用于使NK细胞活化的组合物和方法与流程
相关申请的交叉引用
本申请要求2017年2月15日提交的美国临时专利申请序列号62/459,397的优先权权益,所述美国临时专利申请以引用的方式整体并入本文。
发明背景
天然杀伤(nk)细胞使具有较低表达的i类mhc、cd54和b7h1以及较高表达的cd44的癌干细胞/未分化肿瘤溶解和分化。外周血液淋巴细胞的中等和高度细胞毒性活性与癌症风险降低相关联,并且nk细胞对肿瘤的高度浸润与较好预后相关联,而低活性与癌症风险增加相关联。
对nk细胞的抑制由肿瘤微环境中nk受体的下调介导。先前显示在肿瘤患者中,nk细胞的功能显著降低。已开发若干体外nk扩增技术以允许达成较高治疗性细胞剂量。用饲养细胞诸如表达白介素(il)-15和41bb配体的k562细胞、ebv-tm-lcl、韦尔姆斯氏肿瘤(wilm’stumor)或经照射pbmc刺激外周血液单核细胞(pbmc)或纯化nk细胞群体已产生较大数目的具有足够功能的nk细胞。所产生的nk细胞表达较高水平的nkg2d、天然细胞毒性受体、dnam-1和icam-1。因此,已建立用以获得经离体扩增、经活化和cd3+t细胞经消减nk细胞的各种方法来供临床使用。此外,已确定用hla单倍同一性nk细胞在患有晚期癌症的患者中进行继承性细胞转移具有安全性和功效。另外,临床试验已显示同种异体nk细胞在实体肿瘤的情况下起治疗作用,并且就向患者中转移来说是安全的。
用nk细胞进行的免疫疗法已由于不能获得足够数目的高度功能性nk细胞而受限。此外,不同于来自健康个体的nk细胞,与来自携带肿瘤的人源化小鼠的那些nk细胞类似,患者nk细胞的扩增由于小部分污染性t细胞的扩增而显著受限,所述污染性t细胞由于它们的增殖能力更快而排挤nk细胞。
对nk细胞免疫调节的潜伏机理不了解。存在极大需要来鉴定用于达成改进的nk免疫疗法的治疗性组合物和方法。
技术实现要素:
本发明至少部分地基于发现破骨细胞可在健康人与癌症患者两者中诱导nk细胞的扩增,此进一步使cd8+/cd4+t细胞比率增加。尽管癌症患者相对于健康人通常在体内具有更多nk细胞和更大cd8+/cd4+t细胞比率,但过量nk细胞和cd8+t细胞是短命的(归因于污染性t细胞的扩增可抑制nk细胞功能),并且缺乏活性(例如细胞毒性和细胞因子选择)。然而,破骨细胞可在癌症患者中诱导nk细胞扩增,并且使nk细胞的细胞数目与功能两者均增加(例如通过它们的细胞因子分泌能力所度量)。树突细胞优先促进t细胞的扩增,而破骨细胞优先促进nk细胞的扩增,从而表明选择性扩增t细胞和nk细胞的微环境差异。因此,本发明提供一种用以扩增大量经活化nk细胞以用于免疫治疗策略中的方法。所述细胞可用于通过促进干细胞样/不良分化肿瘤的分化来抑制或消除癌干细胞,并且控制肿瘤生长。
本文提供一种在体外或离体使nk细胞活化的方法,其包括连同破骨细胞(oc)一起在培养基中培养所述nk细胞。nk可为原代nk细胞,任选地,其中它尚未被转化。经活化nk细胞可在4周内扩增约10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25次或更多次群体倍增。培养物可包含多个破骨细胞(oc)和多个nk细胞,例如其中细胞培养物中oc:nk细胞的比率是至少1:2。破骨细胞可使nk细胞细胞毒性增强,例如如通过由nk细胞对口腔鳞状癌干细胞样细胞(oscsc)的溶解所度量或通过51cr释放细胞毒性测定所测量。
另外,破骨细胞可使由nk细胞产生的至少一种细胞因子或趋化因子的产生、分泌和/或功能增强。举例来说,破骨细胞可使由nk细胞对ifn-γ和/或il-12的分泌,和/或由nk细胞对nkg2d、nkp46、nkp44、nkp30、cd94、kir2和kir3中的一者或多者的表达增强。
nk细胞可为从人受试者的癌症样品纯化的细胞。在某些实施方案中,细胞培养物进一步包含也来源于癌症样品的t细胞。在某些所述实施方案中,相对于t细胞,nk细胞可得以优先扩增。可使nk细胞扩增任何时长,例如至少一个月。培养基可被补充以至少一个破骨细胞以继续优先扩增nk细胞。t细胞可分泌ifn-γ,但可不介导细胞毒性,例如如通过例如在51cr释放细胞毒性测定中由t细胞对oscsc的溶解所度量。经扩增nk细胞可能够使cd8+t细胞扩增。相对于cd4+t细胞,由oc扩增的nk细胞也可能够优先扩增cd8+t细胞。在某些实施方案中,方法进一步包括将抗cd3抗体添加至细胞培养物中,例如以进一步增强由nk细胞对ifn-γ的分泌。经活化nk细胞可为分割无反应化的。
在某些实施方案中,方法可进一步包括向细胞培养物中添加包含至少一种选自以下的细菌菌株的组合物:嗜热链球菌(streptococcusthermophiles)、长双歧杆菌(bifidobacteriumlongum)、短双歧杆菌(bifidobacteriumbreve)、婴儿双歧杆菌(bifidobacteriuminfantis)、嗜酸乳酸杆菌(lactobacillusacidophilus)、植物乳酸杆菌(lactobacillusplantarum)、副干酪乳酸杆菌(lactobacillusparacasei)、ke99和保加利亚乳酸杆菌(lactobacillusbulgaricus),任选地,其中所述至少一种细菌菌株可为活的或经声波处理的。举例来说,组合物可包含嗜热链球菌、长双歧杆菌、短双歧杆菌、婴儿双歧杆菌、嗜酸乳酸杆菌、植物乳酸杆菌、副干酪乳酸杆菌、ke99和保加利亚乳酸杆菌。或者或另外,组合物可包含saj2细菌。细胞培养物中saj2细菌浓度与nk细胞和/或oc浓度的比率可为例如i)对于nk细胞:saj2是至少1:2;ii)对于oc:saj2是至少1:4;和/或iii)对于oc:nk细胞:saj2是至少1:2:4。
在某些实施方案中,方法可进一步包括向细胞培养物中添加能够使nk细胞活化的另一药剂。
在某些优选实施方案中,方法包括:i)提供包含破骨细胞(oc)、nk细胞和t细胞的细胞培养物;和ii)培养所述细胞培养物中的所述nk细胞、所述t细胞和所述破骨细胞,由此相对于所述t细胞,优先活化所述nk细胞。
类似地,本文提供一种方法,其包括:i)提供包含树突细胞(dc)、nk细胞和t细胞的细胞培养物;和ii)培养所述细胞培养物中的所述nk细胞、所述t细胞和所述树突细胞,由此相对于所述nk细胞,优先活化所述t细胞。在一实施方案中,nk细胞可为原代nk细胞,任选地,其中所述原代nk细胞尚未被转化。培养物可包含多个破骨细胞(oc)和多个nk细胞,例如其中细胞培养物中oc:nk细胞的比率是至少1:2。破骨细胞可使nk细胞扩增,和/或由nk细胞对il-15的分泌增强。经活化nk细胞可扩增以在4周内达到至少约10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25次或更多次群体倍增。另外,破骨细胞可使nk细胞细胞毒性增强,例如如通过由nk细胞对口腔鳞状癌干细胞样细胞(oscsc)的溶解所度量。细胞细胞毒性可通过51cr释放细胞毒性测定来测量。
在某些实施方案中,破骨细胞(oc)可使由nk细胞产生的至少一种细胞因子或趋化因子的产生、分泌和/或功能增强。举例来说,破骨细胞可使由nk细胞对ifn-γ和/或il-12的分泌增强。破骨细胞可使由nk细胞对nkg2d、nkp46、nkp44、nkp30、cd94、kir2和kir3中的一者或多者的表达增强。nk细胞和/或t细胞可从来自受试者例如人受试者的癌症样品纯化。在某些实施方案中,对nk细胞的优先活化可持续至少一个月。另外,在对nk细胞的优先活化减弱或停止之后,可将至少一个破骨细胞添加至在培养nk细胞之后持续至少一个月的细胞培养物中,由此继续使nk细胞活化。在一些实施方案中,t细胞可分泌ifn-γ,但可不介导细胞毒性。细胞毒性可通过由t细胞对oscsc的溶解所度量,例如通过51cr释放细胞毒性测定来测量。经扩增nk细胞可能够使cd8+t细胞扩增,并且相对于cd4+t细胞,可能够优先扩增cd8+t细胞。在某些实施方案中,相对于cd8+t细胞,由dc扩增的nk细胞可能够优先扩增cd4+t细胞。在其他实施方案中,可将抗cd3抗体添加至细胞培养物中,例如进一步增强由nk细胞对ifn-γ的分泌。另外,经活化nk细胞可为分割无反应化的。
在某些实施方案中,方法可进一步包括向细胞培养物中添加包含至少一种选自以下的细菌菌株的组合物:嗜热链球菌、长双歧杆菌、短双歧杆菌、婴儿双歧杆菌、嗜酸乳酸杆菌、植物乳酸杆菌、副干酪乳酸杆菌、ke99和保加利亚乳酸杆菌,任选地,其中所述至少一种细菌菌株是活的或经声波处理的。举例来说,组合物可包含嗜热链球菌、长双歧杆菌、短双歧杆菌、婴儿双歧杆菌、嗜酸乳酸杆菌、植物乳酸杆菌、副干酪乳酸杆菌、ke99和保加利亚乳酸杆菌。或者或另外,组合物可包含saj2细菌。在某些所述实施方案中,细胞培养物中saj2细菌浓度与nk细胞和/或oc浓度的比率可为i)对于nk细胞:saj2是至少1:2;ii)对于oc:saj2是至少1:4;和/或iii)对于oc:nk细胞:saj2是至少1:2:4。
另外,方法可进一步包括向细胞培养物中添加可能够使nk细胞活化的另一药剂。方法可进一步包括向细胞培养物中添加能够使t细胞活化的另一药剂。
类似地,本文提供一种通过以下方式来治疗患有或被怀疑患有癌症或癌症相关疾病或病症的受试者的癌症或癌症相关疾病或病症的方法:向所述受试者施用治疗有效量的破骨细胞(oc)、包含破骨细胞(oc)的细胞培养物、和/或包含破骨细胞(oc)的细胞培养物的上清液。
破骨细胞可使受试者中的nk细胞扩增增强,任选地,其中破骨细胞使由nk细胞对il-15的分泌增强。在某些实施方案中,增强的nk细胞扩增可为在4周内至少约10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25次或更多次群体倍增。另外,破骨细胞可使nk细胞细胞毒性增强,例如如通过由nk细胞对口腔鳞状癌干细胞样细胞(oscsc)的溶解所度量。细胞细胞毒性可通过51cr释放细胞毒性测定来测量。
在某些实施方案中,破骨细胞可增加或促进由nk细胞产生的至少一种细胞因子或趋化因子的产生、分泌和/或功能。举例来说,破骨细胞可使由nk细胞对ifn-γ和/或il-12的分泌增强。相对于t细胞,破骨细胞可优先活化nk细胞,和/或相对于t细胞,优先增强nk细胞的扩增。在某些实施方案中,对nk细胞的优先扩增可持续至少一个月。t细胞可分泌ifn-γ,但不介导对癌症的细胞毒性,例如如通过在51cr释放细胞毒性测定中由t细胞对oscsc的溶解所度量。
在某些实施方案中,经活化nk细胞可使受试者中的cd8+t细胞扩增,例如相对于cd4+t细胞,可优先扩增cd8+t细胞。受试者也可用抗cd3抗体治疗以进一步增强由nk细胞对ifn-γ的分泌。经活化nk细胞可为分割无反应化的。
在某些实施方案中,治疗方法可进一步包括向受试者施用包含至少一种选自以下的细菌菌株的组合物:嗜热链球菌、长双歧杆菌、短双歧杆菌、婴儿双歧杆菌、嗜酸乳酸杆菌、植物乳酸杆菌、副干酪乳酸杆菌、ke99和保加利亚乳酸杆菌,任选地,其中所述至少一种细菌菌株是活的或经声波处理的。举例来说,组合物可包含嗜热链球菌、长双歧杆菌、短双歧杆菌、婴儿双歧杆菌、嗜酸乳酸杆菌、植物乳酸杆菌、副干酪乳酸杆菌、ke99和保加利亚乳酸杆菌。在某些实施方案中,组合物可包含saj2细菌。
在某些实施方案中,方法可进一步包括向细胞培养物中添加能够使nk细胞活化的另一药剂。
在某些实施方案中,破骨细胞、细胞培养物和/或上清液可以药物组合物形式施用,所述药物组合物可以全身性方式或以局部方式向癌症施用。在某些实施方案中,破骨细胞、细胞培养物和/或上清液可向受试者施用至少两次,例如破骨细胞、细胞培养物和/或上清液可在首次施用以后至少一个月之后向受试者施用。
在一些实施方案中,受试者可为人。
附图说明
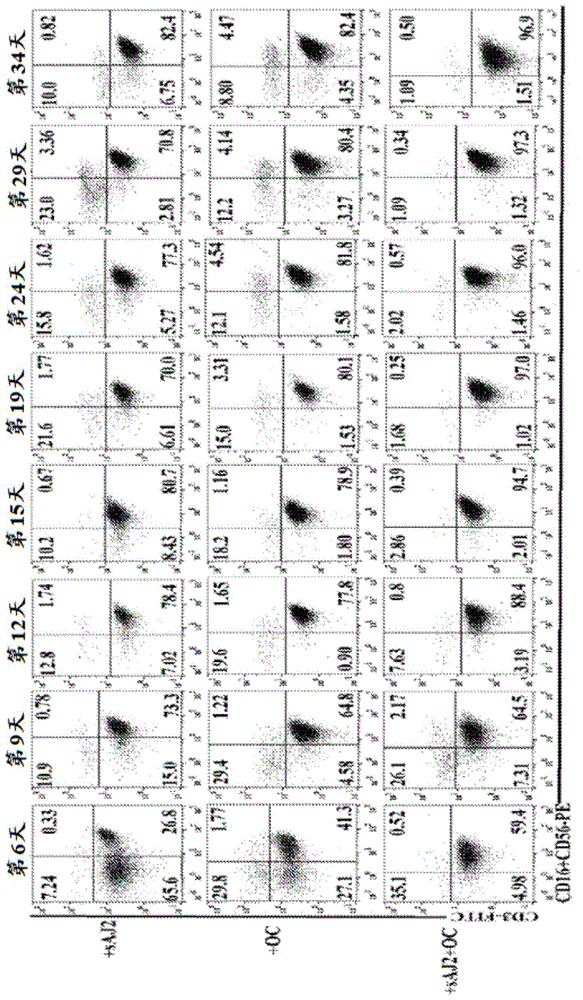
图1包括四个图版,标识为图版a、b、c和d,其显示由破骨细胞对nk活化性配体的较高表达。为产生破骨细胞(oc),在含有巨噬细胞集落刺激因子(m-csf)(25ng/ml)和rankl(25ng/ml)的培养基中培养单核细胞21天。用il-2(1000u/ml)和抗cd16mab(3μg/ml)的组合处理高度纯化nk细胞(1x106个细胞/毫升)18小时,随后使它们与自体oc分别在存在saj2细菌下在1:2:4比率(oc:nk:saj2)下或在不存在saj2细菌下共培养。在第6、9、12、15、19、24、29和34天时,使用流式细胞计量术来分析来自共培养物的含1x104个淋巴细胞的样品中cd3、cd16和cd56的表面表达,并且将培养基更新并补充以rh-il-2(1000u/ml)(图1a)。如图1a中所述来共培养细胞,并且使用显微镜来对经扩增淋巴细胞进行手动计数(图1b)。从健康供者的pbmc分离单核细胞。为产生树突细胞,在含有gm-csf(150ng/ml)和il-4(50ng/ml)的培养基中培养单核细胞8天。如图1a中所述来产生破骨细胞以及使nk细胞纯化,随后使它们分别与自体细胞在saj2细菌存在下在1:2:4比率(靶标细胞:nk:saj2)下共培养。在培养的第6天,移除培养基,并且用rh-il-2处理nk细胞5小时,随后收集上清液,并且使用单一elisa来测定ifn-γ分泌(图1c)。如图1a和图1d中所述来分离单核细胞,产生树突细胞和破骨细胞,如材料和方法中所述来培养oscsc和k562肿瘤细胞系,采用pe缀合抗体和流式细胞计量术,1x104个细胞用于分析mhc-1、cd54、kir2、kir3、klrg1和mica/b表面表达。igg2同种型用作对照(图1d)。
图2包括十个图版,标识为图版a至j,其显示由破骨细胞对nk细胞以及由树突细胞对t细胞的优先扩增和显著功能获得。从人pbmc纯化单核细胞,并且与gm-csf(150ng/ml)和il-4(50ng/ml)一起培养8天以产生dc。为产生破骨细胞,在含有m-csf(25ng/ml)和rankl(25ng/ml)的α-mem培养基中培养单核细胞21天。对于扩增,用il-2(1000u/ml)和抗cd16mab(3μg/ml)的组合处理纯化nk细胞(1x106个细胞/毫升)18小时,随后使它们与自体单核细胞、dc或oc在saj2存在下在1:2:4比率(单核细胞、dc或oc:nk:saj2)下共培养。在图中指示的天数时,使用流式细胞计量术来分析cd3、cd16和cd56的表面表达,并且将培养基更新并补充以rh-il-2(1000u/ml)(图2a)。如图2a中所述来共培养细胞,并且使用显微测定来评估经扩增淋巴细胞的数目(图2b)。使用图2b中总体扩增细胞内nk细胞和t/nkt细胞的百分比(图2a)来测定nk细胞(图2c)和t/nkt(图2d)细胞的数目。如图2a中所述来共培养细胞,并且使用针对口腔鳞状细胞癌干细胞系(oscsc)的标准4小时51cr释放测定来测定在图中所示的天数时的细胞毒性。使用为使30%的oscsc溶解所需的淋巴细胞数的倒数x100来测定溶解单位30/106个细胞(图2e)。在第6、9、12和15天,从如图1a中所述的nk与oc的共培养物收集上清液,并且使用单一elisa来测定ifn-γ分泌(图2f)。如图2a中所述来使来自10名健康供者的nk细胞与自体破骨细胞共培养,并且加以扩增。计算各供者的nk细胞持续31天的累积扩增倍数(图2g),并且基于最终计数与基线计数的比率的对数除以2的对数来计算群体倍增(图2h)。如图2a中所述来产生树突细胞和破骨细胞,并且1x104个细胞用于使用pe缀合抗体和流式细胞计量分析来分析ulbp、kir2、kir3、klrg1和mica/b表面表达。igg2同种型对照抗体用作对照(图2i)。经新鲜分离nk细胞(上排)和如图2a中所述来与自体破骨细胞共培养并加以扩增的nk细胞(下排)用于使用pe缀合抗体来分析cd16、nkp30、nkp44、nkp46、kir2、kir3、cd94和nkg2d表面表达。igg2同种型对照抗体用作对照(图2j)。
图3包括八个图版,标识为图版a至h,其显示不同于nk细胞,从经破骨细胞扩增nk细胞纯化的t细胞不介导针对oscsc的细胞毒性,并且适度分泌ifn-γ。如材料和方法中所述来处理经新鲜纯化nk细胞,并且使其与单核细胞源性自体破骨细胞共培养。使用fitc和pe缀合抗体以及流式细胞计量术,分析来自在第9天时的共培养物的淋巴细胞样品中cd3、cd16、cd56、gl3(tcrγ/δ)、cd4和cd8的表面表达(图3a)。如图1a中所述来处理nk细胞,并且使其与自体破骨细胞共培养,并且在第9天,使用cd3t正性选择试剂盒来将cd3t阳性细胞分选出来,使用cd3、cd16、cd56fitc和pe缀合抗体以及流式细胞计量术来评估cd3t阴性(nk)细胞的纯度(图3b)。用rh-il-2(1000u/ml)处理cd3t阳性细胞和cd3t阴性细胞(cd16阳性细胞)18-20小时,随后使用针对oscsc(图3c)和k562(图3d)细胞系的标准4小时51cr释放测定来测试它们的细胞毒性。使用材料和方法中所述的方法来分别测定针对oscsc和k562的溶解单位30/106个细胞。从培养物收集上清液,并且使用单一elisa来测定ifn-γ分泌(图3e)。如材料和方法中所述来从pbmc纯化nk细胞、cd3t、cd4t、cd8t和γδt细胞,并且用rh-il-2活化18-20小时,随后使用针对oscsc的标准4小时51cr释放测定来测试它们的细胞毒性(图3f)。使用为使30%的oscsc溶解所需的淋巴细胞数的倒数x100来测定溶解单位30/106个细胞(图3f)。如材料和方法中所述来从pbmc纯化nk细胞和t细胞,如材料和方法中所述来处理nk细胞。在使t细胞与自体oc一起培养之前18-20小时,用cd3抗体(1μg/ml)和cd28抗体(3μg/ml)活化t细胞,并且在培养之后第4天使用显微镜来对经扩增淋巴细胞进行手动计数(图3g)。如图3g中所述来使nk细胞和t细胞纯化并与oc一起培养,并且在第4天加以计数,用由oc扩增的淋巴细胞的扩增倍数除以在无oc下的淋巴细胞扩增倍数(图3h)。
图4包括19个图版,标识为图版a至s,其显示在用破骨细胞和saj2细菌各次连续再刺激nk细胞培养物的情况下,nk细胞比例、nk细胞介导的细胞毒性和ifn-γ分泌降低。如图2a中所述来处理经新鲜纯化nk细胞,并且使其与单核细胞源性自体破骨细胞共培养。在图中指示的天数时,使用流式细胞计量分析来分析来自共培养物的1x104个淋巴细胞中cd3、cd16和cd56的表面表达(图4a)。在36天之后,当nk细胞停止扩增时,如图2a中所述来使它们与新鲜自体破骨细胞一起再培养。使用抗体染色和流式细胞计量分析来分析在图中指示的天数时的cd3、cd16和cd56表面表达(图4b)。在第63天,当细胞停止扩增时,如所述来使它们与oc一起再培养,并且分析在图中所示的天数时的cd3、cd16和cd56表面表达(图4c)。使用显微测定来评估经扩增淋巴细胞的数目(图4d、图4g、图4j),并且使用总体扩增细胞(图4d)内nk细胞和t/nkt细胞的百分比来测定nk细胞(图4f、图4i、图4l)和t/nkt(图4e、图4h、图4k)细胞的数目。使用碘化丙锭染色和流式细胞计量分析来测定在第36、59和83天时淋巴细胞的细胞死亡(图4m)。如图2a中所述来处理经新鲜纯化nk细胞,并且使其与自体破骨细胞共培养。接着在6、17和34天共培养(图4n),40和63天共培养(图4o),或76和92天共培养(图4p)之后,使用针对oscsc的标准4小时51cr释放测定来测试淋巴细胞的细胞毒性。使用图2e中所述的方法来测定溶解单位30/106个细胞。收集上清液,使用来自第6、9、12、15、18和21天(图4q);第40、46、51、55和59天(图4r)以及第76、83、92和97天(图4s)的上清液,利用单一elisa来测定ifn-γ分泌。
图5包括八个图版,标识为图版a至h,其显示破骨细胞而非k562或oscsc实质上使nk细胞扩增,并且使nk细胞功能增加。为产生破骨细胞,在含有m-csf(25ng/ml)和rankl(25ng/ml)的培养基中培养单核细胞21天,如材料和方法中所述来培养k562肿瘤细胞系。用il-2(1000u/ml)和抗cd16mab(3μg/ml)的组合处理高度纯化nk细胞(1x106个细胞/毫升)18小时,随后使它们分别与k562和自体oc在saj2细菌存在下在1:2:4比率(oc:nk:saj2)下共培养。在第6、10和13天,使用显微镜来对来自培养物的细胞进行手动计数(图5a)。如图1a中所述来产生破骨细胞,并且如材料和方法中所述来在40戈瑞(gy)下对k562肿瘤细胞系进行照射。如材料和方法中所述来纯化和处理nk细胞,随后使它们分别与经照射k562和经照射自体oc在saj2细菌存在下在1:2:4比率(oc:nk:saj2)下共培养。在第6、10和13天,使用显微镜来对来自培养物的细胞进行手动计数(图5b)。如图1a中所述来纯化nk细胞并与oc和k562一起培养,使用针对oscsc的标准4小时51cr释放测定来测定共培养6天的淋巴细胞的细胞毒性。使用图3f中所述的方法来测定溶解单位30/106个细胞(图5c)。如图5b中所述来纯化nk细胞并与oc和k562一起培养,使用针对oscsc的标准4小时51cr释放测定来测定共培养6天的淋巴细胞的细胞毒性。使用图3f中所述的方法来测定溶解单位30/106个细胞(图5d)。如图5a中所述来纯化nk细胞并与oc和oscsc一起培养,使用针对oscsc的标准4小时51cr释放测定来测定共培养6天的淋巴细胞的细胞毒性。使用图3f中所述的方法来测定溶解单位30/106个细胞(图5e)。如图3a中所述来纯化nk细胞并与oc和k562一起培养,在第3、6、7、10和13天收集上清液,并且使用单一elisa来测定ifn-γ分泌(图5f)。使nk细胞纯化,并且与经照射oc和经照射k562一起培养,在第3、6、7、10和13天收集上清液,并且使用单一elisa来测定ifn-γ分泌(图5g)。使nk细胞纯化,并且与oc和oscsc一起培养,在第1、3、6和8天收集上清液,并且使用单一elisa来测定ifn-γ分泌(图5h)。
图6包括16个图版,标识为图版a至p,其显示相较于来自健康供者的经扩增nk细胞,来自癌症患者的与oc一起培养的纯化nk细胞使t细胞比nk细胞以更大程度扩增,介导低得多的细胞毒性和细胞因子分泌。如图2a中所述来处理来自健康供者和癌症患者的经新鲜纯化nk细胞,并且与单核细胞源性oc共培养。依次使用抗体染色和流式细胞计量分析来分析癌症患者(图6a)和健康供者(图6b)的在第6、9、12、15、18、21、24、27和31天时的扩增细胞上的cd3、cd16和cd56表面表达。使用pi染色和流式细胞计量分析来测定在第19天时,来自癌症患者和健康供者的扩增nk细胞的细胞死亡(图6c)。在6、9、12、15、18、21、24、27和31天共培养之后,使用显微术来对经扩增淋巴细胞进行手动计数(图6d)。使用图6a和图6b中总体扩增细胞内nk细胞和t/nkt细胞的百分比来测定nk细胞(图6e)和t/nkt(图6f)细胞的数目。使用针对oscsc的标准4小时51cr释放测定来测定在第12和15天的淋巴细胞细胞毒性。使用图2e中所述的方法来测定溶解单位30/106个细胞。基于每个nk细胞来使来自图6f的溶解单位标准化(图6h)。从过夜、第6、9、12、15、18、21、24和27天共培养物收集上清液,并且使用单一elisa来测定ifn-γ(图6i)、il-10(图6j)和il-6(图6k)分泌。如图2a中所述来处理来自健康供者、患有扁桃体癌的癌症患者(1号患者)和患有胰腺癌的癌症患者(2号患者)的经新鲜纯化nk细胞,并且与oc共培养。分析来自健康供者和患者nk的第21天以及健康供者的第87天(第3次刺激)的共培养物的淋巴细胞上的cd3、cd16和cd56表面表达(图6l),并且分析cd16阳性细胞内nkp30、nkp44、nkp46、kir2、kir3、cd94和nkg2d表面表达(图6m)。igg2同种型对照抗体用作对照(图6l和图6m)。从在第13天的共培养物收集上清液,并且来自各供者的等量上清液(200μl)用于使oscsc分化过夜,随后测定oscsc上的mhc-i、cd54、cd44和b7h1表面表达水平。igg2同种型对照抗体用作对照(图6n)。使用碘化丙锭染色和流式细胞计量分析来测定未处理oscsc和经nk细胞上清液分化oscsc的细胞死亡(图6o)。用il-2(1000u/ml)处理高度纯化nk细胞,并且用于在4小时51cr释放测定中测定针对未处理oscsc和经nk上清液分化oscsc的细胞毒性。使用图2e中所述的方法来测定溶解单位30/106个细胞(图6p)。
图7包括11个图版,标识为图版a至k,其显示来自癌症患者的纯化nk细胞内的小部分污染性t细胞更快扩增,并且可能由于nk细胞功能降低而排挤nk细胞。如图1a中所述来处理来自健康供者和胰腺癌患者的经新鲜纯化nk细胞,并且与单核细胞源性同种异体(来自不同健康供者)破骨细胞共培养。使用fitc和pe缀合抗体以及流式细胞计量术来分析来自癌症患者(图7a)和健康供者(图7b)的在第6、10、13、17、21、24、28、32和36天时的共培养物的含1x104个淋巴细胞的样品中cd3、cd16和cd56的表面表达。在6、10、13、17、21、24、28和32天共培养之后,使用显微镜来对经扩增淋巴细胞进行手动计数(图7c)。如图7c中所提及来对细胞进行计数,并且基于图7a和图7b中分析的表面表达来调整以测定在每天时的t/nkt细胞(图7d)和nk细胞数目(图7e)。使用针对oscsc的标准4小时51cr释放测定来测定共培养18-20小时、13、20和32天的淋巴细胞的细胞毒性。使用图3f(g)中所述的方法来测定溶解单位30/106个细胞。基于图7a和图7b中分析的表面表达来调整来自图7f的溶解单位以测定由1个nk细胞介导的针对oscsc的细胞毒性(图7h)。从过夜,6、10、13、17、21、24、28和32天共培养物收集上清液,并且使用单一elisa来测定ifn-γ(图7i)、il-10(图7j)和il-6(图7k)分泌。
图8显示来自hu-blt小鼠的脾细胞的cd3t细胞经消减淋巴细胞的表型。通过在6-8周龄免疫损害nod.cb17-prkdcscid/j和nod.cg-prkdcscidil2rgtm1wjl/szj(nsg)小鼠的肾囊下手术植入人胎儿肝和胸腺组织来产生人源化blt(hu-blt;人骨髓/肝/胸腺)小鼠。在组织移植后4-6周,对小鼠进行亚致死照射,并且用从胎儿肝分离的cd34+细胞进行静脉内注射以支持对人骨髓的完全重构。在用cd34+细胞注射之后8-12周,使用血液试样来分析对人免疫系统的重构。在这个实验结束时,通过用抗人cd45、cd3、cd4和cd8抗体染色脾细胞和骨髓细胞来确认人免疫细胞的移入,并且通过流式细胞计量术来分析(数据未显示)。对经成功重构hu-blt小鼠(t细胞的水平和谱系类似于健康供者)用1x106个人oscsc向口底中进行原位注射。再持续3-4周监测疾病进展和重量减轻。将动物处死,从处死动物收集脾,并且如材料和方法中所述来获得单细胞混悬液。使用人cd3t正性选择试剂盒来将cd3t细胞分选出来。在用相应pe缀合抗体、pe-cy5缀合抗体和fitc缀合抗体染色之后,分析流穿细胞(cd3阴性细胞)的人cd3、cd16、cd56、cd45、cd19、cd14表面表达。同种型对照抗体用作对照。
图9包括11个图版,标识为图版a至k,其显示当相较于从健康hu-blt小鼠获得的那些时,来自携带肿瘤的人源化blt小鼠的消减了t细胞的经体外扩增淋巴细胞使t细胞扩增,并且含有较少nk细胞并介导较低细胞毒性。对经重构blt(t细胞的水平和谱系类似于健康供者)用1x106个人oscsc向口底中进行原位注射。再持续4-5周监测疾病进展和重量减轻。将小鼠处死,收集脾,并且如补充材料和方法中所述来获得单细胞混悬液。使用正性选择试剂盒来将cd3+t细胞分选出来,并且在用相应抗体染色之后分析流穿细胞的人cd3/cd16/cd56表面表达。同种型对照抗体用作对照(图9a)。用il-2(1000u/ml)和抗cd16mab(3μg/ml)的组合处理来自hu-blt小鼠的cd3阴性细胞(1x106个细胞/毫升)18小时,随后使它们与oc一起在saj2存在下在1:2:4比率(oc:nk:saj2)下培养。使用流式细胞计量分析来分析在第6、10、14、18和22天的cd3、cd16和cd56表面表达(图9b)。在6、10、18和22天共培养之后,使用显微术来对经扩增淋巴细胞进行手动计数(图9c)。使用总体扩增细胞内nk细胞和t/nkt细胞的百分比来测定nk细胞(图9d)和t/nkt(图9e)细胞的数目。使用针对oscsc的标准4小时51cr释放测定来测定共培养10和18天的nk细胞的细胞毒性,并且使用为使30%的oscsc溶解所需的nk细胞数的倒数x100来测定溶解单位30/106个细胞(图9f)。基于每个nk细胞针对oscsc达成的溶解来使溶解单位标准化和加以调整(图9g)。从在第6、10和13天的共培养物收集上清液,并且使用单一elisa来测定ifn-γ(图9h)、il-10(图9i)和il-6(图9j)分泌。通过心脏穿刺来从hu-blt小鼠进行死后外周血液收集,并且收集血清样品并使用多路阵列来分析ifn-γ、il-10和il-6分泌(图9k)。
图10包括三个图版,标识为图版a至c,其显示由原代nk细胞和经破骨细胞扩增nk细胞分泌的细胞因子、趋化因子和生长因子。从健康供者的外周血液单核细胞(pbmc)获得高度纯化nk细胞和单核细胞,并且用il-2(1000u/ml)处理nk细胞(1x106个细胞/毫升)18小时,随后收集上清液。为产生破骨细胞,在含有m-csf(25ng/ml)和rankl(25ng/ml)的α-mem培养基中培养单核细胞21天。对于扩增,用il-2(1000u/ml)和抗cd16mab(3μg/ml)的组合处理纯化nk细胞(1x106个细胞/毫升)18小时,随后使它们分别与自体破骨细胞在saj2细菌存在下在1:2:4比率(oc:nk:saj2)下共培养。在6天共培养之后收集上清液,并且多路测定用于测定由原代nk细胞和经扩增nk细胞分泌的细胞因子(图10a)、趋化因子(图10b)和生长因子(图10c)水平。
图11包括三个图版,标识为图版a至c,其显示阻断il-12、il-15或两者组合导致降低的nk细胞扩增、nk细胞介导的细胞毒性和细胞因子分泌。如图2a中所述来处理来自健康供者的经新鲜纯化nk细胞并与自体破骨细胞在存在以及不存在分别在100ng/ml和1μg/ml下的抗il12mab、抗il-15mab、或抗il-12mab和抗il-15mab的组合下共培养。每2天对共培养物补充以il-2(1000单位/ml)。在第6、8、12、14和20天,使用显微术来对nk细胞进行计数(图11a)。在第9天和第15天,来自各扩增样品的1x105个nk细胞用于针对oscsc的标准4小时51cr释放中。使用为使30%的oscsc溶解所需的nk细胞数的倒数x100来测定溶解单位30/106个细胞(图11b)。从在第8、12、15和20天的共培养物收集上清液,并且使用单一elisa来测定ifn-γ分泌(图11c)。
图12包括七个图版,标识为图版a至g,其显示添加抗cd3抗体会抑制t细胞扩增,并且使经oc扩增nk细胞增加。用oc使来自健康供者和癌症患者的经新鲜纯化nk细胞扩增27天,随后用rh-il-2和cd3抗体(1μg/ml)处理培养物,随后通过显微评估来测定在第29、31和35天的nk细胞(图12a)和t细胞(图12b)数目(健康供者与患者两者)。测定如图12a和图12b中所述来处理的健康细胞(图12c)和患者细胞(图12d)的前向散射和侧向散射损失。将细胞用pi染色,并且分析dna碎裂(图12e)。来自第31天培养物的淋巴细胞用于针对oscsc的标准4小时51cr释放中。使用为使30%的oscsc溶解所需的nk细胞数的倒数x100来测定溶解单位30/106个细胞(图12f)。从在第35天的共培养物收集上清液,并且使用单一elisa来测定ifn-γ分泌(图12g)。
图13显示在不存在nk细胞下用抗cd3mab处理的纯化t细胞不丧失前向散射和侧向散射。从健康供者的外周血液单核细胞(pbmc)获得高度纯化t细胞和单核细胞,并且用il-2(100u/ml)和cd3抗体(1μg/ml)处理t细胞(1x106个细胞/毫升)18小时,随后使它们分别与自体破骨细胞在saj2细菌存在下在1:2:4比率(oc:t细胞:saj2)下共培养。在培养之后第9天,分析细胞的cd3、cd16和cd56。在不存在nk细胞下,经cd3抗体处理t细胞不丧失前向散射和侧向散射。
图14包括八个图版,标识为图版a至h,其显示经破骨细胞活化nk细胞使cd8+t细胞数目实质上增加。依次使用pe和fitc缀合抗体染色和流式细胞计量分析来分析来自健康供者和癌症患者的pbmc的cd3、cd4和cd8表面表达(图14a)。如图2a中所述来处理来自健康供者和癌症患者的经新鲜纯化nk细胞,并且与oc共培养。使用cd3正性选择试剂盒来从健康供者和癌症患者的pbmc纯化t细胞,并且用rh-il2(100u/ml)以及抗cd3mab(1μg/ml)和抗cd28mab(1μg/ml)的组合活化t细胞18-20小时,随后使它们与oc在saj2存在下在1:2:4比率(oc:t细胞:saj2)下共培养。分析淋巴细胞上的cd3、cd4和cd8表面表达(图14b)。如图2a中所述来从人pbmc纯化单核细胞并产生oc和dc,并且共培养纯化nk细胞,并且使用显微测定来评估经扩增淋巴细胞的数目(图14c),使用图14c中总体扩增细胞内的如图2a中所述的nk细胞和t细胞百分比来测定t细胞(图14d)和nk细胞(图14e)的数目。对淋巴细胞分析cd3+cd4+细胞和cd3+cd8+细胞的表面表达,并且使用图版d中总体t细胞内cd4细胞和cd8细胞的百分比来测定cd3+cd4+t细胞(图14f)和cd3+cd8+t细胞(图14g)的数目。用针对cd45ro、cd62l、cd28、cd44、ccr7和cd127的抗体染色经oc活化nk扩增的t细胞、经dc活化nk扩增的t细胞、经oc扩增/活化t细胞和经dc扩增/活化t细胞,并且通过流式细胞计量术来分析。象限2中的数值表示cd3+t细胞内呈各抗体阳性的细胞的百分比(图14h)。
图15包括两个图版,标识为a和b,其显示经破骨细胞扩增nk细胞在冷冻之后保留它们的细胞因子分泌和细胞毒性功能。如图1a中所述来处理经新鲜纯化nk细胞并与单核细胞源性自体破骨细胞共培养,在培养之后第9天,使经扩增nk细胞冷冻。使nk细胞解冻,并且用rh-il-2(1000u/ml)处理,在培养之后第6天和第9天,收集上清液,并且使用单一elisa来测定ifn-γ分泌(图15a)。如图1a中所述来培养nk细胞,并且使用针对oscsc的标准4小时51cr释放测定来测定在培养之后第6和9天的淋巴细胞细胞毒性。使用图3f中所述的方法来测定溶解单位30/106个细胞(图15b)。
图16包括五个图版,标识为图版a-e,其显示从胰腺(图16b)、结肠(图16c)、口腔(图16d)和前列腺(图16e)癌症患者的外周血液获得的pbmc的数目降低。图16a显示健康受试者相对于患者的情况下的降低。
图17包括五个图版,标识为图版a-e,其显示根据从健康受试者(图17a)、胰腺(图17b)、结肠(图17c)、口腔(图17d)和前列腺(图17e)癌症患者的外周血液获得的pbmc,癌症患者的nk和cd14单核细胞的百分比增加,但t细胞和b细胞显著降低。
图18包括四个图版,标识为图版a-d,其显示相较于健康nk细胞,由患者nk细胞达成的nk细胞细胞毒性降低。
图19包括八个图版,标识为图版a-h,其显示来自患者的经破骨细胞扩增nk细胞以及细胞毒性和ifn-γ分泌。
图20显示在来自胰腺癌患者的非经破骨细胞扩增nk细胞的情况下的细胞因子分泌。
图21包括五个图版,标识为图版a-e,其显示来自健康受试者(图21a)、胰腺(图21b)、结肠(图21c)、口腔(图21d)和前列腺(图21e)癌症患者的经破骨细胞扩增t细胞的ifn-γ分泌。
图22包括六个图版,标识为图版a-f,其显示根据nk细胞相对于t细胞(图22a)、健康受试者(图22b)、胰腺(图22c)、结肠(图22d)、前列腺(图22e)和口腔(图22f)癌症患者,来自nk细胞、t细胞以及经破骨细胞扩增nk细胞和经破骨细胞扩增t细胞的ifn-γ分泌。
图23包括四个图版,标识为图版a-d,其显示来自健康受试者(图23a)、胰腺(图23b)、结肠(图23c)和前列腺(图23d)癌症患者的t细胞ifn-γ分泌。
图24包括六个图版,标识为图版a-f,其显示在累积nk细胞相对于t细胞的情况下,在第0至15天内测定的扩增t细胞的总数相对于扩增nk细胞的总数,所述扩增t细胞通过经由表面受体交联而活化的t细胞的作用来活化(图24a),来自健康受试者(图24b)、胰腺(图24c)、结肠(图24d)、口腔(图24e)和前列腺(图24f)癌症患者的nk细胞。
图25包括五个图版,标识为图版a-e,其显示相较于在第0天至第15天内测定的健康个体(图25a)能力,用以使t细胞扩增的胰腺(图25b)、结肠(图25c)和前列腺(图25d)癌症患者能力。
图26显示相对于健康个体,患者的血清中的细胞因子和趋化因子降低。
图27包括两个图版a-b,其显示相较于健康个体,来自胰腺(图27a)和结肠(图27b)癌症患者的pbmc中cd4t细胞和cd8t细胞的百分比。
图28包括五个图版a-e,其显示相对于健康个体(图28a),胰腺(图28b)、结肠(图28c)、口腔(图28d)和前列腺(图28e)癌症患者的pbmc内cd4t细胞/cd8t细胞的比率。
图29包括五个图版,其显示相对于胰腺(图29b)、结肠(图29c)、口腔(图29d)和前列腺(图29e)癌症患者,健康受试者(图29a)中cd4t细胞与cd8t细胞的比率。
图30包括两个图版a和b,其显示不与(图30a)以及与(图30b)破骨细胞一起培养cd8t细胞和cd4t细胞的影响。
图31显示nk细胞和cd8t细胞的ifn-γ分泌。
图32显示分别由破骨细胞和nk细胞促进的cd8t细胞扩增和cd4t细胞扩增。
图33显示经破骨细胞扩增nk细胞针对癌干细胞/未分化肿瘤的细胞毒性。
图34显示用破骨细胞来扩增nk细胞对nk细胞扩增的影响以及由经破骨细胞扩增细胞达成的nk细胞细胞毒性作用。
图35显示相较于经dc扩增nk细胞,经oc扩增nk细胞中的t细胞具有效应记忆表型。
图36显示相对于经dc扩增nk细胞,经oc扩增nk细胞中具有耗竭表型的t细胞的数目。
图37显示来自胰腺癌患者的经oc扩增nk细胞中的ifn-γ表达性nk细胞。
图38显示由经oc扩增t细胞分泌的cd8和nk特异性细胞因子、共刺激性配体、粒酶、穿孔素以及可溶性fas和fas配体的表格。
图39显示由来自经oc扩增nk细胞培养物的cd8+t细胞分泌的cd8相关细胞因子、趋化因子、共刺激性配体、sfas和fas配体以及粒酶和穿孔素的表格。
图40显示从经nk扩增或经oc扩增cd8细胞分泌的gm-csf、ifn-g、il-10、tnf-a、较低共刺激性配体scd137、粒酶、穿孔素、可溶性fas和fas配体的水平。
图41显示相较于不患有肿瘤的小鼠,来自经口腔肿瘤植入blt小鼠的nk细胞的细胞毒性活性。
图42显示在肿瘤植入之后,在用超动力nk细胞进行的免疫疗法之后,bm、脾和血液中的cd8+t细胞。
图43显示在肿瘤植入之后,在用超动力nk细胞在blt小鼠中进行的免疫疗法之后的血清ifn-γ、il-6、itac、gm-csf和il-8。
图44显示在不同扩增天数时,经破骨细胞扩增患者nk细胞的细胞毒性。
图45显示在不同扩增天数时,在患者的情况下经破骨细胞扩增nk细胞群体中nk细胞的数目。
图46显示相较于来自第15天至第25天的经dc扩增细胞,经破骨细胞扩增nk细胞的数目。
图47显示在不同扩增天数时,相较于经dc扩增nk细胞,经破骨细胞扩增nk细胞的数目。
图48显示在不同扩增天数时,相较于经破骨细胞扩增t细胞,经dc扩增t细胞的数目。
图49显示在不同扩增天数时,相较于经dc扩增nk细胞,经破骨细胞扩增nk细胞的细胞毒性。
图50显示在不同扩增天数时,相较于健康供者nk细胞,由原代非经破骨细胞扩增患者nk细胞和经破骨细胞扩增患者nk细胞达成的ifn-γ分泌。
图51显示在不同扩增天数时,相较于健康供者t细胞,由原代非经破骨细胞扩增患者t细胞和经破骨细胞扩增患者t细胞达成的ifn-γ分泌。
图52显示在不同扩增天数内,当相较于从健康供者nk细胞获得的那些时,由经破骨细胞扩增患者nk细胞达成的ifn-γ分泌。
图53显示在不同扩增天数时,当相较于从健康供者t细胞获得的那些时,在一些患者的情况下,由原代非经破骨细胞扩增患者t细胞和经破骨细胞扩增患者t细胞达成的ifn-γ分泌。
图54显示在不同扩增天数内,当相较于从健康供者t细胞获得的那些时,由原代非经破骨细胞扩增患者t细胞和经破骨细胞扩增患者t细胞(t细胞以正性方式加以选择)达成的ifn-γ分泌(来自所有扩增日的合并分泌)。
图55显示当相较于来自健康供者的经负性选择t细胞时,由经正性选择原代非经破骨细胞扩增患者t细胞和经破骨细胞扩增患者t细胞达成的ifn-γ分泌(来自所有扩增日的合并分泌)。
图56显示当相较于从健康供者获得的nk细胞时,由经破骨细胞扩增t细胞(t细胞以正性方式加以选择)达成的基于每个细胞的ifn-γ分泌。当相较于nk细胞时,用il-2活化的原代经正性选择t细胞分泌更高ifn-γ。
图57显示当相较于从健康供者获得的t细胞时,由原代非经破骨细胞扩增患者t细胞和经破骨细胞扩增患者t细胞(t细胞以正性方式加以选择)达成的基于每个细胞的ifn-g分泌。
图58显示由经破骨细胞扩增患者t细胞(t细胞以正性方式加以选择)和nk细胞达成的基于每个细胞的ifn-g分泌。当相较于用il-2处理的原代nk细胞时,原代非经破骨细胞扩增t细胞在il-2的情况下具有更高ifn-g分泌。
图59显示当相较于来自健康供者的经负性选择nk细胞或经负性选择t细胞时,由经正性选择原代非经破骨细胞扩增t细胞和经破骨细胞扩增t细胞达成的扩增细胞数目增加。
图60显示当相较于从健康供者获得的那些时,由经正性选择原代非经破骨细胞扩增患者t细胞和经破骨细胞扩增患者t细胞达成的扩增细胞数目降低。
图61显示来自胰腺患者血液的血清中细胞因子和趋化因子的水平。
图62包括11个图版,标识为图版a-c,其显示在用/不用aj2喂食下单次注射超动力nk细胞抑制hu-blt小鼠中的肿瘤生长。如材料和方法中所述以及图中所示来产生hu-blt小鼠(图62a)。对hu-blt和nsg小鼠用1x106个人oscsc向口底中进行原位植入,并且在7–10天之后,通过尾部静脉来用1.5x106个超动力nk细胞注射一组hu-blt小鼠,并且监测小鼠的疾病进展。在植入oscsc之前2周以及在植入肿瘤之后,在存在以及不存在nk注射下,每48小时用aj2益生细菌(50亿/天)对另一组hu-blt小鼠喂食直至实验终止(图62b)。基于每周通过将小鼠称重来监测重量减轻。3个代表性实验中的一者显示于这个图中(图62c)。在终止实验后,将小鼠处死,并且在切除之后拍摄肿瘤的照片(图62d),并称重(n=4)(图62e)。如图62b中所示来将小鼠用人oscsc植入并用nk细胞注射,并且用aj2喂食,并且切除肿瘤并进行死后称重(n=4)(图62f)。从hu-blt小鼠和人分离pbmc,并且依次使用抗体染色和流式细胞计量分析来测定cd45+免疫细胞内人cd3(n=5)(图62g)、cd4(n=5)(图62h)、cd8(n=5)(图62i)、cd19(n=3)(图62j)和cd16(n=5)(图62k)的表面表达。
图63包括九个图版,标识为a-i,其显示在用/不用aj2喂食下注射超动力nk细胞在携带肿瘤的hu-blt小鼠的情况下使血液、脾、bm中的nk细胞、增浓nk细胞和纯化cd3+t细胞的ifn-y分泌和细胞毒性功能恢复和增加。如图62b中所述来将hu-blt小鼠用人oscsc植入并用nk细胞注射,并且喂以aj2,并且在nk细胞注射之后一周,通过尾部静脉注射来用pd1抗体(50μg/小鼠)注射小鼠。在处死之后,收集脾(n=5)(图63a)、bm(n=5)(图63b)和外周血液(n=5)(图63c),从各组织制备单细胞混悬液,并且用il-2(1000u/ml)处理7天(对于脾和bm是1x106个细胞/毫升,并且对于pbmc是0.7x106个细胞/毫升)。从脾细胞分离富含nk的细胞,并且用il-2(1000u/ml)处理7天(1x106个细胞/毫升)(n=3)(图63d)。使用针对oscsc的标准4小时51cr释放测定进行细胞毒性测定,并且使用为使30%的oscsc溶解所需的细胞数的倒数x100来测定lu30/106个细胞。将脾细胞(n=5)(图63e)、bm细胞(n=5)(图63f)、pbmc(n=5)(图63g)(对于脾和bm在1x106个细胞/毫升下,并且对于pbmc在0.7x106个细胞/毫升下)各自用il-2(1000u/ml)处理,并且将来自脾细胞的经正性选择cd3+t细胞(n=4)在1x106个细胞/毫升下用il-2(100u/ml)处理(图63h)7天,此后收集上清液,并且使用特异性elisa来测定ifn-γ的水平。相对于从用单独oscsc注射的小鼠获得的那些,测定在来自各组小鼠的各组织的情况下的ifn-y分泌倍数变化(图63e-图63h)。
图64包括两个图版,标识为a和b,其显示在用/不用aj2喂食下单次注射超动力nk细胞使hu-blt小鼠中cd8+t细胞的数目增加。如图62b中所述来将hu-blt小鼠用oscsc植入并用nk细胞注射,并且用aj2喂食,并且依次使用抗体染色和流式细胞计量分析来测定bm细胞(n=3)(图64a)和脾细胞(n=3)(图64b)内人cd8+t细胞的百分比。
图65包括十个图版,标识为a-j,其显示在blt小鼠中在用/不用aj2喂食下单次注射超动力nk细胞介导体内肿瘤分化,使ifn-γ分泌增加并将增加数目的人免疫细胞动员至肿瘤,并且导致离体肿瘤生长降低。如图62b中所述来将hu-blt和nsg小鼠用oscsc植入并用nk细胞注射。在处死之后,收集口腔肿瘤,并且制备单细胞混悬液,并在第0天时培养来自各组的相同数目的细胞(在1x106个细胞/毫升下的总计3x106个细胞)。在第10天,移除上清液并对附着肿瘤细胞进行计数,并且对于后续培养,将各组中的数目调整至从经nk注射小鼠获得的那些,因为所述小鼠达成最小数目的肿瘤扩增。在第10、14、19和20天,测定各组中离体扩增肿瘤细胞的总数。若干代表性实验中的一者显示于这个图中(图65a)。如图62b中所述来将hu-blt小鼠用oscsc或在体外经nk分化的oscsc(分化oscsc)或用针对ifn-γ和tnf-α的抗体处理以阻断分化的经nk分化oscsc植入,随后在hu-blt小鼠中进行nk注射。如图65a中所述,在处死之后,使口腔肿瘤解离,并且制备单细胞并在(处于1x106个细胞/毫升下的总计3x106个细胞)下培养,并且测定扩增肿瘤细胞的数目。若干代表性实验中的一者显示于这个图中(图65b)。如图62b中所述来将hu-blt小鼠用oscsc或分化oscsc或用针对ifn-γ和tnf-α的抗体处理的分化oscsc植入,并且用nk细胞注射和/或用aj2喂食。如图65a中所述,在处死之后,收集口腔肿瘤并培养,并且测定扩增肿瘤细胞的数目(n=7)(图65c)。如图62a中所述来将hu-blt和nsg小鼠用oscsc植入,随后在hu-blt小鼠中进行nk注射。收集口腔肿瘤,并且制备单细胞混悬液。依次使用抗体染色和流式细胞计量分析来测定在培养的第12天时非附着细胞内浸润性hu-cd45c免疫细胞的百分比。三个代表性图中的一者显示为这个图(图65d)。如图4a中所述来培养来自hu-blt和nsg小鼠的口腔肿瘤并用il-2(1000u/ml)处理,并且在图中所示的天数时收集它们的上清液并使用elisa来测定ifn-g的水平。若干代表性图中的一者显示为这个图(图65e)。在用它们的相应抗体染色之后,使用流式细胞计量分析来评估来自hu-blt和nsg小鼠的口腔肿瘤培养物在第10天的人cd54和mhc-i表达。若干代表性实验中的一者显示于这个图中(图65f)。使来自健康人供者的外周血液的纯化nk细胞(1x106个细胞/毫升)未处理或用il-2(1000u/ml)处理18小时,随后将它们添加至从hu-blt小鼠的不同实验组的切除肿瘤加以培养的经51cr标记oscsc中,并且将此与在实验室中在各种效应物与靶标比率下维持的oscsc的培养物进行比较。使用标准4小时51cr释放测定来测定nk细胞介导的细胞毒性。如材料和方法中所述来测定lu30/106个细胞(n=4)(图65g和图65h)。用il-2(1000u/ml)处理如
图65a中所述来自hu-blt小鼠的口腔肿瘤细胞,并且在3和7天之后收集上清液并使用特异性elisa来测定vegf分泌水平。基于从单独经oscsc注射小鼠获得的量来计算由从经nk注射动物(n=6)获得的肿瘤达成的vegf分泌降低(图65i)。在用抗体染色之后,使用流式细胞计量分析来测定从hu-blt小鼠的不同实验组解离的口腔肿瘤内hu-cd45c免疫细胞的浸润性百分比。若干代表性实验中的一者显示于这个图中(图65j)。
图66包括三个图版,标识为a-c,其显示在携带肿瘤的小鼠中在用/不用aj2喂食下单次注射超动力nk细胞使从hu-blt小鼠的外周血液获得的血清内细胞因子、趋化因子和生长因子分泌恢复和增加。如实施例4中所述来获得来自外周血液的血清,并且进行多路阵列以测定ifn-γ的分泌,此处显示四个代表性图中的一者(图66a)。基于从对照hu-blt小鼠获得的值来测定ifn-γ的倍数变化(n=5)(图66b)。多路阵列用于测定从外周血液获得的血清中细胞因子、趋化因子和生长因子分泌(图66c)。
图67包括两个图版,标识为a和b,其显示cddp或伴有和不伴有nac的太平洋紫杉醇诱导用nk上清液分化的oscsc而非经不良分化肿瘤的显著细胞死亡。用il-2(1000u/ml)和抗cd16mab(3mg/ml)的组合处理高度纯化nk细胞18小时,此后在tnf-a抗体(1:100)和ifn-g抗体(1:100)存在下将nk上清液添加至oscsc中,持续5天的时期。此后,使oscsc脱离,并且用/不用顺铂处理18–24小时。接着使用pi染色和流式细胞计量分析来测定oscsc的活力。3个代表性实验中的一者显示于这个图中(图67a)。如图67a中所述来用来自nk细胞的上清液处理oscsc。之后,使肿瘤脱离,并且用/不用nac(20nm)处理24小时,随后用太平洋紫杉醇处理18–24小时。通过pi染色和流式细胞计量分析来测定oscsc活力。3个代表性实验中的一者显示于这个图中(图67b)。
图68包括五个图版,标识为a-e,其显示当相较于采用nk细胞的单独肿瘤植入小鼠的那些时,来自用nk细胞注射或仅用经nk分化oscsc肿瘤植入的携带肿瘤的小鼠的单核细胞或破骨细胞诱导来自自体或同种异体nk-肿瘤共培养物的显著更多ifn-g。如图62b中所述来将hu-blt小鼠用oscsc植入并用nk细胞注射,并且喂以aj2。在处死之后,如本文所述来分离来自脾细胞的nk细胞和来自bm细胞的单核细胞。使自体nk细胞未处理或用il-2(1000u/ml)与单核细胞(nk:单核细胞2:1)组合处理,并且在共培养之后第7天,nk细胞作为效应细胞用于针对oscsc的标准4小时51铬释放测定中。使用为使30%的靶标细胞溶解所需的nk细胞数的倒数x100来测定lu30/106个细胞(图68a)。使自体nk细胞未处理或用il-2(1000u/ml)或用il-2(1000u/ml)和lps(100ng/ml)的组合在不存在以及存在单核细胞(nk:单核细胞2:1)下处理7天,此后收集上清液,并且使用单一elisa来测定ifn-g分泌(图68b)。如实施例4中所述来从纯化hu-blt单核细胞产生oc。用il-2(1000u/ml)和抗cd16mab(3mg/ml)预处理来自健康人供者的纯化同种异体nk细胞18小时,接着使其与hu-bltoc一起在saj2存在下培养(nk:oc:saj22:1:4)。在培养之后,使用显微术来在第5、8、12和15天对培养物中nk细胞的数目进行计数(图68c)。在第5、8、12和15天从培养物收集上清液,并且使用单一elisa来测定ifn-g分泌(图68e)。使用来自图68c的细胞计数,测定在
具体实施方式
本发明部分地涉及一种在体外或离体使nk细胞活化的方法,其包括在包含破骨细胞(oc)的培养基中培养所述nk细胞。类似地,本文提供一种用以在体外或离体使t细胞活化的方法,其包括在包含树突细胞(dc)的培养基中培养所述t细胞。本发明进一步提供一种相对于t细胞,在体外或离体使nk细胞活化的方法,其包括在包含破骨细胞(oc)的培养基中培养所述nk细胞和所述t细胞。本发明进一步提供一种相对于nk细胞,在体外或离体使t细胞活化的方法,其包括在包含树突细胞(dc)的培养基中培养所述nk细胞和所述t细胞。所述经活化nk细胞可用于改进宿主免疫应答,并且可用于治疗疾病(例如癌症)。在一些方面,本发明提供一种通过破骨细胞(oc)来在体内使nk细胞活化,任选地,相对于t细胞,使nk细胞活化的方法。在一些实施方案中,可向受试者施用oc或oc培养上清液以治疗疾病(例如癌症)。在一些实施方案中,可添加益生细菌(例如saj2)以改进oc使nk细胞活化的功能。除破骨细胞(oc)和树突细胞(dc)之外,能够使nk细胞或t细胞活化的其他药剂也可添加至细胞培养物中或向受试者施用,包括任何基因、蛋白质、代谢物等。
i.定义
冠词“一个(种)(a/an)”在本文中用于指代冠词的一个或超过一个(即至少一个)语法对象。举例来说,“一个(种)”要素意指一个(种)要素或超过一个(种)要素。
术语“施用”意图包括允许药剂(例如至少一个破骨细胞(oc)或树突细胞(dc)、包含至少一个破骨细胞(oc)或树突细胞(dc)的细胞培养物、所述细胞培养物的上清液、包含所述oc或dc的任何组合物、至少一个益生细菌、包含所述益生细菌的任何组合物、能够使nk细胞和/或t细胞活化或有助于所述oc或dc和/或益生细菌的功能的其他药剂等,也包括本文所述的各种药剂的经处理(即经分离、经纯化、经浓缩、或在用于治疗用途或其他用途的其他过程之后)形式)执行它的预定功能的施用途径。可使用的用于身体治疗的施用途径的实例包括注射(皮下、静脉内、胃肠外、腹膜内、鞘内等)、口服、吸入和经皮途径。注射可为团式注射或可为连续输注。视施用途径而定,药剂可用所选物质包覆或安置在所选物质中以保护它免遭可不利地影响它执行它的预定功能的能力的天然条件。药剂可单独施用,或与药学上可接受的载体联合施用。药剂也可以前药形式施用,所述前药在体内转化成它的活性形式。
术语“活化(activating/activation)”是指使靶标的功能增强。举例来说,本公开提供一种在体外、离体和/或在体内使nk细胞或t细胞活化的方法,任选地,其中分别相对于t细胞或nk细胞,所述活化是优先的。在本公开中,对细胞的活化是指使所述细胞的功能增强,包括至少增强某一细胞类型(例如nk细胞或t细胞)的各细胞的活性和/或至少一种细胞功能(例如细胞毒性、细胞分裂和/或生长速率等),增强某一细胞类型的细胞数目(例如细胞扩增),或两者。在一些实施方案中,本文所用的药剂使至少一种细胞诸如nk细胞或t细胞活化。在其他实施方案中,相对于另一细胞类型(例如t细胞或nk细胞),本文所用的药剂优先活化一种细胞类型(例如nk细胞或t细胞)。
在本公开中,术语“增强”可与术语“增加”、“上调”、“改进”等互换使用,指代在数量方面对于药剂和/或靶标的功能有意义的任何增加。举例来说,当相比于在对照的情况下的原始数量和/或正常数量(例如正常受试者或不患有疾病或病症(例如癌症)的受试者中某一细胞类型的活性和/或细胞数目),在数量方面的增加以大于用于评估数量的测定的标准误差的数量增大,并且优选是相比于那个原始数量和/或正常数量,增大至少20%、30%、40%、50%、60%、70%、80%、90%、100%、150%、200%、300%、350%、400%、500%、600%、700%、800%、900%、1000%或更多时,某一细胞类型(例如nk细胞或t细胞)的活性和/或细胞数目的增强可为“显著的”。或者,如果相比于原始活性/数量和/或正常活性/数量,某一细胞类型的活性和/或细胞数目的增强是至少约两倍,并且优选是至少约三倍、四倍或五倍或更多倍,那么所述增强可被视为“显著的”。所述“显著性”也可应用于本文所述的任何其他测量参数,诸如表达、抑制、细胞毒性、细胞生长参数等。在一些实施方案中,某一细胞类型(例如nk细胞或t细胞)的活性和/或细胞数目的增强可不是如本文所述的“显著的”,但仍然是对于熟练技术人员来说足以了解它的生物相关性的增加。
除非本文内另外规定,否则术语“抗体”广泛涵盖天然存在形式的抗体(例如igg、iga、igm、ige)和重组抗体诸如单链抗体、嵌合和人源化抗体和多特异性抗体、以及所有前述各物的片段和衍生物,所述片段和衍生物具有至少抗原结合位点。抗体衍生物可包含缀合于抗体的蛋白质或化学部分。
如本文所用的术语“抗体”也包括抗体的“抗原结合部分”(或简称为“抗体部分”)。如本文所用的术语“抗原结合部分”是指抗体的一种或多种保留特异性结合抗原(例如生物标志物多肽或其片段)的能力的片段。已显示抗体的抗原结合功能可通过全长抗体的片段来执行。涵盖在抗体的术语“抗原结合部分”内的结合片段的实例包括(i)fab片段,即由vl、vh、cl和ch1结构域组成的单价片段;(ii)f(ab')2片段,即包含由二硫桥在铰链区处连接的两个fab片段的二价片段;(iii)由vh和ch1结构域组成的fd片段;(iv)由抗体的单臂的vl和vh结构域组成的fv片段,(v)dab片段(ward等,(1989)nature341:544-546),其由vh结构域组成;和(vi)经分离互补决定区(cdr)。此外,尽管fv片段的两个结构域vl和vh由单独基因编码,但可使用重组方法由使得它们能够成为其中vl区和vh区配对以形成单价多肽的单一蛋白质链(称为单链fv(scfv);参见例如bird等(1988)science242:423-426;以及huston等(1988)proc.natl.acad.sci.usa85:5879-5883;以及osbourn等1998,naturebiotechnology16:778)的合成接头来使它们接合。所述单链抗体也意图涵盖在抗体的术语“抗原结合部分”内。可使特定scfv的任何vh和vl序列连接于人免疫球蛋白恒定区cdna或基因组序列以产生编码完整igg多肽或其他同种型的表达载体。vh和vl也可用于使用蛋白质化学或重组dna技术来产生fab、fv或其他免疫球蛋白片段。也涵盖其他形式的单链抗体,诸如微型双功能抗体。微型双功能抗体是二价双特异性抗体,其中vh结构域和vl结构域在单一多肽链上表达,但使用太短以致不允许在同一链上的两个结构域之间配对的接头,由此迫使结构域与另一链的互补结构域配对并产生两个抗原结合位点(参见例如holliger,p.等(1993)proc.natl.acad.sci.usa90:6444-6448;poljak,r.j.等(1994)structure2:1121-1123)。
抗体可为多克隆或单克隆抗体;异种异体、同种异体或同种同基因抗体;或其经修饰形式(例如人源化抗体、嵌合抗体等)。抗体也可为完全人抗体。优选地,本发明的抗体特异性或大致上特异性结合生物标志物多肽或其片段。如本文所用的术语“单克隆抗体”和“单克隆抗体组合物”是指仅含有能够与抗原的特定表位起免疫反应的一个种类的抗原结合位点的抗体多肽的群体,而术语“多克隆抗体”和“多克隆抗体组合物”是指含有能够与特定抗原相互作用的多个种类的抗原结合位点的抗体多肽的群体。单克隆抗体组合物通常显示对它与其起免疫反应的特定抗原的单一结合亲和力。
术语“癌症”或“肿瘤”或“过度增生”是指存在具有致癌细胞典型特征的细胞,所述特征诸如增殖不受控制、永生性、转移潜力、快速生长和增殖速率以及某些特征性形态特征。除非另外陈述,否则所述术语包括化生。在一些实施方案中,所述特征包括以下中的至少一者:使宿主免疫应答沉默、使宿主免疫应答降低和/或避免宿主免疫应答,和/或对宿主细胞(例如nk细胞)溶解和/或分化具有抗性。在一些实施方案中,所述致癌细胞是癌干细胞(例如口腔鳞状癌干细胞(oscsc))。在一些实施方案中,所述细胞部分地或完全地由于至少一个遗传突变而展现所述特征。癌细胞经常呈肿瘤形式,但所述细胞可单独存在于动物内,或可为非致肿瘤性癌细胞诸如白血病细胞。如本文所用,术语“癌症”包括恶变前癌症以及恶性癌症。癌症包括但不限于b细胞癌例如多发性骨髓瘤、瓦尔登斯特伦氏巨球蛋白血症(
在一些实施方案中,癌症是“三重阴性乳腺癌”或“tnbc”,其是指呈雌激素受体(er)阴性、孕酮受体(pr)阴性和人表皮生长因子受体2(her-2)阴性的乳腺癌(pegram等(1998)j.clin.oncol.16:2659-2671;wiggans等(1979)cancerchemother.pharmacol.3:45-48;carey等(2007)clin.cancerres.13:2329-2334)。
在某些实施方案中,癌症是“pi3kβ依赖性癌症”,其可指在功能上依赖于pi3kβ的癌症。举例来说,即使肿瘤组织中的pi3kβ(例如pi3kβmrna、pi3kβ蛋白、新合成的pi3kβ蛋白等)的表达水平与它在正常组织中的表达水平类似,但如果诸如通过使用rnai或任何其他手段来直接或间接抑制pi3kβmrna和/或蛋白质,或缺失pi3kβ基因(例如通过剔除或成簇规律间隔短回文重复序列(crispr)技术)导致抑制肿瘤发生、肿瘤细胞增殖、肿瘤转移或诱导肿瘤细胞分化,那么癌症也是pi3kβ依赖性的。术语“pi3kβ依赖性癌症”也指其中相比于在与pi3kβ依赖性癌症具有相同细胞类型的非癌性细胞中表达的pi3kβ的正常量,pi3kβ在显著更高水平下表达(例如pi3kβmrna、pi3kβ蛋白、新合成的pi3kβ蛋白等)的癌症。
如本文所用的术语“微转移”优选定义为在最大宽度方面测量为大于0.2mm和/或具有大于200个细胞直至2mm的一组汇合癌细胞。更优选地,“微转移”定义为在最大宽度方面是0.2mm至2mm的一组汇合癌细胞(参见edge等(2010)ajcccancerstagingmanualandhandbook(第7版))。对“微转移”的一替代性优选定义是一组至少1000个汇合癌细胞,并且在最宽尺寸方面是至少0.1mm直至在最宽尺寸方面是1mm。在标准对比mri成像或其他临床成像技术的情况下,微转移通常不可见。然而,在某些癌症的情况下,针对肿瘤选择性抗原(例如乳腺癌转移的her2)的放射性抗体允许使微转移显像。其他间接检测方法包括由于vegf诱导的血管渗漏而在脑微转移部位处发生的造影介质渗漏(yano等(2000)cancerres.60:4959-49067;美国专利公布2015/0352113)。更灵敏成像技术也可应用于检测微转移。举例来说,血容量可通过使用替代性造影剂uspio(moldayiron,biopal,worcester,mass.)进行mri来成像以检测微转移(yin等(2009)clin.exp.metastasis.26:403-414)。
术语“对照”是指适于提供与测试样品中的表达产物、细胞数目和/或细胞功能的比较的任何参照标准。在某些实施方案中,对照包括获得“对照样品”,从所述对照样品检测表达产物水平、细胞数目和/或细胞功能,并且与来自测试样品的表达产物水平、细胞数目和/或细胞功能进行比较。这种对照样品可包括任何适合样品,包括但不限于来自具有已知结果的对照癌症患者的样品(可为储存样品或先前样品测量结果);从受试者诸如正常患者或癌症患者分离的正常组织或细胞、从受试者诸如正常受试者或癌症患者分离的经培养原代细胞/组织、从癌症患者的相同器官或身体位置获得的邻近正常细胞/组织、从正常受试者分离的组织或细胞样品、或从寄存处获得的原代细胞/组织。在其他优选实施方案中,对照可包括来自包括但不限于持家基因的任何适合来源的参照标准表达产物水平、来自正常组织(或其他先前分析的对照样品)的表达产物水平范围、来自一组患者或一组具有某一结果(例如存活1年、2年、3年、4年等)或接受某一治疗(例如护理标准癌症疗法)的患者的先前在测试样品内测定的表达产物水平范围。本领域技术人员应了解所述对照样品和参照标准表达产物水平、细胞数目和/或细胞功能可以组合方式作为对照用于本发明方法中。在某些实施方案中,对照可包括正常或非癌性细胞/组织样品。在其他优选实施方案中,对照可包括一组患者,诸如一组癌症患者,或一组接受某一治疗的癌症患者,或一组相对于另一结果,具有一种结果的患者的表达水平、某一细胞类型(例如nk细胞或t细胞)的数目、和/或某一细胞类型的细胞功能。在前述情况下,各患者的特定表达产物水平、细胞数目和/或细胞功能可被指定为百分位表达水平、细胞数目和/或细胞功能,或表示为高于或低于参照标准表达水平、细胞数目和/或细胞功能的平均值或均值。在其他优选实施方案中,对照可包括正常细胞、来自用组合化学疗法治疗的患者的细胞、以及来自患有良性癌症的患者的细胞。在其他实施方案中,对照也可包括测量值,例如相较于群体中持家基因的表达水平或另一细胞类型,同一群体中特定基因的平均表达水平、特定细胞类型(例如nk细胞或t细胞)的平均细胞数目和/或细胞功能。这种群体可包含正常受试者、尚未经受任何治疗(即治疗原初性)的癌症患者、经受护理标准疗法的癌症患者、或患有良性癌症的患者。如由以下数据所显示,在将测试样品中的表达产物水平、细胞数目和/或细胞功能与对照进行比较时,本发明方法不限于使用特定截断点。
术语“确定适于受试者的治疗方案”用于意指确定用于受试者的治疗方案(即单药疗法或用于预防和/或治疗受试者的癌症的不同疗法的组合),所述治疗方案基于或基本上基于或至少部分基于根据本发明的分析的结果而起始、修改和/或终止。一个实例是在手术之后起始辅助疗法,其目的在于使复发风险降低,另一实例将在于修改特定化学疗法的剂量。除根据本发明的分析的结果之外,确定可基于待治疗的受试者的个人特征。在大多数情况下,对适于受试者的治疗方案的实际确定将由主治医师或医生进行。
术语“诊断癌症”包括使用本发明的方法、系统和规程来确定个体中存在或不存在癌症或其亚型。所述术语也包括用于评估个体中的疾病活动性水平的方法、系统和规程。
术语“免疫细胞”是指在免疫应答中起作用的细胞。免疫细胞具有造血起源,并且包括淋巴细胞,诸如b细胞和t细胞;天然杀伤细胞;骨髓细胞,诸如单核细胞、巨噬细胞、嗜酸性粒细胞、肥大细胞、嗜碱性粒细胞和粒细胞。
术语“细胞因子”是指在细胞信号传导中重要的一个广泛和宽松种类的小蛋白质(约5–20kda)。它们的释放对在它们周围的细胞的行为具有影响。细胞因子作为免疫调节剂涉及于自分泌信号传导、旁分泌信号传导和内分泌信号传导中。细胞因子包括趋化因子、干扰素、白介素、淋巴因子和肿瘤坏死因子,并且可另外包括本公开中的激素或生长因子。细胞因子由广泛范围的细胞产生,所述细胞包括免疫细胞如巨噬细胞、b淋巴细胞、t淋巴细胞和肥大细胞,以及内皮细胞、纤维母细胞和各种基质细胞。优选细胞因子例示于本公开的说明书和附图中,例如于表1中。
术语“细胞因子/趋化因子活性”包括细胞因子或趋化因子能够调节至少一种细胞功能。通常,细胞因子或趋化因子调节体液免疫应答与基于细胞的免疫应答之间的平衡,并且它们调控特定细胞群体的成熟、生长和应答性。因此,术语“细胞因子/趋化因子活性”包括细胞因子或趋化因子能够结合它的天然细胞受体,能够调节细胞信号,以及能够调节免疫应答。
术语“免疫应答”包括t细胞介导和/或b细胞介导的免疫应答。示例性免疫应答包括t细胞应答,例如细胞因子产生和细胞性细胞毒性。此外,术语免疫应答包括间接受t细胞活化影响的免疫应答,例如抗体产生(体液应答)和细胞因子应答性细胞例如巨噬细胞的活化。
术语“免疫治疗剂”可包括可刺激宿主免疫系统以产生对受试者的肿瘤或癌症的免疫应答的任何分子、肽、抗体或其他药剂。各种免疫治疗剂适用于本文所述的组合物和方法中。
术语“抑制”包括降低、限制或阻断例如特定作用、功能或相互作用。在一些实施方案中,如果癌症的至少一种症状被缓和、终止、减缓或预防,那么癌症被“抑制”。如本文所用,如果癌症的复发或转移被减轻、减缓、延迟或预防,那么癌症也被“抑制”。类似地,如果相较于参照状态诸如对照如野生型状态,生物功能诸如蛋白质的功能被降低,那么它被抑制。举例来说,如果相较于野生型pi3激酶和/或未与抑制剂接触的pi3激酶,激酶活性由于突变和/或与抑制剂接触而被降低,那么突变pi3激酶或与pi3激酶抑制剂接触的pi3激酶的激酶活性被抑制。所述抑制可诸如通过在特定时间和/或地点施加药剂来诱导,或诸如由于可遗传突变而可为组成型的。所述抑制也可为部分的或完全的(例如相较于参照状态诸如对照如野生型状态,基本上无可测量活性)。基本上完全抑制被称为阻断。
术语“相互作用”在涉及两个分子之间的相互作用时是指分子彼此的物理接触(例如结合)。通常,这种相互作用导致所述分子中的一者或两者具有活性(其产生生物作用)。
“试剂盒”是包括至少一种用于特异性检测和/或影响本发明的标志物的表达的试剂例如探针或小分子的任何制品(例如包装或容器)。试剂盒可以用于进行本发明方法的单元形式推销、销售或售卖。试剂盒可包括一种或多种为使适用于本发明方法中的组合物表达所必需的试剂。在某些实施方案中,试剂盒可进一步包括参照标准,例如编码不影响或调控控制细胞生长、分裂、迁移、存活或凋亡的信号传导路径的蛋白质的核酸。本领域技术人员可设想许多所述对照蛋白质,包括但不限于常见分子标签(例如绿色荧光蛋白和β-半乳糖苷酶)、未由geneontology参考数据库分类在涵盖细胞生长、分裂、迁移、存活或凋亡的任何路径中的蛋白质、或遍在持家蛋白。试剂盒中的试剂可提供于个别容器中,或以两种或更多种试剂的混合物形式提供于单一容器中。此外,可包括描述试剂盒内组合物的使用的指导材料。
术语“新辅助疗法”是指在初级治疗之前给与的治疗。新辅助疗法的实例可包括化学疗法、放射疗法和激素疗法。举例来说,在治疗乳腺癌时,新辅助疗法可允许患有大型乳腺癌的患者经受保乳手术。
生物标志物的“正常”表达和/或活性水平是未受癌症折磨的受试者例如人患者的细胞中的所述生物标志物的表达和/或活性水平。生物标志物的“过度表达”或“显著较高水平表达”是指相比于对照样品(例如来自未患有生物标志物相关疾病的健康受试者的样品)中的所述生物标志物的表达活性或水平,并且优选是相比于若干对照样品中的所述生物标志物的平均表达水平,测试样品中的表达水平增大超过用于评估表达的测定的标准误差,并且优选是相比于所述对照表达活性或水平,增大至少10%,并且更优选是所述对照表达活性或水平的1.2、1.3、1.4、1.5、1.6、1.7、1.8、1.9、2.0、2.1、2.1、2.2、2.3、2.4、2.5、2.6、2.7、2.8、2.9、3、3.5、4、4.5、5、5.5、6、6.5、7、7.5、8、8.5、9、9.5、10、10.5、11、12、13、14、15、16、17、18、19、20倍高或更多。生物标志物的“显著较低水平表达”是指相比于对照样品(例如来自未患有生物标志物相关疾病的健康受试者的样品)中的所述生物标志物的表达水平,并且优选是相比于若干对照样品中的所述生物标志物的平均表达水平,测试样品中的表达水平降低至少10%,并且更优选是所述对照表达水平的1/1.2、1/1.3、1/1.4、1/1.5、1/1.6、1/1.7、1/1.8、1/1.9、1/2.0、1/2.1、1/2.1、1/2.2、1/2.3、1/2.4、1/2.5、1/2.6、1/2.7、1/2.8、1/2.9、1/3、1/3.5、1/4、1/4.5、1/5、1/5.5、1/6、1/6.5、1/7、1/7.5、1/8、1/8.5、1/9、1/9.5、1/10、1/10.5、1/11、1/12、1/13、1/14、1/15、1/16、1/17、1/18、1/19、1/20低或更大降低。可进行相同测定以确定过度活性或活性不足。
nk细胞
天然杀伤细胞或nk细胞是对先天性免疫系统至关重要的一种细胞毒性淋巴细胞类型。nk细胞所起的作用类似于细胞毒性t细胞在脊椎动物适应性免疫应答中的作用。nk细胞提供对受病毒感染细胞的快速应答,在感染之后约3天时起作用,并且对肿瘤形成起应答。通常,免疫细胞检测在受感染细胞表面上呈现的主要组织相容性复合物(mhc),从而触发细胞因子释放,导致溶解或凋亡。然而,nk细胞是独特的,因为它们能够在不存在抗体和mhc下识别应激化细胞,从而允许达成快得多的免疫反应。它们由于以下初始见解而被命名为“天然杀手”:它们不需要活化来杀灭缺失“自身”1类mhc标志物的细胞。这个作用是尤其重要的,因为缺失mhci标志物的有害细胞不能由其他免疫细胞诸如t淋巴细胞检测和破坏。
nk细胞(属于先天性淋巴样细胞的群组)被定义为大颗粒淋巴细胞(lgl),并且构成第三种类的从产生b淋巴细胞和t淋巴细胞的共同淋巴祖细胞分化的细胞。已知的是nk细胞在骨髓、淋巴结、脾、扁桃体和胸腺中分化和成熟,在其中它们接着进入循环中。nk细胞在表型、起源和相应效应物功能方面不同于天然杀伤t细胞(nkt);经常,nkt细胞活性通过分泌ifnγ来促进nk细胞活性。与nkt细胞相比,nk细胞不表达t细胞抗原受体(tcr)或全t标志物cd3或表面免疫球蛋白(ig)b细胞受体,但它们通常在人中表达表面标志物cd16(fcγriii)和cd56,在c57bl/6小鼠中表达nk1.1或nk1.2。目前,nkp46细胞表面标志物构成在人、若干小鼠品系(包括balb/c小鼠)和三个常见猴物种中表达的另一偏好nk细胞标志物。
nk细胞由i类主要组织相容性复合物(mhc)特异性抑制性受体负性调控(karre等,1986;ohlen等,1989)。这些特异性受体结合存在于其他细胞上的i类mhc分子或hla的多态性决定簇,并且抑制nk细胞溶解。在人中,被称为杀伤ig样受体(kir)的受体的家族的某些成员识别各组i类hla等位基因。
kir是存在于包括nk细胞的某些淋巴细胞子组上的一大家族受体。对kir的命名基于细胞外结构域的数目(kir2d或kir3d),以及细胞质尾部是长(kir2dl或kir3dl)还是短(kir2ds或kir3ds)。在人内,在存在于单一个体中的nk群体内,从一个nk细胞至另一nk细胞,存在或不存在给定kir是可变化的。在人群体内,也存在kir分子的相对高水平的多态性,其中某些kir分子存在于一些而非所有个体中。某些kir基因产物在结合于适当配体时导致刺激淋巴细胞活性。经确认刺激性kir全都具有短细胞质尾部,伴有与具有免疫刺激性基序(itam)的衔接分子缔合的带电荷跨膜残基。其他kir基因产物在性质上是抑制性的。
益生细菌
在一些实施方案中,本发明涉及一种能够调控nk细胞功能的包含至少一种益生细菌菌株的组合物。所述益生细菌诱导经活化nk细胞中的显著分割无反应性,从而导致显著诱导ifn-γ和tnf-α。此外,所述益生细菌诱导nk细胞的显著扩增。
许多商业益生菌是可用的,具有使胃肠不适减轻或使免疫系统强化的各种作用。用于本文所述的组合物和方法中的优选益生细菌物种包括那些可商购获得的益生细菌菌株(诸如saj2细菌),尤其是来自链球菌属(streptococcus)(例如嗜热链球菌(s.thermophiles))、双歧杆菌属(bifidobacterium)(例如长双歧杆菌(b.longum)、短双歧杆菌(b.breve)、婴儿双歧杆菌(b.infantis)、短双歧杆菌、婴儿双歧杆菌)和乳酸杆菌属(lactobacillus)(例如嗜酸乳酸杆菌(l.acidophilus)、瑞士乳酸杆菌(l.helveticus)、保加利亚乳酸杆菌(l.bulgaricus)、鼠李糖乳酸杆菌(l.rhamnosus)、植物乳酸杆菌(l.plantarum)和干酪乳酸杆菌(l.casei))的那些。本公开包括向受试者优选是哺乳动物(例如人)施用至少一种益生细菌菌株优选是两种或更多种不同细菌菌株的组合的方法。所述施用可全身或局部(例如直接向肠)进行。一优选施用途径是口服施用。也可使用其他途径(例如经直肠)。对于施用,可施用细菌(例如呈潮湿、经声波处理、经研磨或干燥形式或呈制剂形式)、含有细菌的细菌培养基、或细菌培养基上清液(不含有细菌)。
破骨细胞
破骨细胞是一种源于造血干细胞的骨细胞类型。它们的对骨组织进行再吸收的功能对骨的维持、修复和重塑至关重要。当在骨母细胞骨形成与破骨细胞骨再吸收之间存在平衡时,实现骨体内平衡。破骨细胞通过来自表达rankl的骨母细胞的刺激和它们的由通过icam-1进行的坚固粘附介导的相互作用来成熟。破骨细胞也表达存在于经活化nk细胞上的受体的许多配体。他们报道破骨细胞表达ulbp-1、ulbp-2/5/6和ulbp-3,但表达少许或不表达nk细胞的活化性受体nkg2d的mic-a、mic-b或i类mhc样配体。
相较于树突细胞(dc)和单核细胞,破骨细胞(oc)是nk细胞扩增和功能的显著活化物(tseng等(2015)oncotarget6(24):20002-25)。另外,破骨细胞分泌大量已知会使nk细胞活化的il-12、il-15、ifn-γ和il-18;破骨细胞也表达重要nk活化性配体。本公开提供一种关于如何使高度功能性超动力经破骨细胞扩增nk细胞扩增以致水平显著高于通过其他方法确立的水平的新型策略。若干体外nk扩增技术已被开发来确立较高治疗性细胞剂量,同时使nk细胞的活性和体内增殖潜力加强。这些技术中的一些包括刺激外周血液单核细胞(pbmc),从pbmc纯化nk细胞群体,或使用人脐带血,有时与各种饲养细胞诸如表达膜结合型il-15和41bb配体的k562细胞(k562-mb15-41bbl)、ebv-tm-lcl、韦尔姆斯氏肿瘤或经照射pbmc组合。这些研究已产生具有良好功能的临床相关nk细胞数目。
树突细胞
树突细胞(dc)是哺乳动物免疫系统的抗原递呈细胞(也称为辅助细胞)。它们的主要功能在于加工抗原物质,并且将它在细胞表面上向免疫系统的t细胞递呈。它们充当先天性免疫系统与适应性免疫系统之间的信使。
树突细胞存在于与外部环境接触的那些组织诸如皮肤(在其中存在专门化树突细胞类型,称为朗格汉斯(langerhans)细胞)以及鼻、肺、胃和肠的内衬中。它们也可以不成熟状态见于血液中。一旦活化,它们即向淋巴结迁移,在淋巴结中,它们与t细胞和b细胞相互作用以引发和塑造适应性免疫应答。在某些发育阶段,它们产生分支突出部分,即对细胞给与它的名称的树突。尽管在外观上类似,但这些是不同于神经元的树突的结构。不成熟树突细胞也称为面纱细胞,因为它们具有大型细胞质“面纱”而非树突。
本公开提供一种用以使用破骨细胞来使nk细胞活化,从而导致肿瘤靶标细胞对nk细胞介导的凋亡的敏感性增强以及细胞因子产生增强的新型方法。术语“活化(activate/activation/activating)”是指单独或以组合方式使nk细胞的扩增增强和/或使nk细胞功能活化。术语“nk细胞功能”是指nk细胞的任何功能,诸如细胞毒性和/或细胞因子/趋化因子产生/分泌活性。
术语“预防(prevent/preventing/prevention)”、“防治性治疗”等是指使不患有疾病、病症或疾患,但处于显现所述疾病、病症或疾患的风险下或容易显现所述疾病、病症或疾患的受试者中显现疾病、病症或疾患的概率降低。
术语“对抗癌疗法的响应”或“对用单独或与其他nk免疫疗法组合的包含至少一种益生细菌的组合物进行的疗法的响应”涉及过度增生性病症(例如癌症)对抗癌剂诸如用单独或与其他nk免疫疗法组合的包含至少一种益生细菌的组合物进行的治疗的任何响应,优选涉及在启动新辅助疗法或辅助疗法之后肿瘤质量和/或体积的变化。可评估过度增生性病症响应例如以了解功效,或可评估在新辅助或辅助情况下的过度增生性病症响应,其中可将在全身性干预之后的肿瘤尺寸与初始尺寸和大小进行比较,所述尺寸和大小如通过ct、pet、乳房摄影术、超声或触诊所测量。响应也可通过在活检或手术切除之后对肿瘤进行测径规测量或病理性检查来评估。响应可以定量方式如肿瘤体积变化百分比记录,或以定性方式如“病理性完全响应”(pcr)、“临床完全缓解”(ccr)、“临床部分缓解”(cpr)、“临床稳定疾病”(csd)、“临床进行性疾病”(cpd)或其他定性准则记录。过度增生性病症响应评估可在开始新辅助或辅助疗法之后早期进行,例如在数小时、数天、数周之后或优选在数月之后。响应评估的一典型终点是在终止新辅助化学疗法后或在手术移除残余肿瘤细胞和/或肿瘤床后。这通常在启动新辅助疗法之后3个月。在一些实施方案中,本文所述的治疗性治疗的临床功效可通过测量临床益处率(cbr)来测定。临床益处率通过在疗法结束后至少6个月的时间点测定处于完全缓解(cr)的患者的百分比、处于部分缓解(pr)的患者的数目百分比和患有稳定疾病(sd)的患者的数目百分比的总和来测量。这个公式的简写形式是cbr=超过6个月的cr+pr+sd。在一些实施方案中,特定癌症治疗方案的cbr是至少25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%或更大。用于评估对癌症疗法的响应的额外准则与“存活”相关,所述存活包括以下中的全部:直至死亡的存活期,也称为总体存活期(其中所述死亡可不管原因或与肿瘤相关);“无复发存活期”(其中术语复发应包括局部化复发与远端复发两者);无转移存活期;无疾病存活期(其中术语疾病应包括癌症和与其相关的疾病)。所述存活期的长度可通过参照确定起点(例如诊断或开始治疗的时间)和终点(例如死亡、复发或转移)来计算。此外,用于确定治疗功效的准则可被扩展来包括对化学疗法的响应、存活概率、在给定时期内的转移概率以及肿瘤复发概率。举例来说,为确定适当阈值,可向受试者群体施用特定癌症治疗方案,并且可使结果与在施用任何癌症疗法之前测定的生物标志物测量结果相关联。结局测量结果可为对在新辅助环境下给与的疗法的病理性响应。或者,结果量度诸如总体存活期和无疾病存活期可在已知其生物标志物测量值的癌症疗法之后历经一段时期对受试者加以监测。在某些实施方案中,施用剂量是本领域中已知的用于癌症治疗剂的标准剂量。监测受试者所持续的时期可变化。举例来说,可监测受试者至少2、4、6、8、10、12、14、16、18、20、25、30、35、40、45、50、55或60个月。与癌症疗法的结果相关联的生物标志物测量阈值可使用在本领域中的熟知方法诸如实施例章节中所述的那些来测定。
术语“抗性”是指癌症样品或哺乳动物对癌症疗法的获得性抗性或天然抗性(即对治疗性治疗是非响应性的,或对治疗性治疗具有降低或有限的响应),诸如具有的对治疗性治疗的响应降低25%或更多,例如30%、40%、50%、60%、70%、80%或更多,降低至1/2、1/3、1/4、1/5、1/10、1/15、1/20或更大降低。响应降低可通过与在获得抗性之前的相同癌症样品或哺乳动物进行比较,或通过与已知不具有对治疗性治疗的抗性的不同癌症样品或哺乳动物进行比较来测量。对化学疗法的典型获得性抗性被称为“多药物抗性”。多药物抗性可由p-糖蛋白介导,或可由其他机理介导,或它可在哺乳动物受多药物抗性微生物或微生物的组合感染时发生。确定对治疗性治疗的抗性在本领域中是常规的,并且属于普通熟练临床医师的技能,例如可如同“敏化”一样通过如本文所述的细胞增殖测定和细胞死亡测定来测量。在一些实施方案中,术语“使抗性逆转”意指使用第二药剂与初级癌症疗法(例如化学治疗或放射疗法)组合能够在其中单独所述初级癌症疗法(例如化学治疗或放射疗法)不能相较于未治疗肿瘤的肿瘤体积而产生肿瘤体积统计显著降低的情况下,在相较于未治疗肿瘤的肿瘤体积时,在统计显著水平(例如p<0.05)下产生肿瘤体积显著降低。这通常适用于在未治疗肿瘤以对数节奏生长时进行的肿瘤体积测量。
术语“响应”或“响应性”是指抗癌响应,诸如在使肿瘤尺寸降低或抑制肿瘤生长的意义上。所述术语也可指预后改进,例如如由达到复发的时间增加所反映,所述时间是达到第一复发的时期,删截作为第一事件的第二原发性癌症或无复发迹象的死亡;或总体存活期增加,所述总体存活期是从治疗至由于任何原因的死亡的时期。起响应或具有响应意指存在当暴露于刺激物时获得的有益终点。或者,在暴露于刺激物时,负面或有害症状得以最小化、缓和或减弱。应了解评估肿瘤或受试者将展现有利响应的可能性等效于评估所述肿瘤或受试者将不展现有利响应(也就是将展现缺乏响应或是非响应性的)的可能性。
用于检测或确定至少一种生物标志物的存在或水平的术语“样品”通常是脑组织、脑脊髓液、全血、血浆、血清、唾液、尿、粪便(例如屎)、泪和任何其他体液(例如如上在对“体液”的定义下所述),或组织样品(例如活检体)诸如小肠、结肠样品或手术切除组织。在某些情况下,本发明方法进一步包括在检测或确定至少一种标志物在样品中的存在或水平之前从个体获得样品。
术语“敏化”意指以允许用癌症疗法更有效治疗相关癌症的方式(例如通过用本文所述的组合物治疗)改变癌细胞或肿瘤细胞。在一些实施方案中,正常细胞未在导致正常细胞被过度损伤的程度上受到影响。对治疗性治疗的敏感性增加或敏感性降低根据用于特定治疗的本领域中已知方法和本文在以下所述的方法来测量,所述方法包括但不限于细胞增殖测定(tanigawa等(1982)cancerres.42:2159-2164)、细胞死亡测定(weisenthal等(1984)cancerres.94:161-173;weisenthal等(1985)cancertreat.rep.69:615-632;weisenthallm,kaspers等人编drugresistanceinleukemiaandlymphoma.langhorne,pa:harwoodacademicpublishers,1993:415-432;weisenthal等(1994)contrib.gynecol.obstet.19:82-90)。敏感性或抗性也可通过历经一段时期例如对于人是6个月以及对于小鼠是4-6周测量肿瘤尺寸降低来在动物中测量。如果相较于在不存在组合物或方法下的治疗敏感性或抗性,治疗敏感性增加或抗性降低是25%或更多,例如30%、40%、50%、60%、70%、80%或更多,治疗敏感性增加至2倍、3倍、4倍、5倍、10倍、15倍、20倍或更多,抗性降低至1/2、1/3、1/4、1/5、1/10、1/15、1/20或更大降低,那么所述组合物或方法使对治疗性治疗的响应敏化。确定对治疗性治疗的敏感性或抗性在本领域中是常规的,并且属于普通熟练临床医师的技能。应了解本文所述的用于增强癌症疗法的功效的任何方法都可同等应用于使过度增殖性或另外癌性细胞(例如抗性细胞)对癌症疗法敏感的方法。
术语“特异性结合”是指药剂诸如抗体结合预定靶标诸如抗原。通常,当通过使用作为分析物的目标抗原和作为配体的抗体,在
术语“协同作用”是指两种或更多种抗癌剂(例如用单独或与其他nk免疫疗法组合的包含至少一种益生细菌的组合物的组合进行的治疗)的组合作用可大于单独抗癌剂的单独作用的总和。
术语“受试者”是指任何健康动物、哺乳动物或人、或受癌症折磨的任何动物、哺乳动物或人,所述癌症例如脑转移、肺癌、卵巢癌、胰腺癌、肝癌、乳腺癌、前列腺癌、结肠癌、黑素瘤、多发性骨髓瘤等。术语“受试者”可与“患者”互换。
术语“存活”包括以下中的全部:直至死亡的存活期,也称为总体存活期(其中所述死亡可不管原因或与肿瘤相关);“无复发存活期”(其中术语复发应包括局部化复发与远端复发两者);无转移存活期;无疾病存活期(其中术语疾病应包括癌症和与其相关的疾病)。所述存活期的长度可通过参照确定起点(例如诊断或开始治疗的时间)和终点(例如死亡、复发或转移)来计算。此外,用于确定治疗功效的准则可被扩展来包括对化学疗法的响应、存活概率、在给定时期内的转移概率以及肿瘤复发概率。
术语“治疗作用”是指由药理活性物质在动物,特别是哺乳动物,并且更特别是人中导致的局部或全身作用。因此,所述术语意指意图用于在动物或人中诊断、治愈、缓和、治疗或预防疾病,或增强合乎需要的身体或心理发展和状况的任何物质。短语“治疗有效量”意指这种物质的在可适用于任何治疗的合理益处/风险比率下产生一定所需局部或全身作用的那个量。在某些实施方案中,化合物的治疗有效量将取决于它的治疗指数、溶解性等。举例来说,通过本发明方法发现的某些化合物可以足以产生可适用于所述治疗的合理益处/风险比率的量施用。
如本文所用,术语“不响应”包括癌细胞对疗法的抵抗性或治疗性细胞诸如免疫细胞对刺激例如通过活化性受体或细胞因子进行的刺激的抵抗性。不响应可例如由于暴露于免疫抑制剂或暴露于高剂量的抗原而发生。如本文所用,术语“无反应性”或“耐受性”包括对活化性受体介导的刺激的抵抗性。所述抵抗性通常具有抗原特异性,并且在已停止暴露于耐受抗原之后持续。举例来说,在t细胞的情况下的无反应性(与不响应相对比)的特征在于缺乏细胞因子产生,例如il-2。t细胞无反应性在t细胞暴露于抗原以及在不存在第二信号(共刺激性信号)下接收第一信号(t细胞受体或cd-3介导的信号)时发生。在这些条件下,使细胞重新暴露于相同抗原(即使重新暴露在共刺激性多肽存在下发生)导致不能产生细胞因子,并且因此导致不能增殖。然而,如果与细胞因子(例如il-2)一起培养,那么无反应性t细胞可增殖。举例来说,t细胞无反应性也可根据缺乏由t淋巴细胞产生il-2来观察,如通过elisa或通过使用指示细胞系进行的增殖测定来测量。或者,可使用报道体基因构建体。举例来说,无反应性t细胞未能引发在5’il-2基因增强子控制下由异源性启动子诱导,或由可见于增强子内的ap1序列的多聚体诱导的il-2基因转录(kang等(1992)science257:1134)。
ii.受试者
在某些实施方案中,适于本文公开的组合物和方法的受试者是哺乳动物(例如小鼠、大鼠、灵长类动物、非人哺乳动物、家养动物,诸如狗、猫、母牛、马等),并且优选是人。在其他实施方案中,受试者是癌症动物模型。举例来说,动物模型可为人口腔鳞状癌的原位异种移植物动物模型,或包含癌干细胞(csc)/未分化肿瘤。
在本发明方法的其他实施方案中,受试者尚未经受治疗,诸如化学疗法、放射疗法、靶向疗法和/或抗免疫疗法(诸如nk细胞相关免疫疗法)。在其他实施方案中,受试者已经受治疗,诸如化学疗法、放射疗法、靶向疗法和/或抗免疫疗法(诸如nk细胞相关免疫疗法)。
在某些实施方案中,受试者已进行用以移除癌性组织或癌前组织的手术。在其他实施方案中,癌性组织尚未被移除,例如癌性组织可位于身体的不宜手术区域中,诸如在为生命所必需的组织中,或在手术程序将在其中导致可观患者伤害风险的区域中。
本发明方法可用于治疗受试者的许多不同癌症诸如本文所述的那些,和/或确定受试者的许多不同癌症诸如本文所述的那些对单独或与其他nk免疫疗法组合的包含至少一种益生细菌的组合物的响应性。
iii.抗癌疗法
在一个方面,可施用其他抗癌疗法和/或免疫疗法组合或各种疗法的组合(例如一种或多种pi3kβ选择性抑制剂诸如kin193与一种或多种免疫检查点抑制剂诸如抗pd-1抗体组合,单独或与额外抗癌疗法诸如靶向疗法组合),特别是如果受试者已首先被指示为是对如本文公开的组合物的可能响应者。在其他实施方案中,一旦受试者被指示为不是对所述疗法的可能响应者,即可避免所述疗法,并且替代性治疗方案诸如靶向和/或未靶向抗癌疗法可连同如本文公开的组合物一起施用。
组合疗法也被涵盖,并且可包括例如一种或多种化学治疗剂和放射、一种或多种化学治疗剂和免疫疗法、或一种或多种化学治疗剂、放射和化学疗法,各组合可与如本文公开的疗法一起。如下所述,药剂可以与例如化学治疗剂、激素、抗血管新生剂、经放射性标记化合物或与手术、冷冻疗法和/或放射疗法的组合疗法形式施用。先前治疗方法可与其他形式的常规疗法(例如为熟练技术人员所熟知的癌症护理标准治疗)联合施用,采用与常规疗法连续,在常规疗法之前,或在常规疗法之后的方式。举例来说,这些调节剂可与治疗有效剂量的化学治疗剂一起施用。在其他实施方案中,这些调节剂与化学疗法联合施用以使化学治疗剂的活性和功效增强。医师案头参考(physicians’deskreference,pdr)公开已用于治疗各种癌症的化学治疗剂的剂量。在治疗上有效的这些以上提及的化学治疗药物的给药方案和剂量将取决于所治疗的特定黑素瘤、疾病的程度和为本领域中熟练医师所熟悉的其他因素,并且可由医师确定。
术语“靶向疗法”是指施用与所选生物分子选择性相互作用以由此治疗癌症的药剂。一个实例包括乳腺癌或卵巢癌抗原。
或者,免疫疗法是可包括例如使用癌症疫苗和/或敏化抗原递呈细胞的一种靶向疗法形式。举例来说,溶瘤性病毒是能够感染和溶解癌细胞,同时使正常细胞不受伤害,从而使得它们潜在适用于癌症疗法中的病毒。溶瘤性病毒的复制有助于肿瘤细胞破坏,并且也在肿瘤部位处产生剂量扩增。它们也可充当抗癌基因的载体,从而使得所述抗癌基因被特异性递送至肿瘤部位。免疫疗法可涉及通过施用预先形成的针对癌症抗原或疾病抗原的抗体(例如施用任选连接于化学治疗剂或毒素的针对肿瘤抗原的单克隆抗体)来实现的被动免疫性以短期保护宿主。免疫疗法也可集中于使用细胞毒性淋巴细胞识别的癌细胞系表位。或者,反义多核苷酸、核酶、rna干扰分子、三螺旋多核苷酸等可用于选择性调节与肿瘤或癌症的起始、进展和/或病理相关联的生物分子。
术语“未靶向疗法”是指施用不与所选生物分子选择性相互作用,但治疗癌症的药剂。未靶向疗法的代表性实例包括不限于化学疗法、基因疗法和放射疗法。
在某些实施方案中,使用化学疗法。化学疗法包括施用化学治疗剂。这种化学治疗剂可为但不限于从以下化合物群组之中加以选择的那些:铂化合物、细胞毒性抗生素、抗代谢剂、抗有丝分裂剂、烷基化剂、砷化合物、dna拓扑异构酶抑制剂、紫杉烷、核苷类似物、植物碱和毒素;及其合成衍生物。示例性化合物包括但不限于烷基化剂:顺铂(cisplatin)、曲奥舒凡(treosulfan)和氯乙环磷酰胺(trofosfamide);植物碱:长春花碱(vinblastine)、太平洋紫杉醇(paclitaxel)、多西紫杉醇(docetaxol);dna拓扑异构酶抑制剂:替尼泊苷(teniposide)、克立那托(crisnatol)和丝裂霉素(mitomycin);抗叶酸剂:甲氨蝶呤(methotrexate)、霉酚酸(mycophenolicacid)和羟基脲(hydroxyurea);嘧啶类似物:5-氟尿嘧啶(5-fluorouracil)、脱氧氟尿苷(doxifluridine)和胞嘧啶阿拉伯糖苷(cytosinearabinoside);嘌呤类似物:巯基嘌呤(mercaptopurine)和硫鸟嘌呤(thioguanine);dna抗代谢剂:2'-脱氧-5-氟尿苷、甘氨酸阿非迪霉素(aphidicolinglycinate)和吡唑并咪唑;以及抗有丝分裂剂:大田软海绵素(halichondrin)、秋水仙碱(colchicine)和根霉素(rhizoxin)。也可使用包含一种或多种化学治疗剂的组合物(例如flag、chop)。flag包含氟达拉滨(fludarabine)、胞嘧啶阿拉伯糖苷(ara-c)和g-csf。chop包含环磷酰胺、长春新碱、多柔比星(doxorubicin)和泼尼松(prednisone)。在另外实施方案中,使用parp(例如parp-1和/或parp-2)抑制剂,并且所述抑制剂在本领域中是熟知的(例如奥拉帕尼(olaparib)、abt-888、bsi-201、bgp-15(n-generesearchlaboratories,inc.);ino-1001(inotekpharmaceuticalsinc.);pj34(soriano等,2001;pacher等,2002b);3-氨基苯甲酰胺(trevigen);4-氨基-1,8-萘二甲酰亚胺(trevigen);6(5h)-菲啶酮(trevigen);苯甲酰胺(再颁美国专利36,397);和nu1025(bowman等))。作用机理通常与parp抑制剂能够结合parp以及使它的活性降低相关。parp催化β-烟酰胺腺嘌呤二核苷酸(nad+)转化成烟酰胺和聚adp-核糖(par)。聚(adp-核糖)与parp两者均已与转录调控、细胞增殖、基因组稳定性和癌发生相关联(bouchardv.j.等experimentalhematology,第31卷,第6期,2003年6月,第446-454页(9);hercegz.;wangz.-q.mutationresearch/fundamentalandmolecularmechanismsofmutagenesis,第477卷,第1、2期,2001年6月,第97-110页(14))。聚(adp-核糖)聚合酶1(parp1)是dna单链断裂(ssb)修复中的关键分子(demurciaj.等1997.procnatlacadsciusa94:7303-7307;schreiberv,dantzerf,amejc,demurciag(2006)natrevmolcellbiol7:517-528;wangzq等(1997)genesdev11:2347-2358)。通过抑制parp1功能来剔除ssb修复会诱导dna双链断裂(dsb),其可触发同源性引导的dsb修复有缺陷的癌细胞的合成致死性(bryanthe等(2005)nature434:913-917;farmerh等(2005)nature434:917-921)。化学治疗剂的前述实例是说明性的,并且不意图具有限制性。
在其他实施方案中,使用放射疗法。用于放射疗法中的辐射可为电离辐射。放射疗法也可为γ射线、x射线或质子束。放射疗法的实例包括但不限于外部射束放射疗法、间质植入放射性同位素(i-125、钯、铱)、放射性同位素诸如锶-89、胸部放射疗法、腹膜内p-32放射疗法和/或总体腹部和骨盆放射疗法。对于放射疗法的一般性概述,参见hellman,第16章:principlesofcancermanagement:radiationtherapy,第6版,2001,devita等人编,j.b.lippencottcompany,philadelphia。放射疗法可以外部射束放射或远距疗法形式施用,其中辐射从远距离源头加以引导。放射治疗也可以内部疗法或近距疗法形式施用,其中将放射源接近于癌细胞或肿瘤团块来放置在身体内部。也涵盖使用光动力疗法,其包括施用光敏剂,诸如血卟啉(hematoporphyrin)和它的衍生物、维替泊芬(vertoporfin)(bpd-ma)、酞菁(phthalocyanine)、光敏剂pc4、脱甲氧基-竹红菌素a(demethoxy-hypocrellina);和2ba-2-dmha。
在其他实施方案中,手术干预可物理移除癌性细胞和/或组织。
在其他实施方案中,使用激素疗法。激素治疗性治疗可包括例如激素激动剂、激素拮抗剂(例如氟他胺(flutamide)、比卡鲁胺(bicalutamide)、他莫昔芬(tamoxifen)、雷洛昔芬(raloxifene)、乙酸亮丙瑞林(leuprolideacetate)(利普安(lupron))、lh-rh拮抗剂)、激素生物合成和加工的抑制剂以及类固醇(例如地塞米松(dexamethasone)、类视黄醇(retinoid)、德尔托伊德(deltoids)、倍他米松(betamethasone)、皮质醇(cortisol)、可的松(cortisone)、泼尼松(prednisone)、脱氢睾固酮(dehydrotestosterone)、糖皮质素(glucocorticoid)、矿物类皮质激素(mineralocorticoid)、雌激素(estrogen)、睾酮(testosterone)、孕激素(progestin)、维生素a衍生物(例如全反式视黄酸(全反式视黄酸(atra));维生素d3类似物;抗促孕激素剂(例如米非司酮(mifepristone)、奥那司酮(onapristone))或抗雄激素剂(例如乙酸赛普龙(cyproteroneacetate))。
在其他实施方案中,使用高温,即一种其中使身体组织暴露于高温(多达106℉)的程序。热量可通过使细胞受损或将它们需要用以生存的物质从它们剥夺来有助于使肿瘤收缩。高温疗法可为使用外部和内部加热装置达成的局部性、区域性和全身高温。高温几乎始终与其他疗法形式(例如放射疗法、化学疗法和生物疗法)一起使用以试图使它们的有效性增加。局部高温是指将热量施加于极小区域诸如肿瘤。区域可用来自在身体外部的装置的瞄准肿瘤的高频波在外部加热。为实现内部加热,可使用若干类型的无菌探针中的一者,包括纤细经加热丝线或用温水填充的中空管;植入微波天线;和射频电极。在区域性高温中,对器官或肢体加热。将产生高能量的磁体和装置放置在待加热区域上。在称为灌注的另一方法中,将一些患者血液移除,加热,接着泵送(灌注)至待在内部加热的区域中。全身加热用于治疗已遍及身体进行扩散的转移性癌症。这可使用温水毯、热蜡、感应线圈(如电热毯中的那些)或热腔室(类似于大型孵育器)来实现。高温不导致辐射副作用或并发症的任何显著增加。然而,直接施加于皮肤的热量可在约一半治疗患者中导致不适或甚至显著局部疼痛。这也可导致水疱,其通常会快速愈合。
在其他实施方案中,光动力疗法(也称为pdt、光辐射疗法、光线疗法或光化学疗法)用于治疗一些类型的癌症。
在其他实施方案中,激光疗法用于利用高强度光照来破坏癌细胞。这个技术经常用于减轻癌症的症状诸如流血或阻塞,尤其是在癌症不能通过其他治疗来治愈时。它也可用于通过使肿瘤收缩或破坏来治疗癌症。
用疗法进行的治疗的持续时间和/或剂量可根据特定治疗剂或其组合而变化。适于特定癌症治疗剂的治疗时间将由熟练技术人员所了解。本发明涵盖连续评估各癌症治疗剂的最优治疗时程,其中受试者的如通过本发明方法确定的癌症表型是确定最优治疗剂量和时程的一个因素。
在其他实施方案中,可向受试者施用重组生物标志物多肽及其片段。在一些实施方案中,可构建和施用具有增强的生物性质的融合蛋白。此外,生物标志物多肽及其片段可根据本领域中熟知的药理学方法加以修饰(例如聚乙二醇化、糖基化、寡聚化等)以进一步增强合乎需要的生物活性,诸如生物可用度增加和蛋白水解降解降低。
临床功效可通过本领域中已知的任何方法来测量。举例来说,对疗法诸如如本文公开的组合物的响应涉及癌症例如肿瘤对疗法的任何响应,优选涉及在启动新辅助化学疗法或辅助化学疗法之后肿瘤质量和/或体积的变化。可评估在新辅助或辅助情况下的肿瘤响应,其中可将在全身性干预之后的肿瘤尺寸与初始尺寸和大小进行比较,所述尺寸和大小如通过ct、pet、乳房摄影术、超声或触诊所测量,并且可在组织学方面估计肿瘤的细胞性并与在启动治疗之前获取的肿瘤活检体的细胞性进行比较。响应也可通过在活检或手术切除之后对肿瘤进行测径规测量或病理性检查来评估。响应可以定量方式如肿瘤体积或细胞性变化百分比或使用半定量评分系统诸如残余癌症负荷(symmans等,j.clin.oncol.(2007)25:4414-4422)或米勒-佩恩(miller-payne)评分(ogston等,(2003)breast(edinburgh,scotland)12:320-327)加以记录,以定性方式如“病理性完全响应”(pcr)、“临床完全缓解”(ccr)、“临床部分缓解”(cpr)、“临床稳定疾病”(csd)、“临床进行性疾病”(cpd)或其他定性准则加以记录。肿瘤响应评估可在开始新辅助或辅助疗法之后早期进行,例如在数小时、数天、数周之后或优选在数月之后。响应评估的一典型终点是在终止新辅助化学疗法后或在手术移除残余肿瘤细胞和/或肿瘤床后。
在一些实施方案中,本文所述的治疗性治疗的临床功效可通过测量临床益处率(cbr)来确定。临床益处率通过在疗法结束后至少6个月的时间点测定处于完全缓解(cr)的患者的百分比、处于部分缓解(pr)的患者的数目百分比和患有稳定疾病(sd)的患者的数目百分比的总和来测量。这个公式的简写形式是cbr=超过6个月的cr+pr+sd。在一些实施方案中,特定治疗方案的cbr是至少25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%或更大。
用于评估对如本文公开的疗法的响应的额外准则与“存活”相关,所述存活包括以下中的全部:直至死亡的存活期,也称为总体存活期(其中所述死亡可不管原因或与肿瘤相关);“无复发存活期”(其中术语复发应包括局部化复发与远端复发两者);无转移存活期;无疾病存活期(其中术语疾病应包括癌症和与其相关的疾病)。所述存活期的长度可通过参照确定起点(例如诊断或开始治疗的时间)和终点(例如死亡、复发或转移)来计算。此外,用于确定治疗功效的准则可被扩展来包括对化学疗法的响应、存活概率、在给定时期内的转移概率以及肿瘤复发概率。
举例来说,为确定适当阈值,可向受试者群体施用特定抗癌治疗方案,并且可使结果与在施用任何如本文公开的组合物之前测定的生物标志物测量结果相关联。结局测量结果可为对在新辅助环境下给与的疗法的病理性响应。或者,结果量度诸如总体存活期和无疾病存活期可在已知其生物标志物测量值的疗法之后历经一段时期对受试者加以监测。在某些实施方案中,向各受试者施用相同剂量的治疗性组合物。监测受试者所持续的时期可变化。举例来说,可监测受试者至少2、4、6、8、10、12、14、16、18、20、25、30、35、40、45、50、55或60个月。与如本文公开的疗法的结果相关联的生物标志物测量阈值可使用各种方法诸如实施例章节中所述的那些来测定。
3.药物组合物
本发明提供本文公开的组合物的药学上可接受的组合物。如以下所详述,本发明的药物组合物可以固体或液体形式特别配制以进行施用,所述固体或液体形式包括适合于以下的那些形式:(1)口服施用,例如灌服剂(水性或非水性溶液或混悬液)、片剂、大丸剂、粉剂、颗粒剂、糊剂;(2)例如通过以例如无菌溶液或混悬液形式皮下、肌肉内或静脉内注射进行的胃肠外施用;(3)表面施加,例如以乳膏剂、软膏剂或喷雾剂形式向皮肤施加;(4)阴道内或直肠内,例如以阴道栓剂、乳膏剂或泡沫剂形式;或(5)喷雾,例如以水性气雾剂、脂质体制剂或固体粒子形式。
本文所述的组合物例如益生细菌的组合物可用于口服施用以到达胃肠道,从而对准将益生细菌引入胃肠道的组织中的目标。本发明的治疗性组合物的制剂也可包括其他益生菌剂或促进芽孢萌发和/或细菌生长的营养物。一示例性物质是促双歧杆菌生长性寡糖,其促进有益益生细菌的生长。在某一实施方案中,益生细菌菌株与治疗有效剂量的(优选是广谱)抗生素或抗真菌剂组合。在一些实施方案中,本文所述的组合物被囊封成以肠溶方式包覆的定时释放胶囊或片剂。肠溶性包衣允许胶囊/片剂在它穿过胃肠道时保持完整(即未溶解),直至在某一时间之后和/或直至它到达胃肠道的某一部分(例如小肠)。定时释放组分持续预定时期防止本文所述的组合物中的益生细菌菌株的“释放”。
本发明的治疗性组合物也可包括已知抗氧化剂、缓冲剂和其他试剂诸如着色剂、调味剂、维生素或矿物质。
在一些实施方案中,本发明的治疗性组合物,例如破骨细胞、破骨细胞的细胞培养物和/或破骨细胞的细胞培养物的上清液,可单独或与在生理上可与它向其施用的物种相容的载体组合施用。载体可包含基于固体的干燥物质以用于配制成片剂、胶囊或粉状形式;或载体可包含基于液体或凝胶的物质以用于配制成液体或凝胶形式。载体的特定类型以及最终制剂部分地取决于所选施用途径。本发明的治疗性组合物也可包括多种载体和/或粘合剂。一优选载体是以足以完成1克剂量总重量的量添加的微晶纤维素(mcc)。载体可为基于固体的干燥物质以用于以片剂、胶囊或粉状形式配制,并且可为基于液体或凝胶的物质以用于以液体或凝胶形式配制,所述形式部分地取决于施用途径。用于干燥制剂的典型载体包括但不限于:海藻糖、麦芽糖糊精、米粉、微晶纤维素(mcc)、硬脂酸镁、肌醇、fos、gos、右旋糖、蔗糖和类似载体。基于液体或凝胶的适合载体包括但不限于:水和生理盐溶液;脲;醇和衍生物(例如甲醇、乙醇、丙醇、丁醇);二醇(例如乙二醇、丙二醇等)。优选地,基于水的载体具有中性ph值(即ph7.0)。用于施用本文所述的组合物的其他载体或试剂在本领域中是已知的,例如在美国专利号6,461,607中。本公开的破骨细胞、破骨细胞的细胞培养物和/或破骨细胞的细胞培养物的上清液可使用本领域中已知以及本文所述的局部性和/或全身性施用途径来施用。
本文所述的破骨细胞(oc)或树突细胞(dc)可用于通过本领域中已知的任何施用途径来以任何药学上可接受的组合物形式向受试者施用。举例来说,oc或dc、包含所述oc或dc的细胞培养物、或所述细胞培养物的上清液,任选与额外药剂一起,可通过全身性和/或局部性(例如向癌症或肿瘤组织或在癌症或肿瘤组织附近)注射(例如静脉内)来以药物组合物形式施用。
短语“药学上可接受”在本文中用于指代在合理医学判断的范围内适用于与人类和动物的组织接触而无过度毒性、刺激、过敏应答或其他问题或并发症,与合理益处/风险比率相称的那些试剂、物质、组合物和/或剂型。
如本文所用的短语“药学上可接受的载体”意指将主题化学物质从一个器官或身体部分运载或运送至另一器官或身体部分中涉及的药学上可接受的物质、组合物或媒介物,诸如液体或固体填充剂、稀释剂、赋形剂、溶剂或囊封物质。各载体在可与制剂的其他成分相容,并且不对受试者有损伤的意义上必须是“可接受的”。可充当药学上可接受的载体的物质的一些实例包括:(1)糖,诸如乳糖、葡萄糖和蔗糖;(2)淀粉,诸如玉米淀粉和马铃薯淀粉;(3)纤维素和它的衍生物,诸如羧甲基纤维素钠、乙基纤维素和乙酸纤维素;(4)粉状黄蓍胶;(5)麦芽;(6)明胶;(7)滑石;(8)赋形剂,诸如可可脂和栓剂蜡;(9)油,诸如花生油、棉籽油、红花油、芝麻油、橄榄油、玉米油和大豆油;(10)二醇,诸如丙二醇;(11)多元醇,诸如甘油、山梨糖醇、甘露糖醇和聚乙二醇;(12)酯,诸如油酸乙酯和月桂酸乙酯;(13)琼脂;(14)缓冲剂,诸如氢氧化镁和氢氧化铝;(15)海藻酸;(16)无热原水;(17)等张盐水;(18)林格氏溶液(ringer'ssolution);(19)乙醇;(20)磷酸盐缓冲溶液;和(21)用于药物制剂中的其他无毒可相容物质。
适于口服施用的制剂可呈胶囊、扁囊剂、丸剂、片剂、糖锭(使用调味基质,通常是蔗糖和阿拉伯胶或黄蓍胶)、粉剂、颗粒剂形式,或呈于水性或非水性液体中的溶液或混悬液形式,或呈水包油或油包水液体乳液形式,或呈酏剂或糖浆形式,或呈软锭剂(使用惰性基质,诸如明胶和甘油、或蔗糖和阿拉伯胶)形式和/或呈嗽口剂等形式,各自含有预定量的一种或多种如本文公开的细菌菌株。
本发明也涵盖用于检测和/或调节本文所述的生物标志物的试剂盒。本发明的试剂盒也可包括公开或描述在所公开发明的如本文提供的方法中使用所述试剂盒或所公开发明的抗体的指导材料。试剂盒也可包括额外组分以有助于所述试剂盒被设计所针对的特定应用。举例来说,试剂盒可另外含有检测标记的手段(例如酶标记的酶底物、用以检测荧光标记的过滤套件、适当二级标记诸如绵羊抗小鼠hrp抗体等)和为对照所必需的试剂(例如对照生物样品或标准物)。试剂盒可另外包括缓冲剂和被认定用于所公开发明的方法中的其他试剂。非限制性实例包括用以降低非特异性结合的试剂诸如载体蛋白或清洁剂。
实施例
实施例1:用于实施例2和3的材料和方法
细胞系、试剂和抗体
具有10%胎牛血清(fbs)(geminibio-product)的rpmi1640完全培养基用于细胞培养。口腔鳞状癌细胞(oscc)和口腔鳞状癌干细胞(oscsc)在ucla从患有舌部肿瘤的癌症患者分离[参见参考文献2和33-35;以下所有引用都涉及同一参考文献清单]。具有10%fbs的alpha-mem(lifetechnologies,ca)用于破骨细胞和dc培养。m-csf(biolegend,ca)以及rankl、gm-csf和il-4从peprotech(nj)购买,并且rh-il-2从nih-brb获得。人cd3/cd28t细胞活化剂从stemcelltechnologies购买。
针对mhc-i、kir2、kir3、cd44、cd54、b7h1、cd16、nkg2d、mica/b、klgr1、cd45、cd3/16/56、cd8、cd3、cd28,cd4、gl3、nkp40、nkp30、nkp44、nkp46和cd94的抗体从biolegend(sandiego,ca)购买。ulbp1-6抗体从r&dsystems购买。碘化丙锭(pi)从sigma(st.louis,mo)购买。saj2如先前[36]所述制备。
从人pbmc和hu-blt脾细胞纯化nk细胞和t细胞
如先前[37]所述纯化nk细胞和t细胞。来自hu-blt脾细胞的t细胞使用来自stemcelltechnologies(stemcelltechnologies,vancouver,canada)的分离试剂盒加以正性纯化。
从hu-blt小鼠和人pbmc纯化单核细胞以及产生树突细胞和破骨细胞
由ucla机构审查委员会(irb)核准的书面知情同意书从健康供血者获得,并且所有程序都由ucla-irb核准。如先前[37]所述纯化单核细胞。使用人cd14分离试剂盒(ebioscience,sandiego,ca),从骨髓正性分离来自hu-blt小鼠的单核细胞。基于流式细胞计量分析,各子组都实现大于95%纯度。通过用m-csf(25ng/ml)和rankl(25ng/ml)处理21天来使单核细胞分化成破骨细胞。为获得dc,用gm-csf(150ng/ml)和il-4(50ng/ml)处理单核细胞7天。
nk细胞的扩增
人纯化nk细胞和hu-blt增浓nk细胞用rh-il-2(1000u/ml)和抗cd16mab(3μg/ml)活化18-20小时,随后使它们与饲养细胞和saj2一起共培养。每三天将具有il-2的培养基更新。
肿瘤植入以及从hu-blt小鼠进行的组织制备
本手稿中描述的动物研究根据所有联邦指导方针、州指导方针和当地指导方针,在ucla动物研究委员会(arc)的书面核准下进行。联合免疫缺陷nod.cb17-prkdcscid/j和nod.cg-prkdcscidil2rgtm1wjl/szj(缺乏t细胞、b细胞和天然杀伤细胞的nsg)从jacksonlaboratory购买。人源化blt(hu-blt;人骨髓/肝/胸腺)小鼠如先前以及在图8中所述在nsg背景上制备[38,39]。为建立原位肿瘤,首先用异氟烷与氧组合使小鼠麻醉,并且接着将肿瘤细胞于具有10μlhc基质胶(corning,ny,usa)的混悬液中直接在口底中注射(1x106个细胞)。在肿瘤注射之后四至五周,使小鼠安乐死,并且收集骨髓、脾和血液,并制备单细胞混悬液[40]。
elisa和多路细胞因子阵列试剂盒
单一elisa和多路测定如先前[37]所述进行。
用nk细胞上清液进行的癌干细胞分化
如先前[36]所述制备由nk细胞获得的上清液,并且用于oscsc的分化。由经oc扩增nk细胞获得的第13天上清液用于分化。
表面染色和细胞死亡测定
如先前[37,41,42]所述,通过用抗体或碘化丙锭标记细胞来进行染色。使用beckmancoulterepicsxl细胞仪(brea,ca)进行流式细胞计量术分析,并且使用flowjovx软件(ashland,or)来分析结果。
51cr释放细胞毒性测定
如先前[43]所述进行51cr释放测定。
统计分析
进行未配对或配对双尾史都登t检验(studentt-test)以达成统计分析。采用邦弗伦尼(bonferroni)事后检验的单因素anova用于比较不同组。***(p值<0.001),**(p值0.001-0.01),*(p值0.01-0.05)。
实施例2:经破骨细胞活化超动力nk细胞优先和快速扩增cd8+t细胞,从而导致来自癌症患者和blt人源化小鼠的天然杀伤细胞数目下降
由破骨细胞对nk细胞以及由树突细胞对t细胞的优先扩增和显著功能获得
将破骨细胞、单核细胞和dc对nk细胞扩增和功能的活化作用进行比较。在nk细胞与oc和/或saj2一起共培养之前18-20小时,用il-2和抗cd16mab使nk细胞活化。oc和saj2的组合优先扩增nk细胞,同时维持低比例的t细胞(图1)。接着在用saj2处理的与oc、dc和单核细胞的共培养物之间比较nk培养物中污染性t细胞的扩增速率和水平。nk细胞与oc共培养优先扩增nk细胞,并且污染性t细胞的比率遍及培养的前一个月至两个月保持极低(图2和图4a)。相比之下,dc优先扩增t细胞,并且nk细胞的比例保持低下。尽管单核细胞也能够使t细胞扩增,但t细胞扩增保持低于nk细胞的扩增。在所有三种培养的情况下都存在t细胞比例的初始增加,然而,在后续培养中,在与oc共培养的情况下,t细胞扩增的速率降低,而在与dc一起培养的情况下,t细胞继续扩增,并且实质上增加。也可在与单核细胞共培养的情况下观察到t细胞扩增的稳定状态。(图2a-图2d)。
相比于与单核细胞或dc共培养的nk细胞,与oc共培养的nk细胞能够以显著更大程度使oscsc溶解,并且从第9天至第15天存在显著增加,与在与oc共培养的情况下在第15天nk细胞的较高扩增共相关(图2e)。相较于与单核细胞或dc共培养的nk细胞,与oc一起培养的经il-2和抗cd16mab活化nk细胞分泌显著更高量的ifn-γ(图2f)。在分析nk细胞扩增速率和群体倍增(由最终计数与基线计数的比率的对数除以2的对数加以定义)后,发现在第20天oc使nk细胞扩增至21,000-132,000倍,并且在第31天oc使nk细胞扩增至300,000-5,100,000倍,其中在4周内达到17-21次群体倍增(图2g-图2h)。
将经新鲜分离单核细胞与成熟dc和oc在关键表面受体的表达方面进行比较。当相较于单核细胞时,在dc和oc上,cd54被上调,而在dc和oc上,mhc-i被降低(图1d)。杀伤细胞免疫球蛋白样受体(kir)、klrg1和mica/b在oc上是较高的,在单核细胞上是中等的,并且在dc上是极低的(图2i)。ulbp1-6在单核细胞上是高的,在oc上是中等的,并且在dc上是低的(图2i)。相较于未处理原代nk细胞(图2j上排),经oc扩增nk细胞上的包括cd94和nkg2d的nk细胞受体是更高的(图2j下排)。在经扩增nk细胞上,kir2和kir3表达是中等的(图2j下排)。
从经oc扩增nk细胞纯化的残余t细胞群体不介导细胞毒性,但分泌ifn-γ
来自经oc扩增nk细胞的大多数t细胞污染物是cd8+t细胞(图3a)。将来自第9天经oc扩增nk细胞的t细胞污染物分选出来以获得纯化t细胞和nk细胞。接着使用cd16和cd3/56抗体测试nk细胞的纯度(图3b)。接着用il-2处理nk细胞和t细胞18-20小时,随后将它们用于针对oscsc和k562的51cr释放测定中。从经oc扩增nk细胞分离的cd3+t细胞未能使oscsc(图3c)或k562(图3d)溶解。相较于t细胞,由nk细胞获得的上清液分泌显著更高水平的ifn-γ(图3e)。
用破骨细胞进行的nk细胞扩增在第一个月中保持较高,在第二个月中逐渐降低,并且在第三个月中实质上降低
与oc一起培养的nk细胞持续31-36天进行扩增,同时污染性t细胞的比率保持低下(图4a)。使第36天经扩增nk细胞与oc一起再培养以进行第二轮扩增,并且nk扩增继续持续27天(图4b)。类似地,在用oc进行的第二轮nk细胞扩增中,t细胞扩增速率保持极低(图4b)。使第67天经扩增nk细胞与oc一起再培养以进行第三轮扩增,然而,nk细胞由于t细胞扩增而逐渐丧失(图4c)。在用oc进行的3轮扩增中,不可观察到扩增nk细胞的细胞死亡或可观察到微小水平的细胞死亡(图4m)。从第一轮扩增至第二轮扩增,nk细胞使癌干细胞溶解以及分泌ifn-γ的能力逐渐降低,并且在较大百分比的t细胞在扩增所处的第三轮中,这些功能变为最小(图4n-图4s)。
破骨细胞而非k562或oscsc使nk细胞扩增以及使nk细胞功能增加。
使经活化nk细胞与oscsc、k562、oc、经照射k562或经照射oc一起在saj2存在下培养,并且测定nk扩增和它们的功能的水平(图5a-图5h)。由非照射k562、经照射k562或oscsc诱导的nk细胞扩增和功能(细胞毒性和ifn-g分泌)显著低于由非照射oc或经照射oc诱导的那些(图5)。
由来自患者的nk细胞达成的降低的细胞毒性和较低ifn-γ分泌与t细胞扩增增加相符
当与oc一起培养时,来自癌症患者的纯化nk细胞不能维持nk细胞的扩增,并且实际上,截至第12天,大于半数的扩增细胞是t细胞。此外,截至第31天,仅10%nk细胞剩余在培养物中(图6a和图7a)。此外,当在扩增的31-36天内测定在癌症患者的情况下的经扩增nk细胞和t细胞的总数时,在相较于健康对照时,存在来自癌症患者的较少扩增细胞(图6d和图7c),并且扩增t细胞的水平显著高于nk细胞(图6e-图6f和图7d-图7f)。相比之下,从健康供者分离的nk细胞维持nk细胞的扩增,并且nk扩增水平显著高于t细胞(图6b、图6e、图6f和图7b、图7e、图7f)。在不同培养天数时,当相较于健康nk细胞时,在患者nk细胞的情况下观察到nk细胞不增殖或低得多的增殖(图7f)。不可观察到来自健康供者或患者的扩增细胞的显著细胞死亡,但相比于健康供者,来自患者的细胞的死亡率略微更高(图6c)。
当相较于与oc一起培养的健康nk细胞时,与oc一起培养的患者nk细胞以显著更小程度使oscsc溶解(图6g和图7g)。当基于nk细胞的数目加以标准化时,在相较于来自健康对照的nk细胞时,患者的每个nk细胞的诱导细胞毒性更小(图6h和图7h)。当相较于经oc扩增健康nk细胞时,经oc扩增患者nk细胞分泌显著更少ifn-γ(图6i和图7i)。当相较于健康nk细胞时,经oc扩增口腔癌患者nk细胞分泌显著更少il-10(图6j),而当相较于健康nk细胞时,来自胰腺癌患者的那些nk细胞分泌更高il-10(图7j)。对于由健康nk细胞或癌症患者nk细胞达成的il-6分泌水平,不可观察到显著差异(图6k和图7k)。相较于由破骨细胞扩增的健康对照nk细胞,在患者nk细胞的表面上,nkg2d表面表达的水平是类似的,而cd94表达是更高的,并且kir2、nkp30、nkp44和nkp46表达是更低的(图6l和图6m)。
由患者经扩增nk细胞获得的上清液具有低得多的使oscsc分化的能力
用相等体积的由患者和健康供者获得的第13天上清液处理oscsc18-20小时,随后分析cd44、b7h1、cd54和mhc-i表达水平(图6n)。在mhc-i表达方面存在由健康nk上清液达成的增加至7.1倍,但患者nk上清液仅诱导增加至2.56倍(图6n)。对于cd54表达,相较于用患者nk上清液达成的增加至2.1倍,观察到由健康nk上清液达成的增加至13倍。就b7h1而言,相较于用患者nk上清液达成的增加至1.5倍,观察到由健康nk上清液达成的增加至3.75倍。cd44由健康nk上清液降低,而未观察到由患者nk上清液达成的降低(图6n)。在用nk上清液处理oscsc之后,不可观察到显著细胞死亡(图6o)。如图6p中所示,当用健康nk上清液处理时,观察到nk细胞介导的细胞毒性降低74%,而用患者nk上清液仅可观察到33%降低(图6p)。
人源化小鼠中的口腔肿瘤优先扩增t细胞,从而导致nk细胞毒性丧失,同时保留ifn-γ分泌。
将人源化blt小鼠用口腔肿瘤植入,并且在肿瘤植入之后4周将小鼠处死。收集来自hu-blt小鼠的脾,并且将t细胞分选出来。接着用il-2和抗cd16mab处理含有b细胞的流穿细胞(图8)18-20小时,随后使它们与blt-oc一起培养。尽管携带肿瘤的hu-blt小鼠含有较大百分比的nk细胞(图9a),但扩增导致t细胞逐渐和显著扩增,此在第6天开始,并且持续至第22天,此时,96%的细胞是t细胞,并且仅1.1%是nk细胞。相比之下,来自不患有肿瘤的hu-blt小鼠的初始含有较少nk细胞的流穿细胞达成nk细胞扩增,并且水平从在第6天的28.6%nk细胞升高至在第22天的69%nk细胞(图9b)。当与自体oc一起培养时,在两种动物的情况下,nk细胞的水平从初始培养日开始均得以增加,但无肿瘤小鼠从第0天至第6天增加至10.59倍,而患有肿瘤的小鼠增加至4.56倍(图9b)。相较于不患有肿瘤的小鼠,在携带肿瘤的小鼠的情况下,经扩增淋巴细胞的总数是低下的(图9c),其中大多数是t细胞而非nk细胞(图9d和图9e)。
相比于不患有肿瘤的对照小鼠的富含nk的细胞,来自携带肿瘤的小鼠的富含nk的细胞在与oc共培养时能够以显著更小程度使oscsc溶解(图9f)。此外,当基于每个nk细胞来评估细胞毒性时,相较于来自不患有肿瘤的对照小鼠的nk细胞,在来自携带肿瘤的小鼠的nk细胞的情况下观察到较小细胞毒性(图9g)。当相较于不患有肿瘤的对照小鼠时,来自携带肿瘤的小鼠的富含nk的细胞分泌显著更高ifn-γ(图9h)、更低il-10(图9i)和略微更低il-6(图9j)。相较于不患有肿瘤的对照小鼠,来自携带肿瘤的hu-blt小鼠的外周血液的血清展现ifn-γ、il-10和il-6的分泌增加(图9k)。
il-15部分地介导由破骨细胞对nk细胞的扩增,而il-12导致由nk细胞达成的ifn-γ分泌增加
我们测定了由原代nk细胞和第6天经oc扩增nk细胞分泌的细胞因子、趋化因子、生长因子和配体的水平(图10)。当相较于由用il-2和抗cd16mab活化的原代nk细胞分泌的那些时,由经oc扩增nk细胞分泌的细胞因子、趋化因子、生长因子和配体中的大多数是更高的(图10)。相较于原代nk细胞,观察到由经oc扩增nk细胞达成的50-60倍高的il-12诱导以及20-26倍高的il-15分泌(图10)。添加抗il-12mab和/或抗il-15mab使细胞扩增显著降低,其中1μg/mlil-15抗体具有最大作用(图11a)。用il-12抗体和/或il-15抗体处理不影响在第9天时nk细胞的细胞毒性功能(图11b),但在第15天,nk细胞的细胞毒性功能被显著抑制(图11b)。当相较于il-15抗体处理时,由经oc扩增nk细胞达成的ifn-γ分泌水平由il-12抗体更大程度降低(图11c)。
添加抗cd3抗体会控制t细胞扩增以及使经oc扩增nk细胞增加
用cd3抗体处理淋巴细胞,并且评估在不同天数的nk或t细胞扩增(图12a和图12b)。已丧失前向散射与侧向散射两者的细胞的水平增加(图12c和图12d),并且在经cd3抗体处理细胞的情况下获得dna碎裂升高,如由在细胞周期分析中亚g0/g1峰增加所证实,从而指示t细胞的丧失(图12e)。相应地,在患者与健康供者两者的情况下,nk细胞的水平均升高(图12a-图12b)。丧失前向散射和侧向散射的群体是cd3+t细胞,如通过cd3和cd16mab染色(数据未显示)所确定。在不存在nk细胞下,经抗cd3mab处理t细胞既不丧失前向散射和侧向散射(图13),也不展现细胞死亡(数据未显示)。当在添加抗cd3mab之前和之后评估nk细胞的细胞毒性时,来自健康供者与患者供者两者的nk细胞均具有显著较低细胞毒性,但在健康nk细胞的情况下,水平仍然较高(图12g)。相比之下,在健康nk样品与患者nk样品两者的情况下,ifn-γ分泌水平均显著升高,从而指示由抗cd3mab结合的t细胞诱导nk细胞中的分割无反应性(图12f)。
经破骨细胞活化nk细胞使cd8+t细胞数目实质上增加
当相较于健康个体时,在相较于健康对照时,癌症患者平均来说具有适度更高百分比的cd8+t细胞,以及更低百分比的cd4+t细胞(图14a)。当与破骨细胞一起培养时,在不存在nk细胞下,t细胞未能达成cd8+t细胞扩增,然而,含有极小部分的污染性t细胞的用oc活化的纯化nk细胞使来自健康培养物与患者培养物两者的cd8+t细胞扩增,即使相比于健康nk细胞,患者nk细胞培养物使t细胞更快扩增(图14b)。当将经oc活化nk细胞扩增的t细胞与经dc活化nk细胞扩增的t细胞进行比较时,经oc优先扩增nk细胞介导cd8+t细胞,而经dc扩增nk细胞介导cd4+t细胞(图14c-图14g)。此外,经oc活化nk细胞扩增的cd8+t细胞全都具有活化表型,因为它们呈cd45ro+cd44+cd62l-/低ccr7-/低,从而指示记忆/效应表型(图14h)。
体内和体外研究新近显示破骨细胞是nk细胞的主要活化物。更重要的是,用破骨细胞单一每月刺激能够持续超过两个月维持由健康供者获得的超动力nk细胞的扩增。此外,nk细胞的细胞毒性功能在扩增的第一个月中保持显著较高,并且在扩增的第二个月中下降。不明确的是为何nk细胞功能在刺激的第二个月持续下降,尽管大量nk细胞持续进行扩增。有可能额外信号为在第二个月维持nk细胞细胞毒性所需,和/或需要更频繁补充以oc。
相较于先前所述那些(表1),使用oc作为饲养细胞是用以使大量nk细胞扩增的最佳策略。首先,申请人在第20天获得21,000-132,000倍扩增,并且在第31天获得30-510万倍扩增,其中在扩增的4周内达成17-21次群体倍增,相比于任何先前报道方法,这是高得多的速率(表1)。尽管由于使用不同类型的靶标而难以在各研究之间比较经扩增nk细胞的细胞毒性功能,但本文公开的策略提供具有靶向和溶解癌干细胞,并且分泌ifn-γ的重大能力的大量nk细胞(表1)。此外,原代nk细胞在不具有或具有极小转化概率下得以扩增,所述转化已知会影响nk细胞杀伤潜力,因为大多数nk株系(诸如yt、nk92或nk-l)或经转染nk细胞分别倾向于丧失它们的大部分或一些细胞毒性能力和ifn-γ分泌能力。此外,尽管用以维持nk细胞扩增的抗cd3抗体处理将是一种用以限制t细胞扩增的优异策略,但所述nk细胞的细胞毒性活性由于在nk细胞中诱导的显著分割无反应性而较低。由于ifn-γ分泌较大增加,所以这个策略对于诱导肿瘤的分化可为优异的;然而,对于消除肿瘤,它的最优性小得多。这可为为何在对实体肿瘤的免疫疗法中使用nk细胞在少数测试患者中不导致肿瘤消退的一个原因。
表1
与oc相比,dc刺激纯化nk细胞中少数污染性t细胞的优先扩增,并且t细胞的水平持续升高一个月,由此使nk扩增降低。因此,观察到较低水平的针对oscsc的细胞毒性。引起关注的是,当在采用oc培养物和dc培养物而观察到类似nk细胞比例所处的较早时间点评估nk细胞的细胞毒性时,可观察到由与oc一起培养的nk细胞达成的nk细胞毒性升高(数据未显示)。类似地,当相较于与dc一起培养的那些时,存在由与oc一起培养的nk细胞达成的ifn-γ分泌增加,从而指示基于每个细胞,相较于与dc一起培养的那些,nk细胞在与oc一起培养时分泌更高ifn-γ(图1c)。从经oc扩增nk细胞分选出来的t细胞具有极低细胞毒性,并且用t细胞所见的最小细胞毒性可能归因于污染性nk细胞(图3c和图3b)。在另一方面,t细胞可能释放ifn-γ,但水平显著低于由nk细胞释放的那些(图3e)。当将t细胞的子组cd8、cd4和gdt细胞分选出来并用于细胞毒性测定中时,仅nk细胞能够使这些细胞溶解,并且用t细胞子组所见的残余细胞毒性归因于污染性nk细胞(图3f)。oc相较于dc显示更高活化性nk配体表达,而它们两者均显示类似水平的分化抗原cd54。引起关注的是,当相较于单核细胞时,它们两者均显示较低水平的i类mhc表达(图1d)。此外,il-15分泌似乎对于nk细胞的增殖是重要的(图11a),而il-12对于ifn-γ的分泌是重要的(图11c)。相较于扩增中的较早时间点,当在稍后日评估时,两种细胞因子在nk细胞的细胞毒性功能方面均是重要的(图11b)。
相较于oc,所有测试肿瘤细胞系,无论它们是非照射的还是经照射的,都不长期支持nk细胞的扩增,并且不短期支持nk细胞的扩增,即使可在用k562和oscsc进行的培养中观察到ifn-γ的分泌,这个效应是短暂的(图5)。此外,也将经oc诱导扩增与经照射pbmc进行比较,并且发现它显著次于oc进行的诱导扩增(数据未显示)。当相较于原代nk细胞时,由oc扩增的nk细胞显示高得多水平的活化性受体表达,包括nkg2d、nkp46、nkp44、nkp30、cd94,以及增加的抑制性受体kir2和kir3,伴有低得多的cd16受体表达(图2j,下排)。当相较于健康供者的经oc扩增nk细胞时,癌症患者一般具有低得多的受体表达,并且水平甚至小于在已丧失重大细胞毒性和细胞因子分泌能力,并且允许达成t细胞的扩增的第三轮经刺激健康nk细胞的情况下所见的那些。类似于患者nk细胞,由k562和oscsc扩增的nk细胞是短命的,并且具有低得多的细胞毒性和细胞因子分泌能力。不同于oc,k562或oscsc表达较低水平的nk活化性配体(图1d),并且缺乏导致nk细胞的扩增的关键细胞因子的分泌,因为由oc在这些信号方面达成的增加能够使nk细胞的扩增和功能升高(图10a)。仍然待确定的是由经工程改造k562(表1)达成的所述信号是否次于由oc递送的那些,因为由经工程改造k562细胞达成的nk细胞扩增速率是经oc扩增nk细胞的1/100量值低(表1以及图5a和图5b)。此外,由经工程改造k562连续刺激为维持nk扩增所需,而用oc进行仅一次刺激足以持续超过一个月使超动力nk细胞扩增。此外,不同于原代nk细胞,经oc扩增nk细胞十分良好经得住冷冻温度,并且保留它们的超动力特征和扩增速率而不丧失活力或功能(图15)。
当自体或同种异体oc用于使来自癌症患者的nk细胞扩增时,观察到极其不同概况。癌症患者的oc也在培养早期使t细胞扩增,伴有在不同天数时nk细胞的总体扩增降低。当在oc培养之后评估患者nk细胞的功能时,可观察到每个nk细胞的nk细胞细胞毒性显著丧失,以及ifn-γ分泌降低(图6h和图15g)。这个观察结果是重要的,因为它指示癌症患者的纯化nk培养物中小部分污染性t细胞的较快扩增与nk细胞的细胞毒性功能丧失相关联。因此,nk细胞的丧失也可为癌干细胞的生长和转移提供促进背景。
为测试相比于它们的非携带肿瘤的对应物,从用肿瘤植入的人源化小鼠获得的oc是否类似于癌症患者oc使纯化nk培养物内小部分污染性t细胞更快扩增,我们在口底中植入oscsc并使其生长。在生长五周之后,使小鼠安乐死,并且使t细胞消减,随后使细胞与自体和同种异体oc一起培养,并且测定nk细胞扩增的速率。类似于健康供者nk细胞,来自不患有肿瘤的hu-blt小鼠的nk细胞持续较长时期达成nk细胞扩增,而来自携带肿瘤的小鼠的那些使nk培养物内小部分污染性t细胞更快扩增,从而有利于t细胞扩增超过nk细胞。引起关注的是,类似于在癌症患者的情况下观察到的nk细胞细胞毒性丧失,我们也在用肿瘤植入的hu-blt小鼠的情况下观察到nk细胞细胞毒性显著丧失,此可为t细胞扩增的潜伏机理。然而,当相较于不患有肿瘤的那些时,来自携带肿瘤的小鼠的经oc扩增nk+t细胞分泌更高水平的ifn-γ,从而表明对nk细胞中分割无反应性的潜在诱导以驱动癌干细胞的分化。发现这是事实,因为经nk注射的携带肿瘤的hu-blt小鼠中的肿瘤的单细胞制备物显示较高分化抗原,并且对nk细胞介导的细胞毒性具有抗性。
据假设人源化小鼠中的微环境由于在鼠il-15与人il-15之间缺乏交叉反应性而不有助于nk细胞的扩增,因为添加人il-15会促进nk细胞增加。不同于需要来自破骨细胞的信号达成它们的扩增的nk细胞,t细胞在不存在破骨细胞下快速扩增,并且破骨细胞适度刺激t细胞扩增(图3g和图3h)。有可能的是从小鼠组织微环境接收的信号足以维持t细胞和b细胞增殖,而nk细胞增殖需要来自骨髓性子组的信号。引起关注的是,在外周血液中以及在组织中,nk子组与骨髓性子组两者的频率均降低,而在骨髓中,富含骨髓细胞,t细胞和nk细胞的水平保持类似。在外周中dc是否有利于t细胞的扩增以及在骨髓中破骨细胞是否有利于nk细胞的扩增需要进一步研究。人源化小鼠是人癌症模型的最佳和最接近模型,因为在两种模型中均发现较低百分比以及降低功能的nk细胞潜在促进癌症进展。
最令人激动的研究结果是当在oc存在下培养时,nk细胞能够使cd8+t细胞增加。这个数据指示nk细胞是cd8+t细胞的扩增中的重要效应物,由此允许对表达较高i类mhc的肿瘤细胞的靶向和溶解得以增加。nk细胞是否使抗原特异性cd8+t细胞增加,由此使以抗原特异性方式对肿瘤细胞的溶解增加需要未来研究。癌症患者和人源化小鼠的外周血液/组织中t细胞快速扩增以及nk细胞数目降低对于由nk细胞靶向i类mhc低下靶标以使肿瘤负荷最小化来说可为有害的,所述靶标包括癌干细胞/未分化肿瘤。此外,nk细胞也提供大量ifn-γ来促进癌干细胞的最优分化,以及促进较高i类mhc表达以达成由cd8+t细胞进行靶向。因此,使癌症患者中的nk细胞数目和功能恢复对于建立有效肿瘤控制将是重要的。
实施例3:经破骨细胞活化超动力nk细胞优先和快速扩增超动力cd8+t细胞:在癌症患者和blt人源化小鼠的情况下由经oc扩增nk细胞达成的cd8+t细胞扩增的动力学增加
对实施例2中的研究进行进一步研究,并且结果在以下加以概述。
从经oc扩增nk细胞纯化的残余t细胞群体不介导细胞毒性,但分泌ifn-γ
来自经oc扩增nk细胞的大多数t细胞污染物是cd8+t细胞(补充图s2a)。将来自第9天经oc扩增nk细胞的t细胞污染物分选出来以获得纯化t细胞和nk细胞。接着使用cd16和cd3/56抗体测试nk细胞的纯度(补充图s2b)。接着用il-2处理nk细胞和t细胞18-20小时,随后将它们用于针对oscsc和k562的51cr释放测定中。从经oc扩增nk细胞分离的cd3+t细胞未能使oscsc(补充图s2c)或k562(补充图s2d)溶解。相较于t细胞,由nk细胞获得的上清液分泌显著更高水平的ifn-γ(补充图s2e)。
由来自患者的nk细胞达成的降低的细胞毒性和较低ifn-γ分泌与t细胞扩增增加相符
当与oc一起培养时,来自癌症患者的纯化nk细胞不能维持nk细胞的扩增,并且实际上,截至第12天,大于半数的扩增细胞是t细胞。此外,截至第31天。培养物中剩余细胞中仅10%是nk细胞(图3a和补充图s4a)。此外,当在扩增的31-36天内测定在癌症患者的情况下的经扩增nk细胞和t细胞的总数时,在相较于健康对照时,存在来自癌症患者的较少扩增细胞(图3d和补充图s4c),并且扩增t细胞的水平显著高于nk细胞(图3e-图3f和补充图s4d-补充图s4e)。相比之下,从健康供者分离的nk细胞维持nk细胞的扩增,并且nk扩增水平显著高于t细胞(图3b、图3e、图3f和补充图s4b、补充图s4d、补充图s4e)。在不同培养天数时,当相较于健康nk细胞时,在患者nk细胞的情况下观察到nk细胞不增殖或低得多的增殖(图3e和图s4d)。不可观察到来自健康供者或患者的扩增细胞的显著细胞死亡,但相比于健康供者,来自患者的细胞的死亡率略微更高(图3c)。
当相较于与oc一起培养的健康nk细胞时,与oc一起培养的患者nk细胞以显著更小程度使oscsc溶解(图3g和补充图s4f)。当基于nk细胞的数目加以标准化时,在相较于来自健康对照的nk细胞时,患者的每个nk细胞的诱导细胞毒性更小(图3h和补充图s4g)。当相较于经oc扩增健康nk细胞时,经oc扩增患者nk细胞分泌显著更少ifn-γ(图3i和补充图s4h)。当相较于健康nk细胞时,经oc扩增口腔癌患者nk细胞分泌显著更少il-10(图3j),而当相较于健康nk细胞时,来自胰腺癌患者的那些nk细胞分泌更高il-10(补充图s4i)。对于由健康nk细胞或癌症患者nk细胞达成的il-6分泌水平,不可观察到显著差异(图3k和补充图s4j)。相较于由破骨细胞扩增的患者nk细胞,健康nk细胞上的nkg2d表面表达水平是类似的(图3l)。相较于健康对照,在患者nk细胞的表面上,cd94表达强度更高(图3l)。当相较于健康nk细胞时,在经oc扩增患者nk细胞的表面上,kir2、nkp30、nkp44和nkp46表达更低(图3l),而当相较于健康nk细胞时,在经oc扩增患者nk细胞的表面上,kir3表达是相同的或更低的(图3l)。
经破骨细胞活化nk细胞使cd8+t细胞数目实质上增加
当相较于健康对照时,癌症患者平均来说具有更高百分比的cd8+t细胞,以及更低百分比的cd4+t细胞(图7a)。当与破骨细胞一起培养时,在不存在nk细胞下,t细胞未能达成cd8+t细胞扩增,然而,含有不可检测或极小部分的污染性t细胞的用oc活化的纯化nk细胞使来自健康培养物与患者培养物两者的cd8+t细胞扩增,即使相比于健康nk细胞,患者nk细胞培养物使t细胞更快扩增(图7b)。当相较于从健康对照分离的t细胞时,从患者分离的t细胞具有更高水平的cd45ro和更低cd45ra、cd62l、cd28、ccr7和cd127(图7c)。
经oc活化nk细胞优先扩增cd8+t细胞,而经dc活化nk细胞使cd4+t细胞扩增
为确定在由经oc活化nk细胞相对于经dc活化nk细胞扩增的t细胞的子群体之间是否存在差异,我们分析了在健康供者的情况下cd4和cd8子群体。经oc活化nk细胞优先扩增cd8+t细胞,而经dc活化nk细胞使cd4+t细胞扩增(图7d-图7i)。由经oc活化nk细胞扩增的cd8+t细胞展现较高cd45ro和cd44,低得多的水平的cd62l、ccr7和cd127,以及中等水平的cd28,而由经dc活化nk细胞扩增的cd4+t细胞展现较低水平的cd45ro,中等水平的cd44和较高水平的cd62l、ccr7,其中在cd127方面具有少许变化,以及较低水平的cd28(图7j)。在不存在nk细胞下,由oc或dc活化的t细胞展现的表面概况类似于由经nk活化dc获得的那些,例外之处是cd28表达类似于由经oc活化nk获得的cd28表达(图7j)。在pbmc与在不存在nk细胞下由oc或dc扩增的那些t细胞之间,cd4和cd8在cd3+t细胞内的比例是类似的,并且不可观察到在存在或不存在nk细胞下由oc或dc活化的t细胞上的显著水平的pd-1、tim3或klrg-1(图7k-图7m)。
实施例4:用于实施例5的材料和方法
细胞系、试剂和抗体
补充以10%胎牛血清(fbs)(geminibio-products,ca)的rpmi1640用于免疫细胞的培养。oscsc和oscc在ucla从患者的舌部肿瘤解离并加以生长,并且用补充以10%fbs的rpmi1640培养。重组il-2从nih-brb获得。用于这个研究中的流式细胞计量术抗体从biolegend(sandiego,ca)获得。单克隆抗tnf-a抗体和单克隆抗ifn-g抗体从商业来源获得,或在我们的实验室中制备,并且发现1:100稀释度是用于如先前所述的阻断实验的最优浓度。
人nk细胞和单核细胞的纯化
获得由ucla机构审查委员会(irb)核准的书面知情同意书,并且所有程序都由ucla-irb核准。分离来自健康人供者380的pbmc,并且使用从stemcelltechnologies获得的分离试剂盒纯化nk细胞和单核细胞,如之前所述。基于流式细胞计量分析,发现nk细胞群体和单核细胞群体的纯度分别是95%或更高。
益生细菌
aj2是用于诱导干细胞的分化的8种不同革兰氏阳性益生细菌菌株(嗜热链球菌、长双歧杆菌、短双歧杆菌、婴儿双歧杆菌、嗜酸乳酸杆菌、植物乳酸杆菌、干酪乳酸杆菌(lactobacilluscasei)和保加利亚乳酸杆菌)的组合,并且所述菌株由于它们诱导在nk细胞的情况下促炎性细胞因子与消炎性细胞因子两者的最优分泌的优越能力而加以选择。此外,使各菌株生长,并且在三轮亚克隆之后基于经得住环境压力诸如温度和酸性的能力来选择特定菌落。
破骨细胞的产生以及超动力人源性nk细胞和hu-blt源性nk细胞的扩增
在含有m-csf(25ng/ml)和rankl(25ng/ml)的α-mem培养基400中培养纯化单核细胞21天,或另外加以指定。每3天将培养基用含有m-csf和rankl的新鲜α-mem更新。人纯化nk细胞和hu-blt增浓nk细胞用rh-il-2(1000u/ml)和抗cd16mab(3ug/ml)活化18–20小时,随后使它们与破骨细胞和经声波处理aj2一起共培养。如先前所述每三天将培养基用rh-il-2更新。因为当相较于其他扩增方法时,用破骨细胞和saj2离体扩增的nk细胞具有优越细胞毒性和ifn-g分泌,并且持续较长时期存活,所以它们被称为超动力nk细胞。对于声波处理,将aj2称重,并且在10mg/ml的浓度下再混悬于含有10%fbs的rpmi1640培养基中。使细菌充分涡旋,接着在冰上在6至8幅值下进行声波处理15秒。接着在冰上孵育经声波处理样品30秒。在每五个脉冲之后,获取样品以在显微镜下观察直至至少80%的细胞壁被溶解。确定的是进行约20轮声波处理/冰上孵育以实现完全声波处理。最后,将经声波处理样品(saj2)等分,并且储存在-80摄氏度冷冻器中直至使用。
分析免疫缺陷和人源化小鼠中的人口腔癌细胞生长
在ucla动物研究委员会(arc)的书面核准(2012-101-13a)下进行动物研究。如先前所述在我们的核心研究室中制备人源化blt(hu-blt;人骨髓/肝/胸腺)小鼠。
通过在口底中将肿瘤细胞原位植入8–10周龄nsg小鼠或hu-blt小鼠中来进行口腔肿瘤的体内生长。使用异氟烷使小鼠麻醉,并且接着通过采用10plhc基质胶(corning,ny,usa)在口底中直接注射来转移肿瘤细胞。在手术之后7至10天,小鼠通过尾部静脉来接受
从hu-blt和nsg小鼠的组织解离和培养细胞
为制备小鼠组织的单细胞混悬液以进行后续分析,将动物处死,获得bm、脾、外周血液和口腔肿瘤。立刻将口腔肿瘤切割成1mm3片块并放置至在dmem中含有1mg/ml胶原酶ii(针对脂肪组织)、10u/mldna酶i和1%牛血清白蛋白的消化缓冲液中,并且在37℃烘箱处在150rpm振荡器上孵育20分钟。在消化之后,使样品经40pm细胞滤网过滤,并且在4℃下在1,500rpm下离心10分钟。将集结块再混悬于dmem中,并且进行细胞计数。如先前所述通过使组织消化来获得来自bm和脾的单细胞混悬液。使用对肝素化血液试样进行的ficoll-hypaque离心来获得pbmc。收集含有pbmc的白细胞层,洗涤,并且再混悬于rpmi1640培养基中。
从hu-blt小鼠纯化nk细胞、cd3t细胞和单核细胞
使用t细胞选择试剂盒(stem-celltechnologies)来从hu-blt脾细胞分离cd3ct细胞,并且消减了t细胞的细胞用作富含nk的细胞;使用人cd56c正性选择试剂盒(stem-celltechnologies,canada)来分离来自hu-blt小鼠的nk细胞。使用人cd14正性选择试剂盒(ebioscience,sandiego,ca)来从bm细胞分离来自hu-blt小鼠的单核细胞。
elisa
如先前所述进行单一elisa。为分析和获得细胞因子和趋化因子浓度,通过两倍或三倍稀释由制造商提供的重组细胞因子来产生标准曲线。
表面染色和细胞死亡测定
如先前所述,通过用抗体或碘化丙锭标记细胞来进行染色。将细胞用含有1%bsa的冰冷pbs洗涤两次。将预定最优浓度的特定人单克隆抗体添加至含1x104个细胞的50pl冷bsa中,并且在冰上孵育细胞30分钟。此后,将细胞于冷pbs-bsa中洗涤,并且用pbs-bsa致使达到500pl。对于细胞死亡测定,将处于50pl冷bsa中的1x104个细胞用8pg/ml碘化丙锭染色,并且在冰上孵育细胞10分钟,并用pbs-bsa致使达到500pl。使用beckman485coulterepicsxl细胞仪(brea,ca)进行流式细胞计量术分析,并且在flowjovx软件(ashland,or)中分析结果。
51cr释放细胞毒性测定
如先前所述进行51cr释放测定。简要来说,使不同数目的效应细胞与经51cr标记靶标细胞一起孵育。在4小时孵育时期之后,从各样品收集上清液,并且使用γ计数器加以计数以测定释放的放射性。如下计算特异性细胞毒性百分比:
细胞毒性%d实验cpm--自发性cpm总体cpm--自发性cpm
通过使用为使30%的肿瘤靶标细胞溶解所需的效应细胞数目的倒数来计算lu30/106。
用nk细胞上清液进行的干细胞分化
用抗cd16单克隆抗体(3pg/ml)和il-2(1,000u/ml)的组合处理nk细胞18小时,随后移除上清液,并且用于分化实验。用ifn-yelisa(biolegend,ca,usa)测量由经活化nk细胞产生的ifn-y的量。以逐渐每日添加递增量的nk细胞上清液来使oscsc分化。平均来说,为诱导分化,持续4天添加含有总计3,500pgifn-y的上清液以诱导oscscs的分化和对nk细胞介导的细胞毒性的抗性。之后,将靶标细胞用1xpbs洗涤,使其脱离,并且用于如先前所述的实验。
统计分析
进行未配对双尾史都登t检验以达成统计分析。采用邦弗伦尼事后检验的单因素anova用于比较不同组。(n)表示用于实验的小鼠的数目。对于细胞毒性和细胞因子分析,一式两份或一式三份样品用于评估。以下符号表示各分析内的统计显著性水平,***(p值<0.001),**(p值0.001-0.01),*(p值0.01–0.05)。
实施例5:超动力nk细胞抑制人源化blt小鼠中干细胞样/不良分化口腔肿瘤的体内生长和进展
单次注射超动力nk细胞抑制oscsc肿瘤生长,并且显著改进小鼠的健康状况
产生hu-blt小鼠,并且将人oscsc植入nsg和hu-blt小鼠的口底中(图62a和图62b),并基于每周来监测重量减轻(图62c)。单次注射超动力nk细胞导致用oscsc植入的小鼠的重量减轻较低(图62c)。用oscsc植入并用nk细胞注射的小鼠不展现致病性,并且能够摄取食物;而患有口腔肿瘤的小鼠在不存在nk注射下变为病态,由于肿瘤生长而患有在摄取食物方面的并发症(数据未显示),并且展现快速重量减轻(图62c)。引起关注的是,当相较于携带肿瘤的nsg小鼠时,携带肿瘤的hu-blt小鼠在不注射nk下具有较小重量减轻,从而指示经重构人免疫细胞能够略微而非高效限制肿瘤生长(图62c)。当在肿瘤切除之后比较肿瘤尺寸时,也观察到nk注射在hu-blt小鼠中的治疗作用。相比于经nk注射的携带肿瘤的hu-blt小鼠的那些,来自未进行nk注射的携带肿瘤的hu-blt小鼠的肿瘤是大得多的(图62d和图62e)。相较于在未接受nk治疗的携带肿瘤的小鼠中形成的大肿瘤(图62d–图62f),在经nk注射小鼠或经nk注射/经aj2喂食小鼠中,肿瘤重量实质上在更小程度上得以保持(图62f)。此外,与重量减轻数据一致,当相较于携带肿瘤的nsg小鼠时,携带肿瘤的hu-blt小鼠在不注射nk下具有略微更小肿瘤,从而指示经重构人免疫细胞能够略微而非高效限制肿瘤生长(数据未显示)。
当考虑污染性小鼠cd45c免疫细胞时,hu-blt小鼠展现以hcd45c免疫细胞在bm、脾和100外周血液中达成大于96%-99%重构。hu-blt小鼠的外周血液中cd3ct细胞(图16g)、cd3ccd4ct细胞(图16h)和cd3ccd8ct细胞(图62i)的概况类似于人外周血液的概况;然而,相较于人,在hu-blt小鼠中,b细胞的百分比略微更高(图62j),而nk细胞的百分比较小(图62k)。
在携带肿瘤的小鼠中在所有组织区室内,nk细胞毒性和ifn-y分泌都有损失,并且在注射nk和/或用aj2喂食以及注射pd1抗体的情况下得以恢复
在脾细胞(图63a)、bm源性免疫细胞(图63b)、pbmc(图63c)和cd3+经消减脾细胞(图63d)中,携带肿瘤的小鼠展现较低nk介导细胞毒性。单独用nk注射的携带肿瘤的小鼠与组合用aj2喂食的经nk注射的携带肿瘤的小鼠两者均在所有组织区室中都展现nk细胞毒性升高,其中当对小鼠喂以aj2,用nk细胞和抗pd1抗体注射时观察到最高增加(图63a-图64d)。当通过cd3+正性选择试剂盒来使t细胞从脾细胞消减时,可观察到cd3+经消减脾细胞中如在图63a中所观察到的类似nk细胞毒性概况(图63d)。125携带肿瘤的hu-blt小鼠的脾细胞(图63e和表2)、bm源性免疫细胞(图63f)、pbmc(图63g)和来自脾细胞的cd3+t细胞(图63h)分泌较低ifn-γ。当相较于nk和aj2群组时,与nk细胞和aj2组合注射抗pd1抗体对所有组织区室中的ifn-γ分泌都不具有影响或具有降低的影响(图63e-图63g)。经nk注射的携带肿瘤的小鼠在所有组织区室内都显示ifn-γ分泌升高,其中当也对小鼠喂以aj2时观察到最高增加(图63e-图63h)。当通过cd3+正性选择试剂盒来使t细胞从脾细胞消减时,相较于在不存在nk注射下的携带肿瘤的小鼠或经nk注射的非携带肿瘤的对照小鼠,接受nk注射的携带肿瘤的小鼠具有最高ifn-γ分泌增加(图63i)。
表2
当hu-blt小鼠用nk细胞注射以及喂以aj2时,cd3+cd8+t细胞在各种组织区室中的比例增加
在用单独nk细胞注射或组合以aj2喂食的小鼠的bm(图64a)、脾(图64b)和血液(数据未显示)中,观察到cd3+t细胞或cd3+cd8+t细胞的比例增加(表3和表4)。当考虑在处死时的t细胞百分比时,在用单独nk细胞注射或组合以aj2喂食的携带肿瘤的小鼠中,pbmc、脾和bm展现较高cd3+t细胞百分比,并且bm展现hladr+cd11b+免疫子组升高(表3)。当培养bm和脾细胞时,在用单独nk细胞注射或组合以aj2喂食的携带肿瘤的小鼠中,cd3+t细胞和cd3+cd8+t细胞的百分比增加(表3)。类似地,从用nk注射和/或喂以aj2的携带肿瘤的小鼠解离的肿瘤显示在处死时cd3+t细胞的水平升高(表4)。
表3
表4
nk注射和/或用aj2喂食在hu-blt小鼠中在体内抑制干细胞样口腔肿瘤的生长和进展,并且使csc分化
来自在不注射nk的情况下的nsg(图65a)和hu-blt小鼠的肿瘤细胞快速生长,而在nk细胞的情况下的那些不生长或极缓慢生长(图65a–图65c)。类似地,来自用经nk上清液分化肿瘤植入的hu-blt小鼠的肿瘤不生长(图65c),并且在植入之前用抗ifn-γ抗体和抗tnf-α抗体的组合阻断肿瘤分化使体内肿瘤生长恢复(图65b和图65c)。相较于单独经nk注射小鼠,喂以aj2的经nk注射小鼠中的肿瘤生长程度更小(图65c),并且两者均实质上小于仅接受口腔肿瘤植入的那些(图65c)。当相较于单独肿瘤小鼠时,9至11倍多的hcd45c免疫细胞浸润在用nk细胞注射的携带肿瘤的小鼠的肿瘤中(图65d)。当相较于经nk注射的携带肿瘤的小鼠时,来自未进行nk注射的携带肿瘤的小鼠的肿瘤中的表达表面epcam的上皮细胞的百分比是约5倍高。
当相较于来自携带肿瘤的hu-blt或nsg小鼠的那些时,在不同培养天数时,从用nk细胞注射的携带肿瘤的小鼠的肿瘤获得显著更高ifn-g分泌(图65e)。相较于仅接受肿瘤的hu-blt和nsg小鼠,在从经nk注射的携带肿瘤的小鼠获得的肿瘤上,cd54和mhc-i的表达更高(图65f)。当相较于来自未进行nk注射的携带肿瘤的小鼠的肿瘤时,来自在不存在以及存在用aj2喂食下的经nk注射的携带肿瘤的小鼠的肿瘤对nk细胞介导的细胞毒性具有高度抗性,从而显示它们的分化表型(图65g和图65h)。当在植入之前用抗ifn-γ抗体和抗tnf-α抗体阻断nk介导的对肿瘤细胞的分化时,肿瘤对nk细胞介导的细胞毒性的易感性得以恢复(图65h)。当基于来自携带肿瘤的小鼠的vegf的分泌加以标准化时,来自用nk细胞注射的hu-blt小鼠的口腔肿瘤分泌相对较少vegf(图65i)。来自经nk注射hu-blt小鼠的肿瘤具有较高水平的hcd45c浸润性淋巴细胞,并且最高增加见于用nk细胞注射并用aj2喂食的那些小鼠中(图65j)。大多数浸润性cd45c免疫细胞是cd3+t细胞,其中相比于cd8+t细胞,cd4+t细胞子组具有适度更高比例,并且显示cd3+cd56+cd16cnkt子组。当用il-2处理肿瘤时,在与肿瘤相互作用期间被下调的高强度cd4、cd8和cd16/cd56表面表达得以恢复。
当相较于来自无nk细胞的携带肿瘤的hu-blt小鼠的那些时,来自经nk注射的携带肿瘤的hu-blt小鼠的外周血液的血清展现增加的ifn-γ分泌以及其他细胞因子/趋化因子/生长因子和配体,并且特定来说,ifn-γ分泌通过对小鼠喂以aj2而进一步增强(图66a和图66c)。
cddp或伴有和不伴有nac的太平洋紫杉醇诱导用经il-2+抗cd16mab处理nk上清液分化的oscsc的显著细胞死亡
用nk上清液使oscsc分化导致肿瘤对cddp的显著易感性(图67a)。类似地,太平洋紫杉醇介导经nk上清液分化oscsc的较高细胞死亡,并且nac使太平洋紫杉醇介导的细胞死亡显著增加(图67b)。用抗ifn-γ抗体和抗tnf-α抗体阻断nk介导的oscsc分化使由cddp或伴有或不伴有nac的太平洋紫杉醇诱导的细胞死亡实质上降低(图67b)。用cddp或太平洋紫杉醇和nac处理患者源性分化口腔肿瘤oscc展现较高细胞死亡。
当相较于来自在不存在nk注射下的携带肿瘤的小鼠的那些时,来自经nk注射的携带肿瘤的小鼠的单核细胞和破骨细胞具有使nk细胞活化的更大能力
当相较于在不存在nk细胞注射下用未分化肿瘤植入的那些时,来自经nk注射的携带肿瘤的小鼠和携带经nk上清液分化肿瘤的小鼠的nk细胞在与自体单核细胞一起培养时具有显著增大的细胞毒性(图68a)和ifn-γ分泌(图68b)。通过在植入之前添加针对tnf-α和ifn-γ的抗体来阻断nk介导的肿瘤分化使nk细胞的细胞毒性(图68a)和ifn-γ分泌(图68b)降低。我们接着从hu-blt小鼠的bm源性单核细胞产生oc,并且使其与来自健康人供者的同种异体nk细胞一起培养以研究nk细胞扩增的程度。当相较于从未进行nk注射的携带肿瘤的小鼠获得的那些时,在经nk注射的携带肿瘤的小鼠或携带经nk上清液分化肿瘤的小鼠的情况下,nk细胞扩增(图68c)和ifn-g分泌(图68d和图68e)得以增大(图68c-图68e)。因此,相比于来自在不存在nk注射下的携带肿瘤的小鼠的那些,来自经nk注射的携带肿瘤的小鼠的单核细胞和破骨细胞具有使nk细胞活化的更大能力。
参考文献
1.jewetta,manyg,cacalanon,kosj,tsenghc:naturalkillercellsaseffectorsofselectionanddifferentiationofstemcells:roleinresolutionofinflammation.journalofimmunotoxicology2014,11(4):297-307.
2.tsenghc,buiv,manyg,cacalanon,jewetta:inductionofsplitanergyconditionsnaturalkillercellstopromotedifferentiationofstemcellsthroughcell-cellcontactandsecretedfactors.frontiersinimmunology2014,5:269.
3.burkes,lakshmikantht,coluccif,carbonee:newviewsonnaturalkillercell-basedimmunotherapyformelanomatreatment.trendsinimmunology2010,31(9):339-345.
4.larsensk,gaoy,basseph:nkcellsinthetumormicroenvironment.criticalreviewsinoncogenesis2014,19(1-2):91-105.
5.imaik,matsuyamas,miyakes,sugak,nakachik:naturalcytotoxicactivityofperipheral-bloodlymphocytesandcancerincidence:an11-yearfollow-upstudyofageneralpopulation.lancet2000,356(9244):1795-1799.
6.brunoa,ferlazzog,albinia,noonandm:athinktankoftink/tanks:tumor-infiltrating/tumor-associatednaturalkillercellsintumorprogressionandangiogenesis.journalofthenationalcancerinstitute2014,106(8):dju200.
7.grosse,sunwoojb,buijd:cancerimmunosurveillanceandimmunoeditingbynaturalkillercells.cancerjournal(sudbury,mass)2013,19(6):483-489.
8.mirjacicmartinovickm,babovicn,dzodicrr,jurisicvb,tanicnt,konjevicgm:decreasedexpressionofnkg2d,nkp46,dnam-1receptors,andintracellularperforinandstat-1effectormoleculesinnkcellsandtheirdimandbrightsubsetsinmetastaticmelanomapatients.melanomaresearch2014,24(4):295-304.
9.gubbelsja,felderm,horibatas,belisleja,kapura,holdenh,petries,migneaultm,rancourtc,connorjpetal:muc16providesimmuneprotectionbyinhibitingsynapseformationbetweennkandovariantumorcells.molecularcancer2010,9:11.
10.balsamom,scordamagliaf,pietrag,manzinic,cantonic,boitanom,queirolop,vermiw,facchertif,morettaaetal:melanoma-associatedfibroblastsmodulatenkcellphenotypeandantitumorcytotoxicity.proceedingsofthenationalacademyofsciencesoftheunitedstatesofamerica2009,106(49):20847-20852.
11.castriconir,cantonic,dellachiesam,vitalem,marcenaroe,conter,biassonir,bottinoc,morettal,morettaa:ttansforminggrowthfactorbeta1inhibitsexpressionofnkp30andnkg2dreceptors:consequencesforthenk-mediatedkillingofdendriticcells.proceedingsofthenationalacademyofsciencesoftheunitedstatesofamerica2003,100(7):4120-4125.
12.pietrag,manzinic,rivaras,vitalem,cantonic,petrettoa,balsamom,conter,benellir,minghellisetal:melanomacellsinhibitnaturalkillercellfunctionbymodulatingtheexpressionofactivatingreceptorsandcytolyticactivity.cancerresearch2012,72(6):1407-1415.
13.krockenbergerm,dombrowskiy,weidlerc,ossadnikm,honiga,hauslers,voigth,beckerjc,lengl,steinleaetal:macrophagemigrationinhibitoryfactorcontributestotheimmuneescapeofovariancancerbydown-regulatingnkg2d.journalofimmunology(baltimore,md:1950)2008,180(11):7338-7348.
14.vitalem,cantonic,pietrag,mingarimc,morettal:effectoftumorcellsandtumormicroenvironmentonnk-cellfunction.europeanjournalofimmunology2014,44(6):1582-1592.
15.galloisa,silvai,osmani,bhardwajn:reversalofnaturalkillercellexhaustionbytim-3blockade.oncoimmunology2014,3(12):e946365.
16.herseyp,edwardsa,honeymanm,mccarthywh:lownatural-killer-cellactivityinfamilialmelanomapatientsandtheirrelatives.britishjournalofcancer1979,40(1):113-122.
17.perussiab,ramonic,anegoni,cuturimc,faustj,trinchierig:preferentialproliferationofnaturalkillercellsamongperipheralbloodmononuclearcellscoculturedwithblymphoblastoidcelllines.naturalimmunityandcellgrowthregulation1987,6(4):171-188.
18.rabinowichh,sedlmayrp,herbermanrb,whitesidetl:increasedproliferation,lyticactivity,andpurityofhumannaturalkillercellscoculturedwithmitogen-activatedfeedercells.cellularimmunology1991,135(2):454-470.
19.igarashit,wynbergj,srinivasanr,becknellb,mccoyjp,jr.,takahashiy,suffredinida,linehanwm,caliglurima,childsrw:enhancedcytotoxicityofallogeneicnkcellswithkillerimmunoglobulin-likereceptorligandincompatibilityagainstmelanomaandrenalcellcarcinomacells.blood2004,104(1):170-177.
20.srivastavas,lundqvista,childsrw:naturalkillercellimmunotherapyforcancer:anewhope.cytotherapy2008,10(8):775-783.
21.grasnavarroa,
22.alicie,sutlut,bjorkstrandb,gilljamm,stellanb,nahih,quezadahc,gahrtong,ljunggrenhg,dilberms:autologousantitumoractivitybynkcellsexpandedfrommyelomapatientsusinggmp-compliantcomponents.blood2008,111(6):3155-3162.
23.fujisakih,kakudah,shimasakin,imaic,maj,lockeyt,eldridgep,leungwh,campahad:expansionofhighlycytotoxichumannaturalkillercellsforcancercelltherapy.cancerresearch2009.69(9):4010-4017.
24.bergm,lundqvista,mccoyp,jr,samsell,fany,tawaba,childsr:clinical-gradeexvivo-expandedhumannaturalkillercellsup-regulateactivatingreceptorsanddeathreceptorligandsandhaveenhancedcytolyticactivityagamsttumorcells.cytotherapy2009,11(3):341-355.
25.gargtk,szmaniasm,khanja,hoerlnga,malbroughpa,moreno-bosta,greenwayad,lingojd,lix,yaccobysetal:highlyactivatedandexpandednaturalkillercellsformultiplemyelomaimmunotherapy.haematologica2012,97(9):1348-1356.
26.imaic,iwamotos,campanad:geneticmodificationofprimarynaturalkillercellsovercomesinhibitorysignalsandinducesspecifickillingofleukemiccells.blood2005,106(1):376-383.
27.laptevan,durettag,sunj,rollinsla,huyell,fangj,dandekarv,meiz,jacksonk,verajetal:large-scaleexvivoexpansionandcharacterizationofnaturalkillercellsforclinicalapplications.cytotherapy2012,14(9):1131-1143.
28.koepsellsa,millerjs,mckennadh,jr.:naturalkillercells:areviewofmanufacturingandclinicalutility.transfusion2013,53(2):404-410.
29.millerjs.soigniery,panoskaltsis-mortaria,mcnearneysa,yungh,fautschsk,mckennad,lec,deforte,burnsljetal:successfuladoptivetransferandinvivoexpansionofhumanhaploidenticalnkcellsinpatientswithcancer.blood2005,105(8):3051-3057.
30.iliopouloueg,kountourakisp,karamouzismv,doufexisd,ardavanisa,baxevaniscn,rigatosg,papamichailm,perezsa:aphaseitrialofadoptivetransferofallogeneicnaturalkillercellsinpatientswithadvancednon-smallcelllungcancer.cancerimmunoiogy,immunotherapy:cii2010,59(12):1781-1789.
31.sotiropouloupa,perezsa,gritzapisad,baxevaniscn,papamichailm:interactionsbetweenhumanmesenchymalstemcellsandnaturalkillercells.stemcells2006,24(1):74-85.
32.gellerma,cooleys,judsonpl,ghebrer,carsonlf,argentapa,jonsonal,panoskaltsis-mortaria,curtsingerj,mckennadetal:aphaseiistudyofallogeneicnaturalkillercelltherapytotreatpatientswithrecurrentovarianandbreastcancer.cytotherapy2011,13(1):98-107.
33.tsenghc,arasteha,paranjpea,teruela,yangw,behela,alvaja,walterg,headc,ishikawatoetal:increasedlysisofstemcellsbutnottheirdifferentiatedcellsbynaturalkillercells;de-differentiationorreprogrammingactivatesnkcells.plosone2010,5(7):e11590.
34.tsenghc,inagakia,buivt,cacalanon,kasaharan,manyg,jewetta:differentialtargetingofstemcellsanddifferentiatedglioblastomasbynkcells.journalofcancer2015,6(9):866-876.
35.buivt,tsengh-c,maungpo,kozlowskaa,mannk,topchyanp,jewetta:augmentedifn-γandtnf-αinducedbyprobioticbacteriainnkcellsmediatedifferentiationofstem-liketumorsleadingtoinhibitionoftumorgrowthandreductionininflammatorycytokinerelease;regulationbyil-10.frontiersinimmunology2015,6.
36.buivt,tsenghc,kozlowskaa,maungpo,kaurk,topchyanp,jewetta:augmentedifn-gammaandtnf-alphainducedbyprobioticbacteriainnkcellsmediatedifferentiationofstem-liketumorsleadingtoinhibitionoftumorgrowthandreductionininflammatorycytokinerelease;regulationbyil-10.frontimmunol2015,6:576.
37.jewetta,bonavidab:target-inducedinactivationandcelldeathbyapoptosisinasubsetofhumannkcells.journalofimmunology(baltimore,md:1950)1996,156(3):907-915.
38.shimizus,hongp,arumugamb,pokomol,boyerj,koizumin,kittipongdajap,chena,bristolg,galiczetal:ahighlyefficientshorthairpinrnapotentlydown-regulatesccr5expressioninsystemiclymphoidorgansinthehu-bltmousemodel.blood2010,115(8):1534-1544.
39.vatakisdn,koyarc,nixoncc,weil,kimsg,avancenap,bristolg,baltimored,kohndb,ribasaetal:antitumoractivityfromantigen-specificcd8tcellsgeneratedinvivofromgeneticallyengineeredhumanhematopoieticstemcells.proceedingsofthenationalacademyofsciencesoftheunitedstaresofamerica2011,108(51):e1408-1416.
40.kozlowskaak,kaurk,topchyanp,jewetta:adoptivetransferofosteoclast-expandednaturalkillercellsforimmunotherapytargetingcancerstem-likecellsinhumanizedmice.cancerimmunology,immunotherapy:cii2016.
41.jewetta,cavalcantim,bonavidab:pivotalroleofendogenoustnf-alphaintheinductionoffunctionalinactivationandapoptosisinnkcells.journalofimmunology(baltimore,md:1950)1997,159(10):4815-4822.
42.jewetta,bonavidab:interferon-alphaactivatescytotoxicfunctionbutinhibitsinterleukin-2-mediatedproliferationandtumornecrosisfactor-alphasecretionbyimmaturehumannaturalkillercells.jclinimmunol1995,15(1):35-44.
43.jewetta,wangmy,teruela,poupakz,bostanianz,parknh:cytokinedependentinverseregulationofcd54(icam1)andmajorhistocompatibilitycomplexclassiantigensbynuclearfactorkappabinhep2tumorcellline:effectonthefunctionofnaturalkillercells.humanimmunology2003,64(5):505-520.
44.tsenghc,kanayamak,kaurk,parksh,parks,kozlowskaa,suns,mckennace,nishimurai,jewetta:bisphosphonate-induceddifferentialmodulationofimmunecellfunctioningingivaandbonemarrowinvivo:roleinosteoclast-mediatednkcellactivation.oncotarget2015,6(24):20002-20025.
45.tamyk,martinsonja,doligosak,klingemannhg:exvivoexpansionofthehighlycytotoxichumannaturalkiller-92cell-lineundercurrentgoodmanufacturingpracticeconditionsforclinicaladoptivecellularimmunotherapy.cytotherapy2003,5(3):259-272.
46.magisisters,tsenghc,buivt,kosj,jewetta:regulationofsplitanergyinnaturalkillercellsbyinhibitionofcathepsinscandhandcystatinf.oncotarget2015,6(26):22310-22327.
47.perisicnanutm,saboticj,jewetta,kosj:cysteinecathepsinsasregulatorsofthecytotoxicityofnkandtcells.frontimmunol2014,5:616.
48.cantonic,huergo-zapicol,parodim,pedrazzim,mingarimc,morettaa:nkcells,tumorcelltransition,andtumorprogressioninsolidmalignancies:newhintsfornk-basedimmunotherapy?2016,2016:4684268.
49.tallericor,garofaloc,carbonee:anewbiologicalfeatureofnaturalkillercells:therecognitionofsolidtumor-derivedcancerstemcells.frontimmunol2016,7:179
50.kamiyat,changyh,campanad:expandedandactivatednaturalkillercellsforimmunotherapyofhepatocellularcarcinoma.cancerimmunologyresearch2016.
以引用的方式并入
本文提及的所有出版物、专利和专利申请都据此以引用的方式整体并入本文,所述引用的程度就好像各个别出版物、专利或专利申请被特定地以及个别地指示以引用的方式并入一样。在起冲突的情况下,将以包括本文任何定义的本申请为准。
也以引用的方式整体并入本文的是引用与公共数据库中的条目关联的登录号的任何多核苷酸和多肽序列,诸如由基因组研究所(theinstituteforgenomicresearch,tigr)在万维网上在tigr.org处和/或国家生物技术信息中心(nationalcenterforbiotechnologyinformation,ncbi)在万维网上在ncbi.nlm.nih.gov处维持的那些。
等效物
本领域技术人员将仅使用常规实验就会认识到或能够确定本文所述的本发明的特定实施方案的许多等效物。所述等效物意图由以下权利要求涵盖。
- 还没有人留言评论。精彩留言会获得点赞!