使用GH5木聚糖酶从淀粉生产乙醇的工艺的制作方法
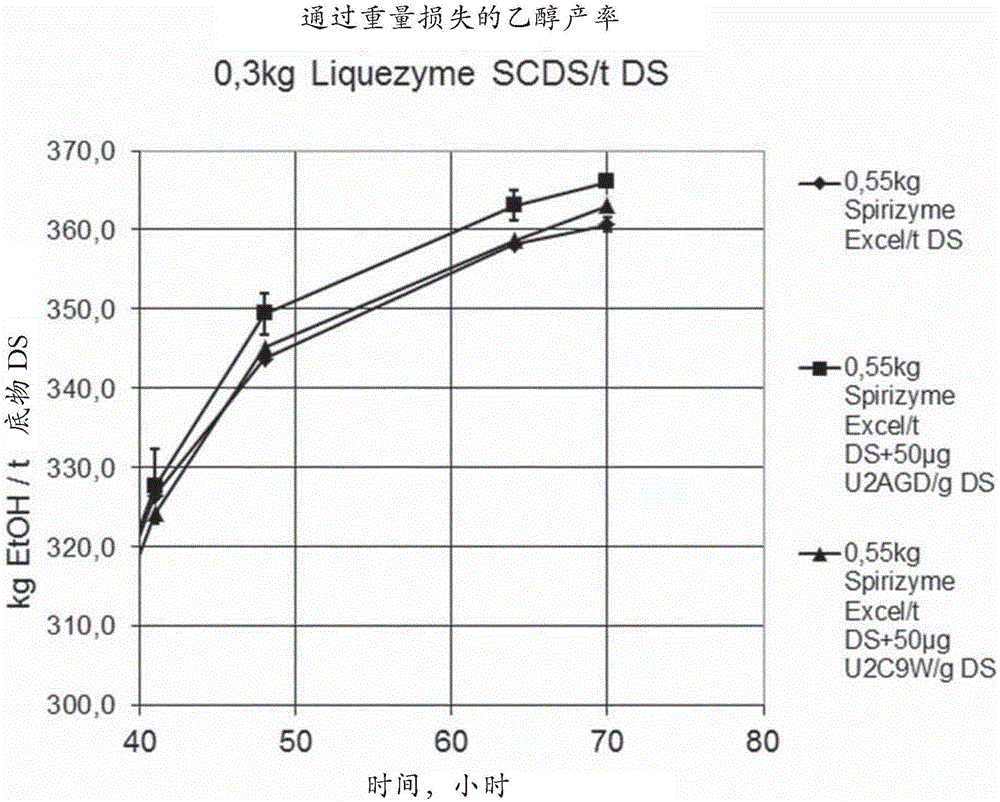
本申请包括计算机可读形式的序列表,将其通过引用结合在此。发明背景发明领域本发明涉及用于从含淀粉材料生产发酵产物的工艺。相关领域描述木聚糖是在所有陆生植物中发现的半纤维素(波普尔(Popper)和陶西(Tuohy),植物生理学(PlantPhysiol.),2010,153:373-383)。它们在次生细胞壁和木材细胞中尤其丰富。在具有II型细胞壁的草中,葡糖醛酸阿拉伯木聚糖是主要的半纤维素,并且在许多基于草的食物和饲料产品中作为可溶性或不可溶性膳食纤维存在。植物木聚糖具有β-1,4-连接的吡喃木糖主链,该主链可以在O2或O3位置以物种和组织特异性方式被阿拉伯糖、葡萄糖醛酸和乙酸取代。黍亚科与经济上重要的物种(例如玉米和高粱)的富含淀粉的种子在它们的细胞壁中具有特殊类型的高度取代的木聚糖。与小麦粉相比,其中在阿拉伯木聚糖主链中超过60%的木糖单元是未被取代的。在玉米核木聚糖中,对应的未被取代的主链木糖的百分比是20%-30%,并且在高粱中,该百分比是35%-40%(惠思曼(Huismann)等人,碳水化合物聚合物(CarbohydratePolymers),2000,42:269-279)。此外,在玉米和高粱中,这些木聚糖侧链可以比在其他木聚糖中常见的单一阿拉伯糖或葡萄糖醛酸取代更长。这种附加的侧链复杂性通常是由于结合至该侧链阿拉伯糖或葡萄糖醛酸的L-和D-半乳糖以及D-木糖造成的。玉米核木聚糖中约每十分之一的阿拉伯糖还被阿魏酸酯化,并且约每四分之一木糖进行乙酰化(阿格尔(Agger)等人,农业与食品化学杂志(J.Agric.FoodChem),2010,58:6141-6148)。所有组合的这些因素使得玉米和高粱中高度取代的木聚糖对抗由传统的木聚糖酶引起的降解。基于序列相似性,将负责木聚糖主链水解的已知的酶分类为酶家族(www.cazy.org)。主要具有内切-木聚糖酶活性的这些酶之前已描述于糖苷水解酶家族(GH)5、8、10、11和30中。家族内的酶共享一些特征,例如3D折叠,并且它们通常共同具有相同的反应机制。一些GH家族具有狭窄或单一底物特异性,然而其他家族具有宽泛的底物特异性。通常使用可商购的GH10和GH11木聚糖酶来分解阿拉伯木聚糖的木糖主链。然而,这种木聚糖酶对侧链位阻敏感,并且虽然它们有效地降解来自小麦的阿拉伯木聚糖,但是它们对在黍亚科物种(例如玉米或高粱)的种子中发现的木聚糖不是非常有效的。在全世界,玉米被用于例如动物饲料中和用于生产生物乙醇,并且因此有需要发现具有木聚糖酶活性的新的多肽,这些多肽能分解细胞壁中的高度支链化的木聚糖主链,以便释放更多陷在细胞壁中的淀粉。发明概述根据本发明,显示出属于GH5家族的若干种木聚糖酶在水解高度支链化的木聚糖主链中是有效的,并且因此这些GH5木聚糖酶可以有利地应用于淀粉乙醇工艺中。因此,本发明涉及用于从含淀粉材料生产发酵产物的工艺,该工艺包括以下步骤:a)使用α-淀粉酶,使含淀粉材料液化;b)使用葡糖淀粉酶使该液化材料糖化;c)使用发酵生物进行发酵,其中在GH5木聚糖酶的存在下进行糖化。附图简述图1显示在糖化期间通过添加GH5木聚糖酶对乙醇产率的影响。将液化醪液分成三个部分并且进行同时糖化发酵。发酵之后,每天测量两次重量损失并且在发酵结束时进行HPLC。定义含阿拉伯木聚糖的材料:术语“含阿拉伯木聚糖的材料”意指包含阿拉伯木聚糖的任何材料。阿拉伯木聚糖是在植物(包括木头和谷物)的初生和次生细胞壁中发现的半纤维素,该阿拉伯木聚糖由两种戊糖、阿拉伯糖和木糖的共聚物组成。该阿拉伯木聚糖链包含大量的1,4-连接的木糖单元。许多木糖单元被2-、3-或2,3-取代的阿拉伯糖残基取代。包含阿拉伯木聚糖的材料的实例是草料、粗粮、种子和谷物(例如来自玉米、燕麦、黑麦、大麦、小麦的整体或通过粉碎、研磨等制备的)、树木或硬木(例如杨树、柳树、桉树、棕榈、枫树、桦树)、竹子、草本和/或木本能源作物、农业粮食和饲料作物、动物饲料产品、木薯皮、可可豆荚、甘蔗、甜菜、槐豆粕、蔬菜或水果果渣、木材废料、树皮、刨花、锯末、木浆、制浆废液、废纸、纸板、建筑和拆除木材废料、工业或城市废水固体或污泥、肥料、来自酿造和/或发酵过程的副产物、湿酒糟、干酒糟、谷物渣、丢糟和甘蔗渣。如在此所定义的草料还包括粗粮。草料是新鲜的植物物料,例如来自草料植物(禾草)和其他草料植物(海草、发芽谷物和豆类植物)或其任何组合的干草和青贮饲料。草料植物的实例是苜蓿(紫花苜蓿)、百脉根、芸苔属植物(例如,羽衣甘蓝、油菜籽(卡诺拉(canola))、芜菁甘蓝(瑞典芜菁)、萝卜)、三叶草(例如,杂三叶、红三叶、地三叶、白三叶)、禾草(例如,百慕大草、雀麦、伪燕麦草、羊茅、石南草(heathgrass)、草地早熟禾、芒草、鸭茅(orchardgrass)、黑麦草、柳枝稷、梯牧草(Timothy-grass))、玉米(玉蜀黍)、大麻、粟、大麦、燕麦、黑麦、高粱、大豆和小麦和蔬菜(如甜菜)。适用于青贮的作物是普通的禾草、三叶草、紫花苜蓿、野豌豆、燕麦、黑麦和玉蜀黍。草料进一步包括来自谷物产物的作物残余物(例如玉米秸秆,来自小麦、大麦、燕麦、黑麦和其他谷物的秸秆),来自蔬菜像甜菜缨(beettop)的残余,来自油籽产物像来自大豆、油菜籽和其他豆类植物的茎和叶子的残余,以及来自用于动物或人类消耗的谷物精致过程或来自燃料生产或其他行业的级分。粗粮一般是具有高水平纤维的干植物物料,例如来自种子和谷物以及作物残余物(例如秸秆、干椰肉(copra)、稻草、谷壳、甜菜废料)的纤维、麸、苞叶。包含阿拉伯木聚糖的材料的优选来源是草料、粗粮、种子和谷物、甘蔗、甜菜和木浆。cDNA:术语“cDNA”意指可以通过从得自真核或原核细胞的成熟的、剪接的mRNA分子进行反转录而制备的DNA分子。cDNA缺乏可以存在于对应基因组DNA中的内含子序列。早先的初始RNA转录本是mRNA的前体,其在呈现为成熟的剪接的mRNA之前要经一系列的步骤进行加工,包括剪接。编码序列:术语“编码序列”意指直接指定多肽的氨基酸序列的多核苷酸。编码序列的边界一般由开放阅读框架决定,该开放阅读框架从起始密码子(如ATG、GTG或TTG)开始并且以终止密码子(如TAA、TAG或TGA)结束。编码序列可以是基因组DNA、cDNA、合成DNA或其组合。对照序列:术语“控制序列”是指表达编码本发明的成熟多肽的多核苷酸所必需的核酸序列。每个控制序列对于编码该多肽的多核苷酸来说可以是天然的(即,来自相同基因)或外源的(即,来自不同基因),或相对于彼此是天然的或外源的。这些调控序列包括但不限于前导序列、聚腺苷酸化序列、前肽序列、启动子、信号肽序列和转录终止子。至少,控制序列包括启动子,以及转录和翻译终止信号。出于引入有利于将这些控制序列与编码多肽的多核苷酸的编码区连接的特异性限制酶切位点的目的,这些控制序列可以提供有多个接头。表达:术语“表达”包括涉及多肽产生的任何步骤,包括但不限于,转录、转录后修饰、翻译、翻译后修饰、以及分泌。表达载体:术语“表达载体”意指直链或环状DNA分子,该分子包括编码多肽的多核苷酸并且可操作地连接至提供用于其表达的控制序列。高度支链化的木聚糖:术语“高度支链化的木聚糖”意指在阿拉伯木聚糖主链中的超过50%的木糖单元是被取代的。这优选地从如在惠思曼(Huismann)等人,碳水化合物聚合物(CarbohydratePolymers),2000,42:269-279中进行的连锁分析来计算。宿主细胞:术语“宿主细胞”意指易于用包括本发明的多核苷酸的核酸构建体或表达载体转化、转染、转导等的任何细胞类型。术语“宿主细胞”涵盖由于复制期间发生的突变而与亲本细胞不同的亲本细胞的任何后代。分离的:术语“分离的”意指处于非天然存在的形式或环境中的物质。分离的物质的非限制性实例包括(1)任何非天然存在的物质;(2)至少部分地从一个或多个或所有与它在自然界中相关的天然存在的成分中除去的任何物质,包括但不限于,任何酶、变体、核酸、蛋白质、肽或辅因子,也就是;(3)相对于在自然界中发现的物质经过人为改变的任何物质;或(4)相对于与它天然相关联的其它组分通过增加该物质的量(例如,在宿主细胞中的重组生产;编码该物质的基因的多个拷贝;以及使用比与编码该物质的基因天然相关联的启动子更强的启动子)而改变的任何物质。成熟多肽:术语“成熟多肽”意指在翻译和任何翻译后修饰如N-末端加工、C-末端截短、糖基化作用、磷酸化作用等之后处于其最终形式的多肽。在一方面,成熟多肽是SEQIDNO:2的氨基酸37至573。SEQIDNO:2的氨基酸1至27是信号肽。SEQIDNO:2的氨基酸28至36是his-标签。在另一方面,成熟多肽是SEQIDNO:4的氨基酸36至582。SEQIDNO:4的氨基酸1至27是信号肽。SEQIDNO:4的氨基酸28至35是his-标签。在另一方面,成熟多肽是SEQIDNO:6的氨基酸36至633。SEQIDNO:6的氨基酸1至27是信号肽。SEQIDNO:6的氨基酸28至35是his-标签。在另一方面,成熟多肽是SEQIDNO:7的氨基酸1至620。在另一方面,成熟多肽是SEQIDNO:8的氨基酸1至865。本领域已知,宿主细胞可以产生由同一多核苷酸表达的两种或更多种不同成熟多肽(即,具有不同C-末端和/或N-末端氨基酸)的混合物。本领域还已知,不同的宿主细胞不同地加工多肽,并且因此一个表达多核苷酸的宿主细胞当与另一个表达相同多核苷酸的宿主细胞相比时可以产生不同的成熟多肽(例如,具有不同的C-末端和/或N-末端氨基酸)。成熟多肽编码序列:术语“成熟多肽编码序列”是指编码具有木聚糖酶活性的成熟多肽的多核苷酸。在一方面,成熟多肽编码序列是SEQIDNO:1的核苷酸109至1719。SEQIDNO:1的核苷酸1至81编码信号肽。SEQIDNO:1的核苷酸82至108编码his-标签。在一方面,成熟多肽编码序列是SEQIDNO:3的核苷酸106至1746。SEQIDNO:3的核苷酸1至81编码信号肽。SEQIDNO:3的核苷酸82至105编码his-标签。在一方面,成熟多肽编码序列是SEQIDNO:5的核苷酸106至1899。SEQIDNO:5的核苷酸1至81编码信号肽。SEQIDNO:5的核苷酸82至105编码his-标签。寡糖组合物:术语“寡糖组合物”意指寡糖和多糖但不包括单单糖和二糖(DP1和DP2)。核酸构建体:术语“核酸构建体”意指单-链或双-链的核酸分子,该核酸分子是从天然存在的基因中分离的,或以本来不存在于自然界中的方式被修饰成含有核酸的区段,或是合成的,该核酸分子包括一个或多个控制序列。可操作地连接:术语“可操作地连接”意指如下配置,其中控制序列相对于多核苷酸的编码序列放置在适当位置,以使得控制序列指导编码序列的表达。溶解的木糖百分比:术语“溶解的木糖百分比”意指,与酶孵育之前存在于底物中的木糖的总量相比,用酶孵育后在上清液中所测量的木糖的量。来自于脱脂脱淀粉玉蜀黍(DFDSM)的溶解的木糖百分比可以如本文中实例3中所描述的进行计算。序列一致性:两个氨基酸序列之间或者两个核苷酸序列之间的关联度通过参数“序列一致性”来描述。出于本发明的目的,使用如在EMBOSS包(EMBOSS:欧洲分子生物学开放软件套件(TheEuropeanMolecularBiologyOpenSoftwareSuite),赖斯(Rice)等人,2000,遗传学趋势(TrendsGenet.)16:276-277)(优选3.0.0版或更新版本)的尼德尔(Needle)程序中所实施的尼德尔曼-翁施(Needleman-Wunsch)算法(尼德尔曼(Needleman)和翁施(Wunsch),1970,分子生物学杂志(J.Mol.Biol.)48:443-453)来确定两个氨基酸序列之间的序列一致性的程度。使用6.1.0版。所用的任选参数是空位开放罚分10、空位延伸罚分0.5,和EBLOSUM62(BLOSUM62的EMBOSS版)替代矩阵。尼德尔标注的“最长的一致性”的输出(使用-非简化选项获得)被用作百分比一致性,并且如下计算:(相同残基x100)/(比对的长度-在比对中的空位总数)出于本发明的目的,使用如在EMBOSS包(EMBOSS:欧洲分子生物学开放软件套件,赖斯(Rice)等人,2000,同上)(优选3.0.0版或更新版本)的尼德尔程序中所实施的尼德尔曼-翁施算法(尼德尔曼(Needleman)和翁施(Wunsch),1970,同上)来确定两个脱氧核糖核苷酸序列之间的序列一致性程度。使用6.1.0版。所用的任选参数是空位开放罚分10、空位延伸罚分0.5,和EDNAFULL(NCBINUC4.4的EMBOSS版)替代矩阵。尼德尔标注的“最长的一致性”的输出(使用-非简化选项获得)被用作百分比一致性,并且如下计算:(一致的脱氧核糖核苷酸x100)/(比对长度-比对中的空位总数)木聚糖酶:术语“木聚糖酶”意指1,4-β-D-木聚糖-木糖水解酶(E.C.3.2.1.8),其催化木聚糖中的1,4-β-D-木糖苷键的内切水解。木聚糖酶活性可以在37℃下在0.01%X-100和200mM磷酸钠(pH6)中用0.2%AZCL-阿拉伯糖基木聚糖作为底物来确定。一个单位的木聚糖酶活性定义为在37℃,pH6在200mM磷酸钠pH6中从作为底物的0.2%AZCL-阿拉伯木聚糖每分钟产生1.0微摩尔天青精。GH5木聚糖酶:在本发明中描述的这些木聚糖酶属于GH5,GH5是具有广泛的底物特异性的GH家族。通过对相关序列的亚家族进行定义已经澄清了GH5内序列之间的关系(阿斯博白(Aspeborg)等人,BMC进化生物学(BMCEvolutionaryBiology),2012,12:186)。将GH家族细分为亚家族显著改进了底物特异性的预测能力,不仅仅是针对GH5(伦巴第(Lombard)等人核酸研究(NucleicAcidsRes.),2014,42:D490-D495)。GH5、GH5_21和GH5_34的亚家族中的两个已经被描述为作用于阿拉伯木聚糖的木聚糖酶。有趣的是,这两个木聚糖酶亚家族不是紧密相关的并且是趋同进化的结果。在这项工作的过程中,已经发现亚家族GH5_35的成员,之前未知功能的亚家族具有木聚糖酶活性。发明详述本发明的诸位发明人已经发现来自糖苷水解酶家族5(本文中被称为GH5)的某些木聚糖酶出乎意料地擅长降解位阻阿拉伯木聚糖的木糖主链,从而释放增加量的木糖。具体地,属于亚家族GH5_21、GH5_34和GH5_35的GH5木聚糖酶都具有获得和降解高度支链化的木聚糖的能力(意指在阿拉伯木聚糖主链中超过50%的木糖单元是被取代的),这些木聚糖对抗由其他GH家族的木聚糖酶引起的木聚糖酶降解。增加降解,并从而增加木糖释放,增加陷在木糖结构中的淀粉的可及性。因此,本发明涉及用于从含淀粉材料生产发酵产物的工艺,该工艺包括以下步骤:a)使用α-淀粉酶,使含淀粉材料液化;b)使用葡糖淀粉酶使该液化材料糖化;c)使用发酵生物进行发酵,其中在GH5木聚糖酶的存在下进行糖化。GH5木聚糖酶在一个实施例中,在本发明的工艺中应用的GH5木聚糖酶选自下组,该组由以下各项组成:亚家族21、34或35。在一个特定的实施例中,该亚家族21GH5木聚糖酶是选自如SEQIDNO:6的氨基酸36至633所示的木聚糖酶,或是与SEQIDNO:6的氨基酸36至633具有至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%一致性的GH5木聚糖酶。在另一个特定的实施例中,该亚家族34GH5木聚糖酶是选自如SEQIDNO:7所示的木聚糖酶,或是与SEQIDNO:7具有至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%一致性的GH5木聚糖酶。在另一个特定的实施例中,该亚家族34GH5木聚糖酶是选自如SEQIDNO:8所示的木聚糖酶,或是与SEQIDNO:8具有至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%一致性的GH5木聚糖酶。在另一个特定的实施例中,该亚家族35GH5木聚糖酶是选自如SEQIDNO:2的氨基酸37至573所示的木聚糖酶,或是与SEQIDNO:2的氨基酸37至573具有至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%一致性的GH5木聚糖酶。在另一个特定的实施例中,该亚家族35GH5木聚糖酶是选自如SEQIDNO:4的氨基酸36至582所示的木聚糖酶,或是与SEQIDNO:4的氨基酸36至582具有至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%一致性的GH5木聚糖酶。从含淀粉材料生产发酵产物由含经糊化淀粉的材料生产发酵产物的工艺在这一方面,本发明涉及用于从含淀粉材料生产发酵产物,尤其是乙醇的工艺,该工艺包括液化步骤及依序地或同时地进行的糖化和发酵步骤。本发明涉及用于从含淀粉材料生产发酵产物的工艺,这些工艺包括以下步骤:i)使用α-淀粉酶,使含淀粉材料液化;ii)使用产碳水化合物源的酶来使该液化材料糖化;iii)使用发酵生物进行发酵,其中在GH5木聚糖酶的存在下进行糖化。发酵产物,如尤其是乙醇,可以可任选地在发酵之后,例如通过蒸馏来回收。在以下“含淀粉材料”部分中列出了多种适合的含淀粉的起始材料。在一个实施例中,该含淀粉材料是玉米或小麦。在以下“酶”部分中列出了多种所考虑的酶。在α-淀粉酶,优选细菌α-淀粉酶,特别是芽孢杆菌α-淀粉酶,如嗜热脂肪芽孢杆菌α-淀粉酶的存在下进行液化。该发酵生物优选地是酵母,优选地是酵母属菌株,尤其是酿酒酵母菌株。在以上“发酵生物”部分中列出了多种适合的发酵生物。在一个优选的实施例中,连续或同时进行步骤ii)和iii)(即,作为SSF工艺)。该水性浆料可以包含含淀粉材料的从10wt.-%-55wt.-%干固体,优选25wt.-%-45wt.-%干固体,更优选30wt.-%-40wt.-%干固体。将该浆料加热至高于初始糊化温度。可以将α-淀粉酶,优选地细菌性α-淀粉酶添加到该浆料中。在一个实施例中,在液化步骤i)中经受α-淀粉酶之前,还将该浆料喷射蒸煮以进一步使该浆料糊化。在步骤(i)期间的温度高于初始糊化温度,例如在80℃-90℃之间,例如85℃左右。在一个实施例中,液化以三步热浆料工艺进行。该浆料被加热到介于60℃到95℃,优选地80℃到90℃之间,并且添加α-淀粉酶以开始液化(稀释)。然后,该浆料在介于95℃到140℃,优选地105℃到125℃之间的温度下进行喷射烹饪1-15分钟,优选地3-10分钟,尤其是约5分钟。该浆料被冷却到60℃-95℃,优选地80℃-90℃,并且再添加α-淀粉酶以结束水解(二次液化)。通常在pH4.5-pH6.5、特别是在pH5-pH6之间进行液化过程。研磨并且液化的淀粉称为“醪液”(mash)。可使用本领域熟知的条件来进行步骤ii)中的糖化。例如,全部糖化工艺可以持续长达从约24小时至约72小时。在一个实施例中,在30℃-65℃之间的温度下,典型地约60℃,在40-90分钟完成了预糖化步骤,随后在同时糖化发酵步骤(SSF)中的发酵过程中进行完全的糖化。糖化典型地在从30℃-70℃、如55℃-65℃,典型地60℃左右的温度下,并且在4与5之间的pH下,一般在约pH4.5下进行。发酵产物生产,特别是乙醇生产中最广泛使用的工艺为同时糖化发酵(SSF)工艺,其中没有糖化的保持阶段。当该发酵生物是酵母,如酿酒酵母的菌株,并且该所希望的发酵产物是乙醇时,SSF通常可以在25℃和40℃之间,如28℃和36℃之间,如在30℃和34℃之间的温度,如约32℃下进行。在一个实施例中,发酵进行6至120个小时,尤其24至96个小时。其它发酵产物可以在本领域技术人员熟知的、适合于所讨论的发酵生物的条件和温度发酵。根据本发明,可以在发酵过程中向上或向下调节温度。在一个实施例中,在发酵过程中添加蛋白酶。蛋白酶的实例可在以下的“蛋白酶”部分中找到。在一个优选的实施例中,以同时糖化发酵工艺进行步骤(ii)和步骤(iii)。在这种优选的实施例中,该过程典型地在介于25℃与40℃之间,如介于28℃与36℃之间,如介于30℃与34℃之间,如约32℃的温度下进行。根据本发明,可以在发酵期间向上或向下调节温度。在一个实施例中,进行同时糖化发酵,这样使得糖水平,如葡萄糖水平保持在低水平,如低于6wt.-%,优选地低于约3wt.-%,优选地低于约2wt.-%,更优选的低于约1wt.-%,甚至更优选的低于约0.5wt.-%或甚至更优选的0.25%wt.-%,如低于约0.1wt.-%。通过简单地采用调整量的酶和发酵生物就可以实现这样的低水平的糖。本领域的普通技术人员可以容易地确定使用多少量的酶和发酵生物。还可以选择酶和发酵生物的所采用的量以在发酵液中维持低麦芽糖浓度。例如,麦芽糖水平可保持在低于约0.5wt.-%或低于约0.2wt.-%。本发明的工艺可以在范围介于3与7之间,优选地从pH3.5到6,或更优选地从pH4到5的pH下进行。在一个实施例中,在发酵过程中添加蛋白酶。蛋白酶的实例可在以下的“蛋白酶”部分中找到。含淀粉材料根据本发明,糖类可以源自含淀粉材料。根据本发明可以使用任何适合的含淀粉的起始材料,包括颗粒状淀粉。通常基于所希望的发酵产物来选择起始材料。适合用于本发明的工艺中的含淀粉起始材料的实例包括整谷粒、玉米、小麦、大麦、黑麦、小黑麦、买罗高梁(milo)、西米、木薯、木薯粉、高粱、稻米、豆类、菜豆和甘薯,或其混合物,或者谷物、或含糖原料,如糖蜜、果实材料、糖甘蔗或糖甜菜、马铃薯。考虑了糯与非糯类型的玉米与大麦两者。在一个具体的实施例中,该含淀粉材料包括玉蜀黍、玉米、小麦、黑麦、大麦、小黑麦、高粱、柳枝稷、粟、珍珠粟、谷子或处于加工过的形式(如研磨的玉米、研磨的小麦、研磨的黑麦、研磨的大麦、研磨的小黑麦、研磨的玉蜀黍、脱脂玉蜀黍、脱脂脱淀粉的玉蜀黍、研磨的高粱、研磨的柳枝稷、研磨的粟、研磨的谷子、研磨的珍珠粟)。具体地,该基于淀粉的材料包括玉米、高粱、小麦、黑麦、大麦、或小黑麦。术语“颗粒状淀粉”意指生的未烹饪的淀粉,即,在谷物、块茎或谷粒中所发现的呈其天然形式的淀粉。淀粉是在植物细胞内作为不溶于水的微小颗粒形式而形成。当放入冷水中时,这些淀粉颗粒可以吸收少量的液体并膨胀。在最多50℃至75℃的温度下,该膨胀可以是可逆的。然而,在更高温度下,开始不可逆膨胀,称为“胶凝化”。在一个实施例中,有待加工的颗粒状淀粉可以是高度精制的淀粉,优选地至少90%、至少95%、至少97%或至少99.5%纯,或它可以为含更粗糙的淀粉的材料,该材料包括经过研磨的整谷粒,包括非淀粉部分在内,如胚芽残余物和纤维。该原材料(如整谷粒)经过研磨以便展开结构并且允许进一步加工。根据本发明,以下两种研磨工艺是优选的:湿式和干式研磨。在干式研磨中,整个谷粒被研磨并且使用。湿式研磨使胚芽与粗粉(淀粉颗粒和蛋白质)良好分离,并且时常应用于使用淀粉水解产物生产糖浆的场所。干式和湿式研磨两者是本领域中众所周知的淀粉加工法并且同等地被涵盖用于本发明的工艺。优选地可以通过干式或湿式研磨来减小该含淀粉材料的粒度,以便暴露出更大的表面积。在一个实施例中,该粒度在0.05mm至3.0mm、优选0.1mm-0.5mm之间,或使得至少30%、优选至少50%、更优选至少70%、甚至更优选至少90%的含淀粉材料适合通过具有0.05mm至3.0mm筛网、优选0.1mm-0.5mm筛网的筛子。发酵生物术语“发酵生物”是指适于生产所希望的发酵产物的任何生物,包括细菌和真菌生物,包括酵母和丝状真菌。尤其是合适的根据本发明所述的发酵生物能够将糖,如葡萄糖、果糖、麦芽糖、木糖、甘露糖和/或阿拉伯糖,直接地或间接地发酵(即转变)成期望的发酵产物。发酵生物的实例包括真菌微生物,例如酵母。优选的酵母包括酵母属的菌株,特别是酿酒酵母或葡萄汁酵母;毕赤酵母属的菌株,特别是树干毕赤酵母或巴斯德毕赤酵母;假丝酵母属的菌株,特别是产朊假丝酵母、阿拉伯糖发酵假丝酵母(Candidaarabinofermentans)、迪丹斯假丝酵母(Candidadiddensii)、萨纳瑞西斯假丝酵母(Candidasonorensis)、休哈塔假丝酵母、假热带假丝酵母(Candidatropicalis)、或博伊丁假丝酵母。其他预期的酵母包括汉逊酵母属的菌株,特别是多形汉逊酵母或异常汉森酵母;克鲁弗酵母属的菌株,特别是马克斯克鲁弗酵母(Kluyveromycesmarxianus)或脆壁克鲁弗酵母;以及裂殖酵母属的菌株,特别是粟酒裂殖酵母。在一个实施例中,该发酵生物是C6糖(己糖)发酵生物,如例如酿酒酵母的菌株。尤其与木质纤维素衍生的材料的发酵相关的,还考虑了C5糖(戊糖)发酵生物。大多数C5糖发酵生物也发酵C6糖。C5糖发酵生物的实例包括毕赤酵母属的菌株,例如物种树干毕赤酵母的菌株。C5糖发酵细菌也是已知的。还有一些酿酒酵母菌株发酵C5(和C6)糖。能够发酵C5糖的酵母属的被遗传修饰过的菌株的实例包括在,例如,霍(Ho)等人,1998,应用与环境微生物学(AppliedandEnvironmentalMicrobiology),第1852-1859页和卡哈马(Karhumaa)等人,2006,微生物细胞工厂(MicrobialCellFactories)5:18中所涉及的那些。在一个实施例中,将该发酵生物添加至发酵中,这样使得每mL的发酵培养基的活发酵生物(例如酵母)计数在从105至1012个的范围内,优选从107至1010个,尤其是约5x107。可商购的酵母包括例如REDSTARTM和ETHANOLREDTM酵母(可获得自富酶泰斯/乐斯富(Fermentis/Lesaffre),美国),FALI(可获得自弗莱施曼酵母(Fleischmann’sYeast),美国),SUPERSTART和THERMOSACCTM新鲜酵母(可获得自乙醇技术(EthanolTechnology),威斯康星州(WI),美国),BIOFERMAFT和XR(可获得自NABC-北美生物产品集团(NABC-NorthAmericanBioproductsCorporation),佐治亚州(GA),美国),GERTSTRAND(可获得自格特·斯特兰德公司(GertStrandAB),瑞典)以及FERMIOL(可获得自帝斯曼食品配料部(DSMSpecialties))。能够从可发酵糖(包括葡萄糖、果糖、麦芽糖、木糖、甘露糖和/或阿拉伯糖)生产所希望的发酵产物的发酵生物优选在精确条件下以具体生长速率生长。当将该发酵生物引入进/添加至发酵培养基中时,接种的发酵生物经过多个阶段。不出现原始生长。此时期称为“停滞期”并且可以被认为是适应时期。在称为“对数期”的接下来的阶段过程中,生长速率逐渐增加。在最大生长期间后,速率停止并且发酵生物进入“静止期”。在另外的时间段后,发酵生物进入“死亡期”,其中活细胞的数目下降。发酵产物术语“发酵产物”意思指在包括使用发酵生物进行的发酵步骤在内的工艺生产的产物。根据本发明预期的发酵产物包括醇类(例如,乙醇、甲醇、丁醇);有机酸(例如,柠檬酸、乙酸、衣康酸、乳酸、葡糖酸);酮类(例如,丙酮);氨基酸(例如,谷氨酸);气体(例如,H2和CO2);抗生素(例如,青霉素和四环素);酶类;维生素(例如,核黄素、B12、β-胡萝卜素);和激素。在优选的实施例中,该发酵产物是乙醇,例如燃料乙醇;饮用乙醇,即,可饮用的酒精;或工业乙醇,或可消费醇工业(例如,啤酒和葡萄酒)、奶制品工业(例如,发酵的奶制品)、皮革工业及烟草工业所使用的产品。优选的啤酒类型包括爱尔啤酒(ale)、烈性黑啤酒(stout)、波特啤酒(porter)、拉格啤酒(lager)、苦啤酒(bitter)、麦芽酒(maltliquor)、低麦芽啤酒(happoushu)、高醇啤酒、低醇啤酒、低热量啤酒、或清淡啤酒。发酵源自用于本发明的工艺中的发酵的植物材料的这些糖可能源自含淀粉材料和/或含木质纤维素材料。基于例如植物材料的种类、可发酵糖、发酵生物和/或所希望的发酵产物确定发酵条件。本领域的普通技术人员可以容易地确定适合的发酵条件。根据本发明,发酵可以在常规使用的条件下进行。优选的发酵过程是厌氧过程。源自含淀粉材料的糖的发酵如上文所提到的,不同种类的发酵生物可以用于发酵源自含淀粉材料的糖。通常使用酵母(例如酿酒酵母)作为发酵生物进行发酵。然而,细菌和丝状真菌也可以用作发酵生物。一些细菌比,例如酿酒酵母具有更高的最适发酵温度。因此,在这些情况下,发酵可以在高达75℃,例如在40℃-70℃之间,如在50℃-60℃之间的温度下进行。然而,还已知的是具有低至室温左右(20℃左右)的显著较低的最适温度的细菌。适合的发酵生物的实例可以发现于上面的“发酵生物”部分中。对于使用酵母生产乙醇,在一个实施例中,该发酵可以持续24至96个小时,特别是持续36至72个小时。在一个实施例中,在20℃至40℃,优选28℃至36℃之间,特别是32℃左右的温度下进行发酵。在一个实施例中,该pH是从pH3至pH6,优选地是约pH4至pH5。尤其考虑的是同时的水解/糖化和发酵(称为“SSF”),其中针对水解/糖化不存在独立的保温阶段,意指可以一起添加该一种或多种水解酶、该一种或多种发酵生物、和钙调素蛋白。然而,应该理解的是,也可以单独地添加钙调素蛋白。当发酵是与糖化同时进行(即,SSF)时,当该发酵生物是酿酒酵母的菌株并且所希望的发酵产物是乙醇时,该温度优选是在20℃至40℃之间,优选是28℃至36℃,特别是约32℃。其他发酵产物可以在本领域的普通技术人员已知的、适于所讨论的发酵生物的温度下发酵。本发明的工艺可以作为分批过程或作为连续过程进行。本发明的发酵工艺可以在超滤系统中进行,其中将渗余物在固体、水和发酵生物的存在下保持在再循环之下,并且其中渗透物是包含所希望的发酵产物的液体。同样考虑的是否在具有超滤膜的连续膜反应器中执行该工艺,并且其中将渗余物在固体、水、一种或多种发酵生物的存在下保持在再循环之下,并且其中渗透物是包含发酵产物的液体。发酵后,可以将发酵生物与发酵的浆料分开并再循环。典型地在3与7之间的范围内的pH,优选从pH3.5至6,如pH5左右下进行发酵。发酵典型地持续24-96小时。根据本发明所述的方法和工艺根据本发明所述的主要的方面涉及,将含淀粉材料水解成糖并随后将这些糖发酵成醇(特别是乙醇)的工艺,其中在水解期间存在GH5木聚糖酶。上面已描述不同的工艺和工艺条件。本发明的具体方面涉及用于从含淀粉材料生产发酵产物的工艺,该工艺包括以下步骤:a)使用α-淀粉酶,使含淀粉材料液化;b)使用葡糖淀粉酶使该液化材料糖化;c)使用发酵生物进行发酵,其中在GH5木聚糖酶的存在下进行糖化。在一个具体的实施例中,该含淀粉材料包括玉蜀黍、玉米、小麦、黑麦、大麦、小黑麦、高粱、柳枝稷、粟、珍珠粟、谷子或处于加工过的形式(如研磨的玉米、研磨的小麦、研磨的黑麦、研磨的大麦、研磨的小黑麦、研磨的玉蜀黍、脱脂玉蜀黍、脱脂脱淀粉的玉蜀黍、研磨的高粱、研磨的柳枝稷、研磨的粟、研磨的谷子、研磨的珍珠粟)。具体地,该基于淀粉的材料包括玉米、高粱、小麦、黑麦、大麦、或小黑麦。该GH5木聚糖酶对高度支链化的木聚糖是特别有效的,并且因此在一个实施例中该含淀粉材料包括高度支链化的木聚糖。该GH5木聚糖酶优选地选自下组,该组由亚家族21、34、或35组成。在一个特定的实施例中,该亚家族21GH5木聚糖酶是选自如SEQIDNO:6的氨基酸36至633所示的木聚糖酶,或是与SEQIDNO:6的氨基酸36至633具有至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%一致性的GH5木聚糖酶。在另一个特定的实施例中,该亚家族34GH5木聚糖酶是选自如SEQIDNO:7所示的木聚糖酶,或是与SEQIDNO:7具有至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%一致性的GH5木聚糖酶。在另一个特定的实施例中,该亚家族34GH5木聚糖酶是选自如SEQIDNO:8所示的木聚糖酶,或是与SEQIDNO:8具有至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%一致性的GH5木聚糖酶。在另一个特定的实施例中,该亚家族35GH5木聚糖酶是选自如SEQIDNO:2的氨基酸37至573所示的木聚糖酶,或是与SEQIDNO:2的氨基酸37至573具有至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%一致性的GH5木聚糖酶。在另一个特定的实施例中,该亚家族35GH5木聚糖酶是选自如SEQIDNO:4的氨基酸36至582所示的木聚糖酶,或是与SEQIDNO:4的氨基酸36至582具有至少75%、至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%一致性的GH5木聚糖酶。在一个实施例中,该发酵产物是醇,特别是乙醇。糖化和发酵可以分开或同时进行。该发酵生物优选地是酵母菌属物种,更特别是酿酒酵母。适合的可商购的酵母菌株已经描述于本文中的“发酵生物”部分中。本发明进一步由下述编号的实施例描述:实施例1.用于从含淀粉材料生产发酵产物的工艺,该工艺包括以下步骤:a)使用α-淀粉酶,使含淀粉材料液化;b)使用葡糖淀粉酶使该液化材料糖化;c)使用发酵生物进行发酵,其中在GH5木聚糖酶的存在下进行糖化。实施例2.根据实施例1所述的工艺,其中该含淀粉材料包括玉蜀黍、玉米、小麦、黑麦、大麦、小黑麦、高粱、柳枝稷、粟、珍珠粟、谷子或处于加工过的形式(如研磨的玉米、研磨的小麦、研磨的黑麦、研磨的大麦、研磨的小黑麦、研磨的玉蜀黍、脱脂玉蜀黍、脱脂脱淀粉的玉蜀黍、研磨的高粱、研磨的柳枝稷、研磨的粟、研磨的谷子、研磨的珍珠粟)。实施例3.根据前述实施例中任一项所述的工艺,其中该含淀粉材料包括高度支链化的木聚糖。实施例4.根据前述实施例中任一项所述的工艺,其中该GH5木聚糖酶选自亚家族21、34、或35。实施例5.根据实施例4所述的工艺,其中该亚家族21GH5木聚糖酶选自如SEQIDNO:6的氨基酸36至633所示的木聚糖酶,或与SEQIDNO:6的氨基酸36至633具有至少75%一致性的GH5木聚糖酶。实施例6.根据实施例4所述的工艺,其中该亚家族34GH5木聚糖酶选自如SEQIDNO:7、SEQIDNO:8所示的木聚糖酶,或与SEQIDNO:7或SEQIDNO:8具有至少75%一致性的木聚糖酶。实施例7.根据实施例4所述的工艺,其中该亚家族35GH5木聚糖酶选自如SEQIDNO:2的氨基酸37至573、SEQIDNO:4的氨基酸36至582所示的木聚糖酶,或与SEQIDNO:2的氨基酸37至573、或SEQIDNO:4的氨基酸36至582具有至少75%一致性的木聚糖酶。实施例8.根据前述实施例中任一项所述的工艺,其中该植物材料包括玉米、高粱、小麦、黑麦、大麦、或小黑麦。实施例9.根据前述实施例中任一项所述的工艺,其中该发酵产物是醇,特别是乙醇。实施例10.根据前述实施例中任一项所述的工艺,其中该发酵生物是酵母,特别是酵母菌属物种,更特别是酿酒酵母。实施例11.根据前述实施例中任一项所述的工艺,其中步骤b)和c)同时进行。实施例12.用于从含淀粉材料产生乙醇的GH5木聚糖酶的用途。实施例13.根据实施例12所述的用途,其中该GH5木聚糖酶选自亚家族21、34、或35。实施例14.根据实施例12-13所述的用途,其中含淀粉材料包括玉米、高粱、小麦、黑麦、大麦、或小黑麦。通过以下实例进一步对本发明进行描述,但不应将其理解为对本发明范围的限制。实例培养基和溶液脱淀粉玉蜀黍(DSM)的制备在53℃下,将107kg的经研磨的玉蜀黍(<10mm)在槽中与253kg的自来水混合以制作浆料。该浆料的温度是47℃并且是pH5.9。用1L的1NNaOH将该pH调至6.15并然后将该槽加热至95℃。在52℃下添加1.119kg的α-淀粉酶(诺维信公司,巴格斯瓦德(Bagsvaerd),丹麦)并在95℃下孵育80分钟。在该孵育结束时测量的pH是6.17。将冷自来水添加到该浆料中并对该浆料进行离心,用Westfalia倾析器CA-225-110(4950±10rpm,流速约600l/h)将该浆料倾析3次,给出64.5kg的污泥。然后收集该污泥,进行冷冻并冻干以给出17.1kg的脱淀粉的玉蜀黍(DSM)。脱脂脱淀粉玉蜀黍(DFDSM)的制备将500mL丙酮添加到100克的脱淀粉的玉蜀黍中,如上所描述的进行制备。将该浆料搅拌5分钟并允许其沉降。倾析该丙酮并重复该程序2次。将该残余物空气干燥过夜以给出在室温下储存的脱脂脱淀粉玉蜀黍(DFDSM)。脱淀粉高粱的制备将整的高粱种子进行研磨并过筛,并将低于0.5mm的部分用于进一步加工。将过筛的部分以20%干物质悬浮于25mMNaOAc(pH5.5)中并进行脱淀粉。该脱淀粉涉及在85℃下用500ppmTermamylSCα-淀粉酶(诺维信公司,巴格斯瓦德(Bagsvaerd),丹麦)进行第一步是,持续20min,接着在65℃下用250ppmAttenuzymeFlex(诺维信公司,巴格斯瓦德,丹麦)过夜孵育。对该浆料进行离心并倾析该液体。在这之后,通过添加MilliQ水和200ppmTermamylSC和200ppmAttenuzymeFlex进行另一个脱淀粉,并在65℃下孵育过夜。通过经过沃特曼F玻璃纤维过滤器的真空过滤,将该高粱纤维从该液体中分离。然后用过量的水洗涤该滤饼若干次以去除可溶性糖。最后将脱淀粉的高粱纤维在65℃的烤箱中干燥,并将该干燥的纤维在咖啡研磨机中进行快速研磨,这样使得粒度一般少于1mm。玉米纤维胶的制备玉米脱淀粉在53℃下,将107kg预研磨的玉米(<1.0mm)与253kg水混合。将该混合物加热至95℃并用1MNaOH将该pH调至6.2。添加1.12kg,Termamyl120L(可从诺维信公司获得的商业α淀粉酶)并且将该反应在约90℃保持3hr。3hr后,添加冷水至总反应重量为600kg。用Westfalia倾析器,CA-225-110,在4950rpm下,600L/小时流速下完成液固分离。使固体纤维部分再浆化并且用水分离两次至总重量为600kg,随后如以上所描述的进行分离。将最后的纤维部分分为较小部分并且冷冻干燥3-4天。Termamyl120L是地衣芽孢杆菌α淀粉酶,然而,也可以使用其他显示出良好的温度稳定性的酸性α淀粉酶。实例包括嗜热脂肪芽孢杆菌α-淀粉酶,该淀粉酶与WO99/19467中披露的SEQIDNO:3阐述的野生型BSGα-淀粉酶氨基酸序列相比具有与δ(181-182)相对应的双缺失并且进一步包含N193F取代(也被表示为I181*+G182*+N193F)。玉米纤维胶提取将20g脱淀粉玉米与0.8g的NaOH和0.8g的Ca(OH)2一起添加至200mL煮沸的MilliQTM水中。将该混合物在96℃下保持1小时,并且然后在6000g下离心20min。该上清液看上去是乳状的并多脂的,并且因此用己烷通过摇动提取脂肪(1份己烷比4份玉米纤维胶)。在静置时间后,通过移液管去除该己烷。第二次提取(CFG2)是通过将球粒溶解于200mL1MNaOH中进行的。将该混合物在96℃下保持1小时,并且然后在6000g下离心20min,并且用4MHCl将上清液调节至pH6。使用旋转蒸发仪(rotavap)浓缩两个CFG部分,并且以90%的终浓度在EtOH中沉淀。来自0.1MNaOH的球粒从此后称为CFG1。由于1MNaOH提取物(CFG2)中的高pH,使用3kDa透析膜在水龙头滴水的情况下将该部分在具有去离子水的2L计量用筒上透析过夜。在应用试验之前,CFG1和CFG2二者在25000g下离心25min并且通过0.44μm针筒式滤器进行过滤。用MettlerToledoHR73确定在CFG2部分中的干物质。测定木糖测定从2.5mg木糖/mL(通过将0.125g木糖溶解于50mL去离子水中进行制备)的储液进行制备从0至125μg木糖/mL的木糖标准曲线。测定原理。通过木糖变旋酶(XMR)使用来自麦格酶国际爱尔兰公司(MegazymeInternationalIreland)的D-木糖测定试剂盒催化D-木糖的α-和β-异头形式的相互转化。在pH7.5,在β-木糖脱氢酶(β-XDH)存在下,通过NAD+将β-D-木糖氧化成D-木糖酸。在该反应中形成的NADH的量与D-木糖的量是化学计量的,并通过在340nm处的吸光度增加进行测量。实例1:GH5木聚糖酶的克隆供体菌株来自解纤维素醋弧菌CD2的木聚糖酶在其公开基因组序列的部分中进行鉴定,该序列如最初在登录号UniProt:E1KC96下所公开的(卢卡斯(Lucas)S.、科普兰(Copeland)A.、拉皮德斯(Lapidus)A.、程(Cheng)J.-F.、布鲁斯(Bruce)D.、古德温(Goodwin)L.、皮特拉克(Pitluck)S.、兰德(Land)M.L.、豪瑟(Hauser)L.、常(Chang)Y.-J.、杰弗里斯(Jeffries)C.、莫塔其(Mouttaki)H.、何(He)Z.、周(Zhou)J.、黑墨(Hemme)C.L.、沃一科(Woyke)T.J.;“解纤维素醋弧菌CD2的基因组草图(ThedraftgenomeofAcetivibriocellulolyticusCD2)”;将(AUG-2010)提交至EMBL/GenBank/DDBJ数据库)。该木聚糖酶还被公开在黑墨(Hemme)CL、莫塔其(Mouttaki)H、李(Lee)YJ、张(Zhang)G、古德温(Goodwin)L、卢卡斯(Lucas)S、科普兰(Copeland)A、拉皮德斯(Lapidus)A、格莱纳德尔里奥(GlavinadelRio)T、泰斯(Tice)H等人:与生物质转化和生物燃料生产相关的多个梭菌基因组的测序(Sequencingofmultipleclostridialgenomesrelatedtobiomassconversionandbiofuelproduction)细菌学杂志(JBacteriol)2010,192(24):6494-6496中。Uniprot:E1KC96是SEQIDNO:7和SEQIDNO:8都是GH5_34木聚糖酶。通过鸟枪基因组测序对来自菌株类芽孢杆菌属物种18054(SEQIDNO:3和4)和伊利诺斯州甜豆杆菌(Paenibacillusillinoisensis)(SEQIDNO:1和2)的木聚糖酶进行鉴定。两种木聚糖酶都属于家族GH5_35。在1991年,从新西兰的热样品中分离出这些菌株。伊利诺斯州甜豆杆菌作为DSMZ被保藏在登录号DSM16232下。通过对宏基因组提取物进行深度测序获得属于GH5_21家族的大象粪便宏基因组木聚糖酶(SEQIDNO:5和6)。使用在德国汉堡的动物园中生活的六岁的雌性亚洲大象(名为“康提(Kandy)”)的粪便。如在生产商的方案中所述,用来自凯杰公司(Qiagen)(希尔登,德国)的QIAampDNA粪便试剂盒进行DNA分离。基因测序,随后的读取和基因发现的装配(即基因功能的注释)对于本领域技术人员是已知的并且该服务是可商购的。克隆实例基于在部分/实例“供体菌株”中提到的核苷酸序列,每个木聚糖酶合成一个密码子优化的合成基因并且是可商购的。对Uniprot:E1KC96(A31-P650)的截短的三模块化版本进行克隆并且从这里开始被称为SEQIDNO:7。如在WO12/025577中所述,使用ClaI和MluI限制酶切位点将该优化程序和购买的合成基因的克隆亚克隆进芽孢杆菌属表达载体中。用克劳氏芽孢杆菌分泌信号(BcSP;具有如下氨基酸序列:MKKPLGKIVASTALLISVAFSSSIASA,该序列源于克劳氏芽孢杆菌的蛋白酶AprH)表达这些木聚糖酶。BcSP分别替代了所有基因中的所有天然分泌信号。将亲和标签序列引入BcSP序列的下游以便于该纯化过程(His标签;分别地具有如下氨基酸序列:针对于来自大象粪便宏基因组,类芽孢杆菌18054和解乙醇芽孢杆菌(Acetivibriocellulolyticus)的木聚糖酶为HHHHHHPR以及针对伊利诺斯州甜豆杆菌木聚糖酶为HQHQHQHPR)。因此,被表达的基因包括BcSP序列,其后是His标签序列,随后是成熟的野生型木聚糖酶序列。这些最终表达质粒(BcSP-His标签-木聚糖酶)被单独地转化到枯草芽孢杆菌表达宿主中。在转化时,将木聚糖酶BcSP-融合基因通过同源重组整合至枯草芽孢杆菌宿主细胞基因组中。在三联启动子系统的控制下表达该基因构建体(如描述于WO99/43835中)。将编码氯霉素乙酰转移酶的基因用作标记物(如描述于狄代丽柯森(Diderichsen)等人,1993,质粒(Plasmid)30:312-315中)。在每ml补充有6毫克氯霉素的LB培养基琼脂上选择转化体。选择一个包含对应的木聚糖酶表达构建体的重组枯草芽孢杆菌克隆,并且在旋转摇床上在500mL带挡板的锥形瓶(Erlenmeyerflask)中进行培养,每个锥形瓶包含100ml基于酵母提取物的培养基。在30℃至37℃下的3-5天培养时间后,通过离心收获包含酶的上清液并通过His-标签纯化对这些酶进行纯化。实例2:GH5木聚糖酶的纯化在5mLHisTrapExcel柱(通用电气医疗集团生命科学部(GEHealthcareLifeSciences))上,使用Ni2+作为金属离子,通过固定化金属层析(IMAC)纯化所有His-标记的酶。该纯化发生在pH8下,并用咪唑对结合蛋白进行洗脱。通过SDS-PAGE对经纯化的酶的纯度进行检查并在缓冲剂交换后通过Abs280nm确定每种酶的浓度。实例3:在底物脱脂脱淀粉玉蜀黍(DFDSM)中的可溶性和不可溶性膳食纤维的测量以及与在酶促孵育后测量的可溶性木糖的相关性将400mg的脱脂脱淀粉玉蜀黍(DFDSM)添加至NaOAc-缓冲液(5mL,pH5)中。将该混合物加热至90℃-100℃之间,然后添加Termamyl300DX(100μL,诺维信公司,巴格斯瓦德(Bagsvaerd),丹麦)并将该混合物孵育1hr。然后冷却该混合物并添加来自黑曲霉的淀粉葡糖苷酶(500μL,目录号E-AMGDF,用于麦格酶总淀粉和膳食纤维(MegazymeTotalStarchandDietaryFiber),麦格酶国际爱尔兰公司(MegazymeInternationalIreland),爱尔兰威克洛郡(Wicklow,Ireland)),并将样品在60℃下孵育过夜(16h)。然后将该混合物冷却并在2500xg在5℃下离心10min。收集该上清液,并将NaOAc-缓冲液(5mL,pH5)添加至残余物中并在2500xg,在5℃下离心10min。将该程序重复两次。然后收集这些上清液,进行汇集,并如在A中所述地分析其可溶性NSP。如在B中所述地分析该残余物的不可溶性NSP。A:可溶的NSP,上清液将这些经汇集的上清液稀释至固定体积,从中取出上清液的5mL等分试样。向该等分试样中添加20.1mL冷的99.9%乙醇并将该混合物置于冰上约15min,用于沉淀具有DP>10的聚合物。在2500xg、5℃下离心10min后,丢弃该上清液。将5mL冷的80%乙醇添加至该沉淀中并将该混合物置于冰上约15min。在2500xg、5℃下离心10min后,丢弃该上清液。通过添加MQ水(7.9mL)、肌醇(0.5mL,内部标准)和12MH2SO4(0.3mL)并在125℃高压灭菌55分钟进行该沉淀物的酸性水解。B:不可溶的NSP,残余物通过添加MQ水(74mL)、肌醇(10mL,内部标准)和12MH2SO4(3mL)并在125℃高压灭菌55分钟,将在AMG处理后获得的沉淀进行水解。GLC分析高压灭菌后,用氢硼化物还原这些样品以产生糖醇(alditolsugars)并过乙酰化将这些糖醇衍生化以变得易挥发用于在具有FID检测器的仪器上的GLC分析(皮特森(Pettersson)等人,(1995)“确定为中性糖残余物、糖醛酸残余物、和硫酸木素的总膳食纤维(乌普萨拉方法(theUppsalamethod)),协作研究”,AOAC国际杂志(J.AOACInt.)78:1030-1044)。相对于肌醇确定这些可溶性或不可溶性糖的浓度。溶解的木糖百分比当DFDSM与酶在40℃下孵育4个小时时,该酶溶解该底物中的木聚糖并且然后进一步通过酸水解这种经溶解的木聚糖。使用D-木糖测定试剂盒(Megazyme,目录号K-木糖)通过分光光度法测量释放的木糖。然后这种木糖(实际上是经酶溶解的木聚糖)与通过如上所述的GLC测量的底物的总木糖的量相关。该DFDSM包含99%不可溶性和1%可溶性木糖,总共14.81%木糖,其代表根据分析在样品(DFDSM)中存在的木糖聚合物(DP>10)的浓度。基于通过Megazyme试剂盒(基于样品重量计算释放)测量的木糖的释放,释放的木糖的量可以如下计算:例如1%释放自400mg的样品等于4mg的木糖。在400mg样品中,有400mgx14.81%木糖,等于59.22mg木糖。总木糖(不溶性+可溶性)释放是4mg/59.22mg的情况,其代表6.75mg木糖的释放,但是应该注意的是该值必须针对非酶补充对照获得的被动释放进行校正。这种校正值在本文中被定义为溶解的木糖百分比。实例4:用GH5木聚糖酶水解脱脂脱淀粉玉蜀黍(DFDSM)将脱脂脱淀粉玉蜀黍(DFDSM,400mg)添加至包含氯化钙(5mM)的pH5的水性乙酸钠(0.1M,3.9mL)溶液中,并将该混合物加热至40℃持续30分钟。添加100μL缓冲液或酶溶液并将该样品在40℃下加热4小时。将该样品冷却至5℃并离心(4000rpm,5℃)10分钟。将1.7mL的该样品转移至艾本德(Eppendorf)管中并通过加热至95℃持续10分钟将该酶灭活。然后将这些样品冷冻直至被水解。解冻该上清液并离心(14000rpm)5分钟。在玻璃管中用Milli-Q水(250μL)稀释该上清液(250μL)并添加HCl(1.63M,2.0mL)。将该反应加热至100℃持续1小时然后在冰浴中冷却。加入NaOH水溶液(1.3M,2.5mL),同时样品在冰上冷却,并且将样品储存在0-5℃下,同时使用木糖测定分析木糖含量。这些结果呈现于表2、表3和表4中。表2:使用GH5木聚糖酶SEQIDNO:2从DFDSM释放木糖表2显示当在不同的酶浓度下用SEQIDNO:2的GH5木聚糖酶孵育脱脂脱淀粉玉蜀黍(DFDSM)时,与空白和商业GH11木聚糖酶RonoxymeWX相比,酸性水解上清液后测量的木糖的量(总的木糖的干物质的%以及溶解的木糖%)。1如实例3中所述,计算溶解的木糖的百分比。2ABC:在不共享大写字母的列中的最小二乘值显著不同(P<0.05,所有对图基-克雷默(Tukey-Kramer)HSD)。表3:使用GH5木聚糖酶SEQIDNO:4从DFDSM释放木糖表3显示当在不同的酶浓度下用SEQIDNO:4的GH5木聚糖酶孵育脱脂脱淀粉玉蜀黍(DFDSM)时,与空白和商业GH11木聚糖酶RonoxymeWX相比,酸性水解上清液后测量的木糖的量(总的木糖的干物质的%以及溶解的木糖%)。1如实例3中所述,计算溶解的木糖的百分比。2ABC:在不共享大写字母的列中的最小二乘值显著不同(P<0.05,所有对图基-克雷默HSD)。表4:使用GH5木聚糖酶SEQIDNO:18从DFDSM释放木糖表4显示当在不同的酶浓度下用SEQIDNO:6的GH5木聚糖酶孵育脱脂脱淀粉玉蜀黍(DFDSM)时,与空白和商业GH11木聚糖酶RonoxymeWX相比,酸性水解上清液后测量的木糖的量(总的木糖的干物质的%以及溶解的木糖%)。1如实例3中所述,计算溶解的木糖的百分比。2ABC:在不共享大写字母的列中的最小二乘值显著不同(P<0.05,所有对图基-克雷默HSD)。正如从表2、表3和表4中可以看出,在从脱脂脱淀粉玉蜀黍释放木糖上,这些GH5木聚糖酶显著地更优于空白或商业的GH11木聚糖酶RonoxymeWX。实例5:玉米纤维胶水解物的酶法制备用于该水解的经纯化的酶是SEQIDNO:2、SEQIDNO:4、SEQIDNO:6、SEQIDNO:7、SEQIDNO:8(来自热纤维梭菌NzyTech(目录号(SKU):CR0061,科雷亚(Correia)等人,生物化学杂志(J.Biol.Chem.)2011,286:22510-22520)的五边形(pentamodular)GH5_34)、Pentopanmono、Pulpzyme、和Shearzyme。针对商业化产品,主要的木聚糖酶是通过阴离子交换色谱纯化的产物并进行测试。在3.1%DM作为最终浓度下用CFG1进行所有的反应。该测定还包含45mMMES缓冲液(pH6)并且在40℃下对于大多数酶经测试的最终的酶浓度为:41mg/L、18mg/L、7.3mg/L、和1.8mg/L持续2小时以及41mg/L持续24hr。由于低热稳定性,在30℃下测定SEQIDNO:7。在95℃下停止所有反应10min并用RI检测(Dionex)在ICS-3000上通过HPLC-SEC进行分析。使用的这些柱是PWXL保护柱并且PWXL-3000和-5000(日本东曹)串联连接。该洗脱液是50mMNaOAc(pH5)并且其流速为0.5ml/min。针对介于803Da-1.3kDa之间的普鲁兰(pullulan)分子量标准物和葡萄糖、麦芽三糖以及麦芽五糖比较保留时间。结果在下面这些表中,给出了该玉米纤维胶的特征以及由酶促处理产生的相应的产物。表5.碱性溶解的玉米纤维胶的特征。Mp(Da)Mn(Da)Mw(Da)多分散性2274921297013023442.3表6.使用在PentopanMonoTM中的主要的GH11木聚糖酶活性产生的玉米纤维胶水解产物的特征。这些计算基于高于DP2的部分。表7.使用在PulpzymeTM中的主要的GH11木聚糖酶活性产生的玉米纤维胶水解产物的特征。这些计算基于高于DP2的部分。表8.使用在ShearzymeTM500L中的主要的GH10木聚糖酶活性产生的玉米纤维胶水解产物的特征。这些计算基于高于DP2的部分。表9.使用SEQIDNO:2的家族GH5_35木聚糖酶产生的玉米纤维胶水解产物的特征。这些计算基于高于DP2的部分。表10.使用SEQIDNO:4的家族GH5_35木聚糖酶产生的玉米纤维胶水解产物的特征。这些计算基于高于DP2的部分。表11.使用SEQIDNO:6的家族GH5_21木聚糖酶产生的玉米纤维胶水解产物的特征。这些计算基于高于DP2的部分。表12.使用SEQIDNO:7的家族GH5_34木聚糖酶产生的玉米纤维胶水解产物的特征。这些计算基于高于DP2的部分。表13.使用SEQIDNO:8的家族GH5_34五边形(pentamodular)阿拉伯木聚糖酶(可从NzyTech获得)产生的玉米纤维胶水解产物的特征。这些计算基于高于DP2的部分。该玉米纤维胶水解产物数据清楚显示所有来自亚家族GH5_21、GH5_34和GH5_35的经测试的GH5木聚糖酶具有降解此种高度取代的木聚糖的能力。在所有测试的条件下,该DP1-DP2部分低于1%。所测试的两种家族GH11木聚糖酶完全不降解木聚糖,而存在于Shearzyme500L中的GH10木聚糖酶仅具有中等的玉米纤维胶降解能力。基于以上结果,测试了在淀粉乙醇工艺中添加GH5木聚糖酶的效果。实例6:使用在糖化/发酵中添加的GH5_35和GH5_21木聚糖酶的乙醇产率提高根据下表,在85℃下,使用商业α淀粉酶,LiquozymeTMSCDS(可从诺维信公司获得),进行来自经研磨的玉米的醪液的液化。将该液化的醪液分成三个部分并且根据下表进行同时糖化发酵。将商业葡糖淀粉酶产品,SpirizymeExcelTM(可从诺维信公司获得)用于糖化并且该酵母是EthanolRedTM(可从拉曼集团获得)。发酵之后,每天测量两次重量损失并且在发酵结束时进行HPLC。该对照样品仅包含这些葡糖淀粉酶。此外,一个样品包含葡糖淀粉酶和GH5_35木聚糖酶(U2AGD/SEQIDNO:4)并且另一个样品包含葡糖淀粉酶和GH5_21木聚糖酶(U2C9W/SEQIDNO:6)。乙醇产率示于图1中并表明了与该对照相比添加GH5木聚糖酶提高了乙醇产率。具体地,该GH5_35木聚糖酶导致约2%的增加。在此描述并且要求保护的本发明不限于在此披露的特定方面的范围,因为这些方面旨在作为本发明若干方面的说明。预期任何等效方面都处于本发明的范围内。实际上,除在此所示和描述的那些之外,本发明的不同修改对于本领域普通技术人员而言从前述描述将变得清楚。这样的修改也旨在落入所附权利要求的范围内。在有冲突的情况下,以包括定义的本披露为准。序列表<110>诺维信公司<120>使用GH5木聚糖酶从淀粉生产乙醇的工艺<130>12986-WO-PCT<160>8<170>PatentIn版本3.5<210>1<211>1722<212>DNA<213>人工的<220><223>编码来自Paenibacillusillinoisensis的GH5_35木聚糖酶的密码子优化基因<400>1atgaagaaaccgttggggaaaattgtcgcaagcaccgcactactcatttctgttgctttt60agttcatcgatcgcatcggctcatcagcaccaacaccagcatcctaggtgggttggcatg120ccgatgggcaaactgcatgtttcaggcaaaaatctggtcaatagcaataatcaaccggtt180ctgctgaatggctggcatcaaccgtcaggcgcatattggacatatcaatcatcaaactat240tatctgaatctgcatggaaataatcgccatgcggcaacactggcatatctgaaagatatc300acagatacatttgcagacacgtcaccgaaatatggctcaaatcatggctggaatatgaat360caggtccgcctgtttattgatcgcgaagatatgggagatgttgcagcaggcacatataac420tttgcaggcgttcaaacagttacacagaatgtcattattccgtatatccagtatgcgaaa480acgaaaggcgtttatgttgttctgggcctggattttacactgaaagatgatcaagcaaca540acagcgagcaatctgcagaaatttaaccaaatttggggctatctggcatcaagaccggaa600atcaaatcagcagataacgtccattttgaactgattaacgaaccggttaaaagctatgca660aatggccattggggaggctataatggcgaaaatgattttgtcgatcattggaatgatctg720cgcaattttcagaacagcattatttcaacaattcgctcacaaggcgcagataatgttatt780tgggcagcaggcctgggctataatcagttttattcactgacagcatcacatccgctgaca840gatccgctgaataactatggctatgcagttcattggtatccgggatatggcgcatatgat900aactttagcattctgcaggatcagtggaacacaaatgttaaagcagcagcagataaatat960ccgattaacatcacggaagtcacatggtttaaaaacaaacctggcgattcagcatactgg1020aatctttttaatggcagcaatgaaggctttggcacaaacacaaaaacgatctttaatgca1080gcaggcaatgttagcattgcagcacatatgaatggctttattctggaaccgggacaaaga1140tcatcatttgcagatccgacagcaggcctgaaatgggatggcgacgcatcaagatcagca1200atgggcagatttctgtttaattggtatcatgaaagagcgcaatcatatcctggcggaaat1260ggcggaggaccgacaacaggcctggtttcaggcgcaacatacaaaattgttgcacgccat1320agcaacaaagtcattgatgtccctggcggacaaaatcaaaacaatctgcaactgcaacag1380tggtcagatctgggaggcaatccgcaaaaatgggttctgacatcaattggcggaggctca1440tatacactgacatcagttaattcaccggacaaagtgattgatattcgcaatggcacactg1500acaaatggcgaagcagttcaactgatgtcaaatctgaatacaacagcgcagcattttaaa1560gtcaatgatctgggcaatggctattggagcattattaacgtcaacagcaacaaagcgatc1620gaagttgaaaatgcatcaacatcagatggcgcaaaactgcagcaaaatacatatacaggc1680gcaacaaatcagcagtggaaatttatcgcagtcagcaattaa1722<210>2<211>573<212>PRT<213>人工的<220><223>来自Paenibacillusillinoisensis的GH5_35木聚糖酶,包括来自克劳氏芽孢杆菌的信号肽和His-标签<400>2MetLysLysProLeuGlyLysIleValAlaSerThrAlaLeuLeuIle151015SerValAlaPheSerSerSerIleAlaSerAlaHisGlnHisGlnHis202530GlnHisProArgTrpValGlyMetProMetGlyLysLeuHisValSer354045GlyLysAsnLeuValAsnSerAsnAsnGlnProValLeuLeuAsnGly505560TrpHisGlnProSerGlyAlaTyrTrpThrTyrGlnSerSerAsnTyr65707580TyrLeuAsnLeuHisGlyAsnAsnArgHisAlaAlaThrLeuAlaTyr859095LeuLysAspIleThrAspThrPheAlaAspThrSerProLysTyrGly100105110SerAsnHisGlyTrpAsnMetAsnGlnValArgLeuPheIleAspArg115120125GluAspMetGlyAspValAlaAlaGlyThrTyrAsnPheAlaGlyVal130135140GlnThrValThrGlnAsnValIleIleProTyrIleGlnTyrAlaLys145150155160ThrLysGlyValTyrValValLeuGlyLeuAspPheThrLeuLysAsp165170175AspGlnAlaThrThrAlaSerAsnLeuGlnLysPheAsnGlnIleTrp180185190GlyTyrLeuAlaSerArgProGluIleLysSerAlaAspAsnValHis195200205PheGluLeuIleAsnGluProValLysSerTyrAlaAsnGlyHisTrp210215220GlyGlyTyrAsnGlyGluAsnAspPheValAspHisTrpAsnAspLeu225230235240ArgAsnPheGlnAsnSerIleIleSerThrIleArgSerGlnGlyAla245250255AspAsnValIleTrpAlaAlaGlyLeuGlyTyrAsnGlnPheTyrSer260265270LeuThrAlaSerHisProLeuThrAspProLeuAsnAsnTyrGlyTyr275280285AlaValHisTrpTyrProGlyTyrGlyAlaTyrAspAsnPheSerIle290295300LeuGlnAspGlnTrpAsnThrAsnValLysAlaAlaAlaAspLysTyr305310315320ProIleAsnIleThrGluValThrTrpPheLysAsnLysProGlyAsp325330335SerAlaTyrTrpAsnLeuPheAsnGlySerAsnGluGlyPheGlyThr340345350AsnThrLysThrIlePheAsnAlaAlaGlyAsnValSerIleAlaAla355360365HisMetAsnGlyPheIleLeuGluProGlyGlnArgSerSerPheAla370375380AspProThrAlaGlyLeuLysTrpAspGlyAspAlaSerArgSerAla385390395400MetGlyArgPheLeuPheAsnTrpTyrHisGluArgAlaGlnSerTyr405410415ProGlyGlyAsnGlyGlyGlyProThrThrGlyLeuValSerGlyAla420425430ThrTyrLysIleValAlaArgHisSerAsnLysValIleAspValPro435440445GlyGlyGlnAsnGlnAsnAsnLeuGlnLeuGlnGlnTrpSerAspLeu450455460GlyGlyAsnProGlnLysTrpValLeuThrSerIleGlyGlyGlySer465470475480TyrThrLeuThrSerValAsnSerProAspLysValIleAspIleArg485490495AsnGlyThrLeuThrAsnGlyGluAlaValGlnLeuMetSerAsnLeu500505510AsnThrThrAlaGlnHisPheLysValAsnAspLeuGlyAsnGlyTyr515520525TrpSerIleIleAsnValAsnSerAsnLysAlaIleGluValGluAsn530535540AlaSerThrSerAspGlyAlaLysLeuGlnGlnAsnThrTyrThrGly545550555560AlaThrAsnGlnGlnTrpLysPheIleAlaValSerAsn565570<210>3<211>1749<212>DNA<213>人工的<220><223>编码来自类芽孢杆菌属18054的GH5_35木聚糖酶的密码子优化基因<400>3atgaagaaaccgttggggaaaattgtcgcaagcaccgcactactcatttctgttgctttt60agttcatcgatagcatcagcacatcatcatcaccatcatcctaggttgacggttccgccc120ggcgccccggccgaggcttggtctggcatgcctacgcccaagcttcatgtcagcggcaac180caactggtaaatgcaaacggacagcctgtcctgctaagcgggtggcatcagccttccggg240tcctactggacgtatcagagcagcagttactacctggaccgtaacggcggaaaccggcat300gccgccaacctggcgtatctcaaggatatcacagacacctttacggacacctcgcccaaa360tacgggaacaaccacggctggtatatgaatcaggttcggctattcattgatcgcgaggat420atgggggatgtcgccgaaggcacttacaactttgctggattgcaggcggttacgcaaaat480gtgattattccgtacattaattatgcgagaacaaaagggctttatgtgacgctcggactg540gattttacgctcaaagacaatcaggcgaccacacaggccaatttggacaagttcaatcag600atttggagttatctagcatccagaccggaaataagaagcgctgacaacgtcatgtttgaa660attattaacgagccggtattgtcgtatgcggacggcagatggggcggtcatccgtccgac720cctcattttatagcattctggaacgatttgcgcagttttcaaaactctatcatctcctcc780attcgcgcacaaggagcggataatgtgatctgggcggccgggctcggctgggatcagtac840taccagttgtgtgcgtcgcatcctctgacggacccgctcaataatgtaggttatgcggtt900cactggtatccgggatatggagcaggggataactattcggtgcttcagcaacaatgggat960acgaatattaagccatgcgccgacaactatccgatcaatataacggaaacgacctggttc1020aagcgactgcctggcgattcggattactggaatttgttcaatggctcgagcgagggcttc1080ggcaaaaatacgaaagcaatctttactgcggcgggcaatgccagcattgccgtccatatg1140aacggctttttattggcgccgggagcaagaagctcatttgccgatccgactgccggtctg1200ttatacgacggaaatacagctcgagacggcatggcccgctttatattcgagtggtattac1260gaaagagcgcagttcttgccgtggaatggcatttggaacggactgttcacaggatcgacc1320tacaagttcgtgaatcgggcaaccggcaaaaatatggatgtgccgggcggacaaaacaat1380aataatctgcaattgaaccaatggacggataatggagcaacggcgcagcgctgggtcgta1440gacgatatgggcactttcaacaatatatatcgcatgaagagcgtcagctcatcggacggc1500aaggtaatggatgtccgcaacggcaccaaaaacaatggagaagccattcagcttatgcag1560gacttctcgaacacggcgcagcgtttccggattattcgacttagcaacggctattggagc1620attattaatgtcaacagcaacaaagcggttgaggtcgccggcggcgcttctcatgacgga1680gcgctattgcaacagaacatgtatcgcggagatcatcatcaacagtggcagctggttcag1740attcaataa1749<210>4<211>582<212>PRT<213>人工的<220><223>来自类芽孢杆菌属的GH5_35木聚糖酶,包括来自克劳氏芽孢杆菌的信号肽和His-标签<400>4MetLysLysProLeuGlyLysIleValAlaSerThrAlaLeuLeuIle151015SerValAlaPheSerSerSerIleAlaSerAlaHisHisHisHisHis202530HisProArgLeuThrValProProGlyAlaProAlaGluAlaTrpSer354045GlyMetProThrProLysLeuHisValSerGlyAsnGlnLeuValAsn505560AlaAsnGlyGlnProValLeuLeuSerGlyTrpHisGlnProSerGly65707580SerTyrTrpThrTyrGlnSerSerSerTyrTyrLeuAspArgAsnGly859095GlyAsnArgHisAlaAlaAsnLeuAlaTyrLeuLysAspIleThrAsp100105110ThrPheThrAspThrSerProLysTyrGlyAsnAsnHisGlyTrpTyr115120125MetAsnGlnValArgLeuPheIleAspArgGluAspMetGlyAspVal130135140AlaGluGlyThrTyrAsnPheAlaGlyLeuGlnAlaValThrGlnAsn145150155160ValIleIleProTyrIleAsnTyrAlaArgThrLysGlyLeuTyrVal165170175ThrLeuGlyLeuAspPheThrLeuLysAspAsnGlnAlaThrThrGln180185190AlaAsnLeuAspLysPheAsnGlnIleTrpSerTyrLeuAlaSerArg195200205ProGluIleArgSerAlaAspAsnValMetPheGluIleIleAsnGlu210215220ProValLeuSerTyrAlaAspGlyArgTrpGlyGlyHisProSerAsp225230235240ProHisPheIleAlaPheTrpAsnAspLeuArgSerPheGlnAsnSer245250255IleIleSerSerIleArgAlaGlnGlyAlaAspAsnValIleTrpAla260265270AlaGlyLeuGlyTrpAspGlnTyrTyrGlnLeuCysAlaSerHisPro275280285LeuThrAspProLeuAsnAsnValGlyTyrAlaValHisTrpTyrPro290295300GlyTyrGlyAlaGlyAspAsnTyrSerValLeuGlnGlnGlnTrpAsp305310315320ThrAsnIleLysProCysAlaAspAsnTyrProIleAsnIleThrGlu325330335ThrThrTrpPheLysArgLeuProGlyAspSerAspTyrTrpAsnLeu340345350PheAsnGlySerSerGluGlyPheGlyLysAsnThrLysAlaIlePhe355360365ThrAlaAlaGlyAsnAlaSerIleAlaValHisMetAsnGlyPheLeu370375380LeuAlaProGlyAlaArgSerSerPheAlaAspProThrAlaGlyLeu385390395400LeuTyrAspGlyAsnThrAlaArgAspGlyMetAlaArgPheIlePhe405410415GluTrpTyrTyrGluArgAlaGlnPheLeuProTrpAsnGlyIleTrp420425430AsnGlyLeuPheThrGlySerThrTyrLysPheValAsnArgAlaThr435440445GlyLysAsnMetAspValProGlyGlyGlnAsnAsnAsnAsnLeuGln450455460LeuAsnGlnTrpThrAspAsnGlyAlaThrAlaGlnArgTrpValVal465470475480AspAspMetGlyThrPheAsnAsnIleTyrArgMetLysSerValSer485490495SerSerAspGlyLysValMetAspValArgAsnGlyThrLysAsnAsn500505510GlyGluAlaIleGlnLeuMetGlnAspPheSerAsnThrAlaGlnArg515520525PheArgIleIleArgLeuSerAsnGlyTyrTrpSerIleIleAsnVal530535540AsnSerAsnLysAlaValGluValAlaGlyGlyAlaSerHisAspGly545550555560AlaLeuLeuGlnGlnAsnMetTyrArgGlyAspHisHisGlnGlnTrp565570575GlnLeuValGlnIleGln580<210>5<211>1902<212>DNA<213>人工的<220><223>编码来自未知生物的GH5_21木聚糖酶的密码子优化基因<400>5atgaagaaaccgttggggaaaattgtcgcaagcaccgcactactcatttctgttgctttt60agttcatcgatagcatcagcacatcatcatcaccatcatcctaggtggcgtggcatgaga120atgccggaactgtttatcaaaggcagatatctgatggcgaaagatatgaatggcaacgat180agcattgttaatctgcatggctttggccaaacatatagcgcgtattttaacggctatgcg240tggtgcaaaaatccggatggctcagttaattggggcaaaacaaaagatgcagcagcatgc300gttaaatggaataaagaacaaattggcctgatgctggatcatggctggaaagttaattgg360ctgagactgcatatggatccggcatggtcaaataatgaaacaaaagtcaatcaatggcag420agccaacatccgggaacatattattcagaaaatctgatcgtcgcgtttgatatgaacctg480tttaaaaaatatctggatgaaatctttattccgatggcggaatatgcgattgaaaacggc540atttatgttgttatgcgtccgcctggcgtttgtccgcaaaaactgacagttggagatgaa600tatcagcagtacctgattaaagtctggacatatgtttgcagccatgaaaaactgaaaaac660aatccgtatattatgtttgaactggcgaacgaaccgatcgatatgaatgatggcaatggc720aattatacgtcatggtcagatggctcacagaaaaactgcacgaaattttttcagaaaatt780gtcgacgaaattagagcagtcggctgcaataacattctgtgggttccgggactggcatat840caacaaaattatcaaggctatgtcaaatacccgattgtcggcgaaaatattggctttgca900gttcattgctatccgggatggtatggctcagattcagaagttgcatcagcagaacaacaa960attgtcacaaacggcaatacgtatgcggattttcaatcaggctggtcagcaagcattgat1020ggcgtttcaaaacttagaccgattatcgtcacagaaatggattgggcaccgaaaaaatac1080aattcatcatggggcaaagcaacgacaggcaaactgggaggcgttggctttggcaataac1140tttaaatacatcatggacaaaacaggcaacgttagctggatgctgtttacagatgcagat1200aaactggcgaaatatgatgattcaaaagcagatggcagcacgtttctgacagatccggaa1260gcatgccctagaccggtttatagatggtataaagaatatgcagaaccgggatggaaattt1320gttgaaacactggcagatgaattttacatgtttccgggaacaaacagcatttttagcccg1380aacatttgggaaaaaggcacactgacaaaaaatgatgatggctcaagaacactggtcaca1440ggccaatatggctttggaggctggaaatttggcggaggcctggatatgtcaggctataaa1500tacctggttctgaacctgacaaaagcaccggcatcaaatcaatggtcactgagactgttt1560gatgtcgataactattggacagacccgtatatgaaagatgtcaaatcaagcacaagagtc1620gttgtcgatctgcagaatatgaaaaatagcaaaggcgttaaagtcgacccgagccatatc1680tatattctgggcctgtggtcaacaggcggaacaccgattacaattaaagatatctatctg1740acaaataatagcgattattcaccggaatcaacaggcatttcagaaacacttgcagaaaaa1800agactggacacaccgatttataacctgtcaggccaaagagttacagaaccgagaaatggc1860catgtctatattcgcaacggcaaaaaattcatttacaaataa1902<210>6<211>633<212>PRT<213>人工的<220><223>包括来自克劳氏芽孢杆菌的信号肽和His-标签的GH5_21木聚糖酶<400>6MetLysLysProLeuGlyLysIleValAlaSerThrAlaLeuLeuIle151015SerValAlaPheSerSerSerIleAlaSerAlaHisHisHisHisHis202530HisProArgTrpArgGlyMetArgMetProGluLeuPheIleLysGly354045ArgTyrLeuMetAlaLysAspMetAsnGlyAsnAspSerIleValAsn505560LeuHisGlyPheGlyGlnThrTyrSerAlaTyrPheAsnGlyTyrAla65707580TrpCysLysAsnProAspGlySerValAsnTrpGlyLysThrLysAsp859095AlaAlaAlaCysValLysTrpAsnLysGluGlnIleGlyLeuMetLeu100105110AspHisGlyTrpLysValAsnTrpLeuArgLeuHisMetAspProAla115120125TrpSerAsnAsnGluThrLysValAsnGlnTrpGlnSerGlnHisPro130135140GlyThrTyrTyrSerGluAsnLeuIleValAlaPheAspMetAsnLeu145150155160PheLysLysTyrLeuAspGluIlePheIleProMetAlaGluTyrAla165170175IleGluAsnGlyIleTyrValValMetArgProProGlyValCysPro180185190GlnLysLeuThrValGlyAspGluTyrGlnGlnTyrLeuIleLysVal195200205TrpThrTyrValCysSerHisGluLysLeuLysAsnAsnProTyrIle210215220MetPheGluLeuAlaAsnGluProIleAspMetAsnAspGlyAsnGly225230235240AsnTyrThrSerTrpSerAspGlySerGlnLysAsnCysThrLysPhe245250255PheGlnLysIleValAspGluIleArgAlaValGlyCysAsnAsnIle260265270LeuTrpValProGlyLeuAlaTyrGlnGlnAsnTyrGlnGlyTyrVal275280285LysTyrProIleValGlyGluAsnIleGlyPheAlaValHisCysTyr290295300ProGlyTrpTyrGlySerAspSerGluValAlaSerAlaGluGlnGln305310315320IleValThrAsnGlyAsnThrTyrAlaAspPheGlnSerGlyTrpSer325330335AlaSerIleAspGlyValSerLysLeuArgProIleIleValThrGlu340345350MetAspTrpAlaProLysLysTyrAsnSerSerTrpGlyLysAlaThr355360365ThrGlyLysLeuGlyGlyValGlyPheGlyAsnAsnPheLysTyrIle370375380MetAspLysThrGlyAsnValSerTrpMetLeuPheThrAspAlaAsp385390395400LysLeuAlaLysTyrAspAspSerLysAlaAspGlySerThrPheLeu405410415ThrAspProGluAlaCysProArgProValTyrArgTrpTyrLysGlu420425430TyrAlaGluProGlyTrpLysPheValGluThrLeuAlaAspGluPhe435440445TyrMetPheProGlyThrAsnSerIlePheSerProAsnIleTrpGlu450455460LysGlyThrLeuThrLysAsnAspAspGlySerArgThrLeuValThr465470475480GlyGlnTyrGlyPheGlyGlyTrpLysPheGlyGlyGlyLeuAspMet485490495SerGlyTyrLysTyrLeuValLeuAsnLeuThrLysAlaProAlaSer500505510AsnGlnTrpSerLeuArgLeuPheAspValAspAsnTyrTrpThrAsp515520525ProTyrMetLysAspValLysSerSerThrArgValValValAspLeu530535540GlnAsnMetLysAsnSerLysGlyValLysValAspProSerHisIle545550555560TyrIleLeuGlyLeuTrpSerThrGlyGlyThrProIleThrIleLys565570575AspIleTyrLeuThrAsnAsnSerAspTyrSerProGluSerThrGly580585590IleSerGluThrLeuAlaGluLysArgLeuAspThrProIleTyrAsn595600605LeuSerGlyGlnArgValThrGluProArgAsnGlyHisValTyrIle610615620ArgAsnGlyLysLysPheIleTyrLys625630<210>7<211>620<212>PRT<213>解纤维素醋弧菌<400>7AlaAspProGlnArgGlyArgProTyrLeuAsnSerAlaArgThrThr151015PheValGlyAspAsnGlyGlnProLeuArgGlyProTyrIleSerThr202530GluTrpThrSerAlaAlaProTyrAspGlnIleAlaArgIleLysAsn354045LeuGlyPheAsnAlaValHisHisTyrAlaGluCysPheAspIleAsn505560TyrProAsnAlaGlySerLysSerProGlyTyrAlaAlaThrGluIle65707580AspLysValValGluArgThrArgGluLeuGlyLeuTyrLeuValMet859095ThrIleGlyAsnGlyAlaAsnAsnGlyAsnHisAsnThrArgTyrAla100105110LysAspPheTrpSerPheTyrSerSerArgTyrAlaAsnGluThrHis115120125ValLeuTyrGluIleHisAsnGluProValAlaTrpGlyProProTyr130135140SerSerThrThrAlaThrProThrGlyAlaValGluMetAsnValAsp145150155160ValTyrLysThrIleArgAlaAsnAlaProLysThrProValLeuIle165170175PheSerTyrSerValPheGlyGlyThrGlyGlyThrThrGluAlaLeu180185190LysAspIleGlnAlaPheAsnSerAlaValPheGlyLysGlnAspAla195200205ValTrpThrAsnGluAlaValAlaPheHisGlyTyrAlaGlyTrpGlu210215220AlaThrSerThrAlaValAspGlyLeuLeuLysAlaGlyTyrProCys225230235240PheMetThrGluTyrAlaGlyGlyAlaTrpGlySerGlyThrGlyGly245250255PheAspIleGlnLeuThrSerGluLeuGluArgMetGlyValSerTrp260265270LeuThrPheGlnTyrIleProProSerGlyValSerAspAspValThr275280285LysProGluTyrPheSerAlaLeuValGluAsnAlaGlyLeuSerTrp290295300LysProAspTyrGlyAsnTrpProAlaAlaArgGlyValHisGlyAsn305310315320GlyGlyLeuProArgLysThrSerThrTrpValAsnAsnPheLeuThr325330335GlyThrThrArgIleGluAlaGluAspPheAspTrpGlyGlyAsnAsp340345350ValSerPheTyrAspLysAspSerGluAsnLysGlyAlaGlnTyrArg355360365LeuAspGluAlaValAspIleGluThrThrLysAspAlaAspGlyGly370375380TyrAsnValGlyTrpIleGluAspGlyGluTrpLeuGluTyrThrIle385390395400TrpValGlnHisProGlyTyrTyrAsnLeuAlaLeuArgValAlaAsn405410415AsnSerGlyGlySerValGlnValAsnPheGlyAsnGlnAspLysThr420425430GlyThrTrpValLeuProValThrGlyGlyValGlnThrTrpLysThr435440445AspThrArgGlnValPheLeuGlySerGlyArgGlnLysLeuArgIle450455460AsnAlaLeuSerGlyGlyPheAsnLeuAsnTrpIleGluLeuSerPro465470475480ValSerThrGlyProIleAlaAspGlyThrTyrLysPheLeuAsnArg485490495AlaAsnThrMetThrLeuGlnGluValThrSerAsnAsnSerIleVal500505510ThrSerThrTyrLysGlyThrAlaAspGlnHisTrpLysIleGlnHis515520525IleGlyGlyGlyGlnTyrArgIleSerSerAlaGlyArgGlyTrpAsn530535540TrpAsnTrpTrpMetGlyPheGlyThrValGlyTrpTrpGlyThrGly545550555560SerGlyThrCysPheIleIleArgProThrGlyAspGlyTyrTyrArg565570575PheValLeuValAsnAspGlyThrAsnLeuGluIleSerAsnAsnAsp580585590SerSerLysIleGluGlyLysAlaTyrHisGluGlyAlaAsnGlnGln595600605TrpAlaIleGlnLeuProSerAlaProValPhePro610615620<210>8<211>865<212>PRT<213>未知的<220><223>来自NZYtech的商业GH5_34木聚糖酶<400>8AlaIleAlaLeuSerIleLeuSerPheIleProAsnArgAlaTyrAla151015SerProGlnArgGlyArgProArgLeuAsnAlaAlaArgThrThrPhe202530ValGlyAspAsnGlyGlnProLeuArgGlyProTyrThrSerThrGlu354045TrpThrAlaAlaAlaProTyrAspGlnIleAlaArgValLysGluLeu505560GlyPheAsnAlaValHisLeuTyrAlaGluCysPheAspProArgTyr65707580ProAlaProGlySerLysAlaProGlyTyrAlaValAsnGluIleAsp859095LysIleValGluArgThrArgGluLeuGlyLeuTyrLeuValIleThr100105110IleGlyAsnGlyAlaAsnAsnGlyAsnHisAsnAlaGlnTrpAlaArg115120125AspPheTrpLysPheTyrAlaProArgTyrAlaLysGluThrHisVal130135140LeuTyrGluIleHisAsnGluProValAlaTrpGlyProProTyrSer145150155160SerSerThrAlaAsnProProGlyAlaValAspMetGluIleAspVal165170175TyrArgIleIleArgThrTyrAlaProGluThrProValLeuLeuPhe180185190SerTyrAlaValPheGlyGlyLysGlyGlyAlaAlaGluAlaLeuLys195200205AspIleArgAlaPheAsnLysAlaValPheGlyAsnGluAsnAlaVal210215220TrpThrAsnGluAlaValAlaPheHisGlyTyrAlaGlyTrpGlnGlu225230235240ThrThrIleAlaValGluGluLeuLeuLysAlaGlyTyrProCysPhe245250255MetThrGluTyrAlaGlyGlyAlaTrpGlySerGlyMetGlyGlyLeu260265270AspValGluLeuThrTyrGluLeuGluArgLeuGlyValSerTrpLeu275280285ThrPheGlnTyrIleProProThrGlyValSerAspAspValThrLys290295300ProGluTyrPheSerAlaLeuValGluAsnSerGlyLeuSerTrpThr305310315320ProAspTyrGlyAsnTrpProAlaAlaArgGlyValTyrGlyAsnGly325330335GlyLeuAlaArgGluThrAlaThrTrpIleAsnAsnPheLeuThrGly340345350ThrThrArgIleGluAlaGluAspPheAspTrpGlyGlyAsnGlyVal355360365SerTyrTyrAspThrAspSerValAsnValGlyGlyGlnTyrArgPro370375380AspGluGlyValAspIleGluLysThrSerAspThrGlyGlyGlyTyr385390395400AsnValGlyTrpIleSerGluGlyGluTrpLeuGluTyrThrIleArg405410415ValArgAsnProGlyTyrTyrAsnLeuSerLeuArgValAlaGlyIle420425430SerGlySerArgValGlnValSerPheGlyAsnGlnAspLysThrGly435440445ValTrpGluLeuProAlaThrGlyGlyPheGlnThrTrpThrThrAla450455460ThrArgGlnValPheLeuGlyAlaGlyLeuGlnLysLeuArgIleAsn465470475480AlaLeuSerGlyGlyPheAsnLeuAsnTrpIleGluLeuSerProIle485490495SerThrGlyThrIleProAspGlyThrTyrLysPheLeuAsnArgAla500505510AsnGlyLysThrLeuGlnGluValThrGlyAsnAsnSerIleIleThr515520525AlaAspTyrLysGlyIleThrGluGlnHisTrpLysIleGlnHisIle530535540GlyGlyGlyGlnTyrArgIleSerSerAlaGlyArgGlyTrpAsnTrp545550555560AsnTrpTrpMetGlyPheGlyThrValGlyTrpTrpGlyThrGlySer565570575SerThrCysPheIleIleSerProThrGlyAspGlyTyrTyrArgIle580585590ValLeuValGlyAspGlyThrAsnLeuGlnIleSerSerGlyAspPro595600605SerLysIleGluGlyLysAlaPheHisGlyGlyAlaAsnGlnGlnTrp610615620AlaIleLeuProValSerAlaProAlaPheProThrGlyLeuSerAla625630635640ValLeuAspSerSerGlyAsnThrAlaAsnLeuThrTrpAsnAlaAla645650655ProGlyAlaAsnSerTyrAsnValLysArgSerThrLysSerGlyGly660665670ProTyrThrThrIleAlaThrAsnIleThrSerThrAsnTyrThrAsp675680685ThrGlyValAlaThrGlyThrLysTyrTyrTyrValValSerAlaVal690695700SerAsnGlyValGluThrLeuAsnSerAlaGluAlaIleLeuGlnTyr705710715720ProLysLeuThrGlyThrValIleGlyThrGlnGlySerTrpAsnAsn725730735IleGlyAsnThrIleHisLysAlaPheAspGlyAspLeuAsnThrPhe740745750PheAspGlyProThrAlaAsnGlyCysTrpLeuGlyLeuAspPheGly755760765GluGlyValArgAsnValIleThrGlnIleLysPheCysProArgSer770775780GlyTyrGluGlnArgMetIleGlyGlyIlePheGlnGlyAlaAsnLys785790795800GluAspPheSerAspAlaValThrLeuPheThrIleThrSerLeuPro805810815GlySerGlyThrLeuThrSerValAspValAspAsnProThrGlyPhe820825830ArgTyrValArgTyrLeuSerProAspGlySerAsnGlyAsnIleAla835840845GluLeuGlnPhePheGlyThrProAlaGlyGluGluAsnAspAspVal850855860His865当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1