定域(ch1、ch2和ch3),也称为重链恒定区。类似地,从n端至c端,每条轻链具有可变区(vl),也称为可变轻域或轻链可变域,接着是恒定轻(cl)域(也称为轻链恒定区)。免疫球蛋白的重链可以归入称为α(iga)、δ(igd)、ε(ige)、γ(igg)或μ(igm)的5类之一,其中一些可以进一步分成亚类,例如γ1(igg1)、γ2(igg2)、γ3(igg3)、γ4(igg4)、α1(iga1)和α2(iga2)。基于其恒定域的氨基酸序列,免疫球蛋白的轻链可以归入称为卡帕(κ)和拉姆达(λ)的两类之一。免疫球蛋白基本由经由免疫球蛋白铰链区连接的两个fab分子和fc域组成。术语“抗体”在本文中以最广义使用且涵盖各种抗体结构,包括但不限于单克隆抗体、多克隆抗体和抗体片段,只要它们展现出期望的抗原结合活性。“抗体片段”指完整抗体外的分子,其包含完整抗体中结合与完整抗体结合的抗原的一部分。抗体片段的例子包括但不限于fv、fab、fab’、fab’-sh、f(ab’)2、双抗体、线性抗体、单链抗体分子(例如scfv)、和单域抗体。对于某些抗体片段的综述,参见hudson等,natmed9,129-134(2003)。对于scfv片段的综述,参见例如plückthun,于thepharmacologyofmonoclonalantibodies,vol.113,rosenburg和moore编,springer-verlag,newyork,pp.269-315(1994);亦参见wo93/16185;和美国专利no.5,571,894和5,587,458。关于包含补救受体结合表位残基且具有延长的体内半衰期的fab和f(ab’)2片段的论述,参见美国专利no.5,869,046。双抗体是具有两个抗原结合位点的抗体片段,其可以是二价或双特异性的。参见例如ep404,097;wo1993/01161;hudson等,natmed9,129-134(2003);和hollinger等,procnatlacadsciusa90,6444-6448(1993)。三抗体和四抗体也记载于hudson等,natmed9,129-134(2003)。单域抗体是包含抗体的整个或部分重链可变域、或整个或部分轻链可变域的抗体片段。在某些实施方案中,单域抗体是人单域抗体(domantis,inc.,waltham,ma;参见例如美国专利no.6,248,516b1)。可以通过各种技术来制备抗体片段,包括但不限于对完整抗体的蛋白水解消化以及通过重组宿主细胞(例如大肠杆菌或噬菌体)产生,如本文中描述的。术语“抗原结合域”指包含特异性结合部分或整个抗原且与其互补的区域的抗体部分。抗原结合域可由例如一个或多个抗体可变域(也称为抗体可变区)提供。具体地,抗原结合域包含抗体轻链可变区(vl)和抗体重链可变区(vh)。术语“可变区”或“可变域”指抗体重链或轻链中牵涉使抗体结合抗原的域。天然抗体的重链和轻链的可变域(分别为vh和vl)一般具有类似的结构,每个域包含4个保守的框架区(fr)和3个高变区(hvr)。参见例如kindt等,kubyimmunology,第6版,w.h.freemanandco.,第91页(2007)。单个vh或vl域可能足以赋予抗原结合特异性。如本文中使用的,术语“高变区”或“hvr”指抗体可变域中序列中高度可变和/或形成结构上定义的环(“高变环”)的每个区域。通常,天然的四链抗体包含六个hvr;三个在vh中(h1、h2、h3),三个在vl中(l1、l2、l3)。hvr一般包含来自高变环和/或来自互补性决定区(cdr)的氨基酸残基,后者具有最高序列变异性和/或涉及抗原识别。除了vh中cdr1外,cdr一般包含形成高变环的氨基酸残基。高变区(hvr)也称为“互补性决定区”(cdr),并且在述及形成抗原结合区的可变区部分时,这些术语在本文中可交换使用。此特定区域已由kabat等,u.s.dept.ofhealthandhumanservices,sequencesofproteinsofimmunologicalinterest(1983)及由chothia等,jmolbiol196:901-917(1987)描述,其中定义包括在彼此比较时氨基酸残基的重叠或子集。然而,应用任一种定义来指抗体或其变体的cdr意图在如本文中定义和使用的术语的范围内。涵盖如由上文引用的每篇参考文献定义的cdr的适宜的氨基酸残基在下表a中列出作为比较。涵盖特定cdr的确切残基数将随着cdr的序列和大小而变化。鉴于抗体的可变区氨基酸序列,本领域技术人员可以常规确定哪些残基构成特定cdr。表a:cdr定义1cdrkabatchothiaabm2vhcdr131-3526-3226-35vhcdr250-6552-5850-58vhcdr395-10295-10295-102vlcdr124-3426-3224-34vlcdr250-5650-5250-56vlcdr389-9791-9689-971表a中所有cdr定义的编号方式依照由kabat等提出的编号惯例(见下文)。2如表a中使用的具有小写字母“b”的“abm”指由oxfordmolecular的“abm”抗体建模软件定义的cdr。kabat等还定义针对可变区序列的编号系统,其可应用于任何抗体。本领域的普通技术人员可以明确地将此“kabat编号”系统归入任何可变区序列,不依赖于序列本身外的任何实验数据。如本文中使用的,“kabat编号方式”指由kabat等,u.s.dept.ofhealthandhumanservices,“sequenceofproteinsofimmunologicalinterest”(1983)提出的编号系统。除非另外说明,提及抗体可变区中特定氨基酸残基位置的编号方式依照kabat编号系统。序列表的多肽序列并不依照kabat编号系统编号。然而,本领域中普通技术人员完全能将序列表的序列编号方式转变成kabat编号方式。“框架”或“fr”指除高变区(hvr)残基外的可变域残基。可变域的fr一般由4个fr域组成:fr1、fr2、fr3和fr4。因而,hvr和fr序列一般以下列顺序出现在vh(或vl)中:fr1-h1(l1)-fr2-h2(l2)-fr3-h3(l3)-fr4。抗体或免疫球蛋白的“类”指其重链拥有的恒定域或恒定区的类型。抗体有5种主要的类:iga、igd、ige、igg和igm,并且这些中数种可以进一步分成亚类(同种型),例如igg1、igg2、igg3、igg4、iga1和iga2。对应于不同免疫球蛋白类的重链恒定域分别称为α、δ、ε、γ和μ。本文中术语“fc域”或“fc区”用于定义免疫球蛋白重链中至少含有恒定区的一部分的c端区域。该术语包括天然序列fc区和变体fc区。虽然igg重链的fc区的边界可以略微变化,但是人igg重链fc区通常定义为自cys226或pro230延伸至重链的羧基端。然而,可以存在或不存在fc区的c端赖氨酸(lys447)。除非本文中另外指定,fc区或恒定区中氨基酸残基的编号方式依照eu编号系统,也称为eu索引,如记载于kabat等,sequencesofproteinsofimmunologicalinterest,第5版publichealthservice,nationalinstitutesofhealth,bethesda,md,1991的。如本文中使用的,fc域的“亚基”指形成二聚体fc域的两个多肽之一,即包含免疫球蛋白重链中能够稳定自身联合的c端恒定区的多肽。例如,iggfc域的亚基包含iggch2和iggch3恒定域。“促进fc域的第一和第二亚基联合的修饰”是降低或防止包含fc域亚基的多肽与相同多肽联合以形成同二聚体的肽主链操作或fc域亚基的翻译后修饰。如本文中使用的,具体地,促进联合的修饰包括对期望联合的两个fc域亚基(即fc域的第一和第二亚基)中的每一个进行的分开的修饰,其中所述修饰彼此互补,从而促进两个fc域亚基的联合。例如,促进联合的修饰可以改变一种或两种fc域亚基的结构或电荷,从而在立体或静电上分别促进它们的联合。如此,异二聚化在包含第一fc域亚基的多肽和包含第二fc域亚基的多肽之间发生,其在融合至每个亚基的别的组分(例如抗原结合模块)不同这一意义上讲可能是不相同的。在一些实施方案中,促进联合的修饰包含在fc域中的氨基酸突变,具体为氨基酸替代。在一个具体的实施方案中,促进联合的修饰包含fc域的两个亚基的每一个中分开的氨基酸突变,具体为氨基酸替代。术语“效应器功能”指那些可归于抗体fc区且随抗体同种型而变化的生物学活性。抗体效应器功能的例子包括:c1q结合和补体依赖性细胞毒性(cdc)、fc受体结合、抗体依赖性细胞介导的细胞毒性(adcc)、抗体依赖性细胞吞噬作用(adcp)、细胞因子分泌、免疫复合物介导的抗原呈递细胞的抗原摄取、细胞表面受体(例如b细胞受体)下调和b细胞活化。如本文中使用的,术语“工程化”视为包括对肽主链的任何操作或对天然存在或重组的多肽或其片段的翻译后修饰。工程化包括对氨基酸序列、糖基化模式或各氨基酸侧链基团的修饰,以及这些办法的组合。如本文中使用的,术语“氨基酸突变”意为涵盖氨基酸替代、缺失、插入和修饰。可以进行取代、缺失、插入和修饰的任意组合来实现最终构建体,只要最终构建体拥有期望的特性,例如降低的对fc受体的结合,或与另一种肽的增加的联合。氨基酸序列缺失和插入包括氨基和/或羧基端缺失和氨基酸插入。具体的氨基酸突变是氨基酸替代。为了改变例如fc区的结合特征,特别优选非保守性的氨基酸替代,即将一个氨基酸用具有不同结构和/或化学特性的另一种氨基酸替换。氨基酸替代包括由非天然存在的氨基酸或由20种标准氨基酸的天然存在的氨基酸衍生物(例如4-羟脯氨酸、3-甲基组氨酸、鸟氨酸、高丝氨酸、5-羟赖氨酸)替换。可以使用本领域中公知的遗传或化学方法生成氨基酸突变。遗传方法可以包括定点诱变、pcr、基因合成等。通过与遗传工程化不同的方法如化学修饰来改变氨基酸侧链基团的方法也可能可用。本文中可使用各种名称来指示同一氨基酸突变。例如,从fc域第329位脯氨酸到甘氨酸的取代可指示为329g、g329、g329、p329g或pro329gly。如本文中使用的,术语“多肽”指由通过酰胺键(也称为肽键)线性连接的单体(氨基酸)构成的分子。术语“多肽”指具有两个或更多个氨基酸的任何链,并且不指特定长度的产物。如此,肽、二肽、三肽、寡肽、“蛋白质”、“氨基酸链”或任何其它用于指具有两个或更多个氨基酸的链的术语均包括在“多肽”的定义中,而且术语“多肽”可以代替这些术语中任一个或与其交换使用。术语“多肽”还意图指多肽的表达后修饰的产物,包括但不限于糖基化、乙酰化、磷酸化、酰化、通过已知的保护性/封闭性基团衍生化、蛋白水解切割、或通过非天然存在的氨基酸的修饰。多肽可以自天然的生物学来源衍生或通过重组技术生成,但不必从指定的核酸序列翻译。它可以以任何方式来生成,包括通过化学合成。本发明的多肽大小可以是约3个以上、5个以上、10个以上、20个以上、25个以上、50个以上、75个以上、100个以上、200个以上、500个以上、1,000个以上、或2,000个以上的氨基酸。多肽可以具有限定的三维结构,尽管它们不必具有此类结构。具有限定的三维结构的多肽被称为折叠的,而不具有限定的三维结构而可以采用大量不同构象的多肽被称为未折叠的。“分离的”多肽或其变体或衍生物意图为不处于其天然环境中的多肽。不需要特定水平的纯化。例如,分离的多肽可以是从其天然或自然环境中取出。就本发明而言,在宿主细胞中表达的重组生成的多肽和蛋白质被视为分离的,已通过任何合适的技术分开、分级、或部分或基本上纯化的天然的或重组的多肽也是如此。关于参照多肽序列的“百分比(%)氨基酸序列同一性”定义为在比对序列并在必要时引入缺口以获取最大百分比序列同一性后,且不将任何保守性取代视为序列同一性的一部分时,候选序列中与参照多肽序列中的氨基酸残基相同的氨基酸残基的百分比。为测定百分比氨基酸序列同一性目的比对可以以本领域技术范围内的多种方式进行,例如使用公众可得到的计算机软件,如blast、blast-2、align或megalign(dnastar)软件。本领域技术人员可决定用于比对序列的适宜参数,包括在比较序列的全长里获得最大比对需要的任何算法。然而,就本文中目的而言,使用序列比较计算机程序align-2来生成%氨基酸序列同一性值。align-2序列比较计算机程序由genentech,inc.创作,并且源代码已与用户文档一起提交到美国版权局(u.s.copyrightoffice),washingtond.c.,20559,其在美国版权注册no.txu510087下注册。align-2程序可从genentech,inc.,southsanfrancisco,california公开获得,或可从源代码汇编。align-2程序应当汇编用于unix操作系统,包括数字unixv4.0d。所有序列比较参数均由align-2程序设定且不改变。在采用align-2进行氨基酸序列比较的情况下,给定的氨基酸序列a对/与/相对给定的氨基酸序列b的%氨基酸序列同一性(或其可以用短语表示为对/与/相对给定的氨基酸序列b具有或包含特定%氨基酸序列同一性的给定氨基酸序列a)如下计算:分数x/y的100倍其中x是由序列比对程序align-2在所述程序对a和b的比对中评为相同匹配的氨基酸残基数,而其中y是b中氨基酸残基的总数。会领会的是,当氨基酸序列a的长度不等于氨基酸序列b的长度时,a对b的%氨基酸序列同一性将不等于b对a的%氨基酸序列同一性。除非另外明确说明,本文中使用的所有%氨基酸序列同一性值如在上一段中描述的那样使用align-2计算机程序获得。术语“多核苷酸”指分离的核酸分子或构建体,例如信使rna(mrna)、病毒衍生的rna或质粒dna(pdna)。多核苷酸可以包含常规的磷酸二酯键或非常规的键(例如酰胺键,如在肽核酸(pna)中发现的)。术语“核酸分子”指任何一种或多种存在于多核苷酸中的核酸区段,例如dna或rna片段。“分离的”核酸分子或多核苷酸意指已从其天然环境取出的核酸分子、dna或rna。例如,就本发明而言,包含在载体中的编码多肽的重组多核苷酸被视为分离的。分离的多核苷酸的别的例子包括在异源宿主细胞中保持的重组多核苷酸或溶液中的(部分或基本上)纯化的多核苷酸。分离的多核苷酸包括在普遍含有该多核苷酸分子的细胞中含有的多核苷酸分子,但该多核苷酸分子存在于染色体外或在不同于其天然染色体位置的染色体位置处。分离的rna分子包括本发明的体内或体外rna转录本,以及正链和负链形式、和双链形式。依照本发明的分离的多核苷酸或核酸还包括合成生成的此类分子。另外,多核苷酸或核酸可以为或可以包括调节元件如启动子、核糖体结合位点或转录终止子。与本发明的参照核苷酸序列具有至少例如95%“相同的”核苷酸序列的核酸或多核苷酸意指该多核苷酸的核苷酸序列与参照序列相同,只不过按照参照核苷酸序列的每100个核苷酸,该多核苷酸序列可以包含多达5处点突变。换言之,为了获得与参照核苷酸序列具有至少95%相同的核苷酸序列的多核苷酸,可以删除或用另一种核苷酸取代参照序列中高达5%的核苷酸,或者可以将参照序列中占总核苷酸的高达5%的数目的核苷酸插入到参照序列中。参照序列的这些变更可以发生在参照核苷酸序列的5’或3’端位置或那些末端位置之间的任何地方,个别分散在参照序列中的残基中或分散在参照序列内的一或多个连续组中。作为一个实际问题,可以使用已知的计算机程序,如上文针对多肽论述的程序(例如align-2)来常规确定任何特定的多核苷酸序列是否与本发明的核苷酸序列为至少80%、85%、90%、95%、96%、97%、98%或99%相同。术语“表达盒”指重组或合成生成的,具有一系列允许特定核酸在靶细胞中转录的指定核酸元件的多核苷酸。可以将重组表达盒掺入质粒、染色体、线粒体dna、质体dna、病毒或核酸片段中。通常,表达载体的重组表达盒部分包含要转录的核酸序列和启动子等。在某些实施方案中,本发明的表达盒包含编码本发明的双特异性抗原结合分子或其片段的多核苷酸序列。术语“载体”或“表达载体”与“表达构建体”同义,并指用于在靶细胞中导入与其可操作联合的特定基因及指导表达的dna分子。该术语包括作为自主复制核酸结构的载体以及掺入到已经接受其导入的宿主细胞的基因组中的载体。本发明的表达载体包含表达盒。表达盒允许转录大量稳定的mrna。一旦表达载体在靶细胞内,就通过细胞转录和/或翻译装置生成基因编码的核糖核酸分子或蛋白质。在一个实施方案中,本发明的表达载体包含表达盒,其包含编码本发明的双特异性抗原结合分子或其片段的多核苷酸序列。术语“宿主细胞”、“宿主细胞系”和“宿主细胞培养物”可交换使用并指已引入外源核酸的细胞,包括此类细胞的后代。宿主细胞包括“转化体”和“经转化的细胞”,其包括初始转化的细胞和自其衍生的后代(不考虑传代数)。后代在核酸内含物上可能与亲本细胞不完全相同,但可以含有突变。本文中包括具有如原始转化细胞中筛选或选择的相同的功能或生物学活性的突变体后代。宿主细胞是能用于生成本发明的双特异性抗原结合分子的任何类型的细胞系统。宿主细胞包括培养的细胞,例如哺乳动物培养细胞如cho细胞、bhk细胞、ns0细胞、sp2/0细胞、yo骨髓瘤细胞、p3x63小鼠骨髓瘤细胞、per细胞、per.c6细胞或杂交瘤细胞、酵母细胞、昆虫细胞和植物细胞等,而且还包括在转基因动物、转基因植物或培养的植物或动物组织中包含的细胞。“激活fc受体”是一种在抗体的fc域衔接后,引发刺激携带该受体的细胞实施效应器功能的信号传导事件的fc受体。人激活fc受体包括fcγriiia(cd16a)、fcγri(cd64)、fcγriia(cd32)和fcαri(cd89)。抗体依赖性细胞介导的细胞毒性(adcc)是一种导致通过免疫效应器细胞对抗体包被的靶细胞裂解的免疫机制。靶细胞是包含fc区的抗体或其衍生物一般经由fc区的n端的蛋白质部分特异性结合的细胞。如本文中使用的,术语“降低的adcc”定义为通过上文定义的adcc机制,以靶细胞周围介质中给定浓度的抗体,在给定的时间内裂解的靶细胞数目的降低,和/或通过adcc机制,实现给定时间内给定数目的靶细胞裂解需要的靶细胞周围介质中抗体浓度的增加。adcc的降低相对于使用相同的标准生产、纯化、配制和贮存方法(其是本领域技术人员已知的)、由同一类型的宿主细胞生成但尚未工程化改造的相同抗体介导的adcc。例如,由在其fc域包含降低adcc的氨基酸替代的抗体所介导的adcc中的降低,是相对于由在fc域中无此氨基酸替代的相同抗体介导的adcc而言。测量adcc的合适测定法是本领域中公知的(参见例如pct公开文本no.wo2006/082515或pct公开文本no.wo2012/130831)。药剂的“有效量”指引起接受其施用的细胞或组织中的生理学变化必需的量。药剂例如药物组合物的“治疗有效量”指有效实现期望的治疗或预防结果的量(以必要的剂量且持续必要的时间)。治疗有效量的药剂例如消除、降低、延迟、最小化或预防疾病的不良作用。“个体”或“受试者”是哺乳动物。哺乳动物包括但不限于驯养的动物(例如牛、绵羊、猫、犬和马)、灵长类(例如人和非人灵长类如猴)、家兔和啮齿动物(例如小鼠和大鼠)。优选地,所述个体或受试者是人。术语“药物组合物”指其形式使得容许其中含有的活性成分的生物学活性有效,且不含对会接受配制剂施用的受试者有不可接受的毒性的别的成分的制剂。“药学可接受载体”指药物组合物中活性成分以外对受试者无毒的成分。药学可接受载体包括但不限于缓冲剂、赋形剂、稳定剂或防腐剂。如本文中使用的,“治疗/处理”(及其语法变体)指试图改变治疗个体中疾病的自然进程,并且可以是为了预防或在临床病理学的过程期间实施的临床干预。治疗的期望效果包括但不限于预防疾病的发生或复发、缓解症状、降低疾病的任何直接或间接病理学后果、预防转移、减缓疾病进展率、改善或减轻疾病状态、及消退或改善的预后。在一些实施方案中,本发明的t细胞活化性双特异性抗原结合分子用于延迟疾病的形成或延缓病症的进展。术语“包装插页”用于指治疗产品的商业化包装中通常含有的说明书,其含有关于适应症、使用、剂量、施用、组合疗法、禁忌症的信息和/或关于使用此类治疗产品的警告。实施方案的详细描述在第一个方面,本发明提供一种t细胞活化性双特异性抗原结合分子,其包含(i)第一抗原结合模块,它是能够特异性结合cd3的fab分子,且包含至少一个选自seqidno:4,seqidno:5和seqidno:6的重链互补决定区(cdr)和至少一个选自seqidno:8,seqidno:9和seqidno:10的轻链cdr;(ii)第二抗原结合模块,它是能够特异性结合靶细胞抗原的fab分子。在一个实施方案中,所述第一抗原结合模块包含包含与选自seqidno:3,seqidno:32和seqidno:33的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与选自seqidno:7和seqidno:31的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个实施方案中,所述第一抗原结合模块包含包含与seqidno:3的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:7的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个特定的实施方案中,所述第二抗原结合模块能够特异性结合cea且包含至少一个选自seqidno:24,seqidno:25和seqidno:26的重链互补决定区(cdr)和至少一个选自seqidno:28,seqidno:29和seqidno:30的轻链cdr。在另一个特定的实施方案中,所述第二抗原结合模块能够特异性结合cea且包含包含与seqidno:23的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:27的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在另一个特定的实施方案中,所述第二抗原结合模块能够特异性结合mcsp且包含至少一个选自seqidno:14,seqidno:15,seqidno:16,seqidno:35,seqidno:37,seqidno:38和seqidno:40的重链互补决定区(cdr)和至少一个选自seqidno:18,seqidno:19,seqidno:20,seqidno:44,seqidno:45,seqidno:48,seqidno:49和seqidno:50的轻链cdr。在另一个特定的实施方案中,所述第二抗原结合模块能够特异性结合mcsp且包含至少一个选自seqidno:14,seqidno:15和seqidno:16的重链互补决定区(cdr)和至少一个选自seqidno:18,seqidno:19和seqidno:20的轻链cdr。在另一个特定的实施方案中,所述第二抗原结合模块能够特异性结合mcsp且包含包含与选自seqidno:13,seqidno:34,seqidno:36,seqidno:39和seqidno:41的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与选自seqidno:17,seqidno:43,seqidno:46,seqidno:47和seqidno:51的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在另一个特定的实施方案中,所述第二抗原结合模块能够特异性结合mcsp且包含包含与seqidno:13的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:17的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个实施方案中,本发明提供t细胞活化性双特异性抗原结合分子,其包含:(i)第一抗原结合模块,它是能够特异性结合cd3的fab分子,包含至少一个选自seqidno:4,seqidno:5和seqidno:6的重链互补决定区(cdr)和至少一个选自seqidno:8,seqidno:9和seqidno:10的轻链cdr;(ii)第二抗原结合模块,它是能够特异性结合cea的fab分子,包含至少一个选自seqidno:24,seqidno:25和seqidno:26的重链互补决定区(cdr)和至少一个选自seqidno:28,seqidno:29和seqidno:30的轻链cdr。在一个实施方案中,本发明提供t细胞活化性双特异性抗原结合分子,其包含:(i)第一抗原结合模块,它是能够特异性结合cd3的fab分子,包含包含与seqidno:3的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:7的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区;(ii)第二抗原结合模块,它是能够特异性结合cea的fab分子,包含包含与seqidno:23的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:27的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个实施方案中,本发明提供t细胞活化性双特异性抗原结合分子,其包含:(i)第一抗原结合模块,它是能够特异性结合cd3的fab分子,包含至少一个选自seqidno:4,seqidno:5和seqidno:6的重链互补决定区(cdr)和至少一个选自seqidno:8,seqidno:9和seqidno:10的轻链cdr;(ii)第二抗原结合模块,它是能够特异性结合mcsp的fab分子,包含至少一个选自seqidno:14,seqidno:15和seqidno:16的重链互补决定区(cdr)和至少一个选自seqidno:18,seqidno:19和seqidno:20的轻链cdr。在一个实施方案中,本发明提供t细胞活化性双特异性抗原结合分子,其包含:(i)第一抗原结合模块,它是能够特异性结合cd3的fab分子,包含包含与seqidno:3的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:7的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区;(ii)第二抗原结合模块,它是能够特异性结合mcsp的fab分子,包含包含与seqidno:13的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:17的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个具体的实施方案中,所述第一抗原结合模块是交换fab分子,其中fab轻链和fab重链的可变区或恒定区任一是交换的。在一个实施方案中,所述第二抗原结合模块是常规fab分子。在一个具体的实施方案中,所述第一抗原结合模块是交换fab分子,其中fab轻链和fab重链的恒定区是交换的,且所述第二抗原结合模块是常规fab分子。在又一个具体的实施方案中,所述第一和第二抗原结合模块彼此融合,任选地经由肽接头。在具体的实施方案中,所述t细胞活化性双特异性抗原结合分子进一步包含由能够稳定联合的第一和第二亚基构成的fc域。在又一个具体的实施方案中,所述t细胞活化性双特异性抗原结合分子中存在不超过一个能够特异性结合cd3的抗原结合模块(即所述t细胞活化性双特异性抗原结合分子提供对cd3的单价结合)。t细胞活化性双特异性抗原结合分子形式t细胞活化性双特异性抗原结合分子的组分可以以多种构造彼此融合。例示性的构造绘于图1、3和5中。在具体的实施方案中,所述t细胞活化性双特异性抗原结合分子包含由能够稳定联合的第一和第二亚基构成的fc域。在一些实施方案中,所述第二抗原结合模块在fab重链的c端融合至所述fc域的第一或第二亚基的n端。在一个此类实施方案中,第一抗原结合模块在fab重链的c端融合至第二抗原结合模块的fab重链的n端。在一个特定的此类实施方案中,t细胞活化性双特异性抗原结合分子基本由以下组成:第一和第二抗原结合模块,由第一和第二亚基构成的fc域和任选地一个或多个肽接头,其中所述第一抗原结合模块在fab重链的c端融合至第二抗原结合模块的fab重链的n端,而第二抗原结合模块在fab重链的c端融合至所述fc域的第一或第二亚基的n端。任选地,第一抗原结合模块的fab轻链和第二抗原结合模块的fab轻链可以另外彼此融合。在另一个此类实施方案中,第一抗原结合模块在fab重链的c端融合至所述fc域的第一或第二亚基的n端。在一个特定的此类实施方案中,t细胞活化性双特异性抗原结合分子基本由以下组成:第一和第二抗原结合模块,由第一和第二亚基构成的fc域和任选地一个或多个肽接头,其中所述第一和第二抗原结合模块各自在fab重链的c端融合至所述fc域的亚基之一的n端。在其他实施方案中,第一抗原结合模块在fab重链的c端融合至所述fc域的第一或第二亚基的n端。在一个具体的此类实施方案中,第二抗原结合模块在fab重链的c端融合至第一抗原结合模块的fab重链的n端。在一个特定的此类实施方案中,t细胞活化性双特异性抗原结合分子基本由以下组成:第一和第二抗原结合模块,由第一和第二亚基构成的fc域和任选地一个或多个肽接头,其中所述第二抗原结合模块在fab重链的c端融合至第一抗原结合模块的fab重链的n端,而第一抗原结合模块在fab重链的c端融合至所述fc域的第一或第二亚基的n端。任选地,第一抗原结合模块的fab轻链和第二抗原结合模块的fab轻链可以另外彼此融合。抗原结合模块可以直接地或经由肽接头融合至fc域或彼此融合,所述肽接头包含一个或多个氨基酸,通常约2-20个氨基酸。肽接头是本领域中已知且本文中记载的。合适的、非免疫原性的肽接头包括例如(g4s)n、(sg4)n、(g4s)n或g4(sg4)n肽接头。“n”一般是介于1和10之间的数字,通常介于2和4之间。特别适用于将第一和第二抗原结合模块的fab轻链彼此融合的肽接头是(g4s)2。适用于连接第一和第二抗原结合模块的fab重链的例示性肽接头是epksc(d)-(g4s)2(seqidno105和106)。另外,接头可包含免疫球蛋白铰链区(的部分)。尤其当抗原结合模块融合至fc域亚基的n端时,其可以用或不用另外的肽接头经由免疫球蛋白铰链区或其部分融合。具有能够特异性结合靶细胞抗原的单一抗原结合模块的t细胞活化性双特异性抗原结合分子(例如如图1小图a、d、f或h中显示的)是有用的,特别是在高亲和力抗原结合模块结合后会预期靶细胞抗原内在化的情况中。在此类情况中,存在超过一个特异于靶细胞抗原的抗原结合模块可能增强靶细胞抗原的内在化,由此降低其利用度。然而,在许多其它情况中,会有利的是具有包含两个或更多个特异于靶细胞抗原的抗原结合模块的t细胞活化性双特异性抗原结合分子(见显示于图1小图b、c、e或g的例子),从而例如优化对靶部位的靶向或允许靶细胞抗原的交联。因而,在某些实施方案中,本发明的t细胞活化性双特异性抗原结合分子还包含第三抗原结合模块,其为能够特异性结合靶细胞抗原的fab分子。在一个实施方案中,所述第三抗原结合模块是常规fab分子。在一个实施方案中,所述第三抗原结合模块能够与第二抗原结合模块特异性结合相同的靶细胞抗原。在一个具体的实施方案中,所述第一抗原结合模块能够特异性结合cd3,而所述第二和第三抗原结合模块能够特异性结合靶细胞抗原。在一个具体的实施方案中,所述第二和第三抗原结合模块是相同的(即它们包含相同的氨基酸序列)。在一个具体的实施方案中,所述第一抗原结合模块能够特异性结合cd3,且所述第二和第三抗原结合模块能够特异性结合cea,其中所述第二和第三抗原结合模块包含至少一个选自seqidno:24,seqidno:25和seqidno:26的重链互补决定区(cdr)和至少一个选自seqidno:28,seqidno:29和seqidno:30的轻链cdr。在一个具体的实施方案中,所述第一抗原结合模块能够特异性结合cd3,且包含至少一个选自seqidno:4,seqidno:5和seqidno:6的重链互补决定区(cdr)和至少一个选自seqidno:8,seqidno:9和seqidno:10的轻链cdr;且所述第二和第三抗原结合模块能够特异性结合cea,其中所述第二和第三抗原结合模块包含至少一个选自seqidno:24,seqidno:25和seqidno:26的重链互补决定区(cdr)和至少一个选自seqidno:28,seqidno:29和seqidno:30的轻链cdr。在一个具体的实施方案中,所述第一抗原结合模块能够特异性结合cd3,其包含至少一个选自seqidno:4,seqidno:5和seqidno:6的重链互补决定区(cdr)和至少一个选自seqidno:8,seqidno:9和seqidno:10的轻链cdr;且所述第二和第三抗原结合模块能够特异性结合cea,其中所述第二和第三抗原结合模块包含至少一个选自seqidno:24,seqidno:25和seqidno:26的重链互补决定区(cdr)和至少一个选自seqidno:28,seqidno:29和seqidno:30的轻链cdr。在一个具体的实施方案中,所述第一抗原结合模块能够特异性结合cd3,其包含包含与选自seqidno:3,seqidno:32和seqidno:33的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与选自seqidno:7和seqidno:31的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区,且所述第二和第三抗原结合模块能够特异性结合cea,其中所述第二和第三抗原结合模块包含包含与seqidno:23的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:27的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个具体的实施方案中,所述第一抗原结合模块能够特异性结合cd3,且包含包含与seqidno:3的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:7的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区,且所述第二和第三抗原结合模块能够特异性结合cea,其中所述第二和第三抗原结合模块包含包含与seqidno:23的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:27的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个实施方案中,所述第一抗原结合模块能够特异性结合cd3,且所述第二和第三抗原结合模块能够特异性结合mcsp,其中所述第二和第三抗原结合模块包含至少一个选自seqidno:14,seqidno:15,seqidno:16,seqidno:35,seqidno:37,seqidno:38和seqidno:40的重链互补决定区(cdr)和至少一个选自seqidno:18,seqidno:19,seqidno:20,seqidno:44,seqidno:45,seqidno:48,seqidno:49和seqidno:50的轻链cdr。在一个具体的实施方案中,所述第一抗原结合模块能够特异性结合cd3,其包含至少一个选自seqidno:4,seqidno:5和seqidno:6的重链互补决定区(cdr)和至少一个选自seqidno:8,seqidno:9和seqidno:10的轻链cdr;且所述第二和第三抗原结合模块能够特异性结合mcsp,其中所述第二和第三抗原结合模块包含至少一个选自seqidno:14,seqidno:15,seqidno:16,seqidno:35,seqidno:37,seqidno:38和seqidno:40的重链互补决定区(cdr)和至少一个选自seqidno:18,seqidno:19,seqidno:20,seqidno:44,seqidno:45,seqidno:48,seqidno:49和seqidno:50的轻链cdr。在一个实施方案中,所述第一抗原结合模块能够特异性结合cd3,其包含至少一个选自seqidno:4,seqidno:5和seqidno:6的重链互补决定区(cdr)和至少一个选自seqidno:8,seqidno:9和seqidno:10的轻链cdr;且所述第二和第三抗原结合模块能够特异性结合mcsp,其中所述第二和第三抗原结合模块包含至少一个选自seqidno:14,seqidno:15和seqidno:16的重链互补决定区(cdr)和至少一个选自seqidno:18,seqidno:19和seqidno:20的轻链cdr。在一个实施方案中,所述第一抗原结合模块能够特异性结合cd3,其包含包含与选自seqidno:3,seqidno:32和seqidno:33的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与选自seqidno:7和seqidno:31的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区,且所述第二和第三抗原结合模块能够特异性结合mcsp,其中所述第二和第三抗原结合模块包含包含与选自seqidno:13,seqidno:34,seqidno:36,seqidno:39和seqidno:41的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与选自seqidno:17,seqidno:43,seqidno:46,seqidno:47和seqidno:51的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个实施方案中,所述第一抗原结合模块能够特异性结合cd3,其包含包含与seqidno:3的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:7的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区,且所述第二和第三抗原结合模块能够特异性结合mcsp,其中所述第二和第三抗原结合模块包含包含与seqidno:13的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:17的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个实施方案中,所述第三抗原结合模块在fab重链的c端融合至所述fc域的第一或第二亚基的n端。在一个更加具体的实施方案中,所述第二和第三抗原结合模块各自在fab重链的c端融合至所述fc域的亚基之一的n端,且所述第一抗原结合模块在fab重链的c端融合至所述第二抗原结合模块的fab重链的n端。任选地,第一抗原结合模块的fab轻链和第二抗原结合模块的fab轻链可以另外彼此融合。所述第二和第三抗原结合模块可以直接或经由肽接头融合至fc域。在一个具体的实施方案中,所述第二和第三抗原结合模块各自经由免疫球蛋白铰链区融合至fc域。在一个特定的实施方案中,所述免疫球蛋白铰链区是人igg11铰链区。在一个实施方案中,所述第二和第三抗原结合模块和fc域是免疫球蛋白分子的部分。在一个具体的实施方案中,所述免疫球蛋白分子是igg类免疫球蛋白。在一个甚至更具体的实施方案中,所述免疫球蛋白是igg1亚类免疫球蛋白。在另一个实施方案中,所述免疫球蛋白是igg4亚类免疫球蛋白。在一个别的具体的实施方案中,所述免疫球蛋白是人免疫球蛋白。在其他实施方案中,免疫球蛋白是嵌合免疫球蛋白或人源化免疫球蛋白。在一个实施方案中,t细胞活化性双特异性抗原结合分子基本由以下组成:能够特异性结合靶细胞抗原的免疫球蛋白分子,和能够特异性结合cd3的抗原结合模块,其中所述抗原结合模块是融合至免疫球蛋白重链之一(任选地经由肽接头)的n端的fab分子,特别是交换fab分子。在一个具体的实施方案中,所述第一和第三抗原结合模块各自在fab重链的c端融合至所述fc域的亚基之一的n端,且所述第二抗原结合模块在fab重链的c端融合至所述第一抗原结合模块的fab重链的n端。在一个特定的此类实施方案中,所述t细胞活化性双特异性抗原结合分子基本由以下组成:第一、第二和第三抗原结合模块,由第一和第二亚基构成的fc域和任选地一个或多个肽接头,其中所述第二抗原结合模块在fab重链的c端融合至第一抗原结合模块的fab重链的n端,且所述第一抗原结合模块在fab重链的c端融合至所述fc域的第一亚基的n端,且其中所述第三抗原结合模块在fab重链的c端融合至所述fc域的第二亚基的n端。任选地,第一抗原结合模块的fab轻链和第二抗原结合模块的fab轻链可以另外彼此融合。在一个实施方案中,本发明提供t细胞活化性双特异性抗原结合分子,其包含:(i)第一抗原结合模块,它是能够特异性结合cd3的fab分子,包含seqidno:4的重链互补决定区(cdr)1,seqidno:5的重链cdr2,seqidno:6的重链cdr3,seqidno:8的轻链cdr1,seqidno:9的轻链cdr2和seqidno:10的轻链cdr3,其中所述第一抗原结合模块是交换fab分子,其中fab轻链和fab重链的可变或恒定区任一(特别是恒定区)是交换的;(ii)第二和第三抗原结合模块,它们各自是能够特异性结合cea的fab分子,包含seqidno:24的重链cdr1,seqidno:25的重链cdr2,seqidno:26的重链cdr3,seqidno:28的轻链cdr1,seqidno:29的轻链cdr2和seqidno:30的轻链cdr3。在一个实施方案中,本发明提供t细胞活化性双特异性抗原结合分子,其包含:(i)第一抗原结合模块,它是能够特异性结合cd3的fab分子,包含包含与seqidno:3的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:7的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区,其中所述第一抗原结合模块是交换fab分子,其中fab轻链和fab重链的可变或恒定区任一(特别是恒定区)是交换的;(ii)第二和第三抗原结合模块,它们各自是能够特异性结合cea的fab分子,包含包含与seqidno:23的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:27的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。在一个实施方案中,本发明提供t细胞活化性双特异性抗原结合分子,其包含:(i)第一抗原结合模块,它是能够特异性结合cd3的fab分子,包含seqidno:4的重链互补决定区(cdr)1,seqidno:5的重链cdr2,seqidno:6的重链cdr3,seqidno:8的轻链cdr1,seqidno:9的轻链cdr2和seqidno:10的轻链cdr3,其中所述第一抗原结合模块是交换fab分子,其中fab轻链和fab重链的可变或恒定区任一(特别是恒定区)是交换的;(ii)第二和第三抗原结合模块,它们各自是能够特异性结合mcsp的fab分子,包含seqidno:14的重链cdr1,seqidno:15的重链cdr2,seqidno:16的重链cdr3,seqidno:18的轻链cdr1,seqidno:19的轻链cdr2和seqidno:20的轻链cdr3。在一个实施方案中,本发明提供t细胞活化性双特异性抗原结合分子,其包含:(i)第一抗原结合模块,它是能够特异性结合cd3的fab分子,包含包含与seqidno:3的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:7的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区,其中所述第一抗原结合模块是交换fab分子,其中fab轻链和fab重链的可变或恒定区任一(特别是恒定区)是交换的;(ii)第二和第三抗原结合模块,它们各自是能够特异性结合mcsp的fab分子,包含包含与seqidno:13的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的重链可变区和包含与seqidno:17的氨基酸序列至少约95%,96%,97%,98%,99%或100%相同的氨基酸序列的轻链可变区。依照上述四个实施方案任一的t细胞活化性双特异性抗原结合分子可进一步包含(iii)由能够稳定联合的第一和第二亚基构成的fc域,其中所述第二抗原结合模块在fab重链的c端融合至所述第一抗原结合模块的fab重链的n端,且所述第一抗原结合模块在fab重链的c端融合至所述fc域的第一亚基的n端,且其中所述第三抗原结合模块在fab重链的c端融合至所述fc域的第二亚基的n端。在本发明的一些t细胞活化性双特异性抗原结合分子中,第一抗原结合模块的fab轻链和第二抗原结合模块的fab轻链彼此融合,任选地经由接头肽融合。根据第一和第二抗原结合模块的构造,第一抗原结合模块的fab轻链可在其c端融合至第二抗原结合模块的fab轻链的n端,或者第二抗原结合模块的fab轻链可在其c端融合至第一抗原结合模块的fab轻链的n端。第一和第二抗原结合模块的fab轻链的融合进一步降低不匹配的fab重链和轻链的错误配对,并且还降低表达本发明的一些t细胞活化性双特异性抗原结合分子需要的质粒数。在某些实施方案中,所述t细胞活化性双特异性抗原结合分子包含其中的第一抗原结合模块的fab轻链可变区与第一抗原结合模块的fab重链恒定区共享羧基末端肽键(即第一抗原结合模块包含交换fab重链,其中重链可变区用轻链可变区替换),所述第一抗原结合模块的fab重链恒定区继而与fc域亚基共享羧基末端肽键的多肽(vl(1)-ch1(1)-ch2-ch3(-ch4)),和其中的第二抗原结合模块的fab重链与fc域亚基共享羧基末端肽键的多肽(vh(2)-ch1(2)-ch2-ch3(-ch4))。在一些实施方案中,所述t细胞活化性双特异性抗原结合分子还包含其中的第一抗原结合模块的fab重链可变区与第一抗原结合模块的fab轻链恒定区共享羧基末端肽键的多肽(vh(1)-cl(1)),和第二抗原结合模块的fab轻链多肽(vl(2)-cl(2))。在某些实施方案中,所述多肽共价连接,例如通过二硫键进行。在备选的实施方案中,所述t细胞活化性双特异性抗原结合分子包含其中的第一抗原结合模块的fab重链可变区与第一抗原结合模块的fab轻链恒定区共享羧基末端肽键(即第一抗原结合模块包含交换fab重链,其中重链恒定区用轻链恒定区替换),所述第一抗原结合模块的fab轻链恒定区继而与fc域亚基共享羧基末端肽键的多肽(vh(1)-cl(1)-ch2-ch3(-ch4)),和其中的第二抗原结合模块的fab重链与fc域亚基共享羧基末端肽键的多肽(vh(2)-ch1(2)-ch2-ch3(-ch4))。在一些实施方案中,所述t细胞活化性双特异性抗原结合分子还包含其中的第一抗原结合模块的fab轻链可变区与第一抗原结合模块的fab重链恒定区共享羧基末端肽键的多肽(vh(1)-cl(1))和第二抗原结合模块的fab轻链多肽(vl(2)-cl(2))。在某些实施方案中,所述多肽共价连接,例如通过二硫键进行。在一些实施方案中,所述t细胞活化性双特异性抗原结合分子包含其中的第一抗原结合模块的fab轻链可变区与第一抗原结合模块的fab重链恒定区共享羧基末端肽键(即第一抗原结合模块包含交换fab重链,其中重链可变区用轻链可变区替换),所述第一抗原结合模块的fab重链恒定区继而与第二抗原结合模块的fab重链共享羧基末端肽键,所述第二抗原结合模块的fab重链继而与fc域亚基共享羧基末端肽键的多肽(vl(1)-ch1(1)-vh(2)-ch1(2)-ch2-ch3(-ch4))。在其他实施方案中,所述t细胞活化性双特异性抗原结合分子包含其中的第一抗原结合模块的fab重链可变区与第一抗原结合模块的fab轻链恒定区共享羧基末端肽键(即第一抗原结合模块包含交换fab重链,其中重链恒定区用轻链恒定区替换),所述第一抗原结合模块的fab轻链恒定区继而与第二抗原结合模块的fab重链共享羧基末端肽键,所述第二抗原结合模块的fab重链继而与fc域亚基共享羧基末端肽键的多肽(vh(1)-cl(1)-vh(2)-ch1(2)-ch2-ch3(-ch4))。仍在其他实施方案中,所述t细胞活化性双特异性抗原结合分子包含其中的第二抗原结合模块的fab重链与第一抗原结合模块的fab轻链可变区共享羧基末端肽键,所述第一抗原结合模块的fab轻链可变区继而与第一抗原结合模块的fab重链恒定区共享羧基末端肽键(即第一抗原结合模块包含交换fab重链,其中重链可变区用轻链可变区替换),所述第一抗原结合模块的fab重链恒定区继而与fc域亚基共享羧基末端肽键的多肽(vh(2)-ch1(2)-vl(1)-ch1(1)-ch2-ch3(-ch4))。在其他实施方案中,所述t细胞活化性双特异性抗原结合分子包含其中的第二抗原结合模块的fab重链与第一抗原结合模块的fab重链可变区共享羧基末端肽键,所述第一抗原结合模块的fab重链可变区继而与第一抗原结合模块的fab轻链恒定区共享羧基末端肽键(即第一抗原结合模块包含交换fab重链,其中重链恒定区用轻链恒定区替换),所述第一抗原结合模块的fab轻链恒定区继而与fc域亚基共享羧基末端肽键的多肽(vh(2)-ch1(2)-vh(1)-cl(1)-ch2-ch3(-ch4))。在这些中的一些实施方案中,所述t细胞活化性双特异性抗原结合分子还包含交换的第一抗原结合模块的fab轻链多肽,其中第一抗原结合模块的fab重链可变区与第一抗原结合模块的fab轻链恒定区共享羧基末端肽键(vh(1)-cl(1)),和第二抗原结合模块的fab轻链多肽(vl(2)-cl(2))。在这些中的其他实施方案中,所述t细胞活化性双特异性抗原结合分子还包含交换的fab轻链多肽,其中第一抗原结合模块的fab轻链可变区与第一抗原结合模块的fab重链恒定区共享羧基末端肽键(vl(1)-ch1(1)),和第二抗原结合模块的fab轻链多肽(vl(2)-cl(2))。仍在这些中的其他实施方案中,所述t细胞活化性双特异性抗原结合分子还包含其中的第一抗原结合模块的fab轻链可变区与第一抗原结合模块的fab重链恒定区共享羧基末端肽键,所述第一抗原结合模块的fab重链恒定区继而与第二抗原结合模块的fab轻链多肽共享羧基末端肽键的多肽(vl(1)-ch1(1)-vl(2)-cl(2)),其中的第一抗原结合模块的fab重链可变区与第一抗原结合模块的fab轻链恒定区共享羧基末端肽键,所述第一抗原结合模块的fab轻链恒定区继而与第二抗原结合模块的fab轻链多肽共享羧基末端肽键的多肽(vh(1)-cl(1)-vl(2)-cl(2)),其中的第二抗原结合模块的fab轻链多肽与第一抗原结合模块的fab轻链可变区共享羧基末端肽键,所述第一抗原结合模块的fab轻链可变区继而与第一抗原结合模块的fab重链恒定区共享羧基末端肽键的多肽(vl(2)-cl(2)-vl(1)-ch1(1)),或其中的第二抗原结合模块的fab轻链多肽与第一抗原结合模块的fab重链可变区共享羧基末端肽键,所述第一抗原结合模块的fab重链可变区继而与第一抗原结合模块的fab轻链恒定区共享羧基末端肽键的多肽(vl(2)-cl(2)-vh(1)-cl(1))。依照这些实施方案的t细胞活化性双特异性抗原结合分子还可以包含(i)fc域亚基多肽(ch2-ch3(-ch4)),或(ii)其中的第三抗原结合模块的fab重链与fc域亚基共享羧基末端肽键的多肽(vh(3)-ch1(3)-ch2-ch3(-ch4))和第三抗原结合模块的fab轻链多肽(vl(3)-cl(3))。在某些实施方案中,所述多肽共价连接,例如通过二硫键进行。依照任一个上文实施方案,t细胞活化性双特异性抗原结合分子的组分(例如抗原结合模块、fc域)可直接地或经由本文中描述或本领域已知的各种接头,特别是包含一个或多个氨基酸(通常约2-20个氨基酸)的肽接头融合。合适的、非免疫原性的肽接头包括例如(g4s)n、(sg4)n、(g4s)n或g4(sg4)n肽接头,其中n一般是介于1和10之间的数字,通常介于2和4之间。fc域t细胞活化性双特异性抗原结合分子的fc域由一对包含免疫球蛋白分子重链域的多肽链组成。例如,免疫球蛋白g(igg)分子的fc域是二聚体,其每个亚基包含ch2和ch3igg重链恒定域。fc域的两个亚基能够彼此稳定联合。在一个实施方案中,本发明的t细胞活化性双特异性抗原结合分子包含不超过一个fc域。在依照本发明的一个实施方案中,t细胞活化性双特异性抗原结合分子的fc域是iggfc域。在一个具体的实施方案中,fc域是igg1fc域。在另一个实施方案中,fc域是igg4fc域。在一个更特定的实施方案中,fc域是包含位置s228(依照kabat的eu编号方式)处的氨基酸替代,特别是氨基酸替代s228p的igg4fc域。此氨基酸替代降低igg4抗体的体内fab臂交换(参见stubenrauch等,drugmetabolismanddisposition38,84-91(2010))。在一个别的具体的实施方案中,fc域是人的。人igg1fc区的一个例示性序列在seqidno:107中给出。促进异二聚化的fc域修饰依照本发明的t细胞活化性双特异性抗原结合分子包含不同的抗原结合模块,其融合至fc域的两个亚基之一个或另一个,如此fc域的两个亚基通常包含在两条不相同的多肽链中。这些多肽的重组共表达和随后二聚化导致两种多肽的数种可能组合。为了改进重组生产中t细胞活化性双特异性抗原结合分子的产率和纯度,如此在t细胞活化性双特异性抗原结合分子的fc域中引入促进期望多肽联合的修饰将是有利的。因而,在具体的实施方案中,依照本发明的t细胞活化性双特异性抗原结合分子的fc域包含促进fc域的第一和第二亚基联合的修饰。人iggfc域的两个亚基之间最广泛的蛋白质-蛋白质相互作用的位点在fc域的ch3域中。如此,在一个实施方案中,所述修饰在fc域的ch3域中。在一个特定的实施方案中,所述修饰是所谓的“节-入-穴”修饰,其包含在fc域的两个亚基之一中的“节”修饰和在fc域的两个亚基的另一个中的“穴”修饰。节-入-穴技术记载于例如us5,731,168;us7,695,936;ridgway等,proteng9,617-621(1996)和carter,jimmunolmeth248,7-15(2001)。一般地,该方法牵涉在第一多肽的界面处引入隆起(“节”)并在第二多肽的界面中引入相应的空腔(“穴”),使得隆起可以置于空腔中从而促进异二聚体形成并阻碍同二聚体形成。通过将来自第一多肽界面的小氨基酸侧链用更大的侧链(例如酪氨酸或色氨酸)替换来构建隆起。在第二多肽的界面中创建具有与隆起相同或相似大小的互补性空腔,其通过将大氨基酸侧链用更小的氨基酸侧链(例如丙氨酸或苏氨酸)替换得到。因而,在一个具体的实施方案中,在t细胞活化性双特异性抗原结合分子fc域的第一亚基的ch3域中,氨基酸残基用具有更大侧链体积的氨基酸残基替换,由此在所述第一亚基的ch3域内生成隆起,其可安置于所述第二亚基的ch3域内的空腔中,而且在fc域的第二亚基的ch3域中,氨基酸残基用具有更小侧链体积的氨基酸残基替换,由此在所述第二亚基的ch3域内生成空腔,其中可安置所述第一亚基的ch3域内的隆起。可以通过改变编码多肽的核酸,例如通过位点专一诱变或通过肽合成来制备隆起和空腔。在一个特定的实施方案中,在fc域第一亚基的ch3域中,第366位的苏氨酸残基用色氨酸残基替换(t366w),而在fc域第二亚基的ch3域中,第407位的酪氨酸残基用缬氨酸残基替换(y407v)。在一个实施方案中,在fc域第二亚基中,另外,第366位的苏氨酸残基用丝氨酸残基替换(t366s)且第368位的亮氨酸残基用丙氨酸残基替换(l368a)。在又一个实施方案中,在fc域的第一亚基中,另外,第354位的丝氨酸残基用半胱氨酸残基替换(s354c),且在fc域的第二亚基中,另外,第349位的酪氨酸残基用半胱氨酸残基替换(y349c)。这两个半胱氨酸残基的引入导致在fc域的两个亚基之间形成二硫桥,进一步稳定了二聚体(carter,jimmunolmethods248,7-15(2001))。在一个具体的实施方案中,将能够结合cd3的抗原结合模块融合(任选地经由能够结合靶细胞抗原的抗原结合模块)至fc域的第一亚基(其包含“节”修饰)。不希望受理论束缚,能够结合cd3的抗原结合模块与fc域的含节的亚基的融合将(进一步)使包含两个能够结合cd3的抗原结合模块的抗原结合分子的生成最小化(两条含节的多肽的空间碰撞)。在一个备选的实施方案中,促进fc域的第一和第二亚基联合的修饰包含介导静电操纵效应(electrostaticsteeringeffect)的修饰,例如如记载于pct公开文本wo2009/089004的。一般地,此方法涉及将在两个fc域亚基界面处的一个或多个氨基酸残基替换为带电荷的氨基酸残基,从而在静电上不利于同二聚体形成而在静电上有利于异二聚化。降低fc受体结合和/或效应器功能的fc域修饰fc域赋予t细胞活化性双特异性抗原结合分子以有利的药动学特性,包括长血清半衰期,其有助于在靶组织中的较好积累和有利的组织-血液分配比。然而,同时它可能导致不想要的t细胞活化性双特异性抗原结合分子对表达fc受体的细胞而非优选的携带抗原的细胞的靶向。此外,fc受体信号传导途径的共激活可能导致细胞因子释放,其与抗原结合分子的t细胞活化性特性以及长半衰期组合,在系统性施用后引起细胞因子受体的过度激活和严重的副作用。(携带fc受体的)免疫细胞而非t细胞的活化甚至会降低t细胞活化性双特异性抗原结合分子的功效,原因是例如通过nk细胞对t细胞的潜在破坏。因而,在具体的实施方案中,相比于天然igg1fc域,依照本发明的t细胞活化性双特异性抗原结合分子的fc域展现出降低的对fc受体的结合亲和力和/或降低的效应器功能。在一个此类实施方案中,所述fc域(或包含所述fc域的t细胞活化性双特异性抗原结合分子)相比于天然igg1fc域(或包含天然igg1fc域的t细胞活化性双特异性抗原结合分子)展现出低于50%,优选低于20%,更优选低于10%且最优选低于5%的对fc受体的结合亲和力,和/或相比于天然igg1fc域(或包含天然igg1fc域的t细胞活化性双特异性抗原结合分子)展现出低于50%,优选低于20%,更优选低于10%且最优选低于5%的效应器功能。在一个实施方案中,所述fc域(或包含所述fc域的t细胞活化性双特异性抗原结合分子)不实质性结合fc受体和/或诱导效应器功能。在一个具体的实施方案中,所述fc受体是fcγ受体。在一个实施方案中,所述fc受体是人fc受体。在一个实施方案中,所述fc受体是活化性fc受体。在一个特定的实施方案中,所述fc受体是活化性人fcγ受体,更具体地为人fcγriiia、fcγri或fcγriia,最具体为人fcγriiia。在一个实施方案中,效应器功能是选自下组的一种或多种:cdc、adcc、adcp和细胞因子分泌。在一个具体的实施方案中,所述效应器功能是adcc。在一个实施方案中,相比于天然igg1fc域,所述fc域展现出基本类似的对新生儿fc受体(fcrn)的结合亲和力。当fc域(或包含所述fc域的t细胞活化性双特异性抗原结合分子)展现出超过约70%,具体为超过约80%,更具体超过约90%的天然igg1fc域(或包含天然igg1fc域的t细胞活化性双特异性抗原结合分子)对fcrn的结合亲和力时实现基本类似的对fcrn的结合。在某些实施方案中,相比于非工程化fc域,fc域工程化改造为具有降低的对fc受体的结合亲和力和/或降低的效应器功能。在具体的实施方案中,所述t细胞活化性双特异性抗原结合分子的fc域包含一处或多处降低fc域对fc受体的结合亲和力和/或效应器功能的氨基酸突变。通常,fc域的两个亚基的每一个中存在相同的一处或多处氨基酸突变。在一个实施方案中,所述氨基酸突变降低fc域对fc受体的结合亲和力。在一个实施方案中,所述氨基酸突变将fc域对fc受体的结合亲和力降低至少2倍、至少5倍、或至少10倍。在有超过一处降低fc域对fc受体的结合亲和力的氨基酸突变的实施方案中,这些氨基酸突变的组合可以将fc域对fc受体的结合亲和力降低至少10倍、至少20倍、或甚至至少50倍。在一个实施方案中,包含工程化fc域的t细胞活化性双特异性抗原结合分子相比于包含非工程化fc域的t细胞活化性双特异性抗原结合分子展现出低于20%,特别是低于10%,更特别地低于5%的对fc受体的结合亲和力。在一个具体的实施方案中,fc受体是fcγ受体。在一些实施方案中,所述fc受体是人fc受体。在一些实施方案中,fc受体是活化性fc受体。在一个特定的实施方案中,fc受体是活化性人fcγ受体,更特定地是人fcγriiia、fcγri或fcγriia,最特定地是人fcγriiia。优选地,对这些受体的每一种的结合是降低的。在一些实施方案中,对补体成分的结合亲和力,特别是对c1q的结合亲和力也是降低的。在一个实施方案中,对新生儿fc受体(fcrn)的结合亲和力没有降低。当fc域(或包含所述fc域的t细胞活化性双特异性抗原结合分子)展现出fc域的非工程化形式(或包含所述fc域的非工程化形式的t细胞活化性双特异性抗原结合分子)对fcrn的结合亲和力的超过约70%时实现基本类似的对fcrn的结合,即保留该fc域对所述受体的结合亲和力。fc域或包含所述fc域的本发明的t细胞活化性双特异性抗原结合分子可以展现出高于约80%和甚至高于约90%的此类亲和力。在某些实施方案中,相比于非工程化fc域,t细胞活化性双特异性抗原结合分子的fc域工程化改造为具有降低的效应器功能。所述降低的效应器功能可包括但不限于下列一种或多种:降低的补体依赖性细胞毒性(cdc)、降低的抗体依赖性细胞介导的细胞毒性(adcc)、降低的抗体依赖性细胞吞噬作用(adcp)、降低的细胞因子分泌、降低的免疫复合物介导的抗原呈递细胞的抗原摄取、降低的对nk细胞的结合、降低的对巨噬细胞的结合、降低的对单核细胞的结合、降低的对多形核细胞的结合、降低的诱导凋亡的直接信号传导、降低的靶物结合的抗体的交联、降低的树突细胞成熟或降低的t细胞启动。在一个实施方案中,所述降低的效应器功能是选自下组的一项或多项:降低的cdc、降低的adcc、降低的adcp和降低的细胞因子分泌。在一个具体的实施方案中,所述降低的效应器功能是降低的adcc。在一个实施方案中,所述降低的adcc小于由非工程化fc域(或包含非工程化fc域的t细胞活化性双特异性抗原结合分子)诱导的adcc的20%。在一个实施方案中,所述降低fc域对fc受体的结合亲和力和/或效应器功能的氨基酸突变是氨基酸替代。在一个实施方案中,fc域包含在选自下组的位置处的氨基酸替代:e233、l234、l235、n297、p331和p329。在一个更特定的实施方案中,fc域包含在选自下组的位置处的氨基酸替代:l234、l235和p329。在一些实施方案中,fc域包含氨基酸替代l234a和l235a。在一个此类实施方案中,fc域是igg1fc域,具体是人igg1fc域。在一个实施方案中,fc域包含在位置p329处的氨基酸替代。在一个更特定的实施方案中,氨基酸替代是p329a或p329g,特别是p329g。在一个实施方案中,fc域包含在位置p329处的氨基酸替代和另一个在选自以下位置处的氨基酸替代:e233、l234、l235、n297和p331。在一个更特定的实施方案中,另外的氨基酸替代是e233p、l234a、l235a、l235e、n297a、n297d或p331s。在具体的实施方案中,所述fc域包含在位置p329、l234和l235处的氨基酸替代。在更具体的实施方案中,fc域包含氨基酸突变l234a、l235a和p329g(“p329glala”)。在一个此类实施方案中,fc域是igg1fc域,具体是人igg1fc域。氨基酸替代的“p329glala”组合几乎完全消除了人igg1fc域的fcγ受体(以及补体)结合,如记载于pct公开文本no.wo2012/130831的,其通过提述完整并入本文。wo2012/130831还描述了制备此类突变体fc域的方法和用于测定其特性如fc受体结合或效应器功能的方法。相比于igg1抗体,igg4抗体展现出降低的对fc受体的结合亲和力和降低的效应器功能。因此,在一些实施方案中,本发明t细胞活化性双特异性抗原结合分子的fc域是igg4fc域,具体是人igg4fc域。在一个实施方案中,所述igg4fc域包含在位置s228处的氨基酸替代,具体为氨基酸替代s228p。为了进一步降低其对fc受体的结合亲和力和/或其效应器功能,在一个实施方案中,所述igg4fc域包含在位置l235处的氨基酸替代,具体为氨基酸替代l235e。在另一个实施方案中,所述igg4fc域包含在位置p329处的氨基酸替代,具体为氨基酸替代p329g。在一个具体的实施方案中,所述igg4fc域包含在位置s228、l235和p329处的氨基酸替代,具体为氨基酸替代s228p、l235e和p329g。此类igg4fc域突变体及其fcγ受体结合特性记载于pct公开文本no.wo2012/130831,其通过提述完整并入本文。在一个具体的实施方案中,相比于天然igg1fc域展现出降低的对fc受体的结合亲和力和/或降低的效应器功能的fc域是包含氨基酸替代l234a、l235a和任选地p329g的人igg1fc域,或包含氨基酸替代s228p、l235e和任选地p329g的人igg4fc域。在某些实施方案中,已消除fc域的n-糖基化。在一个此类实施方案中,所述fc域包含在位置n297处的氨基酸突变,特别是通过丙氨酸(n297a)或天冬氨酸(n297d)替换天冬酰胺的氨基酸替代。除了上文和pct公开文本no.wo2012/130831中描述的fc域外,具有降低的fc受体结合和/或效应器功能的fc域还包括那些具有fc域残基238、265、269、270、297、327和329中一个或多个的取代的fc域(美国专利no.6,737,056)。此类fc突变体包括具有在氨基酸位置265、269、270、297和327的两个或更多个处的取代的fc突变体,包括所谓的“dana”fc突变体,其具有残基265和297到丙氨酸的取代(美国专利no.7,332,581)。可以使用本领域中公知的遗传或化学方法通过氨基酸缺失、取代、插入或修饰来制备突变体fc区。遗传方法可以包括编码dna序列的位点专一性诱变、pcr、基因合成等。正确的核苷酸变化可以通过例如测序来验证。可以容易地测定对fc受体的结合,例如通过elisa或通过使用标准仪器如biacore仪(gehealthcare)的表面等离振子共振(spr)进行,并且fc受体诸如可通过重组表达获得。本文中描述了合适的此类结合测定法。或者,可使用已知表达特定fc受体的细胞系,如表达fcγiiia受体的人nk细胞来估测fc域或包含fc域的细胞活化性双特异性抗原结合分子对fc受体的结合亲和力。可通过本领域中已知的方法来测量fc域或包含fc域的t细胞活化性双特异性抗原结合分子的效应器功能。本文中描述了用于测量adcc的合适的测定法。评估感兴趣分子的adcc活性的体外测定法的其他例子记载于美国专利no.5,500,362;hellstrom等,procnatlacadsciusa83,7059-7063(1986)和hellstrom等,procnatlacadsciusa82,1499-1502(1985);美国专利no.5,821,337;bruggemann等,jexpmed166,1351-1361(1987)。或者,可采用非放射性测定方法(参见例如用于流式细胞术的actitm非放射性细胞毒性测定(celltechnology,inc.mountainview,ca);和cytotox非放射性细胞毒性测定(promega,madison,wi))。对于此类测定法有用的效应细胞包括外周血单个核细胞(pbmc)和天然杀伤(nk)细胞。或者/另外,可体内评估感兴趣分子的adcc活性,例如在动物模型中,如披露于clynes等,procnatlacadsciusa95,652-656(1998)的。在一些实施方案中,fc域对补体成分,特别是对c1q的结合是降低的。因而,在其中fc域工程化为具有降低的效应器功能的一些实施方案中,所述降低的效应器功能包括降低的cdc。可实施c1q结合测定法来测定t细胞活化性双特异性抗原结合分子是否能结合c1q并因此具有cdc活性。参见例如wo2006/029879和wo2005/100402中的c1q和c3c结合elisa。为了评估补体激活,可实施cdc测定法(参见例如gazzano-santoro等,jimmunolmethods202,163(1996);cragg等,blood101,1045-1052(2003);以及cragg和glennie,blood103,2738-2743(2004))。抗原结合模块本发明的抗原结合分子是双特异性的,即它包含至少两种能够特异性结合两种不同抗原性决定簇的抗原结合模块。依照本发明,所述抗原结合模块是fab分子(即由各自包含可变区和恒定区的重链和轻链构成的抗原结合域)。在一个实施方案中,所述fab分子是人的。在另一个实施方案中,所述fab分子是人源化的。在再一个实施方案中,所述fab分子包含人重链和轻链恒定区。至少一个抗原结合模块是交换fab分子。此类修饰防止来自不同fab分子的重链和轻链的错误配对,由此改进了重组生产中本发明的t细胞活化性双特异性抗原结合分子的产率和纯度。在可用于本发明的t细胞活化性双特异性抗原结合分子的具体的交换fab分子中,fab轻链和fab重链的恒定区是交换的。在可用于本发明t细胞活化性双特异性抗原结合分子的另一种交换fab分子中,fab轻链和fab重链的可变区是交换的。在依照本发明的一个具体的实施方案中,所述t细胞活化性双特异性抗原结合分子能够同时结合靶细胞抗原(特别是肿瘤细胞抗原)和cd3。在一个实施方案中,所述t细胞活化性双特异性抗原结合分子能够通过同时结合靶细胞抗原和cd3来交联t细胞和靶细胞。在一个甚至更具体的实施方案中,此类同时结合导致靶细胞特别是肿瘤细胞的裂解。在一个实施方案中,此类同时结合导致t细胞的活化。在其他实施方案中,此类同时结合导致t淋巴细胞,特别是细胞毒性t淋巴细胞的细胞应答,其选自下组:增殖、分化、细胞因子分泌、细胞毒性效应分子释放、细胞毒性活性和活化标志物的表达。在一个实施方案中,t细胞活化性双特异性抗原结合分子对cd3的结合而不同时结合靶细胞抗原不导致t细胞活化。在一个实施方案中,t细胞活化性双特异性抗原结合分子能够将t细胞的细胞毒性活性重定向于靶细胞。在一个具体的实施方案中,所述重定向不依赖于靶细胞的mhc介导的肽抗原呈递和/或t细胞的特异性。具体地,依照本发明任一实施方案的t细胞是细胞毒性t细胞。在一些实施方案中,所述t细胞是cd4+或cd8+t细胞,具体为cd8+t细胞。cd3结合模块本发明的t细胞活化性双特异性抗原结合分子包含至少一个能够结合cd3的抗原结合模块(本文中亦称为“cd3抗原结合模块”或“第一抗原结合模块”)。在一个具体的实施方案中,所述t细胞活化性双特异性抗原结合分子包含不超过一个能够特异性结合cd3的抗原结合模块。在一个实施方案中,所述t细胞活化性双特异性抗原结合分子提供对cd3的单价结合。所述cd3抗原结合模块是交换fab分子,即其中fab重和轻链的可变或恒定区任一是交换的fab分子。在其中有t细胞活化性双特异性抗原结合分子中包含的超过一个能够特异性结合靶细胞抗原的抗原结合模块的实施方案中,能够特异性结合cd3的抗原结合模块优选为交换fab分子且能够特异性结合靶细胞抗原的抗原结合模块是常规fab分子。在一个具体的实施方案中,cd3是人cd3(seqidno:103)或猕猴cd3(seqidno:104),最特别是人cd3。在一个具体的实施方案中,cd3抗原结合模块对于人和猕猴cd3是交叉反应性的(即特异性结合)。在一些实施方案中,所述第一抗原结合模块能够特异性结合cd3的ε亚基。cd3抗原结合模块包含至少一个选自seqidno:4,seqidno:5和seqidno:6的重链互补决定区(cdr)和至少一个选自seqidno:8,seqidno:9和seqidno:10的轻链cdr。在一个实施方案中,所述cd3抗原结合模块包含seqidno:4的重链cdr1、seqidno:5的重链cdr2、seqidno:6的重链cdr3、seqidno:8的轻链cdr1、seqidno:9的轻链cdr2和seqidno:10的轻链cdr3。在一个实施方案中,所述cd3抗原结合模块包含与选自seqidno:3,seqidno:32和seqidno:33的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的重链可变区序列和与选自seqidno:7和seqidno:31的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的轻链可变区序列。在一个实施方案中,所述cd3抗原结合模块包含包含选自seqidno:3,seqidno:32和seqidno:33的氨基酸序列的重链可变区和包含选自seqidno:7和seqidno:31的氨基酸序列的轻链可变区。在一个实施方案中,所述cd3抗原结合模块包含与seqidno:3至少约95%、96%、97%、98%、99%或100%相同的重链可变区序列和与seqidno:7至少约95%、96%、97%、98%、99%或100%相同的轻链可变区序列。在一个实施方案中,所述cd3抗原结合模块包含包含seqidno:3的氨基酸序列的重链可变区和包含seqidno:7的氨基酸序列的轻链可变区。在一个实施方案中,所述cd3抗原结合模块包含seqidno:3的重链可变区序列和seqidno:7的轻链可变区序列。靶细胞抗原结合模块本发明的t细胞活化性双特异性抗原结合分子包含至少一个能够结合靶细胞抗原的抗原结合模块(本文中亦称为“靶细胞抗原结合模块”或“第二”和“第三”抗原结合模块)。在某些实施方案中,所述t细胞活化性双特异性抗原结合分子包含两个能够结合靶细胞抗原的抗原结合模块。在一个具体的此类实施方案中,这些抗原结合模块中的每一个特异性结合相同抗原性决定簇。在一个甚至更加具体的实施方案中,所有这些抗原结合模块是相同的。在一个实施方案中,所述t细胞活化性双特异性抗原结合分子包含能够特异性结合靶细胞抗原的免疫球蛋白分子。在一个实施方案中,所述t细胞活化性双特异性抗原结合分子包含不超过两个能够结合靶细胞抗原的抗原结合模块。所述靶细胞抗原结合模块通常是结合特定的抗原性决定簇且能够将t细胞活化性双特异性抗原结合分子引导至靶部位,例如携带抗原性决定簇的特定类型肿瘤细胞的fab分子,特别是常规fab分子。在某些实施方案中,所述靶细胞抗原结合模块特异性结合细胞表面抗原。在一个具体的实施方案中,所述靶细胞抗原结合模块特异性结合细胞表面抗原的近膜区。在一个特定的此类实施方案中,所述细胞表面抗原是癌胚抗原(cea)且所述近膜区是cea的b3域(seqidno:119的残基208-286)。在另一个特定的此类实施方案中,所述细胞表面抗原是黑素瘤相关硫酸软骨素蛋白聚糖(mcsp)且所述近膜区是mcsp(seqidno:118)的d3域。在某些实施方案中,所述靶细胞抗原结合模块针对与病理学状况有关的抗原,如肿瘤细胞或病毒感染细胞上呈现的抗原。合适的抗原是细胞表面抗原,例如但不限于细胞表面受体。在具体的实施方案中,所述抗原是人抗原。在一个特定的实施方案中,所述靶细胞抗原选自黑素瘤相关硫酸软骨素蛋白聚糖(mcsp,cspg4)和癌胚抗原(cea,ceacam5)。在一些实施方案中,所述t细胞活化性双特异性抗原结合分子包含至少一个特异于黑素瘤关联硫酸软骨素蛋白聚糖(mcsp)的抗原结合模块。在一个实施方案中,所述特异于mcsp的抗原结合模块包含至少一个选自seqidno:14,seqidno:15,seqidno:16,seqidno:35,seqidno:37,seqidno:38和seqidno:40的重链互补决定区(cdr)和至少一个选自seqidno:18,seqidno:19,seqidno:20,seqidno:44,seqidno:45,seqidno:48,seqidno:49和seqidno:50的轻链cdr。在一个实施方案中,所述特异于mcsp的抗原结合模块包含至少一个选自seqidno:14,seqidno:15和seqidno:16的重链互补决定区(cdr)和至少一个选自seqidno:18,seqidno:19和seqidno:20的轻链cdr。在一个实施方案中,所述特异于mcsp的抗原结合模块包含seqidno:14的重链cdr1、seqidno:15的重链cdr2、seqidno:16的重链cdr3、seqidno:18的轻链cdr1、seqidno:19的轻链cdr2和seqidno:20的轻链cdr3。在一个别的实施方案中,所述特异于mcsp的抗原结合模块包含与选自seqidno:13,seqidno:34,seqidno:36,seqidno:39和seqidno:41的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的重链可变区序列和与选自seqidno:17,seqidno:43,seqidno:46,seqidno:47和seqidno:51的氨基酸序列至少约95%、96%、97%、98%、99%或100%相同的轻链可变区序列。在一个别的实施方案中,所述特异于mcsp的抗原结合模块包含包含选自seqidno:13,seqidno:34,seqidno:36,seqidno:39和seqidno:41的氨基酸序列的重链可变区和包含选自seqidno:17,seqidno:43,seqidno:46,seqidno:47和seqidno:51的氨基酸序列的轻链可变区。在一个别的实施方案中,所述特异于mcsp的抗原结合模块包含与seqidno:13至少约95%、96%、97%、98%、99%或100%相同的重链可变区序列和与seqidno:17至少约95%、96%、97%、98%、99%或100%相同的轻链可变区序列,或其保留功能性的变体。在一个实施方案中,所述特异于mcsp的抗原结合模块包含包含seqidno:13的氨基酸序列的重链可变区和包含seqidno:17的氨基酸序列的轻链可变区。在一个实施方案中,所述特异于mcsp的抗原结合模块包含seqidno:13的重链可变区序列和seqidno:17的轻链可变区序列。在一个实施方案中,所述t细胞活化性双特异性抗原结合分子包含与seqidno:12至少约95%、96%、97%、98%、99%或100%相同的多肽序列、与seqidno:53至少约95%、96%、97%、98%、99%或100%相同的多肽序列、与seqidno:54至少约95%、96%、97%、98%、99%或100%相同的多肽序列、和与seqidno:55至少约95%、96%、97%、98%、99%或100%相同的多肽序列。在具体的实施方案中,所述t细胞活化性双特异性抗原结合分子包含至少一个特异于癌胚抗原(cea)的抗原结合模块。在一个实施方案中,所述特异于cea的抗原结合模块包含至少一个选自seqidno:24,seqidno:25和seqidno:26的重链互补决定区(cdr)和至少一个选自seqidno:28,seqidno:29和seqidno:30的轻链cdr。在一个实施方案中,所述特异于cea的抗原结合模块包含seqidno:24的重链cdr1、seqidno:25的重链cdr2、seqidno:26的重链cdr3、seqidno:28的轻链cdr1、seqidno:29的轻链cdr2和seqidno:30的轻链cdr3。在一个别的实施方案中,所述特异于cea的抗原结合模块包含与seqidno:23至少约95%、96%、97%、98%、99%或100%相同的重链可变区序列和与seqidno:27至少约95%、96%、97%、98%、99%或100%相同的轻链可变区序列,或其保留功能性的变体。在一个实施方案中,所述特异于cea的抗原结合模块包含包含seqidno:23的氨基酸序列的重链可变区和包含seqidno:27的氨基酸序列的轻链可变区。在一个实施方案中,所述特异于cea的抗原结合模块包含seqidno:23的重链可变区序列和seqidno:27的轻链可变区序列。在一个实施方案中,所述t细胞活化性双特异性抗原结合分子包含与seqidno:22至少约95%、96%、97%、98%、99%或100%相同的多肽序列、与seqidno:56至少约95%、96%、97%、98%、99%或100%相同的多肽序列、与seqidno:57至少约95%、96%、97%、98%、99%或100%相同的多肽序列、和与seqidno:58至少约95%、96%、97%、98%、99%或100%相同的多肽序列。多核苷酸本发明还提供分离的多核苷酸,其编码如本文中描述的t细胞活化性双特异性抗原结合分子或其片段。在一些实施方案中,所述片段是抗原结合片段。本发明的多核苷酸包括那些与以下所列序列至少约80%、85%、90%、95%、96%、97%、98%、99%或100%相同的多核苷酸:seqidnos63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97和98,包括其功能性片段或变体。可以将编码本发明的t细胞活化性双特异性抗原结合分子的多核苷酸以编码完整t细胞活化性双特异性抗原结合分子的单一多核苷酸表达,或以共表达的多个(例如两个或更多个)多核苷酸表达。由共表达的多核苷酸编码的多肽可以经由例如二硫键或其它手段联合以形成功能性t细胞活化性双特异性抗原结合分子。例如,抗原结合模块的轻链部分可与t细胞活化性双特异性抗原结合分子中包含抗原结合模块重链部分、fc域亚基和任选地(部分的)另一个抗原结合模块的部分由分开的多核苷酸编码。当共表达时,重链多肽将与轻链多肽联合以形成抗原结合模块。在另一个例子中,t细胞活化性双特异性抗原结合分子中包含两个fc域亚基之一和任选地(部分的)一个或多个抗原结合模块的部分可与由t细胞活化性双特异性抗原结合分子中包含两个fc域亚基中另一个和任选地(部分的)抗原结合模块的部分由分开的多核苷酸编码。当共表达时,所述fc域亚基将联合以形成fc域。在一些实施方案中,所述分离的多核苷酸编码完整的依照本文所述发明的t细胞活化性双特异性抗原结合分子。在其它实施方案中,所述分离的多核苷酸编码依照本文所述发明的t细胞活化性双特异性抗原结合分子中包含的多肽。在另一个实施方案中,本发明涉及编码本发明的t细胞活化性双特异性抗原结合分子或其片段的分离的多核苷酸,其中所述多核苷酸包含编码如以下显示的可变区序列的序列:seqidno3,7,13,17,23,27,31,32,33,34,36,39,41,43,46,47或51。在另一个实施方案中,本发明涉及编码t细胞活化性双特异性抗原结合分子或其片段的分离的多核苷酸,其中所述多核苷酸包含编码如以下显示的多肽序列的序列:seqidno22,56,57,58,12,53,54和55。在另一个实施方案中,本发明进一步涉及编码本发明t细胞活化性双特异性抗原结合分子或其片段的分离的多核苷酸,其中所述多核苷酸包含与以下显示的核苷酸序列至少约80%,85%,90%,95%,96%,97%,98%或99%相同的序列:seqidno63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97或98。在另一个实施方案中,本发明涉及编码本发明t细胞活化性双特异性抗原结合分子或其片段的分离的多核苷酸,其中所述多核苷酸包含以下显示的核酸序列:seqidno63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97或98。在另一个实施方案中,本发明涉及编码本发明t细胞活化性双特异性抗原结合分子或其片段的分离的多核苷酸,其中所述多核苷酸包含编码与以下的氨基酸序列至少约80%,85%,90%,95%,96%,97%,98%或99%相同的可变区序列的序列:seqidno3,7,13,17,23,27,31,32,33,34,36,39,41,43,46,47或51。在另一个实施方案中,本发明涉及编码t细胞活化性双特异性抗原结合分子或其片段的分离的多核苷酸,其中所述多核苷酸包含编码与以下的氨基酸序列至少约80%,85%,90%,95%,96%,97%,98%或99%相同的多肽序列的序列:seqidno22,56,57,58,12,53,54或55。本发明涵盖编码本发明t细胞活化性双特异性抗原结合分子或其片段的分离的多核苷酸,其中所述多核苷酸包含编码seqidno3,7,13,17,23,27,31,32,33,34,36,39,41,43,46,47或51的可变区序列及保守性氨基酸替代的序列。本发明还涵盖编码本发明t细胞活化性双特异性抗原结合分子或其片段的分离的多核苷酸,其中所述多核苷酸包含编码以下多肽序列及保守性氨基酸替代的序列:seqidno22,56,57,58,12,53,54或55。在某些实施方案中,所述多核苷酸或核酸是dna。在其它实施方案中,本发明的多核苷酸是rna,例如以信使rna(mrna)的形式。本发明的rna可以是单链或双链的。重组方法可以获得本发明的t细胞活化性双特异性抗原结合分子,例如通过固相肽合成(例如merrifield固相合成)或重组生成进行。对于重组生成,分离一种或多种编码所述t细胞活化性双特异性抗原结合分子(片段)的多核苷酸(例如如上文描述的),并将其插入一种或多种载体中用于在宿主细胞中进一步克隆和/或表达。可以使用常规规程容易分离并测序此类多核苷酸。在一个实施方案中,提供包含一种或多种本发明的多核苷酸的载体(优选为表达载体)。可以使用本领域技术人员公知的方法来构建含有t细胞活化性双特异性抗原结合分子(片段)的编码序列以及适宜的转录/翻译控制信号的表达载体。这些方法包括体外重组dna技术、合成技术和体内重组/遗传重组。参见例如记载于maniatis等,molecularcloning:alaboratorymanual,coldspringharborlaboratory,n.y.(1989);和ausubel等,currentprotocolsinmolecularbiology,greenepublishingassociatesandwileyinterscience,n.y(1989)的技术。表达载体可以是质粒、病毒的一部分或可以是核酸片段。表达载体包含表达盒,其中在与启动子和/或其它转录或翻译控制元件的可操作联合中克隆编码t细胞活化性双特异性抗原结合分子(片段)的多核苷酸(即编码区)。如本文中使用的,“编码区”是核酸中由翻译成氨基酸的密码子组成的一部分。尽管“终止密码子”(tag、tga或taa)不翻译成氨基酸,但可将其视为编码区的一部分(若存在的话),但任何侧翼序列例如启动子、核糖体结合位点、转录终止子、内含子、5’和3’非翻译区等,不是编码区的一部分。两个或更多个编码区可以存在于单一多核苷酸构建体中(例如单一载体上),或存在于分开的多核苷酸构建体中,例如在分开的(不同的)载体上。此外,任何载体可含有单个编码区,或可包含两个或更多个编码区,例如本发明的载体可以编码一种或多种多肽,其经由蛋白水解切割在翻译后或共翻译分开成最终蛋白质。另外,本发明的载体、多核苷酸或核酸可以编码异源编码区,其与编码本发明的t细胞活化性双特异性抗原结合分子(片段)或其变体或衍生物的多核苷酸融合或未融合。异源编码区包括但不限于特殊化的元件或基序,如分泌信号肽或异源功能域。当基因产物例如多肽的编码区与一种或多种调节序列以某种方式联合从而使得该基因产物的表达置于该调节序列的影响或控制下时,即为可操作联合。若诱导启动子功能导致编码期望的基因产物的mrna的转录并且如果两个dna片段之间的连接的性质不干扰表达调节序列指导该基因产物表达的能力或不干扰dna模板被转录的能力,则两个dna片段(如多肽编码区和与其联合的启动子)为“可操作联合的”。如此,如果启动子能够实现编码多肽的核酸的转录,那么该启动子区将是与该核酸可操作联合。所述启动子可以是细胞特异性启动子,其仅在预先确定的细胞中指导dna的实质性转录。除启动子以外,其它转录控制元件例如增强子、操纵基因、阻遏物和转录终止信号能与多核苷酸可操作联合以指导细胞特异性转录。在本文中公开了合适的启动子和其它转录控制区。多种转录控制区是本领域技术人员已知的。这些包括但不限于在脊椎动物细胞中发挥功能的转录控制区,如但不限于来自巨细胞病毒的启动子和增强子区段(例如立即早期启动子,以及内含子-a)、猿病毒40(例如早期启动子)和逆转录病毒(如例如劳斯(rous)肉瘤病毒)。其它转录控制区包括那些自脊椎动物基因如肌动蛋白、热休克蛋白、牛生长激素和家兔β球蛋白衍生的,以及能够控制真核细胞中基因表达的其它序列。另外的合适的转录控制区包括组织特异性启动子和增强子以及诱导型启动子(例如四环素诱导型启动子)。类似地,多种翻译控制元件是本领域普通技术人员已知的。这些包括但不限于核糖体结合位点、翻译起始和终止密码子以及自病毒系统衍生的元件(具体地,内部核糖体进入位点或ires,亦称为cite序列)。表达盒还可以包含其它特征,如复制起点和/或染色体整合元件,如逆转录病毒长末端重复(ltr)或腺伴随病毒(aav)反向末端重复(itr)。本发明的多核苷酸和核酸编码区可以与编码分泌或信号肽的另外的编码区联合,所述分泌或信号肽指导由本发明的多核苷酸编码的多肽的分泌。例如,如果期望分泌所述t细胞活化性双特异性抗原结合分子,那么可以将编码信号序列的dna置于编码本发明t细胞活化性双特异性抗原结合分子或其片段的核酸上游。依照信号假说,由哺乳动物细胞分泌的蛋白质具有信号肽或分泌前导序列,一旦启动将生长的蛋白质链跨越粗面内质网输出,就将该信号肽或分泌前导序列从成熟的蛋白质切去。本领域中普通技术人员知晓由脊椎动物细胞分泌的多肽一般具有融合至多肽n端的信号肽,其从所翻译的多肽切去以生成分泌性或“成熟”形式的多肽。在某些实施方案中,使用天然的信号肽,例如免疫球蛋白重链或轻链信号肽,或该序列的保留指导与其可操作联合的多肽分泌的能力的功能性衍生物。或者,可以使用异源哺乳动物信号肽或其功能性衍生物。例如,可以将野生型前导序列用人组织血纤维蛋白溶酶原激活剂(tpa)或小鼠β-葡糖醛酸糖苷酶的前导序列取代。分泌性信号肽的例示性氨基酸和多核苷酸序列在seqidno108-116给出。可以将编码能用于促进后期纯化(例如组氨酸标签)或辅助标记t细胞活化性双特异性抗原结合分子的短蛋白序列的dna纳入t细胞活化性双特异性抗原结合分子(片段)编码多核苷酸内或其末端。在一个别的实施方案中,提供包含本发明的一种或多种多核苷酸的宿主细胞。在某些实施方案中,提供包含本发明的一种或多种载体的宿主细胞。多核苷酸和载体可以单独地或组合地掺入本文中分别关于多核苷酸和载体所描述的任何特征。在一个此类实施方案中,宿主细胞包含载体(例如已用该载体转化或转染),所述载体包含编码本发明t细胞活化性双特异性抗原结合分子(的部分)的多核苷酸。如本文中使用的,术语“宿主细胞”指任何能工程化以生成本发明的t细胞活化性双特异性抗原结合分子或其片段的细胞系统种类。适用于复制并支持t细胞活化性双特异性抗原结合分子表达的宿主细胞是本领域中公知的。在适当时,可用特定的表达载体转染或转导此类细胞,并且可以培养大量的含载体细胞以用于接种大规模发酵罐,从而获得充足量的t细胞活化性双特异性抗原结合分子用于临床应用。合适的宿主细胞包括原核微生物如大肠杆菌,或各种真核细胞,如中国仓鼠卵巢细胞(cho)、昆虫细胞等。例如,可以在细菌中生成多肽,尤其在不需要糖基化时。在表达后,可以将多肽在可溶性级分中从细菌细胞糊分离并可以进一步纯化。除了原核生物外,真核微生物如丝状真菌或酵母也是适合编码多肽的载体的克隆或表达宿主,包括其糖基化途径已被“人源化”,导致生成具有部分或完全人的糖基化模式的多肽的真菌和酵母菌株。参见gerngross,natbiotech22,1409-1414(2004),和li等,natbiotech24,210-215(2006)。适用于表达(糖基化)多肽的宿主细胞还自多细胞生物体(无脊椎动物和脊椎动物)衍生。无脊椎动物细胞的例子包括植物和昆虫细胞。已鉴定出可与昆虫细胞一起使用的大量杆状病毒株,特别是用于转染草地贪夜蛾(spodopterafrugiperda)细胞。也可以将植物细胞培养物用作宿主。参见例如美国专利no.5,959,177,6,040,498,6,420,548,7,125,978和6,417,429(描述用于在转基因植物中生成抗体的plantibodiestm技术)。脊椎动物细胞也可以用作宿主。例如,适应于在悬液中生长的哺乳动物细胞系可以是有用的。可用的哺乳动物宿主细胞系的其它例子是由sv40转化的猴肾cv1系(cos-7);人胚肾系(293或293t细胞,如例如记载于graham等,jgenvirol36,59(1977))、幼仓鼠肾细胞(bhk)、小鼠塞托利(sertoli)细胞(tm4细胞,如例如记载于mather,biolreprod23,243-251(1980)的)、猴肾细胞(cv1)、非洲绿猴肾细胞(vero-76)、人宫颈癌细胞(hela)、猕猴肾细胞(mdck),buffalo大鼠肝细胞(brl3a)、人肺细胞(w138)、人肝细胞(hepg2)、小鼠乳房肿瘤细胞(mmt060562)、tri细胞(如例如记载于mather等,annalsn.y.acadsci383,44-68(1982)的)、mrc5细胞和fs4细胞。其它可用的哺乳动物宿主细胞系包括中国仓鼠卵巢(cho)细胞,包括dhfr-cho细胞(urlaub等,procnatlacadsciusa77,4216(1980));和骨髓瘤细胞系如yo、ns0、p3x63和sp2/0。对于某些适用于蛋白质生产的哺乳动物宿主细胞系的综述,参见例如yazaki和wu,methodsinmolecularbiology,第248卷(b.k.c.lo编,humanapress,totowa,nj),pp.255-268(2003)。宿主细胞包括培养的细胞,例如哺乳动物培养细胞、酵母细胞、昆虫细胞、细菌细胞和植物细胞等,但还包括在转基因动物、转基因植物或培养的植物或动物组织中包含的细胞。在一个实施方案中,宿主细胞是真核细胞,优选为哺乳动物细胞如中国仓鼠卵巢(cho)细胞、人胚肾(hek)细胞或淋巴样细胞(例如y0、ns0、sp20细胞)。本领域中已知在这些系统中表达外来基因的标准技术。可以将表达包含抗原结合域如抗体的重链或轻链的多肽的细胞工程化改造为使得还表达另一抗体链,从而使得表达的产物是具有重链和轻链两者的抗体。在一个实施方案中,提供了生成依照本发明的t细胞活化性双特异性抗原结合分子的方法,其中所述方法包括在适合于所述t细胞活化性双特异性抗原结合分子表达的条件下培养包含编码t细胞活化性双特异性抗原结合分子的多核苷酸的宿主细胞(如本文中提供的),并从宿主细胞(或宿主细胞培养基)回收所述t细胞活化性双特异性抗原结合分子。所述t细胞活化性双特异性抗原结合分子的组分彼此遗传融合。t细胞活化性双特异性抗原结合分子可设计为使其组分直接彼此融合或经由接头序列间接融合。可依照本领域中公知的方法测定接头的组成和长度,并可以测试功效。t细胞活化性双特异性抗原结合分子不同组分之间的接头序列的例子见于本文中提供的序列。还包含另外的序列以纳入切割位点来分开融合的各个组分(若期望的话),例如内肽酶识别序列。在某些实施方案中,所述t细胞活化性双特异性抗原结合分子的一个或多个抗原结合模块至少包含能结合抗原性决定簇的抗体可变区。可变区可形成天然或非天然存在的抗体及其片段的一部分和自其衍生。生成多克隆抗体和单克隆抗体的方法是本领域中公知的(参见例如harlow和lane,“antibodies,alaboratorymanual”,coldspringharborlaboratory,1988)。非天然存在的抗体可以使用固相肽合成构建生成,可以重组生成(例如如记载于美国专利no.4,186,567的)或可通过例如筛选包含可变重链和可变轻链的组合库获得(参见例如mccafferty的美国专利no.5,969,108)。可以将抗体、抗体片段、抗原结合域或可变区的任何动物种类用于本发明的t细胞活化性双特异性抗原结合分子。可用于本发明的非限制性抗体、抗体片段、抗原结合域或可变区可以是鼠、灵长类或人起源的。如果t细胞活化性双特异性抗原结合分子意图供人使用,那么可以使用嵌合形式的抗体,其中抗体的恒定区来自人。也可以依照本领域中公知的方法制备人源化或全人形式的抗体(参见例如winter的美国专利no.5,565,332)。人源化可以通过各种方法实现,包括但不限于(a)将非人(例如供体抗体)cdr嫁接到人(例如受体抗体)框架和恒定区上,保留或不保留关键的框架残基(例如那些对于保留较好的抗原结合亲和力或抗体功能重要的残基),(b)仅将非人特异性决定区(sdr或a-cdr;对于抗体-抗原相互作用关键的残基)嫁接到人框架和恒定区上,或(c)移植完整的非人可变域,但通过替换表面残基用人样部分来“掩饰(cloak)”它们。人源化的抗体及其制备方法综述于例如almagro和fransson,frontbiosci13,1619-1633(2008),并且还记载于例如riechmann等,nature332,323-329(1988);queen等,procnatlacadsciusa86,10029-10033(1989);美国专利no.5,821,337,7,527,791,6,982,321和7,087,409;jones等,nature321,522-525(1986);morrison等,procnatlacadsci81,6851-6855(1984);morrison和oi,advimmunol44,65-92(1988);verhoeyen等,science239,1534-1536(1988);padlan,molecimmun31(3),169-217(1994);kashmiri等,methods36,25-34(2005)(描述sdr(a-cdr)嫁接);padlan,molimmunol28,489-498(1991)(描述“表面重建”);dall’acqua等,methods36,43-60(2005)(描述“fr改组”);和osbourn等,methods36,61-68(2005)以及klimka等,brjcancer83,252-260(2000)(描述了fr改组的“引导选择”办法)。可以使用本领域中已知的各种技术来生成人抗体和人可变区。人抗体一般记载于vandijk和vandewinkel,curropinpharmacol5,368-74(2001)以及lonberg,curropinimmunol20,450-459(2008)。人可变区能形成通过杂交瘤方法制备的人单克隆抗体的一部分和自其衍生(参见例如monoclonalantibodyproductiontechniquesandapplications,pp.51-63(marceldekker,inc.,newyork,1987))。还可以通过对转基因动物施用免疫原来制备人抗体和人可变区,所述转基因动物已经过修饰以应答抗原激发而生成完整的人抗体或具有人可变区的完整抗体(参见例如lonberg,natbiotech23,1117-1125(2005)。还可以通过分离选自人衍生的噬菌体展示库的fv克隆可变区序列来生成人抗体和人可变区(参见例如hoogenboom等,于methodsinmolecularbiology178,1-37(o’brien等编,humanpress,totowa,nj,2001);和mccafferty等,nature348,552-554;clackson等,nature352,624-628(1991))。噬菌体通常展示抗体片段,作为单链fv(scfv)片段或作为fab片段。在某些实施方案中,将可用于本发明的抗原结合模块工程化改造为具有增强的结合亲和力,其依照例如公开于美国专利申请公开文本no.2004/0132066的方法进行,其完整内容通过提述据此并入。可以经由酶联免疫吸附测定法(elisa)或本领域技术人员熟知的其它技术来测量本发明的t细胞活化性双特异性抗原结合分子结合特定抗原性决定簇的能力,所述其它技术例如表面等离振子共振技术(在biacoret100系统上分析)(liljeblad等,glycoj17,323-329(2000))和传统的结合测定法(heeley,endocrres28,217-229(2002))。可以使用竞争测定法来鉴定与参照抗体竞争对特定抗原的结合的抗体、抗体片段、抗原结合域或可变域,例如与v9抗体竞争对cd3的结合的抗体。在某些实施方案中,此类竞争性抗体结合由参照抗体结合的相同表位(例如线性或构象性表位)。用于定位抗体结合的表位的详细的例示性方法在morris(1996)“epitopemappingprotocols,”于methodsinmolecularbiologyvol.66(humanapress,totowa,nj)中提供。在一种例示性竞争测定法中,将固定化的抗原(例如cd3)在溶液中温育,所述溶液包含结合该抗原的第一标记抗体(例如v9抗体,记载于us6,054,297)和测试其与第一抗体竞争对抗原的结合的能力的第二未标记抗体。第二抗体可以存在于杂交瘤上清液中。作为对照,将固定化的抗原在溶液中温育,所述溶液包含第一标记抗体但没有第二未标记抗体。在允许第一抗体结合抗原的条件下温育后,除去过量的未结合的抗体,并测量与固定化抗原联合的标记物的量。如果与固定化抗原联合的标记物的量在测试样品中相对于对照样品实质性降低,那么这指示第二抗体在与第一抗体竞争对抗原的结合。参见harlow和lane(1988)antibodies:alaboratorymanualch.14(coldspringharborlaboratory,coldspringharbor,ny)。可以通过本领域已知的技术来纯化如本文中描述的那样制备的t细胞活化性双特异性抗原结合分子,所述技术如高效液相层析、离子交换层析、凝胶电泳、亲和层析、大小排阻层析等。用于纯化具体蛋白质的实际条件将部分取决于因素,如净电荷、疏水性、亲水性等,而且对于本领域中的技术人员将是明显的。对于亲和层析纯化,能使用t细胞活化性双特异性抗原结合分子结合的抗体、配体、受体或抗原。例如,对于本发明的t细胞活化性双特异性抗原结合分子的亲和层析纯化,可以使用具有蛋白a或蛋白g的基质。可以使用连续的蛋白a或g亲和层析和大小排阻层析来分离t细胞活化性双特异性抗原结合分子,基本如实施例中描述的。可以通过多种公知的分析方法中的任一种来测定t细胞活化性双特异性抗原结合分子的纯度,包括凝胶电泳、高压液相层析等。例如,如实施例中所描述的那样表达的重链融合蛋白显示为完整且正确装配的,如通过还原性sds-page证明的(见例如图4)。在约mr25,000,mr50,000和mr75,000处解析出三条条带,其对应于预测的t细胞活化性双特异性抗原结合分子轻链、重链和重链/轻链融合蛋白的分子量。测定法通过本领域中已知的各种测定法,可以鉴定、筛选或表征本文中提供的t细胞活化性双特异性抗原结合分子的物理/化学特性和/或生物学活性。亲和力测定法可以依照实施例中提出的方法使用标准的仪器如biacore仪(gehealthcare)通过表面等离振子共振(spr)测定t细胞活化性双特异性抗原结合分子对fc受体或靶抗原的亲和力,而且可以诸如通过重组表达获得受体或靶蛋白。或者,可以例如通过流式细胞术(facs),使用表达特定受体或靶抗原的细胞系来评估t细胞活化性双特异性抗原结合分子对不同受体或靶抗原的结合。一个用于测量结合亲和力的特定的说明性和例示性实施方案记载于下文和下文实施例。依照一个实施方案,通过表面等离振子共振使用t100仪(gehealthcare)于25℃测量kd。为了分析fc部分与fc受体之间的相互作用,通过固定化于cm5芯片上的抗五his抗体(qiagen)来捕捉带his标签的重组fc受体并将双特异性构建体用作分析物。简言之,将羧甲基化的葡聚糖生物传感器芯片(cm5,gehealthcare)依照供应商说明书用n-乙基-n’-(3-二甲基氨丙基)-碳二亚胺氢氯化物(edc)和n-羟基琥珀酰亚胺(nhs)活化。将抗五his抗体用10mm醋酸钠ph5.0稀释至40μg/ml,接着以5μl/分钟的流速注射以实现约6500响应单位(ru)的偶联蛋白。在注射配体后,注射1m乙醇胺以封闭未反应的基团。然后,在4或10nm捕捉fc受体60秒。对于动力学测量,将双特异性构建体的4倍连续稀释液(范围为约500nm至4000nm)在hbs-ep(gehealthcare,10mmhepes,150mmnacl,3mmedta,0.05%表面活性剂p20,ph7.4)中于25℃以约30μl/分钟的流速注射120秒。为了测定对靶抗原的亲和力,通过固定化于活化cm5传感器芯片表面上的抗人fab特异性抗体(gehealthcare)来捕捉双特异性构建体,如对于抗五-his抗体所描述的。偶联蛋白质的最终量为约12000ru。双特异性构建体在300nm捕捉90秒。使靶抗原在从250至1000nm的浓度范围以30μl/min的流速通过流动池达180秒。监测解离达180秒。通过扣除在参照流动池上获得的响应来校正批量折射率(bulkrefractiveindex)差异。使用稳定态响应通过langmuir结合等温线的非线性曲线拟合来导出解离常数kd。使用简单的1对1langmuir结合模型(t100评估软件版本1.1.1)通过同时拟合结合和解离传感图来计算结合速率(k结合)和解离速率(k解离)。平衡解离常数(kd)计算为比率k解离/k结合。参见例如chen等,jmolbiol293,865-881(1999)。活性测定法可以通过各种测定法来测量本发明的t细胞活化性双特异性抗原结合分子的生物学活性,如实施例中描述的。生物学活性可例如包括诱导t细胞的增殖、诱导t细胞中的信号传导、诱导t细胞中活化标志物的表达、诱导通过t细胞的细胞因子分泌、诱导靶细胞如肿瘤细胞的裂解、和诱导肿瘤消退和/或改善存活。组合物、配制剂和施用路径在一个别的方面,本发明提供包含本文中提供的任一种t细胞活化性双特异性抗原结合分子的药物组合物,例如用于以下任一种治疗方法。在一个实施方案中,药物组合物包含本文中提供的任一种t细胞活化性双特异性抗原结合分子以及药学可接受载体。在另一个实施方案中,药物组合物包含本文中提供的任一种t细胞活化性双特异性抗原结合分子以及至少一种另外的治疗剂,例如如下文描述的。还提供以适于体内施用的形式生成本发明的t细胞活化性双特异性抗原结合分子的方法,所述方法包括(a)获得依照本发明的t细胞活化性双特异性抗原结合分子,并(b)将所述t细胞活化性双特异性抗原结合分子与至少一种药学可接受载体配制在一起,由此配制成用于体内施用的t细胞活化性双特异性抗原结合分子制剂。本发明的药物组合物包含治疗有效量的在药学可接受载体中溶解或分散的一种或多种t细胞活化性双特异性抗原结合分子。短语“药学或药理学可接受的”指在所采用的剂量和浓度一般对接受者无毒性,即在适当时对动物如例如人施用时不产生不利、变应性或其它不当反应的分子实体和组合物。根据本公开,制备含有至少一种t细胞活化性双特异性抗原结合分子以及任选地另外的活性成分的药物组合物将是本领域技术人员已知的,如由remington’spharmaceuticalsciences,18thed.mackprintingcompany,1990例示的,其通过提述并入本文。此外,对于动物(例如人)施用,会理解制剂应当满足fda生物标准部门(fdaofficeofbiologicalstandards)或其它国家的相应机构要求的无菌性、热原性(pyrogenicity)、一般安全性和纯度标准。优选的组合物是冻干配制剂或水性溶液。如本文中使用的,“药学可接受载体”包括任何和所有的溶剂、缓冲剂、分散介质、涂料材料、表面活性剂、抗氧化剂、防腐剂(例如抗细菌剂、抗真菌剂)、等张剂、吸收延缓剂、盐、防腐剂、抗氧化剂、蛋白质、药物、药物稳定剂、聚合物、凝胶、粘合剂、赋形剂、崩解剂(disintegrationagent)、润滑剂、甜味剂、芳香剂、染料、此类类似的材料及其组合,如本领域普通技术人员将已知的(参见例如remington’spharmaceuticalsciences,18thed.mackprintingcompany,1990,pp.1289-1329,通过提述并入本文)。除非任何常规载体与活性成分不相容,涵盖其在治疗或药物组合物中的使用。所述组合物可以包含不同类型的载体,这取决于其要以固体、液体还是气雾剂形式施用,以及其对于此类施用路径如注射是否需要是无菌的。可以静脉内、皮内、动脉内、腹膜内、损伤内、颅内、关节内、前列腺内、脾内、肾内、胸膜内、气管内、鼻内、玻璃体内、阴道内、直肠内、肿瘤内、肌内、腹膜内、皮下、结膜下、囊泡内、粘膜、心包内、脐内、眼内、口服、表面(topically)、局部(locally)、通过吸入(例如气雾剂吸入)、注射、输注、连续输注、直接浸洗靶细胞的局部灌注、经由导管、经由灌洗、以乳剂、以液体组合物(例如脂质体)、或通过其它方法或前述项的任意组合施用本发明的t细胞活化性双特异性抗原结合分子(以及任何另外的治疗剂),如本领域中普通技术人员会知晓的(参见例如remington’spharmaceuticalsciences,18thed.mackprintingcompany,1990,通过提述并入本文)。胃肠外施用,特别是静脉内注射,最常用于施用多肽分子如本发明的t细胞活化性双特异性抗原结合分子。胃肠外组合物包括那些设计用于通过注射施用,例如皮下、皮内、损伤内、静脉内、动脉内、肌内、鞘内或腹膜内注射施用的组合物。对于注射,可以将本发明的t细胞活化性双特异性抗原结合分子在水性溶液,优选地在生理学相容的缓冲液中配制,所述生理学相容的缓冲液如汉克(hanks)氏溶液、林格(ringer)氏溶液或生理学盐水缓冲液。溶液可以含有配制剂如悬浮剂、稳定剂和/或分散剂。或者,t细胞活化性双特异性抗原结合分子可以为粉末形式,用于在使用前用合适的媒介物例如无菌无热原水构成。根据需要,通过将本发明的t细胞活化性双特异性抗原结合分子以需要的量掺入到具有下文列举的各种其它成分的合适溶剂中来制备无菌可注射溶液。可以容易地实现无菌,例如通过过滤流过无菌过滤膜进行。一般地,通过将各种无菌活性成分掺入到无菌媒介物中来制备分散剂,所述无菌媒介物含有基础分散介质和/或其它成分。在制备无菌可注射溶液、悬液或乳剂的无菌粉末的情况中,优选的制备方法是真空干燥或冷冻干燥技术,其将活性成分以及任何另外的期望成分的粉末从其先前无菌过滤的液体介质产生。液体介质在必要时应当是适当缓冲的,并且在用充足的盐水或葡萄糖注射前首先使液体稀释液等张。组合物在制备和贮存条件下必须是稳定的,并且针对微生物如细菌和真菌的污染作用提供保护。会领会的是,应当将内毒素污染最少保持于安全水平,例如低于0.5ng/mg蛋白质。合适的药学可接受载体包括但不限于:缓冲剂如磷酸、柠檬酸和其它有机酸;抗氧化剂,包括抗坏血酸和甲硫氨酸;防腐剂(如十八烷基二甲基苄基氯化铵;氯化六甲双铵(hexamethoniumchloride);氯化苯甲烃铵(benzalkoniumchloride);氯化苄乙铵(benzethoniumchloride);酚、丁醇或苯甲醇;烷基对羟基苯甲酸酯如对羟基苯甲酸甲酯或对羟基苯甲酸丙酯;儿茶酚;间苯二酚;环己醇;3-戊醇;和间甲酚);低分子量(低于约10个残基)多肽;蛋白质,如血清清蛋白、明胶、或免疫球蛋白;亲水性聚合物如聚乙烯吡咯烷酮;氨基酸如甘氨酸、谷氨酰胺、天冬酰胺、组氨酸、精氨酸或赖氨酸;单糖、二糖、和其它碳水化合物,包括葡萄糖、甘露糖或糊精;螯合剂如edta;糖如蔗糖、甘露醇、海藻糖或山梨糖醇;形成盐的反荷离子如钠;金属复合物(例如zn-蛋白复合物);和/或非离子型表面活性剂如聚乙二醇(peg)。水性注射悬液可以含有提高悬液粘度的化合物,如羧甲基纤维素钠、山梨糖醇、葡聚糖等。任选地,悬液还可以含有合适的稳定剂或增加化合物溶解度以允许制备高度浓缩的溶液的试剂。另外,可以将活性化合物的悬液制备为合适的油性注射悬液。合适的亲脂溶剂或媒介物包括脂肪油如芝麻油或合成的脂肪酸酯,如乙基cleats或甘油三酯或脂质体。可以将活性成分在例如分别通过凝聚技术或通过界面聚合作用制备的微囊剂,例如羟甲基纤维素或明胶微囊剂和聚-(甲基丙烯酸酯)微囊剂中,在胶体药物投递系统(例如脂质体、清蛋白微球、微乳剂、纳米颗粒和纳米胶囊)或在粗乳液(macroemulsion)中包载。此类技术披露于remington’spharmaceuticalsciences(18thed.mackprintingcompany,1990)。可以制备持续释放的制剂。合适的持续释放制剂的例子包括含多肽的固体疏水性聚合物的半透性基质,该基质为成形制品例如膜或微囊剂的形式。在具体的实施方案中,可以通过在组合物中使用吸收延迟剂如例如单硬脂酸铝、明胶或其组合,来产生可注射组合物的延长吸收。在先前描述的组合物外,t细胞活化性双特异性抗原结合分子还可以配制为贮存(depot)制剂。可以通过植入(例如皮下或肌内)或通过肌内注射来施用此类长效配制剂。如此,例如所述t细胞活化性双特异性抗原结合分子可以用合适的聚合或疏水性材料(例如作为可接受油中的乳剂)或离子交换树脂配制,或配制为微溶性衍生物,例如微溶性盐。可以通过常规的混合、溶解、乳化、包囊、包载或冻干过程来制备包含本发明的t细胞活化性双特异性抗原结合分子的药物组合物。可以以常规方式配制药物组合物,其使用一种或多种有助于将蛋白质加工成可药学使用的制剂的生理学可接受载体、稀释剂、赋形剂或辅助剂。合适的配制剂依赖于选择的施用路径。可以将t细胞活化性双特异性抗原结合分子以游离酸或碱、中性或盐形式配制成组合物。药学可接受盐是基本保留游离酸或碱的生物学活性的盐。这些包括酸加成盐(acidadditionsalt),例如与蛋白质性组合物的游离氨基基团形成的那些,或与无机酸(如例如氢氯酸或磷酸)或与有机酸如乙酸、草酸、酒石酸或扁桃酸形成的。与游离羧基基团形成的盐还可以自无机碱如例如氢氧化钠、钾、铵、钙或铁;或有机碱如异丙胺、三甲胺、组氨酸或普鲁卡因(procaine)衍生。药用盐倾向于比相应的游离碱形式更可溶于水性溶剂和其它质子溶剂中。治疗方法和组合物可以将本文中提供的任一种t细胞活化性双特异性抗原结合分子用在治疗方法中。本发明的t细胞活化性双特异性抗原结合分子可用作免疫治疗剂,例如在癌症的治疗中。对于在治疗方法中的使用,将以与优良医学实践一致的方式配制、给药和施用本发明的t细胞活化性双特异性抗原结合分子。在此背景中考虑的因素包括治疗的特定病症、治疗的特定哺乳动物、个体患者的临床状况、病症的起因、药剂的投递部位、施用方法、施用时间安排以及医学从业人员已知的其它因素。在一个方面,提供用作药物的本发明的t细胞活化性双特异性抗原结合分子。在别的方面,提供用于治疗疾病的本发明的t细胞活化性双特异性抗原结合分子。在某些实施方案中,提供用于治疗方法的本发明的t细胞活化性双特异性抗原结合分子。在一个实施方案中,本发明提供如本文中描述的t细胞活化性双特异性抗原结合分子,用于治疗有此需要的个体中的疾病。在某些实施方案中,本发明提供t细胞活化性双特异性抗原结合分子,用于治疗患有疾病的个体的方法,所述方法包括对所述个体施用治疗有效量的t细胞活化性双特异性抗原结合分子。在某些实施方案中,待治疗的疾病是增殖性病症。在一个具体的实施方案中,所述疾病是癌症。在某些实施方案中,所述方法进一步包括对个体施用治疗有效量的至少一种另外的治疗剂,例如抗癌剂(如果待治疗的疾病是癌症的话)。在别的实施方案中,本发明提供如本文中描述的t细胞活化性双特异性抗原结合分子,用于诱导靶细胞,特别是肿瘤细胞的裂解。在某些实施方案中,本发明提供t细胞活化性双特异性抗原结合分子,用于在个体中诱导靶细胞,特别是肿瘤细胞裂解的方法,该方法包括对该个体施用有效量的t细胞活化性双特异性抗原结合分子以诱导靶细胞的裂解。依照上文任何实施方案的“个体”是哺乳动物,优选是人。在一个别的方面,本发明提供本发明的t细胞活化性双特异性抗原结合分子在制造或制备药物中的用途。在一个实施方案中,所述药物用于治疗有此需要的个体中的疾病。在一个别的实施方案中,所述药物用于治疗疾病的方法,该方法包括对患疾病的个体施用治疗有效量的药物。在某些实施方案中,待治疗的疾病是增殖性病症。在一个具体的实施方案中,所述疾病是癌症。在一个实施方案中,所述方法进一步包括对个体施用治疗有效量的至少一种另外的治疗剂,例如抗癌剂(如果待治疗的疾病是癌症的话)。在一个别的实施方案中,所述药物用于诱导靶细胞,特别是肿瘤细胞的裂解。仍在别的实施方案中,所述药物用于在个体中诱导靶细胞,特别是肿瘤细胞裂解的方法,所述方法包括对该个体施用有效量的药物以诱导靶细胞的裂解。依照上文任何实施方案的“个体”是哺乳动物,优选是人。在一个别的方面,本发明提供用于治疗疾病的方法。在一个实施方案中,所述方法包括对患有此类疾病的个体施用治疗有效量的本发明的t细胞活化性双特异性抗原结合分子。在一个实施方案中,对所述个体施用组合物,其包含药学可接受的形式的本发明的t细胞活化性双特异性抗原结合分子。在某些实施方案中,待治疗的疾病是增殖性病症。在一个具体的实施方案中,所述疾病是癌症。在某些实施方案中,所述方法进一步包括对个体施用治疗有效量的至少一种另外的治疗剂,例如抗癌剂(如果待治疗的疾病是癌症的话)。依照上文任何实施方案的“个体”是哺乳动物,优选是人。在一个别的方面,本发明提供一种用于诱导靶细胞,特别是肿瘤细胞裂解的方法。在一个实施方案中,所述方法包括在存在t细胞,特别是细胞毒性t细胞的情况下使靶细胞与本发明的t细胞活化性双特异性抗原结合分子接触。在一个别的方面,提供一种用于在个体中诱导靶细胞,特别是肿瘤细胞裂解的方法。在一个此类实施方案中,所述方法包括对个体施用有效量的t细胞活化性双特异性抗原结合分子以诱导靶细胞裂解。在一个实施方案中,“个体”是人。在某些实施方案中,所述待治疗的疾病是增殖性病症,特别是癌症。癌症的非限制性例子包括膀胱癌、脑癌、头和颈癌、胰腺癌、肺癌、乳腺癌、卵巢癌、子宫癌、宫颈癌、子宫内膜癌、食道癌、结肠癌、结肠直肠癌、直肠癌、胃癌、前列腺癌、血液癌、皮肤癌、鳞状细胞癌、骨癌和肾癌。其它可使用本发明的t细胞活化性双特异性抗原结合分子治疗的细胞增殖病症包括但不限于位于下列各项中的新生物:腹部、骨、乳房、消化系统、肝、胰、腹膜、内分泌腺(肾上腺、副甲状腺、垂体、睾丸、卵巢、胸腺、甲状腺)、眼、头和颈、神经系统(中枢和外周)、淋巴系统、骨盆、皮肤、软组织、脾、胸区、和泌尿生殖系统。还包括癌症前期状况或损伤以及癌症转移。在某些实施方案中,癌症选自下组:肾细胞癌、皮肤癌、肺癌、结肠直肠癌、乳腺癌、脑癌、头和颈癌。熟练技术人员容易地认可在许多情况下,t细胞活化性双特异性抗原结合分子可能不提供治愈而仅可以提供部分益处。在一些实施方案中,具有一些益处的生理学变化也被视为治疗有益的。如此,在一些实施方案中,提供生理学变化的t细胞活化性双特异性抗原结合分子的量被视为“有效量”或“治疗有效量”。需要治疗的受试者、患者或个体通常为哺乳动物,更特定地为人。在一些实施方案中,对细胞施用有效量的本发明的t细胞活化性双特异性抗原结合分子。在其它实施方案中,对个体施用治疗有效量的本发明的t细胞活化性双特异性抗原结合分子以治疗疾病。为了预防或治疗疾病,本发明的t细胞活化性双特异性抗原结合分子的合适剂量(当单独或与一种或多种其它另外的治疗剂组合使用时)将取决于待治疗疾病的类型、施用路径、患者的体重、t细胞活化性双特异性抗原结合分子的类型、疾病的严重程度和进程、施用t细胞活化性双特异性抗原结合分子是为了预防还是治疗目的、先前或同时的治疗干预、患者的临床史和对t细胞活化性双特异性抗原结合分子的响应、以及主治医师的判断。负责施用的从业人员将在任何事件中确定组合物中活性成分的浓度和用于个体受试者的合适剂量。本文中涵盖各种给药方案,包括但不限于在各个时间点的单次或多次施用、推注施用、和脉冲输注。所述t细胞活化性双特异性抗原结合分子适宜地在一次或一系列治疗里对患者施用。根据疾病的类型和严重程度,约1μg/kg至15mg/kg(例如0.1mg/kg-10mg/kg)的t细胞活化性双特异性抗原结合分子可以是用于对患者施用的起始候选剂量,不管是例如通过一次或多次分开的施用,还是通过连续输注进行。根据上文提及的因素,一种典型的每日剂量的范围可以从约1μg/kg至100mg/kg或更多。对于在数天或更长里的重复施用,根据状况,治疗一般将持续直至发生对疾病症状的期望的抑制。t细胞活化性双特异性抗原结合分子的一种例示性剂量将在约0.005mg/kg至约10mg/kg的范围内。在其它非限制性例子中,剂量还可包括每次施用从约1微克/kg体重、约5微克/kg体重、约10微克/kg体重、约50微克/kg体重、约100微克/kg体重、约200微克/kg体重、约350微克/kg体重、约500微克/kg体重、约1毫克/kg体重、约5毫克/kg体重、约10毫克/kg体重、约50毫克/kg体重、约100毫克/kg体重、约200毫克/kg体重、约350毫克/kg体重、约500毫克/kg体重,至约1000mg/kg体重或更多,以及其中可导出的任何范围。在从本文列出的数量可导出的范围的非限制性例子中,基于上文描述的数目,可以施用约5mg/kg体重至约100mg/kg体重的范围,约5微克/kg体重至约500毫克/kg体重的范围等。如此,可以对患者施用一剂或多剂的约0.5mg/kg、2.0mg/kg、5.0mg/kg或10mg/kg(或其任意组合)。可以间歇地施用此类剂量,例如每周或每3周(例如使得患者接受约2至约20、或例如约6剂的t细胞活化性双特异性抗原结合分子)。可以施用起始较高的加载剂量,继之以一剂或多剂较低剂量。然而,可以使用其它剂量方案。通过常规技术和测定法容易监测该疗法的进行。本发明的t细胞活化性双特异性抗原结合分子一般将以对于实现意图的目的有效的量使用。对于治疗或预防疾病状况的用途,以治疗有效量施用或应用本发明的t细胞活化性双特异性抗原结合分子、或其药物组合物。治疗有效量的确定完全在本领域技术人员的能力以内,尤其根据本文中提供的详细公开内容。对于系统性施用,能从体外测定法如细胞培养测定法初步估算出治疗有效剂量。然后可以在动物模型中配制剂量以达到包含如在细胞培养中测定的ic50的循环浓度范围。可以将此类信息用于更准确地确定人中的可用剂量。使用本领域中公知的技术,还能从体内数据例如动物模型估算出初始剂量。本领域中的普通技术人员能容易地基于动物数据优化对人的施用。可以分别调整剂量量和时间间隔以提供足以维持治疗效果的t细胞活化性双特异性抗原结合分子的血浆水平。通过注射施用的可用患者剂量的范围为从约0.1至50mg/kg/天,通常约0.5至1mg/kg/天。可以通过每日施用多剂实现治疗有效的血浆水平。可以例如通过hplc测量血浆中的水平。在局部施用或选择性摄取的情况中,t细胞活化性双特异性抗原结合分子的有效局部浓度可能与血浆浓度无关。本领域技术人员会能够在无需过度实验的情况下优化治疗有效的局部剂量。本文中描述的t细胞活化性双特异性抗原结合分子的治疗有效剂量一般将提供治疗益处,而不导致实质性毒性。可以通过在细胞培养物或实验动物中的标准药学规程来测定t细胞活化性双特异性抗原结合分子的毒性和治疗功效。可以使用细胞培养测定法和动物研究来测定ld50(对50%的群体致命的剂量)和ed50(在50%的群体中治疗有效的剂量)。毒性和治疗效果之间的剂量比率为治疗指数,其可以表达为ld50/ed50比。优选展现出较大治疗指数的t细胞活化性双特异性抗原结合分子。在一个实施方案中,依照本发明的t细胞活化性双特异性抗原结合分子展现出高治疗指数。可以将从细胞培养测定法和动物研究获得的数据用来制定适用于人的剂量范围。优选地,剂量处于具有很小或无毒性的循环浓度(包含ed50)的范围内。剂量在此范围内可以随多种因素,例如采用的剂量形式、利用的施用路径、受试者的状况等而变化。鉴于患者的状况,各个内科医生可以选择确切的配制剂、施用路径和剂量(参见例如fingl等,1975,于:thepharmacologicalbasisoftherapeutics,ch.1,p.1,通过提述完整并入本文)。用本发明的t细胞活化性双特异性抗原结合分子治疗的患者的主治内科医生将知晓如何及何时终止、中断或调整施用(由于毒性、器官功能障碍等)。相反,如果临床应答不适当(排除毒性),主治内科医生还将知晓如何将治疗调整至更高水平。在感兴趣的病症的管理中的施用剂量的量级将随待治疗状况的严重程度、施用路径等而变化。可以例如部分地通过标准的预后评估方法来评估状况的严重程度。另外,剂量以及可能的给药频率也将随个体患者的年龄、体重和应答而变化。其它药剂和治疗本发明的t细胞活化性双特异性抗原结合分子可以与一种或多种其它药剂在疗法中组合施用。例如,本发明的t细胞活化性双特异性抗原结合分子可以与至少一种另外的治疗剂共施用。术语“治疗剂”涵盖施用以治疗需要此类治疗的个体中的症状或疾病的任何药剂。此类另外的治疗剂可以包含任何适用于所治疗的特定适应征的任何活性成分,优选地具有不会彼此不利影响的互补活性的那些活性成分。在某些实施方案中,另外的治疗剂是免疫调控剂、细胞抑制剂、细胞粘着的抑制剂、细胞毒剂、细胞凋亡的激活剂、或提高细胞对凋亡诱导剂的敏感性的药剂。在一个具体的实施方案中,另外的治疗剂是抗癌剂,例如微管破坏物、抗代谢物、拓扑异构酶抑制剂、dna嵌入剂、烷化剂、激素疗法、激酶抑制剂、受体拮抗剂、肿瘤细胞凋亡的激活剂、或抗血管生成剂。此类其它药剂以对意图目的有效的量适宜地组合存在。此类其它药剂的有效量取决于使用的t细胞活化性双特异性抗原结合分子的量、病症或治疗的类型以及上文所述其它因素。所述t细胞活化性双特异性抗原结合分子一般以与本文中描述的相同的剂量和施用路径、或以本文中描述的剂量的约1至99%、或通过凭经验/临床上确定为合适的任何剂量和任何路径使用。上文记载的此类组合疗法涵盖组合施用(其中在同一组合物或分别的组合物中包含两种或更多种治疗剂)和分开施用,在该情况中,本发明的t细胞活化性双特异性抗原结合分子的施用可以在施用另外的治疗剂和/或辅助剂之前、同时和/或之后发生。本发明的t细胞活化性双特异性抗原结合分子还可以与放射疗法组合使用。制品在本发明的另一个方面,提供含有可用于治疗、预防和/或诊断上文描述的病症的材料的制品。所述制品包含容器和容器上或与容器联合的标签或包装插页。合适的容器包括例如瓶、管形瓶、注射器、iv溶液袋等。所述容器可从多种材料如玻璃或塑料形成。所述容器容纳组合物,其自身或与其它组合物组合对于治疗、预防和/或诊断状况是有效的,并且可以具有无菌的存取口(例如,容器可以是具有由皮下注射针可穿过的塞子的静脉内溶液袋或管形瓶)。组合物中至少一种活性成分是本发明的t细胞活化性双特异性抗原结合分子。标签或包装插页指示该组合物用于治疗选择的状况。此外,所述制品可以包含(a)其中含有组合物的第一容器,其中所述组合物包含本发明的t细胞活化性双特异性抗原结合分子;和(b)其中含有组合物的第二容器,其中所述组合物包含另外的细胞毒性或其它方面治疗剂。本发明的这一实施方案中的制品还可以包含包装插页,其指示该组合物可用于治疗特定状况。或者/另外,所述制品还可以包含第二(或第三)容器,其包含药学可接受缓冲液,如抑菌性注射用水(bwfi)、磷酸盐缓冲盐水、林格(ringer)氏溶液和右旋糖溶液。它可以进一步包含从商业和用户观点看期望的其它材料,包括其它缓冲液、稀释剂、滤器、针、和注射器。实施例下面是本发明的方法和组合物的实施例。应当理解,鉴于上文提供的一般性描述,可以实践各种其他实施方案。通用方法重组dna技术使用标准方法操作dna,如sambrooketal.,molecularcloning:alaboratorymanual;coldspringharborlaboratorypress,coldspringharbor,newyork,1989中描述的。依照制造商的说明书使用分子生物学试剂。关于人免疫球蛋白轻和重链的核苷酸序列的一般信息参见:kabat,e.a.etal.,(1991)sequencesofproteinsofimmunologicalinterest,5thed.,nihpublicationno91-3242。dna测序通过双链测序来测定dna序列。基因合成在需要的情况下,期望的基因区段使用适宜的模板通过pcr来生成或由geneartag(regensburg,germany)通过自动化基因合成从合成的寡核苷酸和pcr产物合成。在确切基因序列不可得的情况下,基于来自最近的同源物的序列设计寡核苷酸引物,并通过rt-pcr从源自适宜组织的rna分离基因。将侧翼为单一限制性内切核酸酶切割位点的基因区段克隆入标准的克隆/测序载体。从经转化的细菌纯化质粒dna,并通过uv分光术测定浓度。通过dna测序确认亚克隆的基因片段的dna序列。基因区段设计为具有合适的限制性位点以允许亚克隆入相应的表达载体。所有构建体均设计为具有编码前导肽的5’端dna序列,该前导肽在真核细胞中将蛋白质靶向分泌。例示性前导肽及其编码多核苷酸序列描绘于seqidno108-116。从pbmc分离原代人泛t细胞通过histopaque密度离心从富集的淋巴细胞制备物(血沉棕黄层)制备外周血单个核细胞(pbmc),所述制备物自当地血库或来自健康人供体的新鲜血液获得。简言之,将血液用无菌pbs稀释并小心在histopaque梯度(sigma,h8889)上铺层。于室温以450xg离心30分钟后(关闭制动器),丢弃含pbmc界面上方的血浆部分。将pbmc转移至新的50mlfalcon管并将管用pbs补充至总体积50ml。将混合物于室温以400xg离心10分钟(开启制动器)。丢弃上清液并将pbmc团粒用无菌pbs清洗两次(离心步骤为4℃,350xg,10分钟)。将所得pbmc群自动计数(vicell)并在温箱中在37℃、5%co2在含有10%fcs和1%l-丙氨酰-l-谷氨酰胺(biochrom,k0302)的rpmi1640培养基中贮存直至测定开始。使用泛t细胞分离试剂盒ii(miltenyibiotec#130-091-156)依照制造商的说明书实施从pbmc富集t细胞。简言之,将细胞团粒以每1千万个细胞40μl冷缓冲液(含0.5%bsa、2mmedta的pbs,无菌过滤)稀释并与每1千万个细胞10μl生物素-抗体混合物一起于4℃温育10分钟。每1千万个细胞添加30μl冷缓冲液和20μl抗生物素磁珠,并将混合物于4℃再温育15分钟。通过添加10-20倍当前体积的缓冲液和300xg,10分钟的随后离心步骤来清洗细胞。将高达1亿个细胞重悬浮于500μl缓冲液。使用ls柱(miltenyibiotec#130-042-401)依照制造商的说明书实施未标记人泛t细胞的磁性分离。将所得t细胞群自动计数(vicell)并在温箱中在37℃、5%co2在aim-v培养基中贮存直至测定开始(不超过24小时)。从pbmc分离原代人幼稚t细胞通过histopaque密度离心从富集的淋巴细胞制备物(血沉棕黄层)制备外周血单个核细胞(pbmc),所述制备物(血沉棕黄层)自当地血库或来自健康人供体的新鲜血液获得。使用幼稚cd8+t细胞分离试剂盒(miltenyibiotec#130-093-244)依照制造商的说明书实施从pbmc富集t细胞,但是跳过最后一个cd8+t细胞分离步骤(亦见关于分离原代人泛t细胞的描述)。从脾细胞分离鼠泛t细胞从c57bl/6小鼠分离脾,转移到装有macs缓冲液(pbs+0.5%bsa+2mmedta)的gentlemacsc管(miltenyibiotech#130-093-237)中,并依照制造商的说明书用gentlemacs解离器解离以获得单细胞悬浮液。使细胞悬浮液通过预分离滤器以除去剩余的未解离组织颗粒。于4℃以400xg离心4分钟后,添加ack裂解缓冲液以裂解红血球(于室温温育5分钟)。将剩余细胞用macs缓冲液清洗两次,计数并用于分离鼠泛t细胞。使用泛t细胞分离试剂盒(miltenyibiotec#130-090-861)依照制造商的说明书实施负(磁性)选择。将所得t细胞群自动计数(vicell)并立即用于进一步测定。从肝素化血液分离原代猕猴pbmc如下通过密度离心从来自健康猕猴供体的新鲜血液制备外周血单个核细胞(pbmc):将肝素化血液用无菌pbs以1:3稀释,并将lymphoprep介质(axonlab#1114545)用无菌pbs稀释至90%。将2份体积的稀释血液在1份体积的稀释密度梯度上铺层,并通过于室温以520xg离心30分钟(无制动)来分离pbmc级分。将pbmc带转移入新的50mlfalcon管并通过于4℃以400xg离心10分钟用无菌pbs清洗。实施一次低速离心以除去血小板(150xg,15分钟,4℃),将所得pbmc群自动计数(vicell)并立即用于进一步测定。靶细胞对于mcsp靶向性双特异性抗原结合分子的评估,使用下列肿瘤细胞系:人黑素瘤细胞系wm266-4(atcc#crl-1676),其衍生自恶性黑素瘤的转移性部位且表达高水平的人mcsp;人黑素瘤细胞系mv-3(来自radboud大学nijmegen医学中心的馈赠),其表达中等水平的人mcsp;人恶性黑素瘤(原发性肿瘤)细胞系a375(ecacc#88113005),其表达高水平的mcsp;人结肠癌细胞系hct-116(atcc#ccl-247),其不表达mcsp;和人高加索人结肠腺癌细胞系ls180(ecacc#87021202),其不表达mcsp。对于cea靶向性双特异性抗原结合分子的评估,使用下列肿瘤细胞系:人胃癌细胞系mkn45(dsmz#acc409),其表达很高水平的人cea;人胰腺腺癌细胞系hpaf-ii(rochenutley的馈赠),其表达高水平的人cea;人原发性胰腺腺癌细胞系bxpc-3(ecacc#93120816),其表达中等水平的人cea;人女性高加索人结肠腺癌细胞系ls-174t(ecacc#87060401),其表达中等水平的人cea;人胰腺腺癌细胞系aspc-1(ecacc#96020930),其表达低水平的人cea;人上皮状胰腺癌细胞系panc-1(atcc#crl-1469),其表达(很)低水平的人cea;人结肠癌细胞系hct-116(atcc#ccl-247),其不表达cea;人腺癌肺泡基底上皮细胞系a549-hucea,其在内部经稳定转染而表达人cea;和鼠结肠癌细胞系mc38-hucea,其在内部经改造而稳定表达人cea。另外,使用人t细胞白血病细胞系jurkat(atcc#tib-152)来评估不同双特异性构建体对细胞上人cd3的结合。实施例1抗mcsp抗体m4-3/ml2的亲和力成熟经由寡核苷酸指导的诱变规程实施亲和力成熟。为此,将重链变体m4-3和轻链变体ml2克隆入噬菌粒载体,与hoogenboom描述的那些类似(hoogenboomettal.nucleicacidsres.1991,19,4133-4137)。如下鉴定要随机化的残基,即首先经由经典的同源性建模产生该抗体的3d模型,然后鉴定重和轻链互补决定区(cdr)的溶剂可及残基。表1所示具有基于三核苷酸合成的随机化的寡核苷酸购自ellabiotech(munich,germany)。经由经典的pcr产生三个独立的子文库,它们包含cdr-h1连同cdr-h2,或cdr-l1连同cdr-l2中的随机化。以另一种办法随机化cdr-l3。经由限制性消化和连接将那些文库的dna片段克隆入噬菌粒,并随后电穿孔入tg1细菌。文库选择如此产生的抗体变体作为每个颗粒内包装的与m13基因iii产物的融合以单价形式从丝状噬菌体颗粒上展示。然后就其生物学活性(在此:结合亲和力)筛选噬菌体展示变体,并将具有一项或多项改进活性的候选者用于进一步的开发。用于制备噬菌体展示文库的方法可参见leeetal.,j.mol.biol.(2004)340,1073-1093。依照下述规程在溶液中用所有亲和力成熟文库进行选择:1.以总体积1ml将每个亲和力成熟文库的约1012个噬菌粒颗粒与100nm生物素化hu-mcsp(d3域)-avi-his(seqidno:118)结合0.5小时,2.通过添加5.4x107个链霉亲合素包被磁珠10分钟来捕捉生物素化hu-mcsp(d3域)-avi-his和特异结合的噬菌体颗粒,3.用5-10x1mlpbs/tween-20和5-10x1mlpbs清洗珠子,4.通过添加1ml100mmtea(三乙胺)10分钟洗脱噬菌体颗粒并通过添加500μl1mtris/hclph7.4进行中和,及5.指数生长中的大肠杆菌tg1细菌的再感染,用辅助噬菌体vcsm13感染并随后peg/nacl沉淀噬菌粒颗粒用于随后的选择轮次。使用恒定的或递减的(从10-7m至2x10-9m)抗原浓度进行3-5轮选择。在第2轮中,使用中性亲合素平板替代链霉亲合素珠实施抗原-噬菌体复合物捕捉。如下通过elisa鉴定特异性结合者:在中性亲合素板上包被每孔100μl10nm生物素化hu-mcsp(d3域)-avi-his。添加含fab的细菌上清液并经由它们的flag标签通过使用抗flag/hrp二抗检测结合性fab。以96孔格式作为可溶性fab片段用细菌表达elisa阳性克隆并使用proteonxpr36(biorad)通过spr分析对上清液进行动力学筛选实验。鉴定所表达之fabs具有最高亲和常数的克隆并对相应噬菌粒测序。表1(始终排除cys和met。在寡核苷酸为反向引物的那些案例中另外排除lys)图2显示亲和力成熟的抗mcsp克隆与非成熟的亲本克隆(m4-3ml2)的比对。只在cdr1和2中实施重链随机化。在cdr1和2中以及独立地在cdr3中实施轻链随机化。在选择期间,出现框架中的一些突变,像克隆g3中的f71y或克隆e10中的y87h。人igg1的生成和纯化将亲和力成熟变体的重和轻链dna序列的可变区亚克隆入各自接受者哺乳动物表达载体,与预插入的恒定重链或恒定轻链同框。抗体表达由mpsv启动子驱动且在cds的3’端携带合成的polya信号序列。此外,每种载体包含ebvorip序列。通过使用聚乙烯亚胺(pei)用哺乳动物表达载体共转染hek293-ebna细胞生成该分子。用相应表达载体以1:1的比率转染细胞。为了转染,将hek293ebna细胞在cdcho培养基中无血清悬浮培养。对于500ml摇瓶中的生成,在转染前24小时接种4亿个hek293ebna细胞。为了转染,将细胞以210xg离心5分钟,用预温热的20mlcdcho培养基置换上清液。在20mlcdcho培养基中混合表达载体至最终200μgdna的量。添加540μlpei溶液后,将混合物涡旋15秒,随后于室温温育10分钟。之后将细胞与dna/pei溶液混合,转移至500ml摇瓶并在具有5%co2气氛的温箱中于37℃温育3小时。温育时间后添加160mlf17培养基并将细胞培养24小时。转染后1天添加1mm丙戊酸和7%给料1(lonza)。培养7天后通过以210xg离心15分钟收集上清液用于纯化,将溶液无菌过滤(0.22μm滤器)并添加终浓度0.01%w/v的叠氮化钠,保存于4℃。通过蛋白a亲和层析从细胞培养物上清液纯化分泌的蛋白质。将上清液加载到用40ml20mm磷酸钠,20mm柠檬酸钠,0.5m氯化钠,ph7.5平衡的hitrap蛋白ahp柱(cv=5ml,gehealthcare)上。通过用至少10个柱体积的20mm磷酸钠,20mm柠檬酸钠,0.5m氯化钠,ph7.5清洗去除未结合的蛋白质。靶蛋白在20个柱体积上从20mm柠檬酸钠,0.5m氯化钠,ph7.5至20mm柠檬酸钠,0.5m氯化钠,ph2.5的梯度期间洗脱。通过添加1/10的0.5m磷酸钠,ph8中和蛋白质溶液。浓缩并过滤靶蛋白,之后加载到用ph6.0的20mm组氨酸,140mm氯化钠溶液平衡的hiloadsuperdex200柱(gehealthcare)上。通过测量280nm的光密度(od)使用基于氨基酸序列计算的摩尔消光系数确定经过纯化的蛋白质样品的蛋白质浓度。通过还原剂存在和缺失下的ce-sds分析来分析分子的纯度和分子量。依照制造商的说明书使用caliperlabchipgxii系统(caliperlifescience)。将2μg样品用于分析。使用tskgelg3000swxl分析型大小排阻柱(tosoh)在25mmk2hpo4,125mmnacl,200mml-精氨酸单盐酸盐,0.02%(w/v)nan3,ph6.7运行缓冲液中于25℃分析抗体样品的聚集体含量。表2:亲和力成熟的抗mcspigg的生成和纯化构建体产量[mg/l]hmw[%]lmw[%]单体[%]m4-3(c1)ml2(g3)43.900100m4-3(c1)ml2(e10)59.500100m4-3(c1)ml2(c5)68.900.899.2亲和力测定proteon分析用通过胺偶联在cm5芯片上固定化的抗人f(ab’)2片段特异性捕捉抗体(jacksonimmunoresearch#109-005-006)及随后从细菌上清液或从经过纯化的fab制备物中捕捉fab,使用proteonxpr36机器(biorad)与25℃通过表面等离振子共振测量kd。简言之,用n-乙基-n’-(3-二甲基氨基丙基)-碳二亚胺盐酸盐(edc)和n-羟基琥珀酰亚胺(nhs)依照供应商的说明书活化羧甲基化右旋糖苷生物传感器芯片(cm5,gehealthcare)。将抗人f(ab’)2片段特异性捕捉抗体用10mm乙酸钠,ph5.0稀释成50μg/ml,之后以流速10μl/min注射来获得大约高达10.000个响应单位(ru)的偶联捕捉抗体。注射捕捉抗体后,注射1m乙醇胺以封闭未反应的基团。对于动力学测量,将来自细菌上清液的fab或经过纯化的fab以流速10μl/min注射300秒,并解离300秒以捕捉基线稳定性。捕捉水平在100-500ru的范围中。在随后的步骤中,以单一浓度或作为在hbs-ep+(gehealthcare,10mmhepes,150mmnacl,3mmedta,0.05%表面活性剂p20,ph7.4)中稀释的浓度系列(取决于克隆亲和力,在100nm至250pm之间的范围中)于25℃以流速50μl/min注射人mcsp(d3域)-avi-his分析物。通过以90μl/min注射甘氨酸ph1.530秒,接着以相同流速注射naoh20秒来再生传感器芯片的表面。使用简单一对一langmuir结合模型(proteonxpr36评估软件或scrubber软件(biologic))通过同时拟合结合和解离传感图计算结合速率(kon)和解离速率(koff)。作为比率koff/kon计算平衡解离常数(kd)。此数据用于确定亲和力成熟变体与亲本抗体的比较性结合亲和力。表3a显示自这些测定法产生的数据。对于轻链选择g3、e10、c5,对于重链选择d6、a7、b7、b8、c1,转换成人igg1形式。由于轻链的cdr1和2的随机化独立于cdr3,所以在igg转换期间组合获得的cdr。在igg形式中再次测量对人mcsp抗原(seqidno:118),此外还有对猕猴同系物(seqidno:117)的亲和力。所使用的方法恰如对fab片段描述的,仅仅使用自哺乳动物生成纯化的的igg。表3a:mcsp亲和力成熟克隆:proteon数据。使用biacoret200通过表面等离振子共振(spr)进行的亲和力测定在biacoret200上于25℃以hbs-ep作为运行缓冲液(0.01mhepesph7.4,0.15mnacl,3mmedta,0.005%表面活性剂p20,biacore,freiburg/germany)实施表面等离振子共振(spr)实验以测定亲和力成熟igg的亲和力和亲合力。为了分析不同抗mcspigg对人和猕猴mcspd3的相互作用的亲合力,在cm5芯片上于ph5.0使用标准胺偶联试剂盒(biacore,freiburg/germany)实施9,500个共振单位(ru)左右的抗5xhis抗体(qiagen)的直接偶联。分别以10μl/min在30nm捕捉抗原60秒。igg以浓度0.0064-100nm和流速30μl/min通过流通池280秒。监测解离180秒。通过扣除参照流通池上获得的响应校正批量折射指数差异。在此,igg流经带固定化抗5xhis抗体的表面,但其上已注射hbs-ep而非人mcspd3或猕猴mcspd3。对于亲和力测量,在带固定化抗人fc的cm5传感器芯片表面上捕捉igg。通过使用标准胺偶联试剂盒(biacore,freiburg/germany)于ph5.09,500个共振单位(ru)左右的直接固定化将捕捉igg偶联至传感器芯片表面。在10nm以30μl/min捕捉igg25秒。在120秒里使人和猕猴mcspd3以浓度2-500nm和流速30μl/min通过流通池。监测解离60秒。对于浓度166和500nm分别监测结合和解离1200和600秒。通过扣除参照流通池上获得的响应校正批量折射指数差异。在此,抗原流经带固定化抗人fc抗体的表面,但其上已注射hbs-ep而非抗mcspigg。使用biacoret200评估软件(vaa,biacoreab,uppsala/sweden)以通过数值积分拟合1:1langmuir结合的速率方程来推导动力学常数。通过表面等离振子共振测量使用biacoret200证实对人和猕猴mcspd3的较高亲和力。此外,亲合力测量显示在二价结合中高达3倍的增加(表3b)。表3b:抗mcspigg对人mcsp-d3和猕猴mcsp-d3的亲和力和亲合力。实施例2包含m4-3(c1)ml2(g3)作为抗mcsp抗体和人源化ch2527作为抗cd3抗体的mcsptcb(2+1crossfab-iggp329glala倒转的)的制备将重和轻链dna序列的可变区亚克隆入各自接受者哺乳动物表达载体,与预插入的恒定重链或恒定轻链同框。抗体表达由mpsv启动子驱动且在cds的3’端携带合成的polya信号序列。此外,每种载体包含ebvorip序列。通过使用聚乙烯亚胺(pei)用哺乳动物表达载体共转染hek293-ebna细胞生成该分子。用相应表达载体以1:2:1:1的比率(“载体重链fc(穴)”:“载体轻链”:“载体轻链crossfab”:“载体重链fc(节)-fabcrossfab”)转染细胞。为了转染,将hek293ebna细胞在cdcho培养基中无血清悬浮培养。对于500ml摇瓶的生成,在转染前24小时接种4亿个hek293ebna细胞。为了转染,将细胞以210xg离心5分钟,用预温热的20mlcdcho培养基置换上清液。在20mlcdcho培养基中混合表达载体至最终200μgdna的量。添加540μlpei溶液后,将混合物涡旋15秒,随后于室温温育10分钟。之后将细胞与dna/pei溶液混合,转移至500ml摇瓶并在具有5%co2气氛的温箱中于37℃温育3小时。温育时间后添加160mlf17培养基并将细胞培养24小时。转染后1天添加1mm丙戊酸和7%给料1(lonza)。培养7天后通过以210xg离心15分钟收集上清液用于纯化,将溶液无菌过滤(0.22μm滤器)并添加终浓度0.01%w/v的叠氮化钠,保存于4℃。通过蛋白a亲和层析从细胞培养物上清液纯化分泌的蛋白质。将上清液加载到用40ml20mm磷酸钠,20mm柠檬酸钠,0.5m氯化钠,ph7.5平衡的hitrap蛋白ahp柱(cv=5ml,gehealthcare)上。通过用至少10个柱体积的20mm磷酸钠,20mm柠檬酸钠,0.5m氯化钠,ph7.5清洗去除未结合的蛋白质。靶蛋白在20个柱体积上从20mm柠檬酸钠,0.5m氯化钠,ph7.5至20mm柠檬酸钠,0.5m氯化钠,ph2.5的梯度期间洗脱。通过添加1/10的0.5m磷酸钠,ph8中和蛋白质溶液。浓缩并过滤靶蛋白,之后加载到用ph6.0的20mm组氨酸,140mm氯化钠溶液平衡的hiloadsuperdex200柱(gehealthcare)上。通过测量280nm的光密度(od)使用基于氨基酸序列计算的摩尔消光系数确定经过纯化的蛋白质样品的蛋白质浓度。通过还原剂存在和缺失下的ce-sds分析来分析分子的纯度和分子量。依照制造商的说明书使用caliperlabchipgxii系统(caliperlifescience)。将2μg样品用于分析。使用tskgelg3000swxl分析型大小排阻柱(tosoh)在25mmk2hpo4,125mmnacl,200mml-精氨酸单盐酸盐,0.02%(w/v)nan3,ph6.7运行缓冲液中于25℃分析抗体样品的聚集体含量。表4a:mcsptcb的生成和纯化汇总。图3显示mcsptcb(2+1crossfab-iggp329glala倒转的)分子的示意图。图4和表4b显示mcsptcb(2+1crossfab-iggp329glala倒转的)分子(seqidno:12,53,54和55)的ce-sds分析。表4b:mcsptcb的ce-sds分析。实施例3包含ch1a1a98/992f1作为抗cea抗体和人源化ch2527作为抗cd3抗体的ceatcb(2+1crossfab-iggp329glala倒转的)的制备将重和轻链dna序列的可变区亚克隆入各自接受者哺乳动物表达载体,与预插入的恒定重链或恒定轻链同框。抗体表达由mpsv启动子驱动且在cds的3’端携带合成的polya信号序列。此外,每种载体包含ebvorip序列。通过使用聚乙烯亚胺(pei)用哺乳动物表达载体共转染hek293ebna细胞生成该分子。用相应表达载体以1:2:1:1的比率(“载体重链fc(穴)”:“载体轻链”:“载体轻链crossfab”:“载体重链fc(节)-fabcrossfab”)转染细胞。为了转染,将hek293ebna细胞在cdcho培养基中无血清悬浮培养。对于500ml摇瓶中的生成,在转染前24小时接种4亿个hek293ebna细胞。为了转染,将细胞以210xg离心5分钟,用预温热的20mlcdcho培养基置换上清液。在20mlcdcho培养基中混合表达载体至最终200μgdna的量。添加540μlpei溶液后,将混合物涡旋15秒,随后于室温温育10分钟。之后将细胞与dna/pei溶液混合,转移至500ml摇瓶并在具有5%co2气氛的温箱中于37℃温育3小时。温育时间后添加160mlf17培养基并将细胞培养24小时。转染后1天添加1mm丙戊酸和7%给料1(lonza)。培养7天后通过以210xg离心15分钟收集上清液用于纯化,将溶液无菌过滤(0.22μm滤器)并添加终浓度0.01%w/v的叠氮化钠,保存于4℃。通过蛋白a亲和层析从细胞培养物上清液纯化分泌的蛋白质。将上清液加载到用40ml20mm磷酸钠,20mm柠檬酸钠,0.5m氯化钠,ph7.5平衡的hitrap蛋白ahp柱(cv=5ml,gehealthcare)上。通过用至少10个柱体积的20mm磷酸钠,20mm柠檬酸钠,0.5m氯化钠,ph7.5清洗去除未结合的蛋白质。靶蛋白在20个柱体积上从20mm柠檬酸钠,0.5m氯化钠,ph7.5至20mm柠檬酸钠,0.5m氯化钠,ph2.5的梯度期间洗脱。通过添加1/10的0.5m磷酸钠,ph8中和蛋白质溶液。浓缩并过滤靶蛋白,之后加载到用ph6.0的20mm组氨酸,140mm氯化钠溶液平衡的hiloadsuperdex200柱(gehealthcare)上。通过测量280nm的光密度(od)使用基于氨基酸序列计算的摩尔消光系数确定经过纯化的蛋白质样品的蛋白质浓度。通过还原剂存在和缺失下的ce-sds分析来分析分子的纯度和分子量。依照制造商的说明书使用caliperlabchipgxii系统(caliperlifescience)。将2μg样品用于分析。使用tskgelg3000swxl分析型大小排阻柱(tosoh)在25mmk2hpo4,125mmnacl,200mml-精氨酸单盐酸盐,0.02%(w/v)nan3,ph6.7运行缓冲液中于25℃分析抗体样品的聚集体含量。表5:ceatcb的生成和纯化汇总。图5显示ceatcb(2+1crossfab-iggp329glala倒转的)分子的示意图。图6和表6显示ceatcb(2+1crossfab-iggp329glala倒转的)分子(seqidno:22,56,57和58)的ce-sds分析。表6:ceatcb的ce-sds分析。峰kda相应的链ceatcb非还原(a)1205.67正确的分子ceatcb还原(b)128.23轻链ch1a1a98/99x2f1236.31轻链ch2527363.48fc(穴)490.9fc(节)在一种备选纯化方法中,通过蛋白a亲和层析(mabselectsure)从收获并澄清的发酵上清液捕捉ceatcb。然后将蛋白a洗脱物进行阳离子交换层析(poros50hs)并随后通过se-hplc和毛细管电泳方式进行分级和分析。合并含产物级分并于室温以结合-洗脱模式进行疏水相互作用层析(butyl-sepharose4ff)。然后通过se-hplc和毛细管电泳方式对由此而来的洗脱物进行分级和分析。合并含产物级分并随后以流过模式进行阴离子交换层析(q-sepharoseff)。使用这种纯化方法获得的材料具有>98%的单体含量。实施例4mcsptcb对mcsp和cd3表达性细胞的结合在mcsp表达性人恶性黑素瘤细胞系(a375)和cd3表达性永生化t淋巴细胞系(jurkat)上测试mcsptcb的结合。简言之,收获细胞,计数,检查生存能力并以2x106个细胞/ml在facs缓冲液(100μlpbs0.1%bsa)中重悬浮。将100μl细胞悬浮液(包含0.2x106个细胞)在圆底96孔板中于4℃与浓度渐增的mcsptcb(2.6pm至200nm)一起温育30分钟,用冷pbs0.1%bsa清洗两次,与pe缀合的affinipuref(ab’)2片段山羊抗人iggfcγ片段特异性二抗(jacksonimmunoresearchlabpe#109-116-170)一起于4℃进一步重温育30分钟,用冷pbs0.1%bsa清洗两次并立即使用facscantoii(softwarefacsdiva)经由门控活的、dapi阴性细胞通过facs进行分析。使用graphpadprism5获得结合曲线(图7小图a,对a375细胞的结合,ec50=3381pm;图7小图b,对jurkat细胞的结合)。实施例5由mcsptcb抗体诱导的t细胞杀伤使用一组表达不同水平的mcsp的肿瘤细胞系(a375=mcsp高,mv-3=mscp中等,hct-116=mcsp低,ls180=mcsp阴性)评估由mcsptcb抗体介导的t细胞杀伤。简言之,用胰蛋白酶/edta收获靶细胞,清洗,并以密度25,000个细胞/孔铺于平底96孔板中。让细胞贴壁过夜。通过将自健康人供体获得的富集淋巴细胞制备物(血沉棕黄层)进行histopaque密度离心来制备外周血单个核细胞(pbmc)。用无菌pbs稀释新鲜血液并在histopaque梯度(sigma,#h8889)上铺层。离心(450xg,30分钟,室温)后,丢弃含pbmc的界面上方的血浆并将pbmc转移至新的falcon管,随后装满50mlpbs。将混合物离心(400xg,10分钟,室温),丢弃上清液,用无菌pbs清洗pbmc团粒两次(离心步骤350xg,10分钟)。对所得pbmc群进行自动计数(vicell)并在含10%fcs和1%l-丙氨酰-l-谷氨酰胺(biochrom,k0302)的rpmi1640培养基中在细胞温箱(37℃,5%co2)中保持直至进一步使用(不超过24小时)。对于杀伤测定法,以标示浓度添加抗体(范围1pm至10nm,一式三份)。以最终效应器对靶(e:t)比率10:1将pbmc添加至靶细胞。于37℃,5%co2温育24小时后通过对由凋亡/坏死细胞释放入细胞上清液中的ldh进行定量(ldh检测试剂盒,rocheappliedscience,#11644793001)来评估靶细胞杀伤。通过将靶细胞与1%tritonx-100一起温育实现对靶细胞的最大裂解(=100%)。最小裂解(=0%)指在没有双特异性构建体的情况下与效应细胞一起共温育的靶细胞。结果显示mcsptcb诱导对mcsp阳性靶细胞系的强的且靶特异性的杀伤,而不杀伤mcsp阴性细胞系(图8,小图a-d)。使用graphpadprism5计算的、与杀伤测定法有关的ec50值见表7。表7:由mcsptcb抗体诱导的对mcsp表达性肿瘤细胞的t细胞介导杀伤的ec50值(pm)。细胞系mcsp受体拷贝数ec50[pm]a37538705812.3mv-32600009.4hct-116367703.7ls180阴性n.d.实施例6由mcsptcb抗体诱导的对mcsp表达性肿瘤细胞的t细胞杀伤后cd8+和cd4+效应细胞上的cd25和cd69上调使用识别t细胞活化标志物cd25(晚期活化标志物)和cd69(早期活化标志物)的抗体通过facs分析评估由mcsptcb抗体介导的对mcsp表达性mv-3肿瘤细胞的t细胞杀伤后cd8+和cd4+t细胞的活化。抗体和杀伤测定法条件基本上如上文所述(实施例5),使用相同的抗体浓度范围(1pm至10nm,一式三份),e:t比率10:1及温育时间24小时。温育后,将pbmc转移至圆底96孔板,以350xg离心5分钟,并用含0.1%bsa的pbs清洗两次。依照供应商的说明书对cd8(fitc抗人cd8,bd#555634),cd4(pecy7抗人cd4,bd#557852),cd69(pe抗人cd69,biolegend#310906)和cd25(apc抗人cd25,bd#555434)实施表面染色。将细胞用150μl/孔含0.1%bsa的pbs清洗两次并使用100μl/孔固定缓冲液(bd#554655)于4℃固定15分钟。离心后,将样品在200μl/孔含dapi的pbs0.1%bsa中重悬浮以便为了facs测定排除死细胞。在bdfacsfortessa上分析样品。结果显示mcsptcb诱导杀伤后cd8+t细胞(图9小图a,b)和cd4+t细胞(图9小图c,d)上活化标志物(cd25,cd69)的强的且靶特异性的上调。实施例7由mcsptcb抗体诱导的对mcsp表达性肿瘤细胞的t细胞杀伤后人效应细胞的细胞因子分泌在杀伤测定法后通过对细胞上清液的facs分析评估由mcsptcb抗体诱导的对mcsp表达性mv-3肿瘤细胞的t细胞杀伤后人pbmc的细胞因子分泌。使用相同的抗体并基本上如上文所述实施杀伤测定法(实施例5和6),使用e:t比率10:1和温育时间24小时。温育时间结束时,将平板以350xg离心5分钟,将上清液转移至新的96孔板中并在-20℃贮存直至后续分析。使用bdcba人可溶性蛋白质flex套组依照制造商的说明书在facscantoii上检测分泌入细胞上清液中的粒酶b、tnfα、ifn-γ、il-2、il-4和il-10。使用下述试剂盒:bdcba人粒酶bflex套组#bd560304;bdcba人tnfflex套组#bd558273;bdcba人ifn-γflex套组#bd558269;bdcba人il-2flex套组#bd558270;bdcba人il-4flex套组#bd558272;bdcba人il-10flex套组#bd558274。结果显示杀伤时mcsptcb诱导il-2、ifn-γ、tnfα、粒酶b和il-10(但无il-4)的分泌(图10,小图a-f)。总的来说,这些实施例显示mcspcd3双特异性抗体·显示优良的对mcsp阳性a375细胞的结合·诱导强的且靶特异性的对mcsp阳性靶细胞系的杀伤,且不杀伤mcsp阴性细胞系·杀伤后诱导cd8+和cd4+t细胞上活化标志物(cd25,cd69)的强的且靶特异性的上调·杀伤时诱导il-2、ifn-γ、tnfα、粒酶b和il-10(非il-4)的分泌。实施例8ceatcb对cea和cd3表达性细胞的结合在已转染的cea表达性肺腺癌细胞(a549-hucea)和cd3表达性永生化人和猕猴t淋巴细胞系(分别为jurkat和hsc-f)上测试ceatcb的结合。使用非靶向性tcb(seqidno:59,60,61和62;见实施例24)作为对照。简言之,收获细胞,计数,检查生存能力并以2x106个细胞/ml在facs缓冲液(100μlpbs0.1%bsa)中重悬浮。将100μl细胞悬浮液(包含0.2x106个细胞)在圆底96孔板中于4℃与浓度渐增的ceatcb(61pm至1000nm)一起温育30分钟,用冷pbs0.1%bsa清洗两次,与fitc缀合的affinipuref(ab’)2片段山羊抗人iggf(ab’)2片段特异性二抗(jacksonimmunoresearchlabfitc#109-096-097)一起于4℃进一步重温育30分钟,用冷pbs0.1%bsa清洗两次,立即使用facscantoii或fortessa(softwarefacsdiva)通过门控活的、pi阴性细胞通过facs进行分析。使用graphpadprism5获得结合曲线(图11小图a,对a549细胞的结合(ec506.6nm);图11小图b,对jurkat细胞的结合;图11小图c,对hsc-f细胞的结合)。实施例9由ceatcb抗体诱导的对cea表达性肿瘤靶细胞的t细胞介导杀伤在hpafii(高cea)、bxpc-3(中等cea)和aspc-1(低cea)人肿瘤细胞上评估由ceatcb抗体诱导的对靶细胞的t细胞介导杀伤。使用hct-116(cea阴性肿瘤细胞系)和非靶向性tcb作为阴性对照。使用人pbmc作为效应器并在与双特异性抗体一起温育后24小时和48小时检测杀伤。简言之,用胰蛋白酶/edta收获靶细胞,清洗,并以密度25,000个细胞/孔铺于平底96孔板中。让细胞贴壁过夜。通过将自健康人供体获得的富集淋巴细胞制备物(血沉棕黄层)进行histopaque密度离心来制备外周血单个核细胞(pbmc)。用无菌pbs稀释新鲜血液并在histopaque梯度(sigma,#h8889)上铺层。离心(450xg,30分钟,室温)后,丢弃含pbmc的界面上方的血浆并将pbmc转移至新的falcon管,随后装满50mlpbs。将混合物离心(400xg,10分钟,室温),丢弃上清液,并用无菌pbs清洗pbmc团粒两次(离心步骤350xg,10分钟)。对所得pbmc群进行自动计数(vicell)并在含10%fcs和1%l-丙氨酰-l-谷氨酰胺(biochrom,k0302)的rpmi1640培养基中在细胞温箱(37℃,5%co2)中保持直至进一步使用(不超过24小时)。对于杀伤测定法,以标示浓度添加抗体(范围6pm至100nm,一式三份)。以最终10:1的e:t比率将pbmc添加至靶细胞。温育24小时和48小时后通过对由凋亡/坏死细胞释放入细胞上清液的ldh(乳酸脱氢酶)定量(ldh检测试剂盒,rocheappliedscience,#11644793001)来评估靶细胞杀伤。通过将靶细胞与1%tritonx-100一起温育实现靶细胞的最大裂解(=100%)。最小裂解(=0%)指在没有双特异性抗体的情况下与效应细胞一起共温育的靶细胞。结果显示ceatcb诱导cea阳性靶细胞的强的且靶特异性的杀伤(图12,小图a-h)。使用graphpadprism5计算的、与杀伤测定法有关的ec50值见表8。表8:cea受体拷贝数和由ceatcb抗体诱导的对cea表达性肿瘤细胞的t细胞介导杀伤的ec50值(pm)。细胞系cea受体拷贝数ec50[pm]48小时hpafii120000-205000667bxpc-3410003785aspc13500-8000846实施例10ceatcb介导的对cea表达性肿瘤靶细胞的杀伤后5天的t细胞增殖和活化在hpafii(高cea)、bxpc-3(中等cea)和aspc-1(低cea)细胞上评估ceatcb介导的对cea表达性肿瘤靶细胞的杀伤后5天检测t细胞增殖和活化。使用hct-116(cea阴性肿瘤细胞系)和非靶向性tcb作为阴性对照。用于增殖测定法的实验条件与实施例9中描述的实验条件相似,但是在96孔平底板中每孔只铺10,000个靶细胞。为了评估t细胞增殖,用cfse(sigma#21888)标记新鲜分离的pbmc。简言之,稀释cfse储液以获得100μm的工作溶液。将90x106个pbmc细胞在90ml预温热的pbs中重悬浮并补充90μlcfse工作溶液。立即混合细胞并于37℃温育15分钟。将10ml预温热的fcs添加至细胞以终止反应。将细胞以400g离心10分钟,在50ml培养基中重悬浮并于37℃温育30分钟。温育后,用温热的培养基清洗细胞一次,计数,在培养基中重悬浮并以10:1的e:t添加至靶细胞进行杀伤测定法和随后的细胞增殖和活化测量。通过对cfse染料稀释的定量评估杀伤5天后cd4和cd8阳性t细胞上的增殖。使用抗人cd25抗体在相同t细胞子集上评估cd25表达。简言之,离心(400xg,4分钟)后,重悬浮细胞,用facs缓冲液清洗,并与25μl稀释的cd4/cd8/cd25抗体混合物一起于4℃温育30分钟(apc/cy7抗人cd4#317418,apc抗人cd8#301014,pe/cy7抗人cd25#302612)。然后清洗细胞三次以去除未结合的抗体,并最终在200μl含碘化丙啶(pi)的facs缓冲液中重悬浮以便为facs测量排除死细胞。使用bdfacscantoii测量荧光。结果显示ceatcb诱导强的且靶特异性的cd8+和cd4+t细胞增殖(图13,小图a-d)及其通过cd25活化标志物上调检测的活化(图13,小图e-h)。实施例11由ceatcb诱导的对cea表达性肿瘤细胞的t细胞介导杀伤后人效应细胞的细胞因子分泌通过杀伤后48小时细胞上清液的facs分析(cba试剂盒)评估由ceatcb诱导的对cea表达性mkn45肿瘤细胞的t细胞介导杀伤后人pbmc的细胞因子分泌。实验条件与实施例9中描述的实验条件相同。温育时间结束时,将板以350xg离心5分钟,将上清液转移入新的96孔板并于-20℃贮存直至后续分析。使用bdcba人可溶性蛋白质flex套组依照制造商的说明书在facscantoii上检测分泌入细胞上清液的(a)ifn-γ、(b)tnfα、(c)粒酶b、(d)il-2、(e)il-6和(f)il-10。使用下述试剂盒:bdcba人il-2bdflex套组#bd558270;bdcba人粒酶bbdflex套组#bd560304;bdcba人tnfflex套组#bd558273;bdcba人ifn-γflex套组#bd558269;bdcba人il-4flex套组#bd558272;bdcba人il-10flex套组#bd558274。结果显示ceatcb介导的杀伤(但是由非靶向性tcb对照介导的杀伤不)诱导ifn-γ、tnfα、粒酶b、il-2、il-6和il-10的分泌(图14,小图a-f)。实施例12在渐增浓度的脱落cea(scea)存在下靶细胞的t细胞介导杀伤在渐增浓度的脱落cea(scea2.5ng/ml至5μg/ml)存在下评估由ceatcb抗体诱导的对cea表达性肿瘤靶细胞(ls180)的t细胞介导杀伤。使用人pbmc作为效应细胞并在与双特异性抗体和scea一起温育后24小时和48小时检测杀伤。简言之,用胰蛋白酶/edta收获靶细胞,清洗,并以密度25,000个细胞/孔铺于平底96孔板中。让细胞贴壁过夜。通过将自健康人供体获得的富集淋巴细胞制备物(血沉棕黄层)进行histopaque密度离心来制备外周血单个核细胞(pbmc)。用无菌pbs稀释新鲜血液并在histopaque梯度(sigma,#h8889)上铺层。离心(450xg,30分钟,室温)后,丢弃含pbmc的界面上方的血浆并将pbmc转移至新的falcon管,随后装满50mlpbs。将混合物离心(400xg,10分钟,室温)后,丢弃上清液,并用无菌pbs清洗pbmc团粒两次(离心步骤350xg,10分钟)。对所得pbmc群进行自动计数(vicell)并在含10%fcs和1%l-丙氨酰-l-谷氨酰胺(biochrom,k0302)的rpmi1640培养基中在细胞温箱(37℃,5%co2)中保持直至进一步使用(不超过24小时)。对于杀伤测定法,以固定浓度1nm使用ceatcb抗体,并以2.5ng至5μg/ml的浓度范围在实验中搀入scea。以最终10:1的e:t比率将pbmc添加至靶细胞。温育24小时和48小时后通过对由凋亡/坏死细胞释放入细胞上清液的ldh(乳酸脱氢酶)定量(ldh检测试剂盒,rocheappliedscience,#11644793001)来评估靶细胞杀伤。通过将靶细胞与1%tritonx-100一起温育实现靶细胞的最大裂解(=100%)。最小裂解(=0%)指在没有双特异性抗体的情况下与效应细胞一起共温育的靶细胞。在scea缺乏下由ceatcb介导的杀伤设置为100%,并将在渐增浓度的scea存在下获得的杀伤对其标准化。结果显示scea对ceatcb介导的对cea表达性靶细胞的杀伤只有较小的影响(图15小图a,b)。上至0.2μg/ml的scea未检测到对t细胞杀伤的影响。高于0.2μg/ml的scea浓度对总体杀伤只有较小的影响(10-50%减少)。实施例13使用人和猕猴pbmc作为效应细胞的对靶细胞的t细胞介导杀伤与ceatcb抗体和作为效应细胞的人pbmc或猕猴pbmc评估一起温育后21小时和40小时评估过表达人cea的a549(肺腺癌)细胞(a549-hcea)的t细胞介导杀伤。简言之,用胰蛋白酶/edta收获靶细胞,清洗,并以密度25,000个细胞/孔铺于平底96孔板中。让细胞贴壁数小时。通过将自健康人供体或健康猕猴获得的富集淋巴细胞制备物(血沉棕黄层)进行histopaque密度离心来制备外周血单个核细胞(pbmc)。对于后者,使用90%histopaque-pbs密度梯度。用无菌pbs稀释新鲜血液并在histopaque梯度(sigma,#h8889)上铺层。离心(对于人pbmc是450xg,30分钟,室温,对于猕猴pbmc是相应的520xg,30分钟,室温)后,丢弃含pbmc的界面上方的血浆并将pbmc转移至新的falcon管,随后装满50mlpbs。将混合物离心(400xg,10分钟,室温),丢弃上清液,并用无菌pbs清洗pbmc团粒两次(离心步骤350xg,10分钟)。为了制备猕猴pbmc,实施另外的低速离心步骤(150xg,15分钟)。对所得pbmc群进行自动计数(vicell)并在含10%fcs和1%l-丙氨酰-l-谷氨酰胺(biochrom,k0302)的rpmi1640培养基中在细胞温箱(37℃,5%co2)中保持直至进一步使用(上至4小时)。对于杀伤测定法,以标示浓度添加抗体(范围6pm至100nm,一式三份)。以最终10:1的e:t比率将pbmc添加至靶细胞。温育21小时和40小时后通过对由凋亡/坏死细胞释放入细胞上清液的ldh(乳酸脱氢酶)定量(ldh检测试剂盒,rocheappliedscience,#11644793001)来评估靶细胞杀伤。通过将靶细胞与1%tritonx-100一起温育实现靶细胞的最大裂解(=100%)。最小裂解(=0%)指在没有双特异性抗体的情况下与效应细胞一起共温育的靶细胞。结果显示使用人(图16,小图a,c)和猕猴(图16,小图b,d)效应细胞(pbmc)二者ceatcb均介导对cea阳性靶细胞的靶特异性杀伤。使用graphpadprism5计算的、与40小时杀伤有关的ec50值对于人pbmc是306pm,而对于猕猴pbmc是102pm。实施例14由ceatcb抗体诱导的对cea表达性人结肠直肠癌细胞系的t细胞介导杀伤与人pbmc和0.8nm,4nm和20nmceatcb抗体一起温育后48小时评估对cea表达性人结肠直肠癌细胞系的t细胞介导杀伤。简言之,从自单一健康供体获得的白细胞椎体分离pbmc。用pbs稀释细胞(1:10)并在50mlfalcon管中在lymphoprep上铺层。离心(1800rpm,25分钟)后,从界面取出pbmc层并用pbs清洗4次。对pbmc计数,在速控冷冻条件下以40x106个细胞/ml冷冻于含10%dmso的fcs中并贮存于液氮中直至进一步使用。为了t细胞杀伤测定法,将来自冻存贮液的肿瘤细胞直接铺于96孔板中。快速温热细胞并立即转移入预温热的培养基中,离心,并重悬浮于完全培养基(dmem、iscoves或rpmi-1640,均补充有10%fcs和1%青霉素/链霉素)中,然后以2.5x104个细胞/孔的密度铺板。然后将平板于37℃在增湿的10%co2温箱中温育并在次日用含1%谷氨酰胺的100μlrpmi2%fcs和50μlceatcb(终浓度范围从6.4至20000pm,1:5滴定步骤,对于每个条件在一式两份孔中)置换培养基。将新鲜融化的pbmc用于测定法(从测定开始的2小时内从冻存管融化)并在每孔中添加50μl(3x105)以给出10:1的效应器:靶(e:t)比率。将tritonx100(4%,50μl)添加至150μl靶细胞以获得最大释放值。将平板于37℃温育48小时并使用乳糖脱氢酶细胞毒性检测试剂盒(roche)依照制造商的说明书测定杀伤活性。将特异性细胞裂解的百分率计算为[样品释放-自发释放]/[最大释放-自发释放]x100。图17,小图a-c显示cea表达(使用qifikit定量受体拷贝数,见下文)和对31种结肠直肠癌细胞系的%杀伤(列于x轴上)之间的相关性。图17,小图d显示cea表达和20nmceatcb的%特异性裂解之间的相关性(spearman相关性=0.7289,p<0.0001,n=31),指示展示高cea受体拷贝数(>50000)的肿瘤细胞被ceatcb有效裂解而展示低cea受体拷贝数(<10000)的一簇细胞在相同实验条件下不被ceatcb裂解。图17,小图e显示cea表达和ceatcb的ec50之间的相关性。尽管相关性不是统计上显著的(spearman相关性=-0.3994,p=0.1006,r2=0.1358),该图清楚地显示了在表达高cea受体拷贝数的肿瘤细胞系上较好的ceatcb效力(即较低ec50值)的样式。为了分析癌细胞系上的cea表面表达,使用qifikit(dakocytomation,glostrup,denmark)校准荧光信号并测定每个细胞的结合位点数目。将细胞与小鼠抗人ceacam5单克隆抗体(对于5x105个细胞是0.5μg,克隆:ci-p83-1,sc-23928,santacruz)一起在冰上温育30分钟,用pbs1x-bsa0.1%清洗两次,接着与随qifikit提供的多克隆异硫氰酸荧光素缀合的山羊抗小鼠抗体一起温育45分钟。使用4’,6-二脒基-2-苯基吲哚(dapi)染色从分析中排除死细胞。在cyantmadp分析仪(beckmancoulter)上分析样品。使用summit4.3软件进行数据分析后获得所有均值荧光强度(mfi)。使用这些mfi利用自校准曲线获得的方程式(qifikit校准珠)测定细胞系上抗体结合位点的相对数目(在结果上称为cea拷贝数)。从冷冻管接种用于t细胞杀伤测定法和cea表面表达定量的结肠直肠癌细胞系。用于维持冷冻储液的方法如brachtetal.(2010)brjcancer103,340-346中所述。实施例15ceatcb在共移植人pbmc的ls174t-fluc2人结肠癌(e:t比率5:1)中的体内抗肿瘤功效对nog(nod/shi-scid/il-2rγnull)小鼠(n=12)皮下注射与人pbmc预混合的1x106个ls174t-fluc2细胞(e:t比率5:1,于pbs中,总体积100μl)。已改造ls174t-fluc2细胞以表达萤光素酶,它容许以非侵入且高度灵敏的方式通过生物发光(bli)监测肿瘤进展。为了评估早期和延迟处理效果,小鼠在皮下共移植肿瘤细胞/pbmc后第1天(早期处理)或第7天(延迟处理)开始两周一次接受静脉内注射0.5或2.5mg/kgceatcb。作为对照,一组小鼠两周一次接受静脉内注射2.5mg/kg与ceatcb具有相同样式的对照tcb(在此情况中mcsptcb充当非靶向对照,因为ls174t-fluc2细胞不表达mcsp),还有一个额外的对照组在第1天开始只接受pbs(媒介)。每周一次用数字测径器测量肿瘤体积。此外,每周一次对小鼠腹膜内注射d-萤光素并用ivisspectrum(perkinelmer)测量活肿瘤细胞的生物发光光发射。施用处理直至肿瘤细胞接种后19天,对应于研究终止的日子。实验结果显示于图18小图a-d。结果显示不同研究组((a,b)早期处理,(c,d)延迟处理)中通过测径器(a和c)和通过生物发光(总通量,b和d)测量的12只小鼠肿瘤体积的平均值和sem。实施例16ceatcb在共移植人pbmc的ls174t-fluc2人结肠癌(e:t比率1:1)中的体内抗肿瘤功效对nog(nod/shi-scid/il-2rγnull)小鼠(n=10)皮下注射与人pbmc预混合的1x106个ls174t-fluc2细胞(见实施例15)(e:t比率1:1,于pbs中,总体积100μl)。为了评估早期和延迟处理效果,小鼠在肿瘤细胞接种后第1天(早期处理)或第7天(延迟处理)开始两周一次接受静脉内注射2.5mg/kgceatcb。作为对照,一组小鼠两周一次接受静脉内注射2.5mg/kgmcsptcb(也见实施例15),还有一个额外的对照组在第1天开始只接受pbs(媒介)。每周一次用数字测径器测量肿瘤体积。此外,每周一次对小鼠腹膜内注射d-萤光素并用ivisspectrum(perkinelmer)测量活肿瘤细胞的生物发光光发射。施用处理直至肿瘤细胞接种后23天,这对应于研究终止的日子。实验结果显示于图19中。结果显示不同研究组(n=10)中通过测径器(a)以及通过生物发光(b)测量的肿瘤体积的平均值和sem。实施例17鼠源化ceatcb在免疫活性hucd3ε/hucea转基因小鼠中的panco2-hucea同位肿瘤模型中的体内功效hucd3ε/hucea转基因小鼠(n=10)接受胰腺内注射总体积10μl于pbs中的2x105个panco2-hucea细胞。由于鼠细胞不表达cea,因此改造鼠胰腺癌细胞系panco2以过表达人cea作为ceatcb的靶抗原。小鼠每周两次静脉内注射0.5mg/kg鼠源化ceatcb或作为对照组的pbs(媒介)并监测存活。每天掌控动物的临床症状并检测不利作用。对动物的终止标准是可见的病态:不整洁的毛,弓背,呼吸问题,运动受损。总体存活结果显示于图20中。结果显示每个时间点存活动物的百分比。使用成对studentt检验比较处理组对pbs对照组的显著性(p=0.078)。实施例18通过表面等离振子共振(spr)测定的ceatcb对cea和cd3的亲和力在biacoret100上于25℃以hbs-ep作为运行缓冲液(0.01mhepesph7.4,0.15mnacl,3mmedta,0.005%表面活性剂p20,biacore,freiburg/germany)实施表面等离振子共振(spr)实验。为了亲和力测量,将ceatcb捕捉到带固定化抗人fab(gehealthcare#28-9583-25)的cm5传感器芯片表面上。通过使用标准胺偶联试剂盒(biacore,freiburg/germany)于ph5.0以约10,000个共振单位(ru)的直接固定化将捕捉用igg偶联至传感器芯片表面。为了分析与人cd3εstalk-fc(节)-avi/cd3δ-stalk-fc(穴)(分别为seqidno120和121)的相互作用,以10μl/min在50nm捕捉ceatcb30秒。cd3ε/cd3δ以浓度0.68-500nm和流速30μl/min通过流通池360秒。监测解离360秒。通过以10μl/min捕捉tcb分子40秒测定ceatcb和重组肿瘤靶抗原人naba-avi-his(包含被人ceacam1的n、a1和a2域围绕的人cea的b3域(ceacam5),具有c-末端avi6his标签;见seqidno:119)之间的相互作用的kd值。抗原以浓度范围0.68至500nm和流速30μl/min流过流通池240秒。测量解离240秒。通过扣除参照流通池上获得的响应校正批量折射指数差异。在此,抗原流经带固定化抗人fab抗体的表面,但其上已注射hbs-ep而非cea。使用biacoret200评估软件(vaa,biacoreab,uppsala/sweden)通过数值积分拟合1:1langmuir结合的速率方程来推导动力学常数。用下述公式计算相互作用的半衰期(t1/2):t1/2=ln2/koff。ceatcb以对于人naba为62nm而对于人cd3ε/cd3δ为75.3nm的kd值结合nm范围中的肿瘤靶标和cd3ε/cd3δ。对naba的单价结合的半衰期是5.3分钟,对cd3ε/cd3δ的结合的半衰期是5.7分钟。动力学数值汇总于表9中。表9:ceatcb对人naba和人cd3ε/cd3δ的亲和力(t=25℃)。抗原tcbkon[1/ms]koff[1/s]t1/2[min]kd[nm]人nabaceatcb3.49x1042.18x10-35.362.4人cd3ε/cd3δceatcb2.69x1042.03x10-35.775.3实施例19通过表面等离振子共振(spr)测定的mscptcb对mcsp和cd3的亲和力在biacoret100上于25℃以hbs-ep作为运行缓冲液(0.01mhepesph7.4,0.15mnacl,3mmedta,0.005%表面活性剂p20,biacore,freiburg/germany)实施表面等离振子共振(spr)实验。为了亲和力测量,将mcsptcb捕捉到带固定化抗人fab(gehealthcare#28-9583-25)的cm5传感器芯片表面上。通过使用标准胺偶联试剂盒(biacore,freiburg/germany)于ph5.0以约7,500个共振单位(ru)的直接固定化将捕捉用igg偶联至传感器芯片表面。以10μl/分钟在30nm捕捉mcsptcb60秒。在90秒里使人和猕猴mcspd3(分别见seqidno118和117)以浓度0.024-50nm和流速30μl/min通过流通池。人和猕猴cd3εstalk-fc(节)-avi/cd3δ-stalk-fc(穴)的浓度范围为1.17-600nm。由于预期与鼠mcspd3(seqidno:122)的相互作用是弱的,所以挑选此抗原的浓度范围在3.9和500nm之间。对所有相互作用监测解离120秒。通过扣除参照流通池上获得的响应校正批量折射指数差异。在此,抗原流经带固定化抗人fab抗体的表面,但其上已注射hbs-ep而非mcsptcb。使用biacoret200评估软件(vaa,biacoreab,uppsala/sweden)通过数值积分拟合1:1langmuir结合的速率方程来推导动力学常数。在稳态中测定mcsptcb与鼠mcspd3的相互作用。用下述公式计算相互作用的半衰期(t1/2):t1/2=ln2/koff。mcsptcb以对于人抗原为0.15nm而对于猕猴抗原为0.12nm的kd值结合pm范围中的肿瘤靶标。重组cd3ε/cd3δ以78nm(人)和104nm(猕猴)的kd值受到mcsptcb结合。对于肿瘤靶标的单价结合的半衰期高达260分钟,对于cd3e/cd3d是2.9分钟。经由亲和力成熟,mcsp抗体获得一些对重组鼠mcspd3的结合。此相互作用的kd值在mm范围中(1.6mm)。动力学数值汇总于表10中。表10:mcsptcb对人、猕猴和鼠mcspd3及人和猕猴cd3ε/cd3δ的亲和力(t=25℃)。kon[1/ms]koff[1/s]t1/2[min]kd[nm]人mcspd33.89x1055.63x10-52050.15猕猴mcspd33.70x1054.39x10-52630.12鼠mcspd3ndndnd1570*人cd3ε/cd3δ4.99x1043.92x10-32.978.7猕猴cd3ε/cd3δ4.61x1044.78x10-32.4104*通过稳态测量确定实施例20ceatcb的热稳定性通过动态光散射(dynamiclightscattering,dls)监测ceatcb的热稳定性。将蛋白质浓度0.5mg/ml的经过过滤的蛋白质样品30μg一式两份施加至dynapro平板读数仪(wyatttechnologycorporation;usa)上。温度以0.05℃/min自25至75℃攀升,伴随着收集径向和总散射强度。结果显示于图21中。于55℃测量ceatcb的聚集温度。实施例21mcsptcb的热稳定性通过动态光散射(dynamiclightscattering,dls)监测mcsptcb的热稳定性。将蛋白质浓度0.5mg/ml的经过过滤的蛋白质样品30μg一式两份施加至dynapro平板读数仪(wyatttechnologycorporation;usa)上。温度以0.05℃/min自25至75℃攀升,伴随收集径向和总散射强度。结果显示于图22中。于55℃测量mcsptcb的聚集温度。实施例22由mcsptcb和mcsp1+1crossmab抗体诱导的对mcsp表达性肿瘤靶细胞的t细胞介导杀伤在a375(高mcsp)、mv-3(中等mcsp)和hct-116(低mcsp)肿瘤靶细胞上评估由mcsptcb和mcsp1+1crossmabtcb(具有与mcsptcb相同的cd3和mcsp结合序列的t细胞活化性双特异性抗体,具有图1小图d中所示分子样式)抗体诱导的对靶细胞的t细胞介导杀伤。ls180(mcsp阴性肿瘤细胞系)用作阴性对照。将靶细胞与抗体和效应细胞(人pbmc)一起温育24小时和48小时后评估肿瘤细胞杀伤。简言之,用胰蛋白酶/edta收获靶细胞,清洗,并以密度25,000个细胞/孔铺于平底96孔板中。让细胞贴壁过夜。通过将自健康人供体获得的富集淋巴细胞制备物(血沉棕黄层)进行histopaque密度离心来制备外周血单个核细胞(pbmc)。用无菌pbs稀释新鲜血液并在histopaque梯度(sigma,#h8889)上铺层。离心(450xg,30分钟,室温)后,丢弃含pbmc的界面上方的血浆并将pbmc转移至新的falcon管,随后装满50mlpbs。将混合物离心(400xg,10分钟,室温),丢弃上清液,并用无菌pbs清洗pbmc团粒两次(离心步骤350xg,10分钟)。对所得pbmc群进行自动计数(vicell)并在含10%fcs和1%l-丙氨酰-l-谷氨酰胺(biochrom,k0302)的rpmi1640培养基中在细胞温箱(37℃,5%co2)中保持直至进一步使用(不超过24小时)。对于杀伤测定法,以标示浓度添加抗体(范围0.01pm至10nm,一式三份)。以最终10:1的e:t比率将pbmc添加至靶细胞。温育24小时和48小时后通过对由凋亡/坏死细胞释放入细胞上清液的ldh(乳酸脱氢酶)定量(ldh检测试剂盒,rocheappliedscience,#11644793001)来评估靶细胞杀伤。通过将靶细胞与1%tritonx-100一起温育实现靶细胞的最大裂解(=100%)。最小裂解(=0%)指在没有双特异性抗体的情况下与效应细胞共温育的靶细胞。结果显示mcsptcb抗体比mcsp1+1crossmabtcb更有效力,因为它在两个时间点以及在所有肿瘤靶细胞上均诱导更强的对mcsp阳性靶细胞的杀伤(图23小图a-h)。使用graphpadprism5计算的、与杀伤测定法有关的ec50值见表11。表11:mcsp受体拷贝数和由mcsptcb抗体诱导的对mcsp表达性肿瘤细胞的t细胞介导杀伤的ec50值(pm)(n.d.=未测定)。细胞系mcsp受体拷贝数ec50[pm]24小时ec50[pm]48小时a3753870580.1n.d.mv-32600001.00.7hct-11636770~6.2e-008~0.09ls180阴性~764n.d.实施例23由mcsptcb和mcsp1+1crossmab抗体诱导的对mcsp表达性肿瘤细胞的t细胞介导杀伤后cd8+和cd4+效应细胞上的cd25和cd69上调使用识别t细胞活化标志物cd25(晚期活化标志物)和cd69(早期活化标志物)的抗体通过facs分析评估由mcsptcb和mcsp1+1crossmab抗体介导的对mcsp表达性肿瘤细胞(a375和mv-3)的t细胞杀伤后cd8+和cd4+t细胞的活化。抗体和杀伤测定法条件基本上如上文所述(实施例22),使用相同的抗体浓度范围(0.01pm至10nm,一式三份),e:t比率10:1及温育时间48小时。温育后,将pbmc转移至圆底96孔板,以350xg离心5分钟,并用含0.1%bsa的pbs清洗两次。依照供应商的说明书实施cd8(fitc抗人cd8,bd#555634)、cd4(pecy7抗人cd4,bd#557852)、cd69(pe抗人cd69,biolegend#310906)和cd25(apc抗人cd25,bd#555434)的表面染色。将细胞用150μl/孔含0.1%bsa的pbs清洗两次并使用100μl/孔固定缓冲液(bd#554655)于4℃固定15分钟。离心后,将样品重悬浮于200μl/孔含dapi的pbs0.1%bsa以便为facs测量排除死细胞。在bdfacsfortessa上分析样品。结果显示mcsptcb诱导杀伤后cd8+t细胞(图24小图a,b(对于a375细胞)和e,f(对于mv-3细胞))和cd4+t细胞(图24小图c,d(对于a375细胞)和g,h(对于mv-3细胞))上活化标志物(cd25,cd69)的强的且靶特异性的上调。就杀伤结果而言,用mcsptcb活化t细胞强于用mcsp1+1crossmab。实施例24包含dp47gs作为非结合抗体和人源化ch2527作为抗cd3抗体的dp47gstcb(2+1crossfab-iggp329glala倒转的=“非靶向tcb”)的制备“非靶向tcb”在以上实验中用作对照。双特异性抗体针对cd3ε但不结合任何其它抗原,并因此不能将t细胞与任何靶细胞交联(且随后不能诱导任何杀伤)。因此将其用作测定法中的阴性对照以监测任何非特异性t细胞活化。将重和轻链dna序列的可变区亚克隆入各自接受者哺乳动物表达载体,与预插入的恒定重链或恒定轻链同框。抗体表达由mpsv启动子驱动且在cds的3’端携带合成的polya信号序列。此外,每种载体包含ebvorip序列。通过使用聚乙烯亚胺(pei)用哺乳动物表达载体共转染hek293ebna细胞生成该分子。用相应表达载体以1:2:1:1的比率(“载体重链fc(穴)”:“载体轻链”:“载体轻链crossfab”:“载体重链fc(节)-fabcrossfab”)转染细胞。为了转染,在cdcho培养基中无血清悬浮培养hek293ebna细胞。为了500ml摇瓶中的生产,在转染前24小时接种4亿个hek293ebna细胞。为了转染,将细胞以210xg离心5分钟,用预温热的20mlcdcho培养基置换上清液。在20mlcdcho培养基中混合表达载体至最终200μgdna的量。添加540μlpei溶液后,将混合物涡旋15秒,随后于室温温育10分钟。之后将细胞与dna/pei溶液混合,转移至500ml摇瓶并在5%co2气氛的温箱中于37℃温育3小时。温育时间后添加160mlf17培养基并将细胞培养24小时。转染后1天添加1mm丙戊酸和7%给料1(lonza)。培养7天后通过以210xg离心15分钟收集上清液用于纯化,将溶液无菌过滤(0.22μm滤器)并以终浓度0.01%w/v添加叠氮化钠,保存于4℃。通过蛋白a亲和层析自细胞培养物上清液纯化分泌的蛋白质。将上清液加载到用40ml20mm磷酸钠,20mm柠檬酸钠,0.5m氯化钠,ph7.5平衡的hitrap蛋白ahp柱(cv=5ml,gehealthcare)上。通过用至少10个柱体积的20mm磷酸钠,20mm柠檬酸钠,0.5m氯化钠,ph7.5清洗去除未结合的蛋白质。靶蛋白在20个柱体积上从20mm柠檬酸钠,0.5m氯化钠,ph7.5至20mm柠檬酸钠,0.5m氯化钠,ph2.5的梯度期间洗脱。通过添加1/10的0.5m磷酸钠,ph8中和蛋白质溶液。浓缩并过滤靶蛋白,之后加载到用ph6.0的20mm组氨酸,140mm氯化钠溶液平衡的hiloadsuperdex200柱(gehealthcare)上。通过测量280nm的光密度(od)使用基于氨基酸序列计算的摩尔消光系数确定经过纯化的蛋白质样品的蛋白质浓度。通过还原剂存在和缺失下的ce-sds分析来分析分子的纯度和分子量。依照制造商的说明书使用caliperlabchipgxii系统(caliperlifescience)。使用2μg样品进行分析。使用tskgelg3000swxl分析型大小排阻柱(tosoh)在25mmk2hpo4,125mmnacl,200mml-精氨酸单盐酸盐,0.02%(w/v)nan3,ph6.7运行缓冲液中于25℃分析抗体样品的聚集体含量。表12:dp47gstcb的产生和纯化汇总。图25和表13显示包含dp47gs作为非结合抗体和人源化ch2527作为抗cd3抗体的dp47gstcb(2+1crossfab-iggp329glala倒转的)(seqidno:59,60,61和62)的ce-sds分析。表13:dp47gstcb的ce-sds分析。峰kda相应的链dp47gstcb非还原(a)1165.22具有2条丢失轻链的分子2181.35具有1条丢失轻链的分子3190.58无n-连接糖基化的正确分子4198.98正确分子dp47gstcb还原(b)127.86轻链dp47gs235.74轻链huch2527363.57fc(穴)493.02fc(节)***尽管为了理解清楚的目的已经通过例示和实施例较为详细地描述了前述发明,但是该描述和实施例不应解释为限制本发明的范围。通过提述明确完整收录本文中引用的所有专利和科学文献的公开内容。序列表<110>罗切格利卡特公司(rocheglycartag)<120>双特异性t细胞活化性抗原结合分子<130>31447<150>ep13156686.1<151>2013-02-26<160>122<170>patentinversion3.5<210>1<211>455<212>prt<213>人工序列<220><223>hccd3ch2527(vh_3-23(12))<400>1gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrpheserthrtyr202530alametasntrpvalargglnalaproglylysglyleuglutrpval354045serargileargserlystyrasnasntyralathrtyrtyralaasp505560servallysglyargphethrileserargaspaspserlysasnthr65707580leutyrleuglnmetasnserleuargalagluaspthralavaltyr859095tyrcysvalarghisglyasnpheglyasnsertyrvalsertrpphe100105110alatyrtrpglyglnglythrleuvalthrvalserseralaserthr115120125lysglyproservalpheproleualaproserserlysserthrser130135140glyglythralaalaleuglycysleuvallysasptyrpheproglu145150155160provalthrvalsertrpasnserglyalaleuthrserglyvalhis165170175thrpheproalavalleuglnserserglyleutyrserleuserser180185190valvalthrvalproserserserleuglythrglnthrtyrilecys195200205asnvalasnhislysproserasnthrlysvalasplyslysvalglu210215220prolyssercysasplysthrhisthrcysproprocysproalapro225230235240gluleuleuglyglyproservalpheleupheproprolysprolys245250255aspthrleumetileserargthrprogluvalthrcysvalvalval260265270aspvalserhisgluaspprogluvallyspheasntrptyrvalasp275280285glyvalgluvalhisasnalalysthrlysproargglugluglntyr290295300asnserthrtyrargvalvalservalleuthrvalleuhisglnasp305310315320trpleuasnglylysglutyrlyscyslysvalserasnlysalaleu325330335proalaproileglulysthrileserlysalalysglyglnproarg340345350gluproglnvaltyrthrleuproproserargaspgluleuthrlys355360365asnglnvalserleuthrcysleuvallysglyphetyrproserasp370375380ilealavalglutrpgluserasnglyglnprogluasnasntyrlys385390395400thrthrproprovalleuaspseraspglyserphepheleutyrser405410415lysleuthrvalasplysserargtrpglnglnglyasnvalpheser420425430cysservalmethisglualaleuhisasnhistyrthrglnlysser435440445leuserleuserproglylys450455<210>2<211>215<212>prt<213>人工序列<220><223>lccd3ch2527(vl_7-46(13))<400>2glnalavalvalthrglngluproserleuthrvalserproglygly151015thrvalthrleuthrcysglyserserthrglyalavalthrthrser202530asntyralaasntrpvalglnglulysproglyglnalaphearggly354045leuileglyglythrasnlysargalaproglythrproalaargphe505560serglyserleuleuglyglylysalaalaleuthrleuserglyala65707580glnprogluaspglualaglutyrtyrcysalaleutrptyrserasn859095leutrpvalpheglyglyglythrlysleuthrvalleuglyglnpro100105110lysalaalaproservalthrleupheproproserserglugluleu115120125glnalaasnlysalathrleuvalcysleuileseraspphetyrpro130135140glyalavalthrvalalatrplysalaaspserserprovallysala145150155160glyvalgluthrthrthrproserlysglnserasnasnlystyrala165170175alasersertyrleuserleuthrprogluglntrplysserhisarg180185190sertyrsercysglnvalthrhisgluglyserthrvalglulysthr195200205valalaprothrglucysser210215<210>3<211>125<212>prt<213>人工序列<220><223>vhcd3ch2527(vh_3-23(12))<400>3gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrpheserthrtyr202530alametasntrpvalargglnalaproglylysglyleuglutrpval354045serargileargserlystyrasnasntyralathrtyrtyralaasp505560servallysglyargphethrileserargaspaspserlysasnthr65707580leutyrleuglnmetasnserleuargalagluaspthralavaltyr859095tyrcysvalarghisglyasnpheglyasnsertyrvalsertrpphe100105110alatyrtrpglyglnglythrleuvalthrvalserser115120125<210>4<211>5<212>prt<213>人工序列<220><223>hcdr1cd3ch2527(vh_3-23(12))<400>4thrtyralametasn15<210>5<211>19<212>prt<213>人工序列<220><223>hcdr2cd3ch2527(vh_3-23(12))<400>5argileargserlystyrasnasntyralathrtyrtyralaaspser151015vallysgly<210>6<211>14<212>prt<213>人工序列<220><223>hcdr3cd3ch2527(vh_3-23(12))<400>6hisglyasnpheglyasnsertyrvalsertrpphealatyr1510<210>7<211>109<212>prt<213>人工序列<220><223>vlcd3ch2527(vl_7-46(13))<400>7glnalavalvalthrglngluproserleuthrvalserproglygly151015thrvalthrleuthrcysglyserserthrglyalavalthrthrser202530asntyralaasntrpvalglnglulysproglyglnalaphearggly354045leuileglyglythrasnlysargalaproglythrproalaargphe505560serglyserleuleuglyglylysalaalaleuthrleuserglyala65707580glnprogluaspglualaglutyrtyrcysalaleutrptyrserasn859095leutrpvalpheglyglyglythrlysleuthrvalleu100105<210>8<211>14<212>prt<213>人工序列<220><223>lcdr1cd3ch2527(vl_7-46(13))<400>8glyserserthrglyalavalthrthrserasntyralaasn1510<210>9<211>7<212>prt<213>人工序列<220><223>lcdr2cd3ch2527(vl_7-46(13))<400>9glythrasnlysargalapro15<210>10<211>9<212>prt<213>人工序列<220><223>lcdr3cd3ch2527(vl_7-46(13))<400>10alaleutrptyrserasnleutrpval15<210>11<211>442<212>prt<213>人工序列<220><223>hcmcspm4-3(c1)$<400>11glnvalglnleuglngluserglyproglyleuvallysprosergln151015thrleuserleuthrcysthrvalserglyglyserilethrsergly202530tyrtyrtrpasntrpileargglnhisproglylysglyleuglutrp354045ileglytyrilethrpheaspglyserasnasntyrasnproserleu505560lysserargvalthrileserargaspthrserlysasnglnpheser65707580leulysleuserservalthralaalaaspthralavaltyrtyrcys859095alaasppheasptyrtrpglyglnglythrleuvalthrvalserser100105110alaserthrlysglyproservalpheproleualaproserserlys115120125serthrserglyglythralaalaleuglycysleuvallysasptyr130135140pheprogluprovalthrvalsertrpasnserglyalaleuthrser145150155160glyvalhisthrpheproalavalleuglnserserglyleutyrser165170175leuserservalvalthrvalproserserserleuglythrglnthr180185190tyrilecysasnvalasnhislysproserasnthrlysvalasplys195200205lysvalgluprolyssercysasplysthrhisthrcysproprocys210215220proalaprogluleuleuglyglyproservalpheleuphepropro225230235240lysprolysaspthrleumetileserargthrprogluvalthrcys245250255valvalvalaspvalserhisgluaspprogluvallyspheasntrp260265270tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu275280285gluglntyrasnserthrtyrargvalvalservalleuthrvalleu290295300hisglnasptrpleuasnglylysglutyrlyscyslysvalserasn305310315320lysalaleuproalaproileglulysthrileserlysalalysgly325330335glnproarggluproglnvaltyrthrleuproproserargaspglu340345350leuthrlysasnglnvalserleuthrcysleuvallysglyphetyr355360365proseraspilealavalglutrpgluserasnglyglnprogluasn370375380asntyrlysthrthrproprovalleuaspseraspglyserphephe385390395400leutyrserlysleuthrvalasplysserargtrpglnglnglyasn405410415valphesercysservalmethisglualaleuhisasnhistyrthr420425430glnlysserleuserleuserproglylys435440<210>12<211>214<212>prt<213>人工序列<220><223>lcmcspml2(g3)<400>12aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnglyileargasntyr202530leuasntrptyrglnglnlysproglylysalaprolysleuleuile354045tyrtyrthrserserleuhisserglyvalproserargphesergly505560serglyserglythrasptyrthrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysglnglntyrseralaleuprotrp859095thrpheglyglnglythrlysvalgluilelysargthrvalalaala100105110proservalpheilepheproproseraspgluglnleulyssergly115120125thralaservalvalcysleuleuasnasnphetyrproarggluala130135140lysvalglntrplysvalaspasnalaleuglnserglyasnsergln145150155160gluservalthrgluglnaspserlysaspserthrtyrserleuser165170175serthrleuthrleuserlysalaasptyrglulyshislysvaltyr180185190alacysgluvalthrhisglnglyleuserserprovalthrlysser195200205pheasnargglyglucys210<210>13<211>112<212>prt<213>人工序列<220><223>vhmcspm4-3(c1)<400>13glnvalglnleuglngluserglyproglyleuvallysprosergln151015thrleuserleuthrcysthrvalserglyglyserilethrsergly202530tyrtyrtrpasntrpileargglnhisproglylysglyleuglutrp354045ileglytyrilethrpheaspglyserasnasntyrasnproserleu505560lysserargvalthrileserargaspthrserlysasnglnpheser65707580leulysleuserservalthralaalaaspthralavaltyrtyrcys859095alaasppheasptyrtrpglyglnglythrleuvalthrvalserser100105110<210>14<211>6<212>prt<213>人工序列<220><223>hcdr1mcspm4-3(c1)<400>14serglytyrtyrtrpasn15<210>15<211>16<212>prt<213>人工序列<220><223>hcdr2mcspm4-3(c1)<400>15tyrilethrpheaspglyserasnasntyrasnproserleulysser151015<210>16<211>3<212>prt<213>人工序列<220><223>hcdr3mcspm4-3(c1)<400>16pheasptyr1<210>17<211>107<212>prt<213>人工序列<220><223>vlmcspml2(g3)<400>17aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnglyileargasntyr202530leuasntrptyrglnglnlysproglylysalaprolysleuleuile354045tyrtyrthrserserleuhisserglyvalproserargphesergly505560serglyserglythrasptyrthrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysglnglntyrseralaleuprotrp859095thrpheglyglnglythrlysvalgluilelys100105<210>18<211>11<212>prt<213>人工序列<220><223>lcdr1mcspml2(g3)<400>18argalaserglnglyileargasntyrleuasn1510<210>19<211>7<212>prt<213>人工序列<220><223>lcdr2mcspml2(g3)<400>19tyrthrserserleuhisser15<210>20<211>9<212>prt<213>人工序列<220><223>lcdr3mcspml2(g3)<400>20glnglntyrseralaleuprotrpthr15<210>21<211>451<212>prt<213>人工序列<220><223>hcceach1a1a98-99<400>21glnvalglnleuvalglnserglyalagluvallyslysproglyala151015servallysvalsercyslysalaserglytyrthrphethrgluphe202530glymetasntrpvalargglnalaproglyglnglyleuglutrpmet354045glytrpileasnthrlysthrglyglualathrtyrvalglugluphe505560lysglyargvalthrphethrthraspthrserthrserthralatyr65707580metgluleuargserleuargseraspaspthralavaltyrtyrcys859095alaargtrpaspphealatyrtyrvalglualametasptyrtrpgly100105110glnglythrthrvalthrvalserseralaserthrlysglyproser115120125valpheproleualaproserserlysserthrserglyglythrala130135140alaleuglycysleuvallysasptyrpheprogluprovalthrval145150155160sertrpasnserglyalaleuthrserglyvalhisthrpheproala165170175valleuglnserserglyleutyrserleuserservalvalthrval180185190proserserserleuglythrglnthrtyrilecysasnvalasnhis195200205lysproserasnthrlysvalasplyslysvalgluprolyssercys210215220asplysthrhisthrcysproprocysproalaprogluleuleugly225230235240glyproservalpheleupheproprolysprolysaspthrleumet245250255ileserargthrprogluvalthrcysvalvalvalaspvalserhis260265270gluaspprogluvallyspheasntrptyrvalaspglyvalgluval275280285hisasnalalysthrlysproargglugluglntyrasnserthrtyr290295300argvalvalservalleuthrvalleuhisglnasptrpleuasngly305310315320lysglutyrlyscyslysvalserasnlysalaleuproalaproile325330335glulysthrileserlysalalysglyglnproarggluproglnval340345350tyrthrleuproproserargaspgluleuthrlysasnglnvalser355360365leuthrcysleuvallysglyphetyrproseraspilealavalglu370375380trpgluserasnglyglnprogluasnasntyrlysthrthrpropro385390395400valleuaspseraspglyserphepheleutyrserlysleuthrval405410415asplysserargtrpglnglnglyasnvalphesercysservalmet420425430hisglualaleuhisasnhistyrthrglnlysserleuserleuser435440445proglylys450<210>22<211>215<212>prt<213>人工序列<220><223>lccea2f1<400>22aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcyslysalaseralaalavalglythrtyr202530valalatrptyrglnglnlysproglylysalaprolysleuleuile354045tyrseralasertyrarglysargglyvalproserargphesergly505560serglyserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcyshisglntyrtyrthrtyrproleu859095phethrpheglyglnglythrlysleugluilelysargthrvalala100105110alaproservalpheilepheproproseraspgluglnleulysser115120125glythralaservalvalcysleuleuasnasnphetyrproargglu130135140alalysvalglntrplysvalaspasnalaleuglnserglyasnser145150155160glngluservalthrgluglnaspserlysaspserthrtyrserleu165170175serserthrleuthrleuserlysalaasptyrglulyshislysval180185190tyralacysgluvalthrhisglnglyleuserserprovalthrlys195200205serpheasnargglyglucys210215<210>23<211>121<212>prt<213>人工序列<220><223>vhceach1a1a98-99<400>23glnvalglnleuvalglnserglyalagluvallyslysproglyala151015servallysvalsercyslysalaserglytyrthrphethrgluphe202530glymetasntrpvalargglnalaproglyglnglyleuglutrpmet354045glytrpileasnthrlysthrglyglualathrtyrvalglugluphe505560lysglyargvalthrphethrthraspthrserthrserthralatyr65707580metgluleuargserleuargseraspaspthralavaltyrtyrcys859095alaargtrpaspphealatyrtyrvalglualametasptyrtrpgly100105110glnglythrthrvalthrvalserser115120<210>24<211>5<212>prt<213>人工序列<220><223>hcdr1ceach1a1a98-99<400>24glupheglymetasn15<210>25<211>17<212>prt<213>人工序列<220><223>hcdr2ceach1a1a98-99<400>25trpileasnthrlysthrglyglualathrtyrvalglugluphelys151015gly<210>26<211>12<212>prt<213>人工序列<220><223>hcdr3ceach1a1a98-99<400>26trpaspphealatyrtyrvalglualametasptyr1510<210>27<211>108<212>prt<213>人工序列<220><223>vlcea2f1<400>27aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcyslysalaseralaalavalglythrtyr202530valalatrptyrglnglnlysproglylysalaprolysleuleuile354045tyrseralasertyrarglysargglyvalproserargphesergly505560serglyserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcyshisglntyrtyrthrtyrproleu859095phethrpheglyglnglythrlysleugluilelys100105<210>28<211>11<212>prt<213>人工序列<220><223>lcdr1cea2f1<400>28lysalaseralaalavalglythrtyrvalala1510<210>29<211>7<212>prt<213>人工序列<220><223>lcdr2cea2f1<400>29seralasertyrarglysarg15<210>30<211>10<212>prt<213>人工序列<220><223>lcdr3cea2f1<400>30hisglntyrtyrthrtyrproleuphethr1510<210>31<211>111<212>prt<213>人工序列<220><223>vlcd3ch2527(vl_7-43(11))<400>31glnthrvalvalthrglngluproserleuthrvalserproglygly151015thrvalthrleuthrcysglyserserthrglyalavalthrthrser202530asntyralaasntrpvalglnglnlysproglyglnalaproarggly354045leuileglyglythrasnlysargalaproglythrproalaargphe505560serglyserleuleuglyglylysalaalaleuthrleuserglyval65707580glnprogluaspglualaglutyrtyrcysalaleutrptyrserasn859095leutrpvalpheglyglyglythrlysleuthrvalleuserser100105110<210>32<211>125<212>prt<213>人工序列<220><223>vhcd3ch2527(vhcomboa49sv93a)<400>32gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrpheserthrtyr202530alametasntrpvalargglnalaproglylysglyleuglutrpval354045serargileargserlystyrasnasntyralathrtyrtyralaasp505560servallysglyargphethrileserargaspaspserlysasnthr65707580leutyrleuglnmetasnserleuargalagluaspthralavaltyr859095tyrcysalaarghisglyasnpheglyasnsertyrvalsertrpphe100105110alatyrtrpglyglnglythrleuvalthrvalserser115120125<210>33<211>125<212>prt<213>人工序列<220><223>vhcd3ch2527(vhcomboa49sv93ar94k)<400>33gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrpheserthrtyr202530alametasntrpvalargglnalaproglylysglyleuglutrpval354045serargileargserlystyrasnasntyralathrtyrtyralaasp505560servallysglyargphethrileserargaspaspserlysasnthr65707580leutyrleuglnmetasnserleuargalagluaspthralavaltyr859095tyrcysalalyshisglyasnpheglyasnsertyrvalsertrpphe100105110alatyrtrpglyglnglythrleuvalthrvalserser115120125<210>34<211>112<212>prt<213>人工序列<220><223>vhmcspm4-3(d6)<400>34glnvalglnleuglngluserglyproglyleuvallysprosergln151015thrleuserleuthrcysthrvalserglyglyserilethrsergly202530tyrtyrtrpasntrpileargglnhisproglylysglyleuglutrp354045ileglytyrilethrpheaspglylysasnasntyrasnproserleu505560lysserargvalthrileserargaspthrserlysasnglnpheser65707580leulysleuserservalthralaalaaspthralavaltyrtyrcys859095alaasppheasptyrtrpglyglnglythrleuvalthrvalserser100105110<210>35<211>15<212>prt<213>人工序列<220><223>hcdr2mcspm4-3(d6)<400>35ilethrpheaspglylysasnasntyrasnproserleulysser151015<210>36<211>112<212>prt<213>人工序列<220><223>vhmcspm4-3(a7)<400>36glnvalglnleuglngluserglyproglyleuvallysprosergln151015thrleuserleuthrcysthrvalserglyglyserilethraspgly202530tyrtyrtrpasntrpileargglnhisproglylysglyleuglutrp354045ileglytyrilethrpheaspglyargasnasntyrasnproserleu505560lysserargvalthrileserargaspthrserlysasnglnpheser65707580leulysleuserservalthralaalaaspthralavaltyrtyrcys859095alaasppheasptyrtrpglyglnglythrleuvalthrvalserser100105110<210>37<211>6<212>prt<213>人工序列<220><223>hcdr1mcspm4-3(a7)<400>37aspglytyrtyrtrpasn15<210>38<211>15<212>prt<213>人工序列<220><223>hcdr2mcspm4-3(a7)<400>38ilethrpheaspglyargasnasntyrasnproserleulysser151015<210>39<211>112<212>prt<213>人工序列<220><223>vhmcspm4-3(b7)<400>39glnvalglnleuglngluserglyproglyleuvallysprosergln151015thrleuserleuthrcysthrvalserglyglyserilethrsergly202530tyrtyrtrpasntrpileargglnhisproglylysglyleuglutrp354045ileglytyrilethrpheaspglyileasnasntyrasnproserleu505560lysserargvalthrileserargaspthrserlysasnglnpheser65707580leulysleuserservalthralaalaaspthralavaltyrtyrcys859095alaasppheasptyrtrpglyglnglythrleuvalthrvalserser100105110<210>40<211>15<212>prt<213>人工序列<220><223>hcdr2mcspm4-3(b7)<400>40ilethrpheaspglyileasnasntyrasnproserleulysser151015<210>41<211>112<212>prt<213>人工序列<220><223>vhmcspm4-3(b8)<400>41glnvalglnleuglngluserglyproglyleuvallysprosergln151015thrleuserleuthrcysthrvalserglyglyserilethrsergly202530tyrtyrtrpasntrpileargglnhisproglylysglyleuglutrp354045ileglytyrilethrpheaspglyargasnasntyrasnproserleu505560lysserargvalthrileserargaspthrserlysasnglnpheser65707580leulysleuserservalthralaalaaspthralavaltyrtyrcys859095alaasppheasptyrtrpglyglnglythrleuvalthrvalserser100105110<210>42<211>112<212>prt<213>人工序列<220><223>vhmcspm4-3<400>42glnvalglnleuglngluserglyproglyleuvallysprosergln151015thrleuserleuthrcysthrvalserglyglyserilethrsergly202530tyrtyrtrpasntrpileargglnhisproglylysglyleuglutrp354045ileglytyrilethrtyraspglyserasnasntyrasnproserleu505560lysserargvalthrileserargaspthrserlysasnglnpheser65707580leulysleuserservalthralaalaaspthralavaltyrtyrcys859095alaasppheasptyrtrpglyglnglythrleuvalthrvalserser100105110<210>43<211>107<212>prt<213>人工序列<220><223>vlmcspml2(e10)<400>43aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalasertyrglyileargglytyr202530leuasntrptyrglnglnlysproglylysalaprolysleuleuile354045tyrtyrthrserserleuhisserglyvalproserargphesergly505560serglyserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrhiscysglnglntyrserlysleuprotrp859095thrpheglyglnglythrlysvalgluilelys100105<210>44<211>11<212>prt<213>人工序列<220><223>lcdr1mcspml2(e10)<400>44argalasertyrglyileargglytyrleuasn1510<210>45<211>9<212>prt<213>人工序列<220><223>lcdr3mcspml2(e10)<400>45glnglntyrserlysleuprotrpthr15<210>46<211>107<212>prt<213>人工序列<220><223>vlmcspml2(e10-g3)<400>46aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalasertyrglyileargglytyr202530leuasntrptyrglnglnlysproglylysalaprolysleuleuile354045tyrtyrthrserserleuhisserglyvalproserargphesergly505560serglyserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrhiscysglnglntyrseralaleuprotrp859095thrpheglyglnglythrlysvalgluilelys100105<210>47<211>107<212>prt<213>人工序列<220><223>vlmcspml2(c5)<400>47aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserargglyileargglutyr202530leuasntrptyrglnglnlysproglylysalaprolysleuleuile354045tyrtyrthrglyserleuhisserglyvalproserargphesergly505560serglyserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysglnglntyrsergluleuprotrp859095thrpheglyglnglythrlysvalgluilelys100105<210>48<211>11<212>prt<213>人工序列<220><223>lcdr1mcspml2(c5)<400>48argalaserargglyileargglutyrleuasn1510<210>49<211>7<212>prt<213>人工序列<220><223>lcdr2mcspml2(c5)<400>49tyrthrglyserleuhisser15<210>50<211>9<212>prt<213>人工序列<220><223>lcdr3mcspml2(c5)<400>50glnglntyrsergluleuprotrpthr15<210>51<211>107<212>prt<213>人工序列<220><223>vlmcspml2(c5-g3)<400>51aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserargglyileargglutyr202530leuasntrptyrglnglnlysproglylysalaprolysleuleuile354045tyrtyrthrglyserleuhisserglyvalproserargphesergly505560serglyserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysglnglntyrseralaleuprotrp859095thrpheglyglnglythrlysvalgluilelys100105<210>52<211>107<212>prt<213>人工序列<220><223>vlmcspml2<400>52aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnglyileargasntyr202530leuasntrptyrglnglnlysproglylysalaprolysleuleuile354045tyrtyrthrserserleuhisserglyvalproserargphesergly505560serglyserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysglnglntyrserlysleuprotrp859095thrpheglyglnglythrlysvalgluilelys100105<210>53<211>214<212>prt<213>人工序列<220><223>lccd3ch2527(crossfab,vl-ch1)<400>53glnalavalvalthrglngluproserleuthrvalserproglygly151015thrvalthrleuthrcysglyserserthrglyalavalthrthrser202530asntyralaasntrpvalglnglulysproglyglnalaphearggly354045leuileglyglythrasnlysargalaproglythrproalaargphe505560serglyserleuleuglyglylysalaalaleuthrleuserglyala65707580glnprogluaspglualaglutyrtyrcysalaleutrptyrserasn859095leutrpvalpheglyglyglythrlysleuthrvalleuserserala100105110serthrlysglyproservalpheproleualaproserserlysser115120125thrserglyglythralaalaleuglycysleuvallysasptyrphe130135140progluprovalthrvalsertrpasnserglyalaleuthrsergly145150155160valhisthrpheproalavalleuglnserserglyleutyrserleu165170175serservalvalthrvalproserserserleuglythrglnthrtyr180185190ilecysasnvalasnhislysproserasnthrlysvalasplyslys195200205valgluprolyssercys210<210>54<211>685<212>prt<213>人工序列<220><223>mcspm4-3(c1)(vh-ch1)-cd3ch2527(crossfabvh-ck)-fc(节)p329glala<400>54glnvalglnleuglngluserglyproglyleuvallysprosergln151015thrleuserleuthrcysthrvalserglyglyserilethrsergly202530tyrtyrtrpasntrpileargglnhisproglylysglyleuglutrp354045ileglytyrilethrpheaspglyserasnasntyrasnproserleu505560lysserargvalthrileserargaspthrserlysasnglnpheser65707580leulysleuserservalthralaalaaspthralavaltyrtyrcys859095alaasppheasptyrtrpglyglnglythrleuvalthrvalserser100105110alaserthrlysglyproservalpheproleualaproserserlys115120125serthrserglyglythralaalaleuglycysleuvallysasptyr130135140pheprogluprovalthrvalsertrpasnserglyalaleuthrser145150155160glyvalhisthrpheproalavalleuglnserserglyleutyrser165170175leuserservalvalthrvalproserserserleuglythrglnthr180185190tyrilecysasnvalasnhislysproserasnthrlysvalasplys195200205lysvalgluprolyssercysaspglyglyglyglyserglyglygly210215220glysergluvalglnleuleugluserglyglyglyleuvalglnpro225230235240glyglyserleuargleusercysalaalaserglyphethrpheser245250255thrtyralametasntrpvalargglnalaproglylysglyleuglu260265270trpvalserargileargserlystyrasnasntyralathrtyrtyr275280285alaaspservallysglyargphethrileserargaspaspserlys290295300asnthrleutyrleuglnmetasnserleuargalagluaspthrala305310315320valtyrtyrcysvalarghisglyasnpheglyasnsertyrvalser325330335trpphealatyrtrpglyglnglythrleuvalthrvalserserala340345350servalalaalaproservalpheilepheproproseraspglugln355360365leulysserglythralaservalvalcysleuleuasnasnphetyr370375380proargglualalysvalglntrplysvalaspasnalaleuglnser385390395400glyasnserglngluservalthrgluglnaspserlysaspserthr405410415tyrserleuserserthrleuthrleuserlysalaasptyrglulys420425430hislysvaltyralacysgluvalthrhisglnglyleuserserpro435440445valthrlysserpheasnargglyglucysasplysthrhisthrcys450455460proprocysproalaproglualaalaglyglyproservalpheleu465470475480pheproprolysprolysaspthrleumetileserargthrproglu485490495valthrcysvalvalvalaspvalserhisgluaspprogluvallys500505510pheasntrptyrvalaspglyvalgluvalhisasnalalysthrlys515520525proargglugluglntyrasnserthrtyrargvalvalservalleu530535540thrvalleuhisglnasptrpleuasnglylysglutyrlyscyslys545550555560valserasnlysalaleuglyalaproileglulysthrileserlys565570575alalysglyglnproarggluproglnvaltyrthrleuproprocys580585590argaspgluleuthrlysasnglnvalserleutrpcysleuvallys595600605glyphetyrproseraspilealavalglutrpgluserasnglygln610615620progluasnasntyrlysthrthrproprovalleuaspseraspgly625630635640serphepheleutyrserlysleuthrvalasplysserargtrpgln645650655glnglyasnvalphesercysservalmethisglualaleuhisasn660665670histyrthrglnlysserleuserleuserproglylys675680685<210>55<211>442<212>prt<213>人工序列<220><223>mcspm4-3(c1)(vh-ch1)-fc(穴)p329glala<400>55glnvalglnleuglngluserglyproglyleuvallysprosergln151015thrleuserleuthrcysthrvalserglyglyserilethrsergly202530tyrtyrtrpasntrpileargglnhisproglylysglyleuglutrp354045ileglytyrilethrpheaspglyserasnasntyrasnproserleu505560lysserargvalthrileserargaspthrserlysasnglnpheser65707580leulysleuserservalthralaalaaspthralavaltyrtyrcys859095alaasppheasptyrtrpglyglnglythrleuvalthrvalserser100105110alaserthrlysglyproservalpheproleualaproserserlys115120125serthrserglyglythralaalaleuglycysleuvallysasptyr130135140pheprogluprovalthrvalsertrpasnserglyalaleuthrser145150155160glyvalhisthrpheproalavalleuglnserserglyleutyrser165170175leuserservalvalthrvalproserserserleuglythrglnthr180185190tyrilecysasnvalasnhislysproserasnthrlysvalasplys195200205lysvalgluprolyssercysasplysthrhisthrcysproprocys210215220proalaproglualaalaglyglyproservalpheleuphepropro225230235240lysprolysaspthrleumetileserargthrprogluvalthrcys245250255valvalvalaspvalserhisgluaspprogluvallyspheasntrp260265270tyrvalaspglyvalgluvalhisasnalalysthrlysproargglu275280285gluglntyrasnserthrtyrargvalvalservalleuthrvalleu290295300hisglnasptrpleuasnglylysglutyrlyscyslysvalserasn305310315320lysalaleuglyalaproileglulysthrileserlysalalysgly325330335glnproarggluproglnvalcysthrleuproproserargaspglu340345350leuthrlysasnglnvalserleusercysalavallysglyphetyr355360365proseraspilealavalglutrpgluserasnglyglnprogluasn370375380asntyrlysthrthrproprovalleuaspseraspglyserphephe385390395400leuvalserlysleuthrvalasplysserargtrpglnglnglyasn405410415valphesercysservalmethisglualaleuhisasnhistyrthr420425430glnlysserleuserleuserproglylys435440<210>56<211>214<212>prt<213>人工序列<220><223>lccd3ch2527(crossfab,vl-ch1)<400>56glnalavalvalthrglngluproserleuthrvalserproglygly151015thrvalthrleuthrcysglyserserthrglyalavalthrthrser202530asntyralaasntrpvalglnglulysproglyglnalaphearggly354045leuileglyglythrasnlysargalaproglythrproalaargphe505560serglyserleuleuglyglylysalaalaleuthrleuserglyala65707580glnprogluaspglualaglutyrtyrcysalaleutrptyrserasn859095leutrpvalpheglyglyglythrlysleuthrvalleuserserala100105110serthrlysglyproservalpheproleualaproserserlysser115120125thrserglyglythralaalaleuglycysleuvallysasptyrphe130135140progluprovalthrvalsertrpasnserglyalaleuthrsergly145150155160valhisthrpheproalavalleuglnserserglyleutyrserleu165170175serservalvalthrvalproserserserleuglythrglnthrtyr180185190ilecysasnvalasnhislysproserasnthrlysvalasplyslys195200205valgluprolyssercys210<210>57<211>694<212>prt<213>人工序列<220><223>ceach1a1a98/99-cd3ch2527(crossfabvh-ck)-fc(节)p329glala<400>57glnvalglnleuvalglnserglyalagluvallyslysproglyala151015servallysvalsercyslysalaserglytyrthrphethrgluphe202530glymetasntrpvalargglnalaproglyglnglyleuglutrpmet354045glytrpileasnthrlysthrglyglualathrtyrvalglugluphe505560lysglyargvalthrphethrthraspthrserthrserthralatyr65707580metgluleuargserleuargseraspaspthralavaltyrtyrcys859095alaargtrpaspphealatyrtyrvalglualametasptyrtrpgly100105110glnglythrthrvalthrvalserseralaserthrlysglyproser115120125valpheproleualaproserserlysserthrserglyglythrala130135140alaleuglycysleuvallysasptyrpheprogluprovalthrval145150155160sertrpasnserglyalaleuthrserglyvalhisthrpheproala165170175valleuglnserserglyleutyrserleuserservalvalthrval180185190proserserserleuglythrglnthrtyrilecysasnvalasnhis195200205lysproserasnthrlysvalasplyslysvalgluprolyssercys210215220aspglyglyglyglyserglyglyglyglysergluvalglnleuleu225230235240gluserglyglyglyleuvalglnproglyglyserleuargleuser245250255cysalaalaserglyphethrpheserthrtyralametasntrpval260265270argglnalaproglylysglyleuglutrpvalserargileargser275280285lystyrasnasntyralathrtyrtyralaaspservallysglyarg290295300phethrileserargaspaspserlysasnthrleutyrleuglnmet305310315320asnserleuargalagluaspthralavaltyrtyrcysvalarghis325330335glyasnpheglyasnsertyrvalsertrpphealatyrtrpglygln340345350glythrleuvalthrvalserseralaservalalaalaproserval355360365pheilepheproproseraspgluglnleulysserglythralaser370375380valvalcysleuleuasnasnphetyrproargglualalysvalgln385390395400trplysvalaspasnalaleuglnserglyasnserglngluserval405410415thrgluglnaspserlysaspserthrtyrserleuserserthrleu420425430thrleuserlysalaasptyrglulyshislysvaltyralacysglu435440445valthrhisglnglyleuserserprovalthrlysserpheasnarg450455460glyglucysasplysthrhisthrcysproprocysproalaproglu465470475480alaalaglyglyproservalpheleupheproprolysprolysasp485490495thrleumetileserargthrprogluvalthrcysvalvalvalasp500505510valserhisgluaspprogluvallyspheasntrptyrvalaspgly515520525valgluvalhisasnalalysthrlysproargglugluglntyrasn530535540serthrtyrargvalvalservalleuthrvalleuhisglnasptrp545550555560leuasnglylysglutyrlyscyslysvalserasnlysalaleugly565570575alaproileglulysthrileserlysalalysglyglnproargglu580585590proglnvaltyrthrleuproprocysargaspgluleuthrlysasn595600605glnvalserleutrpcysleuvallysglyphetyrproseraspile610615620alavalglutrpgluserasnglyglnprogluasnasntyrlysthr625630635640thrproprovalleuaspseraspglyserphepheleutyrserlys645650655leuthrvalasplysserargtrpglnglnglyasnvalphesercys660665670servalmethisglualaleuhisasnhistyrthrglnlysserleu675680685serleuserproglylys690<210>58<211>451<212>prt<213>人工序列<220><223>ceach1a1a98/99(vh-ch1)-fc(穴)p329glala<400>58glnvalglnleuvalglnserglyalagluvallyslysproglyala151015servallysvalsercyslysalaserglytyrthrphethrgluphe202530glymetasntrpvalargglnalaproglyglnglyleuglutrpmet354045glytrpileasnthrlysthrglyglualathrtyrvalglugluphe505560lysglyargvalthrphethrthraspthrserthrserthralatyr65707580metgluleuargserleuargseraspaspthralavaltyrtyrcys859095alaargtrpaspphealatyrtyrvalglualametasptyrtrpgly100105110glnglythrthrvalthrvalserseralaserthrlysglyproser115120125valpheproleualaproserserlysserthrserglyglythrala130135140alaleuglycysleuvallysasptyrpheprogluprovalthrval145150155160sertrpasnserglyalaleuthrserglyvalhisthrpheproala165170175valleuglnserserglyleutyrserleuserservalvalthrval180185190proserserserleuglythrglnthrtyrilecysasnvalasnhis195200205lysproserasnthrlysvalasplyslysvalgluprolyssercys210215220asplysthrhisthrcysproprocysproalaproglualaalagly225230235240glyproservalpheleupheproprolysprolysaspthrleumet245250255ileserargthrprogluvalthrcysvalvalvalaspvalserhis260265270gluaspprogluvallyspheasntrptyrvalaspglyvalgluval275280285hisasnalalysthrlysproargglugluglntyrasnserthrtyr290295300argvalvalservalleuthrvalleuhisglnasptrpleuasngly305310315320lysglutyrlyscyslysvalserasnlysalaleuglyalaproile325330335glulysthrileserlysalalysglyglnproarggluproglnval340345350cysthrleuproproserargaspgluleuthrlysasnglnvalser355360365leusercysalavallysglyphetyrproseraspilealavalglu370375380trpgluserasnglyglnprogluasnasntyrlysthrthrpropro385390395400valleuaspseraspglyserphepheleuvalserlysleuthrval405410415asplysserargtrpglnglnglyasnvalphesercysservalmet420425430hisglualaleuhisasnhistyrthrglnlysserleuserleuser435440445proglylys450<210>59<211>215<212>prt<213>人工序列<220><223>lcdp47gs<400>59gluilevalleuthrglnserproglythrleuserleuserprogly151015gluargalathrleusercysargalaserglnservalserserser202530tyrleualatrptyrglnglnlysproglyglnalaproargleuleu354045iletyrglyalaserserargalathrglyileproaspargpheser505560glyserglyserglythraspphethrleuthrileserargleuglu65707580progluaspphealavaltyrtyrcysglnglntyrglyserserpro859095leuthrpheglyglnglythrlysvalgluilelysargthrvalala100105110alaproservalpheilepheproproseraspgluglnleulysser115120125glythralaservalvalcysleuleuasnasnphetyrproargglu130135140alalysvalglntrplysvalaspasnalaleuglnserglyasnser145150155160glngluservalthrgluglnaspserlysaspserthrtyrserleu165170175serserthrleuthrleuserlysalaasptyrglulyshislysval180185190tyralacysgluvalthrhisglnglyleuserserprovalthrlys195200205serpheasnargglyglucys210215<210>60<211>214<212>prt<213>人工序列<220><223>lccd3ch2527(crossfab,vl-ch1)<400>60glnalavalvalthrglngluproserleuthrvalserproglygly151015thrvalthrleuthrcysglyserserthrglyalavalthrthrser202530asntyralaasntrpvalglnglulysproglyglnalaphearggly354045leuileglyglythrasnlysargalaproglythrproalaargphe505560serglyserleuleuglyglylysalaalaleuthrleuserglyala65707580glnprogluaspglualaglutyrtyrcysalaleutrptyrserasn859095leutrpvalpheglyglyglythrlysleuthrvalleuserserala100105110serthrlysglyproservalpheproleualaproserserlysser115120125thrserglyglythralaalaleuglycysleuvallysasptyrphe130135140progluprovalthrvalsertrpasnserglyalaleuthrsergly145150155160valhisthrpheproalavalleuglnserserglyleutyrserleu165170175serservalvalthrvalproserserserleuglythrglnthrtyr180185190ilecysasnvalasnhislysproserasnthrlysvalasplyslys195200205valgluprolyssercys210<210>61<211>688<212>prt<213>人工序列<220><223>dp47gs(vh-ch1)-cd3ch2527(crossfabvh-ck)-fc(节)p329glala<400>61gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrphesersertyr202530alametsertrpvalargglnalaproglylysglyleuglutrpval354045seralaileserglyserglyglyserthrtyrtyralaaspserval505560lysglyargphethrileserargaspasnserlysasnthrleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alalysglyserglypheasptyrtrpglyglnglythrleuvalthr100105110valserseralaserthrlysglyproservalpheproleualapro115120125serserlysserthrserglyglythralaalaleuglycysleuval130135140lysasptyrpheprogluprovalthrvalsertrpasnserglyala145150155160leuthrserglyvalhisthrpheproalavalleuglnsersergly165170175leutyrserleuserservalvalthrvalproserserserleugly180185190thrglnthrtyrilecysasnvalasnhislysproserasnthrlys195200205valasplyslysvalgluprolyssercysaspglyglyglyglyser210215220glyglyglyglysergluvalglnleuleugluserglyglyglyleu225230235240valglnproglyglyserleuargleusercysalaalaserglyphe245250255thrpheserthrtyralametasntrpvalargglnalaproglylys260265270glyleuglutrpvalserargileargserlystyrasnasntyrala275280285thrtyrtyralaaspservallysglyargphethrileserargasp290295300aspserlysasnthrleutyrleuglnmetasnserleuargalaglu305310315320aspthralavaltyrtyrcysvalarghisglyasnpheglyasnser325330335tyrvalsertrpphealatyrtrpglyglnglythrleuvalthrval340345350serseralaservalalaalaproservalpheilepheproproser355360365aspgluglnleulysserglythralaservalvalcysleuleuasn370375380asnphetyrproargglualalysvalglntrplysvalaspasnala385390395400leuglnserglyasnserglngluservalthrgluglnaspserlys405410415aspserthrtyrserleuserserthrleuthrleuserlysalaasp420425430tyrglulyshislysvaltyralacysgluvalthrhisglnglyleu435440445serserprovalthrlysserpheasnargglyglucysasplysthr450455460histhrcysproprocysproalaproglualaalaglyglyproser465470475480valpheleupheproprolysprolysaspthrleumetileserarg485490495thrprogluvalthrcysvalvalvalaspvalserhisgluasppro500505510gluvallyspheasntrptyrvalaspglyvalgluvalhisasnala515520525lysthrlysproargglugluglntyrasnserthrtyrargvalval530535540servalleuthrvalleuhisglnasptrpleuasnglylysglutyr545550555560lyscyslysvalserasnlysalaleuglyalaproileglulysthr565570575ileserlysalalysglyglnproarggluproglnvaltyrthrleu580585590proprocysargaspgluleuthrlysasnglnvalserleutrpcys595600605leuvallysglyphetyrproseraspilealavalglutrpgluser610615620asnglyglnprogluasnasntyrlysthrthrproprovalleuasp625630635640seraspglyserphepheleutyrserlysleuthrvalasplysser645650655argtrpglnglnglyasnvalphesercysservalmethisgluala660665670leuhisasnhistyrthrglnlysserleuserleuserproglylys675680685<210>62<211>445<212>prt<213>人工序列<220><223>dp47gs(vh-ch1)-fc(穴)p329glala<400>62gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglyphethrphesersertyr202530alametsertrpvalargglnalaproglylysglyleuglutrpval354045seralaileserglyserglyglyserthrtyrtyralaaspserval505560lysglyargphethrileserargaspasnserlysasnthrleutyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alalysglyserglypheasptyrtrpglyglnglythrleuvalthr100105110valserseralaserthrlysglyproservalpheproleualapro115120125serserlysserthrserglyglythralaalaleuglycysleuval130135140lysasptyrpheprogluprovalthrvalsertrpasnserglyala145150155160leuthrserglyvalhisthrpheproalavalleuglnsersergly165170175leutyrserleuserservalvalthrvalproserserserleugly180185190thrglnthrtyrilecysasnvalasnhislysproserasnthrlys195200205valasplyslysvalgluprolyssercysasplysthrhisthrcys210215220proprocysproalaproglualaalaglyglyproservalpheleu225230235240pheproprolysprolysaspthrleumetileserargthrproglu245250255valthrcysvalvalvalaspvalserhisgluaspprogluvallys260265270pheasntrptyrvalaspglyvalgluvalhisasnalalysthrlys275280285proargglugluglntyrasnserthrtyrargvalvalservalleu290295300thrvalleuhisglnasptrpleuasnglylysglutyrlyscyslys305310315320valserasnlysalaleuglyalaproileglulysthrileserlys325330335alalysglyglnproarggluproglnvalcysthrleuproproser340345350argaspgluleuthrlysasnglnvalserleusercysalavallys355360365glyphetyrproseraspilealavalglutrpgluserasnglygln370375380progluasnasntyrlysthrthrproprovalleuaspseraspgly385390395400serphepheleuvalserlysleuthrvalasplysserargtrpgln405410415glnglyasnvalphesercysservalmethisglualaleuhisasn420425430histyrthrglnlysserleuserleuserproglylys435440445<210>63<211>1365<212>dna<213>人工序列<220><223>hccd3ch2527(vh_3-23(12))<400>63gaggtgcagctgctggaatctggcggcggactggtgcagcctggcggatctctgagactg60agctgtgccgccagcggcttcaccttcagcacctacgccatgaactgggtgcgccaggcc120cctggcaaaggcctggaatgggtgtcccggatcagaagcaagtacaacaactacgccacc180tactacgccgacagcgtgaagggccggttcaccatcagccgggacgacagcaagaacacc240ctgtacctgcagatgaacagcctgcgggccgaggacaccgccgtgtactattgtgtgcgg300cacggcaacttcggcaacagctatgtgtcttggtttgcctactggggccagggcaccctc360gtgaccgtgtcatctgctagcaccaagggcccatcggtcttccccctggcaccctcctcc420aagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaa480ccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggct540gtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagc600ttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggac660aagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacct720gaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatg780atctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgag840gtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgg900gaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggac960tggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatc1020gagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgccc1080ccatcccgggatgagctgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttc1140tatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaag1200accacgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtg1260gacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctg1320cacaaccactacacgcagaagagcctctccctgtctccgggtaaa1365<210>64<211>645<212>dna<213>人工序列<220><223>lccd3ch2527(vl_7-46(13))<400>64caggccgtcgtgacccaggaacccagcctgacagtgtctcctggcggcaccgtgaccctg60acatgtggcagttctacaggcgccgtgaccaccagcaactacgccaactgggtgcaggaa120aagcccggccaggccttcagaggactgatcggcggcaccaacaagagagcccctggcacc180cctgccagattcagcggatctctgctgggaggaaaggccgccctgacactgtctggcgcc240cagccagaagatgaggccgagtactactgcgccctgtggtacagcaacctgtgggtgttc300ggcggaggcaccaagctgacagtcctaggtcaacccaaggctgcccccagcgtgaccctg360ttcccccccagcagcgaggaactgcaggccaacaaggccaccctggtctgcctgatcagc420gacttctacccaggcgccgtgaccgtggcctggaaggccgacagcagccccgtgaaggcc480ggcgtggagaccaccacccccagcaagcagagcaacaacaagtacgccgccagcagctac540ctgagcctgacccccgagcagtggaagagccacaggtcctacagctgccaggtgacccac600gagggcagcaccgtggagaaaaccgtggcccccaccgagtgcagc645<210>65<211>375<212>dna<213>人工序列<220><223>vhcd3ch2527(vh_3-23(12))<400>65gaggtgcagctgctggaatctggcggcggactggtgcagcctggcggatctctgagactg60agctgtgccgccagcggcttcaccttcagcacctacgccatgaactgggtgcgccaggcc120cctggcaaaggcctggaatgggtgtcccggatcagaagcaagtacaacaactacgccacc180tactacgccgacagcgtgaagggccggttcaccatcagccgggacgacagcaagaacacc240ctgtacctgcagatgaacagcctgcgggccgaggacaccgccgtgtactattgtgtgcgg300cacggcaacttcggcaacagctatgtgtcttggtttgcctactggggccagggcaccctc360gtgaccgtgtcatct375<210>66<211>15<212>dna<213>人工序列<220><223>hcdr1cd3ch2527(vh_3-23(12))<400>66acctacgccatgaac15<210>67<211>57<212>dna<213>人工序列<220><223>hcdr2cd3ch2527(vh_3-23(12))<400>67cggatcagaagcaagtacaacaactacgccacctactacgccgacagcgtgaagggc57<210>68<211>42<212>dna<213>人工序列<220><223>hcdr3cd3ch2527(vh_3-23(12))<400>68cacggcaacttcggcaacagctatgtgtcttggtttgcctac42<210>69<211>327<212>dna<213>人工序列<220><223>vlcd3ch2527(vl_7-46(13))<400>69caggccgtcgtgacccaggaacccagcctgacagtgtctcctggcggcaccgtgaccctg60acatgtggcagttctacaggcgccgtgaccaccagcaactacgccaactgggtgcaggaa120aagcccggccaggccttcagaggactgatcggcggcaccaacaagagagcccctggcacc180cctgccagattcagcggatctctgctgggaggaaaggccgccctgacactgtctggcgcc240cagccagaagatgaggccgagtactactgcgccctgtggtacagcaacctgtgggtgttc300ggcggaggcaccaagctgacagtccta327<210>70<211>42<212>dna<213>人工序列<220><223>lcdr1cd3ch2527(vl_7-46(13))<400>70ggcagttctacaggcgccgtgaccaccagcaactacgccaac42<210>71<211>21<212>dna<213>人工序列<220><223>lcdr2cd3ch2527(vl_7-46(13))<400>71ggcaccaacaagagagcccct21<210>72<211>27<212>dna<213>人工序列<220><223>lcdr3cd3ch2527(vl_7-46(13))<400>72gccctgtggtacagcaacctgtgggtg27<210>73<211>1326<212>dna<213>人工序列<220><223>hcmcspm4-3(c1)<400>73caggtgcaattgcaggaaagcggccctggcctggtcaagcccagccagaccctgagcctg60acctgcaccgtgtccggcggcagcatcaccagcggctattattggaactggattcggcag120caccccggcaagggcctggaatggatcggctacatcactttcgacggctctaacaactac180aaccccagcctgaagtccagagtgaccatcagccgggacaccagcaagaaccagttcagc240ctgaagctgtccagcgtgacagccgccgacaccgccgtgtactactgcgccgacttcgac300tactggggccagggcaccctggtcaccgtgtccagcgctagcaccaagggcccatcggtc360ttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctg420gtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagc480ggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtg540gtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaag600cccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacaca660tgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttcccccca720aaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggac780gtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcat840aatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtc900ctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaac960aaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaa1020ccacaggtgtacaccctgcccccatcccgggatgagctgaccaagaaccaggtcagcctg1080acctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatggg1140cagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttc1200ctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgc1260tccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccg1320ggtaaa1326<210>74<211>642<212>dna<213>人工序列<220><223>lcmcspml2(g3)<400>74gacatccagatgacccagagccccagcagcctgagcgccagcgtgggcgacagagtgacc60atcacctgccgggccagccagggcatccggaactacctgaactggtatcagcagaagccc120ggcaaggcccccaagctgctgatctactacaccagcagcctgcacagcggcgtgcctagc180cggtttagcggcagcggctccggcaccgactacaccctgaccattagctccctgcagccc240gaggacttcgccacctactactgccagcagtactctgctctgccgtggaccttcggccag300ggaacaaaggtggagatcaagcgtacggtggctgcaccatctgtcttcatcttcccgcca360tctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctat420cccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccag480gagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacg540ctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggc600ctgagctcgcccgtcacaaagagcttcaacaggggagagtgt642<210>75<211>336<212>dna<213>人工序列<220><223>vhmcspm4-3(c1)<400>75caggtgcaattgcaggaaagcggccctggcctggtcaagcccagccagaccctgagcctg60acctgcaccgtgtccggcggcagcatcaccagcggctattattggaactggattcggcag120caccccggcaagggcctggaatggatcggctacatcactttcgacggctctaacaactac180aaccccagcctgaagtccagagtgaccatcagccgggacaccagcaagaaccagttcagc240ctgaagctgtccagcgtgacagccgccgacaccgccgtgtactactgcgccgacttcgac300tactggggccagggcaccctggtcaccgtgtccagc336<210>76<211>18<212>dna<213>人工序列<220><223>hcdr1mcspm4-3(c1)<400>76agcggctattattggaac18<210>77<211>48<212>dna<213>人工序列<220><223>hcdr2mcspm4-3(c1)<400>77tacatcactttcgacggctctaacaactacaaccccagcctgaagtcc48<210>78<211>9<212>dna<213>人工序列<220><223>hcdr3mcspm4-3(c1)<400>78ttcgactac9<210>79<211>321<212>dna<213>人工序列<220><223>vlmcspml2(g3)<400>79gacatccagatgacccagagccccagcagcctgagcgccagcgtgggcgacagagtgacc60atcacctgccgggccagccagggcatccggaactacctgaactggtatcagcagaagccc120ggcaaggcccccaagctgctgatctactacaccagcagcctgcacagcggcgtgcctagc180cggtttagcggcagcggctccggcaccgactacaccctgaccattagctccctgcagccc240gaggacttcgccacctactactgccagcagtactctgctctgccgtggaccttcggccag300ggaacaaaggtggagatcaag321<210>80<211>33<212>dna<213>人工序列<220><223>lcdr1mcspml2(g3)<400>80cgggccagccagggcatccggaactacctgaac33<210>81<211>21<212>dna<213>人工序列<220><223>lcdr2mcspml2(g3)<400>81tacaccagcagcctgcacagc21<210>82<211>27<212>dna<213>人工序列<220><223>lcdr3mcspml2(g3)<400>82cagcagtactctgctctgccgtggacc27<210>83<211>1353<212>dna<213>人工序列<220><223>hcceach1a1a98-99<400>83caggtgcagctggtgcagtctggcgccgaagtgaagaaacctggagctagtgtgaaggtg60tcctgcaaggccagcggctacaccttcaccgagttcggcatgaactgggtccgacaggct120ccaggccagggcctcgaatggatgggctggatcaacaccaagaccggcgaggccacctac180gtggaagagttcaagggcagagtgaccttcaccacggacaccagcaccagcaccgcctac240atggaactgcggagcctgagaagcgacgacaccgccgtgtactactgcgccagatgggac300ttcgcctattacgtggaagccatggactactggggccagggcaccaccgtgaccgtgtct360agcgctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctct420gggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtg480tcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcc540tcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccag600acctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgag660cccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctgggg720ggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacc780cctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaac840tggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtac900aacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggc960aaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatc1020tccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggat1080gagctgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgac1140atcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctccc1200gtgctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcagg1260tggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactac1320acgcagaagagcctctccctgtctccgggtaaa1353<210>84<211>645<212>dna<213>人工序列<220><223>lccea2f1<400>84gatatccagatgacccagtctccatcctccctgtctgcatctgtgggagacagagtcacc60atcacttgcaaggccagtgcggctgtgggtacgtatgttgcgtggtatcagcagaaacca120gggaaagcacctaagctcctgatctattcggcatcctaccgcaaaaggggagtcccatca180aggttcagtggcagtggatctgggacagatttcactctcaccatcagcagtctgcaacct240gaagatttcgcaacttactactgtcaccaatattacacctatcctctattcacgtttggc300cagggcaccaagctcgagatcaagcgtacggtggctgcaccatctgtcttcatcttcccg360ccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttc420tatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcc480caggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctg540acgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcag600ggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt645<210>85<211>363<212>dna<213>人工序列<220><223>vhceach1a1a98-99<400>85caggtgcagctggtgcagtctggcgccgaagtgaagaaacctggagctagtgtgaaggtg60tcctgcaaggccagcggctacaccttcaccgagttcggcatgaactgggtccgacaggct120ccaggccagggcctcgaatggatgggctggatcaacaccaagaccggcgaggccacctac180gtggaagagttcaagggcagagtgaccttcaccacggacaccagcaccagcaccgcctac240atggaactgcggagcctgagaagcgacgacaccgccgtgtactactgcgccagatgggac300ttcgcctattacgtggaagccatggactactggggccagggcaccaccgtgaccgtgtct360agc363<210>86<211>15<212>dna<213>人工序列<220><223>hcdr1ceach1a1a98-99<400>86gagttcggcatgaac15<210>87<211>51<212>dna<213>人工序列<220><223>hcdr2ceach1a1a98-99<400>87tggatcaacaccaagaccggcgaggccacctacgtggaagagttcaagggc51<210>88<211>36<212>dna<213>人工序列<220><223>hcdr3ceach1a1a98-99<400>88tgggacttcgcctattacgtggaagccatggactac36<210>89<211>324<212>dna<213>人工序列<220><223>vlcea2f1<400>89gatatccagatgacccagtctccatcctccctgtctgcatctgtgggagacagagtcacc60atcacttgcaaggccagtgcggctgtgggtacgtatgttgcgtggtatcagcagaaacca120gggaaagcacctaagctcctgatctattcggcatcctaccgcaaaaggggagtcccatca180aggttcagtggcagtggatctgggacagatttcactctcaccatcagcagtctgcaacct240gaagatttcgcaacttactactgtcaccaatattacacctatcctctattcacgtttggc300cagggcaccaagctcgagatcaag324<210>90<211>33<212>dna<213>人工序列<220><223>lcdr1cea2f1<400>90aaggccagtgcggctgtgggtacgtatgttgcg33<210>91<211>21<212>dna<213>人工序列<220><223>lcdr2cea2f1<400>91tcggcatcctaccgcaaaagg21<210>92<211>30<212>dna<213>人工序列<220><223>lcdr3cea2f1<400>92caccaatattacacctatcctctattcacg30<210>93<211>642<212>dna<213>人工序列<220><223>lccd3ch2527(crossfab,vl-ch1)<400>93caggccgtcgtgacccaggaacccagcctgacagtgtctcctggcggcaccgtgaccctg60acatgtggcagttctacaggcgccgtgaccaccagcaactacgccaactgggtgcaggaa120aagcccggccaggccttcagaggactgatcggcggcaccaacaagagagcccctggcacc180cctgccagattcagcggatctctgctgggaggaaaggccgccctgacactgtctggcgcc240cagccagaagatgaggccgagtactactgcgccctgtggtacagcaacctgtgggtgttc300ggcggaggcaccaagctgacagtgctgagcagcgcttccaccaaaggcccttccgtgttt360cctctggctcctagctccaagtccacctctggaggcaccgctgctctcggatgcctcgtg420aaggattattttcctgagcctgtgacagtgtcctggaatagcggagcactgacctctgga480gtgcatactttccccgctgtgctgcagtcctctggactgtacagcctgagcagcgtggtg540acagtgcccagcagcagcctgggcacccagacctacatctgcaacgtgaaccacaagccc600agcaacaccaaggtggacaagaaggtggaacccaagtcttgt642<210>94<211>2055<212>dna<213>人工序列<220><223>mcspm4-3(c1)(vh-ch1)-cd3ch2527(crossfabvh-ck)-fc(节)p329glala<400>94caggtgcaattgcaggaaagcggccctggcctggtcaagcccagccagaccctgagcctg60acctgcaccgtgtccggcggcagcatcaccagcggctattattggaactggattcggcag120caccccggcaagggcctggaatggatcggctacatcactttcgacggctctaacaactac180aaccccagcctgaagtccagagtgaccatcagccgggacaccagcaagaaccagttcagc240ctgaagctgtccagcgtgacagccgccgacaccgccgtgtactactgcgccgacttcgac300tactggggccagggcaccctggtcaccgtgtccagcgctagcacaaagggccccagcgtg360ttccctctggcccctagcagcaagagcacatctggcggaacagccgccctgggctgcctc420gtgaaggactactttcccgagcctgtgaccgtgtcctggaactctggcgccctgacaagc480ggcgtgcacacctttccagccgtgctgcagagcagcggcctgtactctctgagcagcgtg540gtcaccgtgcctagcagcagcctgggcacccagacctacatctgcaacgtgaaccacaag600cccagcaacaccaaagtggacaagaaggtggagcccaagagctgtgatggcggaggaggg660tccggaggcggaggatccgaggtgcagctgctggaatctggcggcggactggtgcagcct720ggcggatctctgagactgagctgtgccgccagcggcttcaccttcagcacctacgccatg780aactgggtgcgccaggcccctggcaaaggcctggaatgggtgtcccggatcagaagcaag840tacaacaactacgccacctactacgccgacagcgtgaagggccggttcaccatcagccgg900gacgacagcaagaacaccctgtacctgcagatgaacagcctgcgggccgaggacaccgcc960gtgtactattgtgtgcggcacggcaacttcggcaacagctatgtgtcttggtttgcctac1020tggggccagggcaccctcgtgaccgtgtcaagcgctagcgtggccgctccctccgtgttt1080atctttcccccatccgatgaacagctgaaaagcggcaccgcctccgtcgtgtgtctgctg1140aacaatttttaccctagggaagctaaagtgcagtggaaagtggataacgcactgcagtcc1200ggcaactcccaggaatctgtgacagaacaggactccaaggacagcacctactccctgtcc1260tccaccctgacactgtctaaggctgattatgagaaacacaaagtctacgcctgcgaagtc1320acccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtgacaag1380acccacacctgtcccccttgtcctgcccctgaagctgctggcggcccttctgtgttcctg1440ttccccccaaagcccaaggacaccctgatgatcagccggacccccgaagtgacctgcgtg1500gtggtggatgtgtcccacgaggaccctgaagtgaagttcaattggtacgtggacggcgtg1560gaagtgcacaacgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtg1620gtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaag1680gtctccaacaaagccctcggcgcccccatcgagaaaaccatctccaaagccaaagggcag1740ccccgagaaccacaggtgtacaccctgcccccatgccgggatgagctgaccaagaaccag1800gtcagcctgtggtgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggag1860agcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggc1920tccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtc1980ttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctcc2040ctgtctccgggtaaa2055<210>95<211>1326<212>dna<213>人工序列<220><223>mcspm4-3(c1)(vh-ch1)-fc(穴)p329glala<400>95caggtgcaattgcaggaaagcggccctggcctggtcaagcccagccagaccctgagcctg60acctgcaccgtgtccggcggcagcatcaccagcggctattattggaactggattcggcag120caccccggcaagggcctggaatggatcggctacatcactttcgacggctctaacaactac180aaccccagcctgaagtccagagtgaccatcagccgggacaccagcaagaaccagttcagc240ctgaagctgtccagcgtgacagccgccgacaccgccgtgtactactgcgccgacttcgac300tactggggccagggcaccctggtcaccgtgtccagcgctagcaccaagggcccctccgtg360ttccccctggcccccagcagcaagagcaccagcggcggcacagccgctctgggctgcctg420gtcaaggactacttccccgagcccgtgaccgtgtcctggaacagcggagccctgacctcc480ggcgtgcacaccttccccgccgtgctgcagagttctggcctgtatagcctgagcagcgtg540gtcaccgtgccttctagcagcctgggcacccagacctacatctgcaacgtgaaccacaag600cccagcaacaccaaggtggacaagaaggtggagcccaagagctgcgacaaaactcacaca660tgcccaccgtgcccagcacctgaagctgcagggggaccgtcagtcttcctcttcccccca720aaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggac780gtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcat840aatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtc900ctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaac960aaagccctcggcgcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaa1020ccacaggtgtgcaccctgcccccatcccgggatgagctgaccaagaaccaggtcagcctc1080tcgtgcgcagtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatggg1140cagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttc1200ctcgtgagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgc1260tccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccg1320ggtaaa1326<210>96<211>642<212>dna<213>人工序列<220><223>lccd3ch2527(crossfab,vl-ch1)<400>96caggccgtcgtgacccaggaacccagcctgacagtgtctcctggcggcaccgtgaccctg60acatgtggcagttctacaggcgccgtgaccaccagcaactacgccaactgggtgcaggaa120aagcccggccaggccttcagaggactgatcggcggcaccaacaagagagcccctggcacc180cctgccagattcagcggatctctgctgggaggaaaggccgccctgacactgtctggcgcc240cagccagaagatgaggccgagtactactgcgccctgtggtacagcaacctgtgggtgttc300ggcggaggcaccaagctgacagtgctgagcagcgcttccaccaaaggcccttccgtgttt360cctctggctcctagctccaagtccacctctggaggcaccgctgctctcggatgcctcgtg420aaggattattttcctgagcctgtgacagtgtcctggaatagcggagcactgacctctgga480gtgcatactttccccgctgtgctgcagtcctctggactgtacagcctgagcagcgtggtg540acagtgcccagcagcagcctgggcacccagacctacatctgcaacgtgaaccacaagccc600agcaacaccaaggtggacaagaaggtggaacccaagtcttgt642<210>97<211>2082<212>dna<213>人工序列<220><223>ceach1a1a98/99-cd3ch2527(crossfabvh-ck)-fc(节)p329glala<400>97caggtgcagctggtgcagtctggcgccgaagtgaagaaacctggcgccagcgtgaaggtg60tcctgcaaggccagcggctacaccttcaccgagttcggcatgaactgggtccgacaggcc120cctggacagggcctggaatggatgggctggatcaacaccaagaccggcgaggccacctac180gtggaagagttcaagggcagagtgaccttcaccaccgacaccagcaccagcaccgcctac240atggaactgcggagcctgagaagcgacgacaccgccgtgtactactgcgccagatgggac300ttcgcctactatgtggaagccatggactactggggccagggcaccaccgtgaccgtgtct360agtgctagcacaaagggccccagcgtgttccctctggcccctagcagcaagagcacatct420ggcggaacagccgccctgggctgcctggtcaaggactactttcccgagcccgtgacagtg480tcctggaactctggcgccctgacaagcggcgtgcacacctttccagccgtgctgcagagc540agcggcctgtactctctgagcagcgtggtcaccgtgcctagctctagcctgggcacccag600acctacatctgcaacgtgaaccacaagcccagcaacaccaaggtggacaagaaggtggaa660cccaagagctgcgatggcggaggcggctccggaggcggaggatccgaggtgcagctgctg720gaatctggcggcggactggtgcagcctggcggatctctgagactgagctgtgccgccagc780ggcttcaccttcagcacctacgccatgaactgggtgcgccaggcccctggcaaaggcctg840gaatgggtgtcccggatcagaagcaagtacaacaactacgccacctactacgccgacagc900gtgaagggccggttcaccatcagccgggacgacagcaagaacaccctgtacctgcagatg960aacagcctgcgggccgaggacaccgccgtgtactattgtgtgcggcacggcaacttcggc1020aacagctatgtgtcttggtttgcctactggggccagggcaccctcgtgaccgtgtcaagc1080gctagcgtggccgctccctccgtgtttatctttcccccatccgatgaacagctgaaaagc1140ggcaccgcctccgtcgtgtgtctgctgaacaatttttaccctagggaagctaaagtgcag1200tggaaagtggataacgcactgcagtccggcaactcccaggaatctgtgacagaacaggac1260tccaaggacagcacctactccctgtcctccaccctgacactgtctaaggctgattatgag1320aaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaag1380agcttcaacaggggagagtgtgacaagacccacacctgtcccccttgtcctgcccctgaa1440gctgctggcggcccttctgtgttcctgttccccccaaagcccaaggacaccctgatgatc1500agccggacccccgaagtgacctgcgtggtggtggatgtgtcccacgaggaccctgaagtg1560aagttcaattggtacgtggacggcgtggaagtgcacaacgccaagacaaagccgcgggag1620gagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactgg1680ctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcggcgcccccatcgag1740aaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgccccca1800tgccgggatgagctgaccaagaaccaggtcagcctgtggtgcctggtcaaaggcttctat1860cccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagacc1920acgcctcccgtgctggactccgacggctccttcttcctctacagcaagctcaccgtggac1980aagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcac2040aaccactacacgcagaagagcctctccctgtctccgggtaaa2082<210>98<211>1353<212>dna<213>人工序列<220><223>ceach1a1a98/99(vh-ch1)-fc(穴)p329glala<400>98caggtgcagctggtgcagtctggcgccgaagtgaagaaacctggagctagtgtgaaggtg60tcctgcaaggccagcggctacaccttcaccgagttcggcatgaactgggtccgacaggct120ccaggccagggcctcgaatggatgggctggatcaacaccaagaccggcgaggccacctac180gtggaagagttcaagggcagagtgaccttcaccacggacaccagcaccagcaccgcctac240atggaactgcggagcctgagaagcgacgacaccgccgtgtactactgcgccagatgggac300ttcgcctattacgtggaagccatggactactggggccagggcaccaccgtgaccgtgtct360agcgctagcaccaagggcccctccgtgttccccctggcccccagcagcaagagcaccagc420ggcggcacagccgctctgggctgcctggtcaaggactacttccccgagcccgtgaccgtg480tcctggaacagcggagccctgacctccggcgtgcacaccttccccgccgtgctgcagagt540tctggcctgtatagcctgagcagcgtggtcaccgtgccttctagcagcctgggcacccag600acctacatctgcaacgtgaaccacaagcccagcaacaccaaggtggacaagaaggtggag660cccaagagctgcgacaaaactcacacatgcccaccgtgcccagcacctgaagctgcaggg720ggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacc780cctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaac840tggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtac900aacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggc960aaggagtacaagtgcaaggtctccaacaaagccctcggcgcccccatcgagaaaaccatc1020tccaaagccaaagggcagccccgagaaccacaggtgtgcaccctgcccccatcccgggat1080gagctgaccaagaaccaggtcagcctctcgtgcgcagtcaaaggcttctatcccagcgac1140atcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctccc1200gtgctggactccgacggctccttcttcctcgtgagcaagctcaccgtggacaagagcagg1260tggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactac1320acgcagaagagcctctccctgtctccgggtaaa1353<210>99<211>645<212>dna<213>人工序列<220><223>lcdp47gs<400>99gaaatcgtgttaacgcagtctccaggcaccctgtctttgtctccaggggaaagagccacc60ctctcttgcagggccagtcagagtgttagcagcagctacttagcctggtaccagcagaaa120cctggccaggctcccaggctcctcatctatggagcatccagcagggccactggcatccca180gacaggttcagtggcagtggatccgggacagacttcactctcaccatcagcagactggag240cctgaagattttgcagtgtattactgtcagcagtatggtagctcaccgctgacgttcggc300caggggaccaaagtggaaatcaaacgtacggtggctgcaccatctgtcttcatcttcccg360ccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttc420tatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcc480caggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctg540acgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcag600ggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt645<210>100<211>642<212>dna<213>人工序列<220><223>lccd3ch2527(crossfab,vl-ch1)<400>100caggccgtcgtgacccaggaacccagcctgacagtgtctcctggcggcaccgtgaccctg60acatgtggcagttctacaggcgccgtgaccaccagcaactacgccaactgggtgcaggaa120aagcccggccaggccttcagaggactgatcggcggcaccaacaagagagcccctggcacc180cctgccagattcagcggatctctgctgggaggaaaggccgccctgacactgtctggcgcc240cagccagaagatgaggccgagtactactgcgccctgtggtacagcaacctgtgggtgttc300ggcggaggcaccaagctgacagtgctgagcagcgcttccaccaaaggcccttccgtgttt360cctctggctcctagctccaagtccacctctggaggcaccgctgctctcggatgcctcgtg420aaggattattttcctgagcctgtgacagtgtcctggaatagcggagcactgacctctgga480gtgcatactttccccgctgtgctgcagtcctctggactgtacagcctgagcagcgtggtg540acagtgcccagcagcagcctgggcacccagacctacatctgcaacgtgaaccacaagccc600agcaacaccaaggtggacaagaaggtggaacccaagtcttgt642<210>101<211>2064<212>dna<213>人工序列<220><223>dp47gs(vh-ch1)-cd3ch2527(crossfabvh-ck)-fc(节)p329glala<400>101gaggtgcaattgttggagtctgggggaggcttggtacagcctggggggtccctgagactc60tcctgtgcagcctccggattcacctttagcagttatgccatgagctgggtccgccaggct120ccagggaaggggctggagtgggtctcagctattagtggtagtggtggtagcacatactac180gcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtat240ctgcagatgaacagcctgagagccgaggacacggccgtatattactgtgcgaaaggcagc300ggatttgactactggggccaaggaaccctggtcaccgtctcgagtgctagcacaaagggc360cccagcgtgttccctctggcccctagcagcaagagcacatctggcggaacagccgccctg420ggctgcctcgtgaaggactactttcccgagcctgtgaccgtgtcctggaactctggcgcc480ctgacaagcggcgtgcacacctttccagccgtgctgcagagcagcggcctgtactctctg540agcagcgtggtcaccgtgcctagcagcagcctgggcacccagacctacatctgcaacgtg600aaccacaagcccagcaacaccaaagtggacaagaaggtggagcccaagagctgtgatggc660ggaggagggtccggaggcggaggatccgaggtgcagctgctggaatctggcggcggactg720gtgcagcctggcggatctctgagactgagctgtgccgccagcggcttcaccttcagcacc780tacgccatgaactgggtgcgccaggcccctggcaaaggcctggaatgggtgtcccggatc840agaagcaagtacaacaactacgccacctactacgccgacagcgtgaagggccggttcacc900atcagccgggacgacagcaagaacaccctgtacctgcagatgaacagcctgcgggccgag960gacaccgccgtgtactattgtgtgcggcacggcaacttcggcaacagctatgtgtcttgg1020tttgcctactggggccagggcaccctcgtgaccgtgtcaagcgctagcgtggccgctccc1080tccgtgtttatctttcccccatccgatgaacagctgaaaagcggcaccgcctccgtcgtg1140tgtctgctgaacaatttttaccctagggaagctaaagtgcagtggaaagtggataacgca1200ctgcagtccggcaactcccaggaatctgtgacagaacaggactccaaggacagcacctac1260tccctgtcctccaccctgacactgtctaaggctgattatgagaaacacaaagtctacgcc1320tgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagag1380tgtgacaagacccacacctgtcccccttgtcctgcccctgaagctgctggcggcccttct1440gtgttcctgttccccccaaagcccaaggacaccctgatgatcagccggacccccgaagtg1500acctgcgtggtggtggatgtgtcccacgaggaccctgaagtgaagttcaattggtacgtg1560gacggcgtggaagtgcacaacgccaagacaaagccgcgggaggagcagtacaacagcacg1620taccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtac1680aagtgcaaggtctccaacaaagccctcggcgcccccatcgagaaaaccatctccaaagcc1740aaagggcagccccgagaaccacaggtgtacaccctgcccccatgccgggatgagctgacc1800aagaaccaggtcagcctgtggtgcctggtcaaaggcttctatcccagcgacatcgccgtg1860gagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggac1920tccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtggcagcag1980gggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaag2040agcctctccctgtctccgggtaaa2064<210>102<211>1335<212>dna<213>人工序列<220><223>dp47gs(vh-ch1)-fc(穴)p329glala<400>102gaggtgcaattgttggagtctgggggaggcttggtacagcctggggggtccctgagactc60tcctgtgcagcctccggattcacctttagcagttatgccatgagctgggtccgccaggct120ccagggaaggggctggagtgggtctcagctattagtggtagtggtggtagcacatactac180gcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacgctgtat240ctgcagatgaacagcctgagagccgaggacacggccgtatattactgtgcgaaaggcagc300ggatttgactactggggccaaggaaccctggtcaccgtctcgagtgctagcaccaagggc360ccctccgtgttccccctggcccccagcagcaagagcaccagcggcggcacagccgctctg420ggctgcctggtcaaggactacttccccgagcccgtgaccgtgtcctggaacagcggagcc480ctgacctccggcgtgcacaccttccccgccgtgctgcagagttctggcctgtatagcctg540agcagcgtggtcaccgtgccttctagcagcctgggcacccagacctacatctgcaacgtg600aaccacaagcccagcaacaccaaggtggacaagaaggtggagcccaagagctgcgacaaa660actcacacatgcccaccgtgcccagcacctgaagctgcagggggaccgtcagtcttcctc720ttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtg780gtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtg840gaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtg900gtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaag960gtctccaacaaagccctcggcgcccccatcgagaaaaccatctccaaagccaaagggcag1020ccccgagaaccacaggtgtgcaccctgcccccatcccgggatgagctgaccaagaaccag1080gtcagcctctcgtgcgcagtcaaaggcttctatcccagcgacatcgccgtggagtgggag1140agcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggc1200tccttcttcctcgtgagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtc1260ttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctcc1320ctgtctccgggtaaa1335<210>103<211>207<212>prt<213>人(homosapiens)<400>103metglnserglythrhistrpargvalleuglyleucysleuleuser151015valglyvaltrpglyglnaspglyasngluglumetglyglyilethr202530glnthrprotyrlysvalserileserglythrthrvalileleuthr354045cysproglntyrproglysergluileleutrpglnhisasnasplys505560asnileglyglyaspgluaspasplysasnileglyseraspgluasp65707580hisleuserleulysgluphesergluleugluglnserglytyrtyr859095valcystyrproargglyserlysprogluaspalaasnphetyrleu100105110tyrleuargalaargvalcysgluasncysmetglumetaspvalmet115120125servalalathrilevalilevalaspilecysilethrglyglyleu130135140leuleuleuvaltyrtyrtrpserlysasnarglysalalysalalys145150155160provalthrargglyalaglyalaglyglyargglnargglyglnasn165170175lysgluargproproprovalproasnproasptyrgluproilearg180185190lysglyglnargaspleutyrserglyleuasnglnargargile195200205<210>104<211>198<212>prt<213>猕猴(macacafascicularis)<400>104metglnserglythrargtrpargvalleuglyleucysleuleuser151015ileglyvaltrpglyglnaspglyasngluglumetglyserilethr202530glnthrprotyrglnvalserileserglythrthrvalileleuthr354045cysserglnhisleuglyserglualaglntrpglnhisasnglylys505560asnlysgluaspserglyaspargleupheleuproglupheserglu65707580metgluglnserglytyrtyrvalcystyrproargglyserasnpro859095gluaspalaserhishisleutyrleulysalaargvalcysgluasn100105110cysmetglumetaspvalmetalavalalathrilevalilevalasp115120125ilecysilethrleuglyleuleuleuleuvaltyrtyrtrpserlys130135140asnarglysalalysalalysprovalthrargglyalaglyalagly145150155160glyargglnargglyglnasnlysgluargproproprovalproasn165170175proasptyrgluproilearglysglyglnglnaspleutyrsergly180185190leuasnglnargargile195<210>105<211>15<212>prt<213>人工序列<220><223>接头<400>105gluprolyssercysglyglyglyglyserglyglyglyglyser151015<210>106<211>15<212>prt<213>人工序列<220><223>接头<400>106gluprolyssercysglyglyglyglyserglyglyglyglyser151015<210>107<211>227<212>prt<213>人(homosapiens)<400>107asplysthrhisthrcysproprocysproalaprogluleuleugly151015glyproservalpheleupheproprolysprolysaspthrleumet202530ileserargthrprogluvalthrcysvalvalvalaspvalserhis354045gluaspprogluvallyspheasntrptyrvalaspglyvalgluval505560hisasnalalysthrlysproargglugluglntyrasnserthrtyr65707580argvalvalservalleuthrvalleuhisglnasptrpleuasngly859095lysglutyrlyscyslysvalserasnlysalaleuproalaproile100105110glulysthrileserlysalalysglyglnproarggluproglnval115120125tyrthrleuproproserargaspgluleuthrlysasnglnvalser130135140leuthrcysleuvallysglyphetyrproseraspilealavalglu145150155160trpgluserasnglyglnprogluasnasntyrlysthrthrpropro165170175valleuaspseraspglyserphepheleutyrserlysleuthrval180185190asplysserargtrpglnglnglyasnvalphesercysservalmet195200205hisglualaleuhisasnhistyrthrglnlysserleuserleuser210215220proglylys225<210>108<211>19<212>prt<213>人工序列<220><223>前导肽<400>108metasptrpthrtrpargileleupheleuvalalaalaalathrgly151015alahisser<210>109<211>57<212>dna<213>人工序列<220><223>前导肽<400>109atggactggacctggagaatcctcttcttggtggcagcagccacaggagcccactcc57<210>110<211>57<212>dna<213>人工序列<220><223>前导肽<400>110atggactggacctggaggatcctcttcttggtggcagcagccacaggagcccactcc57<210>111<211>22<212>prt<213>人工序列<220><223>前导肽<400>111metaspmetargvalproalaglnleuleuglyleuleuleuleutrp151015pheproglyalaargcys20<210>112<211>66<212>dna<213>人工序列<220><223>前导肽<400>112atggacatgagggtccccgctcagctcctgggcctcctgctgctctggttcccaggtgcc60aggtgt66<210>113<211>19<212>prt<213>人工序列<220><223>前导肽<400>113metglytrpsercysileileleupheleuvalalathralathrgly151015valhisser<210>114<211>57<212>dna<213>人工序列<220><223>前导肽<400>114atgggatggagctgtatcatcctcttcttggtagcaacagctaccggtgtgcattcc57<210>115<211>57<212>dna<213>人工序列<220><223>前导肽<400>115atgggctggtcctgcatcatcctgtttctggtggctaccgccactggagtgcattcc57<210>116<211>57<212>dna<213>人工序列<220><223>前导肽<400>116atgggctggtcctgcatcatcctgtttctggtcgccacagccaccggcgtgcactct57<210>117<211>643<212>prt<213>猕猴(macacafascicularis)<400>117leuserleugluglyserargthrleuthrvalcysproglyserval151015glnproleuserserglnthrleuargalaserserseralaglythr202530aspproglnleuleuleutyrargvalvalargglyproglnleugly354045argleuphehisalaglnglnaspserthrglyglualaleuvalasn505560phethrglnalagluvaltyralaglyasnileleutyrgluhisglu65707580metprothrgluprophetrpglualahisaspthrleugluleugln859095leuserserproproalaargaspvalalaalathrleualavalala100105110valserpheglualaalacysproglnargproserhisleutrplys115120125asnlysglyleutrpvalprogluglyglnargalalysilethrmet130135140alaalaleuaspalaserasnleuleualaservalproserprogln145150155160argleugluhisaspvalleupheglnvalthrglnpheproserarg165170175glyglnleuleuvalserglugluproleuhisalaglyglnprohis180185190pheleuglnserglnleualaalaglyglnleuvaltyralahisgly195200205glyglyglythrglnglnaspglyphehispheargalahisleugln210215220glyproalaglyalathrvalalaglyproglnthrserglualaphe225230235240alailethrvalargaspvalasngluargproproglnproglnala245250255servalproleuargilethrargglyserargalaproileserarg260265270alaglnleuservalvalaspproaspseralaproglygluileglu275280285tyrgluvalglnargalaprohisasnglypheleuserleuvalgly290295300glyglyproglyprovalthrhisphethrglnalaaspvalaspser305310315320glyargleualaphevalalaasnglyserservalalaglyvalphe325330335glnleusermetseraspglyalaserproproleuprometserleu340345350alavalaspileleuproseralailegluvalglnleuglnalapro355360365leugluvalproglnalaleuglyargserserleuserglnglngln370375380leuargvalvalseraspargglugluproglualaalatyrargleu385390395400ileglnglyprolystyrglyhisleuleuvalglyglyargproala405410415seralapheserglnleuglnileaspglnglygluvalvalpheala420425430phethrasnpheserserserhisasphispheargvalleualaleu435440445alaargglyvalasnalaseralavalvalasnilethrvalargala450455460leuleuhisvaltrpalaglyglyprotrpproglnglyalathrleu465470475480argleuaspprothrileleuaspalaglygluleualaasnargthr485490495glyservalprohispheargleuleugluglyproarghisglyarg500505510valvalargvalproargalaargthrgluproglyglyserglnleu515520525valgluglnphethrglnglnaspleugluaspglyargleuglyleu530535540gluvalglyargprogluglyargalaproserprothrglyaspser545550555560leuthrleugluleutrpalaglnglyvalproproalavalalaser565570575leuaspphealathrgluprotyrasnalaalaargprotyrserval580585590alaleuleuservalproglualathrargmetglualaglylyspro595600605gluserserthrprothrglygluproglyprometalaserserpro610615620valproalavalalalysglyglypheleuglypheleuglualaasn625630635640metpheser<210>118<211>643<212>prt<213>人(homosapiens)<400>118leuserleulysglyserglnthrleuthrvalcysproglyserval151015glnproleuserserglnthrleuargalaserserseralaglythr202530aspproglnleuleuleutyrargvalvalargglyproglnleugly354045argleuphehisalaglnglnaspserthrglyglualaleuvalasn505560phethrglnalagluvaltyralaglyasnileleutyrgluhisglu65707580metproprogluprophetrpglualahisaspthrleugluleugln859095leuserserproproalaargaspvalalaalathrleualavalala100105110valserpheglualaalacysproglnhisproserhisleutrplys115120125asnlysglyleutrpvalprogluglyglnargalaargilethrval130135140alaalaleuaspalaserasnleuleualaservalproserprogln145150155160argsergluhisaspvalleupheglnvalthrglnpheproserarg165170175glyglnleuleuvalserglugluproleuhisalaglyglnprohis180185190pheleuglnserglnleualaalaglyglnleuvaltyralahisgly195200205glyglyglythrglnglnaspglyphehispheargalahisleugln210215220glyproalaglyalaservalalaglyproglnthrserglualaphe225230235240alailethrvalargaspvalasngluargproproglnproglnala245250255servalproleuargleuthrargglyserargalaproileserarg260265270alaglnleuservalvalaspproaspseralaproglygluileglu275280285tyrgluvalglnargalaprohisasnglypheleuserleuvalgly290295300glyglyleuglyprovalthrargphethrglnalaaspvalaspser305310315320glyargleualaphevalalaasnglyserservalalaglyilephe325330335glnleusermetseraspglyalaserproproleuprometserleu340345350alavalaspileleuproseralailegluvalglnleuargalapro355360365leugluvalproglnalaleuglyargserserleuserglnglngln370375380leuargvalvalseraspargglugluproglualaalatyrargleu385390395400ileglnglyproglntyrglyhisleuleuvalglyglyargprothr405410415seralapheserglnpheglnileaspglnglygluvalvalpheala420425430phethrasnpheserserserhisasphispheargvalleualaleu435440445alaargglyvalasnalaseralavalvalasnvalthrvalargala450455460leuleuhisvaltrpalaglyglyprotrpproglnglyalathrleu465470475480argleuaspprothrvalleuaspalaglygluleualaasnargthr485490495glyservalproargpheargleuleugluglyproarghisglyarg500505510valvalargvalproargalaargthrgluproglyglyserglnleu515520525valgluglnphethrglnglnaspleugluaspglyargleuglyleu530535540gluvalglyargprogluglyargalaproglyproalaglyaspser545550555560leuthrleugluleutrpalaglnglyvalproproalavalalaser565570575leuaspphealathrgluprotyrasnalaalaargprotyrserval580585590alaleuleuservalproglualaalaargthrglualaglylyspro595600605gluserserthrprothrglygluproglyprometalaserserpro610615620gluproalavalalalysglyglypheleuserpheleuglualaasn625630635640metpheser<210>119<211>428<212>prt<213>人工序列<220><223>naba-avi-his<400>119glnleuthrthrglusermetpropheasnvalalagluglylysglu151015valleuleuleuvalhisasnleuproglnglnleupheglytyrser202530trptyrlysglygluargvalaspglyasnargglnilevalglytyr354045alaileglythrglnglnalathrproglyproalaasnserglyarg505560gluthriletyrproasnalaserleuleuileglnasnvalthrgln65707580asnaspthrglyphetyrthrleuglnvalilelysseraspleuval859095asngluglualathrglyglnphehisvaltyrprogluleuprolys100105110proserileserserasnasnserasnprovalgluasplysaspala115120125metalaphethrcysgluprogluthrglnaspthrthrtyrleutrp130135140trpileasnasnglnserleuprovalserproargleuglnleuser145150155160asnglyasnargthrleuthrleuleuservalthrargasnaspthr165170175glyprotyrglucysgluileglnasnprovalseralaasnargser180185190aspprovalthrleuasnvalthrtyrglyproaspthrprothrile195200205serproproaspsersertyrleuserglyalaasnleuasnleuser210215220cyshisseralaserasnproserproglntyrsertrpargileasn225230235240glyileproglnglnhisthrglnvalleupheilealalysilethr245250255proasnasnasnglythrtyralacysphevalserasnleualathr260265270glyargasnasnserilevallysserilethrvalseralaleuser275280285provalvalalalysproglnilelysalaserlysthrthrvalthr290295300glyasplysaspservalasnleuthrcysserthrasnaspthrgly305310315320ileserileargtrpphephelysasnglnserleuproserserglu325330335argmetlysleuserglnglyasnilethrleuserileasnproval340345350lysarggluaspalaglythrtyrtrpcysgluvalpheasnproile355360365serlysasnglnseraspproilemetleuasnvalasntyrasnala370375380leuproglngluasnleuileasnvalaspleugluvalleuphegln385390395400glyproglyserglyleuasnaspilepheglualaglnlysileglu405410415trphisglualaargalahishishishishishis420425<210>120<211>360<212>prt<213>人工序列<220><223>cd3e干-fc(节)-avi<400>120glnaspglyasngluglumetglyglyilethrglnthrprotyrlys151015valserileserglythrthrvalileleuthrcysproglntyrpro202530glysergluileleutrpglnhisasnasplysasnileglyglyasp354045gluaspasplysasnileglyseraspgluasphisleuserleulys505560gluphesergluleugluglnserglytyrtyrvalcystyrproarg65707580glyserlysprogluaspalaasnphetyrleutyrleuargalaarg859095valsergluasncysvalaspgluglnleutyrpheglnglyglyser100105110prolysseralaasplysthrhisthrcysproprocysproalapro115120125gluleuleuglyglyproservalpheleupheproprolysprolys130135140aspthrleumetileserargthrprogluvalthrcysvalvalval145150155160aspvalserhisgluaspprogluvallyspheasntrptyrvalasp165170175glyvalgluvalhisasnalalysthrlysproargglugluglntyr180185190asnserthrtyrargvalvalservalleuthrvalleuhisglnasp195200205trpleuasnglylysglutyrlyscyslysvalserasnlysalaleu210215220proalaproileglulysthrileserlysalalysglyglnproarg225230235240gluproglnvaltyrthrleuproprocysargaspgluleuthrlys245250255asnglnvalserleutrpcysleuvallysglyphetyrproserasp260265270ilealavalglutrpgluserasnglyglnprogluasnasntyrlys275280285thrthrproprovalleuaspseraspglyserphepheleutyrser290295300lysleuthrvalasplysserargtrpglnglnglyasnvalpheser305310315320cysservalmethisglualaleuhisasnhistyrthrglnlysser325330335leuserleuserproglylysserglyglyleuasnaspilepheglu340345350alaglnlysileglutrphisglu355360<210>121<211>325<212>prt<213>人工序列<220><223>cd3d干-fc(穴)<400>121phelysileproileglugluleugluaspargvalphevalasncys151015asnthrserilethrtrpvalgluglythrvalglythrleuleuser202530aspilethrargleuaspleuglylysargileleuaspproarggly354045iletyrargcysasnglythraspiletyrlysasplysgluserthr505560valglnvalhistyrargmetcysargsergluglnleutyrphegln65707580glyasplysthrhisthrcysproprocysproalaprogluleuleu859095glyglyproservalpheleupheproprolysprolysaspthrleu100105110metileserargthrprogluvalthrcysvalvalvalaspvalser115120125hisgluaspprogluvallyspheasntrptyrvalaspglyvalglu130135140valhisasnalalysthrlysproargglugluglntyrasnserthr145150155160tyrargvalvalservalleuthrvalleuhisglnasptrpleuasn165170175glylysglutyrlyscyslysvalserasnlysalaleuproalapro180185190ileglulysthrileserlysalalysglyglnproarggluprogln195200205valcysthrleuproproserargaspgluleuthrlysasnglnval210215220serleusercysalavallysglyphetyrproseraspilealaval225230235240glutrpgluserasnglyglnprogluasnasntyrlysthrthrpro245250255provalleuaspseraspglyserphepheleuvalserlysleuthr260265270valasplysserargtrpglnglnglyasnvalphesercysserval275280285methisglualaleuhisasnhistyrthrglnlysserleuserleu290295300serproglylysserglyglyleuasnaspilepheglualaglnlys305310315320ileglutrphisglu325<210>122<211>658<212>prt<213>小家鼠(musmusculus)<400>122leuserleugluglythrarglysleuthrvalcysprogluserval151015glnproleuserserglnserleuseralaserserserthrglyala202530aspproarghisleuleutyrargvalvalargglyproglnleugly354045argleuleuhisalaglnglnglyseralaglugluvalleuvalasn505560phethrglnalagluvalasnalaglyasnileleutyrgluhisglu65707580metsersergluprophetrpglualahisaspthrileglyleuleu859095leuserserproproalaargaspleualaalathrleualavalmet100105110valserpheaspalaalacysproglnargproserargleutrplys115120125asnlysglyleutrpvalprogluglyglnargalalysilethrval130135140alaalaleuaspalaalaasnleuleualaservalproalasergln145150155160argserarghisaspvalleupheglnvalthrglnpheprothrarg165170175glyglnleuleuvalserglugluproleuhisalaargargprotyr180185190pheleuglnsergluleualaalaglyglnleuvaltyralahisgly195200205glyglyglythrglnglnaspglypheargpheargalahisleugln210215220glyprothrglythrservalalaglyproglnthrserglualaphe225230235240valilethrvalargaspvalasngluargproproglnproglnala245250255serileproleuargvalthrargglyserargalaprovalserarg260265270alaglnleuservalvalaspproaspseralaproglygluileglu275280285tyrgluvalglnargalaprohisasnglypheleuserleualagly290295300aspasnthrglyprovalthrhisphethrglnalaaspvalaspala305310315320glyargleualaphevalalaasnglyserservalalaglyvalphe325330335glnleusermetseraspglyalaserproproileprometserleu340345350alavalaspvalleuproserthrilegluvalglnleuargalapro355360365leugluvalproglnalaleuglyargthrserleuserargglngln370375380leuglnvalileseraspargglugluproaspvalalatyrargleu385390395400thrglnglyproleutyrglyglnleuleuvalglyglyglnproala405410415seralapheserglnleuglnvalaspglnglyaspvalvalpheval420425430phethrasnpheserserserglnasphisphelysvalvalalaleu435440445alaargglyvalasnalaseralathrvalasnvalthrvalglnala450455460leuleuhisvaltrpalaglyglyprotrpproglnglythrthrleu465470475480argleuaspprothrvalleuaspalasergluleualaasnargthr485490495glysermetprohispheargleuleualaglyproargtyrglyarg500505510valvalargvalserglnglyargthrgluserargserasnglnleu515520525valgluhisphethrglnargaspleuglugluglyglnleuglyleu530535540gluvalglylysprogluglyargserthrglyproalaglyasparg545550555560leuthrleugluleutrpalalysglyvalproproalavalalaleu565570575leuaspphealathrgluprotyrhisalaalalyssertyrserval580585590alaleuleuservalproglualavalargthrgluthrglulyspro595600605glyargservalprothrglyglnproglyglnalaalaserserpro610615620valprothralaalalysglyglyvalaspglyleuasnaspilephe625630635640glualaglnlysileglutrphisglualaargalahishishishis645650655hishis当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1