谷氨酸-半胱氨酸连接酶变体和使用其生产谷胱甘肽的方法与流程
1.本公开内容涉及新的谷氨酸-半胱氨酸连接酶变体和使用其生产谷胱甘肽的方法。
背景技术:
2.作为大多数细胞中普遍存在的有机硫化合物,谷胱甘肽(gsh)是由三种氨基酸组成的三肽:甘氨酸、谷氨酸和半胱氨酸。
3.谷胱甘肽以还原型的谷胱甘肽(gsh)和氧化型的谷胱甘肽(gssg)存在于活体中。还原型的谷胱甘肽(gsh)在正常情况下占比较高,主要分布在人体内的肝脏和皮肤细胞中,并且具有以下重要作用:分解和清除活性氧的抗氧化功能、去除例如毒性物质的异生化合物的解毒功能和抑制黑色素产生的美白功能。
4.由于随着衰老过程的进行,谷胱甘肽的生产逐渐减少,而在抗氧化和解毒功能中起重要作用的谷胱甘肽的生产减少促进了作为衰老主要原因的活性氧的积累,因此需要从外部供应谷胱甘肽(sipes ig等人,“the role of glutathione in the toxicity of xenobiotic compounds:metabolic activation of 1,2-dibromoethane by glutathione”,adv exp med biol.1986;197:457-67).
5.具有上述多种功能的谷胱甘肽作为例如药物、保健功能食品和化妆品的多种领域中的物质而受到关注,并且还用于制造调味成分以及食品和饲料添加剂。已知谷胱甘肽对丰富原料的味道和保持浓郁风味有很大的作用,并且可以单独或与其他物质组合用作厚味(kokumi)增味剂。通常,已知厚味物质比例如已知的核酸和味精(msg)的鲜味(umami)物质具有更丰富的风味,并且在成熟期间通过蛋白质分解产生。
6.尽管谷胱甘肽在多个领域的应用需求不断增长,但由于谷胱甘肽的工业生产成本高(因为其合成酶的方法由于生产成本高而尚未商业化,并且培养微生物和从中提取谷胱甘肽的方法的产率很低),因此其市场没有被激活。
技术实现要素:
7.技术问题
8.作为解决上述问题的大量努力的结果,本发明人开发了新型谷氨酸-半胱氨酸连接酶变体,并发现新型谷氨酸-半胱氨酸连接酶引入的菌株的谷胱甘肽生产能力显著提高,从而完成了本公开内容。
9.技术方案
10.本公开内容提供了谷氨酸-半胱氨酸连接酶变体,其中对应于seq id no:1的氨基酸序列的n末端起第86位的氨基酸被不同的氨基酸取代。
11.本公开内容提供了编码该变体的多核苷酸和包含其的载体。
12.本公开内容通过包括以下至少一种来提供生产谷胱甘肽的微生物:变体;编码变体的多核苷酸;和包含多核苷酸的载体。
13.本公开内容提供了生产谷胱甘肽的方法,其包括培养微生物。
14.有利效果
15.本公开内容的新型谷氨酸-半胱氨酸连接酶变体显著增加了谷胱甘肽生产,因此可用于以高产率生产谷胱甘肽。由于以高产率生产谷胱甘肽的酵母,其干燥产物、提取物、培养物和裂解物以及生产的谷胱甘肽具有抗氧化、解毒和免疫力增强作用,它们可以有效地用于化妆品组合物、食品组合物、饲料组合物和药物组合物及其制备。
具体实施方式
16.将详细描述本公开内容。同时,在本公开内容中公开的每个描述和实施方式可以在本文中应用于不同的描述和实施方式。换言之,本公开内容所公开的多种组分的所有组合均包含在本公开内容的范围内。此外,本公开内容的范围不应受以下提供的描述限制。
17.本领域技术人员仅使用常规实验将认识到或能够确定本公开内容的具体实施方式的许多等效物。这样的等效物旨在包含在所附权利要求的范围内。
18.本公开内容的一个方面提供了谷氨酸-半胱氨酸连接酶变体,其中对应于seq id no:1的氨基酸序列的n末端起第86位的氨基酸被不同的氨基酸取代。
19.变体可以是谷氨酸-半胱氨酸连接酶变体,包括在seq id no:1的氨基酸序列中的至少一个氨基酸取代,其中该取代包括对应于seq id no:1的n末端起第86位的氨基酸被不同的氨基酸取代。
20.具体地,变体可以是蛋白质变体,其中seq id no:1的氨基酸序列的n末端起第86位氨基酸被不同的氨基酸取代。
21.本公开内容的“谷氨酸-半胱氨酸连接酶(gcl)”是也称为“谷氨酸-半胱氨酸联接酶”或“γ-谷氨酰半胱氨酸合成酶(gcs)”的酶。已知谷氨酸-半胱氨酸连接酶催化以下反应:
[0022][0023]
此外,由谷氨酸-半胱氨酸连接酶催化的反应被称为谷胱甘肽合成的第一步。
[0024]
作为酵母衍生序列的谷氨酸-半胱氨酸连接酶可以是包括seq id no:1的氨基酸序列的蛋白质,但不限于此。
[0025]
在本公开内容中,seq id no:1的氨基酸序列为gshl基因编码的氨基酸序列,并且也可称为“gsh1蛋白”或“谷氨酸-半胱氨酸连接酶”。构成本公开内容的谷氨酸-半胱氨酸连接酶的氨基酸序列可以从ncbi genbank的已知数据库中获得。例如,氨基酸序列可以来源于酿酒酵母,但不限于此,并且可以没有限制地包括与该氨基酸序列具有相同活性的任何序列。
[0026]
此外,虽然谷氨酸-半胱氨酸连接酶在本公开内容中被定义为包括seq id no:1的氨基酸序列的蛋白质,但不排除可能天然或通过在seq id no:1的氨基酸序列的上游或下游添加无意义的序列而发生的突变、或其天然发生的突变或沉默突变,对普通技术人员显而易见的是,任何具有与包括seq id no:1的氨基酸序列的蛋白质相同或等效活性的蛋白质都属于本公开内容的谷氨酸-半胱氨酸连接酶。
[0027]
例如,本公开内容的谷氨酸-半胱氨酸连接酶可以是包括seq id no:1的氨基酸序列或具有与其至少80%、85%、90%、95%、96%、97%、98%或99%的同源性或同一性的氨
基酸序列的蛋白质。此外,显然,任何具有包括一个或几个氨基酸的缺失、修饰、取代或添加的氨基酸序列的蛋白质都在本公开内容的范围内,只要该氨基酸序列保留上述同源性或同一性以及与蛋白质的作用等效的作用。
[0028]
换言之,尽管在本公开内容中使用了表述“具有预定的seq id no:的氨基酸序列的蛋白质或多肽”和“包括预定的seq id no:的氨基酸序列的蛋白质或多肽”,但是显然,任何具有包括一个或几个氨基酸的缺失、修饰、取代或添加的氨基酸序列的蛋白质也可以用于本公开内容,只要该蛋白质具有与由预定氨基酸序列组成的多肽同一或等效的活性。例如,显然,“包括seq id no:1的氨基酸序列的多肽”属于“由seq id no:1的氨基酸序列组成的多肽”,只要前者具有与后者相同或等效的活性。
[0029]
如本文所用,术语“变体”是指通过与所述序列不同的至少一个氨基酸的保守取代和/或修饰而获得的蛋白质,同时保留了蛋白质的功能或性质,并且可以是谷氨酸-半胱氨酸连接酶变体,其中鉴于本公开内容的目的,对应于seq id no:1的氨基酸序列的n末端起第86位的氨基酸被半胱氨酸以外的氨基酸残基取代。变体不同于通过几个氨基酸的取代、缺失或添加而鉴定的序列。此类变体通常可通过修饰蛋白质的上述氨基酸序列之一并评估修饰蛋白质的性质来鉴定。即,相对于天然蛋白质,变体的能力可以增强。此外,一些变体可能包括至少一个部分例如n末端前导序列或跨膜结构域已被从中去除的变体。其他变体可包括其中一部分已从成熟蛋白质的n和/或c末端去除的变体。术语“变体”也可以与其他术语互换使用,例如修饰、修饰的蛋白质、修饰的多肽、突变体、突变蛋白和趋异(divergent),并且也可以不受限制地使用用于表示变化的任何术语。鉴于本公开内容的目的,与野生型或未修饰的蛋白质相比,变体可以具有增强的活性,但不限于此。
[0030]
如本文所用,术语“保守取代”是指用具有相似结构和/或化学性质的不同氨基酸取代一种氨基酸。该变体可以具有至少一个保守取代,同时保留至少一种生物活性。这种氨基酸取代通常可以基于残基的极性、电荷、溶解性、疏水性、亲水性和/或两亲性质的相似性而发生。
[0031]
变体还可以包括对多肽的性质和二级结构具有最小影响的氨基酸的缺失或添加。例如,多肽可以在蛋白质的n末端缀合到信号(或前导)序列,其共翻译或翻译后指导蛋白质的转移。多肽也可以与另一个序列或接头缀合以鉴定、纯化或合成多肽。
[0032]
在本公开内容中,“用不同的氨基酸取代”没有具体限制,只要取代后的氨基酸与取代前的氨基酸不同。即,用不同的氨基酸取代seq id no:1的氨基酸序列的n末端起第86个氨基酸的半胱氨酸也可以表述为“用半胱氨酸以外的氨基酸取代第86位的氨基酸”。同时,在本公开内容中,显然,除非给出“用不同的氨基酸取代”的表述,否则“用预定氨基酸取代”的表述是指取代后的氨基酸与取代前的氨基酸不同。
[0033]
本公开内容的“谷氨酸-半胱氨酸连接酶变体”也可以称为“具有谷氨酸-半胱氨酸连接酶活性的(变体)多肽”或“gsh1变体”,与修饰之前的蛋白质、野生型多肽或未修饰多肽相比,其可以增加谷胱甘肽生产,但不限于此。
[0034]
在变体中,seq id no:1的氨基酸序列的至少一个氨基酸可以被不同的氨基酸取代。具体地,变体可以包括对应于seq id no:1的氨基酸序列的第86位的氨基酸被半胱氨酸以外的氨基酸取代。不同的氨基酸可以选自甘氨酸、丙氨酸、缬氨酸、亮氨酸、异亮氨酸、甲硫氨酸、苯丙氨酸、色氨酸、脯氨酸、丝氨酸、苏氨酸、酪氨酸、天冬酰胺、谷氨酸、谷氨酰胺、
天冬氨酸、赖氨酸、精氨酸和组氨酸。
[0035]
在本公开内容中,显然,“其中seq id no:1的氨基酸序列的n末端起第86位氨基酸被不同的氨基酸取代的变体”包括其中对应于seq id no:1的氨基酸序列的第86位的氨基酸被不同的氨基酸取代的变体,尽管该氨基酸由于seq id no:1的氨基酸序列的n或c末端或中间的氨基酸的缺失/添加/插入等而位于第86位以外的位置。
[0036]
此外,虽然作为本公开内容的谷氨酸-半胱氨酸连接酶变体的实例公开了其中seq id no:1的氨基酸序列的n末端起第86位的氨基酸被不同氨基酸取代的变体,但是本公开内容的谷氨酸-半胱氨酸连接酶变体不限于seq id no:1的氨基酸序列的变体,显然,在任何具有谷氨酸-半胱氨酸连接酶活性的氨基酸序列中的其中“对应于seq id no:1的氨基酸序列的第86位的氨基酸”被不同氨基酸取代的变体也在本公开内容的谷氨酸-半胱氨酸连接酶变体的范围内。
[0037]
在任何氨基酸序列中,“对应于seq id no:1的氨基酸序列的第86位的氨基酸”可以通过本领域公知的多种序列比对方法进行鉴定。
[0038]
本公开内容的其中对应于seq id no:1的氨基酸序列的n末端起第86位的氨基酸被不同氨基酸取代的谷氨酸-半胱氨酸连接酶变体可以是如下的蛋白质,该蛋白质包括seq id no:1的氨基酸序列或具有与其至少80%、85%、90%、95%、96%、97%、98%或99%的同源性或同一性的氨基酸序列,其中对应于seq id no:1的第86位的氨基酸被不同的氨基酸取代。
[0039]
根据本公开内容的其中seq id no:1的氨基酸序列的n末端起第86位氨基酸被半胱氨酸以外的氨基酸取代的谷氨酸-半胱氨酸连接酶变体可以包括seq id no:3至21的氨基酸序列之一。具体地,变体可以基本上由seq id no:3至21的氨基酸序列之一组成,更具体地,它可以由seq id no:3至21的氨基酸序列之一组成,但不限于此。
[0040]
此外,变体可以包括seq id no:3至21的氨基酸序列之一或包括固定的第86位氨基酸(即,在变体的氨基酸序列中,对应seq id no:3至21的第86位的氨基酸与seq id no:3至21的第86位的氨基酸相同),并且具有与其至少80%的同源性或同一性的氨基酸序列,但不限于此。
[0041]
具体地,本公开内容的变体可以包括具有seq id no:3至21的氨基酸序列之一的多肽和具有与seq id no:3至21的氨基酸序列之一至少80%、90%、95%、96%、97%、98%或99%同源性或同一性的多肽。此外,显然具有包括在第86位以外的位置上的一个或几个氨基酸的缺失、修饰、取代或添加的氨基酸序列的任何蛋白质在本公开内容的范围内,只要该蛋白质保持上述的同源性或同一性以及与变体等效的作用。
[0042]
如本文所用,术语“同源性”或“同一性”是指两个给定氨基酸序列或碱基序列之间的相关程度,并且可以表示为百分比。术语同源性和同一性通常可以互换使用。
[0043]
可以通过标准比对算法确定保守多核苷酸或多肽的序列同源性或同一性,并且可以与其一起使用由程序建立的默认间隙罚分。基本上,在中等或高度严格的条件下,同源或相同的序列可以以整个序列或整个长度的至少约50%、60%、70%、80%或90%彼此杂交。在杂交的多核苷酸中,也可以考虑包括简并密码子而不是密码子的多核苷酸。
[0044]
两个给定多肽或多核苷酸之间的序列同源性、相似性或同一性可以使用任何已知的计算机算法来确定,例如通过使用默认参数的“fasta”程序,如例如pearson等人,(1988)
proc.natl.acad.sci.usa 85:2444所介绍的。可选地,在欧洲分子生物学开放软件套件(the european molecular biology open software suite)(emboss)包(rice et al.,2000,trends genet.16:276-277)(版本5.0.0或更高版本)中进行的needleman-wunsch算法(1970,j.mol.biol.48:443-453)可用于进行确定(包括gcg程序包(devereux,j.et al.,nucleic acids research 12:387(1984))、blastp、blastn、fasta (atschul,s.f.et al.,j molec biol 215:403(1990);guide to huge computers,martin j.bishop,ed.,academic press,san diego,1994;and carillo et al.(1988)siam j applied math 48:1073)。例如,可以使用来自国家生物技术信息中心数据库或clustalw的blast来确定同源性、相似性或同一性。
[0045]
多核苷酸或多肽之间的同源性、相似性或同一性可以通过使用gap计算机程序比较序列信息来确定,例如由needleman等人(1970),j mol biol.48:443介绍的程序,如smith和waterman,adv.appl.math(1981)2:482中所公开的。简而言之,gap程序将相似性定义为相似的比对符号(即核苷酸或氨基酸)的数量除以两个序列中较短的序列中的符号总数。gap程序的默认参数可能包括:(1)二元比较矩阵(包含对于同一性的1的值,和对于非同一性的0的值)和gribskov等人(1986)nucl.acids res.14:6745的加权比较矩阵,在schwartz和dayhoff,eds.,atlas of protein sequence and structure,national biomedical research foundation,pp.353
–
358(1979)中公开(或ednafull(emboss version of ncbi nuc4.4)替代矩阵);(2)对于每个间隙的3.0的罚分和在每个间隙中的每个符号的另外的0.10罚分(或10的间隙开放罚分,和0.5的间隙延伸罚分);和(3)对于端间隙无罚分。
[0046]
此外,两个给定的多核苷酸或多肽之间的序列同源性、相似性或同一性可以通过在限定的严格条件下通过southern杂交比较其序列来鉴定,并且限定的严格杂交条件在本技术的范围内并且可以被通过本领域普通技术人员熟知的方法限定(例如,j.sambrook et al.,molecular cloning,a laboratory manual,2nd edition,cold spring harbor laboratory press,cold spring harbor,new york,1989;f.m.ausubel et al.,current protocols in molecular biology,john wiley&sons,inc.,new york)。本公开内容的另一方面提供了编码变体的多核苷酸。
[0047]
如本文所用,术语“多核苷酸”是指核苷酸的聚合物,其中核苷酸单体通过共价键以长链形式彼此连接,并且通常是指具有一定最小长度的dna或rna链,更具体是编码变体的多核苷酸片段。
[0048]
编码根据本公开内容的蛋白质的变体的多核苷酸可以包括编码具有增强的活性的谷氨酸-半胱氨酸连接酶变体的任何核苷酸序列,但不限于此。
[0049]
编码本公开内容的谷氨酸-半胱氨酸连接酶的基因可以是gshl基因。该基因可以来源于酵母。具体地,该基因可以来源于属于酵母属的微生物,更具体地,可以来源于酿酒酵母。具体地,该基因可以是编码seq id no:1的氨基酸序列的基因,更具体地,是包括seq id no:2的碱基序列的序列,但不限于此。
[0050]
本公开内容的多核苷酸可以包括在编码区中进行的多种修饰,所述修饰通过密码子简并性或考虑到其中表达多肽的活生物体优选的密码子不提供改变从编码区表达的多肽的氨基酸序列。具体地,编码其中对应于seq id no:1的第86位的位置处的氨基酸被不同
氨基酸取代的蛋白质变体的任何多核苷酸序列可以包括在其中而没有限制。例如,本公开内容的多核苷酸可以具有多核苷酸序列,其编码本公开内容的蛋白质变体,具体是包含seq id no:3至21的氨基酸序列之一的蛋白质或与其具有同源性或同一性的多肽,不限于此。同源性或同一性如上所述。
[0051]
此外,多核苷酸可包括与使用已知基因序列例如与核苷酸序列完全或部分互补的核苷酸序列构建的探针在严格条件下杂交的核苷酸序列,以编码蛋白质变体,其中对应于seq id no:1的氨基酸序列的第86位的氨基酸被不同的氨基酸取代,没有限制。
[0052]
术语“严格条件”是指允许多核苷酸之间特异性杂交的条件。这些条件在已知文献(例如,j.sambrook等人)中详细公开。例如,条件可以包括在具有高同源性的基因之间进行杂交,例如,同源性为80%或更大,具体是90%或更大,更具体是95%或更大,甚至更具体是97%或更大,最具体99%或更大,而同源性低于上述同源性的基因之间不进行杂交,或者在盐浓度和温度为60℃、1
×
ssc和0.1%sds,具体是60℃、0.1
×
ssc、0.1%sds,更具体是68℃、0.1
×
ssc和0.1%sds的southern杂交的常规洗涤条件下进行杂交一次,具体地进行杂交两次或三次。然而,严格条件不限于此,并且可以由本领域技术人员根据预期目的适当调整。
[0053]
杂交需要两个多核苷酸具有互补序列,但是根据杂交的严格程度可能存在碱基错配。术语“互补”用于描述能够相互杂交的核苷酸碱基之间的关系。例如,对于dna,腺苷与胸腺嘧啶互补,胞嘧啶与鸟嘌呤互补。因此,本公开内容不仅可以包括基本上相似的核酸序列,还可以包括与整个序列互补的分离的核酸片段。
[0054]
具体地,具有同源性或同一性的多核苷酸可以使用上述杂交条件检测,包括在55℃的tm值下的杂交过程。此外,tm值可能是,但是不限于60℃、63℃或65℃,并且本领域技术人员可根据预期目的适当调整。
[0055]
多核苷酸杂交的适当严格程度可以取决于多核苷酸的长度,并且互补程度及其参数在本领域中是公知的。
[0056]
本公开内容的另一个方面提供了载体,其包括编码蛋白质变体的多核苷酸。
[0057]
如本文所用,术语“载体”是指包括编码靶蛋白的多核苷酸的碱基序列的dna构建体,其可操作地连接到合适的调控序列以在合适的宿主细胞中表达靶蛋白。调控序列可以包括允许启动转录的启动子、用于调控转录的操纵子序列、编码合适的mrna核糖体结合位点的序列以及调控转录和翻译终止的序列。在将载体引入合适的宿主细胞后,它可以独立于宿主基因组进行复制或发挥作用,并且可以整合到基因组本身中。
[0058]
本公开内容中使用的载体没有具体限制,并且可以使用本领域已知的任何载体。作为酵母表达的载体,可以使用整合酵母质粒(yip)和染色体外质粒载体。染色体外质粒载体可以包括游离型酵母质粒(yep)、复制型酵母质粒(yrp)和酵母着丝粒质粒(ycp)。此外,人工酵母染色体(yac)也可以用作本公开内容的载体。作为具体实例,可用载体可包括peschis、pesc-leu、pesc-trp、pesc-ura、gateway pyes-dest52、pao815、pgapz a、pgapz b、pgapz c、pgapαa、pgapαb、pgapαc、ppic3.5k、ppic6 a、ppic6 b、ppic6 c、ppic6αa、ppic6αb、ppic6αc、ppic9k、pyc2/ct、pyd1酵母展示载体、pyes2、pyes2/ct、pyes2/nt a、pyes2/nt b、pyes2/nt c、pyes2/ct、pyes2.1、pyes-dest52、ptef1/zeo、pfld1、pichiapinktm、p427-tef、p417-cyc、pgal-mf、p427-tef、p417-cyc、ptef-mf、pby011、psgp47、psgp46、psgp36、
psgp40、zm552、pag303gal-ccdb、pag414gal-ccdb、pas404、pbridge、pgad-gh、pgad t7、pgbk t7、phis-2、pobd2、prs408、prs410、prs418、prs420、prs428、酵母微米a型(yeast micron a form)、prs403、prs404、prs405、prs406、pyj403、pyj404、pyj405和pyj406,但不限于此。
[0059]
例如,染色体中编码靶蛋白的多核苷酸可以使用用于染色体插入细胞的载体被突变的多核苷酸替换。可以通过本领域已知的任何方法将多核苷酸插入染色体,所述方法例如同源重组,但不限于此。可以进一步包括选择标记以确认染色体插入。选择标记用于选择用载体转化的细胞,即确认所期望的多核苷酸的插入,并且选择标记的实例可以包括提供可选择表型的标记,例如耐药性、营养需求、细胞毒性剂抗性或变体多肽的表面表达。只有表达选择标记的细胞才能在用选择剂处理的环境下存活或显示不同的表型,因此可以选择转化的细胞。
[0060]
如本文所用,术语“转化”是指将包括编码谷氨酸-半胱氨酸连接酶变体的多核苷酸的载体引入宿主细胞,使得由多核苷酸编码的蛋白质在宿主中表达细胞的过程。转化的多核苷酸可以是插入宿主细胞染色体的形式,或者可以是位于染色体外的形式,只要蛋白质在宿主细胞中表达。此外,多核苷酸包括编码谷氨酸-半胱氨酸连接酶变体的dna和rna。多核苷酸可以以任何形式引入宿主细胞,只要将多核苷酸引入宿主细胞并且在其中表达蛋白质。例如,可以以表达盒的形式将多核苷酸引入宿主细胞,该表达盒是包含自我复制所需的所有基本要素的基因构建体。表达盒通常可以包括与多核苷酸有效连接的启动子、转录终止信号、核糖体结合位点和翻译终止信号。表达盒可以是可自我复制的表达载体的形式。此外,可以将多核苷酸以其原始形式引入宿主细胞中并且可操作地连接到在宿主细胞中表达所需的序列,但不限于此。
[0061]
此外,如本文所用,术语“可操作地连接”是指编码本公开内容的多肽的多核苷酸序列和启动并介导多核苷酸序列转录的启动子序列之间的功能连接。根据本公开内容的转化方法包括能够将载体引入宿主细胞的任何方法,并且可以通过根据宿主细胞选择的本领域公知的合适标准技术进行。例如,可以使用电穿孔、磷酸钙(capo4)沉淀、氯化钙(cacl2)沉淀、显微注射、聚乙二醇(peg)法、deae-葡聚糖法、阳离子脂质体法和醋酸锂-dmso法,但本公开内容不限于此。
[0062]
本公开内容可以通过包括以下至少一种来提供生产谷胱甘肽的微生物:变体;编码变体的多核苷酸;和包含多核苷酸的载体。
[0063]
微生物可以是表达变体的微生物或其中引入了变体的微生物。
[0064]
如本文所用,术语“包括变体的微生物”、“其中引入变体的微生物”或“表达变体的微生物”可以指通过增强天然具有低生产谷胱甘肽能力的微生物生产谷胱甘肽的能力或通过向不能生产谷胱甘肽的亲本菌株提供生产谷胱甘肽的能力而制备的微生物。具体地,微生物可以是表达谷氨酸-半胱氨酸连接酶变体的微生物,该变体包括在seq id no:1的氨基酸序列中的至少一个氨基酸突变,并且该氨基酸突变可以包括对应于n末端起第86位的氨基酸被不同的氨基酸取代。此外,微生物可以是表达谷氨酸-半胱氨酸连接酶变体的微生物,其中对应于seq id no:1的氨基酸序列的第86位的氨基酸被不同的氨基酸取代,但不限于此。
[0065]
谷氨酸-半胱氨酸连接酶及其变体如上所述。
[0066]
如本文所用,“待表达/正在表达”的蛋白质是指其中靶蛋白被引入微生物或在微生物中表达的状态。在蛋白质存在于微生物中的情况下,蛋白质的活性与其内源性蛋白质的活性或修饰前的活性相比增强。鉴于本公开内容的目的,“靶蛋白”可以是上述谷氨酸-半胱氨酸连接酶变体。
[0067]
具体地,术语“引入蛋白质”是指将向不具有具体蛋白质的微生物提供该蛋白质的活性,或与该蛋白质的固有活性或修饰前的活性相比增强该蛋白质的活性。例如,引入蛋白质可以指将编码具体蛋白质的多核苷酸引入染色体或将包括编码具体蛋白质的多核苷酸的片段或载体引入微生物,从而表达蛋白质的活性。此外,“活性增强”可以是指微生物的具体蛋白质的活性与固有活性或修饰前的活性相比时增强。术语“固有活性”是指当微生物通过天然或人工遗传变异转化时,亲本菌株在转化之前所具有的具体蛋白质的活性。
[0068]
具体地,根据本公开内容的活性增强可以通过包括以下的方法的至少一种来进行:增加编码蛋白质变体的基因的拷贝数、将突变引入编码蛋白质变体的基因的表达调控序列、用具有更强活性的序列替换编码蛋白质变体的基因的表达调控序列、用编码蛋白质变体的基因替换编码野生型蛋白质的染色体基因、另外将突变引入编码蛋白质变体的基因中以增强蛋白质变体的活性和将蛋白质变体引入微生物中,但不限于此。
[0069]
尽管不具体限于此,但基因拷贝数的增加以与载体可操作地连接的形式或以整合到宿主细胞的染色体中的形式进行。具体地,该方法可以通过向宿主细胞中引入载体来进行,该载体与宿主无关地复制和发挥功能并且与编码本公开内容的蛋白质的多核苷酸可操作地连接。可选地,可以通过向宿主细胞中引入载体来增加基因的拷贝数,该载体将多核苷酸插入宿主细胞的染色体中,并且与多核苷酸可操作地连接。可通过本领域公知的任何方法例如同源重组进行将多核苷酸插入染色体。
[0070]
接下来,可以通过由缺失、插入、非保守取代、保守取代或其任何组合在核苷酸序列中诱导变异以进一步增强表达调控序列的活性,或通过用具有更强活性的核苷酸序列替换核苷酸序列来进行表达调控序列的修饰以增加多核苷酸的表达,但不限于此。表达调控序列可以包括启动子、操纵子序列、核糖体结合位点编码序列和用于调控转录和翻译的序列,但不限于此。
[0071]
比固有启动子更强的启动子可以连接到多核苷酸表达单元的上游,但不限于此。例如,当宿主细胞是酵母时,可用的启动子可以包括tef1启动子、tef2启动子、gal10启动子、gal1启动子、adh1启动子、adh2启动子、pho5启动子、gal1-10启动子、tdh3启动子(gpd启动子)、tdh2启动子、tdh1启动子、pgk1启动子、pyk2启动子、eno1启动子、eno2启动子和tpi1启动子,但不限于此。此外,可以通过由缺失、插入、非保守取代、保守取代或其任意组合在表达调控序列中诱导变异以进一步增强多核苷酸序列的活性,或通过用经修饰以具有更强活性的核苷酸序列替换核苷酸序列来进行所述的染色体上的多核苷酸序列的修饰,但不限于此。
[0072]
通常,引入和增强蛋白质活性可以使相应蛋白质的活性或浓度与野生型或未修饰的微生物菌株的活性或浓度相比增加1%、10%、25%、50%、75%、100%、150%、200%、300%、400%或500%,最大1000%或2000%,但不限于此。
[0073]
如本文所用,术语“未修饰的微生物”不排除具有可能在微生物中自然发生的突变的菌株,并且可以是野生型菌株、不包括蛋白质变体的微生物、或未用包含编码蛋白质变体
的多核苷酸的载体转化的微生物。
[0074]
在本公开内容中,包括谷氨酸-半胱氨酸连接酶变体或编码其的多核苷酸的微生物可以是例如通过用包括多核苷酸的载体转化微生物来制备的重组微生物,但不限于此。重组微生物可以是酵母,例如属于酵母属的微生物,具体是酿酒酵母。例如,微生物可以是登录号为kccm12659p的酿酒酵母菌株,但不限于此。
[0075]
如本文所用,术语“谷胱甘肽”可以与“gsh”互换使用并且指由以下三个氨基酸组成的三肽化合物:谷氨酸、半胱氨酸和甘氨酸。谷胱甘肽可用作药物、保健功能食品、调味成分、食品和饲料添加剂、化妆品等的原料,但不限于此。
[0076]
如本文所用,术语“谷胱甘肽生产微生物”包括通过天然或人工遗传修饰来修饰的微生物,并且可以指具有特定机制的微生物,其经由引入外源基因或通过遗传修饰增强或灭活内源基因来减弱或增强,以便生产谷胱甘肽。鉴于本公开内容的目的,生产谷胱甘肽的微生物可以是指包含谷氨酸-半胱氨酸连接酶并且与野生型或未修饰的微生物相比能够生产大量目标谷胱甘肽的微生物。“谷胱甘肽生产微生物”可以与“生产谷胱甘肽的微生物”、“具有生产谷胱甘肽的能力的微生物”、“生产谷胱甘肽的菌株”、“具有生产谷胱甘肽的能力的菌株”等互换使用。
[0077]
谷胱甘肽生产微生物可以是重组微生物。重组微生物如上所述。
[0078]
谷胱甘肽生产微生物的类型没有具体限制,只要由此生产谷胱甘肽,但可以是属于酵母属的微生物,具体是酿酒酵母,但不限于此。
[0079]
包括变体的谷胱甘肽生产微生物的亲本菌株包括没有具体限制,只要该菌株具有生产谷胱甘肽的能力。微生物可包括用于以下的修饰:增强生物合成途径以增加谷胱甘肽生产能力、释放反馈抑制和使减弱降解途径或生物合成途径的基因失活,并且这种修饰不排除天然发生的修饰。然而,本公开内容不限于此。
[0080]
本公开内容的另一个方面提供生产谷胱甘肽的方法,该方法包括培养微生物。微生物和谷胱甘肽如上所述。谷胱甘肽可以通过培养菌株在菌株中积累。
[0081]
关于用于培养本公开内容的菌株的培养基或其他培养条件,可以使用通常用于培养属于酵母属的微生物的任何培养基而没有限制。具体地,本公开内容的菌株可以在含有适当碳源、氮源、磷源、无机化合物、氨基酸和/或维生素的普通培养基中,在需氧或厌氧条件下培养,同时调整温度、ph等。
[0082]
在本公开内容中,作为碳源,可以使用碳水化合物,例如葡萄糖、果糖、蔗糖和麦芽糖;糖醇,例如甘露醇和山梨醇;有机酸,例如丙酮酸、乳酸和柠檬酸;以及氨基酸如谷氨酸、甲硫氨酸和赖氨酸,但不限于此。此外,可以使用天然有机营养物质,例如淀粉水解物、糖蜜、黑糖蜜、米糠、木薯、甘蔗渣和玉米浆,可以使用碳水化合物,例如葡萄糖和无菌预处理糖蜜(即转化为还原糖的糖蜜),并且也可以使用合适量的任何其他碳源而没有限制。这些碳源可以单独使用或以其至少两种组合使用。
[0083]
作为氮源,可以使用无机氮源,例如氨、硫酸铵、氯化铵、乙酸铵、磷酸铵、碳酸铵和硝酸铵;和有机氮源,例如氨基酸、蛋白胨、nz-胺、肉提取物、酵母提取物、麦芽提取物、玉米浆、酪蛋白水解物、鱼或其降解产物、和脱脂豆饼或其降解产物。这些氮源可以单独使用或以其至少两种组合使用。
[0084]
作为磷源,可以使用磷酸二氢钾、磷酸二钾或与其对应的含钠盐。作为无机化合
物,可以使用氯化钠、氯化钙、氯化铁、硫酸镁、硫酸铁、硫酸锰、碳酸钙等。
[0085]
培养基可进一步包括氨基酸、维生素和/或合适的前体。具体地,可以向菌株的培养基中添加l-氨基酸等。具体地,可以向培养基中添加甘氨酸、谷氨酸和/或半胱氨酸,如果需要,可以进一步向其中添加l-氨基酸如赖氨酸,但本公开内容不限于此。
[0086]
培养基和前体可以以分批或连续过程添加到培养物中,但不限于此。
[0087]
在本公开内容中,在菌株的培养过程期间,可以适当地向培养物中添加例如氢氧化铵、氢氧化钾、氨、磷酸和硫酸的化合物,以调整培养物的ph值。另外,为了抑制培养期间的泡沫形成,也可以添加例如脂肪酸聚乙二醇酯的消泡剂。此外,可以将氧气或含氧气体注入到培养物中以将培养物维持在需氧条件下,并且可以将氮气、氢气或二氧化碳气体注入培养物中以将培养物维持在厌氧和微需氧条件下,而无需为此注入任何其他气体。
[0088]
培养物的温度可以维持在25℃至40℃,更具体地在28℃至37℃,但不限于此。培养可以继续直到获得所期望量的所期望物质,具体地持续1小时至100小时,但不限于此。
[0089]
生产谷胱甘肽的方法可以进一步包括在培养步骤之后的附加过程。可以根据谷胱甘肽的使用目的适当地选择附加过程。
[0090]
具体地,生产谷胱甘肽的方法可以包括在微生物培养之后从选自菌株、干燥产物、提取物、培养产物和其裂解物中的至少一种回收谷胱甘肽。
[0091]
方法可以进一步包括在回收步骤之前或同时裂解菌株。菌株的裂解可以通过本公开内容所涉及领域中常用的任何方法进行,例如通过热处理或通过使用用于裂解的缓冲溶液、超声波仪和弗式压碎器(french press)。同样,裂解步骤可包括通过细胞壁裂解酶、核酸酶、反式核苷酸酶、蛋白酶等进行的酶促反应,但不限于此。
[0092]
鉴于本公开内容的目的,可以通过生产谷胱甘肽的方法制备各自具有高谷胱甘肽含量的干酵母、酵母提取物、酵母提取物混合粉和纯谷胱甘肽。然而,本公开内容不限于此,并且这些产品可以根据期望的产品适当地制备。
[0093]
在本公开内容中,干酵母可以与“菌株的干燥产物”互换使用。干酵母可以通过干燥其中积累谷胱甘肽的酵母菌株来制备,具体地,它可以包含在饲料组合物、食品组合物等中,但不限于此。
[0094]
在本公开内容中,酵母提取物可以与例如“菌株的提取物”的术语互换使用。菌株的提取物可以指从菌株中分离细胞壁后剩余的物质。具体地,菌株的提取物可以指通过裂解细胞获得的组分中的除细胞壁之外的剩余组分。菌株的提取物包括谷胱甘肽和除谷胱甘肽外的选自蛋白质、碳水化合物、核酸和纤维的一种或多种其他组分,但不限于此。
[0095]
可以使用本领域公知的任何合适的方法进行回收步骤,并且可以回收作为靶物质的谷胱甘肽。
[0096]
回收步骤可以包括纯化过程。可以通过仅从菌株中分离谷胱甘肽来进行纯化过程。经由纯化过程,可以制备纯谷胱甘肽。
[0097]
如果需要,制备谷胱甘肽的方法可以进一步包括将赋形剂与选自菌株,其干燥产物、提取物、培养物和裂解物以及从中回收的谷胱甘肽中的一种混合。通过混合步骤,可以制备酵母提取物混合粉末。
[0098]
可以根据期望的用途或形式适当选择和使用赋形剂,所述赋形剂可以例如选自淀粉、葡萄糖、纤维素、乳糖、糖原、d-甘露醇、山梨醇、乳糖醇、麦芽糖糊精、碳酸钙、合成硅酸
铝、磷酸一氢钙、硫酸钙、氯化钠、碳酸氢钠、纯化羊毛脂、糊精、海藻酸钠、甲基纤维素、胶体硅胶、羟丙基淀粉、羟丙基甲基纤维素、丙二醇、酪蛋白、乳酸钙、羧甲基淀粉钠(primojel)和阿拉伯胶,具体地,其可包含选自淀粉、葡萄糖、纤维素、乳糖、糊精、糖原、d-甘露醇和麦芽糊精中的至少一种组分,但不限于此。
[0099]
赋形剂可包括例如防腐剂、湿润剂、分散剂、悬浮剂、缓冲剂、稳定剂或等渗剂,但不限于此。
[0100]
本公开内容的另一方面提供谷氨酸-半胱氨酸连接酶变体用于生产谷胱甘肽的用途。本公开内容的另一方面提供了微生物用于生产谷胱甘肽的用途。谷氨酸-半胱氨酸连接酶变体、微生物和谷胱甘肽如上所述。
[0101]
发明方式
[0102]
在下文中,将参考以下实施例和实验例更详细地描述本公开内容。然而,以下实施例和实验例仅用于举例说明本公开内容,本公开内容的范围不限于此。
[0103]
实施例1:选择谷胱甘肽生产菌株和确认谷胱甘肽生产能力
[0104]
从含有多种菌株的酵母块中获得菌株,并改进其特性以选择具有谷胱甘肽生产能力的菌株。
[0105]
具体地,从大韩民国京畿道的华城、平泽、龙仁等20个地区采集例如大米、大麦、绿豆和燕麦的谷物样品,将其粉碎、揉捏、包裹在布中,用力压成形状,用稻草包裹发酵10天,然后慢慢干燥,制成酵母块。
[0106]
进行以下实验以从制备的酵母块中分离多种菌株。将45ml盐水溶液添加到5g酵母块中并使用混合器粉碎。为了纯净地分离酵母菌株,通过连续稀释将所得物稀释,涂在ypd琼脂上(每1l蒸馏水10g/l酵母提取物、20g/l菌蛋白胨和20g/l葡萄糖),并在30℃下培养48小时。然后,根据菌落形态和显微镜验证,将酵母菌落划线在ypd琼脂上。将25ml ypd肉汤接种在250ml锥形瓶中,将纯净分离的菌株接种于其上,并在30℃和200rpm的摇动培养箱中培养48小时。通过鉴定谷胱甘肽生产来筛选菌株。
[0107]
为了改进主要分离的菌株,在分离的菌株中诱导随机突变。具体地,从酵母块中分离确认具有谷胱甘肽生产能力的菌株并命名为cj-37菌株。将cj-37菌株在固体培养基中培养并接种到肉汤中以获得其培养液,并使用紫外灯将培养液暴露于紫外光下。将暴露于紫外线的培养液平板接种于平板培养基上后,仅分离已形成菌落的突变菌株,鉴定其谷胱甘肽生产。结果,在突变菌株中,选择展现最高谷胱甘肽产量的菌株作为谷胱甘肽生产菌株,命名为cj-5菌株,根据布达佩斯条约于2019年7月31日保藏在韩国微生物培养中心(kccm),并指定为登录号kccm12568p。
[0108]
实施例2:进一步提高谷胱甘肽生产能力的实验为了进一步提高cj-5菌株的谷胱甘肽生产能力,以下列方式诱导突变。
[0109]
将cj-5菌株在固体培养基中培养并接种到肉汤中以获得培养液,并使用紫外灯将培养液暴露于紫外光下。将暴露于紫外光的培养液平板接种于平板培养基上后,仅分离已形成菌落的突变菌株。分离展现最高谷胱甘肽产量的菌株,命名为cc02-2490菌株,根据布达佩斯条约于2020年1月17日保藏在韩国微生物培养中心(kccm),并指定为登录号kccm12659p。作为分析谷胱甘肽生物合成基因gsh1的碱基序列的结果,在增强菌株的谷胱甘肽生产能力方面,证实了由gsh1基因编码的gsh1蛋白的第86位氨基酸半胱氨酸已经被精
氨酸取代。
[0110]
实施例3:gsh1 c86残基突变的实验
[0111]
基于实施例2的结果,考虑到gsh1蛋白的第86位的氨基酸在谷胱甘肽生产中很重要,制备酿酒酵母(s.cerevisiae)cen.pk2-1d和酿酒酵母(s.cerevisiae)cj-5的突变菌株以便表达蛋白质变体,其中gsh1蛋白的第86位的半胱氨酸被不同的氨基酸取代,并且鉴定谷胱甘肽生产的增加。
[0112]
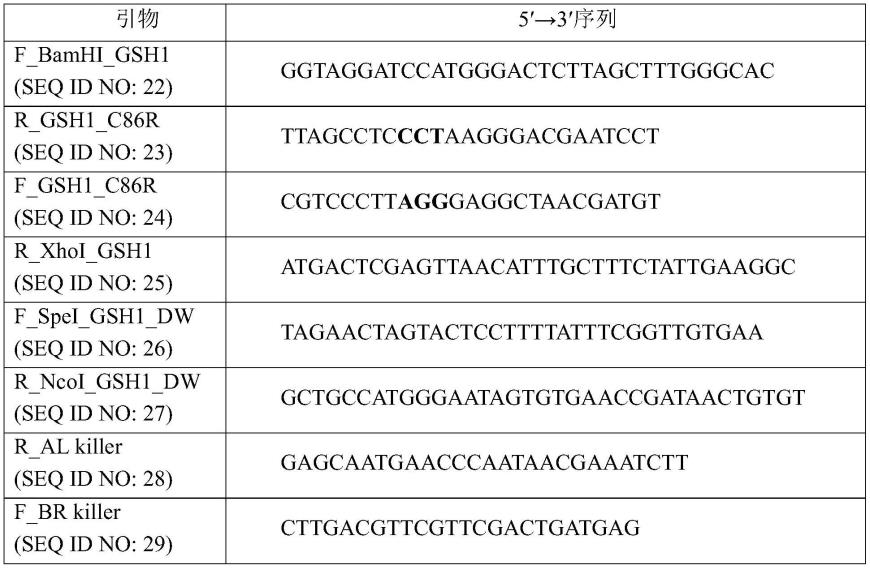
为了制备其中酿酒酵母的gsh1蛋白的第86位的半胱氨酸被精氨酸取代的菌株,参考lee th等人的论文(j.microbiol.biotechnol.(2006),16(6),979-982)使用pwal100和pwbr100质粒。具体地,使用如下的cj-5菌株的基因组dna作为模板进行聚合酶链式反应(pcr)。通过使用引物seq id no:22和23进行pcr,获得gsh1蛋白n末端的部分序列,其包括n末端bamhi侧翼序列、gsh1 orf起始密码子和c86r突变编码序列,并且通过使用引物seq id no:24和25进行pcr,获得gsh1蛋白c末端的部分序列,其包括c末端xhoi侧翼序列、gsh1 orf终止密码子和c86r突变编码序列。接着,作为使用2个序列为模板、利用引物seq id no:22和25进行重叠pcr的结果,获得了gsh1orf片段,其包含编码gsh1修饰蛋白的序列,其中第86位的半胱氨酸被精氨酸取代,以及n末端bamhi和c末端xhoi限制酶序列。用bamhi和xhoi处理orf片段,然后克隆到用相同酶处理的pwal100载体中以制备pwal100-gsh1(c86r)载体。
[0113]
此外,通过使用cj-5菌株的基因组dna作为模板和引物seq.id no:26和27进行pct,获得包括n末端spel和c末端ncol限制酶序列的gsh1 orf终止密码子下游的500bp,并用spei和ncoi限制酶处理。随后,将所得物克隆到用相同酶处理的pwbr100中以制备pwbr100-gsh1载体。
[0114]
为了制备待引入酵母中的最终dna片段,使用如上所述制备的pwal100-gsh1(c86r)载体作为模板和引物seq id no:22和28获得包括编码精氨酸突变的序列和部分klura3的pcr产物,使用pwbr100-gsh1载体作为模板和引物seq id no:29和27,获得包括部分klura3和gsh1终止密码子下游500bp的pcr产物。用相同摩尔比的pcr产物转化酿酒酵母cen.pk2-1d和酿酒酵母cj-5。经由95℃变性5分钟,53℃退火1分钟,72℃聚合持续1分钟/kb进行pcr,酵母的转化按照从来自geitz的论文(nucleic acid research,20(6),1425)中公开的方法改进的醋酸锂法进行。具体地,将od为0.7至1.2的酵母细胞用醋酸锂/te缓冲液洗涤两次,并与pcr产物和单链dna(sigma d-7656)混合。将细胞在醋酸锂/te/40%peg缓冲液中静态培养条件下在30℃下培养30分钟并在42℃下培养15分钟。然后,将细胞在不包括尿嘧啶的sc(2%葡萄糖)琼脂平板中培养直到菌落可见,以获得其中引入了gsh1 c86r突变编码序列和klura3基因的菌株。随后,为了去除klura3,将菌株在2ml ypd中培养过夜,以1/100的比例稀释,平板接种在包含0.1%5-foa的sc(2%葡萄糖)琼脂板上以制备酿酒酵母cen.pk2-1d gsh1 c86r突变的菌株和酿酒酵母cj-5gsh1 c86r突变的菌株,已从其去除了尿嘧啶标记。除了使用引物对seq id no:23和24,其中编码第86位精氨酸的序列被编码不同氨基酸的序列取代,还以相同方式制备了能够表达其中半胱氨酸被精氨酸以外的18种氨基酸取代的gsh1修饰蛋白质的菌株。
[0115]
表1
[0116][0117]
在将如上所述制备的菌株培养26小时后,测量生产的谷胱甘肽(gsh)的浓度并将其列于表2和3中。
[0118]
表2
[0119][0120]
表3
[0121][0122]
基于实验结果,证实了通过用不同氨基酸取代gsh1蛋白的第86位的半胱氨酸获得的谷胱甘肽生产能力与通过野生型gsh1蛋白获得的谷胱甘肽生产能力相比增加了高达27%。
[0123]
基于此,证实可通过用不同氨基酸取代gsh1蛋白的第86位的半胱氨酸制备的gsh1变体具有显著提高的谷胱甘肽生产能力。
[0124]
实施例4:证实另一个cys残基对谷胱甘肽生产的影响
[0125]
作为比较例,对gsh1蛋白的另一个cys残基进行修饰,并鉴定通过该修饰获得的谷胱甘肽生产能力。结果如下表4所示。
[0126]
表4
[0127][0128]
参考实验结果,除了第86位的半胱氨酸以外的半胱氨酸残基对增加谷胱甘肽生产没有影响,反而降低了谷胱甘肽生产。
[0129]
基于此,证实了并非所有存在于蛋白质中的半胱氨酸残基都对增加谷胱甘肽生产
有影响。此外,证实了在本公开内容中开发的新型gsh1变体增加了谷胱甘肽生产。
[0130]
由于以高产率生产谷胱甘肽的酵母,其干燥产物、提取物、培养物和裂解物以及所生产的谷胱甘肽具有抗氧化、解毒和免疫力增强作用,它们可以有效地用于制备化妆品组合物、食品组合物、饲料组合物和药物组合物。
[0131]
本发明的上述描述是为了说明性目的而提供,本领域技术人员应当理解,在不改变本发明的技术构思和基本特征的情况下,可以进行多种变化和修改。因此,很明显,上述实施方式在所有方面都是说明性的,而不是限制本发明。本文公开的多种实施方式不旨在进行限制,其真实范围和精神由所附权利要求指示。本发明仅受所附权利要求的条款以及这些权利要求所享有的等效物的全部范围的限制。
[0132]
[0133]
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1