抑制VEGF‑A受体相互作用的结合蛋白的制作方法
本申请是国际申请日为2009年11月3日的国际申请pct/ep2009/064483进入中国、申请号为200980153604.7的题为“抑制vegf-a受体相互作用的结合蛋白”的发明专利申请的分案申请。
本发明涉及对vegf-a特异的重组结合蛋白,以及编码这类vegf-a结合蛋白的核酸,包括这类蛋白的药物组合物,和这类蛋白在治疗肿瘤和眼病中的用途。
背景技术:
血管生成(从先存在的血管生长出新的血管),是数种病理状况下的关键过程,包括肿瘤生长和眼病,尤其是眼的新生血管形成疾病,诸如老年性黄斑变性(amd)或糖尿病黄斑水肿(dme)(carmeliet,p.,nature438,932–936,2005)。血管内皮生长因子(vegfs)刺激血管生成和淋巴管生成,通过激活内皮细胞中的vegf受体(vegfr)酪氨酸激酶(ferrara,n.,gerber,h.p.andlecouter,j.,naturemed.9,669–676,2003)。
哺乳动物vegf家族由称为vegf-a,vegf-b,vegf-c,vegf-d(亦称为figf)和胎盘生长因子(plgf,亦称为pgf)的5种糖蛋白组成。vegf-a已被证明是抗血管生成疗法的有效靶标(ellis,l.m.andhicklin,d.j.,naturerev.cancer8,579-591,2008)。vegf-a配体结合并活化3种结构类似的iii型受体酪氨酸激酶,称为vegfr-1(亦称为flt1),vegfr-2(亦称为kdr)和vegfr-3(亦称为flt4)。vegf配体对这些酪氨酸激酶受体中的每一种都具有独特的结合特异性,这有助于它们功能的多样性。响应配体结合,vegfr酪氨酸激酶激活不同下游信号传导途径的网络。vegfr-1和vegfr-2主要发现于血管内皮,而vegfr-3大部分发现于淋巴内皮。这些受体都具有胞外结构域,单跨膜区和被激酶插入结构域中断的共有酪氨酸激酶序列。最近,神经毡蛋白(nrp-1)(最初被鉴定为神经元指引介质的脑信号蛋白/脑衰蛋白家族的受体),被证明作为vegf-a的同种型特异性受体。
各种同种型的vegf-a已知由vegf-a基因内8个外显子的选择性剪接产生。所有同种型包含外显子1-5和末端外显子,外显子8。外显子6和7(编码肝素-结合结构域)可以被包括或排除。这产生依据它们的氨基酸数目命名的蛋白家族:vegf-a165,vegf-a121,vegf-a189等。但是,外显子8在核苷酸序列中包含两个3'剪接位点,这可以被细胞使用产生具有相同长度的两个家族的同种型,但c端氨基酸序列不同(varey,a.h.r.etal.,britishj.cancer98,1366-1379,2008)。利用外显子8中最接近序列(导致包括外显子8a)产生vegf-axxx(“xxx”表示成熟蛋白的氨基酸数目)(促血管生成家族的同种型)。最近描述的抗血管生成vegf-axxxb同种型是利用远端剪接位点(从邻近的剪接位点沿着基因66bp更远)产生。这导致剪接出外显子8a,以及产生编码vegf-axxxb家族的mrna序列。vegf-a165是主要的促血管生成同种型,通常在各种人实体瘤中过表达。vegf-a165b是鉴定出的第一个外显子8b编码的同种型,显示具有抗血管生成作用(vareyetal.,在上述引文中;konopatskaya,o.etal.,molecularvision12,626-632,2006)。它是vegf-a的内源抑制形式,其降低vegf-a诱导的内皮细胞增殖和迁移。尽管它可以结合vegfr-2,vegf-a165b结合不引起受体磷酸化或下游信号传导途径的活化。
存在数种方法抑制vegf-a信号传导,包括通过抗体中和配体或受体,以及利用酪氨酸激酶抑制剂阻断vegf-a受体活化和信号传导。vegf-a靶向疗法已被证明是amd,dme,肾细胞癌和肝细胞癌中有效的单一治疗剂,而对转移性结肠直肠癌、非小细胞肺癌和转移性乳腺癌患者来说只有当其与化疗联用时才是有益的(narayanan,r.etal.,natrev.drugdiscov.5,815-816,2005;ellisandhicklin,在上述引文中)。
除了抗体,可以使用其他结合结构域来中和配体或受体(skerra,a.,j.mol.recog.13,167-187,2000;binz,h.k.,amstutz,p.andplückthun,a.,nat.biotechnol.23,1257-1268,2005)。这类新型结合结构域基于设计的重复结构域(wo02/20565;binz,h.k.,amstutz,p.,kohl,a.,stumpp,m.t.,briand,c.,forrer,p.,grütter,m.g.,andplückthun,a.,nat.biotechnol.22,575-582,2004)。wo02/20565描述了如何构建重复蛋白的大文库以及它们的一般应用。尽管如此,wo02/20565既没有公开具有对vegf-axxx的结合特异性的重复结构域的选择,也没有公开特异性结合vegf-axxx的重复结构域的具体重复序列基序。
用现有疗法靶向vegf-a不是在所有患者中都是有效的,或针对所有疾病有效的(例如,表达egfr的癌症)。以下事实甚至变得日益清楚,与vegf-a靶向疗法有关的治疗益处是复杂的,可能涉及多种机制(ellisandhicklin,在上述引文中)。例如,市售的抗vegf药物,诸如bevacizumab(avastin®)或ranibizumab(lucentis®)(参见wo96/030046,wo98/045331和wo98/045332)或正在临床开发的药物,诸如vegf-trap®(wo00/075319),不区分vegf-a的促血管生成形式和抗血管生成形式,因此这两种形式都被它们抑制。因此它们抑制血管生成,并且剥夺健康组织的一种重要存活因子,即vegf-axxxb,产生细胞毒性和剂量限制性副作用,这进而限制功效。现有抗vegf-a疗法的常见副作用是胃肠穿孔,出血,高血压,血栓栓子事件和蛋白尿(kamba,t.andmcdonald,d.m.,br.j.cancer96,1788-95,2007)。因此,现在需要改进的抗血管生成剂来治疗癌症和其他病理状况。
本发明的技术问题是鉴定新的抗血管生成剂,诸如具有对vegf-axxx的结合特异性的重复结构域,用于癌症和其他病理状况(例如眼病诸如amd或dme)的改进治疗。通过提供权利要求所表征的实施方案来解决该技术问题。
技术实现要素:
本发明涉及包括结合结构域的结合蛋白,其中所述结合结构域抑制vegf-axxx结合vegfr-2,并且所述结合结构域在热解折叠时具有高于40℃的中点变性温度(tm),当在pbs中37℃温育1天时在高达10g/l的浓度形成少于5%(w/w)的不溶性聚集体。更具体地,本发明涉及包括至少一个重复结构域的重组结合蛋白,其中所述重复结构域以低于10-7m的kd结合vegf-axxx,并抑制vegf-axxx结合vegfr-2。尤其是这类结合蛋白以低于10nm的ic50值抑制huvec球体(spheroids)的生芽(sprouting),并且这类结合蛋白与vegf-axxxb的相互作用的解离常数kd是其与vegf-axxx相互作用的kd至少10倍高。
尤其是,本发明涉及一种重组结合蛋白,包括具有对vegf-a的特异性的结合结构域,其是重复结构域,例如锚蛋白重复结构域,尤其是包括重复组件的锚蛋白重复结构域,所述重复组件具有锚蛋白重复序列基序
1d23g4tplhlaa56ghleivevllk7gadvna(seqidno:1)
其中1,2,3,4,5,6,和7,彼此独立,代表选自a,d,e,f,h,i,k,l,m,n,q,r,s,t,v,w和y的氨基酸残基。
本发明还涉及一种重组结合蛋白,包括具有对vegf-a的结合特异性的重复结构域,其与本发明的锚蛋白重复结构域具有至少70%的氨基酸序列同一性,或其包括与本发明的锚蛋白重复组件具有至少70%氨基酸序列同一性的重复组件,或其中锚蛋白重复组件的一个或更多个氨基酸残基被在比对锚蛋白重复单位时对应位置上发现的氨基酸残基替换。
本发明进一步涉及包括本发明重组结合蛋白的结合蛋白,其连接有一个或更多个其他的部分,例如,还结合vegfr-2或不同靶的部分,标记部分,促进蛋白纯化的部分,或提供改善的药代动力学的部分,例如聚乙二醇部分。在一些实施方案中,其他的部分是蛋白部分。在某些其他实施方案中,其他的部分是非蛋白的多聚体部分。
本发明进一步涉及编码本发明重组结合蛋白的核酸分子,以及包括一种或更多种上述结合蛋白或核酸分子的药物组合物。
本发明进一步涉及使用本发明的结合蛋白治疗癌症和其他病理状况(例如眼病诸如amd或dme)的方法。
附图说明
图1.挑选的设计的锚蛋白重复蛋白的特异性狗vegf-a164结合。
用粗提物elisa显示挑选的克隆与狗vegf-a164(vegf)和阴性对照蛋白(mbp,大肠杆菌麦芽糖结合蛋白)的相互作用。将生物素化的狗vegf-a164和mbp固定在neutravidin上。编号表示在抗狗vegf-a164或对应的人vegf-a165的核糖体展示中挑选的单个darpin克隆。a=吸光度。白色柱图指示结合狗vegf-a164,黑色柱图显示结合mbp的非特异性背景。
图2.挑选的darpin的球体生长抑制。
在存在各种浓度的(a)darpin#30(seqidno:29),对vegf-axxx具有特异性的darpin,或(b)darpinnc,对vegf-axxx没有特异性的阴性对照darpin的条件下,显示球体生长抑制测定中芽的长度。
图3.vegf-a同种型的特异性识别。
结合蛋白对vegf-a同种型的表面等离子体共振(spr)分析。
(a)和(b):avastin®的spr分析。将250nmavastin®添加到固定有狗vegf-a164(a)或狗vegf-a164b(b)的流动池100秒,然后通过缓冲液流动进行清洗。
(c)和(d):darpin#27(seqidno:16)的spr分析。将250nmdarpin#_27添加到固定有狗vegf-a164(c)或狗vegf-a164b(d)的流动池100秒,然后通过缓冲液流动进行清洗。ru=共振单位。
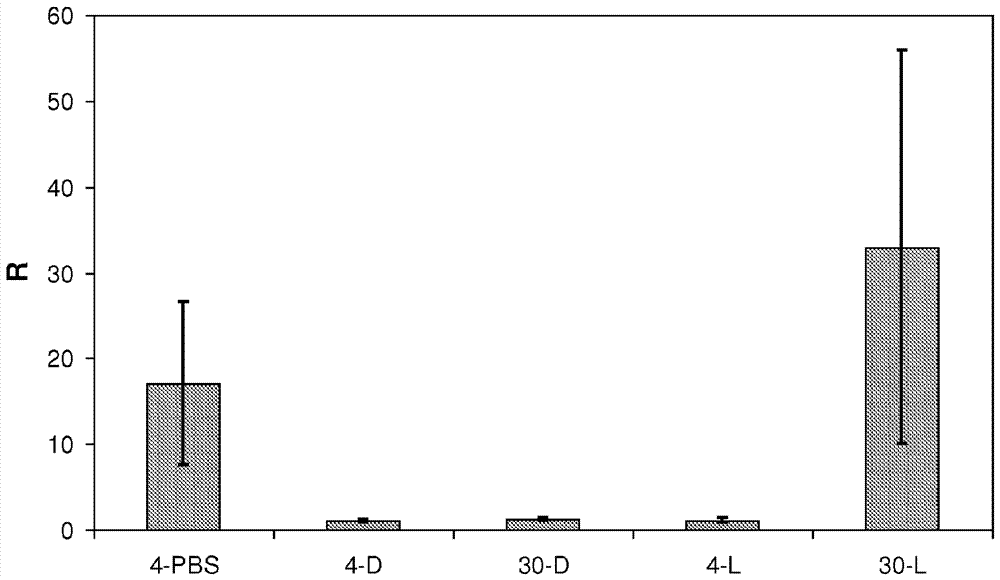
图4.兔眼中人vegf-a165的有效抑制。
血管渗漏兔模型显示与lucentis®相比,darpin在眼中抑制人vegf-a165的功效。在第1天,通过玻璃体内注射将pbs,darpin#30或lucentis®施用到每只兔的一只眼(治疗的眼)。在第4天或第30天,通过玻璃体内注射500ng人vegf-a165攻击每只兔的两只眼。所有眼在vegf-a165注射后48小时进行评估,通过测量静脉注射钠荧光素后1小时所有眼的玻璃体和视网膜中荧光素的含量。
r=治疗眼的荧光素测量值/未治疗眼的荧光素测量值的比例。标准偏差用误差线表示。4-pbs=pbs注射后4天的比例(对照);4-d=darpin#30注射后4天的比例;30-d=darpin#30注射后30天的比例;4-l=lucentis®注射后4天的比例;30-l=lucentis®注射后30天的比例。
具体实施方式
哺乳动物vegf-a以两个家族的选择性剪接同种型存在:(i)促血管生成的“vegf-axxx”同种型,通过外显子8的邻近剪接产生和(ii)抗血管生成的“vegf-axxxb”同种型,通过外显子8的远端剪接产生。优选的,本发明的结合结构域对狗,兔,猴或人来源的促血管生成vegf-axxx是特异性的。更优选的,本发明的结合结构域对人来源的促血管生成vegf-axxx是特异性的。最优选的,本发明的结合结构域对人vegf-a165是特异性的。
术语“蛋白”是指多肽,其中所述多肽的至少一部分具有或能够获得指定的三维结构,通过在其多肽链内和/或之间形成二级、三级或四级结构。如果蛋白包括两个或更多个多肽,单条多肽链可以通过例如两个多肽之间的二硫键非共价或共价相连。蛋白的一部分(其通过形成二级或三级结构单独具有或能够获得指定的三维结构)被称为“蛋白结构域”。这类蛋白结构域是本领域技术人员公知的。
当用于重组蛋白,重组蛋白结构域等等时术语“重组”表示所述多肽是利用相关领域技术人员公知的重组dna技术产生的。例如,可以将编码多肽的重组dna分子(例如通过基因合成产生)克隆入细菌表达质粒(例如pqe30,qiagen)。当这类构建的重组表达质粒被插入细菌(例如大肠杆菌)时,该细菌可以产生所述重组dna编码的多肽。对应产生的多肽被称为重组多肽。
术语“多肽标签”是指与多肽/蛋白连接的氨基酸序列,其中所述氨基酸序列用于所述多肽/蛋白的纯化,检测或靶向,或所述氨基酸序列改进多肽/蛋白的物化性质,或所述氨基酸序列具有效应物功能。结合蛋白的单个多肽标签,部分和/或结构域可以直接或通过多肽接头彼此连接。这些多肽标签都是本领域公知的,并且是本领域技术人员完全可以得到的。多肽标签的实例是小的多肽序列,例如,his,myc,flag,或strep-标签或部分诸如酶(例如酶,像碱性磷酸酶),其允许检测所述多肽/蛋白,或可用于靶向(诸如免疫球蛋白或其片段)和/或用做效应分子的部分。
术语“多肽接头”是指一种氨基酸序列,其能够连接,例如,两个蛋白结构域,多肽标签和蛋白结构域,蛋白结构域和非多肽部分诸如聚乙二醇或两个序列标签。这类其他的结构域,标签,非多肽部分和接头是相关领域的技术人员已知的。一系列实例提供于专利申请wo02/20565的说明书。这类接头的具体实例是可变长度的甘氨酸-丝氨酸-接头;优选的,所述接头具有2-16个氨基酸的长度。
在本发明的背景下,术语“多肽”涉及由多个(即两个或更多个)氨基酸通过肽键连接的一条或更多条链组成的分子。优选的,多肽由通过肽键连接的超过8个氨基酸组成。
术语“结合蛋白”是指包括一个或更多个结合结构域蛋白,下面将进一步解释。优选的,所述结合蛋白包括最多4个结合结构域。更优选的,所述结合蛋白包括最多两个结合结构域。最优选的,所述结合蛋白只包括一个结合结构域。此外,任何这类结合蛋白可能包括其他的不是结合结构域的蛋白结构域,多聚化部分,多肽标签,多肽接头和/或非蛋白聚合物分子。多聚化部分的实例是免疫球蛋白重链恒定区,其配对提供功能性免疫球蛋白fc结构域,以及亮氨酸拉链或包括游离巯基的多肽,其在这样的两个多肽之间形成分子间二硫键。非蛋白聚合物分子的实例是羟乙基淀粉(hes),聚乙二醇(peg),聚丙二醇,或聚氧化烯。
术语“peg化”表示peg部分共价连接于,例如,本发明的多肽。
术语“结合结构域”表示一种蛋白结构域,其显示与蛋白支架相同的“折叠”(三维结构)并具有预定的特性,如下文所述。这类结合结构域可以通过合理的或最常见的组合蛋白工程技术(这是本领域已知的技能)获得(skerra,2000,在上述引文中;binzetal.,2005,在上述引文中)。例如,具有预定特性的结合结构域可以通过包括下列步骤的方法获得:(a)提供显示与下文指定的蛋白支架相同折叠的蛋白结构域的多样集合;和(b)筛选所述多样集合和/或从所述多样集合进行挑选以获得具有所述预定特性的至少一种蛋白结构域。根据使用的筛选和/或挑选系统,蛋白结构域的多样集合可以通过数种方法提供,可以包括使用本领域技术人员公知的方法,诸如噬菌体展示或核糖体展示。
术语“蛋白支架”表示具有暴露表面区域的蛋白,在该区域中氨基酸插入、取代或缺失是高度耐受的。可用于产生本发明结合结构域的蛋白支架的实例是抗体或其片段,诸如单链fv或fab片段,来自金黄色葡萄球菌(staphylococcusaureus)的蛋白a,来自大菜粉蝶(pierisbrassicae)的胆汁三烯结合蛋白或其他脂质运载蛋白,锚蛋白重复蛋白或其他重复蛋白,和人纤连蛋白。蛋白支架是本领域技术人员已知的(binzetal.,2005,在上述引文中;binzetal.,2004,在上述引文中)。
术语“预定特性”是指一种特性,诸如结合靶,阻断靶,活化靶介导的反应,酶活性,以及相关其他特性。根据所需特性的类型,普通技术人员将能够确定筛选和/或挑选具有所需特性的结合结构域的格式和必需步骤。优选的,所述预定特性是结合靶。
优选的,本发明的结合蛋白不是抗体或其片段,诸如fab或scfv片段。抗体及其片段是本领域技术人员公知的。
还优选的,本发明的结合结构域不包括存在于抗体中的免疫球蛋白折叠和/或iii型纤连蛋白结构域。免疫球蛋白折叠是常见的全部-β蛋白折叠,其由排列为两个β片层的约7个反平行β链的双层夹层组成。免疫球蛋白折叠是本领域技术人员公知的。例如,包括免疫球蛋白折叠的这类结合结构域描述于wo07/080392或wo08/097497。
进一步优选的,本发明的结合结构域不包括免疫球蛋白样结构域(如在vegfr-1或vegfr-2中发现)。这类结合结构域描述于wo00/075319。
优选的结合结构域是具有抗血管生成作用的结合结构域。结合结构域的抗血管生成作用可以通过本领域技术人员公知的测定法来确定,诸如实例2描述的huvec球体的生芽测定法。
进一步优选的是包括70-300个氨基酸的结合结构域,尤其是100-200个氨基酸。
进一步优选的是不含游离cys残基的结合结构域。游离的cys残基不参与二硫键的形成。甚至更优选的是不含任何cys残基的结合结构域。
本发明优选的结合结构域是重复结构域或设计的重复结构域,优选的如wo02/20565所述。
尤其优选的结合结构域是设计的锚蛋白重复结构域(binz,h.k.etal.,2004,在上述引文中),优选的如wo02/20565所述。设计的锚蛋白重复结构域的实例显示于实施例中。
下文关于重复蛋白的定义是基于专利申请wo02/20565。专利申请wo02/20565还包含重复蛋白特征、技术和应用的一般说明。
术语“重复蛋白”是指包括一个或更多个重复结构域的蛋白。优选的,所述重复蛋白中的每一个包括最多4个重复结构域。更优选的,所述重复蛋白中的每一个包括最多两个重复结构域。最优选的,每一重复蛋白只包括一个重复结构域。此外,所述重复蛋白可以包括其他的非重复蛋白结构域,多肽标签和/或多肽接头。
术语“重复结构域”是指包括作为结构单位的两个或更多个连续重复单位(组件)的蛋白结构域,其中所述结构单位具有相同折叠并紧密堆积产生。例如,具有节点疏水核(jointhydrophobiccore)的超螺旋结构。
术语“设计的重复蛋白”和“设计的重复结构域”分别是指重复蛋白或重复结构域,通过专利申请wo02/20565说明的创造性方法获得。设计的重复蛋白和设计的重复结构域是合成的,而非来自自然界。它们分别是人工的蛋白或结构域,通过表达相应设计的核酸获得。优选的,表达在真核或原核细胞(诸如细菌细胞)中完成,或使用不含细胞的体外表达系统。
术语“结构单位”是指多肽局部有序的部分,由沿着多肽链彼此邻近的二级结构的两个或更多个区段的三维相互作用形成。这类结构单位显示结构基序。术语“结构基序”是指至少一个结构单位中存在的二级结构元件的三维结构。结构基序是本领域技术人员公知的。单独的结构单位不能获得确定的立体结构;但是,它们的连续排列,例如作为重复结构域中的重复组件,导致相邻单位的相互稳定,以产生超螺旋结构。
术语“重复单位”是指包括一个或更多个天然存在的重复蛋白的重复序列基序的氨基酸序列,其中所述“重复单位”被发现是多个拷贝,其显示对确定蛋白折叠的全部所述基序共有的限定折叠拓扑结构。这类重复单位包括骨架残基和相互作用残基。这类重复单位的实例是犰狳(armadillo)重复单位,富含亮氨酸的重复单位,锚蛋白重复单位,三角形四肽(tetratricopeptide)重复单位,heat重复单位,和富含亮氨酸的变体重复单位。包含两个或更多个这类重复单位的天然存在的蛋白被称作“天然存在的重复蛋白”。当彼此比较时,重复蛋白的单个重复单位的氨基酸序列可以具有很多突变,取代,添加和/或缺失,但仍然基本上保留重复单位的一般模式或基序。
术语“骨架残基”涉及重复单位的氨基酸残基,或重复组件的相应氨基酸残基,其有助于折叠的拓扑结构,即其有助于所述重复单位(或组件)的折叠或有助于相邻单位(或组件)的相互作用。这类贡献可以是与重复单位(组件)中其他残基的相互作用,或对α-螺旋或β-片层中发现的多肽主链构象的影响,或形成线性多肽或环的氨基酸延伸。
术语“靶相互作用残基”是指重复单位的氨基酸残基,或重复组件的相应氨基酸残基,其有助于与靶物质的相互作用。这类贡献可以是与靶物质的直接相互作用,或对其他直接相互作用残基的影响,例如通过稳定重复单位(组件)的多肽构象以允许或增强直接相互作用残基与所述靶的相互作用。这类骨架和靶相互作用残基可以通过以下方法鉴定:对通过物化方法(诸如x射线结晶,nmr和/或cd光谱法)获得的结构数据进行分析,或与结构生物学和/或生物信息学技术人员公知的已知的和相关的结构信息进行比较。
优选的,用于推断重复序列基序的重复单位是同源重复单位,其中重复单位包括相同的结构基序,并且超过70%的所述重复单位的骨架残基是彼此同源的。优选的,超过80%的所述重复单位的骨架残基是同源的。最优选的,超过90%的所述重复单位的骨架残基是同源的。用于确定多肽之间同源性百分比的计算机程序(诸如fasta,blast或gap)是本领域技术人员已知的。进一步优选的,用于推断重复序列基序的重复单位是同源重复单位(获自从靶上挑选的重复结构域),例如描述于实施例1,并具有相同的靶特异性。
术语“重复序列基序”是指氨基酸序列,其从一个或更多个重复单位推断得出。优选的,所述重复单位来自于对相同靶具有结合特异性的重复结构域。这类重复序列基序包括骨架残基位置和靶相互作用残基位置。所述骨架残基位置对应于重复单位的骨架残基的位置。同样,所述靶相互作用残基位置对应于重复单位的靶相互作用残基的位置。重复序列基序包括固定位置和随机位置。术语“固定位置”是指重复序列基序中的氨基酸位置,其中所述位置被设置为特定氨基酸。最常见的,这类固定位置对应于骨架残基的位置和/或靶相互作用残基(对某靶特异的)的位置。术语“随机位置”表示重复序列基序中的氨基酸位置,其中在所述氨基酸位置允许两个或更多个氨基酸,例如,允许常见的二十种天然存在的氨基酸的任意一种,或允许二十种天然存在氨基酸的大部分,诸如除了半胱氨酸以外的氨基酸,或除了甘氨酸、半胱氨酸和脯氨酸以外的氨基酸。最常见的,这类随机位置对应靶相互作用残基的位置。但是,骨架残基的一些位置也可以是随机的。
术语“折叠的拓扑结构”是指所述重复单位的三级结构。通过形成至少部分α-螺旋或β-片层的氨基酸延伸,或形成线性多肽或环的氨基酸延伸,或α-螺旋、β-片层和/或线性的多肽/环的任意组合,确定折叠的拓扑结构。
术语“连续的”是指一种排列,其中重复单位或重复组件串联排列。在设计的重复蛋白中,存在至少2个,通常约2到6个,尤其是至少约6个,通常是20或更多个重复单位。在大多数情况下,重复单位将显示高度的序列同一性(对应位置相同的氨基酸残基)或序列相似性(氨基酸残基是不同的,但具有类似的物化特性),一些氨基酸残基可能是在天然存在的蛋白中发现的不同重复单位中高度保守的关键残基。但是,只要维持常见的折叠拓扑结构,在天然存在的蛋白中发现的不同重复单位之间的氨基酸插入和/或缺失和/或取代造成的高度序列变异性也是可能的。
通过物理化学法诸如x射线结晶、nmr或cd光谱法直接检测重复蛋白的折叠拓扑结构的方法是本领域技术人员公知的。用于鉴定和确定重复单位或重复序列基序或鉴定包括这类重复单位或基序的相关蛋白的家族的方法,诸如同源性查找(blast等),在生物信息学领域是完善的,并且是本领域技术人员公知的。改进初始重复序列基序的步骤可以包括迭代过程。
术语“重复组件”是指设计的重复结构域的重复氨基酸序列,其最初源自天然存在的重复蛋白的重复单位。重复结构域包括的每个重复组件来源于天然存在的重复蛋白的家族或亚家族的一个或更多个重复单位,例如,armadillo重复蛋白或锚蛋白重复蛋白的家族。
“重复组件”可以包括具有在相应重复组件的所有拷贝存在氨基酸残基的位置("固定位置")和具有不同或"随机"氨基酸残基的位置("随机位置")。
术语“加帽组件”是指融合到重复结构域的n或c端重复组件的多肽,其中所述加帽组件与所述重复组件形成紧密的三级相互作用从而提供保护所述重复组件的疏水核在侧面不与来自溶剂的连续重复组件接触的帽。所述n和/或c端加帽组件可以是,或可以来自,邻近于重复单位的天然存在的重复蛋白中发现的加帽单位或其他结构域。术语“加帽单位”是指天然存在的折叠多肽,其中所述多肽限定与重复单位n或c端融合的特定结构单位,其中所述多肽与所述重复单位形成紧密的三级相互作用从而提供保护所述重复单位的疏水核在一侧免于溶剂的帽。这类加帽单位可以具有类似所述重复序列基序的序列。加帽组件和加帽重复描述于wo02/020565。例如,seqidno:21的n端加帽组件由位置1到32的氨基酸编码。此外优选的是在位置5具有甘氨酸或天冬氨酸残基的这类n端加帽组件。
术语“靶”是指单个分子诸如核酸分子,多肽或蛋白,碳水化合物,或任何其他天然存在的分子,包括这类单个分子的任意部分,或两个或更多个这类分子的复合物。靶可以是全细胞或组织样品,或它可以是任何非天然的分子或部分。优选的,靶是天然存在或非天然的多肽或包含化学修饰的多肽,例如通过天然或非天然的磷酸化,乙酰化或甲基化进行的修饰。在本发明的特定应用中,靶是vegf-axxx或vegfr-2。
术语“共有序列”是指一种氨基酸序列,其中所述共有序列是通过多个重复单位的结构和/或序列比对获得的。利用两个或更多个结构和/或序列比对的重复单位,并允许比对中的缺口,有可能确定每个位置最常见的氨基酸残基。共有序列是包括每个位置最常出现的氨基酸的序列。如果在单个位置有两个或更多个氨基酸出现高于平均值,则共有序列可以包括这些氨基酸的亚组。所述两个或更多个重复单位可以获自单个重复蛋白中包括的重复单位,或获自两个或更多个不同的重复蛋白。
共有序列以及确定它们的方法是本领域技术人员公知的。
“保守氨基酸残基”是在共有序列的某些位置发现的氨基酸。如果在所述两个或更多个重复单位中发现具有类似概率的两个或更多个(例如3,4或5个)氨基酸残基,共有氨基酸可以是最常发现的氨基酸之一或所述两个或更多个氨基酸残基的组合。
进一步优选的是非天然存在的结合蛋白或结合结构域。
术语“非天然存在”表示合成的或不是来自自然界的,更具体地,该术语表示由人工制备。术语“非天然存在的结合蛋白”或“非天然存在的结合结构域”表示所述结合蛋白或所述结合结构域是合成的(即通过化学合成从氨基酸产生)或重组的并且不是来自自然界。“非天然存在的结合蛋白”或“非天然存在的结合结构域”分别是通过表达相应设计的核酸获得的人造蛋白或结构域。优选的,表达在真核或细菌细胞中完成,或使用不含细胞的体外表达系统。进一步的,该术语表示所述结合蛋白或所述结合结构域的序列在序列数据库中不作为非人工序列条目存在,例如在genbank,embl-bank或swiss-prot中。这些数据库和其他类似的序列数据库是本领域技术人员公知的。
本发明涉及包括结合结构域的结合蛋白,其中所述结合结构域抑制vegf-axxx结合vegfr-2,并且所述结合蛋白和/或结合结构域在热解折叠时具有高于40℃的中点变性温度(tm),当在磷酸缓冲盐溶液(pbs)中37℃温育1天时,在高达10g/l的浓度形成低于5%(w/w)的不溶性聚集体。
结合结构域可以抑制vegf-axxx结合vegfr-2,通过以以下方式结合vegf-axxx或结合vegfr-2:vegf-axxx和vegfr-2之间的表观解离常数(kd)增加超过102倍,优选的超过103倍,更优选的超过104倍,更优选的超过105倍,和最优选的超过106倍。优选的,结合结构域与vegf-axxx或vegfr-2的相互作用的kd是小于10-7m,优选的小于10-8m,更优选的小于10-9m,更优选的小于10-10m,和最优选的小于10-11m。用于确定蛋白-蛋白相互作用的解离常数的方法,诸如基于表面等离子体共振(spr)的技术,是本领域技术人员公知的。
优选的结合结构域结合vegf-axxx。甚至更优选的是结合人vegf-a165的结合结构域。
术语“pbs”表示包含137mmnacl,10mm磷酸盐和2.7mmkcl并且具有ph7.4的磷酸盐缓冲水溶液。
优选的,结合蛋白和/或结合结构域在热解折叠时具有高于45℃的中点变性温度(tm),更优选的高于50℃,更优选的高于55℃,和最优选的高于60℃。本发明的结合蛋白或结合结构域在生理条件下具有确定的二级和三级结构。这类多肽的热解折叠导致其三级和二级结构的丧失,这可以例如通过圆二色性(cd)测量进行追踪。在热解折叠时结合蛋白或结合结构域的中点变性温度对应于所述蛋白或结构域在热变性时(通过从10℃缓慢升温到约100℃)的生理缓冲液中协同转换的中点的温度。热解折叠时中点变性温度的确定是本领域技术人员公知的。热解折叠时结合蛋白或结合结构域的所述中点变性温度提示所述多肽的热稳定性。
还优选的是当在37℃的pbs中温育超过5天,优选的超过10天,更优选的超过20天,更优选的超过40天,和最优选的超过100天时,在高达20g/l,优选的高达40g/l,更优选的高达60g/l,甚至更优选的高达80g/l,和最优选的高达100g/l的浓度,结合蛋白和/或结合结构域形成低于5%(w/w)的不溶性聚集体。不溶性聚集体的形成可以通过可见沉淀、凝胶过滤或动态光散射(其在不溶性聚集体形成时明显增加)的出现来检测。通过在10’000xg离心10分钟可以从蛋白样品中去除不溶性聚集体。优选的,在37℃的pbs中,在所述温育条件下结合蛋白和/或结合结构域形成低于2%,1%,0.5%,0.2%,0.1%,或0.05%(w/w)的不溶性聚集体。不溶性聚集体的百分比可以通过以下方式确定:分离不溶性聚集体和可溶性蛋白,然后通过标准定量法确定可溶性和不溶性组分中的蛋白量。
还优选的是37℃包含100mm二硫苏糖醇(dtt)的pbs中温育1或10小时,结合蛋白和/或结合结构域不丧失其天然三维结构。
在一个特定实施方案中,本发明涉及一种结合蛋白,其包括抑制vegf-axxx结合vegfr-2的结合结构域,并且具有指定的或优选的中点变性温度以及如上所述的非聚集特性,其中所述结合蛋白以低于100nm的ic50值抑制huvec球体的生芽。
术语“huvec”表示人脐静脉内皮细胞,其可以分离自正常人脐静脉,并对vegf-a刺激有反应。测量huvec球体生芽的测定法(诸如实施例2的描述)是本领域技术人员公知的。
ic50值是实验确定的参数(诸如huvec球体的生芽)的体外50%抑制所需的物质(诸如结合蛋白或结合结构域)的浓度。ic50值可以由本领域技术人员方便的确定(korfft.andaugustinh.g.,j.cellbiol.143(5),1341-52,1998)。
优选的是结合蛋白和/或结合结构域以低于10nm,优选的低于1nm,更优选的低于0.1nm,和最优选的低于0.05nm的ic50值抑制huvec球体的生芽。
进一步优选的是单体结合蛋白和/或结合结构域抑制huvec球体生芽的ic50值低于ranibizumab(lucentis®,genentech的注册商标),bevacizumab(avastin®,genentech的注册商标),aflibercept(vegftrap®,regeneron的注册商标),或pegaptanib(macugen®,pfizer的注册商标)的对应ic50值。
具体来说,本发明涉及一种结合蛋白,其包括抑制vegf-axxx结合vegfr-2的结合结构域,并具有指定的或优选的中点变性温度和如上所述的非聚集特性,其中与所述结合结构域和相应vegf-axxx的相互作用的kd相比,所述结合结构域和vegf-axxxb的相互作用的kd是至少10倍高。
优选的,与所述结合结构域和相应vegf-axxx的相互作用的kd相比,所述结合结构域和vegf-axxxb的相互作用的kd是至少102倍高,优选的103倍高,更优选的104倍高,更优选的105倍高,和最优选的106倍高。
还优选的,结合结构域与vegf-axxxb的相互作用的kd高于103nm,结合结构域与vegf-axxx的相互作用的kd低于10或1nm.。
优选的结合结构域与vegf-b,vegf-c,vegf-d,plgf或pdgf的相互作用的kd高于1nm,优选的高于10nm,更优选的高于102nm,甚至更优选的高于103nm,和最优选的高于104nm。
优选的,vegf-axxx是狗vegf-a164或猴vegf-a165或人vegf-a165,并且vegf-axxxb是狗vegf-a164b或猴vegf-a165b或人vegf-a165b。
另一个优选的实施方案是重组结合蛋白包括结合结构域,其中所述结合结构域抑制vegf-axxx结合vegfr-2,并且所述结合结构域是重复结构域或设计的重复结构域。这类重复结构域可以包括一个,两个,三个或更多个内部重复组件,它们将参与结合vegf-axxx。优选的,这类重复结构域包括n端加帽组件,两到四个内部重复组件,和c端加帽组件。优选的,所述结合结构域是锚蛋白重复结构域或设计的锚蛋白重复结构域。
优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有以下锚蛋白重复序列基序的重复组件
其中1,2,3,4,5,6,和7各自独立的代表选自a,d,e,f,h,i,k,l,m,n,q,r,s,t,v,w和y的氨基酸残基。
尤其优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有以下锚蛋白重复序列基序的重复组件
其中
1代表选自f,t,n,r,v,a,i,k,q,s和y的氨基酸残基;优选的选自f,t,n,r和v;更优选的选自f和t;
2代表选自w,y,h和f的氨基酸残基;优选的选自w,y和h;
3代表选自m,i,f和v的氨基酸残基;优选的选自m和i;
4代表选自h,a,k,g,l,m,n,t,v,w和y的氨基酸残基;优选的选自h,a和k;
5代表选自e,y,f,v,h,i,l,n和r的氨基酸残基;优选的选自e,y,f,v和h;更优选的选自e,y,f和v;和
6代表选自a,n,y,h和r的氨基酸残基。
进一步优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有以下锚蛋白重复序列基序的重复组件
其中
1代表选自t,e,a,d,f,k,n,q,r,s,w和y的氨基酸残基;优选的选自t和e;
2代表选自v,f,y,a,h,i,k,r,t和w的氨基酸残基;优选的选自v,f和y;
3代表选自s,a,n,f和m的氨基酸残基;优选的选自s,a和n;更优选的选自s和a;
4代表选自y,f,s和w的氨基酸残基;
5代表选自a,s,l和y的氨基酸残基;优选的选自a和s;
6代表选自d,n,m,a,i,k和y的氨基酸残基;优选的选自d,n和m;更优选的选自d和n;
7代表选自a,y,h,n和d的氨基酸残基;和
8代表氨基酸残基t或a。
进一步优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有以下锚蛋白重复序列基序的重复组件
其中
1代表选自k,t和y的氨基酸残基;
2代表氨基酸残基n或m;
3代表氨基酸残基t或f;
4代表氨基酸残基s或a;
5代表氨基酸残基h或r;
6代表选自a,y,h和n的氨基酸残基;和
7代表氨基酸残基a或t。
甚至更优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有seqidno:3的锚蛋白重复序列基序的重复组件,其中所述重复组件之前是具有seqidno:2的锚蛋白重复序列基序的重复组件和/或之后是具有seqidno:4的锚蛋白重复序列基序的重复组件。
进一步优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有以下锚蛋白重复序列基序的重复组件
其中
1代表选自a,n,r,v,y,e,h,i,k,l,q,s和t的氨基酸残基;优选的选自a,n,r,v和y;更优选的选自a和r;
2代表选自s,a,n,r,d,f,l,p,t和y的氨基酸残基;优选的选自s,a,n和r;
3代表选自t,v,s,a,l和f的氨基酸残基;优选的选自t,v,s,a和l;更优选的选自t,v和s;
4代表选自w,f和h的氨基酸残基;
5代表选自p,i,a,l,s,t,v和y的氨基酸残基;优选的选自p和i;
6代表选自w,f,i,l,t和v的氨基酸残基;
7代表氨基酸残基l或p;和
8代表选自a,h,n和y的氨基酸残基。
进一步优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有以下锚蛋白重复序列基序的重复组件
其中
1代表选自h,q,a,k,r,d,i,l,m,n,v和y的氨基酸残基;优选的选自h,q,a,k和r;更优选的选自a和r;
2代表选自y,f和h的氨基酸残基;
3代表选自q,f和t的氨基酸残基;
4代表选自w,m,g,h,n和t的氨基酸残基;优选的选自w和m;
5代表选自t,a,m,l和v的氨基酸残基;优选的选自t,a和m;
6代表选自i,l,v,d和t的氨基酸残基;优选的选自i,l和v;和
7代表选自a,h,n和y的氨基酸残基。
甚至更优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有seqidno:6的锚蛋白重复序列基序的重复组件,其中所述重复组件之前是具有seqidno:5的锚蛋白重复序列基序的重复组件。
进一步优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有以下锚蛋白重复序列基序的重复组件
其中
1代表选自k,m,n,r和v的氨基酸残基;
2代表选自y,h,m和v的氨基酸残基;
3代表选自f,l,m和v的氨基酸残基;
4代表选自r,h,v,a,k和n的氨基酸残基;优选的选自r,h,v和a;
5代表选自f,d,h,t,y,m和k的氨基酸残基;优选的选自f,d,h,t和y;和
6代表选自a,h,n和y的氨基酸残基。
进一步优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有以下锚蛋白重复序列基序的重复组件
其中
1代表选自t,a,h,i,n和s的氨基酸残基;
2代表选自f,i,q,r,v和n的氨基酸残基;
3代表选自a,g,n,q和v的氨基酸残基;
4代表氨基酸残基w或y;
5代表选自a,s,t和m的氨基酸残基;
6代表选自n,v,s,f,m和w的氨基酸残基;
7代表选自a,h,n和y的氨基酸残基;和
8代表氨基酸残基t或a。
进一步优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有以下锚蛋白重复序列基序的重复组件
其中
1代表选自k,a,v和n的氨基酸残基;
2代表选自n,i和y的氨基酸残基;
3代表选自t,f,y和w的氨基酸残基;
4代表选自w,d和y的氨基酸残基;
5代表氨基酸残基s或a;
6代表选自d,i和m的氨基酸残基;
7代表选自l,t和y的氨基酸残基;和
8代表选自a,h,y和n的氨基酸残基。
甚至更优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有seqidno:8的锚蛋白重复序列基序的重复组件,其中所述重复组件之前是具有seqidno:7的锚蛋白重复序列基序的重复组件和/或之后是具有seqidno:9的锚蛋白重复序列基序的重复组件。
进一步优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有以下锚蛋白重复序列基序的重复组件
其中
1代表选自l,s和t的氨基酸残基;
2代表选自g,s和c的氨基酸残基;优选的选自g和s;
3代表氨基酸残基s或a;
4代表选自q,s,m和n的氨基酸残基;优选的选自q和s;
5代表选自l,m和q的氨基酸残基;和
6代表选自a,h,n,y和d的氨基酸残基。
进一步优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有以下锚蛋白重复序列基序的重复组件
其中
1代表选自y,h,f,i,l和w的氨基酸残基;优选的选自y和h;
2代表选自m,d,i,l,v的氨基酸残基;优选的选自m和d;
3代表选自g,s和v的氨基酸残基;
4代表氨基酸残基w或f;
5代表选自a,g和t的氨基酸残基;
6代表氨基酸残基d或w;
7代表氨基酸残基l或f;和
8代表选自a,h,n和y的氨基酸残基。
甚至更优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有seqidno:11的锚蛋白重复序列基序的重复组件,其中所述重复组件之前是具有seqidno:10的锚蛋白重复序列基序的重复组件。
进一步优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有以下锚蛋白重复序列基序的重复组件
其中
1代表选自k,s,i,n,t和v的氨基酸残基;优选的选自k和s;
2代表选自k,n,w,a,h,m,q和s的氨基酸残基;优选的选自k和n;
3代表选自f,q,l,h和v的氨基酸残基;优选的选自f,q和l;
4代表氨基酸残基f或t;
5代表氨基酸残基q或h;
6代表氨基酸残基y或s;
7代表选自n,h,y和m的氨基酸残基;优选的选自n和h;和
8代表选自a,h,n和y的氨基酸残基。
进一步优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有以下锚蛋白重复序列基序的重复组件
其中
1代表选自f,y,h和w的氨基酸残基;优选的选自f,y和h;
2代表选自i,m,d和v的氨基酸残基;优选的选自i,m和d;
3代表氨基酸残基f或l;
4代表氨基酸残基l或p;
5代表选自h,l和y的氨基酸残基;和
6代表选自a,h,n,c和y的氨基酸残基。
甚至更优选的是一种重组结合蛋白,其中所述锚蛋白重复结构域或所述设计的锚蛋白重复结构域包括具有seqidno:13的锚蛋白重复序列基序的重复组件,其中所述重复组件之前是具有seqidno:12的锚蛋白重复序列基序的重复组件。
另一个优选的实施方案是一种重组结合蛋白,其包括具有对vegf-axxx的结合特异性的至少一个重复结构域,其中所述重复结构域与选自seqidnos:16,22,23,29,30和33的重复结构域,或选自seqidnos:16,22,23,29,30,33,34,36,39和40的重复结构域竞争结合vegf-axxx。
术语“竞争结合”表示本发明两个不同的结合结构域不能同时结合同一靶,而它们两者能够单独结合同一靶。因此,这两种结合结构域竞争结合所述靶。确定两种结合结构域是否竞争结合靶的方法(诸如竞争性elisa或竞争性spr测量(例如通过使用biorad的proteon装置))是本领域技术人员公知的。
与挑选的重复蛋白竞争结合vegf-axxx的重组结合蛋白可以通过本领域技术人员公知的方法鉴定,诸如竞争性酶联免疫吸附测定(elisa)。
另一个优选的实施方案是一种重组结合蛋白,其包括具有对vegf-axxx的结合特异性的重复结构域,所述vegf-axxx选自seqidnos:14到33,或选自seqidnos:14到40。
进一步优选的是一种重组结合蛋白,其中具有对vegf-axxx的结合特异性的所述重复结构域包括与所述重复结构域组的重复结构域具有至少70%氨基酸序列同一性的氨基酸序列。优选的,所述氨基酸序列同一性是至少75%,更优选的80%,更优选的85%,更优选的90%,和最优选的95%。
进一步优选的是一种重组结合蛋白,其中具有对vegf-axxx的结合特异性的所述重复结构域包括与所述重复结构域组的重复结构域的重复组件具有至少70%氨基酸序列同一性的重复组件。优选的,所述氨基酸序列同一性是至少75%,更优选的80%,更优选的85%,更优选的90%,和最优选的95%。
在本发明包括重复结构域的重组结合蛋白的进一步优选的实施方案中,所述重复结构域的重复组件的一个或更多个氨基酸残基被比对重复单位时在对应位置上发现的氨基酸残基替换。优选的,高达30%的氨基酸残基被替换,更优选的,高达20%,和甚至更优选的,高达10%的氨基酸残基被替换。最优选的,这类重复单位是天然存在的重复单位。甚至更优选的,所述重复结构域具有对vegf-axxx或vegfr-2的结合特异性。
在仍然另一个具体的实施方案中,高达30%的氨基酸残基,更优选的,高达20%,和甚至更优选的,高达10%的氨基酸残基被不是在重复单位的对应位置发现的氨基酸替换。
在进一步的实施方案中,此处描述的任一种vegf-axxx结合蛋白或结构域可以与一种或更多种其他的部分共价结合,包括,例如,也结合
vegfr-2的部分(例如vegfr-2结合多肽),结合不同的靶诸如plgf,人血清白蛋白,细胞受体(例如her2),免疫球蛋白(例如igg1),细胞因子(例如tnf-alpha或白介素)或生长因子以产生双特异性结合剂的部分,标记部分(例如荧光标记诸如荧光素,或放射性示踪剂),促进蛋白纯化的部分(例如小肽标签,诸如his-或strep-标签),提供改善的治疗效果的效应物功能的部分(例如提供抗体依赖性细胞介导的细胞毒性的抗体fc部分,毒性蛋白部分诸如绿脓杆菌(pseudomonasaeruginosa)外毒素a(eta)或小分子毒性剂诸如美登素类或dna烷化剂)或提供改善的药代动力学的部分。可以根据感知的治疗需要评价改善的药代动力学。经常希望增加生物利用度和/或增加给药之间的时间,可能通过增加给药后血清中仍然可用的蛋白的时间。在一些情况下,希望改善蛋白血清浓度随时间的连续性(例如,减少在刚刚施用后和下次施用前不久之间蛋白血清浓度的差异)。倾向于减慢蛋白从血液中清除的部分包括羟乙基淀粉(hes),聚乙二醇(peg),糖(例如唾液酸),良好耐受的蛋白部分(例如fc片段或血清白蛋白),以及对丰富的血清蛋白具有特异性和亲和力的结合结构域或肽,诸如抗体fc片段或血清白蛋白。本发明的重组结合蛋白可以连接到降低多肽在哺乳动物(例如小鼠,大鼠或人)中清除率的部分,相对于未修饰的多肽降低至少于1/3。
一个或更多个聚乙二醇部分可以连接到结合蛋白的不同位置,这类连接可以通过与胺、巯基或其他合适反应基的反应实现。聚乙二醇部分的连接(peg化)可以是定点的,其中将合适的反应基引入蛋白以产生peg化优选发生的位点,或合适的反应基原来就存在于结合蛋白中。硫醇基可以存在于半胱氨酸残基中;和胺基可以是,例如,在多肽n端发现的伯胺或存在于氨基酸侧链的胺基,诸如赖氨酸或精氨酸。在优选的实施方案中,结合蛋白被改造以便在所需位置具有半胱氨酸残基,允许半胱氨酸的定点peg化,例如通过与载有马来酰亚胺功能团的聚乙二醇衍生物反应。聚乙二醇部分的分子量可以变化较大(即从约1kda到约100kda),并且可以是分枝或线性的。优选的,聚乙二醇具有约1到约50kda的分子量,优选的约10到约40kda,甚至更优选的约15到约30kda,和最优选的约20kda。
在进一步的实施方案中,本发明涉及编码特定重组结合蛋白的核酸分子。此外,还涉及包括所述核酸分子的载体。
进一步的,涉及包括一种或更多种上述结合蛋白(尤其是包括重复结构域的重组结合蛋白)的药物组合物,或编码特定重组结合蛋白的核酸分子,以及任选的药学上可接受的载体和/或稀释剂。药学上可接受的载体和/或稀释剂是本领域技术人员已知的,将在下文更详细的解释。甚至进一步的,涉及包括一种或更多种上述重组结合蛋白(尤其是包括重复结构域的结合蛋白)的诊断组合物。
本发明的结合蛋白抑制或预防vegf诱导的病理性血管生成,血管渗漏(水肿),肺动脉高血压,肿瘤形成和/或炎性病症。利用“抑制”表明重组蛋白将所述疾病预防到某种程度,例如到10%或20%,更优选的50%,尤其是70%,80%或90%,或甚至95%。
术语“水肿”表示由血管渗漏引起的状况。炎症期间血管舒张和增加的通透性是主要致病机制。例如,水肿导致中风后的梗塞扩大,有可能在癌症患者中引起危急生命的颅内高压。此外,血浆蛋白的外渗有助于隐性肿瘤的转移性扩散,而气道充血可能造成致命的气喘发作。在炎症期间发生的增加的血管渗漏可能导致呼吸窘迫,腹水,腹膜硬化(在透析患者中),粘连形成(腹部手术)和转移性扩散。
术语“血管生成”表示形成新血管的基本过程。人的初始血管生成期发生在胚胎发育的前三个月,但血管生成还作为正常的生理过程在组织生长期间发生,诸如肌肉或脂肪的增加以及月经周期和怀孕期间。
术语“病理性血管生成”是指数种疾病状态的维持和进展期间血管的形成和生长。病理性血管生成的具体实例发现于血管(动脉粥样硬化,血管瘤,血管内皮瘤),骨和关节(类风湿性关节炎,滑膜炎,骨和软骨损伤,骨髓炎,血管翳生长,骨赘形成,肿瘤和转移瘤),皮肤(疣,脓性肉芽肿,毛发生长,卡波西氏肉瘤,疤痕瘢痕疙瘩,变应性水肿,肿瘤),肝,肾,肺,耳和其他上皮细胞(炎性和感染性过程包括肝炎,肾小球肾炎,肺炎;和哮喘,鼻息肉,耳炎,移植病症,肝再生病症,肿瘤和转移瘤),子宫,卵巢和胎盘(子宫内避孕器造成的功能障碍性子宫出血,卵泡囊肿形成,卵巢超刺激综合征,子宫内膜异位,肿瘤),脑、神经和眼睛(早产儿的视网膜病,糖尿病性视网膜病,脉络膜和其他眼内病症,白质软化,肿瘤和转移瘤),工作超负荷造成的心脏和骨骼肌,脂肪组织(肥胖症),内分泌器官(甲状腺炎,甲状腺肿大,胰移植病症),造血作用(aids中的kaposi综合征),血液恶性肿瘤(白血病),和淋巴管(肿瘤转移,淋巴组织增生病症)。
术语“视网膜局部缺血病”表示视网膜的血液和氧供给减少,视网膜的外周部分失去它们的营养来源并终止正常功能。视网膜局部缺血病的具体实例是视网膜病。引起视网膜病的常见疾病是糖尿病性视网膜病,视网膜中央静脉阻塞,颈动脉狭窄,和镰状细胞视网膜病。糖尿病性视网膜病是糖尿病患者视觉丧失的主要原因。在局部缺血性视网膜中,存在新血管的生长(新生血管形成)。这些血管经常生长在视网膜的表面,视神经中,眼睛前部的虹膜上。新血管无法代替必需营养物的流动,相反,可能造成许多问题诸如玻璃体出血,视网膜剥离和不受控制的青光眼。存在这些问题是因为新血管是脆弱和易于出血的如果在其早期发现,利用全视网膜光凝术有时可以阻滞增生性糖尿病性视网膜病。但是,一些情况下,玻璃体切割(vitrectomy)手术是唯一的选择。
除了这些视网膜病外,眼睛的血管病还包括眼新生血管形成病,诸如黄斑变性和糖尿病黄斑水肿(dme)。黄斑变性源于斑点下脉络膜血管的新生血管生长。存在两种类型的黄斑变性∶干性和湿性。尽管湿性黄斑变性只包括全部黄斑变性的15%,但几乎所有的湿性黄斑变性都致盲。此外,湿性黄斑变性几乎总是源于干性黄斑变性。一旦一只眼染上湿性黄斑变性,该病症通常感染另一只眼。湿性黄斑变性经常被称为湿性-amd的年龄相关性湿性黄斑变性,因为它主要发现在老人中。
糖尿病性视网膜病(dr)和dme是大部分发达国家的劳动年龄人口失明的主要原因。世界范围内糖尿病患者数量的增加表明dr和dme仍将是未来数年内视力丧失以及相关功能性损伤的主要原因。数种生化机制(包括蛋白激酶c-β活化,增加的血管内皮生长因子产生,氧化应激,和胞内山梨糖醇和高度糖基化最终产物的积聚)可能导致表现为dr/dme的血管破坏。这些途径的抑制有希望干预dr和dme。
术语“肺部高压”表示肺部动脉的血压异常高的病症。在没有心脏或肺的其他疾病情况下,它被称为原发性肺动脉高压。肺部小动脉的弥散性狭窄是作为病理性动脉生成的结果发生的,然后是作为对血流阻碍增加的应答的肺动脉高压。发病率是100'000人中8个。但是,肺动脉高压还可以作为慢性阻塞性肺病(copd)的并发症(诸如肺气肿,慢性支气管炎或弥漫性间质纤维化)以及在哮喘样copd患者中发生。copd的发病率是10'000人中大约5个。
此外,本发明的结合蛋白可用于治疗炎症和更具体的来说炎性病症。
本文使用的术语“炎症”表示,对活组织损伤的局部反应,尤其是小血管,它们的内容物以及它们的相关结构的局部反应。血液组分穿过血管壁进入组织是炎症的标志,如此形成的组织收集物被称为渗出液或水肿。破坏活组织的任何有害步骤,例如感染细菌,过热,冷,机械性损伤诸如挤压,酸,碱金属,辐射,或感染病毒都可以引起炎症,与涉及的器官或组织无关。应清楚,分类到“炎性疾病”的疾病以及范围从烧伤到肺炎,麻疯病,肺结核,和类风湿性关节炎的组织反应都是“炎症”。
本发明的结合蛋白可用于治疗肿瘤形成。术语“肿瘤”表示异常组织团,其无明显原因的来自先前存在的体细胞,没有预期功能,特征在于自发和无限生长的倾向。肿瘤与炎性或其他肿胀完全不同,因为肿瘤内的细胞在其形态和其他特征方面是异常的。异常细胞,即通常组成肿瘤的细胞种类,与正常细胞的不同之处在于经历下列改变中的一种或更多种:(1)肥大,或单个细胞大小的增加;(2)增生或指定区域内细胞数目的增加;(3)退行性变化,或细胞生理特征向更原始或未分化类型的退化。肿瘤可以是良性的,例如脂肪瘤,血管瘤,骨瘤,软骨瘤,和腺瘤。恶性肿瘤的实例是癌(诸如乳腺瘤,呼吸道和胃肠道、内分泌腺和生殖泌尿系统的癌),肉瘤(在结缔组织中,包括纤维组织,脂肪(脂)组织,肌肉,血管,骨,和软骨),癌肉瘤(在上皮和结缔组织两者中),白血病和淋巴瘤,神经组织(包括脑)瘤,和黑色素瘤(着色皮肤细胞的癌症)。本发明结合蛋白抗肿瘤的使用还可以结合本领域已知的任何其他肿瘤疗法诸如放疗,光动力疗法,化疗或手术。
药物组合物包括如上所述的结合蛋白和药学上可接受的载体,赋形剂或稳定剂(remington'spharmaceuticalsciences16thedition,osol,a.ed.[1980])。技术人员已知的合适载体,赋形剂或稳定剂是盐水,ringer's溶液,葡萄糖溶液,hank's溶液,非挥发油,油酸乙酯,5%葡萄糖盐溶液,增强等渗性和化学稳定性的物质,缓冲剂和防腐剂。其他合适的载体包括自身不诱导产生对接受组合物的个体有害的抗体的任何载体,诸如蛋白,多糖,聚乳酸,聚乙醇酸,聚氨基酸和氨基酸共聚物。药物组合物还可以是组合制剂,包括其他的活性剂,诸如抗癌剂或抗血管生成剂(例如人vegf-axxxb;优选的,人vegf-a165b)。
优选的用于治疗眼病的药物组合物包括如上所述的结合蛋白和去污剂诸如聚山梨酯20(例如约0.04%),缓冲剂诸如组氨酸,磷酸盐或乳酸和糖诸如蔗糖或海藻糖。优选的,这类组合物包括如上所述的结合蛋白和pbs。所述药物组合物可以局部施用,或局部施用到眼睛的一部分或注射到眼中,例如注射入结膜下,眼球周围或眼球后间隙或直接注射入眼。可选的,可以通过肠胃外施用将所述组合物全身施用。优选的,通过玻璃体内注射将所述药物组合物应用于眼。还优选的,所述药物组合物被局部应用于眼,并作为滴眼剂。滴眼剂可用于角膜(眼中心的澄清部分),从而允许分子渗入眼中。为了治疗感染眼睛后部的疾病,最希望当在结膜下或眼球周围注射时结合蛋白能够穿透巩膜。结合蛋白的施用可以在调节眼睛表面以提高分子穿透的预备步骤之后进行。优选的,通过穿透增强子调节上皮层诸如角膜上皮以允许例如上文描述的分子的足量和快速穿透。本发明结合蛋白抗眼病的使用还可以结合本领域已知的任何其他疗法诸如光动力疗法。
用于体内施用的制剂必须是无菌或消毒的。这可以通过过滤除菌膜进行过滤方便地实现。
可以制备缓释制剂。在本发明的一个实施方案中,可以使用眼内植入物来提供本发明的结合蛋白。缓释制剂的合适实例包括固态疏水多聚体(包含本发明多肽)的半渗透基质,所述基质采用成形产品的形式,例如膜或微胶囊。缓释基质的实例包括聚酯,水凝胶(例如,聚(2-羟乙基-异丁烯酸酯),或聚(乙烯醇)),聚丙交酯,l-谷氨酸和gamma-乙基-l-谷氨酸酯的共聚物,不可降解的亚乙基-醋酸乙烯酯,可降解的乳酸-羟基乙酸共聚物诸如luprondepot®(由乳酸-羟基乙酸共聚物和醋酸亮丙瑞林组成的注射用微球),和聚-d-(-)-3-羟丁酸。
可以使用技术人员已知的任何合适方法施用药学组合物。优选的施用途径是肠胃外。在肠胃外施用中,本发明的药物将与上文所述的药学上可接受的赋形剂一起被配制为单位剂量的注射形式,诸如溶液,悬液或乳液。施用的剂量和方式将取决于待治疗的个体和具体疾病。通常,施用药物组合物使得给予本发明结合蛋白的剂量为1μg/kg到20mg/kg,更优选的10μg/kg到5mg/kg,最优选的0.1到2mg/kg。优选的,其作为快速浓注剂量给药。还可以使用连续输注,包括通过渗透性微型泵的连续皮下输送。这样的话,药物组合物的输注剂量为5到20μg/kg/分钟,更优选的7到15μg/kg/分钟。尤其是,通过注射入眼睛来施用药物组合物,使得给予本发明结合蛋白的剂量为0.1mg到10mg每次注射,更优选的0.3到6mg每次注射,最优选的1mg到4mg每次注射。此外,通过滴眼剂给眼睛施用药物组合物,使得施用于眼的单滴溶液包含本发明结合蛋白的浓度为10到120mg/ml,更优选的20到100mg/ml,最优选的40到80mg/ml。
在本发明的另一个实施方案中,如上所述抑制vegf-axxx活性的结合蛋白,可以与抑制plgf活性的结合蛋白或小分子(具有与上述针对vegf-axxx的相同plgf抑制水平)联用。该实施方案基于以下事实:在vegf-axxx水平增加的位置发现plgf是生成血管的。此外,如上所述抑制vegf-axxx活性的结合蛋白,可以与抑制以下物质活性的结合蛋白或小分子联用:血小板衍生生长因子(pdgf),vegf-c或蛋白vegf家族的其他成员,肿瘤坏死因子alpha(tnfalpha),delta-配体样4(dii4),白介素6(il-6),神经毡蛋白或促血管生成素2(ang2)。
本发明还提供新的治疗方法。一方面,提供治疗视网膜病的方法,所述方法包括给需要其的患者施用治疗有效量的本发明结合蛋白,尤其是抑制人vegf-axxx和人vegfr-2相互作用、但不抑制vegf-axxxb和人vegfr-2相互作用的结合蛋白,并且所述结合蛋白抑制vegfr-2介导的血管生成。
本发明还涉及使用所述结合蛋白抑制细胞中vegf-a生物活性或抑制vegfr-2介导的生物活性的方法。细胞可以位于体内或离体,并且可以是,例如,活生物体的细胞,培养的细胞或组织样品中的细胞。该方法包括将所述细胞与本文公开的vegf-a/vegfr-2相互作用抑制结合蛋白中的任一种按照足以抑制这类生物活性的量和时间进行接触。
本发明提供治疗患有对vegf-axxx或vegfr-2的抑制产生应答的状况的受试者的方法。这类方法包括给所述受试者施用有效量的本文所述结合蛋白。状况可以是特征为不当血管生成的状况。状况可以是高增殖(hyperproliferative)状况。适于治疗的状况(或病症)的实例包括自身免疫病,炎性病症,视网膜病(尤其是增殖性视网膜病),和癌症,尤其是上述疾病的一种。本文描述的任一种结合蛋白可用于制备治疗下述病症的药物,尤其是选自以下组的病症∶自身免疫病,炎性病症,视网膜病,和癌症。适于治疗的优选状况(或病症)是第一线转移性肾细胞癌,复发的多形性胶质母细胞瘤,辅助(adjuvant)结肠癌,辅助her2-阴性乳腺癌,辅助her2-阳性乳腺癌,辅助非小细胞肺癌,弥散性大b细胞淋巴瘤,第一线晚期胃癌,第一线her2-阴性转移性乳腺癌,第一线her2-阳性转移性乳腺癌,第一线转移性卵巢癌症,胃肠间质肿瘤,高风险类癌瘤,激素难治性前列腺癌,新诊断的多形性胶质母细胞瘤,转移性头和颈部癌症,复发的铂敏感性卵巢癌,第二线转移性乳腺癌,泛发性小细胞肺癌,非鳞状、非小细胞肺癌以及先前治疗的cns转移瘤和复发的多发性骨髓瘤,前列腺癌,非小细胞肺癌(nsclc),结肠直肠癌和胰腺癌,晚期卵巢癌(aoc),具有征兆性恶性腹水的aoc患者和非霍奇金氏淋巴瘤。
本发明的重组结合蛋白可以通过以下数种方法获得和/或进一步发展,诸如噬菌体表面展示(wo90/02809,wo07/006665)或细菌细胞表面展示(wo93/10214),核糖体展示(wo98/48008),质粒展示(wo93/08278)或使用共价的rna-重复蛋白杂合构建体(wo00/32823),或胞内表达和挑选/筛选诸如通过蛋白互补测定(wo98/341120)。这类方法是本领域技术人员已知的。
用于本发明重组结合蛋白的挑选/筛选的锚蛋白重复蛋白库可以根据本领域技术人员已知的方案(wo02/020565,binz,h.k.etal.,jmb,332,489-503,2003,和binzetal.,2004,在上述引文中)获得。用于vegf-axxx特异性darpins挑选的这类文库的使用在实施例1中给出。类似的,上文描述的锚蛋白重复序列基序可以用于构建锚蛋白重复蛋白的库(其可用于vegf-axxx特异性darpins的挑选或筛选)。此外,利用标准重组dna技术(例如wo02/020565,binzetal.,2003,在上述引文中,和binzetal.,2004,在上述引文中),本发明的重复结构域可以从本发明的重复组件和合适的加帽组件模块化组装(forrer,p.,etal.,febsletters539,2-6,2003)。
本发明不局限于在实施例描述的具体实施方案。可以按照如下所述的一般概述使用和处置其他来源。
实施例
下文公开的所有起始原料和试剂都是本领域技术人员已知的,可以商购获得或利用公知的技术制备。
材料
化学制剂购自fluka(switzerland)。寡核苷酸购自microsynth(switzerland)。除非另有说明,dna聚合酶、限制性内切酶和缓冲液购自newenglandbiolabs(usa)或fermentas(lithuania)。克隆菌株和蛋白生成菌株是大肠杆菌xl1-blue(stratagene,usa)。vegf变体购自r&dsystems(minneapolis,usa),或在中国仓鼠卵巢细胞或巴斯德毕赤酵母中产生并根据标准步骤进行纯化(rennel,e.s.etal.,europeanj.cancer44,1883-94,2008;2008;pichia表达系统购自invitrogen)。生物素化的vegf变体通过化学法获得,利用标准的生物素化试剂和方法将生物素部分偶联到纯化的vegf变体的伯胺(pierce,usa)。
分子生物学
除非另有说明,根据描述的方案(sambrookj.,fritsche.f.andmaniatist.,molecularcloning:alaboratorymanual,coldspringharborlaboratory1989,newyork)实施方法。
设计的锚蛋白重复蛋白文库
n2c和n3c设计的锚蛋白重复蛋白文库被描述(wo02/20565;binzetal.2003,在上述引文中;binzetal.2004,在上述引文中)。n2c和n3c中的数字描述加帽组件n端和c端之间存在的随机重复组件的数目。用于定义重复单位和组件内位置的命名法是基于binzetal.2004,在上述引文中,改变是重复组件和重复单位的边界迁移一个氨基酸位置。例如,binzetal.2004(在上述引文中)的重复组件的位置1对应于本文重复组件的位置2,所以binzetal.2004(在上述引文中)的重复组件的位置33对应于本文下一个重复组件的位置1。
所有dna序列都通过测序确认,所有描述蛋白的计算分子量都通过质谱确认。
实施例1∶具有对vegf-axxx的结合特异性的包括重复结构域的结合蛋白的挑选
利用核糖体展示(hanes,j.andplückthun,a.,pnas94,4937-42,1997),许多设计的具有对vegf-axxx的结合特异性的锚蛋白重复蛋白(darpins)选自binz等2004(在上述引文中)描述的n2c或n3cdarpin文库。通过粗提物elisa评价挑选的克隆对特异性(vegf-axxx)和非特异性(mbp,大肠杆菌麦芽糖结合蛋白)靶标的结合,表明vegf-axxx结合蛋白被成功挑选(图1)。seqidno:14到40构成所选结合蛋白的氨基酸序列,所述结合蛋白包括具有对vegf-axxx的结合特异性的重复结构域。所选结合剂的序列分析显示某些挑选的结合剂家族固有的特定锚蛋白重复序列基序。具有对vegf-axxx的结合特异性的重复结构域中存在的这类锚蛋白重复序列基序提供于seqidno:1到13。
通过核糖体展示挑选vegf-axxx特异性锚蛋白重复蛋白
通过核糖体展示(hanesandplückthun,在上述引文中)进行vegf-axxx特异性锚蛋白重复蛋白的挑选,使用狗vegf-a164或人vegf-a165作为靶蛋白,如所述的设计的锚蛋白重复蛋白文库(wo02/020565,binzetal.,2003,在上述引文中和binzetal.,2004,在上述引文中)和确立的方案(zahnd,c.,amstutz,p.andplückthun,a.,nat.methods4,69-79,2007)。使用确立的方案(binzetal.2004,在上述引文中),利用n2c和n3cdarpin文库对狗或人vegf变体(包括生物素化的变体,固定于neutravidin或链霉亲和素)进行核糖体展示挑选循环。每轮挑选后逆转录(rt)-pcr循环的数目恒定的从40减少到30,以适应结合剂富集得到的产出。对狗vegf的4轮初步挑选得到纳摩亲和力darpins的混合物,这通过单个克隆的elisa和spr测量显示。为了发现具有进一步改善的亲和力的darpins,对固定于neutravidin或链霉亲和素的生物素化人或狗vegf进行额外的解离速率筛选(off-rateselections),采用第二轮和第三轮初始核糖体展示挑选后的混合物,然后是对人vegf的结合速率筛选(on-rateselection)。
如粗提物elisa所示,挑选的克隆特异性结合vegf-axxx
使用标准步骤,利用darpin表达细胞的粗制大肠杆菌提取物,通过酶联免疫吸附测定(elisa)鉴定特异性结合vegf-axxx的所选单个darpins。所选克隆被克隆入pqe30(qiagen)表达载体,转化大肠杆菌xl1-blue(stratagene),然后在包含1ml生长培养基(2yt包含1%葡萄糖和100µg/ml氨苄青霉素)的96深孔板(单孔中每个克隆)中37℃生长过夜。将100µl过夜培养物接种到新的96深孔板中的1ml新鲜2yt(包含50µg/ml氨苄青霉素)。在37℃温育2h后,利用iptg(1mm终浓度)诱导表达并持续3h。收集细胞,重悬于100µlb-perii(pierce),并在室温振荡条件下温育15分钟。然后,添加900µlpbs-tb(pbs添加0.2%bsa,0.1%tween20,ph7.4),通过离心去除细胞碎片。将100µl每种裂解的克隆加入neutravidin包被的maxisorp板的孔(包含通过它们的生物素部分固定的vegf-axxx变体或无关mbp),并在室温温育1h。用pbs-t(pbs添加0.1%tween20,ph7.4)彻底清洗后,使用单克隆抗rgs(his)4抗体(34650,qiagen)作为一抗和缀合碱性磷酸酶的多克隆山羊抗小鼠抗体(a3562,sigma)作为第二试剂通过标准elisa法将板显色。然后使用4-硝基苯基磷酸二钠(4npp,fluka)作为碱性磷酸酶的底物检测结合。在405nm测量显色。用于鉴定结合vegf-axxx的darpins的示例性粗提物elisa的结果显示于图1。通过这种粗制细胞提取物elisa筛选数百个克隆显示,超过上百个不同的darpins具有对vegf-axxx的特异性。挑选这些结合蛋白用于进一步分析。挑选的特异性结合vegf-axxx的锚蛋白重复结构域的氨基酸序列的实例提供于seqidno:14到40。
从具有对vegf-axxx的结合特异性的所选重复结构域推定重复序列基序
通过本领域技术人员已知的序列分析工具进一步分析所选重复结构域(具有对vegf-axxx的结合特异性)的氨基酸序列(wo02/020565;forreretal.,2003,在上述引文中;forrer,p.,binz,h.k.,stumpp,m.t.andplückthun,a.,chembiochem,5(2),183-189,2004)。但是,与wo02/020565(其中使用天然存在的重复基序推导重复序列基序)不同,本文从挑选的具有对vegf-axxx的结合特异性的重复结构域的重复单位推导出重复序列基序。从而确定包括常见重复序列基序的所选重复结构域的家族。具有对vegf-axxx的结合特异性的重复结构域中存在的这类重复序列基序提供于seqidno:1到13。
darpins的高水平和可溶性表达
为了进一步分析,将在所述粗制细胞提取物elisa中显示特异性vegf-axxx结合的挑选克隆在大肠杆菌xl1-blue细胞中表达,并通过标准步骤使用它们的his标签进行纯化。使用25ml静止过夜培养物(lb,1%葡萄糖,100mg/l氨苄青霉素;37℃)接种1升培养物(相同培养基)。在a(600)=0.7时,用0.5mmiptg诱导培养物,并在37℃温育4h。将培养物离心,所获沉淀重悬于40mltbs500(50mmtris–hcl,500mmnacl,ph8)并进行超声处理。将裂解物再次离心,向所获上清中添加甘油(10%(v/v)终浓度)和咪唑(20mm终浓度)。根据制造商的说明书(qiagen,germany),在ni-氮川三乙酸柱(2.5ml柱体积)上纯化蛋白。根据sds-15%page估算,从1升大肠杆菌培养物可以纯化出高达200mg的高可溶性darpins(具有对vegf-axxx的结合特异性),其纯度>95%。这种纯化的darpins用于进一步表征。
实施例2∶在球体生长测定中确定挑选的具有对vegf-axxx的结合特异性的darpins的ic50值
向埋入胶原基质的huvec球体添加vegf-axxx导致球体生芽。添加vegf-axxx的抑制剂将阻断芽形成,这可以通过芽的数目和长度在统计上定量。通过添加不同浓度的抑制剂和恒定量的vegf,可以确定ic50。
vegf-axxx特异性darpins对球体生芽的抑制
根据标准步骤(korffetal.,在上述引文中)完成球体生长测定。挑选具有对vegf-axxx的特异性的darpins,并按实施例1所述纯化到>96%纯度。人脐静脉细胞在单层培养中生长到汇合。胰酶消化后,将细胞悬液置于悬滴条件以形成球体,即大约500个有组织聚集的huvecs。将球体埋入胶原基质,并用vegf-a165刺激以起始芽生长。另外添加生芽抑制剂以观察它们对生芽抑制的作用。使用图形软件定量每个球体的芽数目和芽长度。
来自2个示例性球体生芽测定的结果显示于图2a(具有对vegf-axxx的结合特异性的darpin#30)和图2b(darpinnc,没有对vegf-axxx的结合特异性的阴性对照darpin;例如darpine3_5(binzetal.,2005,在上述引文中))。在该测定各种表现最好的darpins显示范围为10到50pm的ic50值,而平行实验中avastin®,lucentis®和macugen®显示范围为150到500pm的ic50值。
实施例3∶通过表面等离子体共振分析确定darpin#27与avastin®相比的靶特异性
将狗vegf-a164或狗vegf-a164b固定于流动池,并分析darpin#27(seqidno:16)和avastin®与固定靶的相互作用。
表面等离子体共振(spr)分析
使用proteon装置(biorad)测量spr。运行缓冲液是20mmhepes,ph7.4,150mmnacl和0.005%tween20。将约1200ru的狗vegf-a164或狗vegf-a164b固定于glc芯片(biorad)。测量相互作用,60μl/min流速的5分钟缓冲液流动,浓度为250nm的avastin®或darpin#27的100秒注射,以及具有缓冲液流动的数分钟解离速率测量。从测量值减去未包被参照池的信号。
结果显示于图3a(avastin与狗vegf-a164的相互作用),图3b(avastin与狗vegf-a164b的相互作用),图3c(darpin#27与狗vegf-a164的相互作用)和图3d(darpin#27与狗vegf-a164b的相互作用)。avastin与两种固定的vegf同种型均明显相互作用,而darpin#27只显示与vegf-a164而非vegf-a164b的相互作用。
实施例4∶血管渗漏兔模型中darpin#30抑制vegf-a165的体内功效
通过玻璃体内注射将聚乙二醇化的darpin#30(seqidno:29)或lucentis®施用于兔的眼睛以检测它们抑制血管渗漏(通过人vegf-a165的后续玻璃体内的注射诱导)的功效。
兔中血管渗漏抑制的测量
在第一天,通过玻璃体内注射将pbs、聚乙二醇化的darpin#30(125μg)或等摩尔量的lucentis®(162μg)施用于每只兔的一只眼(治疗眼)。在第4天或第30天,通过玻璃体内注射500ng人vegf-a165攻击每只兔的治疗眼。在vegf-a165注射后48小时评价所有动物的两只眼,通过静脉注射钠荧光素(50mg/kg动物体重,10%(w/v)溶于0.9%(w/v)盐溶液)后1h测量所有眼睛中的荧光素含量。计算每只动物的治疗眼与未治疗眼的荧光量的比值。比值为1相当于治疗眼中不存在额外的荧光渗漏,比值大于1表示治疗眼比未治疗对照眼更多的荧光渗漏。
聚乙二醇化darpin的制备
利用单个cys残基和马来酰亚胺化学法将蛋白peg化是本领域技术人员公知的,可以根据确立的方案(例如来自pierce)进行。使用标准层析法将包括附加c端接头(gggsgggsc,seqidno:41)的darpin#30纯化到接近均质。使用dtt将蛋白完全还原,并通过凝胶过滤进行纯化以去除dtt,将缓冲液更换为pbs。将溶于pbs的peg-马来酰亚胺(甲氧基-聚(乙二醇)-氧代丙胺基-丙基马来酰亚胺;nof,no.sunbrightme-200ma)与溶于pbs的darpin在室温下混合2-4小时,其中peg-马来酰亚胺大约15%摩尔过量。然后使用标准阴离子交换层析,将聚乙二醇化的darpin与未反应的darpin和未反应的peg部分分离。
结果显示于图4。在通过玻璃体内注射施用聚乙二醇化的darpin#30和lucentis®后4天,它们能够保护兔眼免于vegf-a165诱导的血管渗漏。尽管如此,只有聚乙二醇化的darpin#30,而不是lucentis®,在直至玻璃体内注射后30天仍能够保护兔眼免于vegf-a165诱导的血管渗漏。
序列表
<110>molecularpartnersag
binz,hanskaspar
forrer,patrik
stumpp,michaeltobias
<120>抑制vegf-a受体相互作用的结合蛋白
<130>p368a
<150>ep08168166.0
<151>2008-11-03
<160>41
<170>patentinversion3.5
<210>1
<211>33
<212>prt
<213>人工的
<220>
<223>锚蛋白重复序列基序
<220>
<221>misc_feature
<222>(1)..(1)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(3)..(4)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(14)..(15)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(27)..(27)
<223>xaa可以是任何天然存在的氨基酸
<400>1
xaaaspxaaxaaglyxaathrproleuhisleualaalaxaaxaagly
151015
hisleugluilevalgluvalleuleulysxaaglyalaaspvalasn
202530
ala
<210>2
<211>33
<212>prt
<213>人工的
<220>
<223>锚蛋白重复序列基序
<220>
<221>misc_feature
<222>(1)..(1)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(3)..(4)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(14)..(15)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(27)..(27)
<223>xaa可以是任何天然存在的氨基酸
<400>2
xaaaspxaaxaaglytrpthrproleuhisleualaalaxaaxaagly
151015
hisleugluilevalgluvalleuleulysxaaglyalaaspvalasn
202530
ala
<210>3
<211>33
<212>prt
<213>人工的
<220>
<223>锚蛋白重复序列基序
<220>
<221>misc_feature
<222>(1)..(1)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(3)..(4)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(14)..(15)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(27)..(27)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(33)..(33)
<223>xaa可以是任何天然存在的氨基酸
<400>3
xaaaspxaaxaaglyxaathrproleuhisleualaalaxaaxaagly
151015
hisleugluilevalgluvalleuleulysxaaglyalaaspvalasn
202530
xaa
<210>4
<211>33
<212>prt
<213>人工的
<220>
<223>锚蛋白重复序列基序
<220>
<221>misc_feature
<222>(1)..(1)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(3)..(4)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(12)..(12)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(17)..(17)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(27)..(27)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(33)..(33)
<223>xaa可以是任何天然存在的氨基酸
<400>4
xaaaspxaaxaaglytrpthrproleuhisleuxaaalaaspleugly
151015
xaaleugluilevalgluvalleuleulysxaaglyalaaspvalasn
202530
xaa
<210>5
<211>33
<212>prt
<213>人工的
<220>
<223>锚蛋白重复序列基序
<220>
<221>misc_feature
<222>(1)..(1)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(3)..(4)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(14)..(15)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(18)..(18)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(27)..(27)
<223>xaa可以是任何天然存在的氨基酸
<400>5
xaaaspxaaxaaglyxaathrproleuhisleualaalaxaaxaagly
151015
hisxaagluilevalgluvalleuleulysxaaglyalaaspvalasn
202530
ala
<210>6
<211>33
<212>prt
<213>人工的
<220>
<223>锚蛋白重复序列基序
<220>
<221>misc_feature
<222>(1)..(1)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(3)..(4)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(14)..(15)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(27)..(27)
<223>xaa可以是任何天然存在的氨基酸
<400>6
xaaaspxaaxaaglyxaathrproleuhisleualaalaxaaxaagly
151015
hisleugluilevalgluvalleuleulysxaaglyalaaspvalasn
202530
ala
<210>7
<211>33
<212>prt
<213>人工的
<220>
<223>锚蛋白重复序列基序
<220>
<221>misc_feature
<222>(1)..(1)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(3)..(4)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(14)..(15)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(27)..(27)
<223>xaa可以是任何天然存在的氨基酸
<400>7
xaaaspxaaxaaglytrpthrproleuhisleualaalaxaaxaagly
151015
hisleugluilevalgluvalleuleulysxaaglyalaaspvalasn
202530
ala
<210>8
<211>33
<212>prt
<213>人工的
<220>
<223>锚蛋白重复序列基序
<220>
<221>misc_feature
<222>(1)..(1)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(3)..(4)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(14)..(15)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(27)..(27)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(33)..(33)
<223>xaa可以是任何天然存在的氨基酸
<400>8
xaaaspxaaxaaglyxaathrproleuhisleualaalaxaaxaagly
151015
hisleugluilevalgluvalleuleulysxaaglyalaaspvalasn
202530
xaa
<210>9
<211>33
<212>prt
<213>人工的
<220>
<223>锚蛋白重复序列基序
<220>
<221>misc_feature
<222>(1)..(1)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(3)..(4)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(12)..(12)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(14)..(15)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(27)..(27)
<223>xaa可以是任何天然存在的氨基酸
<400>9
xaaaspxaaxaaglyxaathrproleuhisleuxaaalaxaaxaagly
151015
hisleugluilevalgluvalleuleulysxaaglyalaaspvalasn
202530
ala
<210>10
<211>33
<212>prt
<213>人工的
<220>
<223>锚蛋白重复序列基序
<220>
<221>misc_feature
<222>(1)..(1)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(5)..(5)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(14)..(15)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(18)..(18)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(27)..(27)
<223>xaa可以是任何天然存在的氨基酸
<400>10
xaaaspphelysxaaaspthrproleuhisleualaalaxaaxaagly
151015
hisxaagluilevalgluvalleuleulysxaaglyalaaspvalasn
202530
ala
<210>11
<211>33
<212>prt
<213>人工的
<220>
<223>锚蛋白重复序列基序
<220>
<221>misc_feature
<222>(1)..(1)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(3)..(3)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(5)..(6)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(13)..(15)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(27)..(27)
<223>xaa可以是任何天然存在的氨基酸
<400>11
xaaaspxaaleuxaaxaathrproleuhisleualaxaaxaaxaagly
151015
hisleugluilevalgluvalleuleulysxaaglyalaaspvalasn
202530
ala
<210>12
<211>33
<212>prt
<213>人工的
<220>
<223>锚蛋白重复序列基序
<220>
<221>misc_feature
<222>(1)..(1)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(3)..(4)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(6)..(6)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(10)..(10)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(14)..(15)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(27)..(27)
<223>xaa可以是任何天然存在的氨基酸
<400>12
xaaaspxaaxaaglyxaathrproleuxaaleualaalaxaaxaagly
151015
hisleugluilevalgluvalleuleulysxaaglyalaaspvalasn
202530
ala
<210>13
<211>33
<212>prt
<213>人工的
<220>
<223>锚蛋白重复序列基序
<220>
<221>misc_feature
<222>(1)..(1)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(3)..(4)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(8)..(8)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(17)..(17)
<223>xaa可以是任何天然存在的氨基酸
<220>
<221>misc_feature
<222>(27)..(27)
<223>xaa可以是任何天然存在的氨基酸
<400>13
xaaaspxaaxaaglytrpthrxaaleuhisleualaalaaspleugly
151015
xaaleugluilevalgluvalleuleulysxaaglyalaaspvalasn
202530
ala
<210>14
<211>159
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>14
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
pheasptrpmetglytrpthrproleuhisleualaalahisglugly
354045
hisleugluilevalgluvalleuleulysasnglyalaaspvalasn
505560
alathraspvalserglytyrthrproleuhisleualaalaalaasp
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnthrlysaspasnthrglytrpthrproleuhisleuseralaasp
100105110
leuglyhisleugluilevalgluvalleuleulystyrglyalaasp
115120125
valasnalaglnasplyspheglylysthralapheaspileserile
130135140
aspasnglyasngluaspleualagluileleuglnlysalaala
145150155
<210>15
<211>159
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>15
glyseraspleuglylyslysleuleuglualaalaargvalglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
pheasptrpmetglytrpthrproleuhisleualaalahisglugly
354045
hisleugluilevalgluvalleuleulysasnglyalaaspvalasn
505560
alathraspvalserglytyrthrproleuhisleualaalaalaasp
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnthrlysaspasnthrglytrpthrproleuhisleuseralaasp
100105110
leuglyhisleugluilevalgluvalleuleulystyrglyalaasp
115120125
valasnalaglnasplyspheglylysthralapheaspileserile
130135140
aspasnglyasngluaspleualagluileleuglnlysalaala
145150155
<210>16
<211>159
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>16
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
pheasptrpmetglytrpthrproleuhisleualaalahisglugly
354045
hisleugluilevalgluvalleuleulysasnglyalaaspvalasn
505560
alathraspvalserglytyrthrproleuhisleualaalaalaasp
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnthrlysaspasnthrglytrpthrproleuhisleuseralaasp
100105110
leuglyargleugluilevalgluvalleuleulystyrglyalaasp
115120125
valasnalaglnasplyspheglylysthralapheaspileserile
130135140
aspasnglyasngluaspleualagluileleuglnlysalaala
145150155
<210>17
<211>159
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>17
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
pheasptrpmetglytrpthrproleuhisleualaalahisglugly
354045
hisleugluilevalgluvalleuleulysasnglythraspvalasn
505560
alathraspvalserglytyrthrproleuhisleualaalaalaasp
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnthrlysaspasnthrglytrpthrproleuhisleuseralaasp
100105110
leuglyhisleugluilevalgluvalleuleulyshisglyalaasp
115120125
valasnalaglnasplyspheglylysthralapheaspileserile
130135140
aspasnglyasngluaspleualagluileleuglnlysalaala
145150155
<210>18
<211>159
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>18
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
pheasptrpmetglytrpthrproleuhisleualaalahisglugly
354045
hisleugluilevalgluvalleuleulysasnglyalaaspvalasn
505560
alathraspvalserglytyrthrproleuhisleualaalaalaasp
65707580
glyhisleugluilevalgluvalleuleulyshisglyalaaspval
859095
asnthrlysaspasnthrglytrpthrproleuhisleuseralaasp
100105110
leuglyhisleugluilevalgluvalleuleulysasnglyalaasp
115120125
valasnalaglnasplyspheglylysthralapheaspileserile
130135140
aspasnglyasngluaspleualagluileleuglnlysalaala
145150155
<210>19
<211>159
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>19
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
pheasptrpmetglytrpthrproleuhisleualaalahisglugly
354045
hisleugluilevalgluvalleuleulysasnglyalaaspvalasn
505560
alathraspvalserglytyrthrproleuhisleualaalaalaasp
65707580
glyhisleugluilevalgluvalleuleulyshisglyalaaspval
859095
asnthrlysaspasnthrglytrpthrproleuhisleuseralaasp
100105110
leuglyhisleugluilevalgluvalleuleulysasnglyalaasp
115120125
ileasnalaglnasplyspheglylysthralapheaspileserile
130135140
aspasnglyasngluaspleualagluileleuglnlysalaala
145150155
<210>20
<211>159
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>20
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
pheasptrpmetglytrpthrproleuhisleualaalahisglugly
354045
hisleugluilevalgluvalleuleulysasnglyalaaspvalasn
505560
alathraspvalserglytyrthrproleuhisleualaalaalaasp
65707580
glyhisleugluilevalgluvalleuleulyshisglyalaaspval
859095
asnthrthraspasnthrglytrpthrproleuhisleuseralaasp
100105110
leuglyhisleugluilevalgluvalleuleulystyrglyalaasp
115120125
valasnalaglnasplyspheglylysthralapheaspileserile
130135140
aspasnglyasngluaspleualagluileleuglnlysalaala
145150155
<210>21
<211>159
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>21
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
pheasptyrmetglytrpthrproleuhisleualaalahisasngly
354045
hismetgluilevalgluvalleuleulystyrglyalaaspvalasn
505560
alaserasptyrserglytyrthrproleuhisleualaalaalaasp
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnthrlysaspasnthrglytrpthrproleuhisleuseralaasp
100105110
leuglyhisleugluilevalgluvalleuleulystyrglyalaasp
115120125
valasnalaglnasplyspheglylysthralapheaspileserile
130135140
aspasnglyasngluaspleualagluileleuglnlysalaala
145150155
<210>22
<211>159
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>22
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
valasptyrileglytrpthrproleuhisleualaalaalatyrgly
354045
hisleugluilevalgluvalleuleulystyrseralaaspvalasn
505560
alagluaspphealaglytyrthrproleuhisleualaalaserasn
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnthrlysaspasnthrglytrpthrproleuhisleuseralaasp
100105110
leuglyhisleugluilevalgluvalleuleulystyrglyalaasp
115120125
valasnthrglnasplyspheglylysthralapheaspileserile
130135140
aspasnglyasngluaspleualagluileleuglnlysalaala
145150155
<210>23
<211>159
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>23
glyseraspleuglylyslysleuleuglualaalaargthrglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
thrasptyrmetglytrpthrproleuhisleualaalalysvalgly
354045
hisleugluilevalgluvalleuleulystyrglyalaaspvalasn
505560
alagluasptyrasnglytyrthrproleuhisleualaalaalamet
65707580
glyhisleugluilealagluvalleuleulystyrglyalaaspval
859095
asnthrlysaspasnthrglytrpthrproleuhisleuseralaasp
100105110
leuglyhisleugluilevalgluvalleuleulysasnglyalaasp
115120125
valasnalaglnasplyspheglylysthralapheaspileserile
130135140
aspasnglyasngluaspleualagluileleuglnlysalaala
145150155
<210>24
<211>126
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>24
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
argaspserthrglytrpthrproleuhisleualaalaprotrpgly
354045
hisprogluilevalgluvalleuleulysasnglyalaaspvalasn
505560
alaalaasppheglnglytrpthrproleuhisleualaalaalaval
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnalaglnasplyspheglylysthralapheaspileserileasp
100105110
asnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>25
<211>126
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>25
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
argaspserthrglytrpthrproleuhisleualaalaprotrpgly
354045
hisprogluilevalgluvalleuleulysasnglyalaaspvalasn
505560
alaalaasppheglnglytrpthrproleuhisleualaalaalaval
65707580
glyhisleugluilevalgluvalleuleulyshisglyalaaspval
859095
asnalaglnasplyspheglylysthralapheaspileserileasp
100105110
asnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>26
<211>126
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>26
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnthr
202530
alaaspserthrglytrpthrproleuhisleualaalaprotrpgly
354045
hisprogluilevalgluvalleuleulysasnglyalaaspvalasn
505560
alahisasptyrglnglytrpthrproleuhisleualaalathrleu
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnalaglnasplyspheglylysthralapheaspileserileasp
100105110
asnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>27
<211>126
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>27
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnthr
202530
alaaspserthrglytrpthrproleuhisleuvalalaprotrpgly
354045
hisprogluilevalgluvalleuleulyshisglyalaaspvalasn
505560
thrhisasptyrglnglytrpthrproleuhisleualaalathrleu
65707580
glyhisleugluilevalgluvalleuleuargtyrglyalaaspval
859095
asnalaglnasplyspheglylysthralapheaspileserileasp
100105110
asnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>28
<211>126
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>28
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnthr
202530
alaaspserthrglytrpthrpromethisleualaalaprotrpgly
354045
hisprogluilevalgluvalleuleulyshisglyalaaspvalasn
505560
alaglnasppheglnglytrpthrproleuhisleualaalaalaile
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnalaglnasplyspheglylysthralapheaspileserileasp
100105110
asnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>29
<211>126
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>29
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnthr
202530
alaaspserthrglytrpthrproleuhisleualavalprotrpgly
354045
hisleugluilevalgluvalleuleulystyrglyalaaspvalasn
505560
alalysasppheglnglytrpthrproleuhisleualaalaalaile
65707580
glyhisglngluilevalgluvalleuleulysasnglyalaaspval
859095
asnalaglnasplyspheglylysthralapheaspileserileasp
100105110
asnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>30
<211>126
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>30
glyseraspleuglylyslysleuleuglualaalaargvalglygln
151015
aspaspgluvalargileleumetalaaspglyalaaspvalasnala
202530
seraspphelysglyaspthrproleuhisleualaalaserglngly
354045
hisleugluilevalgluvalleuleulystyrglyalaaspvalasn
505560
alatyraspmetleuglytrpthrproleuhisleualaalaaspleu
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnalaglnaspargpheglylysthralapheaspileserileasp
100105110
asnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>31
<211>126
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>31
glyseraspleuglylyslysleuleuglualaalaargvalglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
seraspphelysglyaspthrproleuhisleualaalaserglngly
354045
hisleugluilevalgluvalleuleulysasnseralaaspvalasn
505560
alapheaspleuleuglytrpthrproleuhisleualaalaaspleu
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnalaglnasplyspheglylysthralapheaspileserileasp
100105110
asnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>32
<211>126
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>32
glyseraspleuglylyslysleuleuglualaalaargvalglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
leuaspphelysglyaspthrproleuhisleualaalaalasergly
354045
hisleugluilevalgluvalleuleulysasnglyalaaspvalasn
505560
alahisaspmetleusertrpthrproleuhisleualaglyaspleu
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnalaglnaspargpheglylysthralapheaspileserileasp
100105110
asnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>33
<211>126
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>33
glyseraspleuglylyslysleuleuglualavalargalaglygln
151015
aspaspgluvalargileleumetthrasnglyalaaspvalasnala
202530
lysaspglnpheglyphethrproleuglnleualaalatyrasngly
354045
hisleugluilevalgluvalleuleulystyrglyalaaspvalasn
505560
alapheaspilepheglytrpthrproleuhisleualaalaaspleu
65707580
glyhisleugluilevalgluvalleuleulysasnglyalaaspval
859095
asnalaglnasplyspheglyargthralapheaspileserileasp
100105110
asnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>34
<211>127
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>34
glyseraspleuglylyslysleuleuglualavalargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
seraspasnglnglythrthrproleuhisleualaalaserhisgly
354045
hisleugluilevalgluvalleuleulystyrglyalaaspvalasn
505560
aspalahisaspaspleuglytrpthrproleuhisleuseralaasp
65707580
leuglyhisleugluilevalgluvalleuleulystyrglyalaasp
859095
valasnalaglnasplyspheglylysthralapheaspileserile
100105110
aspasnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>35
<211>127
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>35
glyseraspleuglylyslysleuleuglualathrargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
seraspasnglnglythrthrproleuhisleualaalaserhisgly
354045
hisleugluilevalgluvalleuleulystyrglyalaaspvalasn
505560
aspalahisaspaspleuglytrpthrproleuhisleualaalaasp
65707580
leuglyhisleugluilevalgluvalleuleulystyrglyalaasp
859095
valasnalaglnasplyspheglylysthralapheaspileserile
100105110
aspasnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>36
<211>126
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>36
glyseraspleuglylyslysleuleuglualaalaargvalglygln
151015
aspaspgluvalargileleumetalaaspglyalaaspvalasnala
202530
seraspphelysglyaspthrproleuhisleualaalaserglngly
354045
hisleugluilevalgluvalleuleulystyrglyalaaspvalasn
505560
alatyraspmetleuglytrpthrproleuhisleualaalaaspleu
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnalaglnaspargpheglylysthralapheaspileserileasp
100105110
asnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>37
<211>126
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>37
glyseraspleuglylyslysleuleuglualaalaargvalglygln
151015
aspaspgluvalargileleumetalaasnaspalaaspvalasnala
202530
seraspphelysglyaspthrproleuhisleualaalaserglngly
354045
hisleugluilevalgluvalleuleulystyrglyalaaspvalasn
505560
alatyraspmetleuglytrpthrproleuhisleualaalaaspleu
65707580
glyhisleugluilevalgluvalleuleulyshisglyalaaspval
859095
asnalaglnasplyspheglylysthralapheaspileserileasp
100105110
asnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>38
<211>126
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>38
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnthr
202530
leuaspphelysseraspthrproleuhisleualaalaalasergly
354045
hisleugluilevalgluvalleuleulysasnglyalaaspvalasn
505560
alahisaspmetleusertrpthrproleuhisleualaglyaspleu
65707580
glyhisleugluilevalgluvalleuleulyshisglyalaaspval
859095
asnalaglnasplyspheglylysthralapheaspileserileasp
100105110
asnglyasngluaspleualagluileleuglnlysalaala
115120125
<210>39
<211>159
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>39
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
lysaspiletyrglyargthrproleuhisleualaalaleuhisgly
354045
hisprogluilevalgluvalleuleulystyrglyalaaspvalasn
505560
alaasnasptyrtrpglythrthrserleuhisleuvalalailetrp
65707580
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
859095
asnalavalaspaspileglyglnthrproleuhisleualaalaala
100105110
trpglyhisleugluilevalgluvalleuleulyshisglyalaasp
115120125
valasnalaglnasplyspheglylysthralapheaspileserile
130135140
aspasnglyasngluaspleualagluileleuglnlysalaala
145150155
<210>40
<211>158
<212>prt
<213>人工的
<220>
<223>锚蛋白重复结构域
<400>40
glyseraspleuglylyslysleuleuglualaalaargalaglygln
151015
aspaspgluvalargileleumetalaasnglyalaaspvalasnala
202530
asnasptyraspglymetthrproleuhisleualaalametglugly
354045
hisleugluilevalgluvalleuleulystyrglyalaaspvalasn
505560
alaasnasphistyrglyphethrproleuhisleualatrpthrgly
65707580
argleugluilevalgluvalleuleulysasnglyalaaspvalasn
859095
alaalaaspvalpheglyargthrproleuhisleualaalathrser
100105110
glyhisleugluilevalgluvalleuleulystyrglyalaaspval
115120125
asnalaglnasplyspheglylysthralapheaspileserileasp
130135140
asnglyasngluaspleualagluileleuglnlysalaala
145150155
<210>41
<211>9
<212>prt
<213>人工的
<220>
<223>gs接头
<400>41
glyglyglyserglyglyglysercys
15
- 还没有人留言评论。精彩留言会获得点赞!
- Mdm2和p53相互作用的抑制剂的制作方法
- 用于抑制pf4和rantes相互作用的拮抗剂的制作方法
- Mdm2及p53间相互作用的抑制剂的制作方法
- 单独的和与PDE5抑制剂相组合的sGC刺激剂、sGC活化剂用于治疗系统性硬化症(SSc)的用途的制作方法
- 用于抑制bcl2与结合配偶体相互作用的化合物的制作方法
- 用于抑制bcl2与结合配偶体相互作用的化合物的制作方法
- 用于抑制bcl2与结合配偶体相互作用的化合物的制作方法
- 抑制下游 u P A R相互作用并结合了尿激酶类纤溶酶原激发剂( u P A)与其受体( u P ...的制作方法
- 灵芝酸作为p53-MDM2相互作用抑制剂的用途
- Mdm2与p53间相互作用的抑制剂的制作方法