乙酰葡萄糖胺磷酸变位酶AtAGM编码基因及其酶、制备、应用与酶活性检测方法与流程
本发明涉及一种乙酰葡萄糖胺磷酸变位酶AtAGM的基因序列及其制备方法,尤其涉及该酶在核苷酸糖及磷酸己糖异构体生产中的应用。本发明还提供了该乙酰葡萄糖胺磷酸变位酶的重组质粒和重组基因工程菌株,并且提供了检测己糖磷酸变位酶活性的新方法以及利用该酶进行磷酸己糖生产和分离的方法。
背景技术:
乙酰葡萄糖胺(GlcNAc,N-乙酰-β-D-氨基葡萄糖)是葡萄糖的一种氨基糖衍生物。在生物体内,是仅次于葡萄糖的一种具有重要生理生化功能的单糖。常以UDP-GlcNAc形式参与主要包括神经系统和免疫系统在内的诸多生物反应过程和生物体内多种糖缀合物的结构组成。在生物体内,生成UDP-GlcNAc的主要途径是氨基己糖途径(Hexosamine biosynthesis pathway)。这一途径是糖酵解的分支,都以己糖代谢途径的中间产物果糖-6-磷酸(Fructose-6-P)为起始底物,在多种酶的协同催化作用下,最终合成UDP-GlcNAc。根据合成过程中催化反应的顺序及合成途径所涉及酶的来源不同,分为真核,原核以及拟菌病毒的UDP-GlcNAc合成途径。目前,氨基己糖途径已经被广泛的研究,但是植物中的氨基己糖途径中的第三个酶乙酰葡萄糖胺磷酸变位酶至今尚未研究和应用,所以植物中的氨基己糖途径仍未被打通。
另外UDP-GlcNAc作为生物体内重要的活性核苷糖,可被大量用于寡糖的生产,进而被开发成医药品或功能性材料。但由于其生产方法的限制,目前其供应量仍较小,价格也很高。目前已经有研究使用无细胞催化的多步酶催化反应体系,偶联了葡萄糖激酶,AGM与GlcNAc-1-P尿苷转移酶(GlmU)三个酶,以GlcNAc作为底物生产UDP-GlcNAc。无论是体内表达体系还是体外表达体系,来自酵母的AGM l的可溶表达水平均很低,影响了产物合成的效率和成本。所以提供一种高活性、高表达的AGM具有十分重要的意义。
磷酸己糖的价格十分的昂贵,目前主要是化学方法合成,生产步骤繁琐。Sigma以及Santa Cruz等公司的储量很少甚至断货。由于合成方法的限制,导致磷酸己糖不同的异构体之间价格差异巨大(几十甚至上百倍);如GlcN-6-P的价格29元/mg,而GlcN-1-P的价格则是460/mg。所以探索新的磷酸己糖的制备方法,特别是价格昂贵的磷酸己糖的制备方法势在必行。
相对于氨基己糖途径中的其他三个酶,己糖磷酸变位酶(真核AGM,原核GlmM)研究相对较少。导致此现象的主要原因是其活性的检测存在一定的困难,一般采用偶联法检测,即偶联此途径中的其他酶,通过检测终产物UDP-GlcNAc的产生来间接反映己糖磷酸变位酶的活性。但是由于偶联的酶只能通过实验室自己制备,其酶活及性质无法保证,使得己糖磷酸变位酶的活性无法准确测定。
针对上述乙酰葡萄糖胺磷酸变位酶的研究现状及其在应用中存在的问题,本发明首次公开了一种来源于拟南芥(Arabidopsis thaliana)的乙酰葡萄糖胺磷酸变位酶(Arabidopsis thaliana N-acetylpHospHoglucosamine mutase,AtAGM)的基因序列及其制备方法。该酶活性高,酶学性质优良,可以通过调节反应条件来控制反应发生方向(正逆反应),并且制备方法简单,易操作,将其应用于无细 胞酶法催化合成UDP-GlcNAc,或生产己糖磷酸异构体,可大大提高UDP-GlcNAc和己糖磷酸异构体的制备效率,降低生产成本,克服现有技术的不足。同时本发明还提供了一种己糖磷酸变位酶的酶活测定方法,与现有方法相比,灵敏度高,结果准确、可靠。
技术实现要素:
本发明的第一个目的是提供一种植物来源的乙酰葡萄糖胺磷酸变位酶AtAGM及其编码基因。
本发明的第二个目的是提供一种制备乙酰葡萄糖胺磷酸变位酶AtAGM的方法。
本发明的第三个目的是提供含有所述的乙酰葡萄糖胺磷酸变位酶AtAGM的重组表达质粒和重组基因工程菌株。
本发明的第四个目的是提供一种新型己糖磷酸变位酶的活性检测方法。
本发明的第五个目的是提供一种乙酰葡萄糖胺磷酸变位酶AtAGM用于UDP-GlcNAc合成生产。
本发明的第六个目的是提供一种制备分离生产磷酸己糖异构体的方法。
本发明所提供的乙酰葡萄糖胺磷酸变位酶AtAGM,来源于拟南芥(Arabidopsis thaliana),其氨基酸序列具有如下特征中的一种或二种:
1)序列表中的SEQ ID NO.2从氨基端开始的第1-556氨基酸残基序列,为有活性的乙酰葡萄糖胺磷酸变位酶AtAGM的氨基酸序列,556-564为His-Tag的氨基酸序列。
2)将序列表中的SEQ ID NO.2从氨基端开始的第1-556位氨基酸残基进行一个或两个以上氨基酸取代、缺失或添加而形成具有乙酰葡萄糖胺磷酸变位酶活性不变的氨基酸序列。
本发明还提供了上述乙酰葡萄糖胺磷酸变位酶AtAGM的编码基因,来源于拟南芥(Arabidopsis thaliana),其核苷酸序列具有如下特征中的一种或二种以上:
1)序列表中SEQ ID NO.1的脱氧核糖核酸(DNA)序列;
2)编码序列表中SEQ ID NO.2氨基酸序列的脱氧核糖核酸(DNA)序列;
3)对序列表中SEQ ID NO.1的脱氧核糖核酸(DNA)序列进行一个或两个以上核苷酸取代、缺失或添加而得到的编码具有乙酰葡萄糖胺磷酸变位酶活性的核苷酸序列。
本发明的乙酰葡萄糖胺磷酸变位酶AtAGM的氨基酸序列及其核苷酸编码序列也可以根据预测的AtAGM的氨基酸序列及其核苷酸编码序列人工合成获得。
制备重组酶AtAGM的方法,是将乙酰葡萄糖胺磷酸变位酶AtAGM的编码基因克隆入重组表达载体,导入宿主细胞,获得重组表达的乙酰葡萄糖胺磷酸变位酶
上述乙酰葡萄糖胺磷酸变位酶AtAGM的编码基因,其核苷酸序列具有如下特征中的一种或二种以上:
1)具有序列表中SEQ ID NO.1的脱氧核糖核酸(DNA)序列,
2)编码SEQ ID NO.2氨基酸序列的脱氧核糖核酸(DNA)序列,
3)对序列表中SEQ ID NO.1的脱氧核糖核酸(DNA)序列进行一个或两个以上核苷酸取代、缺失或添加而得到的编码具有乙酰葡萄糖胺磷酸变位酶活性的核苷酸序列。
所述的重组表达乙酰葡萄糖胺磷酸变位酶AtAGM的表达载体可以是大肠杆菌表达载体、酵母表达载体、枯草杆菌表达载体、乳酸菌表达载体、链霉菌表达载体、噬菌体载体、丝状真菌表达载体、植物表达载体、昆虫表达载体、或哺乳动物细胞表达载体等。
用于重组表达乙酰葡萄糖胺磷酸变位酶AtAGM的重组菌或转基因细胞系,可以是大肠杆菌宿主细胞(如Escherichia coli BL21、Escherichia coli JM109、Escherichia coli DH5α等)、酵母菌宿主细胞(如Saccharomyces cerevisiae、Pichia pastoris、Kluyveromyces lactis等)、枯草杆菌宿主细胞(如Bacillus subtilis R25、Bacillus subtilis 9920等)、乳酸菌宿主细胞(如Lactic acid bacteria COCC101等)、放线菌宿主细胞(如Streptomyces spp.等)、丝状真菌宿主细胞(如Trichoderma viride,Trichoderma reesei,Aspergillus niger、Aspergillus nidulans等)、昆虫细胞(如Bombyx mori,Antharaea eucalypti等)或哺乳动物细胞(如中国仓鼠卵巢细胞CHO,幼小仓鼠肾脏细胞BHK、中国仓鼠肺细胞CHL等)。
上述的乙酰葡萄糖胺磷酸变位酶在磷酸己糖与核苷酸糖生产中可以应用,包括以下应用中的一或二种以上:
1)在实现磷酸己糖1,6位异构体的转变,生产相应同分异构体中的应用;
2)在UDP-GlcNAc(UDP-GlcN;UDP-Glc)等核苷酸糖的生产中的应用;
所述乙酰葡萄糖胺磷酸变位酶AtAGM对己糖-1-P在低温下(10-20℃)和碱性PH范围(7-9)活性最高,并抑制逆反应进行,可以用于产生对应的同分异构体己糖-6-P。AtAGM对己糖-6-P在稍高温度(20-30℃)和酸性PH范围(5-7)活性最高,可抑制逆反应进行,可以用于产生对应的同分异构体己糖-1-P或者催化合成相应的核苷酸糖(UDP-GlcNAc,UDP-GlcN,UDP-Glc等)。
使用离子交换色谱对磷酸己糖底物和变位产物的量进行同步检测。具体色谱检测条件为:
所用的离子交换色谱系统是DIONEX ICS-3000,离子交换色谱柱CarboPac PA-100column(4×250mm),与电化学检测器。所用的流动相:A,100mM NaOH水溶液;B,800mM乙酸钠与100mM NaOH水溶液。流动相洗脱条件是0-5min,90%A+10%B;6-15min,10%-90%B;16-18min,10%A+90%B;19-20min,90%A+10%B.总流速是0.5ml/min,检测柱温是30℃,进样体积是20μL。
所述的己糖磷酸变位酶活性检测方法,可以用于检测不同来源的己糖磷酸变位酶如乙酰葡萄糖胺磷酸变位酶(AGM),葡萄糖胺磷酸变位酶(GlmM),葡萄糖磷酸变位酶(PGM)。以及以磷酸己糖作为底物的酶,包括GlN-6-P合成酶(GFA),GlcN-6-P乙酰转移酶(GNA),GlN-1-P乙酰转移酶(GlmU)。
本发明的乙酰葡萄糖胺磷酸变位酶AtAGM的基因序列是通过PCR技术从拟南芥基因组中克隆得到。该基因编码区长1710bp,编码556个氨基酸,属于己糖磷酸变位酶(α-D-pHospHohexomutases)家族。大肠杆菌重组表达获得的AtAGM,蛋白分子量在61.5KDa,对不同的己糖磷酸均有活性。
本发明提供的AtAGM活力高,对GlcNAc-6-P的活力是来源于真菌的AGM的438倍,对GlcNAc-1-P的活力是来源于酵母的AGM的50倍。同时具有良好的可大规模应用的酶学性质,正逆反应具有不同的最适反应温度与不同的最适反应pH,所以可以通过调节反应体系的pH或者反应的温度,来促进目的异构体的积累,可以用于磷酸己糖异构体的生产。
本发明提供的检测己糖磷酸变位酶活性的新方法,操作简便,灵敏度远远超过传统的偶联法,而且还有可检测正、逆反应的巨大优势。此方法对多对磷酸己 糖异构体具有良好的分离效果,不仅可以用于己糖磷酸变位酶活性的检测,还可以用于磷酸己糖异构体的生产分离。
附图说明
图1:乙酰葡萄糖胺磷酸变位酶AtAGM表达及纯化的SDS-PAGE图。各泳道加入的样品分别是:泳道1-E.coli BL21(DE3)/pET23a-AtGAM未诱导菌体;泳道2-E.coli BL21(DE3)/pET23a-AtGAM诱导后菌体;泳道3-预染蛋白分子量标准;泳道4-200mmol/L咪唑洗脱液。
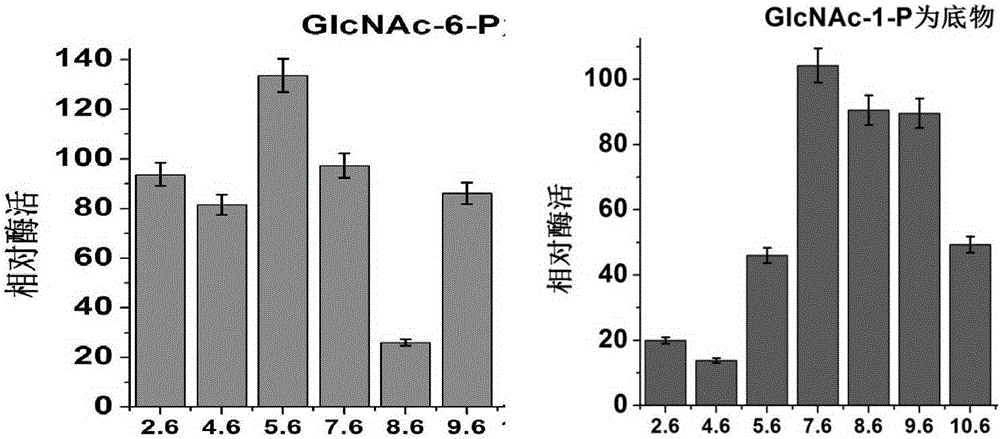
图2:AtGAM酶在不同反应体系pH下的相对活性柱状图。
图3:HAPEC-PAD对不同的磷酸己糖异构体的分离效果。
图4:HAPEC-PAD方法与传统偶联方法的比较效果。
具体实施方式
实施例1 乙酰葡萄糖胺磷酸变位酶AtAGM全长基因的克隆
参照RNA提取试剂盒(博迈德生物,货号RN0112)操作步骤提取拟南芥叶片的mRNA。对The National Center for Biotechnology Information(NCBI)数据库中乙酰葡萄糖胺磷酸变位酶序列分析后,设计引物Agm-F:5’-CACCCCCATATGATGGACGAGATCCAAATAGC-3’;Agm-R:5’-CTCGAGGCTTGAACCAAGGAAGCTTTTGACG-3’扩增编码乙酰葡萄糖胺磷酸变位酶AtAGM的成熟蛋白的基因序列,以提取的拟南芥的RNA反转的cDNA为模板进行PCR扩增。PCR反应条件为:94℃2min,1个循环;94℃30s,55℃30s,72℃2min,30个循环;72℃5min,1个循环。PCR产物进行琼脂糖凝胶电泳分析后,对目的片段进行切胶回收,经双酶切后连接到原核表达载体pET23a上后测序。
实施例2 乙酰葡萄糖胺磷酸变位酶AtAGM基因序列分析
测序的结果采用GenBank数据库中的Basic Local Alignment Search Tool(BLAST)分析,Vector NTI Suite 8.0软件进行多序列比对,分析序列信息。
获得的乙酰葡萄糖胺磷酸变位酶基因(命名为AtAGM)编码区长1698bp,其核苷酸序列如SEQ ID NO 1所示。AtAGM编码564个氨基酸和一个终止密码子,其氨基酸序列如SEQ ID NO 2所示,蛋白质理论分子量为61.5kDa,预测等电点为5.35。AtAGM的核苷酸序列在拟南芥的基因组中被称为DNA-DAMAGE-REPAIR/TOLERATION 101(DRT101),位于拟南芥的五号染色体上(locus-tag="AT5G18070"),具有乙酰葡萄糖胺磷酸变位酶功能,但是仅仅具有序列信息,其活性及性质未被研究。乙酰葡萄糖胺磷酸变位酶AtAGM属于己糖磷酸变位酶(α-D-pHospHohexomutases)家族。
实施例3 AtAGM基因在大肠杆菌中的重组表达及纯化
测序结果表明,在pET23a上插入SEQ ID NO 1所示的AtAGM基因,且插入方向正确,证明了构建的重组质粒正确,将该重组质粒命名为pET23a-AtAGM。
将pET23a-AtAGM转化大肠杆菌菌株BL21(DE3)进行诱导表达。以1%的接种量将过夜培养的种子液加入新鲜的LB培养基中于37℃进行扩大培养,当菌液的OD600nm=0.6-0.8时加入IPTG,使其终浓度为0.5mM,16℃进行过夜诱导。菌体破碎后高速离心取上清进行镍柱纯化,梯度咪唑洗脱(20-500mM咪唑,20mM Tris-HCl,PH7.6)。用聚丙烯酰胺凝胶电泳检测乙酰葡萄糖胺磷酸变位酶AtAGM的表达及纯化情况,电泳使用12%的分离胶,80V电压压平后,换120V电压跑完分离胶。结果如图1所示,纯化后的乙酰葡萄糖胺磷酸变位酶AtAGM 在电泳胶上呈单一条带,且位置与预测的分子量相吻合。
SEQ ID NO.1
ATGGACGAGATCCAAATAGCTTCAATCCTCAAATCATCTGAGCTTTTTCCGATTCCACAAGGCGTCAAGCTTTCGTATGGAACAGCTGGAT-TCAGAGGCGATGCAAAGTTATTGGAATCAACTGTGTATAGAGTTGGGATTCTCTCAGCTCTCCGATCACTTAAGCTTGGATCAGCCACCGTCGGGCTTATGATCACAGCTTCGCATAACAAAGTCTCTGACAATGGCATTAAAGTTTCAGATCCATCTGGTTTTATGCTTTCTCAGGAATGGGAGCCTTTTGCAGATCAGATCGCTAACGCATCTTCTCCTGAAGAACTCGTTTCGTTGATTAGAAAATTCATGGAGAAGGAAGAGATTGCAATCGGAGAGAATAATAAAGGTGCAGAGGTTTGGTTGGGAAGAGATACTAGACCTA-GTGGTGAATCACTTCTCAGAGCTGGTGAGATCGGAGTTGGTTCAATTTTGGGATCTGTTGCGAT-TGACATTGGGATTTTGACAACTCCGCAATTGCATTGGATGGTTAGAGCTAAGAATAAAGGTCTTAAGGCAACTGAGAATGATTACTTTGAGAATCTATCTACTTCGTTTAGGTGTTTGATTGATTTGATTCCAAGCAGTGGAAATGATAAGTTGGAGATTAGCAAATTGCTTGTAGATGGTGCTAACGGTGTAGGTGGACAGAAGATTGAGAAGCTAAGAGGGTCTTTGAGTAATTTAGATGTTGAGATTCGTAACACAGGGAGAGATGGTGGTGTGCTTAATGAAGGTGTAGGTGCTGATTTTGTGCAGAAAGAAAAGGTTTTGCCTGTAGGATTTGGGTTTAAGGATGTTGGGATGAGGTGTGCGAGTTTGGATGGTGATGCAGATCGATTGGTTTACTTTTACATTCCTTCAGATTCTTCTGAAAAGGTTGAGCTACTTGACGGTGATAAGATTCTGTCTTTGTTTGCTCTCTTCATCAAAGAGCAACTAAATGCTCTGGAGGATGATGAAGAAAGGAAGCAGTCTCGTCTTGGTGTTGTGCAGACAGCTTACGCGAATGGTGCGTCTACTGATTACCTAAAGCATTTGGGTTTAGATGTTGTTTTTGCTAAAACTGGAGTTAAGCATTTACACGAGAAAGCAGCAGAGTTTGATATTGGAATCTACTTTGAAGCTAATGGCCACGGGACTATTCTCTTCTCGGAATCTTTCCTATCTTGGTTAGTTTCCAAACAAAAGGATCTTACGGCTAAAGGTCAGGGTGGTTCTGAAGAGCACAAAGCTGTTTCTAGACTAATGGCGGTGAGTAATCTGATTAACCAAGCGGTAGGTGATGCTCTAAGTGGAGTGCTCTTGGTTGAAGTGATTCTACAACACCTGGGATGGTCGATAGAGAAGTGGAATGAGCTATACAAGGACCTTCCTAGCAGGCAGATCAAGGTCGAAGTTCCAGATAGAACAGCGGTTGTGACCACAAGCGAAGAAACCGAGGCTCTGAGACCTATGGGGATTCAAGATGCTATTAATTCTGAAATCAAGAAGTACTCGCGTGGCAGAGCTTTTATAAGGCCATCGGGTACAGAAGATGTGGTGAGAGTATATGCAGAGGCTTCCACTCAAGAAGATGCTGATTCTTTGGCTAATTCTGTGGCTCAGCTCGTCAAAAGCTTCCTTGGTTCAAGCCTCGAGCACCACCACCACCACCACTGA
SEQ ID NO.2
MDEIQIASILKSSELFPIPQGVKLSYGTAGFRGDAKLLESTVYRVGILSALRSLKLGSATVGLMITASHNKVSDNGIKVSDPSGFMLSQEWEPFADQIANASSPEELVSLIRKFMEKEEIAIGENNKGAEVWLGRDTRPSGESLLRAGEIGVGSILGSVAIDIGILTTPQLHWMVRAKNKGLKATENDYFENLSTSFRCLIDLIPSSGNDKLEISKLLVDGANGVGGQKIEKLRGSLSNLDVEIRNTGRDGGVLNEGVGADFVQKEKVLPVGFGFKDVGMRCASLDGDADRLVYFYIPSDSSEKVELLDGDKILSLFALFIKE QLNALEDDEERKQSRLGVVQTAYANGASTDYLKHLGLDVVFAKTGVKHLHEKAAEFDIGIYFEANGHGTILFSESFLSWLVSKQKDLTAKGQGGSEEHKAVSRLMAVSNLINQAVGDALSGVLLVEVILQHLGWSIEKWNELYKDLPSRQIKVEVPDRTAVVTTSEETEALRPMGIQDAINSEIKKYSRGRAFIRPSGTEDVVRVYAEASTQEDADSLANSVAQLVKSFLGSSLEHHHHHH-
实施例4 乙酰葡萄糖胺磷酸变位酶AtAGM的酶学性质
(1)乙酰葡萄糖胺磷酸变位酶AtAGM的活力测定
测定AtAGM酶活的通用体系如下:不同的磷酸己糖底物(GlcNAc-1-P,GlcNAc-6-P,GlcN-1-P,GlcN-6-P,Glc-1-P,Glc-6-P)作为底物,每组底物加入反应体系(300μL):20mM PBS,pH 7.0,10mM MgSO4,20μM Glc-1,6-2P,加入适量的重组酶AtAGM,反应20min。将反应体系立即煮沸后,除蛋白,每组底物加入200μL 200mM NaOH。用HAPEC-PAD方法检测体系中底物的消耗与产物的生成情况。由于AtAGM催化的反应是可逆反应,并且还有中间产物己糖-1,6-2P的产生,所以酶活(nmol/min/mg)用1mg蛋白每分钟消耗的底物量(nmol)来表示。蛋白浓度使用碧云天BCA蛋白浓度测定试剂盒进行测定。
(2)乙酰葡萄糖胺磷酸变位酶AtAGM底物特异性
由于AtAGM底物选择性较广,可以催化GlcNAc-6-P与GlcNAc-1-P;GlcN-6-P与GlcN-1-P;Glc-6-P与Glc-1-P几对异构体之间的转换。并且AtAGM催化的是可逆反应,所以采用以上六种底物来检测AtAGM的底物特异性。采用实施例4(1)中提到的通用体系,以0.1mM不同的磷酸己糖底物(GlcNAc-1-P,GlcNAc-6-P,GlcN-1-P,GlcN-6-P,Glc-1-P,Glc-6-P)作为底物,每组底物反应加入2.5μg AtAGM,于30℃水浴中反应30min.蛋白浓度使用碧云天BCA蛋白浓度测定试剂盒进行测定。
结果如表1所示,在如上的反应条件下,相同的底物浓度,AtAGM对不同底物的活力不同。一般规律是逆反应的活力大于正反应(己糖-1-P的活性高于相应的己糖-6-P)。其中以GlcNAc-1-P的活力最高,而GlcNAc-6-P的活力最低。
表1:乙酰葡萄糖胺磷酸变位酶AtAGM的底物特异性。
(3)温度对重组酶AtAGM的影响
采用实施例4(1)中提到的通用体系,以30μM的GlcNAc-1-P或GlcNAc-6-P为底物,分别在4-80℃下按照标准方法测定重组酶的活性,5μg酶在不同温度(具体温度见表2)下的相对活性如表2A,B所示。以灭活的酶为对照,以反应最高酶活力为100%计算相对酶活。以GlcNAc-6-P作为底物的最适反应温度为30℃(表2A),以GlcNAc-1-P作为底物的最适反应温度是10℃(表2B)。AtAGM催化正逆反应的最适温度不同,可以根据此特性调节反应温度来控制目的异构体的积累,这对应用于生产中具有十分重大的意义。
表2A:温度对重组酶AtAGM活性的影响,正反应:以GlcNAc-6-P作为底物
表2B:温度对重组酶AtAGM活性的影响,逆反应:以GlcNAc-1-P作为底物
(4)pH对重组酶AtGAM的影响
采用实施例4(1)中提到的通用体系,在30℃的条件下,分别以30μM的GlcNAc-1-P或GlcNAc-6-P为底物,在PH3.6-10.6(pH3.6-5.6HAc-NaAc,PH6.6-7.6Na2HPO4-NaH2PO4,PH8.6Tris-HCl,PH9.6-10.6Gly-NaOH)反应体系中,5μg酶按照标准检测方法测定其活性。根据酶在不同pH下的相对活性绘制柱状图,确定酶的最适反应pH。以灭活的酶作为对照,以活性最高的值为100%,测定在各个反应pH下酶的相对活性。
结果如图2所示,以GlcNAc-6-P为底物时的最适pH范围在中性偏酸性范围内,而以GlcNAc-1-P为底物的最适PH范围在中性偏碱性范围内。AtAGM催化正逆反应的最适pH不同,可以根据此特性调节反应pH来控制目的异构体的积累,这对应用于生产中具有具有十分重大的意义。
(5)EDTA、SDS及金属离子等对AtAGM活性的影响
采用实施例4(1)中提到的通用体系,以30μM的GlcNAc-6-P作为底物,反应体系中设定各种金属离子浓度在10mM,按照标准方法检测酶活力。以灭活的酶作为对照,以活性最高的值为100%。结果如表3所示,Mg2+对AtAGM的酶活具有明显的提高作用,另外Ca2+与K+也有较大的提高作用,一些离子如Fe2+,Fe3+,Cu2+等对反应有明显的抑制作用。
表3:金属离子对AtAGM酶活的影响(GlcNAc-6-P为底物)
实施例5 一种检测己糖磷酸变位酶活性新方法的建立
经过多次探索,利用离子交换色谱与电化学检测器(HAPEC-PAD)可以成功的将多对磷酸己糖的1,6位磷酸异构体成功分离。
所用的离子交换色谱系统包括戴安Bio-LC梯度混匀泵,GM-3(4mm)梯度混匀器,离子交换色谱柱CarboPac PA-100column(4×250mm),AgCl参比电极与电化学检测器。用于检测的脉冲电位变化为:t=0s,E=0.10v;t=0.20s,E=0.10v;t=0.40s,E=0.10v;t=0.41s,E=-2.00v;t=0.42s,E=-2.00v;t=0.43s,E=0.60v;t=0.44s,E=-0.10v;t=0.50s,E=-0.10v.所用的流动相:A,100mM NaOH水溶液;B,800mM乙酸钠与100mM NaOH水溶液。流动相洗脱条件是0-5min,90%A+10%B;6-15min,10%-90%B;16-18min,10%A+90%B;19-20min,90%A+10%B.总流速是0.5ml/min,检测柱温是30℃,进样体积是20μL。
利用以上的检测方法,磷酸己糖的保留时间:GlcNAc-1-P是12.17min,GlcNAc-6-P是14.30min,GlcN-1-P是11.55min,GlcN-6-P是13.75min,Glc-1-P是12.05min,Glc-6-P是13.72min。磷酸己糖的检测限:GlcNAc-1-P是2.747pmol,GlcNAc-6-P是1.365pmol,GlcN-1-P是0.512pmol,GlcN-6-P是0.415pmol,Glc-1-P是1.486pmol,Glc-6-P是0.868pmol.仪器的响应值与磷酸己糖的浓度在2-400μM浓度范围内线性极好(R2>0.999),在1-15000μM内保持线性关系(R2>0.99)(附图2所示)。
基于HAPEC-PAD方法对磷酸己糖异构体的良好的分离效果,推测可以用于己糖磷酸变位酶家族酶活性的检测。AtAGM可以催化GlcNAc-6-P与GlcNAc-1-P;GlcN-6-P与GlcN-1-P;Glc-6-P与Glc-1-P几对异构体之间的转换。利用HAPEC-PAD方法可以很直观的观察到其底物的消耗与产物的生成情况。当以Glc-1-P为底物的时候,利用传统的偶联法与HAPEC-PAD同时检测其酶动力学参数,结果相差无几,说明HAPEC-PAD可以用于己糖磷酸变位酶酶活的检测(附图3及表4所示)。
表4:用偶联法与HAPEC-PAD检测AtAGM酶动力学,以Glc-1-P为底物。
另外,当用于检测GlmM(结核杆菌来源,催化GlcN-6-P与GlcN-1-P的转变)酶活的时候,HAPEC-PAD所检测的酶活要远高于传统偶联法,原因在于偶联反应的酶存在底物抑制的情况(表5所示)。所以HAPEC-PAD与传统偶联法相比较,更加的方便快捷,而且灵敏度高,正逆反应都可以检测,更为直观。
表5:用偶联法与HAPEC-PAD检测MtGlmM的酶活。
实施例6 一种生产UDP-GlcNAc与己糖磷酸异构体的方法
目前已经有研究使用无细胞催化的多步酶催化反应体系,偶联了葡萄糖激酶YpgR,AGM与GlmU三个酶,以GlcNAc作为底物生产UDP-GlcNAc(2010,浙江大学)。将三个酶分别用无细胞体系表达后再进行合成反应,40mM GlcNAc在20℃下反应24h后,生成4.1mM的UDP-GlcNAc。由于其使用酵母来源的AGM,在其表达体系中的可溶性表达较低,而且酶活力较低,成为整个酶催化体系的限速步骤,严重影响了UDP-GlcNAc的产量。
植物来源的AtAGM在16℃过夜诱导情况下,可溶性表达量达到80%。在30℃下反应30min,AtAGM对GlcNAc-6-P的酶活是130.08±1.67nmol/min.mg(实施案例4)。来自于枯草芽孢杆菌的YpgR的酶活是120nmol/min/mg,来自于大肠杆菌的GlmU的酶活是290nmol/min/mg。当YpgR,AtAGM,GlmU分别加入100mg、100mg和50mg混匀,40mM GlcNAc反应24h可产生17.28mM的UDP-GlcNAc。
表6:AGM对己糖磷酸异构体的转化率,30℃下反应20min。
对于己糖磷酸异构体的生产,以GlcNAc-1-P作为底物,30℃,反应20min就可产生相应的GlcNAc-6-P,其转化率可达90%以上(表5所示)。以GlcNAc-6-P作为底物,30℃,反应20min就可产生相应的GlcNAc-1-P,其转化率可达80%以上(表6所示)。具体反应条件见实施案例4。可以利用液相分离产物峰,即可得到相应的己糖磷酸异构体。
- 还没有人留言评论。精彩留言会获得点赞!