阿魏菇多糖作为制备树突状细胞疫苗佐剂的应用的制作方法
本发明属于医药领域,具体涉及阿魏菇多糖作为制备树突状细胞疫苗佐剂的应用。
背景技术:
树突状细胞(dendritic cell;DC)是机体专职的抗原递呈细胞,可以识别、捕获和加工抗原,并将抗原递呈给naïve T细胞,激活抗原特异性的免疫反应,从而连接固有免疫系统和获得性免疫系统。在抗原递呈过程中,不同的信号参与了抗原特异性T细胞的激活,包括MHC-多肽复合物、DC和T细胞表面共刺激分子之间的相互作用、DC分泌的细胞因子等。DC分泌的白细胞介素-12(IL-12)可促进CD4+ Th1型免疫反应及细胞毒性T淋巴细胞反应,这两种免疫反应在肿瘤治疗中起着重要作用。DC疫苗(DC-based vaccine)是一种具有前景的肿瘤治疗性疫苗形式,在临床上可以诱导产生抗原特异性的免疫反应,安全性高。然而,由于DC未达到最佳成熟状态,并且分泌的IL-12水平较低,导致DC疫苗的临床效果有限。因此,需要通过佐剂提高DC成熟状态及IL-12分泌水平,从而提高DC疫苗的临床效果。
佐剂(如钟样受体激动剂:toll-like receptor (TLR) agonists)被广泛应用于新型疫苗(如多肽/蛋白疫苗、DNA疫苗及DC疫苗等),以提高疫苗的免疫原性。但是大部分佐剂具有毒副作用,不适合临床应用。目前美国FDA及欧洲批准用于临床应用的佐剂种类非常少,仅包括铝化合物佐剂、MF59、AS03、AF03及AS04。因此,急需开发新型、安全、高效的免疫佐剂增强疫苗的免疫原性,提高抗原特异性免疫反应,发挥疫苗良好的预防和治疗效果。
中草药已经有几千年的临床应用历史,临床效果与其有效成分对机体的免疫调节作用,尤其是对抗原递呈细胞的成熟状态及功能的调节密切相关,其安全性得到了充分的验证,并具有多效性,无依赖性等特征,是筛选安全、高效佐剂的理想来源。
技术实现要素:
本发明的目的是将从中药阿魏菇(Pleurotus ferulae)中提取纯化得到的阿魏菇多糖(PFPS)作为树突状细胞疫苗佐剂,刺激树突状细胞成熟,从而促进树突状细胞疫苗免疫反应,提高抗肿瘤效果。
为了解决上述技术问题,本发明提供了如下的技术方案:
阿魏菇多糖作为制备树突状细胞疫苗佐剂的应用。
优选地,所述阿魏菇多糖为均一多糖,分子量为1500~1600kDa。
阿魏菇多糖的提取纯化方法,包括:
(1)采用水提醇沉法得到阿魏菇粗多糖;
(2)将阿魏菇粗多糖溶解于水中,然后过二乙氨基乙基纤维素柱,流动相为水和/或盐水。
优选地,所述盐水为浓度为0.05~0.3mol/L的氯化钠溶液。
优选地,所述盐水为浓度为0.05mol/L的氯化钠溶液。
附图说明
附图用来提供对本发明的进一步理解,并且构成说明书的一部分,与本发明的实施例一起用于解释本发明,并不构成对本发明的限制。在附图中:
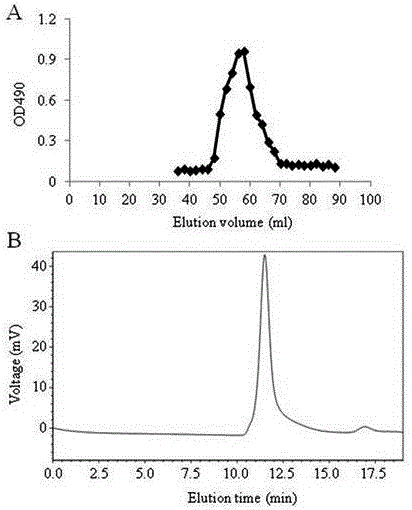
图1 (A)阿魏菇粗多糖的洗脱曲线及洗脱液多糖含量,(B)不同流动相所得的洗脱液体外刺激DC成熟的活性。
图2 (A)为PFPS-0.05的纯化及分子量检测过丙烯葡聚糖凝胶S-300HR的洗脱曲线,(B)PFPS-0.05的高效凝胶渗透色谱图。
图3 为PFPS-0.05对DC成熟及细胞因子表达的影响。收集体外培养的不成熟DC,用10、50、100 μg/ml的PFPS-0.05 或者 20 ng/ml的LPS刺激12小时。(A)通过流式细胞术检测DC表面CD40、CD80、 CD86和MHC II的表达。图中显示了这些蛋白的平均荧光强度(MFI)。(B)酶联免疫吸附法(ELISA)测定DC培养上清中白细胞介素-12(IL-12)及肿瘤坏死因子-α(TNF-α)的表达量。数据来自4个独立的实验,并进行单因素方差分析,*P<0.05、**P<0.01、***P <0.001,处理组与未处理组比较得出。
图4 为TLR4抑制剂对DC成熟及细胞因子表达的影响。收集体外培养的不成熟DC,用TLR4抑制剂(TAK-242)进行预处理,然后用10、50 、100 μg/ml的PFPS-0.05 或者 20 ng/ml的LPS刺激12小时。(A)通过流式细胞术检测DC表面CD40的表达。上边重叠峰图为CD40表达水平,下边为CD40的平均荧光强度(MFI)。(B)ELISA测定DC培养上清中IL-12及TNF-α的表达量。数据来自3个独立的实验,并进行成对t检验。
图5 为TLR4信号通路分子磷酸化水平检测。收集体外培养的不成熟DC,用50 μg/ml的PFPS-0.05进行刺激,在不同时间点收集DC,提取细胞质及细胞核蛋白,通过Western blot检测TLR4信号通路分子的磷酸化水平。
图6 为PFPS-0.05刺激的DC与人乳头瘤病毒(HPV)多肽孵育后(HPV+PFPS+DC)对肿瘤的治疗效果。肿瘤模型建立后,治疗分为早期治疗及晚期治疗,早期治疗组在第5天及第12天注射HPV+PFPS+DC,晚期治疗组在第12天及第19天注射HPV+PFPS+DC。对照组分别为在第5天及第12天注射PFPS+DC,或不治疗组。(A)左边为肿瘤体积生长曲线,右边为计算的曲线下面积(AUC)。(B)实验结束后,分离肿瘤并称重。数据进行单因素方差分析,*P<0.05、**P<0.01,治疗组与不治疗组比较得出。
图7 为肿瘤小鼠脾脏中CD4+和CD8+ T细胞比例及亚群比例。实验结束后,分离脾脏细胞,通过流式细胞术检测CD4+(A)和CD8+(B)T细胞比例及亚群比例。*P<0.05、**P<0.01,治疗组与不治疗组比较得出。(C)CD4+和CD8+ Tem细胞比例与肿瘤体积相关性。
图8 为HPV特异性细胞免疫反应及骨髓来源的抑制性细胞(MDSCs)和巨噬细胞的比例。实验结束后,分离脾脏细胞。(A)脾脏细胞用HPV多肽刺激过夜,通过流式细胞术检测HPV特异性细胞免疫反应。(B)CD8+ IFN-γ+ 细胞比例与肿瘤体积相关性。(C)MDSCs(CD11b+Gr-1+)及巨噬细胞(CD11b+Gr-1-)在肿瘤小鼠脾脏中的比例。(D)MDSCs比例与肿瘤体积相关性。*P<0.05、**P<0.01、***P<0.001,治疗组与不治疗组比较得出。
具体实施方式
以下结合附图对本发明的优选实施例进行说明,应当理解,此处所描述的优选实施例仅用于说明和解释本发明,并不用于限定本发明。
阿魏菇多糖的提取纯化
称取100g阿魏菇冻干粉,用1 L 95%乙醇浸提2小时去脂,5000 rpm/min离心10 min收集沉淀,沉淀用1 L去离子水重悬,60℃水浴2小时,超声20min助溶,6000 rpm/min离心15 min,收集上清,滤渣反复浸提4次后,合并上清液,在60℃水浴温度下真空旋转浓缩至1/10体积后,用Sevage试剂法:按浓缩液:Sevage试剂(氯仿:正丁醇=4:1)=3:1比例混合,磁力搅拌30min后,6000rpm/min 离心10 min,保留上清,反复脱蛋白6次,所得上清加无水乙醇至终浓度为80%, 4℃过夜,6000rpm/min离心10 min,沉淀用无水乙醇,丙酮各洗涤一次,自然干燥3小时后得粗多糖。
称取360 mg粗多糖重新溶解于去离子水中(浓度约5 mg/ml),经0.22 μm超滤杯过滤后,真空旋转浓缩至终浓度约为20 mg/ml,过DEAE-52纤维素柱层析(2.6×66 cm),依次用去离子水、0.05 M、0.1 M和0.3 M NaCl溶液阶段洗脱柱子,流速为1.0 ml/min,5 ml/tube收集洗脱液,采用苯酚硫酸法在490 nm处测定多糖含量,绘制洗脱曲线(图1A)。按照洗脱峰收集合并组分,透析冷冻干燥,分别命名为PFPS-W、PFPS-0.05、PFPS-0.1及PFPS-0.3,多糖纯度分别为59%、96%、70%及35%。
阿魏菇多糖刺激DC成熟活性检测
以不同浓度的纯化的PFPS处理体外培养的不成熟DC,12小时后通过流式细胞术检测DC表面CD40的表达(图1B)。可以看出,PFPS-W、PFPS-0.05、PFPS-0.1均具有刺激DC成熟的活性,其中盐洗脱的多糖刺激DC成熟的活性显著高于水洗脱的多糖。
PFPS-0.05的分子量测定
基于多糖纯度及刺激DC成熟活性,我们选取0.05 M NaCl洗脱的PFPS-0.05进行进一步的分析。PFPS-0.05经0.22 μm滤膜过滤后,过丙烯葡聚糖凝胶S-300HR,以去离子水进行洗脱,3 ml/tube收集洗脱液,流速为0.5 ml/min, 苯酚硫酸法测定吸光值,绘制洗脱曲线。根据洗脱曲线,得到一个纯洗脱峰(图2A)。我们采用高效凝胶渗透色谱法(high performance gel-permeation chromatography)检测了PFPS-0.05的均一度及分子量检测(图2B)。移动相为0.71%的硫酸钠,流动速率为0.5 ml/min。结果显示,PFPS-0.05为成分均一的多糖。以分子量为9.75, 36.8, 135.35, 300.6 and 2000 kDa的右旋糖苷标准品制备标准曲线,计算PFPS-0.05的分子量大约为1500~1600 kDa。
PFPS-0.05促进DC成熟及细胞因子表达
以不同浓度(10、50和100 mg/ml)的PFPS-0.05刺激体外培养的不成熟DC,12小时后收集细胞,采用流式细胞术检测DC表面CD40、CD80、CD86和MHC II分子的表达。如图3A所示,PFPS-0.05显著提高了CD40及CD86的表达水平,并呈剂量依赖性。CD80及MHC II的表达水平也有一定的提高。收集上述细胞培养上清,采用酶联免疫吸附法(ELISA)测定白细胞介素-12(IL-12)及肿瘤坏死因子-α(TNF-α)的表达量。如图3B所示,PFPS-0.05显著增加了IL-12及TNF-α的分泌水平,水平与阳性对照细菌脂多糖(LPS)相当。结果显示,PFPS-0.05促进了DC的成熟及细胞因子表达。
PFPS-0.05通过TLR4信号通路刺激DC成熟
为了验证PFPS-0.05是否通过TLR4信号通路刺激DC成熟,我们用TLR4抑制剂(TAK-242)对DC进行预处理,然后用不同浓度的PFPS-0.05或LPS刺激。12小时后检测CD40表达及细胞因子分泌。如图4A&B所示,TAK-242预处理显著抑制了PFPS-0.05及LPS诱导的CD40的表达及IL-12和TNF-α的分泌水平,结果暗示PFPS-0.05通过TLR4信号通路刺激了DC成熟。
我们进一步检测了TLR4信号通路下游分子的活性状态。50 mg/ml的PFPS-0.05处理后,在不同时间点收集细胞,提取细胞质及细胞核蛋白,通过Western blot检测TLR4信号通路分子的磷酸化水平。如图5所示,JNK和ERK在10 min时被磷酸化,30 min时磷酸化水平开始降低。p38也在10 min时被磷酸化,并持续到了240 min。我们也观察到,在30 min时被磷酸化并且持续到了240 min。结果显示,PFPS-0.05激活了TLR4下游MAPK及信号通路。
PFPS-0.05刺激的DCs与人乳头瘤病毒(HPV)多肽孵育后(HPV+PFPS+DCs)抑制了肿瘤的生长
为了检测PFPS-0.05刺激DC的抗肿瘤效果,我们用TC-1细胞建立了小鼠肿瘤模型。肿瘤模型建立后,治疗分为早期治疗及晚期治疗,早期治疗组在第5天及第12天注射HPV+PFPS+DCs(HPV+PFPS+DCs early),晚期治疗组在第12天及第19天注射HPV+PFPS+DCs(HPV+PFPS+DCs late)。对照组分别为在第5天及第12天注射PFPS+DCs,或不治疗组。每组8只小鼠,每隔一天测量一次肿瘤大小。对照组在27天时死了一只小鼠,肿瘤体积为3097 mm3,PFPS+DCs组在17天时死了一只小鼠,肿瘤体积为242 mm3,HPV+PFPS+DCs late组在31天时死了一只小鼠,肿瘤体积为1742 mm3,而HPV+PFPS+DCs early组所有小鼠均存活到了实验结束。如图6A所示,与对照组相比,早期治疗组和晚期治疗组均显著抑制了肿瘤的生长,抑制率分别为93%和62%。实验结束后,处死肿瘤小鼠,分离肿瘤,拍照并称重。如图6B所示,早期治疗组和晚期治疗组的肿瘤显著小于对照组,肿瘤重量也显著低于对照组。
HPV+PFPS+DCs改变了T细胞的数量及活性状态,诱导产生了HPV特异性的细胞免疫反应,并且降低了骨髓来源的抑制性细胞(MDSCs)的比例
采用流式细胞术,检测了肿瘤小鼠脾脏中T细胞的数量及活性状态,HPV特异性的细胞免疫反应及MDSCs的比例。如图7A&B所示,与对照组相比,早期治疗组和晚期治疗组小鼠脾脏中CD4+和CD8+ T细胞的比例显著上升。以CD44和CD62L为标志物,检测T细胞活性状态。与对照组相比,HPV+PFPS+DCs early组CD4+ Tem(效应记忆细胞),CD8+ Tem和Tcm(中央记忆细胞)细胞的比例显著上升。进而发现,CD4+和CD8+ Tem细胞的比例与肿瘤体积呈显著负相关性(图7C)。
我们接下来检测了HPV特异性的细胞免疫反应,HPV+PFPS+DCs early及late在一定程度上增加了CD4+ T细胞反应,但显著增加了CD8+ T细胞反应(图8A),并与肿瘤体积呈显著负相关性(图8B)。与对照组相比,HPV+PFPS+DCs early显著降低了MDSCs的比例,但没有改变巨噬细胞的比例(图8C)。我们也观察到MDSCs的比例与肿瘤体积呈显著正相关性(图8D)。结果显示,HPV+PFPS+DCs增强了T细胞的活性状态,诱导产生了HPV特异性的细胞免疫反应,降低了MDSCs的比例,从而抑制了肿瘤的生长。
最后应说明的是:以上所述仅为本发明的优选实施例而已,并不用于限制本发明,尽管参照前述实施例对本发明进行了详细的说明,对于本领域的技术人员来说,其依然可以对前述各实施例所记载的技术方案进行修改,或者对其中部分技术特征进行等同替换。凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
- 还没有人留言评论。精彩留言会获得点赞!