催化紫杉烷类化合物中C9位羟基化的生物酶、核酸分子、生物材料及其应用
本技术涉及植物代谢生物学与合成生物学,具体涉及催化紫杉烷类化合物中c9位羟基化的生物酶、核酸分子、生物材料及其应用。
背景技术:
1、紫杉烷类是从植物中分离得到的抗肿瘤活性成分,并对已取得的活性成分的化合物进行结构修饰,合成的一系列衍生物,是重要的一线抗肿瘤药物。紫杉烷类药物主要包括紫杉醇(式iii所示化合物)、多西他赛、卡巴他赛,以及具有紫杉烷骨架结构的衍生物。当前,获取紫杉醇类原料药的主要生产方式为半合成法,即首先通过提取天然前体巴卡亭ⅲ(baccatinⅲ)和10-去乙酰巴卡亭ⅲ(10-deacetylbaccatinⅲ,10-dab),然后化学合成紫杉醇,但该方法所用的前体物质仍依赖于从红豆杉的树皮或树叶细胞中提取,受制于红豆杉稀缺资源,无法完全解决供应问题。
2、
3、巴卡亭ⅲ和10-去乙酰巴卡亭ⅲ目前只能依赖于从稀缺红豆杉中提取的原因在于其合成通路中仍有几个关键酶未能解析,其中之一便是催化c9位氧化反应的酶。由于巴卡亭ⅲ含有7个单氧化位点,氧化顺序未知、氧化底物未知,给催化c9位氧化反应的酶(即c9羟化酶,t9αh)的筛选带来极大的困难,导致该酶在过去三十年中都没能鉴定得到。
4、基于上述现状,急需全面解析紫杉醇合成的关键酶,尤其是解析天然前体物质巴卡亭ⅲ和10-去乙酰巴卡亭ⅲ合成的关键酶,如c9羟化酶,继而采用合成生物学手段和代谢工程策略,在植物或者微生物底盘中重构紫杉醇合成通路,以期大量生产紫杉醇,彻底解决紫杉醇供不应求的难题。
技术实现思路
1、有鉴于此,本发明提供了催化紫杉烷类化合物中c9位羟基化的生物酶、核酸分子、生物材料及其应用。该生物酶(又称c9羟化酶,t9αh)的成功解析,有助于实现紫杉醇的全生物合成,提高巴卡亭iii生物合成的产量,解决紫杉醇供不应求的难题。
2、为了实现上述发明目的,本发明提供以下技术方案:
3、第一方面,本发明提供了一种生物酶t9αh,其氨基酸序列包括如下序列中的至少一种:
4、a1)seq id no:1~14所示氨基酸序列中的至少一种;
5、a2)seq id no:1~14所示氨基酸序列中的至少一种的n端和/或c端连接标签得到的融合氨基酸序列;
6、a3)seq id no:1~14所示氨基酸序列中的至少一种经过一个或多个氨基酸残基的取代和/或缺失和/或添加得到的具有相同功能的氨基酸序列;
7、a4)与seq id no:1~14所示氨基酸序列中的至少一种具有至少60%同一性且具有相同功能的氨基酸序列。
8、任选的,a4)所述的氨基酸序列来源于红豆杉属,包括但不限于欧洲红豆杉taxusbaccata l.、短叶红豆杉taxus brevifolia nutt.、灰岩红豆杉taxus calcicolal.m.gao&mich.加拿大红豆杉taxus canadensis marshall、中国红豆杉taxuschinensis(pilg.)rehd.、密叶红豆杉taxus contorta griff.、东北红豆杉taxuscuspidata siebold
9、&zucc.、佛罗里达红豆杉taxus floridana nutt.ex chapm.、高山红豆杉taxusflorinii spjut、中美红豆杉taxus globosa schltdl.、南方红豆杉taxus mairei(lemée&lév.)s.y.hu、phyton氏红豆杉taxus phytonii spjut、苏门达腊红豆杉taxus sumatrana(miq.)de laub.、喜马拉雅红豆杉taxus wallichiana zucc.、黄山红豆杉taxushuangshan type、峨眉红豆杉taxus emei type、秦岭红豆杉taxus qinling type.、云南红豆杉taxus yunnanensis w.c.cheng&l.k.fu、曼地亚红豆杉taxus×media rehder、欧美红豆杉taxus×hunnewelliana rehder。
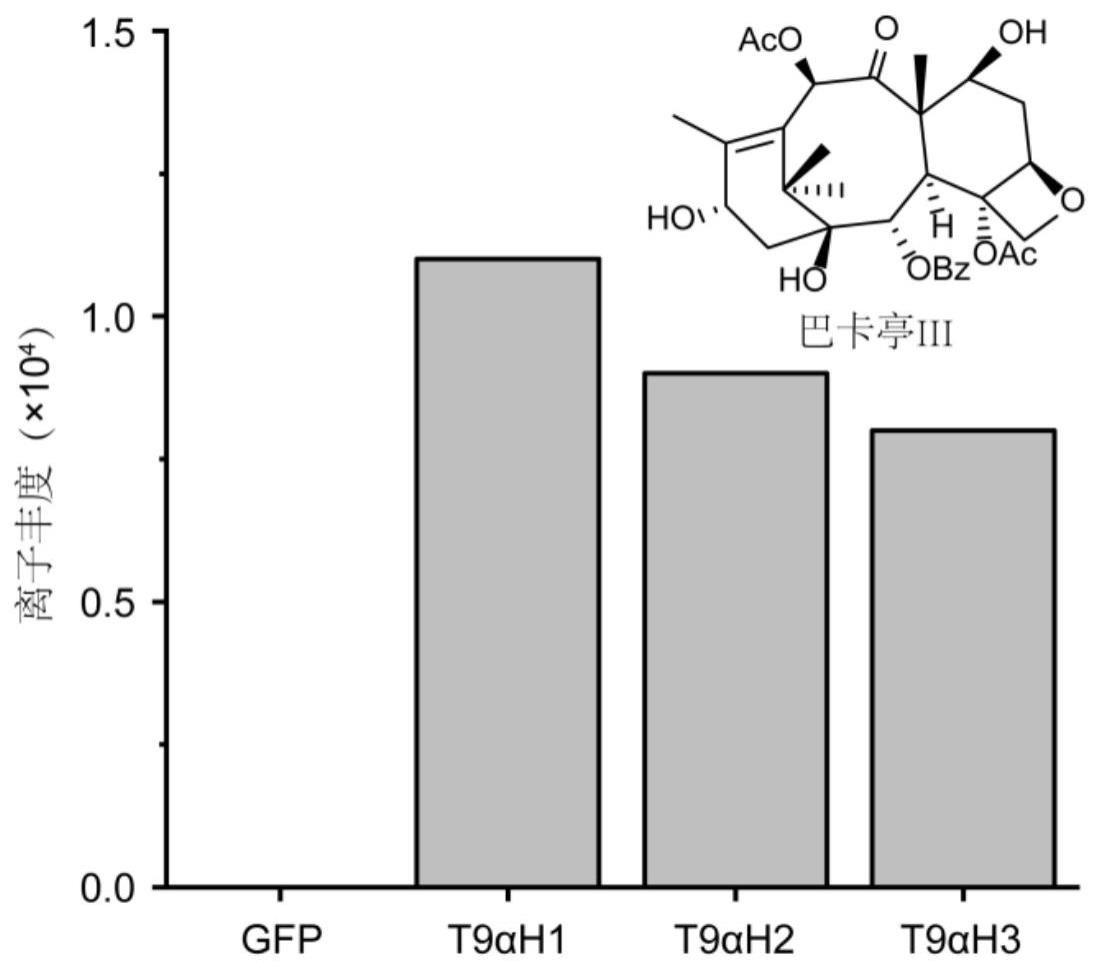
10、本技术创造性地从红豆杉中鉴定得到一种具有催化紫杉烷类化合物合成过程中c9位的羟基化反应的生物酶,本技术将其命名为t9αh。功能分析发现,t9αh可以催化具有紫杉醇碳骨架结构、c9位未羟基化的化合物中c9位发生羟基化,从而打通了巴卡亭iii或其前体物质等紫杉烷类中间体的生物合成路径,可进一步实现紫杉醇等紫杉烷类的全生物合成。
11、在本发明具体实施例中,合成紫杉素时,该酶能够催化紫杉二烯-5α,10β,13α-三醇(taxadiene-5α,10β,13α-triol)转化为紫杉二烯-5α,9α,10β,13α-四醇(taxadiene-5α,9α,10β,13α-tetraol),实现c9位的羟基化反应。
12、在本发明另一具体实施例中,合成巴卡亭iii(紫杉醇前体物质)时,该酶能够催化具有紫杉醇碳骨架结构、c9位未羟基化的化合物中c9位发生羟基化,成功合成了巴卡亭iii。
13、进一步地,本发明获得了t9αh的同源蛋白和突变体,证明同源蛋白和突变体同样能够催化c9位的羟基化。
14、在本发明实施方式中,a3)中的氨基酸序列为a1)中所示的氨基酸序列中添加、缺失、替换一个或多个(例如可以为1个、2个、3个、4个、5个、6个、7个、8个、9个、10个或更多个)氨基酸残基且具有相同功能的氨基酸序列。
15、在本发明实施方式中,a4)中的氨基酸序列为与a1)中所示的氨基酸序列具有至少60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或者99%同一性且具有相同功能的氨基酸序列。
16、在本发明的具体实施方式中,上述14条t9αh的氨基酸序列的序列相似度为63.15%-100%,保守域/位点为保守的psrf基序(氨基酸423-426)、高度保守的结合血红素的pfg元件(氨基酸439-441)、etmr盐桥(氨基酸369-372)、exxr基序以及关键的半胱氨酸(氨基酸447);突变体的关键氨基酸位点包括l75、g128、p129、f132、v136、v142、l228、t231、l232、f305、h308、a309、d312、t313、s316、p376、a377、f378、g379、s380、f381、r382、m402、f485、p486、p487。上述氨基酸是以单字母缩写形式表示的,氨基酸以及英文名、英文缩写见表4。
17、在本发明的具体实施方式中,上述生物酶为分离的生物酶。
18、第二方面,本发明提供了编码上述生物酶的核酸分子,其核苷酸序列包括如下序列中的至少一种:
19、b1)seq id no:15~28所示核苷酸序列中的至少一种;
20、b2)seq id no:15~28所示核苷酸序列中的至少一种的互补序列、简并序列或同源序列,同源序列为与seq id no:15~28所示核苷酸序列具有至少75%同一性的核苷酸序列;
21、b3)在严格条件下与b1)或b2)所示的核苷酸序列杂交,且能够编码相同功能蛋白质的核苷酸序列。
22、在本发明的实施方式中,互补序列为按照碱基互补配对原则形成的互补序列,互补序列可以为与b1)所示核苷酸序列具有相同功能的不完全互补序列或完全互补序列。
23、在本发明的实施方式中,简并序列是指改变b1)所示核苷酸序列一个或多个核苷酸后,改变的核苷酸序列位置对应编码的氨基酸种类不变,不会影响核酸分子的功能和表达水平。
24、在本发明的实施方式中,同源序列为与b1)中所示的核苷酸序列具有至少75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或者99%同一性且具有相同功能的核苷酸序列。例如突变基因、等位基因或衍生物。
25、任选的,b2)所述核酸分子的核苷酸序列来源于红豆杉属,包括但不限于欧洲红豆杉taxus baccata l.、短叶红豆杉taxus brevifolia nutt.、灰岩红豆杉taxus calcicolal.m.gao&mich.加拿大红豆杉taxus canadensis marshall、中国红豆杉taxuschinensis(pilg.)rehd.、密叶红豆杉taxus contorta griff.、东北红豆杉taxuscuspidata siebold&zucc.、佛罗里达红豆杉taxus floridana nutt.ex chapm.、高山红豆杉taxus florinii spjut、中美红豆杉taxus globosa schltdl.、南方红豆杉taxusmairei(lemée&lév.)s.y.hu、phyton氏红豆杉taxus phytonii spjut、苏门达腊红豆杉taxus sumatrana(miq.)de laub.、喜马拉雅红豆杉taxus wallichiana zucc.、黄山红豆杉taxus huangshan type、峨眉红豆杉taxus emei type、秦岭红豆杉taxus qinlingtype.、云南红豆杉taxus yunnanensis w.c.cheng&l.k.fu、曼地亚红豆杉taxus×mediarehder、欧美红豆杉taxus×hunnewelliana rehder。
26、任选的,所述核酸分子为合成的和/或分离的核酸分子。
27、示例性地,“严格条件”是指探针将与其靶序列杂交至可探测程度超过与其它序列杂交(如至少2倍于背景)的条件。严格条件具有序列依赖性,且因环境的不同而不同。通过控制杂交和/或洗涤条件的严格性,可以鉴定与探针100%互补的靶序列。可选择地,可以调节严格条件以允许一些序列错配,使得探测到较低程度的相似性。
28、在本发明的具体实施方式中,编码t9αh的核苷酸序列的序列相似度为75.48%-100%。
29、第三方面,本发明提供了一种生物材料,该生物材料为下述c1)至c3)中的任一种:
30、c1)含有上述核酸分子的表达盒;
31、c2)含有上述核酸分子或c1)所述表达盒的载体;
32、c3)含有上述核酸分子、c1)所述表达盒、c2)所述载体中的至少一种的宿主细胞。
33、任选的,宿主细胞包括微生物细胞、植物细胞、动物细胞、藻类细胞中的至少一种。
34、在本发明的实施方式中,c1)所述表达盒进一步包含一个可操作地与上述核酸分子相连的启动子。
35、在本发明的实施方式中,c2)的载体中,对于载体的种类不作具体限定,可以根据需要选择适合的载体。例如载体包括但不限于pfastbac1、pyes2、pyes2.1、pesc-ura、pesc-trp、pesc-leu、pesc-his、pgex2t、ptaex3、pusa、pymb0、pht43、pet28b、pij702、pucp19、peaq-ht、pymb03或pht43,优选peaq-ht。
36、在本发明的实施方式中,c3)的宿主细胞中,微生物细胞包括但不限于链霉菌、假单孢菌、芽孢杆菌、酵母细胞、大肠杆菌中的至少一种;植物细胞包括但不限于烟草细胞、青蒿细胞、拟南芥细胞、小立碗藓细胞、地钱细胞、番茄细胞、人参细胞、棉花细胞、甘蔗细胞、马铃薯细胞、玉米细胞、小麦细胞、水稻细胞、萝卜细胞、莴苣细胞中的至少一种;动物细胞包括但不限于昆虫细胞、哺乳动物细胞、果蝇细胞、线虫细胞、鱼类细胞中的至少一种;藻类细胞包括但不限于蓝藻细胞、绿藻细胞中的至少一种。
37、本发明还提供了一种工程菌株,该工程菌株包含上述核酸分子、表达盒、载体中的至少一种。在本发明的实施方式中,该工程菌株可为酵母工程菌株、大肠杆菌工程菌株。
38、在本发明的实施方式中,植物细胞和/或动物细胞包括非生物性转换(non-biologically transformed)的重组植物细胞和/或重组动物细胞。
39、任选的,植物细胞或动物细胞可通过细胞培养方式表达生物酶、生产目标产物。
40、在上述生物材料中,在体内和体外将核酸分子、表达盒、载体转化宿主细胞的方法包括但不限于电穿孔、聚乙二醇(peg)转化、脂质转染、热休克、磷酸钙沉淀、病毒介导、显微注射。
41、第四方面,本发明提供了一种用于产生宿主细胞的方法,该方法包括用上述核酸分子、生物材料中的表达盒或载体中的至少一种转化宿主细胞。
42、第五方面,本发明提供了一种用于产生植物或植物细胞的方法,该方法包括用上述核酸分子、生物材料中的表达盒或载体中的至少一种转化植物或植物细胞。
43、第六方面,本发明提供了一种产生上述生物酶的方法,包括:将上述核酸分子,或生物材料中的表达盒、载体中的至少一种,转化宿主细胞,使宿主细胞产生所述生物酶。
44、第七方面,本发明提供了上述生物酶、核酸分子或生物材料中的任一种在如下d1)、d2)、d3)和/或d4)中的应用:
45、d1)在催化紫杉烷类化合物和/或其中间体的前体物质中的c9位的羟基化;紫杉烷类化合物和/或其中间体的前体物质为c9位未发生羟基化和氧化的前体物质;
46、d2)在合成紫杉素和/或其中间体中的应用;
47、d3)在合成巴卡亭iii和/或其中间体中的应用;
48、d4)在合成紫杉烷类化合物和/或其中间体中的应用。
49、在本发明实施方式中,紫杉烷类化合物包括但不限于紫杉醇及其衍生物、紫杉素及其衍生物、多西他赛及其衍生物、卡巴他赛及其衍生物中的至少一种。
50、在本发明实施方式中,紫杉烷类化合物中间体包括但不限于紫杉二烯-六醇-六乙酸酯及其衍生物、1-去羟基巴卡亭iv及其衍生物、巴卡亭ⅲ及其衍生物、10-去乙酰巴卡亭ⅲ及其衍生物、紫杉醇及其衍生物、巴卡亭iv及其衍生物、巴卡亭vi及其衍生物、巴卡亭vii及其衍生物、巴卡亭ix及其衍生物、巴卡亭v及其衍生物、紫杉素及其衍生物、巴卡亭i及其衍生物、紫杉二烯-2α,5α,7β,9β,10β,13α-六醇-5α-乙酸酯及其衍生物、5α-乙酸酯-taxuspine f及其衍生物、紫杉二烯-六醇-四乙酸酯及其衍生物、taxuspine f及其衍生物中的至少一种。
51、在本发明实施方式中,所述应用包括以下几个方面:(1)本发明所提供的生物酶的氨基酸序列或至少部分序列的多肽可能在去除或替代某些氨基酸之后仍有生物活性甚至有新的生物学活性,或者提高了产量或优化了蛋白动力学特征或其他致力于得到的性质;(2)本发明所提供的核酸分子的核苷酸序列或至少部分核苷酸序列被修饰或突变,修饰或突变的途径包括密码子优化、插入、缺失,聚合酶链式反应(pcr),易错pcr,不同序列的重新连接,序列的不同部分或与其他来源的同源序列进行定向进化,或通过化学试剂诱变等。(3)本发明所提供的核苷酸序列或至少部分核苷酸序列的克隆基因通过合适的表达体系在外源宿主中表达以得到相应的酶或其他更高的生物活性或产量。(4)在受体宿主细胞中过表达上述多核苷酸或蛋白质,得到目的宿主细胞,与受体宿主相比,所述目的宿主的紫杉醇和/或其中间体的产量增加。过表达受体宿主中上述多核苷酸或蛋白质的方式包括启动子编辑技术、密码子优化、利用强启动子、插入内含子、利用病毒载体、融合蛋白技术中的一种或多种,以及可实现过表达目的的其他方式。(5)本发明所提供的核苷酸序列或至少部分核苷酸序列的基因或基因簇可以通过遗传重组来构建重组质粒以获得新型生物合成途径,也可以通过插入、置换、缺失或失活进而获得新型生物合成途径。
52、第八方面,本发明提供了一种在体内或体外合成紫杉烷类化合物和/或其中间体的方法,以紫杉烷类化合物和/或其中间体的前体物质中的至少一种为底物,在上述生物酶的催化作用下,c9位发生羟基化;紫杉烷类化合物和/或其中间体的前体物质为c9位未发生羟基化和氧化的前体物质。
53、在本发明实施方式中,紫杉烷类化合物和/或其中间体的前体物质包括如式i和/或式ii所示的化合物:
54、
55、其中,r1、r3、r4各自独立地选自h、-oh、-oac;r2、r6各自独立地选自h、-oh、-oac、-obz;r5、r7各自独立地选自h、-oh、-oac、=o。
56、在本发明具体实施方式中,紫杉烷类化合物和/或其中间体的前体物质包括但不限于taxa-4(5),11(12)-二烯、taxa-4(20),11(12)-二烯-5α-ol、紫杉二烯-5α,10β,13α-三醇和/或其衍生物中的至少一种。
57、在本发明实施方式中,合成紫杉烷类化合物和/或其中间体的生物酶还包括txs、tat、t5αh、t13αh、t2αh、t7βh、tot、tbt、t10βh、t14βh、dbat、bapt、t2’oh、dbtnbt、pam、coaligase、tax9、tax14、tax19中的至少一种。
58、其中,本发明的生物酶(t9αh)、txs、tat、t5αh、t13αh、t2αh、t7βh、tot、tbt为从头开始合成巴卡亭iii所需的生物酶;dbat、bapt、t2’oh、dbtnbt、pam、coa ligase这几种生物酶为从巴卡亭iii开始合成紫杉醇等紫杉烷类化合物所需的生物酶。合成其它紫杉烷类的酶包括t10βh、t14βh、tax9、tax14、tax19。
59、在本发明实施方式中,体内包括微生物细胞内、藻类细胞内、植物细胞内和/或动物细胞内。
60、在本发明实施方式中,在体内合成紫杉烷类化合物和/或其中间体,包括使上述核酸分子在微生物细胞内、藻类细胞内、植物细胞内和/或动物细胞内表达本发明生物酶;以紫杉烷类化合物和/或其中间体的前体物质为底物,在上述生物酶的催化作用下,合成紫杉烷类化合物和/或其中间体。任选的,所述底物由微生物细胞、藻类细胞内、植物细胞和/或动物细胞生产。任选的,所述底物可人为额外添加。
61、在本发明实施方式中,在体内合成含氧杂环化合物,包括使上述核酸分子在微生物细胞内、藻类细胞内、植物细胞内和/或动物细胞内表达本发明生物酶;以紫杉烷类化合物和/或其中间体的前体物质为底物,在上述生物酶的催化作用下,合成紫杉烷类化合物和/或其中间体。任选的,所述底物由微生物细胞、藻类细胞内、植物细胞和/或动物细胞生产。任选的,所述底物可人为额外添加。
62、第九方面,本发明提供了一种植物或其植物部分,植物为如下植物之一:
63、e1)上述植物细胞生长形成的植物;
64、e2)上述方法生产获得的植物;
65、e3)所述e1)-e2)中任一植物自交所形成的后代,以及后代生长形成的植物;
66、e4)所述e1)-e2)中任一植物与其它品种杂交所形成的后代,以及后代生长形成的植物;
67、上述的植物部分为根、茎、叶、花、果实、花粉或种子。
68、与现有技术相比,本发明具有的有益效果为:
69、本技术创造性地从红豆杉中鉴定得到了t9αh,该t9αh可以催化紫杉烷类化合物合成过程中c9位的羟基化反应。例如,合成紫杉醇时,该酶能够催化紫杉二烯-5α,10β,13α-三醇(taxadiene-5α,10β,13α-triol)转化为紫杉二烯-5α,9α,10β,13α-四醇(taxadiene-5α,9α,10β,13α-tetraol),实现c9位的羟基化反应;
70、进一步地,本技术将上述t9αh基因,与其他已鉴定的催化酶基因在植物细胞(例如烟草等)或微生物细胞(例如大肠杆菌、酿酒酵母等)等模式生物体系中共同表达,最终能够在植物细胞或微生物细胞中从头合成前体物质巴卡亭iii;
71、进一步地,本技术填补了数十年来关于紫杉醇等紫杉烷类化合物生物合成的最后几步酶促反应,最终解析了紫杉醇等紫杉烷类化合物的完整生物合成通路。这不仅为生产紫杉醇或其他紫杉烷类化合物提供了一个新的工业生产途径,还可以通过对通路酶的改造,利用低价值的前体物质来高效地获得大量的紫杉醇等紫杉烷类,这不仅满足绿色环保的要求,还能够极大地降低紫杉醇相关药物的价格,让更多人的人能够用得起相关的药物,从而降低病患的家庭负担。
- 还没有人留言评论。精彩留言会获得点赞!