包含蒽环类衍生物的结合蛋白药物缀合物的制作方法
本发明涉及包含蒽环类毒素衍生物的结合蛋白药物缀合物。
背景技术:
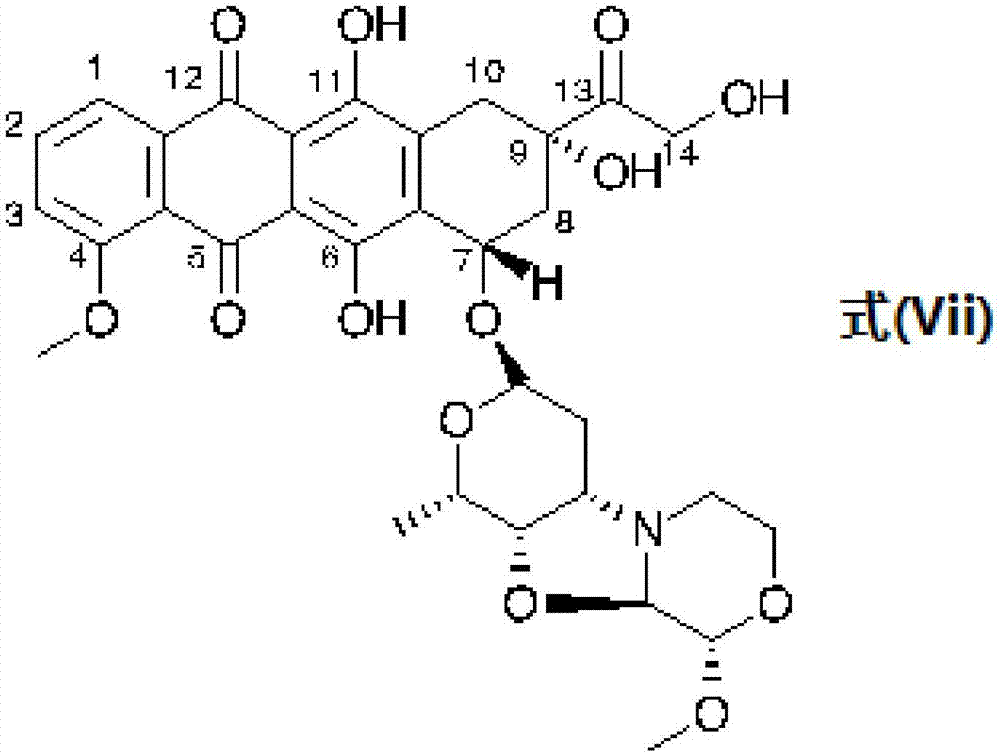
:小分子量毒素(mw优选<2'500道尔顿)与蛋白质(特别是特异于肿瘤细胞的抗体)的共价缀合物是特异性地靶向癌细胞对其进行破坏的有力工具。因此,此类结合蛋白药物缀合物(bpdc),特别是抗体药物缀合物(adc),对于用于癌症的治疗而言有着高度的医疗和商业兴趣。为了开发有效且安全的bpdc或adc以用于癌症治疗,需要解决若干方面:首先,所述结合蛋白或抗体需要特异于几乎不会(或理想情况下不会)由正常或健康组织细胞表达的给定的肿瘤特异性抗原(tsa)。其次,药物和结合蛋白之间的共价键(或连接)需要具有在环化中足够稳定的功能性,防止毒素载荷在血流中不期望的释放,但是其在结合至和/或内化至癌细胞中时必需要有效释放药物。第三,毒素载荷必需要具有足够高的毒性或效力,以便有效破坏癌细胞,即便是假如可能只有有限量的tsa在癌细胞上得到表达因而只有有限量的adc得以内化、或者说假如在结合至癌细胞时(或内化至癌细胞内时)毒素载荷的释放效率不够高。经tsa而成功靶向癌症的第一个方面依赖于对于靶向生物学和开发用于其特异性结合的靶向分子的深入理解,而与最佳接头以及毒素载荷相关的第二和第三方面,则一般适用于结合蛋白药物缀合物(bpdc)或抗体药物缀合物(adc)的有效性。所有目前用于临床试验的adc,以及经fda许可用于癌症治疗的两种adc(来自takeda的抗-cd30adc(本妥昔单抗(brentuximab)-vedotin)、和来自roche/genentech的抗-her-2adc(曲妥珠单抗(trastuzumab)-emtansine,或t-dm1)(参见perez等人2014)),是通过将包括马来酰亚胺接头化学的毒性载荷化学缀合至抗体赖氨酸残基的伯氨基、或游离硫醇基(通过抗体链内二硫键桥的温和还原而产生的)而产生的。所述化学缀合具有两种局限:首先,已经发现基于化学马来酰亚胺的接头在人血清白蛋白的存在下与不期望的不稳定性相关联,并因而导致在用含马来酰亚胺-接头的adc治疗的患者血液循环中毒素的释放(参见alley等人,2008)。第二,通过马来酰亚胺接头化学的典型化学缀合会导致异质性的bpdc或adc,因为不能控制与哪个氨基或硫醇基发生缀合。因此,会获得每个抗体共价结合药物数目的高斯分布,从而缀合的adc具有范围在3.5至4之间的平均药物抗体比(dar)。然而,个别缀合物可以不具有附着至抗体的药物(dar=0)、或者在半胱氨酸缀合物的情形具有达8个缀合至抗体的药物(dar=8),甚至是在赖氨酸缀合物的情形具有每个抗体更多药物(>dar10)。因此典型的化学缀合的adc代表了展现出不同功能特性的不同分子的异质性混合物(参见panowski等人,2014),这在开发用于治疗癌症患者的adc时的监管角度来看,显然是不期望的。因此,对于提供经位点特异性缀合的(且因而在药物抗体比方面是均质的)adc或bpdc是具有商业和医疗方面的需求的。此外,对于提供具有更稳定的药物抗体连接的adc或bpdc(其在血液循环中比传统的基于马来酰亚胺接头化学更稳定)是具有商业和医疗方面的需求的。另外,对于提供比目前市售的adc或bpdc有更高的效力且更低的副作用的adc或bpdc是具有商业和医疗方面的需求的。本发明的一般特征本发明解决了这些问题。其提供了用于结合蛋白药物缀合物的新的毒素,以及任选外加将这些毒素通过避免典型的马来酰亚胺接头化学而以位点特异性的方式缀合至所述结合蛋白的新技术。这两种特征的一般优势将在下文进行讨论:接头技术上文提到的经化学缀合的且含马来酰亚胺-接头的bpdc或adc的血清不稳定性和异质性,都代表了这些药物在癌症患者中显著的安全性负担,因为这两者都会在患者中增加此类adc的毒素的非特异性释放("药物解离(de-drugging)")。一方面,典型的马来酰亚胺接头可以被人血清中的游离硫醇(特别是人血清白蛋白的半胱氨酸-34,其作为最丰富的血清蛋白在人血清中提供了最高浓度的游离硫醇)所破坏。人血清白蛋白的半胱氨酸-34通过所谓的回归迈克尔反应(retro-michaelreaction)的方式(其将毒素转移并共价结合至人血清白蛋白(hsa))而破坏马来酰亚胺接头的硫醚键。该毒素-hsa缀合物继而可以使毒素在血液循环中或在身体中分布而无任何肿瘤选择性(参见:alley等人,2008)。另一方面,在化学缀合的、异质性的adc中的较高dar的种类已知由于这些adc的较高疏水性以及聚集的倾向而具有较短的血清半衰期。因此这些较高dar的种类在将这些adc结合至靶向阳性的癌细胞之前,会经受更快的血清清除、降解和毒素释放。此外,也已知较高dar的种类会导致更快的药物解离,因为个体缀合位点会具有不同的药物解离动力学(取决于携带毒素的氨基酸的结构背景)。上文所述的化学缀合的adc的负担阻碍了将adc开发至临床的成功,虽然通过将其缀合至肿瘤特异性的抗体而将高度强力的细胞毒素递送至癌细胞这一概念是令人信服的。在2000年经fda许可的首个adc,来自pfizer/wyeth的(吉姆单抗奥佐米星),由于毒素相关的副作用而需要在fda许可10年后退市(http://www.fda.gov/newsevents/newsroom/pressannouncements/ucm216448.htm)。因此,化学缀合的adc的负担将目前对adc的开发努力局限在具有中等细胞毒性的毒素,例如基于微管蛋白聚合抑制性多拉司他汀/澳瑞他汀的以及基于美登素的药物。实际上,目前在临床评估的所有adc中有超过90%携带有与单甲基澳瑞他汀e(mmae)或f(mmaf)或美登素(例如dm1或dm4)相关的毒素(参见:mullard(2013))。然而,微管蛋白聚合抑制性药物不能达到低于纳摩尔范围的效力,因为微管蛋白(细胞骨架的成份)是高度丰富的细胞内蛋白靶标,从而许多药物需要扩散或转运至细胞内,以便将细胞分裂和存活所必须的细胞内细胞骨架的代谢关闭。微管蛋白聚合抑制性药物的中等效力,可以耐受缀合物某种程度上的"药物解离",并且其对于分化中的和有丝分裂的细胞的特异性作用使得具有此种特定作用模式的毒素在adc的开发中最为流行。然而,为了特异性地针对具有低tsa表达水平的肿瘤会需要强力得多的毒素。因此,依然在临床前开发中的更新的adc策略,会涉及具有高细胞毒性和不同作用方式的毒素,特别是损坏dna的毒素,如倍癌霉素(参见:doktor等人(2014)和吡咯并苯并二氮杂(pbd)(参见:hartley&hochhauser(2012))。蒽环类衍生物用于bpdc或adc作为载荷的dna插入毒素高度有兴趣的类别是蒽环类,因为其在癌症治疗中作为化疗药物已经证实的临床验证(参见:minotti(2004))。蒽环类是具有高抗肿瘤活性的红色聚酮化合物,最初衍生自链霉菌(streptomyces)种类。在过去40年间已经描述了许多衍生物,包括常规用于各种实体癌症和血液癌症作为化疗药物的一些衍生物,例如阿霉素(doxorubicin,也称作阿霉素(adriamycin))、道诺霉素、表柔比星、伊达比星、或戊柔比星。甚至有一种使用阿霉素作为毒性载荷的,处于i/ii期临床试验用于多发性骨髓瘤和其它血液癌症的抗-cd74adc,米拉珠单抗-阿霉素(参见:clinicaltrials.gov识别号:nct01101594)。所有基于蒽环类的化疗药物都已知对肿瘤细胞显示有限的效力,因而游离的药物对大部分的肿瘤细胞具有μmol/ml范围的ic50(crooke和prestayko,1981)。虽然有目前正在临床试验评估中的第一个阿霉素-adc的实例,但是使用常规的蒽环类作为adc策略的毒性载荷很可能依然是具有挑战性的。大约十年前,新的蒽环类衍生物,称作pnu-159682,已经被描述为奈莫柔比星的代谢物(参见:quintieri等人(2005)clin.cancerres.11,1608-1617),其最近有报道为展现出在皮摩尔至飞摩尔的范围对体外细胞杀伤极高的效力(对于一个卵巢癌(a2780)和一个乳腺癌(mcf7)细胞系)(wo2012/073217a1,caruso等人)。蒽环类衍生物pnu-159682的结构(如上述现有技术文献中所公开的)已经在图2中公开作为参考,并且具有用于四环糖苷配基结构的反应性碳的正式蒽环类编号体系。基于上文所述的化学缀合adc的局限(关于异质性化学缀合adc中的马来酰亚胺-接头不稳定性以及较高dar的种类的药物解离),预期高度强力的蒽环类毒素,如pnu-159682,在典型的化学缀合情况中将会是有很大问题的(由于在靶向肿瘤细胞之前在血液循环中对毒素的释放)。因此,需要强力的毒素(如例如pnu-159682)、具有给定的药代动力学特性和延长的血清稳定性的同质性adc,以便避免、或最小化患者血液循环中来自过早释放的毒素的副作用。然而,同时也依然需要可以对特征在于低靶向表达的肿瘤细胞有特异性的杀伤。虽然通过传统的马来酰亚胺接头方式而产生的使用pnu-159682作为载荷的adc此前已有公开(wo2009/099741a1,cohen等人),但是在此现有技术文献中并未提供功能数据。在化学缀合的且异质性的含马来酰亚胺-接头缀合物的情况中,以不同的接头和间隔结构连接至抗体的pnu-159682对肿瘤细胞的最早功能数据在现有技术文献wo2010/009124a2(beria等人)中公开,但是却并未提供安全性和药代动力学数据。优选的实施方案根据第一个优选的实施方案,描述了蒽环类(pnu)衍生物缀合物,其含有pnu-159682衍生物(缺乏c14碳和附着的蒽环类特征性的四环糖苷配基结构的羟基)。作为第二个优选实施方案,描述了蒽环类(pnu)衍生物缀合物,其缺乏c13和c14碳(具有蒽环类特征性的四环糖苷配基结构的c13处的羰基官能(function)和c14处的羟基)。在这些实施方案中,所述蒽环类(pnu)衍生物缀合物包含具有以下式(i)或式(ii)的蒽环类pnu-159682的衍生物:所述缀合物在其波状线处包含接头结构,所述接头结构可具有不同的元素x-l1-l2-l3-y,其中l1-l3代表接头,且l1-l3中的两个是任选存在的,并且其中x和y各自进一步代表一或多个任选存在的接头。两种衍生物都与pnu-159682(其是蒽环类奈莫柔比星的代谢物且首次由quintieri等人(2005)公开)有显著不同。c13和c14碳(具有其c13处的羰基官能和c14处的羟基)是pnu-159682必须的结构特征,其不是本文公开的衍生物缀合物的一部分。出乎意料且首次发现的是,证明了不具有碳14和附着的蒽环类特征性的四环糖苷配基结构的羟基的pnu-衍生物展现出细胞毒性,例如,在位点特异性缀合的抗体药物缀合物中。其优选的实施方案在图3a、6a和6b中示出。根据本发明的另一实施方案,提供了结合蛋白药物缀合物(bpdc),其具有以下式:·其中l1-l3代表接头,且l1-l3中的两个必须的,·其中x和y各自代表一或多个任选存在的接头,·其中bp是结合蛋白,以及·其中n是≥1和≤10之间的整数。在此构建物中,若干接头可形成单一链,其将一个毒素缀合至一个结合蛋白,和/或若干接头可以连接若干毒素至一个结合蛋白。同样的,接头可以将同一结合蛋白的两个或更多个亚基缀合至两个或更多个毒素分子。任选存在的接头x可以是任何现有技术中已知的化学接头结构,其已用于adc以使得毒素在内化至癌细胞内时能够特异性释放(参见例如ducry&stump(2010)或mccombs等人(2015)。此类接头的一些实例在现有技术中有描述,仅以示例性的方式提供而并非是要进行限制,显示如下。接头l1、l2和l3在下文讨论。任选存在的接头y可以是具有达20个氨基酸的任意氨基酸链,其使得结合蛋白能够最佳缀合至接头x、l1、l2、l3或其变体(特别是l3)的单一链。另外,提供了接头结构,其使得pnu-衍生物能够位点特异性地缀合至适宜的结合蛋白,例如优选缀合至抗体。所述衍生物因而可用于产生位点特异性缀合的、同质性的结合蛋白药物缀合物,其可用于治疗应用,如抗癌治疗。根据蒽环类(pnu)衍生物另一优选的实施方案,所述接头结构作为l2包含寡聚甘氨酸肽(glyn),其直接或通过另一接头l1的方式偶合至所述蒽环类衍生物,以这样的方式从而寡聚甘氨酸(glyn)肽具有游离的氨基末端,并且其中n是在≥1和≤21之间的整数。在每种情况中,(gly)n(在本文也称作(gly)n-nh2或glyn-区段)是寡聚甘氨酸肽-区段。在一个特别优选的实施方案中,n是≥3和≤10之间的整数,优选是≥3和≤6之间的整数。最优选n=5。如本文中已经公开的那样,本文所公开的蒽环类(pnu)衍生物是pnu-159682的衍生物,其缺乏碳原子13和14或者仅缺乏具有附着的官能团的碳14。对于式(i),一个优选的实施方案是,寡聚甘氨酸肽(gly)n(≥1且≤21,优选n=3或n=5)通过亚烷基二氨基接头(nh2-(ch2)m-nh2,m≥1且≤11,优选m=2)的方式缀合至蒽环类衍生物,其通过第一个酰胺键至碳13的方式缀合至蒽环类衍生物以及通过第二个酰胺键的方式缀合至寡聚甘氨酸肽的羧基末端。用于产生位点特异性缀合的蒽环类(pnu)衍生物缀合物的优选的化合物pnu-eda-gly5在图3a中示出。对于式(ii),优选的实施方案是,寡聚甘氨酸肽(gly)n(≥1且≤21,优选n=3或n=5)直接偶合至pnu衍生物的环a(或蒽环类糖苷配基结构的碳9),从而碳13的羰基代表了甘氨酸肽接头的羧基末端。用于产生位点特异性缀合的蒽环类(pnu)衍生物缀合物的优选的化合物pnu-gly5在图6a中示出。对于式(ii),另一优选的实施方案是,寡聚甘氨酸肽(gly)n(≥1且≤21,优选n=3或n=5)直接缀合至pnu衍生物的环a(或蒽环类糖苷配基结构的碳9),其中有亚烷基胺接头-(ch2)m-nh2,m≥1且≤11,优选m=2),其通过酰胺键的方式缀合至寡聚甘氨酸肽的羧基末端。用于产生位点特异性缀合的pnu-衍生物缀合物的优选的化合物pnu-ea-gly5在图6b中示出。在下文中,根据上文所述的蒽环类衍生物缀合物也称作“pnu-eda-glyn-nh2”、“pnu-glyn-nh2”或“pnu-ea-glyn-nh2”,或者也分别缩写为"pnu-eda-glyn"、"pnu-glyn"、或"pnu-ea-glyn",或者在其优选的实施方案中分别具有5个甘氨酸残基,"pnu-eda-gly5"、"pnu-gly5"、或"pnu-ea-gly5"。本发明还提供了结合蛋白药物缀合物(bpdc),其包含根据上文公开的蒽环类衍生物缀合物,该衍生物还包含通过另外的酰胺键的方式缀合至寡聚甘氨酸肽(glyn)的游离氨基末端的结合蛋白。根据蒽环类(pnu)衍生物或结合蛋白药物缀合物(bpdc)的另一实施方案,寡聚甘氨酸肽(glyn)(命名为l2)通过亚烷基二氨基接头(命名为l1的)方式缀合至式(i)的蒽环类衍生物,该亚烷基二氨基接头通过第一个酰胺键的方式缀合至蒽环类衍生物,而其通过第二个酰胺键的方式缀合至寡聚甘氨酸肽的羧基末端,亚烷基二氨基接头和寡聚甘氨酸肽的所述缀合物具有下式(v),其中波状线表示连接至式(i)的蒽环类衍生物。m是在≥1和≤11之间的整数,并且n是在≥1和≤21之间的整数。优选m是在≥2和≤4之间的整数,最优选m=2(亚乙基二氨基,eda)。使用亚烷基二氨基接头以使得(gly)n接头能附着而用于转肽酶(sortase)缀合,从而偶合可以经过(gly)n肽的c-末端来发生,由此提供最终毒素-接头加合物的游离n-末端,以用于转肽酶缀合。应当理解的是,亚烷基二氨基接头中的任何ch2亚甲基都可以被另一稳定的键取代,例如-o-(醚)、-s-(硫醚)、-nh-(胺)、或任何其它烷基,、杂烷基、芳基或杂芳基基团、或其任意组合,以便实现本发明。根据蒽环类(pnu)衍生物或结合蛋白药物缀合物(bpdc)的另一实施方案,寡聚甘氨酸肽(glyn)直接偶合至式(ii)蒽环类衍生物的环a(或碳9)。其说明参见图6a。根据蒽环类(pnu)衍生物或结合蛋白药物缀合物(bpdc)的另一实施方案,寡聚甘氨酸肽(glyn)通过亚烷基氨基接头(命名为l1)的方式缀合至式(ii)的蒽环类衍生物,该亚烷基氨基接头通过酰胺键的方式缀合至寡聚甘氨酸肽的羧基末端,亚烷基氨基接头和寡聚甘氨酸肽的所述缀合物具有下式(vi)其中波状线表示连接至式(ii)的蒽环类衍生物,其中m是在≥1和≤11之间的整数,且是在≥1和≤11之间的整数。优选m是在≥2和≤4之间的整数,最优选m=2(亚乙基氨基,ea)。使用亚烷基氨基接头以使得(gly)n接头能附着而用于转肽酶缀合,从而偶合可以经过(gly)n肽的c-末端来发生,由此提供最终毒素-接头加合物的游离n-末端,以用于转肽酶缀合。应当理解的是,亚烷基氨基接头中的任何ch2亚甲基都可以被另一稳定的键取代,例如-o-(醚)、-s-(硫醚)、-nh-(胺)、或任何其它烷基、杂烷基、芳基或杂芳基基团、或其任意组合,以便实现本发明。在结合蛋白药物缀合物(bpdc)的另一实施方案中,接头结构l3包含从转肽酶酶识别基序的特异性切割而获得的肽基序。如在本文的别处以及在wo2014140317(其内容援引加入本文)所公开的那样,转肽酶(也称作转肽酶转肽酶)形成了一组通过识别并切割特异性的分选信号(包含特定的肽基序)而修饰表面蛋白的原核生物酶。此肽基序在本文中也称作“转肽酶酶识别基序”、“转肽酶标签”或“转肽酶识别标签”。通常,给定的转肽酶具有一或多个识别的转肽酶酶识别基序。转肽酶可以是天然存在的、或者可以经历过遗传工程化(doerr等人,2014)。在结合蛋白药物缀合物(bpdc)的优选实施方案中,所述转肽酶酶识别基序包含五肽。在结合蛋白药物缀合物(bpdc)的优选实施方案中,所述转肽酶酶识别基序包含以下氨基酸序列中的至少一个(显示为n-末端->c-末端):·lpxtg·lpxsg,和/或·laxtg。前两个转肽酶酶识别基序由野生型金黄色葡萄球菌(staphylococcusaureus)转肽酶a所识别。第二个也由来自金黄色葡萄球菌的工程化的转肽酶a4s9所识别,而第三个由来自金黄色葡萄球菌的工程化的转肽酶a2a-9所识别(doerr等人,2014)。在所有三种情况中,x可以是20个成肽氨基酸中的任意一个。这些转肽酶酶识别基序,例如通过遗传融合而融合至结合蛋白的c-末端,或者其结构域或亚基,并与其共表达。所述融合可直接完成,或者经本文别处描述的另外的接头y来间接完成。值得注意的是,一旦整合至接头结构中并缀合至l2,l3则缺乏转肽酶酶识别基序的第5个氨基酸残基(c-末端的g)。在表1中,所述c-末端的g因而在括号中示出。如果转肽酶酶识别基序是五肽,则l3因此是四肽。在转肽酶缀合之前,转肽酶酶识别基序还可以另外携带其它标签,如his-标签、myc-标签或strep-标签(参见wo2014140317的图4a,其内容援引加入本文),融合至转肽酶酶识别基序的c-末端。然而,由于转肽酶酶识别基序的第4和第5个氨基酸之间的肽键在转肽酶介导的缀合时被切割,这些另外的标签将最终从完全缀合的bpdc中去除。转肽酶酶识别基序可缀合至(gly)n接头,所述(gly)n接头通过本文和wo2014140317中公开的转肽酶技术的方式而缀合至蒽环类衍生物。在缀合过程中,从(gly)n接头释放一个甘氨酸残基。值得提及的是,虽然这三种肽区段以典型的n-末端->c-末端方向在上文示出,l残基是通过肽键的方式融合至结合蛋白c-末端(或融合至接头y的c-末端)的残基。l3的第5个氨基酸残基(g)在缀合至(gly)n肽时被去除,而l3的第4个t或s氨基酸残基是实际上缀合至(gly)n肽的n-末端残基。下表因而给出了本发明结合蛋白-药物缀合物(bpdc)的优选实施方案的概览,其中显示了l1–l3。表1:典型的接头结构如讨论中值得注意的是,一旦整合至接头结构中并缀合至l2,l3则缺乏第5个氨基酸残基(c-末端的g)。在表1中,所述c-末端的g因而显示在括号中。根据结合蛋白药物缀合物(bpdc)的另一实施方案,蒽环类(pnu)衍生物通过一或多个接头的方式缀合至结合蛋白的羧基末端、或缀合至其结构域或亚基的羧基末端。在另一优选的实施方案中,寡聚甘氨酸(glyn)肽接头中的n是在≥3和≤11之间的整数,更优选在≥3和≤7之间,优选n=3、或n=5。最优选寡聚甘氨酸(glyn)肽接头中的n是5。在一个优选的实施方案中,载荷是式(i)之一。在第二个优选的实施方案中,载荷是式(ii)之一。根据结合蛋白药物缀合物(bpdc)的另一实施方案,结合蛋白通过酰胺键的方式缀合至寡聚甘氨酸肽(glyn)的游离氨基末端。根据结合蛋白药物缀合物(bpdc)的另一实施方案,结合蛋白是选自以下组的至少一种:·抗体,·经修饰的抗体形式,·抗体衍生物或保留靶标结合特性的片段,·基于抗体的结合蛋白,·寡聚肽结合蛋白,和/或·抗体模拟物。术语“结合蛋白”,如本文所用,等价于发明人其它出版物中所使用的术语“免疫配体”,包括附录1,其提供了关于转肽酶酶缀合技术的进一步技术细节、公开和实施。"抗体",也同义地称作"免疫球蛋白"(ig),一般包含四个多肽链,两个重链(h)和两个轻链(l),并且因此是多聚蛋白,或其等价的ig同源物(例如,骆驼纳米抗体,其仅包含重链,可衍生自重链或轻链的单结构域抗体(dab));包括其全长功能性突变体、变体或衍生物(包括但不限于鼠源的、嵌合的、人源化的、和完全的人抗体,其保留ig分子的基本表位结合特征,并且包括双重特异性的、双特异性的、多特异性的、和双重可变结构域的免疫球蛋白;免疫球蛋白分子可以是任何类别的(例如,igg、ige、igm、igd、iga和igy)、或亚类的(例如,igg1、igg2、igg3、igg4、iga1和iga2)以及异型(allotype)。倘若结合蛋白是抗体,结合蛋白药物缀合物是抗体药物缀合物(adc)。在下文中,根据本发明的adc也称作“pnu-eda-glyn-ab”、“pnu-glyn-ab”或“pnu-ea-glyn-ab”。"基于抗体的结合蛋白",如本文所用,可以表示含有至少一个抗体衍生的vh、vl、或ch免疫球蛋白结构域的任何蛋白(在其它非免疫球蛋白、或非抗体衍生成分的背景下)。此类基于抗体的蛋白包括但不限于(i)结合蛋白的fc-融合蛋白,包括受体或受体成分(具有全部或部分的免疫球蛋白ch结构域,(ii)结合蛋白,其中vh和或vl结构域偶合至其它的分子骨架,或(iii)分子,其中免疫球蛋白vh,和/或vl,和/或ch结构域以天然存在的抗体或抗体片段中不常见的方式组合和/或组装。"抗体衍生物或片段",如本文所用,涉及分子,其包含至少一个衍生自非全长抗体的多肽链,包括但不限于(i)fab片段,其是由可变轻链(vl)、可变重链(vh)、恒定轻链(cl)和恒定重链1(ch1)结构域组成的单价片段;(ii)f(ab')2片段,其是包含两个fab片段(在铰链区通过二硫桥连接)的二价片段;(iii)fab(fd)片段的重链部分,其由vh和ch1结构域组成;(iv)可变片段(fv)的片段,其由抗体单一臂的vl和vh结构域组成,(v)结构域抗体(dab)的片段,其包含单一可变结构域;(vi)分离的互补决定区(cdr);(vii)单链fv片段(scfv);(viii)双价抗体,其是二价的、双特异性的抗体,其中vh和vl结构域在单一多肽链上表达,但使用很短的接头从而使得同一链上的两个结构域之间不能配对,由此迫使结构域与另一链的互补结构域配对,并创建了两个抗原结合位点;和(ix)线性抗体,其包含一对串联的fv节段(vh-ch1-vh-ch1),其与互补轻链多肽一起,形成一对抗原结合区;和(x)免疫球蛋白重链和/或轻链的其它非全长部分、或其突变体、变体、或衍生物,单独的或以任何组合的形式。在任意情况中,所述衍生物或片段保留靶标结合特性。术语"经修饰的抗体形式",如本文所用,涵盖了抗体-药物-缀合物、多亚烷基氧化物修饰的scfv、单价抗体(monobody)、双价抗体、骆驼抗体、结构域抗体、双特异性的或三特异性的抗体、iga、或通过j链和分泌成分结合的两个igg结构、鲨鱼抗体、新世界灵长类构架区+非新世界灵长类的cdr、铰链区去除的igg4抗体、具有两个改造至ch3结构域的额外结合位点的igg、具有改变的fc区以增强对fc-γ受体亲和性的抗体、包含ch3+vl+vh的二聚化的构建物、以及类似的等。术语"抗体模拟物",如本文所用,是指不属于免疫球蛋白家族的蛋白,并且甚至是非蛋白如适体、或合成聚合物。一些类型具有类抗体的β-折叠结构。"抗体模拟物"或"另外的骨架"相对于抗体的潜在优势是更好的稳定性、更高的组织渗透性、对热和酶更高的稳定性、和相对低廉的生产成本。一些抗体模拟物可以在大的文库中提供,其提供了针对每种可设想靶标的特异性结合候选。与抗体一样,靶标特异性抗体模拟物可以通过使用高通量筛选(hts)技术以及用已建立的展示技术(如噬菌体展示、细菌展示、酵母或哺乳动物展示)来进行开发。目前已开发的抗体模拟物涵盖例如锚蛋白重复序列蛋白(称作darpins)、c-型凝集素、金黄色葡萄球菌的a结构域蛋白、转铁蛋白、脂笼蛋白、纤连蛋白的第10个iii型结构域、kunitz结构域蛋白酶抑制剂、泛素衍生的结合蛋白(称作affilin)、γ晶状体衍生的结合物、半胱氨酸结(knot)或结肽(knottin)、基于硫氧还蛋白a骨架的结合物、sh-3结构域、stradobody、通过二硫键和ca2+稳定的膜受体的“a结构域”、基于ctla4的化合物、fynsh3、和适体(结合至特异性靶标分析的肽分子)。术语“寡聚肽结合蛋白”,如本文所用,涉及具有以高亲和性结合至给定靶标的能力的寡聚肽。术语“寡聚”是指具有5至50个氨基酸残基的肽。根据结合蛋白药物缀合物(bpdc)的另一实施方案,所述结合蛋白结合选自以下组的至少一种实体:·受体,·抗原,·生长因子,·细胞因子,和/或·激素。此列表定义了结合蛋白可以结合的不同类型的靶标。如本文所用,术语"受体"是指细胞表面分子,优选这样的细胞表面分子,其(i)结合特异性的、或特异性的组的信号分子(也即受体,如例如,vegf受体),和/或(ii)具有已知的配体(也即孤儿受体,如例如her2/neu)。天然受体在细胞群体的表面上表达,或者它们仅代表此种分子的细胞外结构域(无论此种形式天然存在与否),或者在血浆中进行天然结合功能的可溶性分子(或在细胞中或器官中)。优选此种受体是参与特定致病过程的信号级联的成员(例如,属于生长因子信号级联的受体),或者是在参与病理学过程的细胞和颗粒(例如癌细胞)的表面表达。如本文所用,术语"抗原"是指有能力诱导特异性的免疫应答的物质,并且可包括表面蛋白或蛋白复合物(例如离子通道)。很多时候,抗原与致病实体(例如癌细胞)相关联。如本文所用,术语"细胞因子”是指由许多细胞分泌的小的细胞信号蛋白分子,并且是在细胞间通讯中广泛使用的信号分子类别。细胞因子可以分类为蛋白、肽、或糖蛋白;术语"细胞因子"涵盖了由遍及身体的多样化胚胎学来源的细胞所产生的大的且多样化的调节剂家族。如本文所用,术语"生长因子”涉及能够刺激细胞生长、增殖和细胞分化的天然存在的物质。通常生长因子是蛋白或类固醇激素。生长因子对于许多细胞过程的调节是重要的。如本文所用,术语"激素”涉及由身体一部分的细胞、腺体、或器官所释放的化合物,其发出讯息影响生物体其它部分的细胞。该术语涵盖肽激素、脂质和磷脂衍生的激素包括类固醇激素、和一元胺。在结合蛋白结合受体或抗原的情况中,结合蛋白药物缀合物(bpdc)可以例如导向载荷(例如毒素)所递送至的特异性的位点,例如,导向病原性实体,例如,癌细胞。因而,毒素或化疗剂的全身毒性得以降低,而毒素或化疗剂在作用位点的局部浓度得以提高,由此提供更好的效率却又降低了副作用。另外,各自的信号级联可以通过结合蛋白的结合来抑制。在载荷是标志物的情况中,该标志物因而可用于标志特异性的位点,例如,通过结合蛋白检测特征为给定表面抗原的癌细胞,以用于诊断。在结合蛋白结合生长因子、细胞因子、和/或激素的情况中,结合蛋白药物缀合物(bpdc)可以例如导向生长因子、细胞因子或激素通常所结合的位点,以便以位点特异性的方式递送载荷。另外,各自的信号级联可以通过结合蛋白的结合来抑制。如本文所用,术语"结合"是指结合蛋白(例如,抗体、或抗体片段、以及其靶标)之间熟知的相互作用或其它非随机的关联。优选此种结合反应特征在于对靶标的高亲和性和/或灵敏性。优选所述结合反应特征在于解离常数(kd)≤10-3m,优选≤10-4m、≤10-5m、≤10-6m、≤10-7m、≤10-8m、≤10-9m、以及最优选≤10-10。根据另一个实施方案,结合蛋白具有至少两个亚基。在此实施方案中,一个亚基可以缀合至本文公开的蒽环类pnu-159682的衍生物(参见图3a和6a和6b)。优选至少两种不同的药物可以位点特异性地缀合至至少两个亚基。此选择提供了多功能的工具箱,其可用来创建大量各种各样不同的结合蛋白-药物构建物。优选所述至少两种不同的药物是干扰不同细胞通路的药物。这意味着,在本文公开的蒽环类衍生物缀合物之后,第二种毒素可以缀合至同一结合蛋白的另一亚基。此种实施方案可以,例如,通过将两种不同的药物分别缀合至全长抗体2个轻链的每一个、以及缀合至全长抗体2个重链来实现,其中通过利用两种不同的转肽酶,识别不同的转肽酶识别基序(“转肽酶标签”),外加在重链和轻链(所述重链和轻链包含用于所述不同转肽酶的各自识别基序)处包含不同c-末端修饰的抗体。以此方式,可以创建抗体药物缀合物,每个包含两个全长ig轻链和ig重链,含有共价附着至所述重链和轻链的不同的载荷。此种实施方案导致优选至少两个亚基的位点特异性缀合,用于产生结合蛋白药物缀合物(具有位点特异性,且相等的载荷缀合至每个所述亚基)。在结合蛋白药物缀合物(bpdc)的一个实施方案中,结合蛋白结合her-2。优选所述结合蛋白是特异于her-2的抗体。在此实施方案中,所述her-2特异性的抗体优选:a)包含曲妥珠单抗的cdr区域1-6(人源化的hu4d5),b)包含曲妥珠单抗的重链可变结构域和轻链可变结构域,c)与a)或b)的区域或结构域具有90%或更高的氨基酸序列相同性,d)是曲妥珠单抗、或者其靶标结合片段或衍生物,和/或e)与曲妥珠单抗竞争结合her-2。抗-her-2单克隆抗体曲妥珠单抗结合her-2的结构域iv。优选所述抗-her-2抗体包含图11a的igh和igl链的一级氨基酸序列(seqidno1和2)。曲妥珠单抗的序列也在drugbank登录号db00072(biod00098,btd00098)中有公开,其援引加入本文,以及在imgt数据库(vh:http://www.imgt.org/3dstructure-db/cgi/details.cgi?pdbcode=7637&part=chain&chain=7637h&vl:http://www.imgt.org/3dstructure-db/cgi/details.cgi?pdbcode=7637&part=chain&chain=7637l)中有公开。在结合蛋白药物缀合物(bpdc)的另一实施方案中,所述结合蛋白结合cd30。优选所述结合蛋白是特异于cd30的抗体。在此实施方案中,所述抗体优选:a)包含本妥昔单抗的cdr区域1-6(嵌合的cac10),b)包含本妥昔单抗的重链可变结构域和轻链可变结构域,c)与a)或b)的区域或结构域具有90%或更高的氨基酸序列相同性,d)是本妥昔单抗、或者其靶标结合片段或衍生物,和/或e)与本妥昔单抗竞争结合cd30。本妥昔单抗(克隆cac10)的序列,其是us2008213289a1中公开的已经获得许可的药物adcetris/本妥昔单抗vedotin的抗体组分。优选所述抗-cd30抗体包含图11的igh和igl链的一级氨基酸序列(seqidno3和4)。优选在这些实施方案中,所述毒素是式(i)之一,·l1是亚乙基二氨基接头,·l2是寡聚甘氨酸(glyn)肽接头(且n是优选的5个氨基酸的长度),和·l3表示经加工的转肽酶标签五肽基序的氨基酸残基1-4(也即缺少c-末端的g残基(第5个氨基酸残基),其在转肽酶介导的缀合至(gly)n肽时被去除,·接头x缺失,和·y是ig轻链的c-末端和l3之间的5个氨基酸的接头,优选具有氨基酸序列ggggs。或者,所述毒素是式(ii)之一,而·l1是亚乙基氨基接头,·l2是寡聚甘氨酸(glyn)肽接头(且n是优选的5个氨基酸的长度),·l3表示经加工的转肽酶标签五肽基序的氨基酸残基1-4(也即缺少c-末端的g残基(第5个氨基酸残基),其在转肽酶介导的缀合至(gly)n肽时被去除,·接头x缺失,和·y是ig轻链的c-末端和l3之间的5个氨基酸的接头,优选具有氨基酸序列ggggs。本发明还提供了用于制备上文所述结合蛋白药物缀合物(bpdc)的方法,其中携带有转肽酶酶识别基序的结合蛋白,通过转肽酶的方式,缀合至至少一种蒽环类衍生物缀合物,其携带有作为l2的寡聚甘氨酸肽(glyn)。所述转肽酶技术,其优点(位点特异性的缀合、毒素和结合蛋白之间经化学计算定义的关系、缀合的高效率)在申请wo2014140317a1(其内容援引加入本文)中有详细解释。对于转肽酶标签的进一步解释可在上文中找到。本发明优选的实施方案是,通过smac技术将pnu-衍生物载荷缀合至结合蛋白的c-末端,且优选缀合至抗体或免疫球蛋白链(到至少一个ig轻链或ig重链)的c-末端。这通过产生用于结合蛋白或免疫球蛋白亚基的哺乳动物细胞表达构建物来实现,所述表达构建物编码紧接在结合蛋白的c-末端之后的用于转肽酶的c-末端五肽识别基序、或多聚结合蛋白(如例如抗体)的多肽亚基。应理解的是,金黄色葡萄球菌的转肽酶a的五肽基序(其是lpxtg或lpxsg并且其此前已经提过)仅仅作为非限制性实例来提供,并且可以由来自其它物种或其它类别的转肽酶(如来自金黄色葡萄球菌的转肽酶b,其识别五肽基序npqtn)所识别的其它五肽基序来取代。也可以使用由工程化的转肽酶所识别的识别基序,如例如laetg(由dorr等人(2014)近来描述的金黄色葡萄球菌转肽酶a的经改造版本所识别)。wo2014140317还提供了关于转肽酶缀合技术的技术细节、公开以及实施,其也称作smac技术(转肽酶介导的抗体缀合技术)。此技术使得两个实体能够缀合,一个以(gly)n区段为标志(如本文上文对于毒素的讨论),且一个以所谓的转肽酶标签(其是可以附着至例如结合蛋白的肽标签)为标志。这些转肽酶标签是寡聚肽,通常是五肽基序,其融合至第一实体(此处:结合蛋白),所述第一实体要缀合至第二实体(此处:蒽环类衍生物),以这样的方式从而使所述转肽酶标签寡聚肽的c-末端保持游离。如wo2014140317中所公开的,这可以通过从表达载体(编码用于五肽转肽酶标签的另外的氨基酸)表达结合蛋白来实现。此种转肽酶标签是例如lpxtg或lpxsg(对于来自金黄色葡萄球菌的转肽酶a)、lpxsg(对于来自金黄色葡萄球菌的工程化的转肽酶a4s9,在dorr等人,2014中有描述)、或laxtg(对于来自金黄色葡萄球菌的经改造的转肽酶a2a9,在dorr等人,2014中有描述),其中x是20个天然存在氨基酸中的任意一个。然而,此类转肽酶标签对于来自其它细菌物种的转肽酶或对于转肽酶类别(如wo2014140317中、以及如现有技术(spirig等人2011)中公开的)可以在序列上有不同。第二实体包含具有游离n末端(-nh2)的甘氨酸-区段(glyn-区段),该glyn-区段是寡聚-甘氨酸肽。优选n是在≥1和≤21之间的整数。在一个特别优选的实施方案中,n是≥3和≤10之间的整数,优选n=3或n=5。最优选n=5。转肽酶继而能够通过转肽反应的方式将两个实体彼此融合,在此期间c-末端氨基酸残基(例如,lpxtg中的g)被切割掉,并继而被所述甘氨酸区段的第一个甘氨酸所取代。在另一个优选的实施方案中,所述五肽识别基序可以直接附加至免疫球蛋白轻链或重链最后的天然存在的c-末端氨基酸,其在人免疫球蛋白κ轻链的情况中是c-末端半胱氨酸残基,其在人免疫球蛋白λ轻链的情况中是c-末端丝氨酸残基,以及其在人免疫球蛋白igg1重链的情况中可以是c-末端赖氨酸残基(由人fcγ1cdna编码)。然而,另一个优选的实施方案也是直接将转肽酶五肽基序附加至倒数第二个c-末端的甘氨酸残基(由人fcγ1cdna编码),因为通常抗体重链的末端赖氨酸残基在哺乳动物细胞中由翻译后修饰所剪去。因此,在超过90%的情况中,天然存在的人igg1缺乏igg1重链的c-末端赖氨酸残基。在另一个优选的实施方案中,所述五肽识别基序可以被附加至人免疫球蛋白igg1重链的c-末端,其中由人fcγ1cdna所编码的c-末端赖氨酸残基被除了赖氨酸之外的氨基酸残基所取代。我们此前已经描述过,在一些情况中(例如在igκ轻链的c-末端,(beerli等人2015),在结合蛋白的c-末端和转肽酶标签l3之间添加额外的氨基酸是有益的。这已经显示出会改善转肽酶对载荷缀合至结合蛋白的酶缀合效率。在igκ轻链的情况中,观察到通过在igκ轻链的最后的c-末端半胱氨酸残基和转肽酶标签之间添加5个氨基酸(ggggs)改善缀合的动力学,从而使igκ轻链和ig重链的c-末端能够以相似的动力学进行缀合(参见:beerli等人(2015)。因此,另一优选的实施方案是在结合蛋白或抗体亚基最后的c-末端氨基酸和转肽酶标签l3之间任选包括≥1和≤21之间个氨基酸的接头y。本发明还提供了上文所述的、或通过上文所述方法产生的结合蛋白药物缀合物用于治疗人类或动物对象的用途,其中所述人类或动物对象:·患有给定的病理状况,·具有发展出给定的病理状况的风险,和/或·诊断为给定的病理状况。本发明还提供了上文所述的结合蛋白药物缀合物用于制备药物的用途,所述药物用于治疗人类或动物对象,其中所述人类或动物对象:·患有给定的病理状况,·具有发展出给定的病理状况的风险,和/或·诊断为给定的病理状况。优选所述病理状况是肿瘤疾病。更优选所述肿瘤疾病是:·具有由ihc或ish确定的,her-2表达评分1+、2+、或3+的癌症,该癌症优选是乳腺癌,·由ihc、elisa、或流式细胞术确定的,cd30阳性的癌症,优选淋巴瘤、更优选霍奇金淋巴瘤(hl)或全身性间变性大细胞淋巴瘤(salcl)。对her-2状况的确定可以例如根据asco/cap指引(其在wolff等人2013中有描述)来进行确定。对cd30状况的确定可以例如根据young2014的方法来进行确定。本发明还提供了药物组合物,其包含上文所述、或由上文所述方法产生的结合蛋白药物缀合物(bpdc),以及至少一种其它的药物可接受的成分。进一步描述为了克服用于产生bpdc和adc的常规马来酰亚胺接头化学的主要局限,我们此前开发了用于产生bpdc或adc的酶方法(使用序列特异性的转肽酶,其中或是用转肽酶、或所谓的剪切内含肽(参见:wo2014140317a1))。具体而言,可以得到证明的是,小分子载荷通过转肽酶进行的位点特异性缀合(在抗体的情况中),指的是smac-技术(转肽酶介导的抗体缀合技术),导致了与化学缀合的adc在体外杀伤癌细胞方面具有同样效力的adc(若使用同样的结合蛋白和同样的载荷)。另外,若使用同样的靶向抗体(抗-her-2曲妥珠单抗)和同样的毒素载荷(dm1),smac-技术产生的特异于her-2靶标的adc在异种移植模型中导致了具有类似效力的肿瘤衰退(wo2014140317a1)。然而,首先,在smac-产生的adc中,未利用马来酰亚胺接头化学,以及第二,缀合反应是以位点特异性的方式对抗体的igh或igl链的c-末端进行的,从而获得了更加同质性的adc。在smac-技术的情况中,位点特异性的缀合可以通过例如金黄色葡萄球菌的重组转肽酶a来实现,该重组转肽酶a特异性地识别lpxtg或lpxsg五肽基序(x=20种天然存在的氨基酸中的任意一种),并且可以附加至要用于缀合的重组抗体。转肽酶a继而使用寡聚甘氨酸-区段作为亲核试剂,来催化转肽作用,通过该作用寡聚甘氨酸的氨基实现了对lpxtg或lpxsg五肽基序的苏氨酸或丝氨酸与甘氨酸之间肽键的亲核攻击。这导致了肽键的断裂和寡聚甘氨酸肽的n-末端甘氨酸之间新肽键的形成(参见图1),也即导致转肽作用。虽然已经显示出,通过转肽酶介导的缀合所产生的曲妥珠单抗-dm1缀合物具有与化学缀合的dm1缀合物(t-dm1、或已经应用于临床)相当的效力,更高效力的smac-技术产生的adc还未能实现(wo2014140317a1)。这是没有预期到的,因为利用了同样的靶向抗体和同样的载荷。基于此以及用不同单克隆抗体(特异性地结合其它tsa,其在癌细胞上可能以比her-2靶标低的水平表达、或者其在adc结合时可能内化效率较低(数据未显示))的其它实验,表现出来的是,需要具有比美登素高的效力、和/或具有潜在的不同作用模式的毒性载荷,来产生显著有效的adc。此外,所述载荷必须要能够修改为在至少一个反应基团的修饰,允许添加寡聚甘氨酸肽,以使得载荷能够转肽酶缀合至lpxtg-或lpxsg-修饰的结合蛋白。最后,如果利用了更高效力的毒素,修饰应导致甘氨酸-区段和载荷之间的稳定连接,以便防止毒素载荷在血液循环中不期望的释放,但同时毒素应当依然导致对癌细胞的有效杀伤(在bpdc或adc特异性结合和内化至肿瘤细胞内时)。对现有技术在smac-技术背景中描述的不同毒性载荷的实证评估,已经导致了这样的发现,具有高度效力的奈莫柔比星的蒽环类衍生物(称作pnu-159682(quintieri等人,2005)(也参见图2),其已经用乙二胺-间隔子进行了修饰,以便允许添加五甘氨酸区段),能够通过smac技术很有效地缀合至lpxtg修饰的抗体,产生几乎完全缀合的adc(基于通过hic(疏水相互作用层析)和反相层析(数据未显示)对产物的分析)。此外,如果此经修饰的pnu-159682衍生物,称作pnu-eda-gly5,经smac缀合至各种单克隆抗体(如下文提供的实施例所示),则实现了对肿瘤细胞的高度效力和tsa-依赖的肿瘤细胞杀伤。具体而言,用smac-技术缀合的pnu-eda-gly5缀合物在体外可以有效地杀伤her-2低表达人乳腺癌细胞,而美登素-毒素缀合物是几乎无效的。这证明了pnu-eda-gly5衍生物用于产生有效力的bpdc和adc(优选含有pnu-eda-gly5、或任何具有寡聚甘氨酸肽(有至少两个甘氨酸附着其上)的pnu-衍生物)的潜在作用。此外,这证明了含有优选pnu-eda-gly5、或任何具有寡聚甘氨酸肽的pnu-衍生物作为载荷的bpdc和adc用于治疗癌症疾病的作用。虽然蒽环类衍生物pnu-159682(图2)及其在化学缀合和adc情况下的用途已经在现有技术中有描述(例如wo2009099741a1、wo2010009124、wo2012073217,本文中提供作为参考),如本文所公开的类似于pnu-eda-glyn的化合物、或含adc的转肽酶-缀合的pnu-eda-glyn,在现有技术中并未有描述,而具有eda间隔子和glyn接头的pnu-衍生物的特定结构也并未在任何现有技术文献中有公开或要求保护。其中pnu衍生物经肽键(而不是酯键或马来酰亚胺接头)稳定连接至蛋白的稳定加合物,已经证明在体内稳定性和药代动力学行为方面是优越的(由于肽键在血清中一般的高稳定性),如下文实施例中所公开的那样。另外,在smac-技术缀合之后预期会展现出稳定药物缀合物的具有glyn-区段的pnu-衍生物在图6a和图6b中公开。实验和附图本发明已经在附图和前述说明书中进行了详细的说明和描述,但是此类说明和描述应当理解为是说明性的或示例性的而非限制性的;本发明不限于所公开的实施方案。本领域技术人员在实施所要求保护的发明时,从对附图、公开以及所附权利要求的研究,可以理解和实现所公开实施方案的其它变体。在权利要求中,措辞“包含”并不排除其它元素或步骤,而不定冠词“a”或“an”并不排除复数形式。仅仅是某些量度在互相不同的从属权利要求中被提及的情况,并不表示这些量度的组合不能用于有利的进行。权利要求中的任何参考迹象都不应理解为是对范围的限制。本文公开的所有氨基酸序列都是以从n-末端到c-末端显示的;本文公开的所有核酸序列都是显示为5'->3'。实施例1:通过转肽酶介导的抗体缀合技术(smac技术)产生位点特异性c-末端化的pnu-eda-glyn-载荷缀合的单克隆抗体本妥昔单抗和曲妥珠单抗特异于人cd30靶标的单克隆抗体本妥昔单抗(克隆cac10)的重链和轻链可变区序列从专利us2008213289a1获得,商业抗体herceptin(曲妥珠单抗)、或从其衍生的adc中所含有的人her-2特异性曲妥珠单抗抗体的那些序列,源自在线imgt数据库(vh:http://www.imgt.org/3dstructure-db/cgi/details.cgi?pdbcode=7637&part=chain&chain=7637h&vl:http://www.imgt.org/3dstructure-db/cgi/details.cgi?pdbcode=7637&part=chain&chain=7637l。嵌合mabcac10和人源化的mab曲妥珠单抗是用其重链和轻链产生的,其中所述重链和轻链使用本领域技术人员已知的方法在c-末端标记有转肽酶a识别序列以及另外的strepii亲和性纯化标签(hc标签序列:lpetggwshpqfek;lc标签序列:ggggslpetggwshpqfek)(参见图11a&11b)。蒽环类衍生物pnu-eda-gly5(图3a)由levanabiopharma,sandiego,ca提供,其根据图3的合成方案经亚乙基二氨基(eda)接头而将五甘氨酸肽合成至pnu159682的羰基。为此,首先用naio4在60%甲醇中于室温rt进行3小时,将商业可得的pnu159682氧化,以获得其羧酸(图3b中的1)。之后,将二氯甲烷(dcm)中的n-羟基琥珀酰亚胺(nhs,46mg,400μmol)和乙基(二甲基氨丙基)碳二亚胺(edc,100mg,523μmol)添加至6ml的dcm中的1的溶液(51mg,81μmol)。30分钟后,将混合物用水洗涤(2x6ml),经na2so4干燥并蒸发。继而将残余物溶解在2ml的二甲基甲酰胺(dmf)中,之后添加胺(图3b中的2,55mg,81μmol,作为三氯乙酸盐),随后添加n,n-二异丙基乙胺(diea,50μl)。将混合物搅拌1小时,之后添加哌啶(40μl),随后另外再进行20分钟的搅拌。通过hplc纯化混合物以给出pnu-eda-gly5(图3b中的3,34mg,44%)作为红色固体;msm/z955.2(m+h)。通过将lpetg-标记的mab[10μm]与pnu-eda-gly5[200μm]在0.62μm转肽酶a的存在下在50mmhepes、150mmnacl、5mmcacl2、ph7.5中于25℃温育3.5h,来将pnu-eda-gly5缀合至mab。通过将反应穿过经25mm磷酸钠ph7.5平衡的proteinahitrap柱(gehealthcare)来将反应终止,随后用5个柱体积(cv)的缓冲液洗涤。用5cv的洗脱缓冲液(0.1m琥珀酸,ph2.8)洗脱结合的缀合物,其有1cv的部分被收集至管中(含有25%v/v1mtris碱来中和酸)。通过g25柱使用nap25(gehealthcare)柱根据厂商指引汇集含蛋白部分,并将其配制在10mm琥珀酸钠ph5.0、100mg/ml海藻糖、0.1%%w/v聚山梨醇酯或磷酸酯20中。每种缀合物的聚积成分通过层析在tosohtskgelg3000swxl7.8mmx30cm,5μm柱上以0.5ml/min在10%ipa、0.2m磷酸钙、0.25m氯化钙、ph6.95中运行来进行评估。药物负载通过疏水相互作用层析(hic)和反相层析二者来进行评估。在tosohbutyl-npr4.6mmx3.5cm、2.5μm柱上以0.8ml/min运行(具有在a-1.5m(nh4)2so4,25mmnapi,ph=6.95±0.05与b-75%25mmnapi,ph=6.95±0.05,25%ipa之间12分钟的线性梯度)来进行hic。在polymerlabsplrp2.1mmx5cm、5μm柱上以1ml/min/80℃运行(具有0.05%tfa/h2o与0.04%tfa/ch3cn之间25分钟的线性梯度)来进行反相层析。首先通过与dtt在ph8.0于37℃温育15分钟来将样品还原。两种基于pnu-eda-gly5的adc都主要是单体,并且分别具有接近理论最大值4的药物-对-抗体-比例。表2总结了adc制备的结果。mab靶标hc标签lc标签%单体dar本妥昔单抗cd30有有99.64.0曲妥珠单抗her-2有有98.23.9表2.所制备的基于pnu-eda-gly5的adc的总结。hc,重链;lc,轻链;%单体,%单体含量;dar,药物-对-抗体-比例。实施例2:用转肽酶a-缀合的本妥昔单抗-pnu-eda-gly5和曲妥珠单抗-pnu-eda-gly5adc进行的体外细胞毒性测定使用karpas-299(表达高水平cd30的非霍奇金淋巴瘤细胞系)、和l428(表达低至中等水平cd30的霍奇金淋巴瘤细胞系),对本妥昔单抗-pnu-eda-gly5的细胞毒性进行研究(图4)。作为对照,将cac10-pnu-eda-gly5的效力与商业可得的cd30-特异性cac10-vcpab-mmae缀合物(作为阳性对照)和商业可得的her-2-特异性曲妥珠单抗-dm1缀合物(作为阴性对照)的效力进行对比。为此,将细胞在100μlrpmi/10%fcs中以每孔104细胞的密度铺在96孔平板上,并在37℃于加湿的温育器以5%的co2大气进行生长。在一天的温育之后,将25μl的培养基小心地从每孔移除并用每种adc在生长培养基中25μl的3.5-倍系列稀释来取代,导致最终的adc浓度范围在20μg/ml至0.25ng/ml。每个稀释进行一式两份。在另外4天之后,将平板从温育器移出,并平衡至室温。在大约30分钟之后,将100μl的luminescentsolution(promega,cat.nog7570)添加至每个孔,并且在450rpm摇动5分钟之后,随后是10分钟无摇动温育,在tecaninfinityf200上对发光进行了测量(每孔1秒的积分时间)。如预期的那样,用作阳性对照的抗-cd30adc有力地杀伤cd30hikarpas-299细胞(具有8.2ng/ml的ec50)(图4a),但却不足以杀伤cd30lol428细胞(图4b)。相反,用作阴性对照的抗-her-2adc未展示出特异性的细胞杀伤,且在每种细胞系都是无效的(图4)。显著地,转肽酶-缀合的adccac10-pnu-eda-gly5有力地杀伤cd30hikarpas-299细胞(且有6.9ng/ml的ec50值)(图4a)。cac10-pnu-eda-gly5仅在较高浓度杀伤cd30lol428细胞,类似于所使用的对照adc,表明此adc的效力实际上是特异性的并且是通过cd30结合来介导的(图4b)。因而,pnu-eda-gly5的转肽酶-介导的缀合产生具有很高效力的adc,甚至超过了参考adc的效力。使用skbr3细胞(过表达her-2的人乳腺癌细胞系)、和t47d细胞(天然表达低水平her-2的乳腺癌细胞系),对smac-产生的曲妥珠单抗-pnu-eda-gly5adc对于肿瘤细胞的杀伤效力进行研究,并将此与商业可得的her-2-特异性的adc曲妥珠单抗-dm1缀合物(图5)进行比较。为此,将细胞于100μldmem/10%fcs中以每孔104细胞的密度铺在96孔平板上,并且确切如上文那样进行测定。如预期的那样,阳性对照adc有力地杀伤过表达her-2的人skbr3乳腺癌细胞(且有23.7ng/ml的ec50)(图5a),而在杀伤her-2lot47d细胞方面是无效的(图5b)。显著的是,通过smac-技术产生的曲妥珠单抗-pnu-eda-gly5展现了优越的细胞毒性并且不仅杀伤过表达her-2的skbr3细胞、还杀伤her2lot47d细胞(且分别具有4.8和11.0ng/ml的ec50值)(图5)。因而,转肽酶-介导的pnu-eda-gly5与曲妥珠单抗的缀合产生具有很高效力的adc,超过了商业可得的且fda-许可的参考adc的效力,并且甚至也对her2lo人乳腺癌细胞有效。实施例3:与含有马来酰亚胺接头的曲妥珠单抗emtansine相比,转肽酶a-缀合的cac10-pnu-eda-gly5adc的体外血清稳定性在基于elisa的血清稳定性测定中,评估了本妥昔单抗-pnu-eda-gly5(cac10-pnu-eda-gly5)和kadcylaadc的体外血清稳定性。简言之,将cac10-pnu-eda-gly5在小鼠(sigma,m5905)、大鼠(sigma,r9759)和人血清(sigma,h6914)中进行了稀释,并在37℃温育。在第0、3、7、14天于液氮中将样品速冻,并储存在-80℃直至elisa分析。对于啮齿动物血清,cac10-pnu-eda-gly5血清样品的系列稀释在elisa平板上进行捕获,所述elisa平板包被有2μg/ml的小鼠抗-pnumab(通过用人igg-pnu缀合物免疫小鼠并用bsa-pnu缀合物筛选而内部生产)以结合adc、或包被有抗-人fcf(ab’)2(jacksonimmunoresearch)以结合总igg,并用hrp-缀合的抗-人iggf(ab’)2(jacksonimmunoresearch)的1:2500稀释来进行检测。对于灵长类血清,将2μg/ml的重组人cd30(sinobiologicals,10777-h08h)包被在elisa平板上,并使用hrp-缀合的抗-人iggf(ab’)2(jacksonimmunoresearch)或1μg/ml小鼠抗-pnuigg(内部生产)的1:2500稀释、随后是hrp-缀合的抗-小鼠fcf(ab’)2(jacksonimmunoresearch),来用于分别检测总igg和adc。在kadcyla的情况中,使用与上文相同的方案来确定在小鼠、大鼠和人血清中的稳定性(但却用内部生产的抗-美登素mab来结合adc)。adc和总igg的血清浓度通过与已知浓度相同adc的样品的比较,而从样品滴度的一半最大值来计算。图7a显示了优越的稳定性cac10-pnu-eda-gly5adc,特别是与含有马来酰亚胺接头的kadcyla相比(图7b),且在所测试的四个物种的任何血清中在整个实验期间adc水平都几乎没有降低。通过将0-14天的时间点与单相指数衰减函数(控制为达到0的最终浓度)拟合,在每种血清中确定了cac10-pnu-eda-gly5和kadcyla的半衰期值。kadcyla的半衰期在小鼠、大鼠和人血清中,分别为3.7天、4.4天和2.9天,而cac10-pnu-eda-gly5在小鼠、大鼠和人血清中的半衰期超过14天。实施例4:转肽酶a-缀合的ac10-gly5-pnu在小鼠中的体内稳定性在室温将ac10-gly5-pnuadc解冻,并在无菌pbs中稀释至0.2mg/ml以用于1mg/kg的剂量浓度。将样品以5ml/kg的体积静脉注射至9只雌性swisswebster小鼠中。在1h、24h、72h、7、天、14天、和21天后从动物收集血液。根据伦理标准个体动物仅用于两次至少间隔一周的血液采集时间点。因而,三只小鼠在1h和7天后接受血液采集,三只不同小鼠在24h和14天后接受血液采集,而另外三只不同小鼠在72h和21天后接受血液采集,总共每组9只小鼠。对每组的动物,在第一次采集期间通过下颌下静脉柳叶刀穿刺收集大约200μl的血液,而在最终的采集期间通过下颌下静脉柳叶刀穿刺收集大约600μl血液(末端出血)。将所有血液收集至含有k2-edta的管中。通过在1500g离心10分钟而将血浆从血液中分离,并将血浆转移至无菌冷冻小管中用于储存在-80℃直至如实施例4中所述通过elisa分析。图8中的数据显示出通过smac-技术产生的adc的高度稳定性。对于整个实验期间,adc的浓度仅略低于对总igg进行的那些测量,这表明药物和抗体之间的接头在体内稳定。通过将第3天和第21天的时间点与单相指数衰减函数(控制为达到0的最终浓度)拟合,在缓慢相中确定了体内半衰期为对于总igg和adc分别为8.3天和7.8天。实施例5:对于表达her-2的emt-6克隆的描述和表征使用工程化为过表达人her-2的鼠乳腺肿瘤细胞系emt-6来研究抗-her-2adc的细胞毒性。将emt-6细胞在补加有10%(v/v)的fcs(胎牛血清)、1%(v/v)的10,000iu/ml青霉素-链霉素和1%(v/v)的200mml-谷氨酰胺的dmem(dulbecco’smodifiedeaglemedium-高葡萄糖)中作为单层进行培养。将emt-6细胞用编码人her-2和嘌呤霉素抗性标志物的表达载体进行电穿孔,并使用本领域技术人员已知的方法筛选稳定表达人her-2的细胞集合。her-2表达通过流式细胞术确认。简言之,在胰蛋白酶化之后,将106细胞在facs管中离心;将所获得的颗粒在补加有2%的fcs的pbs(磷酸盐缓冲盐水)中重悬浮。继而将细胞与抗-her-2抗体曲妥珠单抗温育(30min,4℃),随后是离心和洗涤(3ml的pbs补加有2%fcs)。继而如前所述将细胞重悬浮,并与抗-人igg抗体(fcγ-特异性的)pe(ebioscience)在黑暗中温育(30min,4℃),之后洗涤(4mlpbs补加有2%fcs)。继而在facscalibur(bd)进行流式细胞术。her-2-转染的emt-6细胞是通过流式细胞术分选的单个细胞(使用facsariaii来分离单个细胞克隆)。将这些细胞铺展,并且通过流式细胞术验证her-2表达。图9显示了选择用于体内研究(实施例6)的克隆的facs分析数据。实施例6:转肽酶a-缀合的曲妥珠单抗-pnu-eda-gly5adc在原位乳腺癌模型中的体内效力在her-2-阳性乳腺癌的免疫活性原位小鼠模型中,对曲妥珠单抗-pnu-eda-gly5的体内效力进行评估。为此,将表达人her-2(实施例6)的106emt6小鼠乳腺癌细胞(此前确认适合体内生长),移植至雌性balb/c小鼠的右乳腺脂肪垫。此外,对照动物植入有her-2-阴性的emt6细胞。在下文中,通过卡尺测量原发肿瘤的体积。在13天后,当达到了100-150mm3的平均肿瘤体积时,将携带有肿瘤的动物根据肿瘤大小随机分至每组6只动物的组。通过静脉内注射参考adc(15mg/kg)、曲妥珠单抗-pnu-eda-gly5(1mg/kg)或载剂对照,来在同一天(第13天,也即随机分组那天)和7天后(第20天)对动物进行处理。通过卡尺监测肿瘤大小,并将肿瘤大小达到1000-1500mm3的动物处死(图10)。载剂对照小鼠中的肿瘤迅速生长,并在细胞移植后的30天内达到了大约1000mm3的平均大小(图10a)。用处理在大部分动物中对肿瘤生长只有很小的影响。6只动物中只有1只展现出肿瘤生长的显著延缓(图10c)。作为鲜明的对比,在所有用曲妥珠单抗-pnu-eda-gly5处理的动物中,肿瘤在处理期间持续退化,并且在细胞移植后的第30天基本上检测不到(图10d)。在第60天大部分动物都没有可检测到的肿瘤,而仅在1只动物中于大约第40天观测到肿瘤的复发。值得注意的是,曲妥珠单抗-pnu-eda-gly5的抗肿瘤活性是高度特异性的,并且对携带有her-2-阴性肿瘤的小鼠的处理并未导致肿瘤退化(图10b)。综合来看,所述数据证明了转肽酶-介导的位点特异性缀合的曲妥珠单抗-eda-gly5-pnuadc产生出具有体内肿瘤细胞杀伤活性(远优于参照adc)的adc。附图标注图1:位点特异性的转肽酶介导的抗体缀合(smac-技术)的示意图。单克隆抗体需要用c-末端lpxtg转肽酶标签来产生。需要产生毒素载荷以含有寡聚甘氨酸肽区段(glyn-区段),其具有一定数目的连续的甘氨酸残基(n≥1且≤21、优选n≥3且≤10、优选n=3或n=5、最优选n=5)。来自金黄色葡萄球菌的转肽酶a酶特异性地识别lpxtg五肽基序,并催化寡聚甘氨酸肽区段转肽为lpxtg的苏氨酸-甘氨酸肽键,由此在苏氨酸和寡聚甘氨酸区段的n-末端甘氨酸之间产生新的稳定肽键。图2:现有技术中描述的pnu-159682的结构(例如wo2009099741、或quintieri等人(2005)),包括用于四环糖苷配基结构的反应性碳的正式蒽环类编号体系。图3:(a)pnu衍生物-eda-gly5的结构,本文称为“pnu-eda-gly5”,用于使用本文实施例中所公开的转肽酶来通过smac-技术缀合至经c-末端lpetg转肽酶标记的单克隆抗体。(b)蒽环类衍生物pnu-eda-gly5的合成方案。图4:所示adc对于在细胞表面表达高水平cd30靶标的人非霍奇金淋巴瘤细胞系karpas-299(a)、以及对在细胞表面表达很低水平cd30靶标的人霍奇金淋巴瘤细胞系l428细胞(b)的细胞毒性作用的剂量反应。adcetris是指商业可得的抗-cd30adc本妥昔单抗-vedotin。kadcyla是指商业可得的抗-her-2/neuadct-dm1(曲妥珠单抗-emtansine)。两种细胞系对于her-2/neu都是阴性的,因此kadcyla是作为阴性对照adc,其不应当影响靶标特异性通路中的细胞杀伤。将细胞与adc的系列稀释温育4天,之后添加luminescentsolution(promega),并通过在tecaninfinityf200上测量发光来对细胞进行定量。图5:所示adc对于表达高水平her-2/neu的人乳腺癌细胞系skbr3(a)、以及对于表达低水平her-2/neu的人乳腺癌细胞系t47d(b)的细胞毒性作用的剂量反应。将细胞与adc的系列稀释温育4天,之后添加luminescentsolution(promega),并通过在tecaninfinityf200上测量发光来对细胞进行定量。图6:另外的pnu-159682相关的蒽环类衍生物,其可用于通过smac-技术而位点特异性地缀合至lpxtg-标记的结合蛋白或抗体,以产生bpdc或adc。仅有具有gly5-区段的优选版本示出。6a示出了衍生物,其中gly5氨基酸区段经过其羧基末端直接偶合至pnu衍生物的四环糖苷配基结构的a环。6b示出了衍生物,其中优选的亚乙基-氨基接头和gly5氨基酸区段直接偶合至pnu衍生物的四环糖苷配基结构的a环。图7(a)本妥昔单抗-pnu-eda-gly5adc(标记为“cac10-pnuadc”)和总igg在小鼠(a)、大鼠(b)、人类(c)血清中14天期间的体外浓度测量。(b)曲妥珠单抗-emtansineadc和总igg在小鼠(a)、大鼠(b)和人类(c)血清中14天期间的体外浓度测量。图8:在小鼠中施用ac10-gly5-pnuadc之后21天期间6个时间点测量的adc和总igg的体内血浆浓度。图9:在与抗-her-2抗体曲妥珠单抗温育并继而与含荧光团的抗-人igg抗体(fcγ-特异性的)pe温育之后,选择用于体内研究的emt-6her-2克隆的facs分析数据。图10:在her2-阳性乳腺癌的免疫活性原位小鼠模型中,对her-2-特异性adc的体内评估。表达人her-2(a,c,d)或不相关抗原ror-1的emt6小鼠乳腺癌细胞在balb/c小鼠的乳腺脂肪垫中生长。在第13天和20天,将动物经静脉注射用对照(a)、1mg/kg曲妥珠单抗-pnu159682(b,d)、或15mg/kgkadcyla(c)进行处理。对肿瘤生长进行检测直至动物由于伦理援引必须处死。图11a&b:含有曲妥珠单抗(a)和本妥昔单抗(b)pnu-毒素衍生物的用于所述研究的adc中经c-末端smac-技术tm缀合的igh和igl链的氨基酸组成,包含图3b中所示的pnu衍生物通过gly5-区段的氨基经肽键连接至转肽酶标签(以粗体突出)的第4个氨基酸(在转肽酶缀合之后)。参考文献beerlietal.(2015)plosone10,e0131177dorretal.(2014)pnas111,13343-8quintieriletal(2005),clincancerres.2005feb15;11(4):1608-17.roguskaetal.(1994)pnas91,pp969-971perezetal.(2014)drugdiscoverytoday19,pp.869-881alleyetal.(2008)bioconjug.chem.19,pp.759-765panowskietal.(2014)mabs6,pp.34-45wolffacetal(2013),jclinoncol.2013nov1;31(31):3997-4013youngkh(2014)clinicaladvancesinhematology&oncology,volume12,issue4,supplement10spirigetal.(2011)mol.microbiol.82,1044-1059ducry&stump(2010)bioconjug.chem.21,5-13,mccombsetal.(2015)theaapsjournal17,339-351mullard(2013)naturerevdrugdisc12,329-332doktoretal.(2014)molcancerther13,2618-2629hartley&hochhauser(2012)curr.opin.pharmacol.12,398-402minotti(2004)pharmacol.rev56,185-229cancerandchemotherpay-antineoplasticagentsvol.iii,stanleyt.crookeandarchiew.prestayko(eds.),academicpress1981).序列表<110>恩比伊治疗股份公司<120>包含蒽环类衍生物的结合蛋白药物缀合物<130>nd40432<160>4<170>patentinversion3.5<210>1<211>459<212>prt<213>artificial<220><223>trastuzumab-hc-lpetggggg-pnu-toxin<400>1gluvalglnleuvalgluserglyglyglyleuvalglnproglygly151015serleuargleusercysalaalaserglypheasnilelysaspthr202530tyrilehistrpvalargglnalaproglylysglyleuglutrpval354045alaargiletyrprothrasnglytyrthrargtyralaaspserval505560lysglyargphethrileseralaaspthrserlysasnthralatyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095serargtrpglyglyaspglyphetyralametasptyrtrpglygln100105110glythrleuvalthrvalserseralaserthrlysglyproserval115120125pheproleualaproserserlysserthrserglyglythralaala130135140leuglycysleuvallysasptyrpheprogluprovalthrvalser145150155160trpasnserglyalaleuthrserglyvalhisthrpheproalaval165170175leuglnserserglyleutyrserleuserservalvalthrvalpro180185190serserserleuglythrglnthrtyrilecysasnvalasnhislys195200205proserasnthrlysvalasplyslysvalgluprolyssercysasp210215220lysthrhisthrcysproprocysproalaprogluleuleuglygly225230235240proservalpheleupheproprolysprolysaspthrleumetile245250255serargthrprogluvalthrcysvalvalvalaspvalserhisglu260265270aspprogluvallyspheasntrptyrvalaspglyvalgluvalhis275280285asnalalysthrlysproargglugluglntyrasnserthrtyrarg290295300valvalservalleuthrvalleuhisglnasptrpleuasnglylys305310315320glutyrlyscyslysvalserasnlysalaleuproalaproileglu325330335lysthrileserlysalalysglyglnproarggluproglnvaltyr340345350thrleuproproserargaspgluleuthrlysasnglnvalserleu355360365thrcysleuvallysglyphetyrproseraspilealavalglutrp370375380gluserasnglyglnprogluasnasntyrlysthrthrproproval385390395400leuaspseraspglyserphepheleutyrserlysleuthrvalasp405410415lysserargtrpglnglnglyasnvalphesercysservalmethis420425430glualaleuhisasnhistyrthrglnlysserleuserleuserpro435440445glylysleuprogluthrglyglyglyglygly450455<210>2<211>228<212>prt<213>artificial<220><223>trastuzumab-lc-ggggs-lpetggggg-pnu-toxin<400>2aspileglnmetthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnaspvalasnthrala202530valalatrptyrglnglnlysproglylysalaprolysleuleuile354045tyrseralaserpheleutyrserglyvalproserargphesergly505560serargserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysglnglnhistyrthrthrpropro859095thrpheglyglnglythrlysvalgluilelysargthrvalalaala100105110proservalpheilepheproproseraspgluglnleulyssergly115120125thralaservalvalcysleuleuasnasnphetyrproarggluala130135140lysvalglntrplysvalaspasnalaleuglnserglyasnsergln145150155160gluservalthrgluglnaspserlysaspserthrtyrserleuser165170175serthrleuthrleuserlysalaasptyrglulyshislysvaltyr180185190alacysgluvalthrhisglnglyleuserserprovalthrlysser195200205pheasnargglyglucysglyglyglyglyserleuprogluthrgly210215220glyglyglygly225<210>3<211>456<212>prt<213>artificial<220><223>brentuximab-hc-lpetggggg-pnu-toxin<400>3glnileglnleuglnglnserglyprogluvalvallysproglyala151015servallysilesercyslysalaserglytyrthrphethrasptyr202530tyrilethrtrpvallysglnlysproglyglnglyleuglutrpile354045glytrpiletyrproglyserglyasnthrlystyrasnglulysphe505560lysglylysalathrleuthrvalaspthrserserserthralaphe65707580metglnleuserserleuthrsergluaspthralavaltyrphecys859095alaasntyrglyasntyrtrpphealatyrtrpglyglnglythrgln100105110valthrvalseralaalaserthrlysglyproservalpheproleu115120125alaproserserlysserthrserglyglythralaalaleuglycys130135140leuvallysasptyrpheprogluprovalthrvalsertrpasnser145150155160glyalaleuthrserglyvalhisthrpheproalavalleuglnser165170175serglyleutyrserleuserservalvalthrvalproserserser180185190leuglythrglnthrtyrilecysasnvalasnhislysproserasn195200205thrlysvalasplyslysvalgluprolyssercysasplysthrhis210215220thrcysproprocysproalaprogluleuleuglyglyproserval225230235240pheleupheproprolysprolysaspthrleumetileserargthr245250255progluvalthrcysvalvalvalaspvalserhisgluaspproglu260265270vallyspheasntrptyrvalaspglyvalgluvalhisasnalalys275280285thrlysproargglugluglntyrasnserthrtyrargvalvalser290295300valleuthrvalleuhisglnasptrpleuasnglylysglutyrlys305310315320cyslysvalserasnlysalaleuproalaproileglulysthrile325330335serlysalalysglyglnproarggluproglnvaltyrthrleupro340345350proserargaspgluleuthrlysasnglnvalserleuthrcysleu355360365vallysglyphetyrproseraspilealavalglutrpgluserasn370375380glyglnprogluasnasntyrlysthrthrproprovalleuaspser385390395400aspglyserphepheleutyrserlysleuthrvalasplysserarg405410415trpglnglnglyasnvalphesercysservalmethisglualaleu420425430hisasnhistyrthrglnlysserleuserleuserproglylysleu435440445progluthrglyglyglyglygly450455<210>4<211>232<212>prt<213>artificial<220><223>brentuximab-lc-ggggs-lpetggggg-pnu-toxin<400>4aspilevalleuthrglnserproalaserleualavalserleugly151015glnargalathrilesercyslysalaserglnservalasppheasp202530glyaspsertyrmetasntrptyrglnglnlysproglyglnpropro354045lysvalleuiletyralaalaserasnleugluserglyileproala505560argpheserglyserglyserglythraspphethrleuasnilehis65707580provalgluglugluaspalaalathrtyrtyrcysglnglnserasn859095gluaspprotrpthrpheglyglyglythrlysleugluilelysarg100105110thrvalalaalaproservalpheilepheproproseraspglugln115120125leulysserglythralaservalvalcysleuleuasnasnphetyr130135140proargglualalysvalglntrplysvalaspasnalaleuglnser145150155160glyasnserglngluservalthrgluglnaspserlysaspserthr165170175tyrserleuserserthrleuthrleuserlysalaasptyrglulys180185190hislysvaltyralacysgluvalthrhisglnglyleuserserpro195200205valthrlysserpheasnargglyglucysglyglyglyglyserleu210215220progluthrglyglyglyglygly225230当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1
- 联苯胺衍生物、其制造方法和电子照相感光体与制造工艺
- 包含环糊精衍生物的可注射美法仑组合物及其制备和使用方法与制造工艺
- 一种黄芪甲苷衍生物及其制备方法和应用与制造工艺
- (Z,E)5‑OH‑3‑(苯基衍生物‑次甲基)‑二氢黄酮‑7‑O‑糖苷及其制备及其应用的制造方法与工艺
- 喹唑啉衍生物的制造方法与工艺
- 吡唑衍生物及其作为大麻素受体介体的用途的制造方法与工艺
- Harrisotone A衍生物的组合物在防治肝脏损伤药物中的应用的制造方法与工艺
- 水生态处理中淤泥及衍生物的再利用处理系统的制造方法与工艺
- Schiglautone A衍生物的组合物用于制备抗缺氧药物的制作方法
- Schiglautone A衍生物的组合物用于制备抗红细胞低下性贫血药物的制作方法