布渣叶黄烷酮‑2‑羟基化酶及其编码基因与应用的制作方法
本发明属于基因工程技术领域。更具体地,涉及一种布渣叶黄烷酮-2-羟基化酶及其编码基因与应用。
背景技术:
植物类黄酮是一类广泛存在的小分子化合物,在植物体内大多以糖苷形式存在,其O-苷不胜枚举,而C-苷历历可数。黄酮C-苷的糖配基大多直接连接在其A环的C-6/C-8位,因母核各异,其种类也不尽相同。长期以来,植物类黄酮以其广泛的生理活性和良好的药食两用价值使得其应用研究如火如荼,其中大量的工作主要集中了各种黄酮类成份的分离纯化、结构鉴定、活性评价及产品开发上。但相关的基础研究,比如植物类黄酮次生代谢调控的研究,尤其是黄酮C-苷的次生代谢调控研究,相关的研究报道屈指可数。
近年来,随着研究的不断深入,黄酮 C-苷及其生物活性引起了越来越多的关注。从文献报道可知,其苷元以芹菜素或木犀草素较为常见,其糖配基以葡萄糖、鼠李糖、木糖或阿拉伯糖等居多,如牡荆苷、异牡荆苷、维采宁-2、夏佛塔苷、异夏佛塔苷等。研究发现,黄酮碳苷通常具有较好的抗炎、抗氧化、抗肿瘤、降血糖、降血压、护肝等活性。
黄酮C-苷化学性质较O-苷稳定,其化学合成难度也较大。迄今,获取黄酮C-苷的主要途径仍然是从植物原料中提取分离。随着现代生物技术的发展,植物次生代谢逐渐成为研究热点之一。而今,通过调控相应的生物合成关键酶基因的表达,以提高特定代谢产物的累积,在植物基因工程上的应用已越来越成熟,如青蒿素,人参皂苷等。在前期研究中发现,广东凉茶植物布渣叶中黄酮C-苷含量较高,组份较多,且大多数都是芹黄素-C-苷(Apigenin C-glycosides, ACGs)。因此,探索布渣叶ACGs生物合成相关酶的研究具有重要的意义。
技术实现要素:
本发明要解决的技术问题是克服现有布渣叶ACGs生物合成相关酶及其基因的研究不足,提供一种与其ACGs生物合成相关的关键酶基因,并进行了原核表达和功能验证,为进一步研究布渣叶ACG生源途径奠定了基础。
本发明的目的是提供一种布渣叶黄烷酮-2-羟基化酶MpF2H及其编码基因。
本发明另一目的是提供所述布渣叶黄烷酮-2-羟基化酶及其编码基因的应用。
本发明再一目的是提供所述布渣叶黄烷酮-2-羟基化酶的制备方法。
本发明上述目的通过以下技术方案实现:
一种编码布渣叶黄烷酮-2-羟基化酶的DNA分子(即布渣叶黄烷酮-2-羟基化酶编码基因MpF2H),其核苷酸序列如SEQ ID NO.1或SEQ ID NO.2所示(SEQ ID NO.1为DNA全长序列,SEQ ID NO.2为cDNA序列,即开放阅读框)。
一种在严格条件下能够与SEQ ID NO.1或SEQ ID NO.2所示序列杂交且编码具有相同功能蛋白质的DNA分子,也应在本发明的保护范围之内。
优选地,上述严格条件可为:在6×SSC、0.5% SDS的溶液中,在65℃下杂交,然后用2×SSC、0.1% SDS,以及1×SSC、0.1% SDS各洗膜一次。
一种与SEQ ID NO.1或SEQ ID NO.2所示序列至少具有80%、至少具有85%、至少具有90%、至少具有95%、至少具有96%、至少具有97%、至少具有98%或至少具有99%同源性且编码具有相同功能蛋白质的DNA分子,也应在本发明的保护范围之内。
一种布渣叶黄烷酮-2-羟基化酶,其氨基酸序列如SEQ ID NO.3所示。
一种将SEQ ID NO.3所示氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加所得具有相同功能的蛋白质,也应在本发明的保护范围之内。
本发明在前期研究发现布渣叶中黄酮C-苷含量较高的基础上,同时,综合布渣叶的转录组和代谢组数据,经差异基因表达和趋势分析,筛选发现了与其ACGs生物合成相关的关键酶基因 F2H(Flavanone 2-hydroxylase,MpF2H),并首次从布渣叶中克隆了 MpF2H基因cDNA 序列,并进行了原核表达和功能验证,为牡荆苷、异牡荆苷和维采宁-2等黄酮碳苷类成分的制备提供了一种新的途径。
另外,一种含有上述DNA分子的重组载体、表达盒、转基因细胞系或重组菌,也应在本发明的保护范围之内。
优选地,所述重组载体为将SEQ ID NO.1或SEQ ID NO.2所示序列插入pET30a(+)载体的Bam HI和 Hind III酶切位点间得到的载体,命名为pET30- MpF2H。
一种扩增上述DNA分子全长或其任意片段的引物对,也应在本发明的保护范围之内。
优选地,一种扩增SEQ ID NO.1或SEQ ID NO.2所示序列的引物对,其正反向引物序列分别如SEQ ID NO.4和SEQ ID NO.5所示。
另外,上述布渣叶黄烷酮-2-羟基化酶在作为或制备黄烷酮-2-羟基化酶制剂方面的应用,所述布渣叶黄烷酮-2-羟基化酶、上述DNA分子或上述重组载体、表达盒、转基因细胞系或重组菌在催化柚皮素生成2-羟基柚皮素方面的应用,以及上述DNA分子在制备布渣叶黄烷酮-2-羟基化酶中的应用,也都应在本发明的保护范围之内。
具体地,可以通过含有所述DNA分子的重组载体、表达盒、转基因细胞系或重组菌制备布渣叶黄烷酮-2-羟基化酶。
一种制备布渣叶黄烷酮-2-羟基化酶的方法,包括如下步骤:培养含有布渣叶黄烷酮-2-羟基化酶基因MpF2H的重组菌,收集培养物,得到布渣叶黄烷酮-2-羟基化酶。
更具体地,所述制备布渣叶黄烷酮-2-羟基化酶的方法,包括如下步骤:
S1.构建重组载体
将布渣叶黄烷酮-2-羟基化酶编码基因MpF2H的序列插入至pET30a(+)质粒的Bam HI和 Hind III酶切位点之间,得到重组载体pET30a-MpF2H;
S2. MpF2H蛋白的表达
将重组载体pET30a-MpF2H导入感受态细胞,得到重组菌;在重组菌的培养体系中加入异丙基硫代-β-D-半乳糖苷(IPTG),振荡诱导培养;
S3. MpF2H蛋白的纯化
S31.步骤S2所述振荡诱导培养6~8小时后收集培养物,离心收集菌体沉淀,加入Tris–HCl,超声破碎;
S32.步骤S31破碎后的产物离心,取上清液,即为MpF2H蛋白粗提取物,经过纯化得到MpF2H蛋白。
其中,优选地,步骤S1的具体方法为:
以布渣叶叶片cDNA为模板,利用正向引物MpF2H-F(序列如SEQ ID NO.4所示)和反向引物MpF2H-R(序列如SEQ ID NO.5所示)进行PCR扩增基因MpF2H片段,再用限制性内切酶 Bam HI 和 Hind III 对PCR扩增产物和pET-30a (+)质粒进行双酶切,经过连接酶连接(T4 DNA 连接酶 18℃连接过夜)后得到重组质粒pET30a-MpF2H。
其中,优选地,所述PCR扩增的反应体系为:cDNA 模板 1μL,MpF2H-F 1μL,MpF2H-R 1μL,10×ExTaq buffer 5μL,2.5mmol/L dNTP 4μL,ExTaq 酶 0.25μL,加水至50μL。
优选地,所述PCR扩增的反应程序为(降落PCR):95℃预变性 5 min,循环内98℃ 10s变性,从65℃每个循环降1℃退火,72℃延伸2min,10个循环;在55℃退火,进行20个循环;PCR反应循环后72℃延伸7 min,然后4℃保存。
另外,优选地,步骤S2所述感受态细胞为E.coli trans1 T1感受态细胞,得到重组菌E.coli trans1 T1-pET30a-MpF2H。
优选地,步骤S2中培养重组菌所用培养基为含50~100μg/ml(更优选为50μg/ml)卡拉霉素的LB培养基。
优选地,步骤S2中异丙基硫代-β-D-半乳糖苷(IPTG)的终浓度为0.5μM。
优选地,步骤S2中,重组菌的培养过程为:先37℃振荡培养至OD600=0.6~0.8后,加入IPTG,28℃振荡诱导培养。
更优选地,步骤S2中,重组菌的培养过程为:将重组菌接种于含50μg/ml卡拉霉素的LB培养基中,37℃振荡培养过夜,以1:100比例接种于新鲜的含抗生素卡拉霉素的培养基中,37℃振荡培养2~3h,待OD600=0.6~0.8 后,加入异丙基硫代-β-D-半乳糖苷(IPTG,终浓度为0.5μM),在28℃振荡诱导培养。
优选地,步骤S31所述离心的条件为12000r/min,4℃,离心1min。
优选地,步骤S31所述菌体沉淀还需重悬于PBS(pH 7.5)缓冲液中洗涤2次后,再加入Tris–HCl。步骤S31所述Tris–HCl的浓度为0.1mM,pH7.5。
优选地,步骤S31所述超声破碎的条件为:置于碎冰上超声破碎,20KHz、120W,开2秒、停10秒,循环10次。
优选地,步骤S32所述离心的条件为12000r/min,4℃,离心30min。
优选地,步骤S32所述纯化利用His-Bind蛋白纯化回收试剂盒进行。
本发明具有以下有益效果:
本发明首次从布渣叶中克隆得到布渣叶黄烷酮-2-羟基化酶 (MpF2H)基因,体外实验表明,MpF2H具有催化柚皮素生成2-羟基柚皮素的活性,是一种黄烷酮-2-羟基化酶,为进一步研究布渣叶ACGs生源途径奠定了基础;研究结果对其次生代谢调控及种质创新研究也将具有非常重要科学意义与应用价值。
利用本发明,可以以MpF2H基因为基础,通过基因工程技术来提高布渣叶等植物中牡荆苷、异牡荆苷和维采宁-2等黄酮碳苷类成分的含量,或通过生物合成技术制备牡荆苷、异牡荆苷和维采宁-2等黄酮碳苷类成分,为该类黄酮碳苷类成分的制备合成提供了一种新的途径。
附图说明
图1为重组表达载体pET30a-MpF2H的图谱。
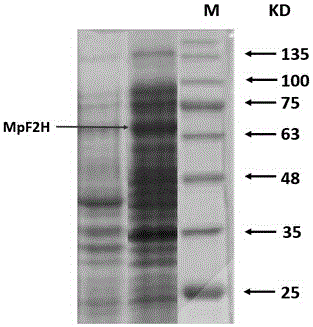
图2为MpF2H蛋白质的SDS-聚丙烯酰胺凝胶电泳结果。
具体实施方式
以下结合说明书附图和具体实施例来进一步说明本发明,但实施例并不对本发明做任何形式的限定。除非特别说明,本发明采用的试剂、方法和设备为本技术领域常规试剂、方法和设备。
除非特别说明,本发明所用试剂和材料均为市购。
以下实施例所用试剂盒材料来源:
同源重组克隆试剂盒( pEASY-Uni Seamless Cloning andAssembly Kit)、蛋白Marker、大肠杆菌感受态Trans1-T1、pET30a(+)载体以及DNA凝胶回收试剂盒购自北京全式金生物技术有限公司;RNA反转录试剂盒、BamH I限制性内切酶购自宝生物(大连)工程有限公司(TaKaRa)公司。引物合成和测序由广州生工有限公司完成。柚皮素和2-羟基柚皮素购自Chemface(武汉天植生物技术有限公司)。
实施例1 布渣叶黄烷酮-2-羟基化酶基因MpF2H的克隆
1、利用植物总 RNA 提取试剂盒提取布渣叶叶片的总 RNA,反转录得到cDNA为模板,用正向引物MpF2H-F(序列如SEQ ID NO.4所示)和反向引物MpF2H-R(序列如SEQ ID NO.5所示)进行PCR扩增。
正向引物:MpF2H-F(序列如SEQ ID NO.4所示)
5’-CGGGATCCATGATGCCCATGATACTTGAAT-3’;
反向引物:MpF2H-R(序列如SEQ ID NO.5所示)
5’-CCCAAGCTTTTAGGTTGCAAAGAGAGTGGGA3’。
其中,所述PCR扩增的反应体系为 :
cDNA 模板 1μL,MpF2H-F 1μL,MpF2H-R 1μL,10×ExTaq buffer 5μL,2.5mmol/L dNTP 4μL,ExTaq 酶 0.25 μL,加水至 50μL。
所述的PCR反应条件为(降落PCR):95℃预变性 5 min,循环内98℃ 10s变性,从65℃每个循环降1℃退火,72℃延伸2min,10个循环;在55℃退火,进行20个循环;PCR反应循环后72℃延伸7 min,然后4℃保存。
2、PCR 产物用1%琼脂糖凝胶电泳检测分析,用 PCR 产物纯化试剂盒纯化。用限制性内切酶 Bam HI 和 Hind III 对PCR纯化产物和pET-30a (+)质粒进行双酶切,T4 DNA 连接酶16℃连接过夜后转化大肠杆菌 Trans1-T1感受态细胞,在卡拉抗性平板上进行筛选,经菌液 PCR 和电泳检测后,挑选阳性克隆测序。
3、经过测序分析,得到布渣叶黄烷酮-2-羟基化酶基因MpF2H的序列,如SEQ ID NO.1所示(1826bp),其开放阅读框如SEQ ID NO.2所示(1557bp),编码的蛋白命名为布渣叶黄烷酮-2-羟基化酶MpF2H,该蛋白的氨基酸序列如SEQ ID NO.3所示(518aa)。
实施例2 布渣叶黄烷酮-2-羟基化酶MpF2H的表达和纯化
1、重组载体的构建
提取布渣叶叶片总RNA,反转录得到cDNA为模板,用用正向引物MpF2H-F(序列如SEQ ID NO.4所示)和反向引物MpF2H-R(序列如SEQ ID NO.5所示)进行PCR扩增。
PCR反应体系为:cDNA 模板 1μL,MpF2H-F 1μL,MpF2H-R 1μL,10×ExTaq buffer 5μL,2.5mmol/L dNTP 4μL,ExTaq 酶 - 0.5μL,加水至50μL。
反应条件为(降落PCR):95℃预变性 5 min,循环内98℃ 10s变性,从65℃每个循环降1℃退火,72℃延伸2min,10个循环;在55℃退火,进行20个循环;PCR反应循环后72℃延伸7 min,然后4℃保存。
PCR产物用纯化试剂盒纯化。用限制性内切酶 Bam HI 和 Hind III 对PCR纯化产物和pET-30a (+)质粒进行双酶切,T4 DNA 连接酶 18℃连接过夜后得到重组质粒,该重组载体为将MpF2H基因序列插入pET30a(+)质粒的Bam HI和 Hind III酶切位点间得到的载体,命名为pET30a-MpF2H(如图1所示)。
2、MpF2H蛋白的表达
(1)将目标质粒pET30a-MpF2H导入E.coli trans1 T1感受态细胞,得到重组菌E.colitrans1 T1-pET30a-MpF2H。
将空载体pET30a导入E.coli trans1 T1感受态细胞,得到对照菌E.coli trans1 T1-pET30a。
(2)将重组菌E.coli trans1 T1-pET30a-MpF2H接种于含50μg/ml卡拉霉素的LB培养基中,37℃振荡培养过夜,以1:100比例接种于新鲜的含抗生素卡拉霉素的培养基中,37℃振荡培养2~3h,待OD600=0.6~0.8 后,取1ml菌液作为诱导前对照,然后加入异丙基硫代-β-D-半乳糖苷(IPTG,终浓度为0 .5μM ),在28℃振荡诱导培养。以E.coli trans1 T1-pET30a为阴性对照。
(3)诱导培养6~8小时后,收集培养物,置于碎冰上超声破碎,20KHz,120W,开2秒,停10秒,循环10次,离心收集上清液。将上清液进行SDS-聚丙烯酰胺凝胶电泳,结果如图2所示,泳道M为Protein Marker(17-180kDa),可以看出,在分子量约为70 kD处,出现一条明显的特异蛋白质表达条带,与理论值一致,表明表达得到蛋白MpF2H(如图2所示)。
3、MpF2H蛋白纯化
(1)将重组菌E.coli trans1 T1-pET30a-MpF2H接种于含50μg/ml卡拉霉素的LB培养基中,37℃振荡培养过夜,以1:100比例接种于新鲜的含抗生素卡拉霉素的培养基中,37℃振荡培养2~3h,待OD600=0.6~0.8 后,加入异丙基硫代-β-D-半乳糖苷(IPTG,终浓度为0 .5μM ),在28℃振荡诱导培养,6~8小时后收集培养物,12000r/min,4℃,离心1min,收集菌体沉淀,重悬于PBS(pH 7.5)缓冲液中,洗涤2次,再加入0.1mM Tris–HCl(pH 7.5),置于碎冰上超声破碎,20KHz,120W,开2秒,停10秒,循环10次,12000r/min,4℃,离心30min,吸取上清液作为MpF2H蛋白粗提取物,置冰上备用。
(2)按照His-Bind蛋白纯化回收试剂盒说明书对上述粗提物进行纯化,得到MpF2H蛋白。
实施例3 布渣叶黄烷酮-2-羟基化酶MpF2H及其基因的功能研究
1、按照实施例2的方法将布渣叶黄烷酮-2-羟基化酶基因MpF2H表达出布渣叶黄烷酮-2-羟基化酶MpF2H,对其功能进行研究。
MpF2H蛋白功能验证:
以柚皮素为反应底物进行体外酶功能验证。在200μL反应体系中包括:0.1 M甘氨酸(KOH, pH 7.9),0.1 mM柚皮素,0.5 mM还原性的谷胱甘肽,1 mM NADPH,50μg MpF2H蛋白纯化产物。混匀,水浴30℃,孵育6~12h,加入400μL冰冷的甲醇终止反应,12000rpm离心10min,取上清,过0.22μm滤膜,进行LC-ESI-QTOF-MS分析。以柚皮素和2-羟基柚皮素对照品(购自Chemface)为对照,根据精确分子量、MS/MS碎片质荷比和色谱保留时间等进行目标产物鉴定。
UPLC-ESI-QTOF-MS分析条件如下:
液相条件:流动相为0.1%甲酸-乙腈(A),0.1%甲酸-水(B),流速0.2 mL/min,梯度洗脱程序为0min 7%B,3min 18%B,3.5min 7%B,5.5min 7%B;
质谱条件:离子源:ESI+,电离模式:正离子模式;离子源温度:150℃;脱溶剂温度450℃;毛细管电压:2500V脱溶剂气流速900L/hr,锥孔气流速50L/hr,裂解能20-30eV;高分辨质谱分析参数值如下表1所示。
表1高分辨质谱分析值
结果发现,柚皮素和2-羟基柚皮素标准品保留时间分别为4.24min 和3.52min,上述MpF2H酶促反应体系在4.24min检出2-羟基柚皮素相应特征峰,而对照反应体系没有检测到相应的特征峰。
对MpF2H酶促反应体系的特征峰进行质谱碎片分析,相应的分子离子精确分子量(m/z)和碎片离子精确分子量(m/z)为与标准品MS和MS/MS一致;至此可以确定酶促反应后确有2-羟基柚皮素生成。
结果表明,尽管相应的诱导表达条件、蛋白的分离纯化条件以及酶促反应体系可以进一步优化,但MpF2H融合蛋白确实具有催化柚皮素合成2-羟基柚皮素的活性,说明该基因为黄烷酮-2-羟基化酶编码基因。因此,可以以MpF2H基因为基础,通过基因工程技术来提高布渣叶等植物中牡荆苷、异牡荆苷和维采宁-2等黄酮碳苷类成分的含量,或通过生物合成技术制备牡荆苷、异牡荆苷和维采宁-2等黄酮碳苷类成分。
SEQUENCE LISTING
<110> 广东药科大学
<120> 布渣叶黄烷酮-2-羟基化酶及其编码基因与应用
<130>
<160> 5
<170> PatentIn version 3.3
<210> 1
<211> 1826
<212> DNA
<213> 布渣叶黄烷酮-2-羟基化酶基因MpF2H的DNA全长序列
<400> 1
ttccctctag aataattttg tttaacttta agaaggagat atacatatgc accatcatca 60
tcatcattct tctggtctgg tgccacgcgg ttctggtatg aaagaaaccg ctgctgctaa 120
attcgaacgc cagcacatgg acagcccaga tctgggtacc gacgacgacg acaaggccat 180
ggctgatatc ggatccatga tgcccatgat acttgaattc ctatcatatc tagctccctt 240
agttgtttca tttcttctcg ttaagagaat actcatctct accaacaagc aaggtcctaa 300
tgccaagcag cttcttcccc caggcccaat cgccttaccg atcattggcc acctccatct 360
cctgggcccc ttccttcatc aaaccttccg caagctctcc tcgcgctacg gtcccttaat 420
gtatcttcgt cttggctctg ttggctgcgt ggtggcttcc aatccagagt ttgccaaaga 480
gcttctcaaa acttacgagc tcgccttcgc tgcccgcatg cacaccgctg ccatcaccca 540
cctgacctac aactcgtcct ttgccttcgc accgtacgga acgtactgga aattcatcag 600
aaagttcagc acgtacgagc tcctaggcaa ccgtactctc gcccagtttc ttcccgttcg 660
aaccaaggaa ttgcaccatc tccttcagtt tgttcttggc aaggctaaag caggggaaag 720
cgtgaattta acccaagagc tgttgaaact aaccaacaac accatatcgc agatgatgct 780
gagcatgcgc tgttcgggga caggaaaccc tgccgactcg gttagaactc ttgtgcgaga 840
ggtgacagag atcttcggag agttcaacat ctcagatagc atatggtttt tgaaaaactg 900
ggacttgcag ggattcagaa agagattcga agacctgcat aggaggtttg atgcattgtt 960
ggagatgatt atgagagaac gtgagcgagt aagatcagaa agcaagcaaa agaaaggcga 1020
caatgttaac aaggtaaaag atttcctgga catcatgctt gacgttttgg aaaaggataa 1080
ctcggagtcg gaggtagatt ttaccagaaa tcacatcaag gccttaattt tggatttctt 1140
cacagccgct acagatacaa cggcaattgt acttgaatgg gcaatggcag agttgatcaa 1200
ccacccgaag ctactaaaaa tagctcagca agaaattgat caagttgtgg gaaaaggcag 1260
gttggtggaa gaatctgaca gcccacatct ccattacatc caagccatta ttaaagaaac 1320
atttcggctt cacccaccag tcccaatgat caataggaaa tcaatccaaa catgccaggt 1380
taagggatac acaatccctg ctgaatgtat ggtgtttgtg aacgtttggg ctatcggaag 1440
agatcccaag gtttggacag atccattgaa gtttcagcct gagagattcc tgaaatccga 1500
tgattctata gatgttagag ggcaacatta cgagctgttg cctttcgggt cagggaggag 1560
gggctgccct ggcgcttctt tggcgttgca ggagctgccc accactttgg ctgccatgat 1620
tcagtgcttt gactggaagg tggcgaatgg cgttgttgac atggttgaaa ggcctggact 1680
tacggctccc agggctaagg atctcgagtg tgttcctgtt gtacgcttca ctcccactct 1740
ctttgcaacc taaaagcttg cggccgcact cgagcaccac caccaccacc actgagatcc 1800
ggctgctaac aaagcccgaa agaagc 1826
<210> 2
<211> 1557
<212> DNA
<213> 布渣叶黄烷酮-2-羟基化酶基因MpF2H的cDNA序列
<400> 2
atgatgccca tgatacttga attcctatca tatctagctc ccttagttgt ttcatttctt 60
ctcgttaaga gaatactcat ctctaccaac aagcaaggtc ctaatgccaa gcagcttctt 120
cccccaggcc caatcgcctt accgatcatt ggccacctcc atctcctggg ccccttcctt 180
catcaaacct tccgcaagct ctcctcgcgc tacggtccct taatgtatct tcgtcttggc 240
tctgttggct gcgtggtggc ttccaatcca gagtttgcca aagagcttct caaaacttac 300
gagctcgcct tcgctgcccg catgcacacc gctgccatca cccacctgac ctacaactcg 360
tcctttgcct tcgcaccgta cggaacgtac tggaaattca tcagaaagtt cagcacgtac 420
gagctcctag gcaaccgtac tctcgcccag tttcttcccg ttcgaaccaa ggaattgcac 480
catctccttc agtttgttct tggcaaggct aaagcagggg aaagcgtgaa tttaacccaa 540
gagctgttga aactaaccaa caacaccata tcgcagatga tgctgagcat gcgctgttcg 600
gggacaggaa accctgccga ctcggttaga actcttgtgc gagaggtgac agagatcttc 660
ggagagttca acatctcaga tagcatatgg tttttgaaaa actgggactt gcagggattc 720
agaaagagat tcgaagacct gcataggagg tttgatgcat tgttggagat gattatgaga 780
gaacgtgagc gagtaagatc agaaagcaag caaaagaaag gcgacaatgt taacaaggta 840
aaagatttcc tggacatcat gcttgacgtt ttggaaaagg ataactcgga gtcggaggta 900
gattttacca gaaatcacat caaggcctta attttggatt tcttcacagc cgctacagat 960
acaacggcaa ttgtacttga atgggcaatg gcagagttga tcaaccaccc gaagctacta 1020
aaaatagctc agcaagaaat tgatcaagtt gtgggaaaag gcaggttggt ggaagaatct 1080
gacagcccac atctccatta catccaagcc attattaaag aaacatttcg gcttcaccca 1140
ccagtcccaa tgatcaatag gaaatcaatc caaacatgcc aggttaaggg atacacaatc 1200
cctgctgaat gtatggtgtt tgtgaacgtt tgggctatcg gaagagatcc caaggtttgg 1260
acagatccat tgaagtttca gcctgagaga ttcctgaaat ccgatgattc tatagatgtt 1320
agagggcaac attacgagct gttgcctttc gggtcaggga ggaggggctg ccctggcgct 1380
tctttggcgt tgcaggagct gcccaccact ttggctgcca tgattcagtg ctttgactgg 1440
aaggtggcga atggcgttgt tgacatggtt gaaaggcctg gacttacggc tcccagggct 1500
aaggatctcg agtgtgttcc tgttgtacgc ttcactccca ctctctttgc aacctaa 1557
<210> 3
<211> 518
<212> PRT
<213> 布渣叶黄烷酮-2-羟基化酶MpF2H氨基酸序列
<400> 3
Met Met Pro Met Ile Leu Glu Phe Leu Ser Tyr Leu Ala Pro Leu Val
1 5 10 15
Val Ser Phe Leu Leu Val Lys Arg Ile Leu Ile Ser Thr Asn Lys Gln
20 25 30
Gly Pro Asn Ala Lys Gln Leu Leu Pro Pro Gly Pro Ile Ala Leu Pro
35 40 45
Ile Ile Gly His Leu His Leu Leu Gly Pro Phe Leu His Gln Thr Phe
50 55 60
Arg Lys Leu Ser Ser Arg Tyr Gly Pro Leu Met Tyr Leu Arg Leu Gly
65 70 75 80
Ser Val Gly Cys Val Val Ala Ser Asn Pro Glu Phe Ala Lys Glu Leu
85 90 95
Leu Lys Thr Tyr Glu Leu Ala Phe Ala Ala Arg Met His Thr Ala Ala
100 105 110
Ile Thr His Leu Thr Tyr Asn Ser Ser Phe Ala Phe Ala Pro Tyr Gly
115 120 125
Thr Tyr Trp Lys Phe Ile Arg Lys Phe Ser Thr Tyr Glu Leu Leu Gly
130 135 140
Asn Arg Thr Leu Ala Gln Phe Leu Pro Val Arg Thr Lys Glu Leu His
145 150 155 160
His Leu Leu Gln Phe Val Leu Gly Lys Ala Lys Ala Gly Glu Ser Val
165 170 175
Asn Leu Thr Gln Glu Leu Leu Lys Leu Thr Asn Asn Thr Ile Ser Gln
180 185 190
Met Met Leu Ser Met Arg Cys Ser Gly Thr Gly Asn Pro Ala Asp Ser
195 200 205
Val Arg Thr Leu Val Arg Glu Val Thr Glu Ile Phe Gly Glu Phe Asn
210 215 220
Ile Ser Asp Ser Ile Trp Phe Leu Lys Asn Trp Asp Leu Gln Gly Phe
225 230 235 240
Arg Lys Arg Phe Glu Asp Leu His Arg Arg Phe Asp Ala Leu Leu Glu
245 250 255
Met Ile Met Arg Glu Arg Glu Arg Val Arg Ser Glu Ser Lys Gln Lys
260 265 270
Lys Gly Asp Asn Val Asn Lys Val Lys Asp Phe Leu Asp Ile Met Leu
275 280 285
Asp Val Leu Glu Lys Asp Asn Ser Glu Ser Glu Val Asp Phe Thr Arg
290 295 300
Asn His Ile Lys Ala Leu Ile Leu Asp Phe Phe Thr Ala Ala Thr Asp
305 310 315 320
Thr Thr Ala Ile Val Leu Glu Trp Ala Met Ala Glu Leu Ile Asn His
325 330 335
Pro Lys Leu Leu Lys Ile Ala Gln Gln Glu Ile Asp Gln Val Val Gly
340 345 350
Lys Gly Arg Leu Val Glu Glu Ser Asp Ser Pro His Leu His Tyr Ile
355 360 365
Gln Ala Ile Ile Lys Glu Thr Phe Arg Leu His Pro Pro Val Pro Met
370 375 380
Ile Asn Arg Lys Ser Ile Gln Thr Cys Gln Val Lys Gly Tyr Thr Ile
385 390 395 400
Pro Ala Glu Cys Met Val Phe Val Asn Val Trp Ala Ile Gly Arg Asp
405 410 415
Pro Lys Val Trp Thr Asp Pro Leu Lys Phe Gln Pro Glu Arg Phe Leu
420 425 430
Lys Ser Asp Asp Ser Ile Asp Val Arg Gly Gln His Tyr Glu Leu Leu
435 440 445
Pro Phe Gly Ser Gly Arg Arg Gly Cys Pro Gly Ala Ser Leu Ala Leu
450 455 460
Gln Glu Leu Pro Thr Thr Leu Ala Ala Met Ile Gln Cys Phe Asp Trp
465 470 475 480
Lys Val Ala Asn Gly Val Val Asp Met Val Glu Arg Pro Gly Leu Thr
485 490 495
Ala Pro Arg Ala Lys Asp Leu Glu Cys Val Pro Val Val Arg Phe Thr
500 505 510
Pro Thr Leu Phe Ala Thr
515
<210> 4
<211> 30
<212> DNA
<213> 正向引物MpF2H-F
<400> 4
cgggatccat gatgcccatg atacttgaat 30
<210> 5
<211> 31
<212> DNA
<213> 反向引物MpF2H-R
<400> 5
cccaagcttt taggttgcaa agagagtggg a 31
- 还没有人留言评论。精彩留言会获得点赞!