利用细胞膜穿透肽而不依赖于基因重组的植物基因组编辑技术的制作方法
1.本发明涉及使用新复合物的基因组编辑技术。详细而言,本发明涉及不依赖于基因重组而将基因组编辑酶和核酸直接引入到植物细胞内的技术。
2.本技术对作为日本专利申请的特愿2020-211301主张优先权,通过参照使特愿2020-211301的全部内容与本技术一体化。
背景技术:
[0003]“基因组编辑技术”作为改变生物功能的革新技术,在基础研究到应用研究(例如医疗或农作物育种)的广泛领域利用。然而,在以植物为对象的现有的基因组编辑技术中,存在“操作需要大量时间和繁杂度”的致命的课题。作为该最主要的理由,可列举:现有的植物基因组编辑技术依赖于基因重组技术。在现有的植物基因组编辑技术中,可采取以下方法:通过基因重组将基因组编辑盒基因(合成基因组编辑酶和靶标基因特异性核酸的基因)引入到植物细胞内诱发基因组编辑,之后选择缺失了基因组编辑酶基因的细胞,从而获得基因组编辑细胞。然而,对于无法适用基因重组技术的植物种类无法引入基因组编辑盒基因,因此目前可进行基因组编辑的植物种类仅限于一部分。另外,一旦将作为外源基因的基因组编辑酶基因引入植物细胞内,所以只要未去除该外源基因,就视为基因重组生物,因此直接进行商业利用非常困难。外源基因的去除仅通过重复交配即可进行,因此如上所述基因组编辑盒基因的去除存在需要大量时间的问题。
[0004]
为了克服如上所述的现有植物基因组编辑技术的诸多问题,不依赖于基因重组的基因组编辑技术的开发正在积极进行。具体而言,提出了如下的方法:通过将基因组编辑酶以蛋白的状态直接引入到植物细胞内,不经基因重组而诱发基因组编辑(非专利文献1)。然而,由于植物细胞具有带强负电荷的细胞壁,所以在想要将蛋白引入到植物细胞内的情况下,碱性蛋白被细胞壁捕获,而另一方面酸性蛋白被细胞壁排斥。虽然可利用粒子枪或电穿孔法强制性地将蛋白引入到植物细胞内,但需要特殊、高价的专用装置。迄今为止,完全没有简便地将蛋白引入到具有细胞壁的状态的植物细胞内的通用方法。因此,也完全没有将基因组编辑酶简便地引入到植物细胞内的方法,这样的现状是极大地推迟利用了基因组编辑技术的植物功能改变(特别是农作物育种)的主要原因。
[0005]
现有技术文献非专利文献非专利文献1:svitashev s等人, nat. commun. 2016; 7: 13274。
技术实现要素:
[0006]
发明所要解决的课题人们在寻求不依赖于基因重组、而且可在广泛的植物种类中简便地进行植物基因组编辑的技术。
[0007]
用于解决课题的手段本发明人为了解决上述课题反复进行深入研究,发现了:通过使用包含基因组编辑酶和细胞膜穿透肽(以下,有时称为“cpp”)的复合物、以及包含基因组编辑盒(基因组编辑酶和靶标基因特异性核酸)和cpp的复合物,无需基因重组即可将基因组编辑盒直接引入到植物细胞内,从而完成了本发明。
[0008]
即,本发明提供以下内容:(1) 复合物,其是包含基因组编辑酶和cpp的复合物,其中,cpp与基因组编辑酶融合。
[0009]
(2) 复合物,其是包含基因组编辑酶、靶标基因特异性核酸和cpp的复合物,其中,cpp与基因组编辑酶和/或靶标基因特异性核酸融合。
[0010]
(3) (1)或(2)所述的复合物,其中,cpp与基因组编辑酶和/或靶标基因特异性核酸进行共价结合。
[0011]
(4) (3)所述的复合物,其中,cpp与基因组编辑酶进行共价结合。
[0012]
(5) (2)所述的复合物,其中,聚阳离子部分与cpp融合,聚阳离子部分与靶标基因特异性核酸进行静电结合。
[0013]
(6) (5)所述的复合物,其中,聚阳离子部分与cpp进行共价结合。
[0014]
(7) (6)所述的复合物,其中,聚阳离子部分为聚阳离子肽。
[0015]
(8) (7)所述的复合物,其中,聚阳离子肽包含10个以上的赖氨酸残基或10个以上的精氨酸残基。
[0016]
(9) (1)~(8)中任一项所述的复合物,其中,cpp的80%以上的氨基酸残基为组氨酸残基,cpp的长度为8个氨基酸~数十个氨基酸。
[0017]
(10) (9)所述的复合物,其中,cpp的所有的氨基酸残基均为组氨酸残基。
[0018]
(11) (1)~(10)中任一项所述的复合物,该复合物还包含信号序列。
[0019]
(12) (1)~(11)中任一项所述的复合物,该复合物还包含亚结构域。
[0020]
(13) 基因组编辑方法,该方法包括:将(1)~(12)中任一项所述的复合物引入到细胞内。
[0021]
(14) (13)所述的方法,其中,细胞为植物细胞、藻类细胞、丝状菌细胞或酵母细胞。
[0022]
(15) 基因组编辑用试剂盒,其包含(1)~(12)中任一项所述的复合物或其构成成分。
[0023]
(16) (15)所述的试剂盒,该试剂盒用于植物、藻类、丝状菌或酵母的基因组编辑。
[0024]
发明效果根据本发明,无需基因重组,即可将基因组编辑酶或基因组编辑盒直接引入到植物细胞内。因此,即使对于无法适用基因重组技术的植物种类,也可引入基因组编辑酶或基因组编辑盒,也可进行基因组编辑。只需在培养基中孵育本发明的复合物和植物细胞,即可将基因组编辑酶或基因组编辑盒引入到植物细胞内。另外,本发明的基因组编辑的效率高。总之,通过使用本发明的复合物,无需基因重组,即可对广泛的植物种类简便且高效地进行基因组编辑。通过本发明得到的基因组编辑植物不具有外源基因,不属于基因重组生物,因此可立即进行商业利用,商业价值非常高。
[0025]“将基因组编辑酶直接引入到植物细胞内”是指将基因组编辑酶作为具有活性的蛋白引入到植物细胞内。“将基因组编辑盒直接引入到植物细胞内”是指将基因组编辑酶与靶标基因特异性核酸的复合物引入到植物细胞内,将基因组编辑酶与核酸的复合物作为具有活性的蛋白复合物引入到植物细胞内。
附图说明
[0026]
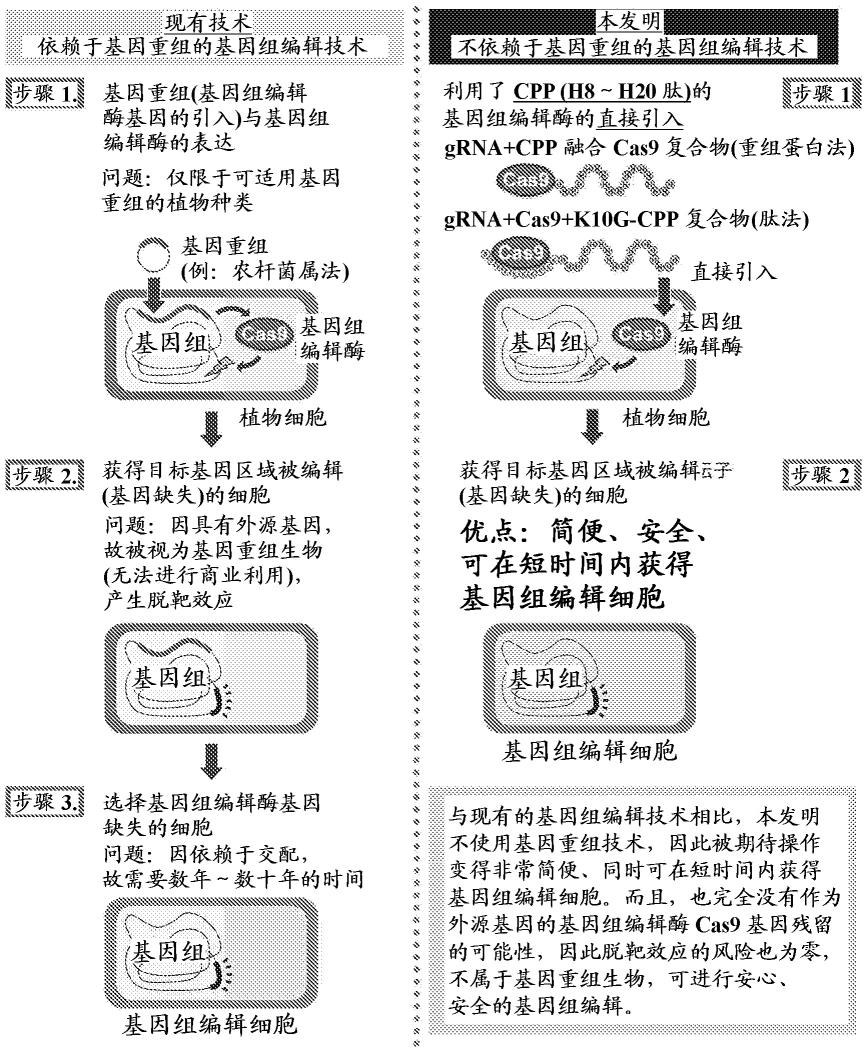
[图1] 图1是用于比较说明使用了本发明的复合物的基因组编辑法与现有方法的图。
[0027]
[图2] 图2是显示在实施例中作为靶标基因的日本柳杉镁螯合酶基因(cjchli基因)的核苷酸序列的图。图中,“grna1”和“grna2”分别表示grna1和grna2的识别位点。
[0028]
[图3] 图3是显示在实施例中确认到基因组编辑(基因缺失)的grna1所识别的区域的核苷酸序列的图。发生了基因组编辑(基因缺失)的位点用
“‑”
表示。另外,样品表示处理植物细胞时的条件,读数表示实施了核苷酸序列分析的序列数和确认到基因组编辑(基因缺失)的序列数,效率表示基因组编辑效率。下划线表示显著确认到基因组编辑(基因缺失)的条件。
[0029]
[图4] 图4是显示在实施例中确认到基因组编辑(基因缺失)的grna2所识别的区域的核苷酸序列的图。发生了基因组编辑(基因缺失) 的位点用
“‑”
表示。另外,样品表示处理植物细胞时的条件,读数表示实施了核苷酸序列分析的序列数和确认到基因组编辑(基因缺失)的序列数,效率表示基因组编辑效率。下划线表示显著确认到基因组编辑(基因缺失)的条件。
[0030]
[图5] 图5是显示在实施例中确认到基因组编辑(基因缺失)的grna2所识别的区域的核苷酸序列的图。发生了基因组编辑(基因缺失)的位点用
“‑”
表示。另外,样品表示处理植物细胞时的条件,读数表示实施了核苷酸序列分析的序列数和确认到基因组编辑(基因缺失)的序列数,效率表示基因组编辑效率。下划线表示显著确认到基因组编辑(基因缺失)的条件。
[0031]
[图6] 图6是显示在实施例中确认到基因组编辑(基因缺失)的grna2所识别的区域的核苷酸序列的图。发生了基因组编辑(基因缺失)的位点用
“‑”
表示。另外,样品表示处理植物细胞时的条件,读数表示实施了核苷酸序列分析的序列数和确认到基因组编辑(基因缺失)的序列数,效率表示基因组编辑效率。下划线表示显著确认到基因组编辑(基因缺失)的条件。
[0032]
[图7] 图7是显示在实施例中确认到基因组编辑(基因缺失)的grna2所识别的区域的核苷酸序列的图。发生了基因组编辑(基因缺失)的位点用
“‑”
表示。另外,样品表示处理植物细胞时的条件,读数表示实施了核苷酸序列分析的序列数和确认到基因组编辑(基因缺失)的序列数,效率表示基因组编辑效率。下划线表示显著确认到基因组编辑(基因缺失)的条件。
[0033]
[图8a] 图8a是表示在实施例中作为靶标基因的水稻e3泛素蛋白连接酶gw2基因(osgw2基因)的核苷酸序列的图。图中,“grna3”表示grna3的识别位点。
[0034]
[图8b] 图8b是图8a的续图,表示在实施例中作为靶标基因的水稻e3泛素蛋白连接酶gw2基因(osgw2基因)的核苷酸序列。
[0035]
[图9] 图9是表示在实施例中确认到基因组编辑(基因缺失)的grna3所识别的区域的核苷酸序列的图。发生了基因组编辑(基因缺失)的位点用
“‑”
表示。另外,样品表示处理植物细胞时的条件,读数表示实施了核苷酸序列分析的序列数和确认到基因组编辑(基因缺失)的序列数,效率表示基因组编辑效率。下划线表示显著确认到基因组编辑(基因缺失)的条件。
[0036]
[图10] 图10是表示在实施例中确认到基因组编辑(基因缺失)的grna3所识别的区域的核苷酸序列的图。发生了基因组编辑(基因缺失)的位点用
“‑”
表示。另外,样品表示处理植物细胞时的条件,读数表示实施了核苷酸序列分析的序列数和确认到基因组编辑(基因缺失)的序列数,效率表示基因组编辑效率。下划线表示显著确认到基因组编辑(基因缺失)的条件。
[0037]
[图11] 图11是表示在实施例中确认到基因组编辑(基因缺失)的grna3所识别的区域的核苷酸序列的图。发生了基因组编辑(基因缺失)的位点用
“‑”
表示。另外,样品表示处理植物细胞时的条件,读数表示实施了核苷酸序列分析的序列数和确认到基因组编辑(基因缺失)的序列数,效率表示基因组编辑效率。下划线表示显著确认到基因组编辑(基因缺失)的条件。
[0038]
[图12] 图12是表示在实施例中确认到基因组编辑(基因缺失)的grna3所识别的区域的核苷酸序列的图。发生了基因组编辑(基因缺失)的位点用
“‑”
表示。另外,样品表示处理植物细胞时的条件,读数表示实施了核苷酸序列分析的序列数和确认到基因组编辑(基因缺失)的序列数,效率表示基因组编辑效率。下划线表示显著确认到基因组编辑(基因缺失)的条件。
[0039]
[图13] 图13是表示在实施例中确认到基因组编辑(基因缺失)的grna3所识别的区域的核苷酸序列的图。发生了基因组编辑(基因缺失)的位点用
“‑”
表示。另外,样品表示处理植物细胞时的条件,读数表示实施了核苷酸序列分析的序列数和确认到基因组编辑(基因缺失)的序列数,效率表示基因组编辑效率。下划线表示显著确认到基因组编辑(基因缺失)的条件。
具体实施方式
[0040]
本发明在1个方案中提供复合物,该复合物包含基因组编辑酶和cpp,其中cpp与基因组编辑酶融合。关于cpp的融合在后面进行说明。该方案的复合物中的基因组编辑酶的典型例子是talen或zfn。
[0041]
本发明在另一方案中提供复合物,该复合物包含基因组编辑酶、靶标基因特异性核酸和cpp,其中cpp与基因组编辑酶和/或靶标基因特异性核酸融合。
[0042]
cpp的融合只要不妨碍向植物细胞内引入复合物和基因组编辑,则可以是任何方式。融合例如可以是通过肽键等共价键进行的融合,也可以是通过静电结合、范德华力等非共价键进行的融合。在cpp与基因组编辑酶进行共价结合的情况下,共价键可以是任一种形式,典型的是肽键。
[0043]
基因组编辑酶与cpp可处于任一种位置关系。cpp可与基因组编辑酶的n末端融合,也可与基因组编辑酶的c末端融合,还可与基因组编辑酶的n末端和c末端的双方融合,或者可与基因组编辑酶的除n末端和c末端以外的氨基酸残基融合。优选cpp与基因组编辑酶的n
末端或c末端融合。基因组编辑酶与cpp的融合可通过接头进行。各种接头均已知且可使用。优选的接头不会妨碍向植物细胞内引入本发明的复合物和基因组编辑。在通过肽键进行的融合的情况下,作为接头的例子,可列举:由1个~几个甘氨酸残基构成的肽等,但并不限于这些。1个基因组编辑酶上可融合1个cpp,也可融合2个以上的cpp。
[0044]
可使cpp与靶标基因特异性核酸融合。融合可在核酸的3’末端进行,也可在5’末端进行,还可在除这些以外的部分、例如在核酸的糖部分和/或碱基部分进行。优选融合在核酸的3’末端进行。融合可采用例如有机合成等已知方法。1个核酸上可融合1个cpp,也可融合2个以上的cpp。
[0045]
各种cpp均已知。用于本发明的cpp只要可将本发明的复合物直接引入到植物细胞内、且不妨碍基因组编辑即可,可以是任一种肽。作为本发明中可使用的cpp的例子,可列举:富碱性氨基酸(例如精氨酸、赖氨酸、组氨酸)的肽、或者聚组氨酸等,但并不限于这些。
[0046]
作为本发明中可使用的cpp的其他例子,可列举:长度为几个氨基酸以上、且构成氨基酸的半数以上为组氨酸的肽。这样的肽具有优异的细胞膜穿透性。
[0047]
对本发明中可使用的cpp的长度没有特别限定,典型的是几个氨基酸以上,例如几个氨基酸~数十个氨基酸。例如可以是6个氨基酸~40个氨基酸、7个氨基酸~30个氨基酸、8个氨基酸~20个氨基酸,例如可以是6个氨基酸、7个氨基酸、8个氨基酸、9个氨基酸、10个氨基酸、11个氨基酸、12个氨基酸、13个氨基酸、14个氨基酸、15个氨基酸、16个氨基酸、17个氨基酸、18个氨基酸、19个氨基酸、20个氨基酸、21个氨基酸、22个氨基酸、23个氨基酸、24个氨基酸、25个氨基酸、26个氨基酸、27个氨基酸、28个氨基酸、29个氨基酸、30个氨基酸,或者可以是30个氨基酸以上。在本说明书中,几个氨基酸是指2个、3个、4个、5个、6个、7个、8个或9个氨基酸。在本说明书中,数十个氨基酸是指10~100个范围的任意的氨基酸数。
[0048]
作为本发明中可使用的cpp的优选例子,可列举:构成氨基酸残基的约80%以上为组氨酸残基、更优选构成氨基酸残基的约90%以上为组氨酸残基、进一步优选所有的构成氨基酸残基均为组氨酸残基的肽(聚组氨酸)。聚组氨酸的长度与以上说明的cpp的长度相同。
[0049]
构成本发明中可使用的cpp的除组氨酸以外的氨基酸残基可以是任一种氨基酸残基。优选构成本发明中可使用的cpp的除组氨酸以外的氨基酸残基为精氨酸、赖氨酸等碱性氨基酸残基、或者具有与组氨酸类似的特性的氨基酸残基。构成本发明中可使用的cpp的氨基酸残基可以是天然氨基酸残基、非天然氨基酸残基、修饰氨基酸残基、或合成氨基酸残基。氨基酸的合成或修饰可由本领域技术人员适当进行。
[0050]
本发明中可使用的cpp可通过fmoc固相合成法等肽合成法或基因重组法等已知方法来调制。
[0051]
通过使用细胞膜穿透性高的肽(以上说明的肽),可提高本发明的复合物向植物细胞内的引入效率,其结果,基因组编辑效率提高。
[0052]
用于本发明的基因组编辑酶可以是任一种基因组编辑酶,没有特别限定。基因组编辑酶已知有各种酶。作为可用于本发明的基因组编辑酶的例子,可列举:cas9、cas12、cas13、casφ、tid等cas家族的核酸酶、talen、zfn等核酸酶、活化诱导胞苷脱氨酶(aid)或target-g等脱氨酶等,但并不限于这些。在本说明书中,基因组编辑酶包含野生型和突变型。突变型基因组编辑酶包含自然突变体和人工突变体两者。突变型基因组编辑酶可以是与原始的酶相比编辑效率增大的酶、效率减少的酶或效率消失的酶。突变型基因组编辑酶
的制造方法已知,可列举:基因重组、肽化学合成、化学修饰等,但并不限于这些方法。另外,基因组编辑酶可以是切割单链dna的酶,也可以是切割双链dna的酶。
[0053]
在本发明的特别的具体例子中,基因组编辑酶为cas9。在本发明的其他的特别具体例子中,基因组编辑酶是通过功能改变而调节了切割活性的与cas9结合的aid。
[0054]
在基因组编辑酶为talen或zfn的情况下,与靶标基因一并设计这些蛋白。这种设计可通过本领域技术人员所已知的方法进行。
[0055]
靶标基因特异性核酸是可在靶标基因中的想要发生突变的位点配置基因组编辑酶的核酸。作为靶标基因特异性核酸的典型例子,可列举:指导rna (grna),但并不限于此。靶标基因特异性核酸可考虑靶标基因的核苷酸序列,利用已知方法进行设计、制作。可使用crisprdirect、crispr-p2.0、geneious、ape等已知软件设计grna。优选靶标基因特异性核酸对基因组编辑酶呈特异性,通过与基因组编辑酶进行孵育而与基因组编辑酶形成复合物。作为这样的核酸与基因组编辑酶的例子,可列举:grna与cas9,但并不限于这些。
[0056]
通过本发明进行基因组编辑的植物包含所有种类的植物。植物包含种子植物以及蕨类植物和苔藓植物。种子植物包括被子植物和裸子植物。被子植物包含双子叶类和单子叶类。双子叶类包含合瓣花类和离瓣花类。作为合瓣花类的例子,可列举:菊科、杜鹃花科、唇形科、茄科、旋花科、胡麻科、报春花科、桔梗科等的植物,但并不限于这些。作为离瓣花类的例子,可列举:十字花科、蔷薇科、山茶科、石竹科、马齿苋科、杨梅科、葫芦科、芸香科、伞形科、豆科、樟科等的植物,但并不限于这些。作为单子叶类的例子,可列举:鸢尾科、禾本科、灯心草科、天南星科、姜科、鸭跖草科、凤梨科、芭蕉科、百合科、兰科等的植物,但并不限于这些。作为裸子植物的例子,可示例:柏科、松科、杉树科、红豆杉科、银杏科、苏铁科等的植物,但并不限于这些。作为蕨类植物的例子,可列举:紫萁、欧洲蕨、狼尾蕨、粗茎鳞毛蕨、问荆、木贼等,但并不限于这些。作为苔藓植物的例子,可列举:地钱、桧叶金发藓、光藓、泥炭藓等,但并不限于这些。
[0057]
可利用本发明将基因组编辑酶引入到各种食用植物、园艺植物、观赏用植物、建材用树木、林荫树或防风林用树木等中,进行基因组编辑。作为基因组编辑的应用例,可列举:品种改良或遗传学研究等,但并不限于这些。
[0058]
本发明的复合物不仅用于植物的基因组编辑,还可用于动物、丝状菌、酵母、细菌和放线菌等微生物、以及藻类的基因组编辑。
[0059]
上述复合物可利用化学合成法或基因重组法等已知方法来制造。例如,利用使用编码基因组编辑酶的dna和编码cpp的dna的融合物的基因重组法得到基因组编辑酶与cpp的融合物,孵育该融合物和靶标基因特异性核酸,从而可得到上述复合物。孵育通常是在水溶液中、室温~约37℃下进行。水溶液可以是缓冲液。根据需要,可利用柱层析等已知方法纯化复合物。
[0060]
作为通过非共价键融合有cpp的本发明的复合物的例子,可列举:聚阳离子部分与cpp融合、且聚阳离子部分与靶标基因特异性核酸进行静电结合的复合物。在上述复合物中,基因组编辑盒与cpp通过聚阳离子部分进行融合。
[0061]
聚阳离子部分是在生理学条件下具有带2个以上的正电荷的基团的部分,其与靶标基因特异性核酸进行静电结合。生理学条件下例如可以是植物细胞可存活或增殖的ph条件下或植物细胞中的ph条件下。
[0062]
静电结合是指在生理学条件下带负电荷的核酸与带正电荷的聚阳离子部分通过静电引力进行结合。
[0063]
聚阳离子部分与cpp的融合只要不妨碍向植物细胞内引入本发明的复合物和基因组编辑,则可以是任何方式。融合例如可通过共价键、静电结合、基于范德华力的键来进行。典型的是,聚阳离子部分与cpp的融合通过共价键来进行。共价键的典型例子为肽键。聚阳离子部分与cpp可处于任一种位置关系。聚阳离子部分可与cpp的n末端结合,也可与cpp的c末端结合,还可与cpp的n末端和c末端的双方结合,或者可与cpp的除n末端和c末端以外的氨基酸残基结合。聚阳离子部分与cpp的结合可通过接头进行。已知有各种接头且均可使用。优选接头不会妨碍向植物细胞内引入复合物和基因组编辑。在通过肽键进行的融合的情况下,作为接头的例子,可列举:由1个~几个甘氨酸残基构成的肽等,但并不限于这些。1个cpp上可融合1个聚阳离子部分,也可融合2个以上的聚阳离子部分。另外,在1个聚阳离子部分上可融合1个cpp,也可融合2个以上的cpp。
[0064]
聚阳离子部分只要不妨碍向植物细胞内引入本发明的复合物和基因组编辑,则可以是任一种。作为聚阳离子部分的例子,可列举:在生理学条件下带正电荷的肽(优选聚阳离子肽)、寡糖、阳离子性聚合物等,但并不限于这些。肽和寡糖可以是野生型,也可以是突变型或修饰物。突变型肽和突变型寡糖、以及它们的修饰物具有与原始的肽和寡糖同等或更高的与靶标基因特异性核酸的静电结合能力。阳离子性聚合物可以来自天然,也可化学合成。
[0065]
聚阳离子肽是在生理学条件下带有正电荷的具有2个以上的氨基酸残基的肽,这样的肽已知。作为聚阳离子肽的例子,可列举富碱性氨基酸(例如赖氨酸、精氨酸、组氨酸)的肽,但并不限于这些。聚阳离子肽的长度只要不妨碍向植物细胞内引入复合物和基因组编辑,则没有特别限定,典型的是几个氨基酸~数十个氨基酸,例如可以是6个氨基酸~40个氨基酸,可以是8个氨基酸、9个氨基酸、10个氨基酸、11个氨基酸、12个氨基酸、13个氨基酸、14个氨基酸、15个氨基酸、16个氨基酸、17个氨基酸、18个氨基酸、19个氨基酸、20个氨基酸、或20个氨基酸以上。作为聚阳离子肽的例子,可列举:由赖氨酸和/或精氨酸残基构成的肽等。作为由赖氨酸和/或精氨酸残基构成的肽的具体例子,可列举:k8、k9、k10、k11、k12、r8、r9、r10、r11、r12等。作为聚阳离子肽的其他具体例子,可列举:由几个kh重复序列构成的肽。聚阳离子肽并不限于以上例子。构成本发明中可使用的聚阳离子肽的氨基酸残基可以是天然氨基酸残基、非天然氨基酸残基、修饰氨基酸残基或合成氨基酸残基。氨基酸的合成或修饰可由本领域技术人员适当进行。
[0066]
作为带正电荷的寡糖的例子,可列举:葡糖胺、果糖胺、半乳糖胺、甘露糖胺等己糖胺的聚合物,例如壳聚糖等,但并不限于这些。带正电荷的寡糖的糖残基数只要不妨碍向植物细胞内引入本发明的复合物和基因组编辑,则没有特别限定。
[0067]
作为阳离子性聚合物的例子,可列举:聚乙烯亚胺、聚丙烯亚胺、聚(β-氨酯)、聚乳酸/聚乙醇酸、2-羟丙基甲基丙烯酰胺等,但并不限于这些。阳离子性聚合物的长度只要不妨碍向植物细胞内引入本发明的复合物和基因组编辑,则没有特别限定。
[0068]
聚阳离子肽、带正电荷的寡糖和阳离子性聚合物可利用本领域技术人员已知的方法进行制造或从天然提取。
[0069]
上述复合物可利用已知的方法来制造。例如,可通过孵育(i)孵育基因组编辑酶和
靶标基因特异性核酸而得到的复合物(基因组编辑盒)和(ii)聚阳离子部分与cpp的融合物以利用核酸的负电荷和聚阳离子部分的正电荷进行静电结合,来制作上述复合物。孵育通常是在水溶液中、室温~约37℃下进行。水溶液可以是缓冲液。聚阳离子部分与cpp的融合物可通过已知方法、例如fmoc法等肽合成法或基因重组法等来制作。根据需要,可利用柱层析等已知方法纯化复合物。
[0070]
作为聚阳离子肽与cpp的融合物的具体例子,可列举:k10(g)h8、k10(g)h12、k10(g)h16、k10(g)h20、r10(g)h20等,但并不限于这些。需要说明的是,括弧文字表示可存在、也可不存在甘氨酸残基。
[0071]
本发明的复合物可进一步包含信号序列。信号序列也称为信号肽。通过在本发明的复合物中包含信号序列,可使本发明的复合物局部存在于所期望的细胞内区域。各种信号序列均已知,作为信号序列的例子,可列举:核移位信号序列(nls)、线粒体移位信号序列(mls)、叶绿体移位信号序列(cls)等,但并不限于这些。可根据细胞内的所期望的局部存在位置选择信号序列,与本发明的复合物融合。可使用还包含信号序列的本发明的复合物,在细胞内的所期望的位置进行基因组编辑。例如,使用包含核移位信号的本发明的复合物,无需基因重组,即可在核内进行基因组编辑。使用包含线粒体移位信号序列的本发明的复合物,无需基因重组,即可在线粒体内进行基因组编辑。使用包含叶绿体移位信号序列的本发明的复合物,无需基因重组,即可在线粒体内进行基因组编辑。
[0072]
信号序列可与基因组编辑酶、靶标基因特异性核酸、亚结构域(后述)等本发明的复合物的任何部分融合。信号序列的融合方式只要使复合物局部存在于细胞内的所期望的位置,而且不妨碍向植物细胞内引入复合物和基因组编辑,则可以是任何方式。融合例如可通过肽键等共价键进行,也可通过静电结合、范德华力等非共价键进行。典型的是,信号序列通过共价键与复合物融合。在信号序列与基因组编辑酶进行共价结合的情况下,共价键可以是任何形式,典型的是肽键。
[0073]
基因组编辑酶与信号序列可处于任一种位置关系。信号序列可与基因组编辑酶的n末端融合,也可与基因组编辑酶的c末端融合,还可与基因组编辑酶的n末端和c末端的双方融合,或者与基因组编辑酶的除n末端和c末端以外的氨基酸残基融合。或者信号序列可插入至基因组编辑酶的氨基酸序列内。优选信号序列与基因组编辑酶的n末端或c末端融合。更优选信号序列与基因组编辑酶的n末端融合。基因组编辑酶与信号序列的融合可通过接头进行。各种接头均已知且可使用。优选的接头使复合物局部存在于细胞内的所期望的位置,且不妨碍向植物细胞内引入复合物和基因组编辑。在通过肽键进行的融合的情况下,作为接头的例子,可列举:由1个~几个甘氨酸残基构成的肽等,但并不限于这些。1个基因组编辑酶上可融合1个信号序列,也可融合2个以上的信号序列。
[0074]
可使信号序列与亚结构域融合。融合可在亚结构域的n末端进行,也可在c末端进行,或者可在除n末端和c末端以外的氨基酸残基上进行。融合例如可采用基因重组或有机合成等已知方法。信号序列与亚结构域的融合可通过接头进行。在1个亚结构域上可融合1个信号序列,也可融合2个以上的信号序列。
[0075]
可使信号序列与靶标基因特异性核酸融合。融合可在核酸的3’末端进行,也可在5’末端进行,还可在除这些以外的部分、例如核酸的糖部分和/或碱基部分进行。优选融合在核酸的3’末端进行。融合可采用例如有机合成等已知方法。信号序列与核酸的融合可通
过接头进行。在1个核酸上可融合1个信号序列,也可融合2个以上的信号序列。
[0076]
本发明的复合物可进一步包含亚结构域。在本说明书中,亚结构域是指具有功能的蛋白。通过使用这样的本发明的复合物,可进行所期望的各种类型的基因组编辑。对亚结构域的种类没有特别限定,例如可列举:碱基替换酶、dna甲基化酶、dna脱甲基化酶、转录活化酶、转录抑制酶等。本领域技术人员可选择适当的亚结构域以用于本发明的复合物。例如,使用包含碱基替换酶作为亚结构域的本发明的复合物,无需基因重组,即可进行基因组的碱基替换。使用包含dna甲基化酶作为亚结构域的本发明的复合物,无需基因重组,即可进行基因组的甲基化。使用包含dna脱甲基化酶作为亚结构域的本发明的复合物,无需基因重组,即可进行基因组的脱甲基化。使用包含转录活化酶作为亚结构域的本发明的复合物,无需基因重组,即可进行基因组的转录活化。使用包含转录抑制酶作为亚结构域的本发明的复合物,无需基因重组,即可进行基因组的转录抑制。
[0077]
信号序列和亚结构域可均包含在本发明的复合物中。通过使用这样的本发明的复合物,可在所期望的细胞内区域内进行所期望的类型的基因组编辑。例如,使用包含核移位信号序列和碱基替换酶的本发明的复合物,无需基因重组,即可进行核基因组的碱基替换。通过使用包含核移位信号序列和dna甲基化酶的本发明的复合物,无需基因重组,即可进行核基因组的甲基化。使用包含核移位信号序列和dna脱甲基化酶的本发明的复合物,无需基因重组,即可进行核基因组的脱甲基化。使用包含核移位信号和转录活化酶的本发明的复合物,无需基因重组,即可进行核基因组的转录活化。使用包含线粒体移位信号序列和碱基替换酶的本发明的复合物,无需基因重组,即可进行线粒体基因组的碱基替换。使用包含叶绿体移位信号序列和转录抑制酶的本发明的复合物,无需基因重组,即可进行叶绿体基因组的转录抑制。
[0078]
亚结构域可与本发明的复合物的任何部分融合。典型的是,亚结构域与基因组编辑酶融合。亚结构域可与基因组编辑酶的n末端融合,也可与基因组编辑酶的c末端融合,还可与基因组编辑酶的n末端和c末端的双方融合,或者可与基因组编辑酶的除n末端和c末端以外的氨基酸残基融合。优选亚结构域与基因组编辑酶的n末端或c末端融合。
[0079]
基因组编辑酶与亚结构域的融合方式只要不妨碍亚结构域的功能,而且不妨碍向植物细胞内引入复合物和基因组编辑,则可以是任何方式。融合例如可通过肽键等共价键进行,也可通过静电结合、范德华力等非共价键进行。典型的是,信号序列通过共价键与复合物融合。在信号序列与基因组编辑酶进行共价结合的情况下,共价键可以是任何形式,典型的是肽键。
[0080]
基因组编辑酶与亚结构域的融合可通过接头进行。各种接头均已知且可使用。优选的接头不妨碍亚结构域的功能,不妨碍向植物细胞内引入复合物和基因组编辑。在通过肽键进行的融合的情况下,作为接头的例子,可列举:由1个~几个甘氨酸残基构成的肽等,但并不限于这些。1个基因组编辑酶上可融合1个亚结构域,也可融合2个以上的亚结构域。
[0081]
cpp可与亚结构域融合。关于亚结构域与cpp的融合方式、融合部分,适用关于基因组编辑酶与cpp的融合的说明。
[0082]
在包含亚结构域的本发明的复合物中,在cpp与亚结构域融合、亚结构域与基因组编辑酶融合的情况下,可理解为cpp经由亚结构域与基因组编辑酶融合。因此,在本说明书中,“cpp与基因组编辑酶融合”包括如上所述的情况。
[0083]
上述说明虽然是关于将本发明的复合物引入到植物细胞内,但由于本发明的复合物对于所有的生物种类的细胞穿透性均高,因此不仅可引入到植物内,还可引入到动物、丝状菌、细菌、放线菌、酵母、藻类等细胞内,可用于广泛的生物种类的基因组编辑。尤其是本发明的复合物对于具有细胞壁的植物细胞、藻类细胞、丝状菌细胞和酵母细胞的穿透性高,因此适合于这些生物种类的基因组编辑。
[0084]
因此,本发明在又一方案中提供基因组编辑方法,该方法包括:将本发明的复合物引入到细胞内。可通过在培养基中孵育本发明的复合物和细胞,而将本发明的复合物引入到细胞内。向细胞内引入本发明的复合物的方法、培养基的种类、孵育的条件可根据细胞的种类由本领域技术人员适当选择、变更。在该方案的基因组编辑方法中,典型的是,细胞为植物细胞。
[0085]
在进行植物的基因组编辑的情况下,可将本发明的复合物引入到所有形态的植物细胞、所有的植物组织内。例如,可将本发明的复合物引入到植物的叶、茎、茎尖、冬芽、根、种子、孢子、花粉、培养细胞等中。在本说明书中,有时将植物的叶、茎、茎尖、冬芽、根、种子、孢子、花粉、培养细胞等统称为植物细胞。
[0086]
在本发明的基因组编辑方法中,可引入1种以上的靶标基因特异性核酸。另外也可引入1种以上的基因组编辑酶。即,在基因组编辑中可使用1种本发明的复合物,也可使用2种以上的本发明的复合物。
[0087]
本发明在又一方案中提供基因组编辑用试剂盒,其包含本发明的复合物或其构成成分。作为本发明的复合物的构成成分,可示例:cpp、聚阳离子部分、基因组编辑酶与cpp的融合物、聚阳离子部分与cpp的融合物等。在该试剂盒中可组合本发明的复合物的构成成分而得到本发明的复合物。通常,该试剂盒附有操作说明书。可使用本发明的试剂盒进行基因组编辑的生物种类如上,没有特别限定。本发明的试剂盒适合用于具有细胞壁的植物、藻类、丝状菌和酵母的基因组编辑。
[0088]
以下,参照图1对使用本发明的复合物的基因组编辑方法(重组蛋白法/肽法)和现有的基因组编辑方法进行比较说明。以下的说明是具体地说明本发明以容易理解,并不限定本发明的范围。
[0089]
现有的基因组编辑技术存在如下所述的问题:由于依赖于基因重组技术,所以只可进行可适用基因重组的植物种类的基因组编辑;由于基因组编辑后的植物具有外源基因,所以基因组编辑后的植物被视为基因重组生物,商业利用非常困难;外源基因的去除仅可通过重复交配进行,基因组编辑盒的去除需要大量时间。
[0090]
本发明涉及:通过使cpp (例如h8~h20肽)与基因组编辑盒(例如基因组编辑酶cas9和grna)融合而得到的复合物、以及通过将该复合物直接引入到植物细胞内而进行的植物的基因组编辑方法等。cpp的融合可通过共价键或非共价键(例如静电结合)进行。融合方法的具体例子有:调制将基因组编辑酶cas9和cpp以基因工程学方式融合而成的重组蛋白并进行利用的方法(重组蛋白法);以及使cpp与基因组编辑盒进行静电结合的方法(肽法)。可利用这些方法调制本发明的复合物。
[0091]
根据重组蛋白法,例如可使用大肠杆菌表达系统调制cas9-cpp重组融合蛋白(例
如cas9与h8~h20的cpp的融合蛋白),与用于切割靶标区域的grna一同孵育以形成复合物,将如此操作而得到的复合物引入到植物细胞内进行基因组编辑。
[0092]
根据肽法,例如有以下方法:孵育基因组编辑酶cas9和grna以形成复合物,使聚阳离子部分与cpp的融合物(例如k10、h8~h20与cpp的融合物)与上述复合物的核酸进行静电结合(利用grna的碱基的负电荷和聚阳离子部分的赖氨酸残基的正电荷)。可将如此操作而得到的复合物引入到植物细胞内进行基因组编辑。
[0093]
两种方法均不依赖于基因重组,而将基因组编辑酶盒直接引入到植物细胞内,因此可完全克服现有的基因组编辑技术的诸多问题,而且还可简便地且在短时间内获得基因组编辑细胞。
[0094]
本说明书中的术语只要没有特别说明,则解释为在生物学、生化学、化学、药学、医学等领域通常所理解的意义。
[0095]
本说明书中的数值可包含其数值
±
5%、
±
10%或
±
20%的范围的数值。
[0096]
本说明书中的氨基酸的标记采用已知的单字母法或三字母法。在本说明书中表示肽的情况下,在以单字母法表示的氨基酸的右边带有数字。例如h20是指由20个组氨酸残基构成的肽。k10是指由10个赖氨酸残基构成的肽。k10h20是指在由10个赖氨酸残基构成的肽的c末端结合有由20个组氨酸残基构成的肽的n末端而得到的肽。k10gh20是指从n末端向c末端结合有由10个赖氨酸残基构成的肽、1个甘氨酸残基和由20个组氨酸残基构成的肽而得到的肽。在本说明书中,肽可包含除肽键以外的键。只要没有特别说明,则肽中的氨基酸残基间的键为肽键。
[0097]
本说明书中,用连字符(-)表示基因组编辑酶与cpp的融合物。例如,cas9-h20是指使cpp(h20)与基因组编辑酶cas9的c末端结合而得到的融合物。只要没有特别说明,则基因组编辑酶与cpp的键为肽键。
[0098]
以下,给出实施例,以进一步详细且具体地说明本发明,但实施例并不限定本发明的范围。
实施例
[0099]
1) 供试细胞作为表达cas9和cpp融合cas9的大肠杆菌,使用bl21(de3)株。作为植物细胞,使用作为木本植物的日本柳杉(cryptomeria japonica)的愈伤组织和来自愈伤组织的细胞,以及作为草本植物的水稻(oryza sativa)的培养细胞。在日本柳杉愈伤组织的传代中,使用1/2md琼脂培养基,以1周间隔进行传代。在来自日本柳杉愈伤组织的细胞的悬浮和试验中使用1/2md液体培养基,在遮光、25℃、120rpm下振荡培养。在水稻培养细胞的传代中,使用ms液体培养基,以1周间隔进行传代。在水稻培养细胞的悬浮和试验中使用ms液体培养基,在遮光、27℃、120rpm下振荡培养。
[0100]
2) cpp融合cas9的表达调制融合有作为细胞膜穿透肽(cpp)的h8、h12、h16和h20肽的cas9-h8、cas9-h12、cas9-h16和cas9-h20作为重组蛋白。表达质粒使用pet24b,宿主大肠杆菌使用bl21(de3)株。重组蛋白在20℃、18小时的条件下在菌体内表达,用co (钴)离子固定化树脂(ge healthcare公司制造)纯化菌体破碎液。
[0101]
3) cas9的表达调制融合有不具有细胞膜穿透能力的肽标签即flag标签(dykddddk)以代替cpp的cas9作为重组蛋白。表达质粒使用pet24b,宿主大肠杆菌使用bl21(de3)株。重组蛋白在20℃、18小时的条件下在菌体内表达,用抗flag抗体固定化树脂(mbl公司制造)来纯化菌体破碎液。
[0102]
4) grna+cpp融合cas9复合物的调制(用于重组蛋白法的复合物的调制)将溶于sec缓冲液(20mm的hepes-koh、500mm的kcl、ph7.5)的cpp融合cas9 (cas9-h8、cas9-h12、cas9-h16、cas9-h20) (20μm)和溶于duplex缓冲液(30mm的hepes-koh、100mm的乙酸钾、ph7.5)的grna (20μm)等量混合,在室温下孵育15分钟,从而调制了grna+cpp融合cas9复合物(10μm)。利用同样的方法,调制了grna+cas9复合物(10μm)。使用3种grna (grna1、grna2、grna3) (以下的实验也相同)。这些grna以日本柳杉镁螯合酶基因(其核苷酸序列见seq id no: 1):cjchli基因的特定位点(图2中,表示为grna1、grna2的位点)和水稻e3泛素蛋白连接酶gw2基因(其核苷酸序列见seq id no: 2):osgw2基因的特定位点(图8中,表示为grna3的位点)为靶标。grna1位点和grna2位点分别以seq id no: 1的第56~78位、第1094~1116位的碱基表示。grna3位点以seq id no: 2的第1796~1818位的碱基表示。grna1、grna2使用已知的软件(ape)来设计,grna3使用已知的软件(crisprdirect和crispr-p2.0)来设计。
[0103]
5) grna+cas9+cpp复合物的调制(用于肽法的复合物的调制)将溶于sec缓冲液(20mm的hepes-koh、500mm的kcl、ph7.5)的cas9 (20μm)和溶于duplex缓冲液(30mm的hepes-koh、100mm的乙酸钾、ph7.5)的grna (20μm)等量混合,在室温下孵育15分钟,从而调制了grna+cas9复合物(10μm)。然后,相对于grna+cas9复合物,混合等量的溶于duplex缓冲液(30mm的hepes-koh、100mm的乙酸钾、ph7.5)的k10g-cpp (k10gh8、k10h12、k10h16、k10h20肽) (20、200、2000μm),在室温下孵育60分钟,从而调制了grna+cas9+cpp复合物。使用的grna为上述grna1、grna2、grna3。
[0104]
6) 基因组编辑试验将传代1周后的360μl日本柳杉细胞(在1/2md液体培养基中为20mg/ml)或360μl水稻细胞(在ms液体培养基中为20mg/ml)分别注入到聚苯乙烯制的5ml falcon圆底管中。相对于该360μl日本柳杉细胞或360μl水稻细胞,混合40μl上述调制的grna+cpp融合cas9复合物或grna+cas9+cpp复合物。之后,在遮光、25℃、120rpm下振荡培养24~72小时。在上述实验条件下,确认到荧光修饰后的grna+cpp融合cas9复合物或grna+cas9+cpp复合物被摄入到日本柳杉细胞/水稻细胞内。
[0105]
通过离心(500g
×
10分钟、4℃)回收培养后的日本柳杉细胞或水稻细胞,用1/2md液体培养基洗涤多次。使用植物细胞用基因组dna提取试剂盒(dnas-ici!-p:rizo inc.公司制造),从日本柳杉细胞或水稻细胞中提取基因组dna,通过pcr扩增基因组编辑的靶标基因区域。通过将该pcr产物供于扩增子序列分析,评价基因组编辑的有无。另外,克隆pcr产物,通过供于sanger序列分析,评价基因组编辑的有无。
[0106]
7) 结果在用重组蛋白法(grna+cpp融合cas9复合物)处理的日本柳杉细胞或用肽法(grna+cas9+cpp复合物)处理的日本柳杉细胞中,确认到在单独的grna或grna+cas9复合物处理
细胞中没有检测到的基因组编辑(基因缺失)。另一方面,在用重组蛋白法(grna+cpp融合cas9复合物)处理的水稻细胞中,确认到在grna+cas9复合物处理细胞内没有检测到的基因组编辑(基因缺失)。
[0107]
使用与靶标基因(cjchli基因)对应的2种grna (grna1或grna2)的结果,对于重组蛋白法(grna+cpp融合cas9复合物)确认到4种基因组编辑(基因缺失) (图3、图4、图7)。另一方面,对于肽法(grna+cas9+cpp复合物)确认到6种基因组编辑(基因缺失) (图3、图4、图5、图6、图7)。另外,对于重组蛋白法(grna+cpp融合cas9复合物),在使用4种cpp融合cas9 (cas9-h8、cas9-h12、cas9-h16、cas9-h20)的情况下,确认到基因组编辑(基因缺失)。对于肽法(grna+cas9+cpp复合物),在使用4种k10g-cpp (k10gh8、k10gh12、k10gh16、k10gh20肽)的情况下,确认到基因组编辑(基因缺失)。
[0108]
使用与靶标基因(osgw2基因)对应的grna3的结果,对于重组蛋白法(grna+cpp融合cas9复合物)确认到5种基因组编辑(基因缺失) (图8、图9、图10、图11、图12)。对于重组蛋白法(grna+cpp融合cas9复合物),在使用3种cpp融合cas9 (cas9-h8、cas9-h16、cas9-h20)的情况下,确认到基因组编辑(基因缺失)。
[0109]
确认到基因组编辑(基因缺失)的区域与grna的识别区域一致,由此确认到:在靶标区域可诱导基因组编辑。
[0110]
由这些结果证实了使用本发明的复合物的基因组编辑的效果。即,通过利用本发明的复合物,证实了向植物细胞内引入了难以直接引入的基因组编辑酶和诱导基因组编辑。
[0111]
通过利用本发明,实现不依赖于基因重组的农作物的品种改良。通过本发明得到的农作物不属于基因重组生物,因此商品价值高。特别是,本发明的实施例中使用的日本柳杉是每一代的时间非常长的农作物,在利用现有的依赖于基因重组的基因组编辑技术的情况下,通过交配去除外源基因(基因组编辑酶cas9基因)需要数十年单位的时间,因此品种改良不现实。认为本发明即使在这样的世代时间长的农作物的基因组编辑(利用了基因组编辑的分子育种,例如品种改良)中也发挥巨大的威力。
[0112]
工业实用性本发明可用于农业、林业、食品、药品,以及植物的研究、育种、品种改良等领域。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1