一种产物特异性提高的环糊精葡萄糖基转移酶及制备方法与流程
本发明涉及一种产物特异性提高的环糊精葡萄糖基转移酶及制备方法,属于遗传工程和酶工程领域。
背景技术:
染料木素(又名金雀异黄素、染料木黄酮等)被认为是一种活性功能最高的大豆异黄酮类物质。在豆科植物中,染料木素经常以其葡萄糖苷衍生物即染料木苷(又名4',5,7-三羟异黄酮-7-糖苷)的形式存在。染料木素在人体及动物细胞中具有广泛的药理学功效。主要表现为:1)具有化学防癌(乳腺癌和前列腺癌等)作用。染料木素具有类雌性激素及抗激素作用,可以抑制肿瘤细胞合成过程中相关酶的活性,在肿瘤细胞形成过程中抑制肿瘤血管增生,延缓或阻止肿瘤变成癌细胞。2)可以预防心血管疾病。染料木素会激发低密度的脂蛋白受体产生正向调节作用,同时可以促进胆固醇的清除,抑制血小板的凝集,对于动脉粥样硬化等疾病具有预防和治疗作用。3)可以预防绝经后疾病。染料木素是典型的植物雌激素,所具有的雌激素活性能够缓解妇女更年期综合症及预防绝经后疾病。4)抗骨质疏松作用。染料木素的雌激素活性能够激活雌激素受体,提高成骨细胞活性;另外还可以增加骨密度,抑制骨量丢失,对骨质疏松具有较好的改善作用。
然而,染料木素具有很强疏水性,几乎不溶于水,在一般的有机溶媒中溶解度较差,而易溶于二甲基亚砜等有机溶剂。由于染料木素在水溶液中的溶解度极低,不仅限制了其作为食品添加剂、化妆品以及其他水溶性产品的应用,而且也大大降低了其作为口服药物及静脉注射药剂的药用效果,限制了其医药行业中的应用。因此,如何提高染料木素在水溶液中的溶解度,成为目前国内外关注的焦点。其中研究较多的是染料木素的糖基化衍生物。据报道二葡萄糖基染料木素和三葡萄糖基染料木素在水中的溶解度分别是染料木素的3700倍和44000倍。并且产物所连糖链越长,水溶性越好。糖基化染料木素相比与染料木素具有如下优点:1)与染料木素具有相似的生理生化功能;2)在体内水解为可被人体吸收的葡萄糖与染料木素,安全性较高;3)与染料木素相比水溶性明显的改善,拓展了其的应用范围。因此,长链糖基化染料木素衍生物具有更加广泛的应用前景。环糊精葡萄糖基转移酶(ec2.4.1.19)是目前常见的催化糖基化反应的酶。但是,由于环糊精葡萄糖基转移酶(cgtase)合成长链糖基染料木素效率较低,因此,急需找到提高环糊精葡萄糖基转移酶对长链糖基化染料木素合成效率的方法以推动与染料木素糖基衍生物相关行业的快速发展。
技术实现要素:
[技术问题]
本发明要解决的技术问题是提供一种染料木素糖基化效率高的环糊精葡萄糖基转移酶(cgtase,ec2.4.1.19)。
[技术方案]
为解决上述问题,本发明提供了一种环糊精葡萄糖基转移酶突变体,所述环糊精葡萄糖基转移酶突变体与氨基酸序列如seqidno.1所示的环糊精葡萄糖基转移酶(genbankjx412224)相比,第174位亮氨酸进行了突变。
在本发明的一种实施方式中,所述环糊精葡萄糖基转移酶来源于软化类芽孢杆菌(peanibacillusmacerans)。
在本发明的一种实施方式中,编码所述环糊精葡萄糖基转移酶的核苷酸序列如seqidno.2所示。
在本发明的一种实施方式中,所述环糊精葡萄糖基转移酶突变体与氨基酸序列如seqidno.1所示的环糊精葡萄糖基转移酶相比,第174位亮氨酸突变为了脯氨酸、酪氨酸或天冬氨酸。
本发明还提供了编码上述环糊精葡萄糖基转移酶突变体的基因。
本发明还提供了含有上述基因的重组质粒。
本发明还提供了含有上述重组质粒的基因工程菌。
本发明还提供了一种构建上述基因工程菌方法,所述方法为将编码上述环糊精葡萄糖基转移酶突变体的基因连接到质粒,得到重组质粒;将重组质粒转化大肠杆菌,得到基因工程菌。
本发明还提供了一种生产环糊精葡萄糖基转移酶的方法,所述方法为以上述基因工程菌为生产菌株,将上述基因工程菌接种至发酵培养基中进行发酵,得到发酵液;从发酵液中分离得到环糊精葡萄糖基转移酶。
本发明还提供了上述环糊精葡萄糖基转移酶突变体在染料木素的糖基化方面的应用。
本发明还提供了上述环糊精葡萄糖基转移酶突变体在食品、化工或纺织领域的应用。
[有益效果]
本发明提供了3个染料木素糖基化效率高的环糊精葡萄糖基转移酶突变体l174p、l174y和l174d,相比于野生型环糊精葡萄糖基转移酶,它们利用麦芽糊精为糖基供体生产糖基化染料木素时,长链糖基化染料木素合成产量分别提高了0.7、1.2和1.8倍,更利于糖基化染料木素的工业化生产。
附图说明
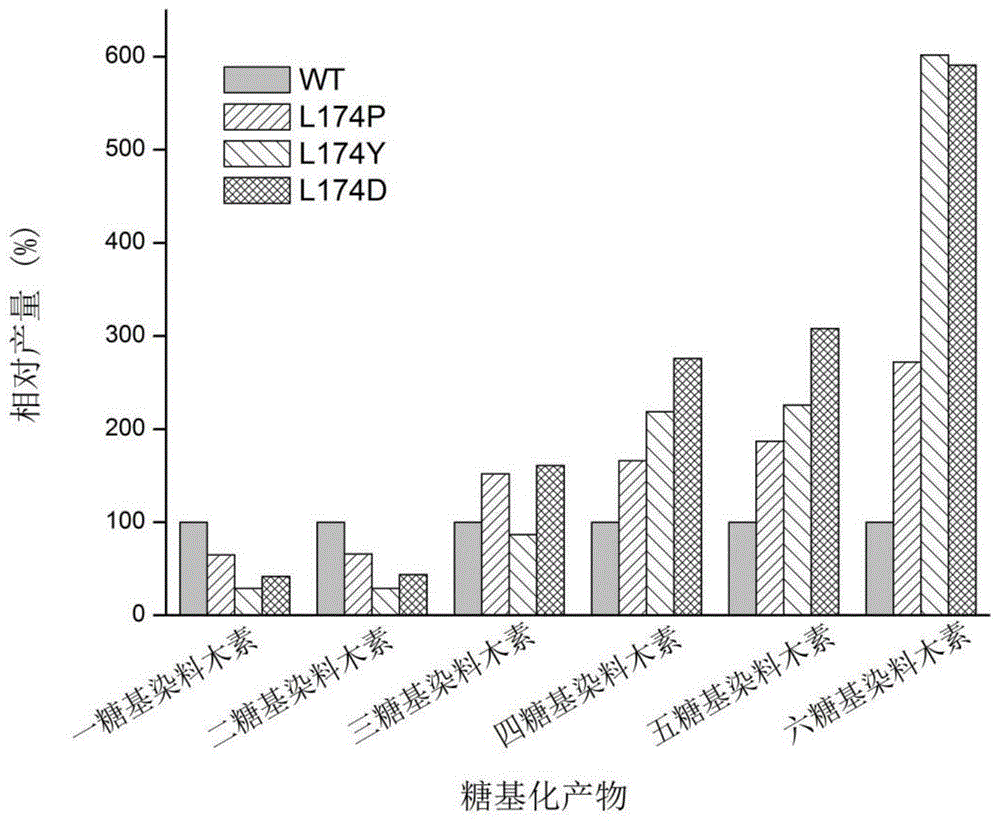
图1:野生型环糊精葡萄糖基转移酶和不同环糊精葡萄糖基转移酶突变体利用麦芽糊精为糖基供体生产不同糖基化染料木素的相对产量。
具体实施方式
下面结合具体实施例对本发明进行进一步的阐述。
下述实施例中涉及的大肠杆菌jm109以及大肠杆菌e.colibl21(de3)购自北纳生物,pet-20b(+)质粒购自novagen公司(上述菌株大肠杆菌e.colibl21(de3)可以购买得到,不需要进行用于专利程序的保藏)。
下述实施例中涉及的培养基如下:
lb液体培养基:酵母粉5.0g·l-1、胰蛋白胨10.0g·l-1、nacl10.0g·l-1、氨苄青霉素100μg·l-1。
lb固体培养基:酵母粉5.0g·l-1、胰蛋白胨10.0g·l-1、nacl10.0g·l-1、琼脂粉15g·l-1、氨苄青霉素100μg·l-1。
实施例1:染料木素糖基化效率高的环糊精葡萄糖基转移酶突变体l174p、l174y或l174d
环糊精葡萄糖基转移酶突变体l174p、l174y或l174d与氨基酸序列如seqidno.1所示的环糊精葡萄糖基转移酶(genbankjx412224)相比,第174位亮氨酸分别突变为了脯氨酸、酪氨酸或天冬氨酸。突变可通过化学全合成或pcr的方式完成。
实施例2:染料木素糖基化效率高的环糊精葡萄糖基转移酶突变体l174p、l174y或l174d的制备方法
化学合成编码氨基酸序列如seqidno.1所示的环糊精葡萄糖基转移酶的基因(基因的核苷酸序列如seqidno.2所示);将获得的基因与pet-20b(+)质粒经双酶切(ncoi和xhoi)后进行连接,转化大肠杆菌jm109,转化产物涂布于lb固体培养基,于37℃培养8h,在lb固体培养基上挑取转化子,接入lb液体培养基培养,于37℃培养10h后提取质粒,将此质粒进行序列测定,获得测序正确的重组质粒pet20b-cgt;将测序正确的重组质粒pet20b-cg转化大肠杆菌e.colibl21(de3),获得重组大肠杆菌pet20b-cgt/e.colibl21。
利用全质粒pcr技术,以获得的重组质粒pet20b-cgt为模板进行定点突变;pcr产物经经dpni处理后转化大肠杆菌jm109,转化产物涂布于lb固体培养基(含100μg/ml氨苄青霉素),于37℃培养8h,在lb固体培养基上挑取转化子,接入lb液体培养基(含100μg/ml氨苄青霉素)培养,于37℃培养10h后提取质粒,将此质粒进行序列测定,获得测序正确的含有编码环糊精葡萄糖基转移酶突变体l174p、l174y或l174d的基因的重组质粒;将测序正确的重组质粒转化大肠杆菌e.colibl21(de3),即获得含有编码突变体环糊精葡萄糖基转移酶突变体l174p、l174y或l174d的基因的重组大肠杆菌pet20b-cgt/e.colibl21-1、pet20b-cgt/e.colibl21-2或pet20b-cgt/e.colibl21-3;
其中,引入a174p突变的引物如下:
正向引物:5'-gacaccgctggcccgttccat-3',下划线为突变碱基(seqidno.3);
反向引物:5'-gttgtgatggaacgggccagc-3',下划线为突变碱基(seqidno.4);
引入a174y突变的引物如下:
正向引物:5'-gacaccgctggctacttccat-3',下划线为突变碱基(seqidno.5);
反向引物:5'-gttgtgatggaagtagccagc-3',下划线为突变碱基(seqidno.6);
引入a174d突变的引物如下:
正向引物:5'-gacaccgctggcgacttccat-3',下划线为突变碱基(seqidno.7);
反向引物:5'-gttgtgatggaagtcgccagc-3',下划线为突变碱基(seqidno.8);
pcr反应体系均为:5×primestarbuffer(mg2+plus)5μl,2.5mmdntps4μl,10μm正向引物1μl,10μm反向引物1μl,模板dna1μl,2.5u/μlprimestartaqhs0.5μl,加入双蒸水至50μl;
pcr产物扩增条件均为:98℃预变性3min;随后进行98℃10s,57℃15s,72℃6min,30个循环;最后72℃保温10min。
将获得的重组大肠杆菌pet20b-cgt/e.colibl21、重组大肠杆菌pet20b-cgt/e.colibl21-1、pet20b-cgt/e.colibl21-2或pet20b-cgt/e.colibl21-3分别涂布于lb固体培养基(含100μg/ml氨苄青霉素),于37℃培养8~10h,获得单菌落;挑取单菌落接入lb液体培养基(含100μg/ml氨苄青霉素),于37℃培养12~14h,获得种子液;将种子液按照4%(v/v)的接种量接入lb液体培养基(含100μg/ml氨苄青霉素),于30℃、120rpm培养至od600=0.6后,在发酵液中加入终浓度为0.01mm的iptg,于25℃、120rpm继续诱导培养90h,得到发酵液;将发酵液于4℃、1000rpm离心20min后,收集发酵上清液;在发酵上清液中加入70%固体硫酸铵盐析过夜,4℃、10000rpm离心20min,取沉淀物用适量含20mm磷酸钠、0.5m氯化钠、20mm咪唑、ph7.4的缓冲液a溶解,并在缓冲液a中透析过夜后,通过0.22μm膜过滤后制成上样样品;ni亲和柱用缓冲液a平衡后,将上样样品吸入ni柱,使之完全吸附后,分别用缓冲液a、含20~480mm咪唑的缓冲液a、含480mm咪唑的缓冲液a的洗脱,流速1ml/min,检测波长为280nm,分部收集含环糊精葡萄糖基转移酶酶活的洗脱液;活力组分在50mm磷酸钠缓冲液(ph=6)中透析过夜后,得到环糊精葡萄糖基转移酶突变体l174p、l174y或l174d的纯酶,并冻干备用。
实施例3:环糊精葡萄糖基转移酶突变体l174p、l174y或l174d的酶活分析以及染料木素糖基化效率分析
1、酶活分析
1.1、α-环化活力
以野生型环糊精葡萄糖基转移酶为对照,取实施例2获得的环糊精葡萄糖基转移酶突变体l174p、l174y或l174d的纯酶0.1ml,加入装有0.9ml预先用50mm磷酸缓冲液(ph6.5)配制的浓度为30g·l-1的可溶性淀粉溶液中,在40℃下反应10min后,加入1.0ml1.0m的盐酸停止反应,再加入1.0ml用50mm磷酸缓冲液配制的0.1mm甲基橙,在16℃下保温20min,在505nm下测定吸光度,得到实施例2获得的环糊精葡萄糖基转移酶突变体l174p、l174y或l174d的α-环化活力,检测结果见表1。
环糊精葡萄糖基转移酶α-环化活力的定义:在该条件下每分钟生成1μmolα-环糊精所需酶量为一个酶活力单位。
1.2、水解活力
以野生型环糊精葡萄糖基转移酶为对照,取实施例2获得的环糊精葡萄糖基转移酶突变体l174p、l174y或l174d的纯酶0.1ml,加入装有0.9ml预先用50mm磷酸缓冲液(ph6.5)配制的浓度为10g·l-1的可溶性淀粉溶液中,在50℃下反应10min后,用dns法测定反应液中还原糖浓度,得到实施例2获得的环糊精葡萄糖基转移酶突变体l174p、l174y或l174d的水解活力,检测结果见表1。
环糊精葡萄糖基转移酶水解活力的定义:在该条件下每分钟生成1μmol还原糖所需酶量为一个酶活力单位。
1.3、歧化活力
将含有6mm供体底物4-硝基苯基-α-d-麦芽庚糖-4-6-o-亚乙基(eps)和10mm受体底物麦芽糖的10mm柠檬酸缓冲液(ph6.0)在50℃下保温10min后,以野生型环糊精葡萄糖基转移酶为对照,将0.1ml实施例2获得的环糊精葡萄糖基转移酶突变体l174p、l174y或l174d的纯酶添加至柠檬酸缓冲液(ph6.0)中进行反应,每0.5min取100μl反应液,在反应液中加入20μl1.2m的盐酸(4℃)后,将反应液在60℃保温10min使环糊精葡萄糖基转移酶突变体失活,接着,在反应液中加入20μl1.2m的naoh中和,得到检测样品,将检测样品添加至磷酸缓冲液(ph7.0)中,并在磷酸缓冲液(ph7.0)加入60μl(1u)α-糖苷酶,37℃反应60min后,在反应液中加入1ml1m的碳酸钠使反应液ph升至8以上,在401nm下测定吸光度(ε401=18.4mm-1),得到实施例2获得的环糊精葡萄糖基转移酶突变体l174p、l174y或l174d的歧化活力,检测结果见表1。
环糊精葡萄糖基转移酶歧化活力的定义:在该条件下每分钟转化1μmol4-硝基苯基-α-d-麦芽庚糖-4-6-o-亚乙基(eps)所需酶量为一个酶活力单位。
由表1可知,环糊精葡萄糖基转移酶突变体l174p、l174y和l174d的α-环化活力较野生型环糊精葡萄糖基转移酶有所下降,分别降低了18.2%、45.5%和38%;
环糊精葡萄糖基转移酶突变体l174p、l174y和l174d的水解活力较野生型环糊精葡萄糖基转移酶变化不大,略有上升;
环糊精葡萄糖基转移酶突变体l174p、l174y和l174d的歧化活力较野生型环糊精葡萄糖基转移酶均有提高,分别提高了37%,89%和42%。
表1环糊精葡萄糖基转移酶突变体l174p、l174y和l174d的α-环化活力、水解活力和歧化活力
2、染料木素糖基化效率分析
将染料木素(购自sigma公司)溶解于二甲基亚砜(dmso)中配制成终浓度为7.5g/l的染料木素溶液;将麦芽糊精(购自上海生工生物工程有限公司)溶解于pbs缓冲液(50mm,ph6.5)中配制成终浓度为40g/l的麦芽糊精溶液;以野生型环糊精葡萄糖基转移酶为对照,取实施例2获得的环糊精葡萄糖基转移酶突变体l174p、l174y或l174d的纯酶0.1ml,溶解于pbs缓冲液(50mm,ph6.5)中配制成终浓度为15u/l的cgtase酶液;分别取300μl染料木素溶液,500μl麦芽糊精溶液和200μlcgtase酶液混合于2ml的带盖小管内,放于40℃、120rpm摇床缓慢振荡24h,得到反应液。
通过hplc检测反应液中不同糖基化染料木素(此处糖基化染料木素为一糖基化染料木素、二糖基化染料木素、三糖基化染料木素四糖基化染料木素、五糖基化染料木素和六糖基化染料木素)的摩尔含量,并且,以野生型环糊精葡萄糖基转移酶反应获得的反应液中不同糖基化染料木素产物的摩尔含量分别作为100%,计算不同环糊精葡萄糖基转移酶突变体反应得到的反应液中不同糖基化染料木素产物的相对产量(%),检测结果见图1和表2;
其中,通过hplc检测反应液中不同糖基化染料木素的摩尔含量的方法为:反应液通过0.22μm滤膜过滤,使用amethystc18-h柱(4.6×250mm,sepax,america)检测(具体检测条件见表3);不同长链糖基化染料木素的绝对产量=不同长链糖基化染料木素的摩尔含量×不同长链糖基化染料木素的分子量,不同短链糖基化染料木素的绝对产量=不同短链糖基化染料木素的摩尔含量×不同短链糖基化染料木素的分子量。
由图1可表2可知,环糊精葡萄糖基转移酶突变体l174p、l174y和l174d生产短链糖基化染料木素的效率较野生型环糊精葡萄糖基转移酶有所下降,生产长链糖基化染料木素的效率较野生型环糊精葡萄糖基转移酶有所提高,其中,环糊精葡萄糖基转移酶突变体l174p、l174y和l174d生产四糖基染料木素的相对产量较野生型环糊精葡萄糖基转移酶分别提高了0.7、1.2和1.8倍,生产五糖基染料木素的相对产量较野生型环糊精葡萄糖基转移酶分别提高了0.9、1.3和2.1倍,生产六糖基染料木素的相对产量较野生型环糊精葡萄糖基转移酶分别提高了1.7、5.0和4.9倍,而其余突变体无此性质。
表2不同环糊精葡萄糖基转移酶突变体反应得到的反应液中六糖基染料木素的相对产量
表3hplc检测反应液中短链糖基化染料木素以及长链糖基化染料木素的含量的条件
虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动与修饰,因此本发明的保护范围应该以权利要求书所界定的为准。
序列表
<110>江南大学
<120>一种产物特异性提高的环糊精葡萄糖基转移酶及制备方法
<160>8
<170>patentinversion3.3
<210>1
<211>687
<212>prt
<213>人工序列
<400>1
serproaspthrservalaspasnlysvalasnpheserthraspval
151015
iletyrglnilevalthraspargphealaaspglyaspargthrasn
202530
asnproalaglyaspalapheserglyaspargserasnleulysleu
354045
tyrpheglyglyasptrpglnglyileileasplysileasnaspgly
505560
tyrleuthrglymetglyvalthralaleutrpileserglnproval
65707580
gluasnilethrservalilelystyrserglyvalasnasnthrser
859095
tyrhisglytyrtrpalaargaspphelysglnthrasnaspalaphe
100105110
glyaspphealaasppheglnasnleuileaspthralahisalahis
115120125
asnilelysvalvalileaspphealaproasnhisthrserproala
130135140
aspargaspasnproglyphealagluasnglyalaleutyraspasn
145150155160
glyserleuleuglyalatyrserasnaspthralaglyleuphehis
165170175
hisasnglyglythrasppheserthrilegluaspglyiletyrlys
180185190
asnleutyraspleualaaspileasnhisasnasnasnalametasp
195200205
alatyrphelysseralaileaspleutrpleuglymetglyvalasp
210215220
glyileargpheaspalavallyshismetpropheglytrpglnlys
225230235240
serphevalserseriletyrglyglyasphisprovalphethrphe
245250255
glyglutrptyrleuglyalaaspglnthraspglyaspasnilelys
260265270
phealaasngluserglymetasnleuleuasppheglutyralagln
275280285
gluvalarggluvalpheargasplysthrgluthrmetlysaspleu
290295300
tyrgluvalleualaserthrgluserglntyrasptyrileasnasn
305310315320
metvalthrpheileaspasnhisaspmetaspargpheglnvalala
325330335
glyserglythrargalathrgluglnalaleualaleuthrleuthr
340345350
serargglyvalproalailetyrtyrglythrgluglntyrmetthr
355360365
glyaspglyaspproasnasnargalametmetthrserpheasnthr
370375380
glythrthralatyrlysvalileglnalaleualaproleuarglys
385390395400
serasnproalailealatyrglythrthrthrgluargtrpvalasn
405410415
asnaspvalleuileilegluarglyspheglyserseralaalaleu
420425430
valalaileasnargasnserseralaalatyrproileserglyleu
435440445
leuserserleuproalaglythrtyrseraspvalleuasnglyleu
450455460
leuasnglyasnserilethrvalglyserglyglyalavalthrasn
465470475480
phethrleualaalaglyglythralavaltrpglntyrthralapro
485490495
gluthrserproalaileglyasnvalglyprothrmetglyglnpro
500505510
glyasnilevalthrileaspglyargglypheglyglythralagly
515520525
thrvaltyrpheglythrthralavalthrglyserglyilevalser
530535540
trpgluaspthrglnilelysalavalileprolysvalalaalagly
545550555560
lysthrglyvalservallysthrserserglythralaserasnthr
565570575
phelysserpheasnvalleuthrglyaspglnvalthrvalargphe
580585590
leuvalasnglnalaasnthrasntyrglythrasnvaltyrleuval
595600605
glyasnalaalagluleuglysertrpaspproasnlysalailegly
610615620
promettyrasnglnvalilealalystyrprosertrptyrtyrasp
625630635640
valservalproalaglythrlysleuaspphelyspheilelyslys
645650655
glyglyglythrvalthrtrpgluglyglyglyasnhisthrtyrthr
660665670
thrproalaserglyvalglythrvalthrvalasptrpglnasn
675680685
<210>2
<211>2064
<212>dna
<213>人工序列
<400>2
tcaccggacacctcagtggacaataaagttaacttcagcaccgatgttatctaccagatc60
gtcacggaccgttttgcggatggtgaccgcaccaacaatccggcaggcgatgctttcagc120
ggtgaccgttctaatctgaaactgtattttggcggtgattggcagggcattatcgataaa180
attaacgacggttacctgaccggcatgggtgtgacggcgctgtggatcagccaaccggtg240
gaaaacatcacctcagttatcaaatactcgggcgtcaacaatacgtcttatcatggttac300
tgggcccgtgattttaaacagaccaacgacgcgtttggcgatttcgccgactttcaaaat360
ctgattgataccgcacatgctcacaacattaaagtggttatcgatttcgccccgaaccac420
acctctccggcagatcgcgacaatccgggctttgcagaaaatggtgctctgtatgataac480
ggctcactgctgggtgcatactcgaatgacaccgctggcctgttccatcacaacggcggt540
acggattttagtaccattgaagacggtatctataaaaatctgtacgatctggctgacatc600
aaccataacaataacgcgatggatgcctatttcaaatcagcaattgacctgtggctgggc660
atgggtgttgatggcatccgctttgacgcggtcaaacacatgccgttcggttggcagaaa720
tcgtttgtgagcagcatttatggcggtgatcacccggtttttaccttcggcgaatggtat780
ctgggtgctgatcagacggatggcgacaatatcaaatttgcgaacgaatctggtatgaat840
ctgctggattttgaatatgcacaagaagtccgtgaagtgtttcgcgataaaacggaaacc900
atgaaagacctgtacgaagtgctggcctcaaccgaatcgcagtatgattacattaataac960
atggtgaccttcatcgacaatcacgatatggaccgttttcaggttgcgggctcaggtacg1020
cgcgccaccgaacaagcgctggcactgacgctgacctcgcgtggcgttccggcgatttat1080
tacggcaccgaacagtatatgacgggcgatggtgacccgaataaccgcgccatgatgacg1140
agtttcaataccggcaccacggcatataaagtgattcaagcactggctccgctgcgtaaa1200
tccaacccggcaatcgcctacggcaccaccaccgaacgttgggtgaataacgatgttctg1260
attatcgaacgcaaatttggtagttccgcggccctggtcgccattaatcgcaactcatcg1320
gcagcttatccgatcagtggtctgctgagcagcctgccagcgggcacctactccgatgtg1380
ctgaatggcctgctgaatggtaacagcattaccgtgggctctggcggtgcggttacgaac1440
tttaccctggcagcgggcggcaccgcagtttggcagtatacggctccggaaaccagcccg1500
gcgatcggtaatgtcggtccgacgatgggccaaccgggtaacattgtgacgatcgatggt1560
cgtggtttcggcggtacggctggcaccgtgtactttggtacgaccgcggtcaccggcagt1620
ggtattgtgtcctgggaagatacgcagattaaagcggtcatcccgaaagtggcagctggc1680
aaaaccggtgtcagcgtgaaaacgagttccggcaccgccagtaatacgttcaaatccttt1740
aacgttctgaccggtgatcaggttacggtccgctttctggtcaaccaagcgaataccaac1800
tatggcacgaatgtttacctggtcggcaacgcggccgaactgggttcctgggacccgaat1860
aaagccattggtccgatgtataaccaggttatcgcaaaatacccgagctggtattacgat1920
gtgagcgttccggcgggcaccaaactggacttcaaattcattaaaaaaggcggtggcacg1980
gtgacctgggaaggtggcggtaaccatacctacacgaccccggcgagcggcgttggcacg2040
gtgacggtggattggcaaaattaa2064
<210>3
<211>21
<212>dna
<213>人工序列
<400>3
gacaccgctggcccgttccat21
<210>4
<211>21
<212>dna
<213>人工序列
<400>4
gttgtgatggaacgggccagc21
<210>5
<211>21
<212>dna
<213>人工序列
<400>5
gacaccgctggctacttccat21
<210>6
<211>21
<212>dna
<213>人工序列
<400>6
gttgtgatggaagtagccagc21
<210>7
<211>21
<212>dna
<213>人工序列
<400>7
gacaccgctggcgacttccat21
<210>8
<211>21
<212>dna
<213>人工序列
<400>8
gttgtgatggaagtcgccagc21
- 还没有人留言评论。精彩留言会获得点赞!