NY-ESO-1有作为增强CA9免疫原性分子佐剂的作用的制作方法
本发明涉及生物医药领域,特别是涉及NY-ESO-1有作为增强CA9免疫原性分子佐剂的作用。
背景技术:
据推断,最开始对于肿瘤特异性抗原(TAA)的自身免疫应答反应可能有先天性免疫系统受体的参与。坏死肿瘤细胞会释放内源性因子或是例如热休克蛋白,高迁移率族蛋白B1(HMGB-1),尿酸这样的损伤相关模式分子。这些损伤模式分子分别通过CD91,晚期糖基化终末产物受体,TLR2/TLR4,以及白介素-1(IL-1)受体向先天免疫系统发出警报。TAA本身通常被认为是上述“危险信号”的旁观者,担任抗肿瘤免疫应答反应起始阶段的佐剂。根据这个例子可以说明,自发的免疫反应是由于存在于癌细胞中11000个基因突变而产生的新肽所引起的。然而,迄今为止,所发现的人类的TAA通常是非突变的自身蛋白。我们因此推断,来自于一些已知的TAA的内在因子也可能在体内最初阶段的抗肿瘤免疫应答反应中发挥着作用。
在寻找那些宿主和肿瘤中有助于促成抗肿瘤的免疫反应的内在因子的过程中,我们将关注点放在了NY-ESO-1身上。NY-ESO-1是一种具有超强免疫原性的非突变肿瘤睾丸抗原。针对于NY-ESO-1的自发性抗体以及T细胞免疫反应能够在患有NY-ESO-1表达的肿瘤病人身上常常检测的到,这些病人包括那些免疫能力低的癌症晚期的老年患者。此外,自发性NY-ESO-1抗原在黑色素瘤中的减少,表明了抗NY-ESO-1的免疫反应可能会影响肿瘤的自然史。与其他TAA相比,NY-ESO-1的免疫原性不是由于其更高的表达水平。事实上,至少在黑色素肿瘤中的NY-ESO-1的表达水平要显著低于黑色素细胞分化抗原,如gp100,MART-1,TRP-1和TRP-2,,以及其他的肿瘤/睾丸抗原,如基于新鲜肿瘤样本实时定量PCR分析的MAGE-1和MAGE-3。
我们最近研究了NY-ESO-1与在不成熟的树突细胞,巨噬细胞,单核细胞表面上补体Cq1受体(在文献中被视为等同于钙网织蛋白)间特殊的联系,从而说明了NY-ESO-1与先天性免疫系统之间的特异性相互作用。在这篇文章中,说明了NY-ESO-1的免疫原性,与它自身的聚合结构以及小鼠体内中的TLR4有关。
技术实现要素:
据推断,最开始对于肿瘤特异性抗原(TAA)的自身免疫应答反应可能有先天性免疫系统受体的参与。坏死肿瘤细胞会释放内源性因子或是例如热休克蛋白,高迁移率族蛋白B1(HMGB-1),尿酸这样的损伤相关模式分子。这些损伤模式分子分别通过CD91,晚期糖基化终末产物受体,TLR2/TLR4,以及白介素-1(IL-1)受体向先天免疫系统发出警报。TAA本身通常被认为是上述“危险信号”的旁观者,担任抗肿瘤免疫应答反应起始阶段的佐剂。在寻找那些宿主和肿瘤中有助于促成抗肿瘤的免疫反应的内在因子的过程中,我们将关注点放在了NY-ESO-1身上。NY-ESO-1是一种具有超强免疫原性的非突变肿瘤睾丸抗原。针对于NY-ESO-1的自发性抗体以及T细胞免疫反应能够在患有NY-ESO-1表达的肿瘤病人身上常常检测的到,这些病人包括那些免疫能力低的癌症晚期的老年患者。但NY-ESO-1作为免疫分子佐剂的作用未清楚。
鉴于以上所述现有技术,本发明第一方面提供NY-ESO-1蛋白甚至在存在有常规浓度的β-巯基乙醇的loading buffer中也能形成多聚结构。
相关数据显示,从细菌及哺乳动物细胞中获得的NY-ESO-1蛋白甚至在存在有常规浓度的β-巯基乙醇的loading buffer中也能形成多聚结构(图1A,B)。
本发明第二方面提供NY-ESO-1的聚合反应由其分子间的二硫键介导。
相关实验显示,NY-ESO-1野生型蛋白清楚的显示出了存在二聚体,四聚体甚至130 kDa的多聚体的形式。ESOcs1、ESOcs2显示出的主要为单体及二聚体结构形式,而ESOcs3的结果显示出主要为单体。说明NY-ESO-1的聚合反应是由于其分子间的二硫键造成的。
本发明第三方面提供NY-ESO-1在体外与人的及鼠的未成熟的树突细胞结合与其自身的聚合结构有关。
相关数据显示,NY-ESO-1野生型蛋白清楚的显示出了存在二聚体,四聚体甚至130 kDa的多聚体的形式。ESOcs1、ESOcs2显示出的主要为单体及二聚体结构形式,而ESOcs3的结果显示出主要为单体。在癌细胞中存在的NY-ESO-1,同样采用Western blot分析其在含有常规浓度的β-巯基乙醇的loading buffer中蛋白聚合情况,发现NY-ESO-1主要表现为三聚体及四聚体,很少发现存在二聚体及单体形式的现象(突变1C)。表明NY-ESO-1在体外与人的及鼠的未成熟的树突细胞结合是与其自身的聚合结构有关的。
本发明第四方面提供TLR4影响了NY-ESO-1与骨髓中DC细胞的体外结合。
我们分别采用了从野生型小鼠及TLR4和TLR2基因敲除的小鼠骨髓中提取出来的树突细胞来进行NY-ESO-1的结合实验,从而进行对比。结果显示出了TLR4主要影响了NY-ESO-1与骨髓中DC细胞的体外结合,而TLR2则没有显著的影响(图2A)。作为NY-ESO-1与TLR4之间直接联系的证据,显示,鼠的TLR4与野生型NY-ESO-1聚合物及钙网蛋白形成明显的高分子聚合物一起沉淀下来(图2C)。与全细胞裂解液中的情况相比,这些高分子聚合物所包含的多聚体NY-ESO-1并不是以单体或者低分子量的结构域与CRT和TLR4移动结合的。随着聚合物的越来越低聚化,TLR4与NY-ESO-1的结合量也随之减少(图2D)。这些实验结果为NY-ESO-1能够与人、鼠的树突细胞结合是通过细胞表面的TLR4这一结论提供了依据。
本发明第五方面提供NY-ESO-1与人和鼠的未成熟的树突细胞结合还可通过钙网蛋白的作用。
实验采用了从野生型小鼠及TLR4和TLR2基因敲除的小鼠骨髓中提取出来的树突细胞来进行NY-ESO-1的结合实验,从而进行对比。结果显示出了TLR4主要影响了NY-ESO-1与骨髓中DC细胞的体外结合,而TLR2则没有显著的影响(图2A)。因为NY-ESO-1与人和鼠的未成熟的树突细胞结合还通过了钙网蛋白的作用。
本发明第六方面提供聚合物低聚化减少了TLR4与NY-ESO-1的结合量。
相关数据显示,随着聚合物的越来越低聚化,TLR4与NY-ESO-1的结合量也随之减少(图2D)。实验结果表明聚合物低聚化减少了TLR4与NY-ESO-1的结合量。
本发明第七方面提供免疫球抗体反应由NY-ESO-1的聚合体结构和宿主体内的TLR4参与。
相关数据显示,ESOcs1的引起的抗体的效价有某些程度上的减弱,然而ESOcs2, ESOcs3所引起的抗体其效价几乎检测不到。在TLR4基因敲除的小鼠中(图3B),由野生型质粒所编码诱导的NY-ESO-1抗体的效价也有明显的减弱,而β-gal特异性抗体的效价始终与在野生型小鼠中保持一致。表明NY-ESO-1抗体反应的TLR4依赖性。在抗体效价测定中显示,在野生型小鼠中NY-ESO-1聚合体结果突变(半胱氨酸-丝氨酸替换)与抗体效价有某些联系,但是在TLR4基因敲除的小鼠中,这种联系没有体现出来,例如,ESOcs1 在野生型小鼠能够诱导产生相当高的抗体反应,而在TLR4基因敲除的小鼠中却没有此现象,从而假设ESOcs1与野生型 NY-ESO-1与的免疫原性相同。与此相反,ESOcs2所诱导的抗体在TLR4基因敲除的小鼠中表现了高的抗体效价,而在野生型小鼠中没有检测到抗体,此现象与野生型的NY-ESO-1不同,但很像HMGB-1的免疫原性很像。ESOcs3在两种小鼠中只展现出了很小的免疫原性。说明免疫球抗体反应的是由NY-ESO-1的聚合体结构和宿主体内的TLR4来参与的。
本发明第八方面提供Art v1(wtArt)和CA9基因免疫原性的提高依赖于NY-ESO-1基因融合表达。
相关实验显示,当Art与NY-ESO-1融合表达时,能够显著提高wtArt的免疫原性,并且使免疫反应加快,注射完抗原后加快转入到产生抗体的阶段(图4A)。另外,NY-ESO-1与人的CA9基因融合表达的质粒,pcDNA-CA9-ESO,与单独编码CA9基因的质粒pcDNA-CA9,通过肌肉注射使产生免疫反应,比较两者的免疫程度。结果显示,通过注射融合质粒Balb/c所产生的免疫反应,产生了高效价的NY-ESO-1抗体,而CA9抗体的效价相对较弱,反过来,在同样的情况下只编码CA9基因的质粒并没有产生任何抗体反应。因为pcDNA-CA9, pcDNA-ESO这两种质粒在之间相同的条件下,也并没有引起任何的免疫反应,从而说明了这种CA9基因免疫原性的提高是依赖于于NY-ESO-1基因进行了融合表达。
综上所述,本发明发明人证实了NY-ESO-1有作为增强CA9免疫原性分子佐剂的作用。发现NY-ESO-1蛋白甚至在存在有常规浓度的β-巯基乙醇的loading buffer中也能形成多聚结构;NY-ESO-1的聚合反应由其分子间的二硫键介导; NY-ESO-1在体外与人的及鼠的未成熟的树突细胞结合与其自身的聚合结构有关;TLR4影响了NY-ESO-1与骨髓中DC细胞的体外结合;NY-ESO-1与人和鼠的未成熟的树突细胞结合还可通过钙网蛋白的作用;聚合物低聚化减少了TLR4与NY-ESO-1的结合量;免疫球抗体反应由NY-ESO-1的聚合体结构和宿主体内的TLR4参与;Art v1(wtArt)和CA9基因免疫原性的提高依赖于NY-ESO-1基因融合表达。表明NY-ESO-1有作为增强CA9免疫原性分子佐剂的作用。
附图说明:
图1:NY-ESO-1的DC-结合特性与它由二硫键支撑的聚合物结构有关。
A:用质粒编码NY-ESO-1转染的人293细胞(第1列),GFP质粒转染到人293细胞(第2列和第4列),融合了N-末端FLAG标签的NY-ESO-1 的质粒转染到人293细胞(FLAG-ESO,第3列)。将细胞裂解液进行SDS聚丙烯酰胺凝胶电泳,免疫印迹与mab131(第1列和第2列)和M2单克隆抗体(第3列和第4列)分别对NY-ESO-1和FLAG抗原决定基发生特异反应。值得注意的是,NY-ESO-1单体(单箭头注明)的分子量为18 kDa;而它的二聚体(双箭头注明)和四聚体分别在免疫印迹中出现36和72 kDa的条带。
B:部分纯化的野生型(第4列),esocs1 NY-ESO-1,esocs2,和esocs3蛋白(分别为1–3列),用免疫印迹(Western blot)分析。所有的蛋白质样品进行聚丙烯酰胺凝胶电泳(SDS-tuPAGE),然后使131抗体进行Western blot检测。值得注意的是,他的标签NY-ESO-1单体及其变体均出现20 kDa。
C:分别为NY-ESO-1在黑色素瘤株397,624,2984,HEK293,黑色素瘤株M202,f5.1,f5.4,和F5.6(从第1列到第8列)中的Western blot分析。单体和二聚体分别以单双箭头注明。经过PCR后,HEK293,M202,和f5.1均检测不到NY-ESO-1表达。GAPDH表示在每列蛋白总数数量相等的阳性对照。
D:相比较于野生型NY-ESO-1聚合蛋白,由半胱氨酸到丝氨酸的突变引起的NY-ESO-1低聚物变体能够降低小鼠和人类树突状细胞结合。ESO(1–74)有可通过单克隆抗体mAb131识别的B细胞表位,用于检测并作为阴性对照。用蛋白酶K处理的NY-ESO-1蛋白可作为另一个对照。等NY-ESO-1摩尔浓度的gp100蛋白也作为阴性对照(数据未显示)。实验重复使用从三个不同的捐助者获得的树突状细胞。未成熟的DC与NY-ESO-1在体外的明显结合的范围通常从8到80%不等,很大程度上是由于供体差异和实验之间的变化。因此,通常在树突状细胞结合分析中没有标准差。
E: NY-ESO-1结合人类树突状细胞的点阵图显示在(E)图中,包括散点图。MW,分子量。
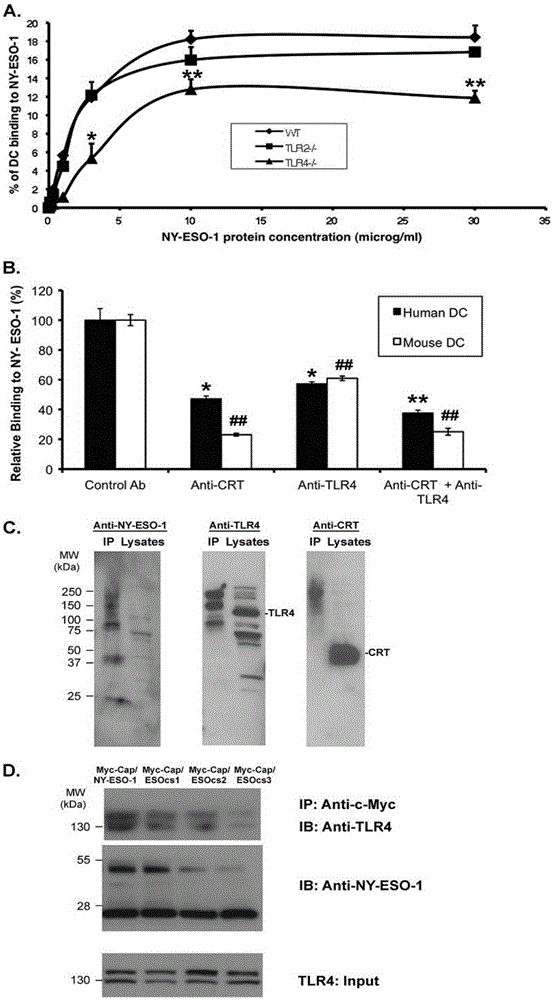
图2: 鼠及人的TLR4 参与到NY-ESO-1与未成熟树突细胞结合的过程中
A:从野生型,TLR2基因敲除和 TLR4基因敲除的骨髓中获得的未成熟的树突细胞用来研究与NY-ESO-1的结合,按照之前所介绍的浓度。用FITC标记的抗体mAb131,来能够稳定的检测到细胞。实验结果要是三个平行的带有方差的平均值,并进行t检验。当p值<0.05时,视为差异显著,当p值<0.01时,视为差异极显著。这个实验要从三只不同的老鼠身上至少重复出两次以上的同样的结果。然而,实验中存在一定的实验差异,尽管实验结果显示的是树突细胞与NY-ESO-1的相对比率。
B:抑制NY-ESO-1与人和鼠的未成熟的树突细胞结合是通过使用人的TLR4及CRT多克隆抗体来进行的,但是并没有控制对照β-actin进行处理。树突细胞结合实验标准是在3u g/ml的NY-ESO-1蛋白条件下进行的,单独的使用抗体或结合。实验至少得到一次同样得重复实验结果。
C:聚合物NY-ESO-1与鼠得TLR4和CRT形成复杂得大分子结构。Myc-Cap细胞的裂解产物与Myc标记的NY-ESO-1结合,用来做免疫共沉淀反应,用c-Myc的多克隆抗体进行检测,结果为每组的第一列。然而总的细胞裂解液在每组的第二行。利用非变性的聚丙烯酰胺凝胶点用来进行wb,左边的组用了抗-NY-ESO-1抗体,中间组是用了兔的抗TLR4抗体,右边组用了兔的抗CRT抗体。请观察NY-ESO-1,TLR4,CRT在沉淀复合物及细胞裂解液中的位置。
D:共同沉淀下来的NY-ESO-1和鼠的TLR4与多聚物NY-ESO-1联系。在每一组,1-4到包含了Myc-Cap细胞的裂解产物与编码NY-ESO-1,ESOcs1,ESOcs2,ESOcs3基因的逆转录病毒结合。沉淀产物也难怪抗c-Myc抗体来分离出NY-ESO-1及它的突变体,之后用兔的TLR4抗体(左上方),NY-ESO-1及它的突变体的mAb131抗体(左下方),TLR4在全细胞裂解液中在底部的图中
图3:NY-ESO-1和它的野生型变体以及TLR4基因敲除小鼠的免疫原性。
从C57BL / 10(A)和TLR4和C57BL/10scnj(B)免疫小鼠产生的分级转化IgG抗体,是通过基因枪引入无内毒素质粒产生的。绘制每组三只小鼠平均OD值是基于用ELISA对特异性抗体的目标的检测,即NY-ESO-1,HMGB-1,GAL蛋白。将前、后免疫血清都用ELISA稀释五倍。如果在免疫之后对特定的抗原目标OD值增加超过2倍,,表明血清为阳性。整个实验重复一次,获得了类似的结果,而关键免疫做了第三次重复。
图4:NY-ESO-1的体内免疫佐剂作用
A:基因枪注射质粒,造成免疫反应,产生艾蒿花粉主要过敏原Art 1抗体之后的两周,将收集到的血清稀释1-50倍。在第二次免疫之后饿两周,将血清稀释1-1250倍。编码wtArt, wtArt –ESO融合,及密码子替换的Art(重组)Art质粒用来造成每组3只小鼠的免疫,同样实验结果要重复两次。
B:CA9特定的抗体是由pcDNA-CA9-ESO编码产生的CA9-ESO融合蛋白所产生的,并不是是由pcDNA-CA9所编码的产生的单独的CA9基因所产生的。免疫小鼠所产生的血清与带有gfp质粒(第1列),CA9基因的质粒(第2,4列),NY-ESO-1基因的质粒(第3,5列)的293细胞裂解液进行wb。全CA9以及NY-ESO-1蛋白都在图中展示出来,在第2列的颜色稍微淡一些的阳性条带可能是CA9的转录变体。每组3只老鼠,从采用三组老鼠血清池获得结果。
C:从3只老鼠/每组获得的抗CA9抗体通过ELISA来分析重组的人CA9。一种以pcDNA-CA9为基础的质粒编码,在相对于在盐离子存在下pcDNA-CA9或是在pcDNA-CA9与pcDNA-ESO共同存在的情况下,CA9-ESO融合基因能够显著提高抗体的效价。所有的无内毒素的质粒都是经过肌肉注射到Balb/c小鼠体内。此实验结果重复1次。
图5. 一种假定的机制在癌症患者中NY-ESO-1蛋白的免疫原性
在癌症环境中,从坏死的癌细胞中产生的NY-ESO-1与钙网蛋白-Toll样受体4(CRT-TLR4 )复合物在树突细胞表面发生联系。由于CRT没有跨膜结构域,与TLR4进行复合。NY-ESO-1与CRT-TLR4复合物连接,从而引起了吞噬反应及产生危险信号。树突细胞产生了NY-ESO-1短肽以及分泌出促炎症因子和趋化因子。NY-ESO-1不会在健康人的身上引起免疫反应,因为它的表达最开始室受到睾丸免疫特权区域的限制。
具体实施方式:
以下通过特定的具体实例说明本发明的实施方式,本领域技术人员可由本说明书所揭露的内容轻易地了解本发明的其他优点与功效。本发明还可以通过另外不同的具体实施方式加以实施或应用,本说明书中的各项细节也可以基于不同观点与应用,在没有背离本发明的精神下进行各种修饰或改变。
在进一步描述本发明具体实施方式之前,应理解,本发明的保护范围不局限于下述特定的具体实施方案;还应当理解,本发明实施例中使用的术语是为了描述特定的具体实施方案,而不是为了限制本发明的保护范围;在本发明说明书和权利要求书中,除非文中另外明确指出,单数形式“一个”、“一”和“这个”包括复数形式。
当实施例给出数值范围时,应理解,除非本发明另有说明,每个数值范围的两个端点以及两个端点之间任何一个数值均可选用。除非另外定义,本发明中使用的所有技术和科学术语与本技术领域技术人员通常理解的意义相同。除实施例中使用的具体方法、设备、材料外,根据本技术领域的技术人员对现有技术的掌握及本发明的记载,还可以使用与本发明实施例中所述的方法、设备、材料相似或等同的现有技术的任何方法、设备和材料来实现本发明。
除非另外说明,本发明中所公开的实验方法、检测方法、制备方法均采用本技术领域常规的相关融合质粒构建法,质粒转染法,Western blotting,免疫共沉淀,ELISA,树突状细胞结合法,数据分析及相关领域的常规技术。
据推断,最开始对于肿瘤特异性抗原(TAA)的自身免疫应答反应可能有先天性免疫系统受体的参与。坏死肿瘤细胞会释放内源性因子或是例如热休克蛋白,高迁移率族蛋白B1(HMGB-1),尿酸这样的损伤相关模式分子。这些损伤模式分子分别通过CD91,晚期糖基化终末产物受体,TLR2/TLR4,以及白介素-1(IL-1)受体向先天免疫系统发出警报。TAA本身通常被认为是上述“危险信号”的旁观者,担任抗肿瘤免疫应答反应起始阶段的佐剂。在寻找那些宿主和肿瘤中有助于促成抗肿瘤的免疫反应的内在因子的过程中,我们将关注点放在了NY-ESO-1身上。NY-ESO-1是一种具有超强免疫原性的非突变肿瘤睾丸抗原。针对于NY-ESO-1的自发性抗体以及T细胞免疫反应能够在患有NY-ESO-1表达的肿瘤病人身上常常检测的到,这些病人包括那些免疫能力低的癌症晚期的老年患者。但NY-ESO-1作为免疫分子佐剂的作用尚未清楚。
NY-ESO-1与在不成熟的树突细胞,巨噬细胞,单核细胞表面上补体Cq1受体间存在特殊的联系,NY-ESO-1与先天性免疫系统之间的特异性相互作用。通过相关实验,我们发现NY-ESO-1蛋白甚至在存在有常规浓度的β-巯基乙醇的loading buffer中也能形成多聚结构;NY-ESO-1的聚合反应由其分子间的二硫键介导; NY-ESO-1在体外与人的及鼠的未成熟的树突细胞结合与其自身的聚合结构有关;TLR4影响了NY-ESO-1与骨髓中DC细胞的体外结合;NY-ESO-1与人和鼠的未成熟的树突细胞结合还可通过钙网蛋白的作用;聚合物低聚化减少了TLR4与NY-ESO-1的结合量;免疫球抗体反应由NY-ESO-1的聚合体结构和宿主体内的TLR4参与;Art v1(wtArt)和CA9基因免疫原性的提高依赖于NY-ESO-1基因融合表达。证实了NY-ESO-1有作为增强CA9免疫原性分子佐剂的作用。
相关融合质粒构建法
在之前的文章中介绍过的只有1-74位氨基酸残基的NY-ESO-1,被称为ESO(1-74)。为了在NY-ESO-1 cDNA中引入定点突变,利用PCR扩增来将75,76,78位的半胱氨酸换成丝氨酸。使用下列的一对磷酸化的引物:正向引物, 5-TCCTCCAGATCCGGGGCCAGGGGGCCGGAGAG(下划线表示突变)和反向引物,5-TCCATTCAGCCCTGAAGCCGCGCCGCCAT。PCR的模板为之前介绍过的表达载体PET-28-NY-ESO-1,用高保真PCR扩增系统(Invitrogen)进行扩增。PCR产物连接在载体上使其形成一个能够编码ESOcs1的新的质粒。用同样的方法引入在氨基酸152和165号位上从半胱氨酸到丝氨酸的突变,被称为ESOcs2,使用下列一对引物:正向引物,5-TTGATGTGGATCACGCAGTCCTTTCTGCCCGTG,反向引物,5-CAGGGAAAGCTGCTGGAGAGAGGAGCTGATG(下划线部分表示突变的核苷酸)。同样,能够产生以上5种75,76,78,152及165位半胱氨酸-丝氨酸替换突变的质粒(命名为ESOcs3),也用上述方法进行构建。
质粒转染法
HEK293及黑色素瘤细胞系397,624,2984从美国国家癌症研究所获得,均保存在含10%胎牛血清的DMEM培养液中。黑色素瘤细胞系M202,F5.1,F5.4及F5.6原发肿瘤细胞的早期培养物,由Ali Jazirehi (UCLA)赠与。利用Lipofectamine 2000(Invitrogen)系统转染,将质粒转入到293细胞中。Western blotting实验利用ECL发光系统(R&D公司,Minneapolis, MN)
免疫印迹法(Western blotting)
免疫印迹法(Western blotting)实验利用ECL发光系统(R&D公司,Minneapolis, MN)。
免疫共沉淀法
免疫共沉淀是小鼠的Myc-CaP细胞系进行的,即能够表达小鼠的TLR4及CRT,此细胞是Charles Sawyers(Memorial Sloan Kettering Cancer Institute, New York, NY)赠与。Myc-Cap是通过导入了逆转录病毒来编码NY-ESO-1基因,并且这个逆转录病毒融合了c-Myc标签。从10cm的培养皿中获得细胞,并用含有蛋白酶抑制剂的50 mM Tris-HCl (pH 7.4), 150 mM NaCl, 5 mM MgCl2/CaCl2来裂解细胞。细胞裂解液与抗体过夜孵育,采用特异性结合有c-Myc标签磁珠将特异性蛋白分离了出来,通过非变性凝胶或是还原型SDS-PAGE来分析全蛋白。
细胞因子酶联免疫吸附测定(ELISA)法
Verikine TM鼠干扰素α酶联免疫试剂盒从PBL InterferonSource公司(皮斯卡塔韦,新泽西州)购买。根据生产商的说明书,利用该试剂盒测定细胞因子水平。
树突状细胞结合法
人的树突状细胞是从孤立的CD14(Miltenyi Biotech)得到的,即在1000单位/毫升人的GM-CSF (Amgen,Thousand Oaks, CA)和1000单位/毫升人的IL-4(Schering-Plough, San Francisco, CA)中保存6天的阳性单核细胞。在Iscove’s medium (Invitrogen)培养液中加入10%人类雄性血清(BioCheMed, Winchester, VA)用于树突状细胞的培养。按照之前的实验计划,小鼠的树突状细胞是从C57BL/10小鼠和相应的TLR2和TLR4基因敲除小鼠的骨髓细胞产生的(The Jackson Laboratory, Bar Harbor, ME)。
按照之前的描述,所有的树突状细胞结合实验都在冰上进行,以防止由不成熟的树突状细胞导致的目标蛋白的内部活化。如果没有特别注明,我们使用的蛋白浓度分别为3, 10, 和1μg/ml,目的是维持NY-ESO-1以及它的突变体,gp100, ESO(1–74)这些蛋白所使用的摩尔浓度保持一致。大约105从鼠和人中获得的预先预冷的,不成熟的树突细胞与上述蛋白在50ul的含有5%的胎牛血清的PBS缓冲液中混合,来消除非特异性反应。在冰上孵育20min后,将样品洗两次,然后加入FITC标记的二次抗体,此单克隆抗体要么是针对NY-ESO-1的抗体要么是针对多组氨酸标签(Sigma/Aldrich)的抗体。细胞孵育20min,清洗2遍,并进行流式细胞分析。
相关数据分析方法
实施例1
NY-ESO-1蛋白甚至在存在有常规浓度的β-巯基乙醇的loading buffer中也能形成多聚结构。
实验发现从细菌及哺乳动物细胞中获得的NY-ESO-1蛋白甚至在存在有常规浓度的β-巯基乙醇的loading buffer中也能形成多聚结构(图1A,B)。
实施例2
NY-ESO-1的聚合反应由其分子间的二硫键介导。
将75,76,78,152及165位的氨基酸残基中的半胱氨酸替换成丝氨酸。三种带有氨基酸替换的NY-ESO-1分别为:在75,76,78位半胱氨酸-丝氨酸替换的ESOcs1,在152,165位替换的ESOcs2以及以上五个位点都替换突变的ESOcs3。将这些蛋白进行纯化并用Western blot来比较它们之间的齐聚反应状态(图1B)。结果显示出,野生型蛋白清楚的显示出了存在二聚体,四聚体甚至130 kDa的多聚体的形式。ESOcs1、ESOcs2显示出的主要为单体及二聚体结构形式,而ESOcs3的结果显示出主要为单体。这个实验说明了NY-ESO-1的聚合反应是由于其分子间的二硫键造成的。
实施例3
NY-ESO-1在体外与人的及鼠的未成熟的树突细胞结合与其自身的聚合结构有关。
将这些蛋白进行纯化并用Western blot来比较它们之间的齐聚反应状态(图1B)。结果显示出,野生型蛋白清楚的显示出了存在二聚体,四聚体甚至130 kDa的多聚体的形式。ESOcs1、ESOcs2显示出的主要为单体及二聚体结构形式,而ESOcs3的结果显示出主要为单体。这个实验说明了NY-ESO-1的聚合反应是由于其分子间的二硫键造成的。在癌细胞中存在的NY-ESO-1,同样采用Western blot分析其在含有常规浓度的β-巯基乙醇的loading buffer中蛋白聚合情况,发现NY-ESO-1主要表现为三聚体及四聚体,很少发现存在二聚体及单体形式的现象(突变1C)。实验表明NY-ESO-1在体外与人的及鼠的未成熟的树突细胞结合是与其自身的聚合结构有关的。
实施例4
TLR4影响了NY-ESO-1与骨髓中DC细胞的体外结合。
我们分别采用了从野生型小鼠及TLR4和TLR2基因敲除的小鼠骨髓中提取出来的树突细胞来进行NY-ESO-1的结合实验,从而进行对比。结果显示出了TLR4主要影响了NY-ESO-1与骨髓中DC细胞的体外结合,而TLR2则没有显著的影响(图2A)。因为NY-ESO-1与人和鼠的未成熟的树突细胞结合还通过了钙网蛋白的作用,它能够使TLR4基因敲除的小鼠依然保持着与树突细胞结合的作用。采用抗-TLR4抗体处理能够阻止NY-ESO-1与人,鼠的树突细胞结合,此现象也可以通过采用抗-CRT抗体处理(图2B);同时,采用β-actin的抗体作为阴性对照,此抗体对NY-ESO-1与树突细胞的结合没有影响。在与上述相同的实验条件下,用两种抗体同时对TLR4及CRT进行处理,并没有显示出明显对NY-ESO-1/DC细胞之间联系的协同抑制作用。作为NY-ESO-1与TLR4之间直接联系的证据,显示,鼠的TLR4与野生型NY-ESO-1聚合物及钙网蛋白形成明显的高分子聚合物一起沉淀下来(图2C)。与全细胞裂解液中的情况相比,这些高分子聚合物所包含的多聚体NY-ESO-1并不是以单体或者低分子量的结构域与CRT和TLR4移动结合的。这一结果与研究不同突变的NY-ESO-1(图1D)与未成熟的DC细胞结合的结果一致,随着聚合物的越来越低聚化,TLR4与NY-ESO-1的结合量也随之减少(图2D)。这些实验结果为NY-ESO-1能够与人、鼠的树突细胞结合是通过细胞表面的TLR4这一结论提供了依据。
实施例5
NY-ESO-1与人和鼠的未成熟的树突细胞结合还可通过钙网蛋白的作用。
实验采用了从野生型小鼠及TLR4和TLR2基因敲除的小鼠骨髓中提取出来的树突细胞来进行NY-ESO-1的结合实验,从而进行对比。结果显示出了TLR4主要影响了NY-ESO-1与骨髓中DC细胞的体外结合,而TLR2则没有显著的影响(图2A)。因为NY-ESO-1与人和鼠的未成熟的树突细胞结合还通过了钙网蛋白的作用。
实施例6
聚合物低聚化减少了TLR4与NY-ESO-1的结合量。
相关数据显示,鼠的TLR4与野生型NY-ESO-1聚合物及钙网蛋白形成明显的高分子聚合物一起沉淀下来(图2C)。与全细胞裂解液中的情况相比,这些高分子聚合物所包含的多聚体NY-ESO-1并不是以单体或者低分子量的结构域与CRT和TLR4移动结合的。这一结果与研究不同突变的NY-ESO-1(图1D)与未成熟的DC细胞结合的结果一致,随着聚合物的越来越低聚化,TLR4与NY-ESO-1的结合量也随之减少(图2D)。实验结果表明聚合物低聚化减少了TLR4与NY-ESO-1的结合量。
实施例7
免疫球抗体反应由NY-ESO-1的聚合体结构和宿主体内的TLR4参与。
我们采用标准的基因枪程序方法将无内毒素的编码野生型NY-ESO-1基因以及其不同突变基因的质粒(包括自然存在的变种LAGE-1b)转入到C57BL/10及C57BL/10ScNJ(TLR4-/-)中,来使动物产生免疫。质粒所编码的β-gal及膜锚定蛋白HMGB-1作为对照。每间隔两个星期做一次免疫反应,共两次。在第二次免疫反应后的两周,将小鼠处死,取其血清来分析其抗体效价。野生型小鼠的抗原特异性,抗体分类可通过质粒编码的NY-ESO-1序列及阳性对照质粒pSport-β-gal快速而有效的区分开来(图3A)。注意到了ESO(1-74)蛋白中,其包含了被NY-ESO-1及其变体共享的占主导地位的B细胞表位,用来测定抗体,并通过由低聚变体诱导的抗体来使与野生型NY-ESO-1相结合的潜在差异降到最低。ESOcs1的引起的抗体的效价有某些程度上的减弱,然而ESOcs2, ESOcs3所引起的抗体其效价几乎检测不到。在TLR4基因敲除的小鼠中(图3B),由野生型质粒所编码诱导的NY-ESO-1抗体的效价也有明显的减弱,而β-gal特异性抗体的效价始终与在野生型小鼠中保持一致。表明NY-ESO-1抗体反应的TLR4依赖性,而免疫的过程(皮肤)和原理(基因枪将质粒注入体内)不受TLR4支配。奇怪的是,一种早前报道过的与TLR4有某些联系的蛋白HMGB-1,由编码HMGB-1基因的质粒做引起的免疫反应只引起了TLR4基因缺失小鼠体内的少量的抗体(从图3B中可以看出,HMGB-1的抗体ELISA结果比NY-ESO-1, β-gal的背景小很多),但是在能够与TLR4反应的野生型小鼠体内没有检测到抗体。在抗体效价测定中显示,在野生型小鼠中NY-ESO-1聚合体结果突变(半胱氨酸-丝氨酸替换)与抗体效价有某些联系,但是在TLR4基因敲除的小鼠中,这种联系没有体现出来,例如,ESOcs1 在野生型小鼠能够诱导产生相当高的抗体反应,而在TLR4基因敲除的小鼠中却没有此现象,从而假设ESOcs1与野生型 NY-ESO-1与的免疫原性相同。与此相反,ESOcs2所诱导的抗体在TLR4基因敲除的小鼠中表现了高的抗体效价,而在野生型小鼠中没有检测到抗体,此现象与野生型的NY-ESO-1不同,但很像HMGB-1的免疫原性很像。ESOcs3在两种小鼠中只展现出了很小的免疫原性。这个实验说明了免疫球抗体反应的是由NY-ESO-1的聚合体结构和宿主体内的TLR4来参与的。
实施例8
Art v1(wtArt)和CA9基因免疫原性的提高依赖于NY-ESO-1基因融合表达。
我们利用小鼠研究了NY-ESO-1对于过敏原免疫反应的分子佐剂的作用。一个NY-ESO-1基因与艾蒿花粉主要过敏原基因融合表达的质粒通过基因枪转入到了Balb/c小鼠内。众所周知,Art v1(wtArt)在体内小鼠有着很低的免疫原性,例如,将野生型的Art基因疫苗注入到Balb/c小鼠体内,并没有展示出其免疫原性,但是可以通过改变某些密码子来变为哺乳动物能够编码的密码子及可得到克服(重组Art)。当Art与NY-ESO-1融合表达时,能够显著提高wtArt的免疫原性,并且使免疫反应加快,注射完抗原后加快转入到产生抗体的阶段(图4A)。并且,通过这个一次免疫反应产生的抗体比通过重组Art产生抗体更具有深远的意义,因为两者产生的抗体都达到了同样的水平,但后者是通过两次免疫才产生抗体。
另一个例子,NY-ESO-1与人的CA9基因融合表达的质粒,pcDNA-CA9-ESO,与单独编码CA9基因的质粒pcDNA-CA9,通过肌肉注射使产生免疫反应,比较两者的免疫程度。结果显示,通过注射融合质粒Balb/c所产生的免疫反应,产生了高效价的NY-ESO-1抗体,而CA9抗体的效价相对较弱,反过来,在同样的情况下只编码CA9基因的质粒并没有产生任何抗体反应。因为pcDNA-CA9, pcDNA-ESO这两种质粒在之间相同的条件下,也并没有引起任何的免疫反应,从而说明了这种CA9基因免疫原性的提高是依赖于于NY-ESO-1基因进行了融合表达。
综上所述,本发明发明人证实了NY-ESO-1有作为增强CA9免疫原性分子佐剂的作用。发现NY-ESO-1蛋白甚至在存在有常规浓度的β-巯基乙醇的loading buffer中也能形成多聚结构;NY-ESO-1的聚合反应由其分子间的二硫键介导; NY-ESO-1在体外与人的及鼠的未成熟的树突细胞结合与其自身的聚合结构有关;TLR4影响了NY-ESO-1与骨髓中DC细胞的体外结合;NY-ESO-1与人和鼠的未成熟的树突细胞结合还可通过钙网蛋白的作用;聚合物低聚化减少了TLR4与NY-ESO-1的结合量;免疫球抗体反应由NY-ESO-1的聚合体结构和宿主体内的TLR4参与;Art v1(wtArt)和CA9基因免疫原性的提高依赖于NY-ESO-1基因融合表达。表明NY-ESO-1有作为增强CA9免疫原性分子佐剂的作用。
综上所述,本发明有效克服了现有技术中的种种缺点而具高度产业利用价值。
上述实施例仅例示性说明本发明的原理及其功效,而非用于限制本发明。任何熟悉此技术的人士皆可在不违背本发明的精神及范畴下,对上述实施例进行修饰或改变。因此,举凡所属技术领域中具有通常知识者在未脱离本发明所揭示的精神与技术思想下所完成的一切等效修饰或改变,仍应由本发明的权利要求所涵盖。
- 还没有人留言评论。精彩留言会获得点赞!