小麦育性恢复基因及其应用的制作方法
本发明属于植物生物技术领域,具体涉及植物隐性核雄性不育基因的克隆,及其雄性不育系的繁殖方法和在杂交育种中的应用,更具体地涉及一个隐性核雄性不育基因及其启动子的克隆,及其在杂交育种中的应用。
技术背景
杂种优势是生物界的普遍现象,杂交种育种是选育新品种的主要途径,是近代育种工作最重要方法。与水稻、玉米、高粱等相比,小麦杂种优势利用的研究相对滞后,近十年来其平均产量的增长也趋于停滞、甚至出现连续多年下滑的局面。小麦是自花授粉作物,小麦杂种优势利用的核心问题是高效生产小麦杂交种的技术体系,国内外大量科学家为此已做出巨大的努力,并取得一系列重要成果。综合近50年来的研究进展,小麦杂种优势利用研究主要集中于:核质互作雄性不育的利用(“三系法”)、化学杀雄技术的利用(“化杀法”)和光温敏核雄性不育的利用(“两系法”)。三系法由于不育系难以繁殖、恢复源较窄、细胞质副效应等原因,未能在生产上大面积应用。化杀法避开了恢复与保持间的相互关系,曾被认为是一种很有希望的小麦杂交制种新技术,但由于其在制种过程中稳定性差、制种成本高及环境污染等多方面原因,在实际生产上也难以推广利用。基于光温敏的两系法虽然具有制种成本低、恢复源广泛,较易获得优势组合等优点,但也面临着两大关键问题——环境因素的不稳定性对不育系育性的影响和利用光温敏特性所选育的小麦不育系十分有限。
隐性核雄性不育突变体用于作物杂种优势利用,其不育性具有易被恢复而不易被保持的特性。与细胞质雄性不育杂交小麦体系相比,隐性核雄性不育突变体用于杂交小麦开发具有以下优点:1)不存在外源细胞质的负面影响,杂种f1优势强;2)父本系对杂种f1的育性恢复度高;3)不育系、保持系、父本系的育种亲本材料选择不受特定恢/保关系的限制,选材范围极广、种质资源利用率高,有利于选育高配合力的杂交种。但是,按照常规方法无法实现纯合核不育系种子的大量有效生产。因此,针对小麦杂种优势利用的现状,建立高效小麦雄性不育繁育新体系,是杂交小麦获得成功应用的最关键因素之一。
小麦显性核雄性不育系由于其自身的遗传特点,既找不到完全的恢复系,也找不到完全的保持系,比如1972年在我国发现的太谷核不育系(即ms2),因此只适用于常规育种轮回选择和回交育种手段,不能用作杂交小麦育种的亲本。而隐性核雄性不育材料与任何正常材料杂交,f1代均可育,任何育性正常的材料都是其恢复系,只要解决核不育性的标识和有效保持的问题,就能应用于新一代小麦杂交育种技术。
小麦基因组巨大(17gb),约是人类的5倍,水稻的40倍,拟南芥的100倍;组成极其复杂,由a、b、d三个具有部分同源关系的染色体组组成,每个染色体组由7对染色体构成,共有21对染色体,是典型的异源六倍体(zhangzb,etal.,2002),并且约75%是简单重复序列(rachelb,etal.,2012;iwgsc,2014)。近年来,虽然小麦及其近缘种的基因组测序工作取得了很大进展,但截至目前,仍然没有完整的参考基因组序列公布(vogeljp,etal.,2010;theinternationalbarleygenomesequencingconsortium,2012;rachelb,etal.,2012;linghq,etal.,2013;jiaj,etal.,2013;iwgsc,2014)。如此复杂的基因组使得小麦功能基因的研究工作异常困难,目前为止国际范围内小麦突变体成功克隆基因的例子只有寥寥几个。
本发明通过流式细胞术和高通量测序的方法,成功克隆了育性恢复基因frg1,该基因可以完全恢复兰州核雄性不育突变体或其等位突变体的雄性育性,为构建新型小麦杂交育种技术体系奠定了基础。同时,为解决小麦现有的“三系”和两系”杂交技术所存在不育系育性不稳定、杂交品种资源受局限、制种技术复杂、制种成本高等技术瓶颈问题,提供了更多的可能性。本发明所提供的基因及不育系的繁殖和保持方法,对小麦杂交育种工作具有重要的意义和应用价值。
技术实现要素:
本文提到的所有参考文献都通过引用并入本文。
除非有相反指明,本文所用的所有技术和科学术语都具有与本发明所属领域普通技术人员通常所理解的相同的含义。除非有相反指明,本文所使用的或提到的技术是本领域普通技术人员公知的标准技术。材料、方法和例子仅作阐述用,而非加以限制。
本发明提供了一个育性恢复基因frg1(fertilityrestorationgene1),所述育性恢复基因的核苷酸序列选自下列组的序列之一:
(a)如seqidno:1或2所示的核苷酸序列;
(b)其编码氨基酸序列如seqidno:3所示的核苷酸序列;
(c)在严谨条件下能够与(a)或(b)中所述序列的dna杂交的dna序列;或
(d)与(a)-(c)所述序列有至少80%(优选为至少85%)序列相似性,且具有育性恢复功能的dna序列;或
(e)与(a)-(d)之任一所述序列互补的dna序列。
本领域技术人员应该知晓,本发明所述的育性恢复基因还包括与frg1基因的核苷酸序列或蛋白序列高度同源,并且具有同样的育性调控或恢复功能的同源基因序列。所述高度同源且具有育性调控功能的的同源基因包括在严谨条件下能够与具有seqidno:1或2所示序列的dna杂交的dna序列。或是其编码的氨基酸序列与seqidno:3所示的蛋白氨基酸序列具有85%以上相似性的核苷酸序列。本文中使用的“严谨条件”是公知的,包括诸如在含400mmnacl、40mmpipes(ph6.4)和1mmedta的杂交液中于53℃-60℃杂交12-16小时,然后在62℃-68℃下用含0.5×ssc、和0.1%sds的洗涤液洗涤15-60分钟。
上述同源基因还包括与seqidno:1或2所示序列的全长有至少80%、85%、90%、95%、98%、或99%序列相似性,且具有育性调控功能的dna序列,可以从任何植物中分离获得。其中,序列相似性的百分比可以通过公知的生物信息学算法来获得,包括myers和miller算法、needleman-wunsch全局比对法、smith-waterman局部比对法、pearson和lipman相似性搜索法、karlin和altschul的算法。这对于本领域技术人员来说是公知的。
本发明还提供了一种表达盒,所述表达盒含有本发明所公开的育性恢复基因的dna序列,所述育性恢复基因的核苷酸序列选自下列组的序列之一:
(a)如seqidno:1或2所示的核苷酸序列;
(b)其编码氨基酸序列如seqidno:3所示的核苷酸序列;
(c)在严谨条件下能够与(a)或(b)中所述序列的dna杂交的dna序列;或
(d)与(a)-(c)所述序列有至少80%(优选为至少85%)序列相似性,且具有育性恢复功能的dna序列;或
(e)与(a)-(d)之任一所述序列互补的dna序列。
具体地,上述表达盒中的育性恢复基因还可操作性的连有一个可驱动其表达的启动子,所述启动子包括但不限于组成型表达启动子、诱导型启动子、组织特异表达启动子、或时空特异表达启动子。更具体地,所述启动子是一个花粉特异表达启动子。优选地,所述花粉特异表达启动子的核苷酸序列如seqidno:4所示。
本发明上述表达盒,还进一步的包含一个花粉失活基因,所述花粉失活基因可以干扰植株中含有该花粉失活基因的雄性配子的功能或形成。所述花粉失活基因包括但不限于barnase基因、淀粉酶基因、dam甲基化酶等。更具体的,所述花粉失活基因是玉米a淀粉酶基因,优选其核苷酸序列如seqidno:6所示。
本发明上述表达盒,还进一步的包含一个筛选基因,所述筛选基因可以用于将含有该表达盒的植株、植物组织细胞或载体筛选出来。所述筛选基因包括但不限于抗生素抗性基因、或是抗除草剂基因、或是荧光蛋白基因等。具体地,所述筛选基因包括但不限于:氯霉素抗性基因、潮霉素抗性基因、链霉素抗性基因、奇霉素抗性基因、磺胺类抗性基因、草甘磷抗性基因、草丁膦抗性基因、bar基因、红色荧光基因dsred、mcherry基因、青色荧光蛋白基因、黄色荧光蛋白基因、荧光素酶基因、绿色荧光蛋白基因等。
本发明还公开了一种植物育性调控的方法,所述方法通过将育性恢复基因转入到兰州核雄性不育突变体(周宽基等,1996)或其等位突变体中,使兰州核雄性不育突变体或其等位突变体的雄性育性恢复,其中所述的育性恢复基因的核苷酸序列选自下列组的序列之一:
(a)如seqidno:1或2所示的核苷酸序列;
(b)其编码氨基酸序列如seqidno:3所示的核苷酸序列;
(c)在严谨条件下能够与(a)或(b)中所述序列的dna杂交的dna序列;或
(d)与(a)-(c)所述序列有至少80%(优选为至少85%)序列相似性,且具有育性恢复功能的dna序列;或
(e)与(a)-(d)之任一所述序列互补的dna序列。
本发明中所述的兰州核雄性不育突变体,又称为兰州核不育或兰州核不育系或兰州核不育突变体。
本发明还公开了一种雄性不育系的保持方法,所述方法以兰州核雄性不育突变体或其等位突变体为转化受体材料,将紧密连锁的3个目标基因转化至该不育突变体受体植株中。所述3个目标基因分别是育性恢复基因frg1、花粉失活基因和筛选标记基因。其中,育性恢复基因frg1可使不育的转化受体育性恢复,花粉失活基因可使含有转化的外源基因的花粉失活,即失去授精能力,筛选基因可以用于转基因种子或组织和非转基因种子或组织的分拣,分拣出的非转基因种子用作不育系生产杂交种,转基因种子用作保持系来源源不断地、稳定地生产不育系。
在本发明中,所述兰州核雄性不育突变体或其等位突变体也可以称为不育系或雄性不育系或兰州核雄性不育系、或等位不育系。
上述雄性不育系的保持方法中,所述的花粉失活基因包括但不限于barnase基因、淀粉酶基因、dam甲基化酶等。更具体的,所述花粉失活基因是玉米a淀粉酶基因zm-aa,优选其核苷酸序列如seqidno:6所示。所述花粉失活基因与偏好于雄性配子表达的启动子相连。更具体地,所述偏好于雄性配子表达的启动子包括但不限于pg47启动子、zm13启动子等。所述筛选基因可以用于将含有该表达盒的植株或载体筛选出来。所述筛选基因包括但不限于抗生素抗性基因、或是抗除草剂基因、或是荧光蛋白基因等。具体地,所述筛选基因包括但不限于:氯霉素抗性基因、潮霉素抗性基因、链霉素抗性基因、奇霉素抗性基因、磺胺类抗性基因、草甘磷抗性基因、草丁膦抗性基因、bar基因、红色荧光基因dsred、mcherry基因、青色荧光蛋白基因、黄色荧光蛋白基因、荧光素酶基因、绿色荧光蛋白基因等。
更具体地,本发明还公开了一种雄性不育系的繁殖方法,所述方法包括以下步骤:
(a)向兰州核雄性不育系或其等位不育系中转入下述载体,以获得含有下述载体的保持系,所述载体包含:育性恢复基因frg1,所述育性恢复基因frg1可以恢复兰州核雄性不育系或其等位不育系的雄性生育力;和花粉失活基因,所述花粉失活基因表达时,会干扰植株中含有该花粉失活基因的雄性配子的功能或形成,从而使得所述植株中产生的具有活性的雄性配子都是不含所述载体的;和筛选基因,所述筛选基因可以用于转基因种子或组织和非转基因种子或组织的分拣。
(b)将转入上述载体后形成的保持系植株自交,同时产生不含载体的兰州核雄性不育系或其等位不育系种子和含载体的保持系种子;或是将保持系植株的花粉赶到兰州核雄性不育系或其等位不育系植株上,使兰州核雄性不育系或其等位不育系授粉繁殖出兰州核雄性不育系或其等位不育系种子。
上述雄性不育系的繁殖方法中,所述的花粉失活基因包括但不限于barnase基因、淀粉酶基因、dam甲基化酶等。更具体的,所述花粉失活基因是玉米a淀粉酶基因zm-aa,优选其核苷酸序列如seqidno:6所示。所述花粉失活基因与偏好于雄性配子表达的启动子相连。更具体地,所述偏好于雄性配子表达的启动子包括但不限于pg47启动子、zm13启动子等。所述筛选基因可以用于将含有该表达盒的植株或载体筛选出来。所述筛选基因包括但不限于抗生素抗性基因、或是抗除草剂基因、或是荧光蛋白基因等。具体地,所述筛选基因包括但不限于:氯霉素抗性基因、潮霉素抗性基因、链霉素抗性基因、奇霉素抗性基因、磺胺类抗性基因、草甘磷抗性基因、草丁膦抗性基因、bar基因、红色荧光基因dsred、mcherry基因、青色荧光蛋白基因、黄色荧光蛋白基因、荧光素酶基因、绿色荧光蛋白基因等。
本发明还公开了一种保持系的生产方法,所述方法包括以下步骤:
(a)向兰州核雄性不育系或其等位不育系中转入下述载体,即获得了兰州核雄性不育系或其等位不育系的保持系,所述载体包含:育性恢复基因frg1,所述育性恢复基因frg1可以恢复兰州核雄性不育系或其等位不育系的雄性生育力;和花粉失活基因,所述花粉失活基因表达时,会干扰植株中含有该花粉失活基因的雄性配子的功能或形成,从而使得所述植株中产生的可育雄性配子都是不含所述载体的;和筛选基因,所述筛选基因可以用于转基因种子和非转基因种子的分拣。
上述保持系的生产方法中,所述的花粉失活基因包括但不限于barnase基因、淀粉酶基因、dam甲基化酶等。更具体的,所述花粉失活基因是玉米a淀粉酶基因zm-aa,优选其核苷酸序列如seqidno:6所示。所述花粉失活基因与偏好于雄性配子表达的启动子相连。更具体地,所述偏好于雄性配子表达的启动子包括但不限于pg47启动子、zm13启动子等。所述筛选基因可以用于将含有该表达盒的植株或载体筛选出来。所述筛选基因包括但不限于抗生素抗性基因、或是抗除草剂基因、或是荧光蛋白基因等。具体地,所述筛选基因包括但不限于:氯霉素抗性基因、潮霉素抗性基因、链霉素抗性基因、奇霉素抗性基因、磺胺类抗性基因、草甘磷抗性基因、草丁膦抗性基因、bar基因、红色荧光基因dsred、mcherry基因、青色荧光蛋白基因、黄色荧光蛋白基因、荧光素酶基因、绿色荧光蛋白基因等。
本发明还公开了一种保持系的繁殖方法,所述方法包括以下步骤:
(a)向兰州核雄性不育系或其等位不育系中转入下述载体,即获得了兰州核雄性不育系或其等位不育系的保持系,所述载体包含:育性恢复基因frg1,所述育性恢复基因frg1可以恢复兰州核雄性不育系或其等位不育系的雄性生育力;和花粉失活基因,所述花粉失活基因表达时,会干扰植株中含有该花粉失活基因的雄性配子的功能或形成,从而使得所述植株中产生的可育雄性配子都是不含所述载体的;和筛选基因,所述筛选基因可以用于转基因种子和非转基因种子的分拣;和
(b)将转入上述载体后形成的保持系植株自交,即按1:1的比例繁殖获得了不含载体的兰州核雄性不育系或其等位不育系种子和含载体的保持系种子。
本发明还公开了一种种子的生产方法,所述方法包括:
(a)向兰州核雄性不育系或其等位不育系中引入下述载体,获得兰州核雄性不育系或其等位不育系的保持系,所述载体包含:育性恢复基因frg1,所述育性恢复基因frg1可以恢复兰州核雄性不育系或其等位不育系的雄性生育力;和花粉失活基因,所述花粉失活基因表达时,会干扰植株中含有该花粉失活基因的雄性配子的功能或形成,从而使得所述植株中产生的可育雄性配子都是不含所述载体的。
(b)将转入上述载体后的保持系植株自交;和
(c)自交后即获得含有所述载体的保持系种子和不含载体的兰州核雄性不育系或其等位不育系种子。
本发明上述的雄性不育系的繁殖或保持方法、保持系的生产方法或繁殖方法、种子的生产方法等中,其中步骤(a)也可以是向普通的植株中引入含有育性恢复基因frg1、花粉失活基因和筛选基因的载体,获得含有所述载体的转基因植株后,再与兰州核雄性不育系或其等位不育系杂交,经过定向选育,获得背景为兰州核雄性不育系或其等位不育系、并且含有所述载体的保持系植株。
本发明上述的雄性不育系的繁殖方法或保持方法、保持系的生产方法或繁殖方法、种子的生产方法等中,其中所述的育性恢复基因的核苷酸序列选自下列组的序列之一:
(a)如seqidno:1或2所示的核苷酸序列;
(b)其编码氨基酸序列如seqidno:3所示的核苷酸序列;
(c)在严谨条件下能够与(a)或(b)中所述序列的dna杂交的dna序列;或
(d)与(a)-(c)所述序列有至少80%(优选为至少85%)序列相似性,且具有育性恢复功能的dna序列;或
(e)与(a)-(d)之任一所述序列互补的dna序列。
上述育性恢复基因frg1还可操作性的连有一个花粉特异表达的启动子,可以驱动frg1基因在植物花粉中的表达。所述花粉特异表达的启动子选自由ms26、np1、msp1、pair1、pair2、zep1、mell、pss1、tdr、udt1、gamyb4、ptc1、api5、wda1、cyp704b2、ms26、ms22、dpw、mads3、osc6、rip1、csa、aid1、5126或ms45等育性调控基因的启动子构成的组之一。更具体的,所述花粉特异表达启动子的核苷酸序列如seqidno:4所示。上述育性恢复基因frg1还可操作性的连有一个终止子,所述终止子可以是已经公开的任一个基因的终止子,具体地,其中一个终止子的核苷酸序列如seqidno:5所示。本发明上述的雄性不育系的繁殖或保持方法、保持系的生产方法或繁殖方法、种子的生产方法等中,所述的花粉失活基因包括但不限于barnase基因、淀粉酶基因、dam甲基化酶等。更具体的,所述花粉失活基因是玉米a淀粉酶基因zm-aa,优选其核苷酸序列如seqidno:6所示。所述花粉失活基因与偏好于雄性配子表达的启动子相连。更具体地,所述偏好于雄性配子表达的启动子包括但不限于pg47启动子、zm13启动子等。
本发明上述的雄性不育系的繁殖或保持方法、保持系的生产方法或繁殖方法、种子的生产方法等中,其中所述的筛选基因包括但不限于抗生素抗性基因、除草剂抗性基因或荧光基因。具体地,所述筛选基因包括但不限于:氯霉素抗性基因、潮霉素抗性基因、链霉素抗性基因、奇霉素抗性基因、磺胺类抗性基因、草甘磷抗性基因、草丁膦抗性基因、bar基因、红色荧光基因dsred、mcherry基因、青色荧光蛋白基因、黄色荧光蛋白基因、荧光素酶基因、绿色荧光蛋白基因等。
本发明还提供了一种花粉特异表达启动子,其核苷酸序列如seqidno:4所示。将seqidno:4与报告基因gus相连,构建载体转化水稻和小麦,检测分析转基因植株中的gus表达活性和表达模式,通过对转基因植株的根、茎、叶和花进行gus染色分析,结果发现本发明所提供的启动子驱动gus基因在植物花粉中表达。说明本发明所提供的seqidno:4是一个花粉特异性表达的启动子。
本发明所提供的植物花粉特异表达启动子,含有序列表中如seqidno:4所示的核苷酸序列,或包含与seqidno:4中所列核苷酸序列具有90%以上相似性的核苷酸序列,或包含来源于seqidno:4序列上的500个及500以上连续的核苷酸片段,并且可以驱动与该启动子操作性连接的核苷酸序列在植物花粉中的表达。含有上述序列的表达载体、转基因细胞系以及宿主菌等均属于本发明的保护范围。扩增本发明所公开的seqidno:4启动子的任一核苷酸片段的引物对也在本发明的保护范围之内。
本发明所述的“启动子”是指一种dna调控区域,其通常包含能指导rna聚合酶ii在特定编码序列的合适转录起始位点起始rna合成的tata盒。启动子还可包含其它识别序列,这些识别序列通常位于tata盒的上游或5’端,通常被称为上游启动子元件,起调控转录效率的作用。本领域技术人员应该知晓,虽然已经鉴定了针对本发明公开的启动子区域的核苷酸序列,但是分离和鉴定处于本发明鉴定的特定启动子区域的tata盒上游区域的其它调控元件也在本发明的范围内。因此,本文公开的启动子区域通常被进一步界定为包含上游调控元件,例如用于调控编码序列的组织表达性和时间表达功能的那些元件、增强子等。以相同的方式,可以鉴定、分离出使得能在目标组织(例如雄性组织)中进行表达的启动子元件,将其与其它核心启动子一起使用,以验证雄性组织优先的表达。核心启动子指起始转录所需的最小限度的序列,例如被称为tata盒的序列,这是编码蛋白质的基因的启动子通常都具有的。因此,可选地,frg1基因的上游启动子可与其自身的或来自其它来源的核心启动子关联使用。
核心启动子可以是任何一种已知的核心启动子,例如花椰菜花叶病毒35s或19s启动子(美国专利no.5,352,605)、泛素启动子(美国专利no.5,510,474)、in2核心启动子(美国专利no.5,364,780)或玄参花叶病毒启动子。
所述基因启动子的功能可以通过以下方法进行分析:将启动子序列与报告基因可操作性连接,形成可转化的载体,再将该载体转入植株中,在获得转基因后代中,通过观察报告基因在植物各个组织器官中的表达情况来确认其表达特性;或者将上述载体亚克隆进用于瞬时表达实验的表达载体,通过瞬时表达实验来检测启动子或其调控区的功能。
用来测试启动子或调控区域功能的适当表达载体的选择将取决于宿主和将该表达载体引入宿主的方法,这类方法是本领域普通技术人员所熟知的。对于真核生物,在载体中的区域包括控制转录起始和控制加工的区域。这些区域被可操作地连接到报告基因,所述报告基因包括yfp、uida、gus基因或荧光素酶。包含位于基因组片段中的推定调控区的表达载体可以被引入完整的组织,例如阶段性花粉,或引入愈伤组织,以进行功能验证。
此外,本发明的启动子还可与并非frg1基因的核苷酸序列相连,以表达其它异源核苷酸序列。本发明的启动子核苷酸序列及其片段和变体可与异源核苷酸序列一起组装在一个表达盒中,用于在目的植株中表达,更具体地,在该植株的雄性器官中表达。所述表达盒有合适的限制性酶切位点,用于插入所述启动子和异源核苷酸序列。这些表达盒可用于对任何植株进行遗传操作,以获得想要的相应表型。
本发明所公开的花粉特异表达启动子,可用于驱动下列异源核苷酸序列的表达,以使转化的植株获得雄性不育的表型,所述异源核苷酸序列可编码促使碳水化合物降解的酶或修饰酶、淀粉酶、脱支酶和果胶酶,更具体的如barnase基因、玉米a淀粉酶基因、生长素基因、rotb、细胞毒素基因、白喉毒素、dam甲基化酶,或是显性的雄性不育基因。
在某些实施方式中,本发明中所提到的可操作性地连接在本发明启动子下游的核苷酸序列,其中所述的“核苷酸序列”可以是操作性连接于本文所公开的启动子之后的结构基因、调节基因、结构基因的反义基因、调节基因的反义基因或者能够干扰内源基因表达的小rna。
本发明还提供了一个转录终止子序列,所述转录终止子的核苷酸序列如seqidno:5所示,具有终止基因转录表达的功能。
本发明还提供了一种表达盒、载体或工程菌株,所述表达盒、载体或工程菌株中包含了本发明所提供的花粉特异表达启动子seqidno:4。具体地,可以将本发明所提供的育性恢复基因frg1的核苷酸序列构建到本发明所提供的启动子seqidno:4的下游,从而驱动该育性基因在转化受体植株中的表达。
本发明的所提供的花粉特异表达启动子可用于外源基因在花粉中的特异性表达,从而避免该外源基因在植物其他组织中持续表达所带来的不利影响,还可以用于植物花粉生长发育相关基因的功能分析和鉴定;可用于雄性不育系和保持系的创建;并可应用于花粉败育实验中,从而避免由植物转基因漂移或花粉逃逸所带来的生物安全问题,对植物雄性不育系和保持系的创造具有重要意义。
本发明所提供的frg1基因的核苷酸序列和启动子序列或表达盒可被插入载体、质粒、酵母人工染色体、细菌人工染色体或其他适合转化进宿主细胞中的任何载体中。优选的宿主细胞是细菌细胞,尤其是用于克隆或储存多核苷酸、或用于转化植物细胞的细菌细胞,例如大肠杆菌、根瘤土壤杆菌和毛根土壤杆菌。当宿主细胞是植物细胞时,表达盒或载体可插入至被转化的植物细胞的基因组中。插入可以是定位的或随机的插入。
本发明所述的将核苷酸序列、载体或表达盒转入植株或引入植株或对植株进行转化,均指通过常规的转基因方法,将核苷酸序列、载体或表达盒转入到受体细胞或受体植株中。植物生物技术领域技术人员已知的任何转基因方法均可被用于将重组表达载体转化进植物细胞中,以产生本发明的转基因植物。转化方法可包括直接和间接的转化方法。合适的直接方法包括聚乙二醇诱导的dna摄入、脂质体介导的转化、使用基因枪导入、电穿孔、以及显微注射。所述转化方法也包括农杆菌介导的植物转化方法等。
与现有技术相比,本发明具有如下的有益效果:本发明提供了一种育性恢复基因frg1及其启动子,及将该基因用于兰州核雄性不育系或其等位不育系的繁殖和保持的方法。本发明所提供的育性恢复基因,小麦隐性核雄性不育系的育性保持和不育系的繁殖,对小麦的杂交育种生产来说,具有重大的生产推广价值和应用价值,本发明提供的育性恢复基因解决了兰州核雄性不育系或其等位不育系的繁殖和保持问题,对于突破并改良现有的“三系”和“两系”杂交技术具有重要意义。
参考文献:
周宽基,周文麟,王淑英(1996)4e-ms小麦雄性核不育、保持体系的建立.中国农业科学29:93theinternationalbarleygenomesequencingconsortium(2012)aphysical,geneticandfunctionalsequenceassemblyofthebarleygenome.nature491:711
theinternationalwheatgenomesequencingconsortium(iwgsc)(2014)achromosome-baseddraftsequenceofthehexaploidbreadwheat(triticumaestivum)genome.science345:1251788
jiaj,zhaos,kongx,etal.(2013)aegilopstauschiidraftgenomesequencerevealsagenerepertoireforwheatadaptation.nature496:91
linghq,zhaos,liud,etal.(2013)draftgenomeofthewheata-genomeprogenitortriticumurartu.nature496:87
rachelb,manuels,matthiasp,etal.(2012)analysisofthebreadwheatgenomeusingwhole-genomeshotgunsequencing.nature491:705
vogeljp,garvindf,mocklertc,etal.(2010)genomesequencingandanalysisofthemodelgrassbrachypodiumdistachyon.nature463:763
zhangzb,xup(2002)reviewedonwheatgenome.hereditas24:389
附图说明
图1是4ags-ms双端体异附加系的gish分析。以长穗偃麦草基因组dna为探针进行杂交,红色荧光信号为4ags染色体;染色体用dapi染色呈深蓝色。
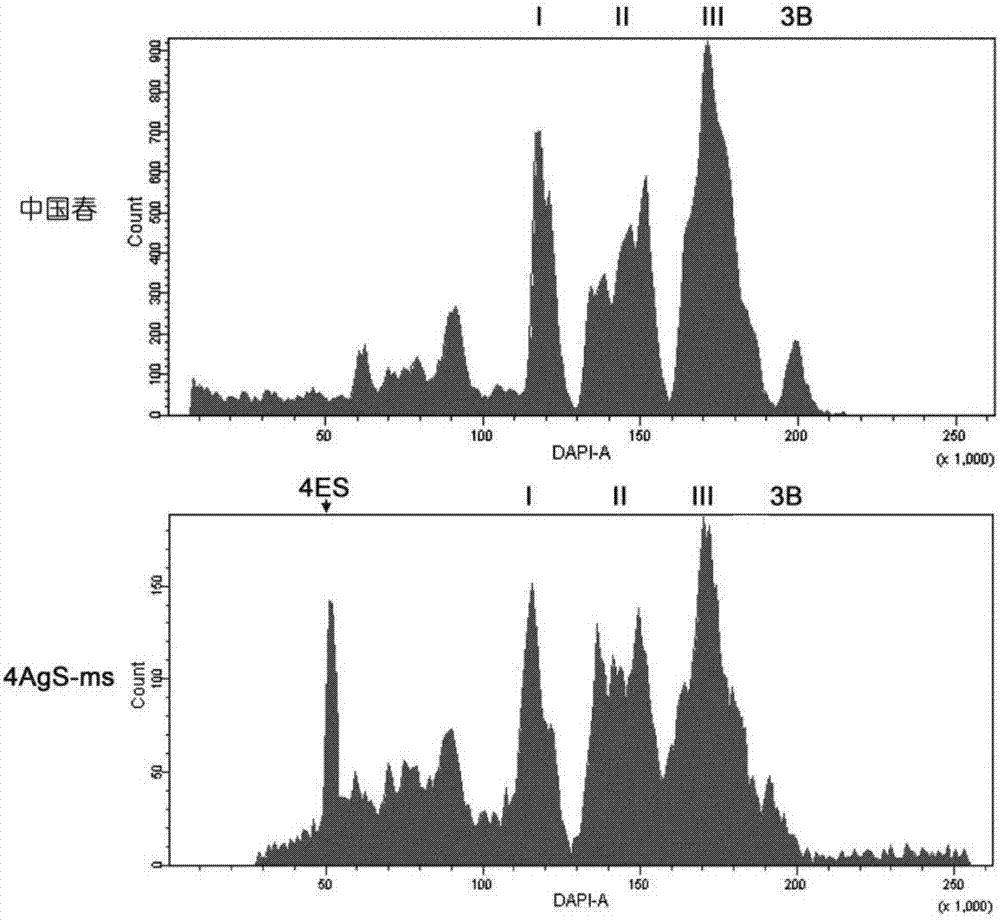
图2是中国春与4ags-ms双端体异附加系的流式核型图。上图为中国春材料,下图为4ags-ms双端体异附加系。横坐标为相对荧光强度,纵坐标为染色体数目。ⅰ、ⅱ、ⅲ和3b标记不同大小小麦染色体组群。4ags标记来自成穗偃麦草的4ag染色体。
图3是亲本材料表达值高于兰州核不育材料的基因的百分比在chr4b分布情况。
具体实施方式
下面对本发明的实施例作详细说明,本实施例在以本发明技术方案为前提下进行实施,给出了详细的实施方式和具体的操作过程,但本发明的保护范围不限于下述的实施例。
实施例1、4ags-ms双端体异附加系的选育
兰州核雄性不育突变体是在春小麦品种间杂交组合的杂种f4代群体中发现的。将其与包括中国春在内的9个小麦品系杂交,f1代自交,观察f2代不育株与可育株的分离比,均符合1:3的比例,表明小麦兰州核雄性不育突变体是典型的单基因控制的隐性突变(周宽基等,1996)。
蓝粒二体异附加系是在小麦基因组中附加了两条长穗偃麦草4ag染色体的小麦品系,籽粒为蓝色是由于附加的4ag染色体含有蓝粒基因ba。将蓝粒二体异附加系小麦的花粉授予兰州核雄性不育突变体,获得浅蓝粒杂种种子,f1植株能正常结实。经过有目标的选育获得了遗传背景为兰州核不育纯合突变体并附加了一条长穗偃麦草的4ag染色体的株系,命名为4ag-ms单体异附加系。该株系籽粒为蓝色的、自交可育,表明杂交进入兰州核雄性不育突变体的长穗偃麦草4ag染色体中含有育性恢复基因。
由于4ag-ms单体异附加系中只存在一条4ag染色体,因此其自交获得的种子存在蓝粒(4ag+)和白粒(4ag-)的分离,其中蓝粒的植株可育,白粒的植株不育。经过对4ag-ms单体异附加系的多代自交选育,获得了一个籽粒为白粒、植株为可育的株系。gish分析显示(图1),该材料中的4ag染色体发生断裂,导致大部分4ag染色体丢失,只留下一小部分保留在小麦染色体组中,因此将该株系命名为4ags-ms双端体异附加系。
4ags-ms双端体异附加系中4ags染色体的长度比小麦所有染色体都显著短,测量后估计其大小约为最长小麦染色体的1/4,大概250mb左右。由于4ags-ms双端体异附加系材料表现为白粒、可育,表明丢失的4ag染色体含有蓝粒基因ba,保留的4ag含有育性恢复基因fertilityrestorationgene1(frg1)。
实施例2、利用流式细胞术分离4ag染色体
采用双阻断法对根尖细胞进行细胞周期同步化处理,再经过甲醛固定、机械匀浆和dapi染色等步骤后,上流式细胞仪。
首先进行流式核型分析以确认需要分离的染色体大小,以4ags-ms双端体异附加系为实验组,中国春为对照。小麦染色体根据其大小差异,核型图通常形成4个独立的峰(vranaetal.,2000):ⅰ峰包含1d、4d、6d和7d共4条染色体;ⅱ峰包含1a、3a、6a、2d、3d和5d共6条染色体;ⅲ峰包含2a、4a、5a、7a、1b、2b、4b、5b、6b和7b共10条染色体;3b最大,独立成峰。对比中国春的核型图,可见4ags-ms双端体异附加系材料中除了标准核型图外,多出一个明显小的峰,根据相对荧光强度测算其大小在250mb左右,这个峰就是要分离的目标染色体(图2)。
经过多次试验,我们合计分离了200万条4ag染色体。以长穗偃麦草基因组dna为探针通过gish分析鉴定分离染色体的纯度为88%。
实施例3、4ag染色体的高通量测序和序列拼接
通过流式细胞仪分离出来的染色体是高度浓缩状态的染色质,不能直接应用于高通量测序,必须通过蛋白酶k消化处理将dna从染色体中分离出来。220万条4es染色体共纯化获得约500ngdna,完整性好。
以100ng4es基因组为模板,用qiagenrepl1-gsinglecellkit进行基因扩增,合计获得30μg扩增产物。将扩增产物进行酶切、克隆、测序,31个克隆中无来自大肠杆菌及人类污染,有1个克隆能够与小麦基因组序列完全匹配,余下克隆推测很可能来自长穗偃麦草4ag,比例为97%。
考虑到小麦族物种基因组都有重复序列比例高、拼接困难的特点(小麦中重复序列比例约为80%),因此我们采取了二代和三代测序相结合的策略。二代测序构建不同的插入片段库,分别为300bp、500bp、2kb和4kb种片段,双向2×125bp测序,测序量分别为20gb、14gb、6gb和6gb,合计测序46gb,数据量覆盖该条染色体184倍。三代测序构建10kb插入片段库,获取5~10kb读长,有效帮助整条染色体序列的拼接。
利用上述数据,首先通过platanus组装二代测序数据,获得了大小为212mb、n50为30kb的参考基因组序列。在此基础上利用sspacelr添加三代数据进行gapfilling,最终获得总大小为234m,n50为48k,scaffold数目为17302条的参考基因组(表1)。
表1、4ag参考基因组的拼接
实施例4、4ags-ms双端体异附加系的转录组测序
取4ags-ms双端体异附加系的单核期花药进行转录组测序,同时以兰州核不育突变体和相应亲本作为对照。illuminahi-seq2000双向2×100bp测序,每个样品各测约12g有效数据。对数据进行质量控制,分别是:去除接头序列和切除5’端前14bp序列、切除序列两端质量值的碱基、去除序列长度小于50bp的测序序列以及去除来自于人类、大肠杆菌、小麦线粒体和叶绿体基因组、禾本科核糖体rrna的污染,最后得到干净的测序数据。
实施例5、利用转录组和共线性获得候选基因
利用4ags-ms双端体异附加系转录组数据比对到17302条4agsscaffold上,并控制比对参数为100%一致性,得到2150个基因。也就是说4ags-ms双端体异附加系中4ags染色体上有2150个基因在单核期花药中表达。
由于流式细胞术分离4ags染色体纯度为88%,因此组装的4agsscaffold中会有少量来自小麦基因组的污染,根据要寻找的fertilityrestoregene1(frg1)基因位于4ags染色体上,只在4ags-ms双端体异附加系中表达,在对照材料兰州核不育突变体和相应亲本中均不表达的原则,去除在兰州核不育材料和/或相应亲本中也有表达的基因。由于表达偏高的有可能是重复序列、表达偏低的可信度差,因此去除4ags-ms双端体异附加系表达列表中10%与90%分位数的表达值以外的基因。经过上述两步筛选,候选基因从2150减少到374个基因。比对参数为100%一致性。
以iwgsc发布的小麦序列作为参考基因组,使用序列比对工具tophat2将兰州核不育材料和相应亲本的转录组数据分别比对到参考基因组上,最大允许2个错配,比对后统计每个locus中分别来自2个样品的reads数目,转换成rpkm,计算每个locus代表的基因在2个样品中的表达丰度差异,fdr值小于0.001的即被认定为具有显著性差异。结果显示亲本与兰州核不育突变体之间的差异显著:将每半条染色体均分成100份,计算每一份内所有基因中,在亲本中表达值显著高于兰州核不育材料的基因所占比例。结果发现,4b短臂远端2-7%区间内,几乎所有基因在亲本材料中均显著高表达,这一值显著高于整条染色体上24%的水平(图3)。由此我们推断,兰州核不育突变体是由于4bs染色体远端约2-7%缺失所导致。
由于长穗偃麦草的4ags染色体与小麦4bs染色体存在一定共线性,将374个基因全长(exon+intron)比对到tgac-4bs参考基因组上,有且仅有189个基因在4bs中有同源基因。比对工具为blastn,比对参数为blastn默认参数。其中2-7%区段内共有8个基因,这8个基因就是长穗偃麦草fertilityrestoregene(frg1)的候选基因。
这8个候选基因进行功能注释及在4ags-ms双端体异附加系及对照材料中的表达丰度信息见下表2。
表2、候选基因表达信息和功能注释
实施例6、转基因互补
将8个候选基因的基因组dna序列构建到pahc20载体上,利用基因枪法,转化4ag-ms单体异附加系植株幼胚。对t0代转基因植株中,不带有4ag染色体的植株(即兰州核不育纯合突变背景)进行花粉育性观察,结果只有连有基因id为cuff.199的基因组dna序列的载体能够恢复兰州核不育突变体雄性不育的表型,详细结果见下表3。因此,基因id为cuff.199所对应的基因为长穗偃麦草中的育性恢复基因(fertilityrestorationgene1,frg1),其基因组dna序列如seqidno:1所示,cds序列如seqidno:2所示,蛋白序列如seqidno:3所示,启动子序列如seqidno:4所示,终止子序列如seqidno:5所示。
表3、候选基因转基因t0代植株花粉育性观察
实施例6.frg1基因启动子表达载体的构建和功能分析
以4ags-ms双端体异附加系dna为模板,扩增frg1基因启动子2265bp,序列如seqidno:4所示,扩增产物通过in-fusion方法连入pahc20-gus载体,获得pahc20-pfrg1-gus表达载体。
利用基因枪法将pahc20-pfrg1-gus质粒转化小麦幼胚,获得16株转基因阳性植株。对转基因阳性植株的根、茎、叶和不同发育时期的花进行gus染色分析,发现frg1基因启动子能驱动gus在小麦花粉中特异表达,说明frg1基因启动子为花粉特异表达型启动子。
实施例7.frg1基因在新一代小麦杂交育种技术中的应用
frg1基因可以用于新一代杂交育种技术,该技术的核心思想是:以小麦隐性核雄性不育突变体为转化受体材料,通过将紧密连锁的3个目标基因转化至不育突变体中,其中,育性恢复基因可使转化受体育性恢复,花粉失活基因可使含有外源基因的花粉失活,即失去授精能力,种子标记基因可以用于转基因种子和非转基因种子的分拣,分拣出的非转基因种子即为不育系,而转基因种子用作保持系。也可以通过用保持系给不育系授粉杂交,可以在不育系上结实,由此繁殖不育系。而保持系通过自交可以源源不断地得以繁殖。由于该技术利用生物技术生产非转基因产品,解决了小麦杂交制种过程中面临的瓶颈问题,即三系法资源利用率低而两系法中不育系育性不稳定的问题。
本发明上述的杂交育种技术适用于兰州核雄性不育突变体及其等位突变体的繁殖和保持。根据以上原理,发明人首先分别对表达载体内的zmbt1-zmaa、frg1和mcherryw三个表达盒单独进行了小麦转化,并进一步对各个表达盒的功能进行了验证。结果表明各个表达盒单独转化小麦时,都能够工作良好,达到预期的设计效果。
进一步,发明人通过装配下述dna元件,构建了pahc20-frg1-aa-mcherryw载体:
1)以pahc20载体为基础;
2)frg1基因表达盒,目标基因frg1及其启动子和终止子均来自长穗偃麦草,其中frg1基因的启动子序列如seqidno:4所示,其终止子序列如seqidno:5所示,frg1基因的基因组dna序列如seqidno:1所示,其核苷酸序列编码的蛋白氨基酸序列如seqidno:3所示;
3)基因表达盒pg47:zmbt1-zmaa-in2-1,目标基因为zmaa,转运肽为zmbt1,zmbt1-zmaa(其核苷酸序列如seqidno:6所示)的开放读码框连接于启动子pg47(其核苷酸序列如seqidno:7所示)的下游、终止子in2-1(其核苷酸序列如seqidno:8所示)的上游。
4)基因表达盒camv35s增强子-ltp2:mcherryw-pinii,mcherryw基因(seqidno:9)的开放读码框连接于camv35s增强子-ltp2启动子(seqidno:10)和pinii终止子(seqidno:11)之间,重组成mcherryw的基因表达盒(camv35s增强子-ltp2:mcherryw-pinii);
小麦转化:利用基因枪法将质粒pahc20-frg1-aa-mcherryw转化4ag-ms单体异附加系植株幼胚,经过筛选、分化、壮苗生根、pcr鉴定等过程,得到基因型为兰州核不育位点纯合突变、无长穗偃麦草4ag染色体、且转基因为单拷贝的转基因阳性植株。
转基因小麦植株的花粉育性检测:对上述植株进行花粉活性检测。具体做法为:对转基因植株和非转基因植株各取一朵花,每朵花取1个花药,置于载玻片中央,滴加一滴1%的i2-ki溶液,用镊子和解剖针释放花粉后,盖上盖玻片,在显微镜下观察、计数不育花粉数和花粉总数,可以着色为深蓝色的为可育花粉,而不能够着色的为不育花粉。结果显示,在非转基因植株中,不育花粉的比例小于2%,而多个转基因植株中不育花粉的比例为50%左右,表明本发明所提供的载体能够达到预期的花粉失活功能。
转基因小麦植株的荧光种子与非荧光种子分离分析:对植株所结t1代种子进行荧光分离比例调查,结果表明这些种子均显示1:1分离比,即携带外源基因的荧光种子和不携带外源基因的非荧光种子表现为1:1分离,表明本发明所提供的载体各元件作为整体表达良好,可以实现创制和繁殖不育系的目的;其中,frg1基因可以恢复雄性不育突变体受体的育性zmbt1-zmaa基因和mcherryw基因的表达可以分别实现预期的花粉失活功能和种子荧光标记功能。
sequencelisting
<110>未名兴旺系统作物设计前沿实验室(北京)有限公司
北京大学
<120>小麦育性恢复基因及其应用
<130>
<160>11
<170>patentinversion3.3
<210>1
<211>1878
<212>dna
<213>长穗偃麦草(elytrigiaelongata)
<400>1
atggagagatcccggcgcctgctgctcgtggcgggcctgctggcggcgctgctcccggcg60
gcggcggccgcgttcgggccgcagccgggggcgccgtgcgagcccacgctgctggcgacg120
caggtggcgctcttctgcgcgcccgacatgcccaccgcgcagtgctgcgagcccgtcgtc180
gccgccgtcgacctcggcggcggcgtcccctgcctctgccgcgtcgccgcggagccgcag240
ctcgtcatggcgggcctcaacgccacccacctcctcacgctctacagctcctgcggcggc300
ctccgccccggcggcgcccacctcgccgccgcctgcgaaggtacgcaccgcacgtcgcct360
ccttccctccctccctctacgtgccgattttctgtgctcgctttcctgcttacctagtag420
tacgtagtttcccatggcttctcgagtcgctatagtgctccgatttgggtcggtttcctc480
gctgttctaccggatctgtcggcacggcgcgcggcgtcgggttctcgccgtctcccgtgg540
cgagcgacctgcgcagcgcgcgcacggcctagctagcttcttcataccgctgtaccttga600
gatatacggagcgatttagggtctactctgagtatttcgtcatcgtagaacgcatgtgcc660
acgcgcgattgtttcatcgattctagatctgtgtttgttcccgcgagttaagatggatct720
agcgccgtacgcagatgcggaggccttgcggtctctgtagctcgagttatcttatctact780
gtcgttcgactacagtatttgcctgcttccttttcactgggtttatcgtgcagtagtagt840
agccatgtccacgccttcttgttttgaggcgatcatcgtggacatacgtttcctgctaca900
gatttgaggagcactttgtttcaaactgcaacgcagctttgctttctgcagtatcttctg960
ccttgttttgttctgtgcagtacctcttgcttggtcaaaactgaaaacgcttgctgtttg1020
atcggcagagcaaaagcttgccgtgcttttcgctctgcagtgcatcgcctctgcgtcttt1080
tcccaaacatttccgcgttgatcctctggcggcactgcttttttgcatgcggtttccgta1140
gccttcctatttcgtgaaaaaaggttgggtcaaatcaaatggatcgcctattggcagagc1200
agcagcagcaacagatagctggctgtctcgcagctttggcagaatcggtctgtggccatc1260
tgtcccctgccaccgttttcctgatatttgtttctctcatctcatcttgcctgccactgt1320
gcttcttttcttgttgcgcacgtcgtcacctctactttctttccagatttgtttgctttt1380
gagatacggacgaacggctggtaattaactttggttgttgttgttactgtggattttgga1440
cgcaggacccgctcccccggccgccgtcgtcagcagccccccacccccgccaccgtccgc1500
cgcacctcgccgcaagcagccagcgcgtacgaaccctccctctctctctcgcctgcatct1560
cgctctgtttatctatcttcatatgttgatcagccttgtttacatactgacatgtgctct1620
ggatcggttttcgcagacgacgcaccgccaccgccgccgtcgaccgagaagccagcgtcc1680
ccgccgccccaggagcacgacggcgccgccccccgcgccaaggccgcccccgcccaggcg1740
gctacctccccgctcgcgcccgctgccgccaccgccccgccgccccaggcgccgcactcc1800
gccgcggcgacggcgtcgtccaaggcggccttcttcttcgtcgccacggccatgctcggc1860
ctctacatcatcctctga1878
<210>2
<211>663
<212>dna
<213>长穗偃麦草(elytrigiaelongata)
<400>2
atggagagatcccggcgcctgctgctcgtggcgggcctgctggcggcgctgctcccggcg60
gcggcggccgcgttcgggccgcagccgggggcgccgtgcgagcccacgctgctggcgacg120
caggtggcgctcttctgcgcgcccgacatgcccaccgcgcagtgctgcgagcccgtcgtc180
gccgccgtcgacctcggcggcggcgtcccctgcctctgccgcgtcgccgcggagccgcag240
ctcgtcatggcgggcctcaacgccacccacctcctcacgctctacagctcctgcggcggc300
ctccgccccggcggcgcccacctcgccgccgcctgcgaaggacccgctcccccggccgcc360
gtcgtcagcagccccccacccccgccaccgtccgccgcacctcgccgcaagcagccagcg420
cacgacgcaccgccaccgccgccgtcgaccgagaagccagcgtccccgccgccccaggag480
cacgacggcgccgccccccgcgccaaggccgcccccgcccaggcggctacctccccgctc540
gcgcccgctgccgccaccgccccgccgccccaggcgccgcactccgccgcggcgacggcg600
tcgtccaaggcggccttcttcttcgtcgccacggccatgctcggcctctacatcatcctc660
tga663
<210>3
<211>220
<212>prt
<213>长穗偃麦草(elytrigiaelongata)
<400>3
metgluargserargargleuleuleuvalalaglyleuleualaala
151015
leuleuproalaalaalaalaalapheglyproglnproglyalapro
202530
cysgluprothrleuleualathrglnvalalaleuphecysalapro
354045
aspmetprothralaglncyscysgluprovalvalalaalavalasp
505560
leuglyglyglyvalprocysleucysargvalalaalagluprogln
65707580
leuvalmetalaglyleuasnalathrhisleuleuthrleutyrser
859095
sercysglyglyleuargproglyglyalahisleualaalaalacys
100105110
gluglyproalaproproalaalavalvalserserpropropropro
115120125
proproseralaalaproargarglysglnproalahisaspalapro
130135140
proproproproserthrglulysproalaserproproproglnglu
145150155160
hisaspglyalaalaproargalalysalaalaproalaglnalaala
165170175
thrserproleualaproalaalaalathralaproproproglnala
180185190
prohisseralaalaalathralaserserlysalaalaphephephe
195200205
valalathralametleuglyleutyrileileleu
210215220
<210>4
<211>2265
<212>dna
<213>长穗偃麦草(elytrigiaelongata)
<400>4
gctttgcctacatactctagagaagttcaacccatgatccataagcaagctctcaattgc60
tgttttgagtgtcaaagatgtggtgtttgcaacatataaaatacgaagaaacctctcaac120
cacccttcctattttaggtacacatcacttgattcatggacaagtattgagaaacaacta180
tcacctagctctttcatcatgcgcctagtagtttctttggcacaagggttgatgaggtat240
gtttgtatatcttgagctatcattttattgttccttgaagcattcttgagaaccaccttg300
ttcacttcttcagaaatctctactagccattgtataagctcaaggaaattacctttatgc360
agtgaaacttctttccatcatgtccacgaaaagctaacccttgatgcaaaagaaaccttg420
gacattaaaaggaatatgtcaagccagctatatataaaaccttgtcttgtttggtggttg480
aaaaaatgactctatgattgttgtttttggtgcaatgaagaaattgtatttctcttgtgc540
tttgttgtgtgcactgttgattccaccaacatgtttcaaaaaactatccttgtgattcta600
atttctaaatcacccgttcacaaaaaatctccacaaacatgcaaattgtccttgaaaaga660
taacatacaagtagaaagcatcatcttttttactatactcaagccaactataaggcttaa720
accatttagctacaaatatcgatgcacacctccggtggggtgttgtggaaaagcatgtta780
ttttggccgataagccccttttacagtgtatcctcttctaattctattcagatcattaac840
atcagctgtgattgacatcctcttcccaagattagattcacgcaattgaacatcataaac900
cacatcttcaatgtcatcctcttcctatatattttcagattattggcttgcttcgttctc960
aatatcaggttctatgaatggacttctgttgatgccactaataatttgtagttgttgtgg1020
aatataaattgaacgggagctcatggtgctatgaacttgattggatgagaaattgttcta1080
cagctccacttgctgctcaacctaaatacatgcttggatttcttcccagctctagcacat1140
aaagttttcaaattaatgtttccaccacataaagttttgaaatccacaaatacttttttt1200
agtacatgaacatttttctaatatacggtgaagatttttcatatacaaactgatcgtttt1260
aatatatggtaaaaattggtgtaatatatgctgaaatgttttcaaatacatattgaatat1320
atttataataaatggtgagcatttctgtaatacatgatgaccattttaaaaatacatatc1380
gaacatttcataatatacgatgaacagttttataatactcgatgaacatattttggagtt1440
ctgaacaattttttttcaaaaacacaagccattttccaggaaaaaacaaaagcaaaagaa1500
atgagaaacccaaaaaacaaaaaagcaaagcagaacaaaacaaaacaaaacaaaacagag1560
aaatctacacggaaaatgaaacagaaaaagtaaagaaagaacccgaactgggccaggcaa1620
tgtttgcaacggcctcgctcttcctgaacaagaaggtcagtccatgggctgcttccagta1680
cacgggcctcgctgtgtggcaacacgccatgtaatagttttcgcgggaatccaacgtcgg1740
tctggtgacttctggtgcctaccagaactctccacaagctcccacaggacccacgcagcc1800
gtctgatcagatcagcacgaagcacgaactttgaagcgcggcgatattttctctccccag1860
cctccgcctcgcccgacgacgctgcgctgcagcaccatttcaatttcaaaatcaaaatcg1920
aaaacggaaaagctttctcgcatcccgaggagaggcggttacgcgcgccagaggagcacg1980
agagaggccaccccacccacccccgccctcacgtgccgcgctcgcaccccccgcggccgc2040
atccgggccgtccgcgcggacagctggccgcgccccacccgaaccgacgcccaggatcga2100
gcgagagcggcgcgcccgaggcttggcttagcgtccacggctcctccggctatataaccc2160
gcccgccacccgctccccctccggcattccattccgccaccgcacccccactccaccaca2220
aaccctagcgagcgagcgagagggagagaccgccccgccccgccg2265
<210>5
<211>628
<212>dna
<213>长穗偃麦草(elytrigiaelongata)
<400>5
gtggccgaccccgaggccatggtccgtccagttgcagtagagtgctcgtcgtcttgttcc60
gtttcatgcttgtcgccatttgaggttcgtttctgcagtccggtcgaagaagacggtgga120
ttttgagtagtagtagtagctatcgttggcaggagtatggagttcatgtgtcctcggtcg180
cctagttttggtctcaagtagtactgtctgtccgccgtgtttgggtgtttgcgattcctc240
tggttagatgaaccactgctatgtgatcgatcgatatgatctgaatggaatggatcaagt300
tttgcgttctgctgatgatgtgatatgcttcttcgtatatactcatgctcgacctatcta360
tgttctcccatttgaatttgtggagcaacagtttggcatgcttttgctctgctatgaatg420
aatgcttcttgcatgcatcttgtctttgcttaatttgaactgtagaacggatgcagttct480
gatttcttcatatgcatatgctctgtatgtgttcatctcttcgaatttattatgtagcaa540
cagtttgtagcttttcattctgctatgaatgaatgcctcttgcatgttgtctttgcttaa600
tttgtactagcagaatgcagaccttgcg628
<210>6
<211>1487
<212>dna
<213>人工合成
<400>6
atggcggcgacaatggcagtgacgacgatggtgacgaggagcaaggagagctggtcgtca60
ttgcaggtcccggcggtggcattcccttggaagccacgaggtggcaagaccggcggcctc120
gagttccctcgccgggcgatgttcgccagcgtcggcctcaacgtgtgcccgggcgtcccg180
gcggggcgcgacccgcgggagcccgatcccaaggtcgtccgggcggcctgcggcctggtc240
caggcacaagtcctcttccaggggtttaactgggagtcgtgcaagcagcagggaggctgg300
tacaacaggctcaaggcccaggtcgacgacatcgccaaggccggcgtcacgcacgtctgg360
ctgcctccaccctcgcactccgtctcgccacaaggctacatgccaggccgcctatacgac420
ctggacgcgtccaagtacggcacggcggcggagctcaagtccctgatagcggcgttccac480
ggcaggggcgtgcagtgcgtggcggacatcgtcatcaaccaccggtgcgcggaaaagaag540
gacgcgcgcggcgtgtactgcatcttcgagggcgggactcccgacgaccgcctggactgg600
ggccccgggatgatctgcagcgacgacacgcagtactcggacgggacggggcaccgcgac660
cgggcgaggggttcgcggcggcgcccgacatcgaccacctcaacccgcgcgtgcagcggg720
agctctccgcctggctcaactggctcaggtccgacgccgtggggttcgacggctggcgcc780
tcgacttcgccaagggctactcgccggccgtcgccagaatgtacgtggagagcacggggc840
cgccgagcttcgtcgtcgcggagatatggaactcgctgagctacagcggggacggcaagc900
cggcgcccaaccaggaccagtgccggcaggagctgctggactggacgcgggccgtcggcg960
ggcccgccatggcgttcgacttccccaccaagggcctgctgcaggcgggcgtgcaggggg1020
agctgtggcggctgcgcgacagctccggcaacgcggccggcctgatcgggtgggcgcccg1080
agaaggccgtcaccttcgtcgacaaccatgacaccgggtcgacgcagaagctctggccgt1140
tcccatccgacaaggtcatgcagggctacgcctacatcctcacccatccaggagtcccct1200
gcattttctacgaccacatgttcgactggaacctgaagcaggagatatccacgctgtctg1260
ccatcagggcgcggaacggcatccgcgccgggagcaagctgcggatcctcgtggcggacg1320
cggacgcgtacgtggccgtcgtcgacgagaaggtcatggtgaagatcgggacaaggtacg1380
gcgtgagcagcgtggtcccgtcggatttccacccggcggcgcacggcaaggactactgcg1440
tctgggagaaagcgagcctccgcgtcccggcggggcgccacctctag1487
<210>7
<211>2736
<212>dna
<213>玉米(zeamays)
<400>7
tgcaccggacactgtctggtggcataccagacagtccggtgtgccagatcagggcaccct60
tcggttcctttgctcctttgcttttgaaccctaactttgatcgtttattggtttgtgttg120
aacctttatgcacctgtggaatatataatctagaacaaactagttagtccaatcatttgt180
gttgggcattcaaccaccaaaattatttataggaaaaggttaaaccttatttccctttca240
atctccccctttttggtgattgatgccaacacaaaccaaagaaaatatataagtgcagaa300
ttgaactagtttgcataaggtaagtgcataggttacttagaattaaatcaatttatactt360
ttacttgatatgcatggttgctttcttttattttaacattttggaccacatttgcaccac420
ttgttttgttttttgcaaatctttttggaaattctttttcaaagtcttttgcaaatagtc480
aaaggtatatgaataagattgtaagaagcattttcaagatttgaaatttctccccctgtt540
tcaaatgcttttcctttgactaaacaaaactccccctgaataaaattctcctcttagctt600
tcaagagggttttaaatagatatcaattggaaatatatttagatgctaattttgaaaata660
taccaattgaaaatcaacataccaatttgaaattaaacataccaatttaaaaaatttcaa720
aaagtggtggtgcggtccttttgctttgggcttaatatttctccccctttggcattaatc780
gccaaaaacggagactttgtgagccatttatactttctccccattggtaaatgaaatatg840
agtgaaagattataccaaatttggacagtgatgcggagtgacggcgaaggataaacgata900
ccgttagagtggagtggaagccttgtcttcgccgaagactccatttccctttcaatctac960
gacttagcatagaaatacacttgaaaacacattagtcgtagccacgaaagagatatgatc1020
aaaggtatacaaatgagctatgtgtgtaatgtttcaatcaaagtttcgagaatcaagaat1080
atttagctcattcctaagtttgctaaaggttttatcatctaatggtttggtaaagatatc1140
gactaattgttctttggtgctaacataagcaatctcgatatcacccctttgttggtgatc1200
cctcaaaaagtgataccgaatgtctatgtgcttagtgcggctgtgttcaacgggattatc1260
cgccatgcagatagcactctcattgtcacataggagagggactttgctcaatttgtagcc1320
atagtccctaaggttttgcctcatccaaagtaattgcacacaacaatgtcctgcggcaat1380
atacttggcttcggcggtagaaagagctattgagttttgtttctttgaagtccaagacac1440
cagggatctccctagaaactgacaagtccctgatgtgctcttcctatcaattttacaccc1500
tgcccaatcggcatctgaatatcctattaaatcaaaggtggatcccttggggtaccaaag1560
accaaatttaggagtgtaaactaaatatctcatgattcttttcacggccctaaggtgaac1620
ttccttaggatcggcttggaatcttgcacacatgcatatagaaagcatactatctggtcg1680
agatgcacataaatagagtaaagatcctatcatcgaccggtataccttttggtctacgga1740
tttacctcccgtgtcgaggtcgagatgcccattagttcccatgggtgtcctgatgggctt1800
ggcatccttcattccaaacttgttgagtatgtcttgaatgtactttgtttggctgatgaa1860
ggtgccatcttggagttgcttgacttgaaatcctagaaaatatttcaacttccccatcat1920
agacatctcgaatttcggaatcatgatcctactaaactcttcacaagtagatttgttagt1980
agacccaaatataatatcatcaacataaatttggcatacaaacaaaacttttgaaatggt2040
tttagtaaagagagtaggatcggctttactgactctgaagccattagtgataagaaaatc2100
tcttaggcattcataccatgctgttggggcttgcttgagcccataaagcgcctttgagag2160
tttataaacatggttagggtactcactatcttcaaagccgagaggttgctcaacatagac2220
ctattcaccccatttgatcacttttttggtccttcaggatctaatagttatgtataattt2280
agagtctcttgtttaatggccagatatttctaattaatctaagaatttatgatatttttt2340
aattttttatcatgtctgatgagaattaacataaaggctcaattgggtcctgaattaata2400
atagagtgaaaattaatccagaggctctattagaaccttcaattagtaataccaagatat2460
atataagatagtagagtatagtttaaatgttggcattgttcattctttcttttgttattt2520
aatttatgctttccacggtggttagtggttacttctgaagggtccaaataatgcatgaag2580
agtttgaggacaagaagtctgccctaaaaatagcgatgcaaaggcatggtgtccaagcca2640
tacatatagcgcactaattttatcagcagaacaatggtatttataggtcctagtgcccag2700
gcaacaagagacacgaataaagcatcgatcacgaca2736
<210>8
<211>344
<212>dna
<213>玉米(zeamays)
<400>8
tgacaaagcagcattagtccgttgatcggtggaagaccactcgtcagtgttgagttgaat60
gtttgatcaataaaatacggcaatgctgtaagggttgttttttatgccattgataataca120
ctgtactgttcagttgttgaactctatttcttagccatgccaagtgcttttcttattttg180
aataacattacagcaaaaagttgaaagacaaaaaaaaaaacccccgaacagagtgctttg240
ggtcccaagctactttagactgtgttcggcgttccccctaaatttctccccctatatctc300
actcacttgtcacatcagcgttctctttcccctatatctccacg344
<210>9
<211>711
<212>dna
<213>人工合成
<400>9
atggtgtccaagggcgaggaggacaacatggccatcatcaaggagttcatgcgcttcaag60
gtgcacatggagggctccgtgaacggccacgagttcgagatcgagggcgagggcgagggc120
cgcccctacgagggcacccaaaccgccaagctcaaggtgaccaagggtggccccctcccc180
ttcgcctgggacatcctctccccacaattcatgtacggctccaaggcctacgtgaagcac240
cccgccgacatccccgactacctcaagctctccttccccgagggcttcaagtgggagcgc300
gtgatgaacttcgaggacggcggcgtggtgaccgtgacccaagactcctccctccaagac360
ggcgagttcatctacaaggtgaagctccgcggcaccaacttcccctccgacggccccgta420
atgcaaaagaagaccatgggctgggaggcctcctccgagcggatgtaccccgaggacggc480
gccctcaagggcgagatcaagcaaaggctcaagctcaaggacggcggccactacgacgcc540
gaggtgaagaccacctacaaggccaagaagcccgtgcaactccccggcgcctacaacgtg600
aacatcaagctcgacatcacctcccacaacgaggactacaccatcgtggagcaatacgag660
cgcgccgagggccgccactccaccggcggcatggacgagctgtacaagtga711
<210>10
<211>1149
<212>dna
<213>人工合成
<400>10
cgtcaacatggtggagcacgacacgcttgtctactccaaaaatatcaaagatacagtctc60
agaagaccaaagggcaattgagacttttcaacaaagggtaatatccggaaacctcctcgg120
attccattgcccagctatctgtcactttattgtgaagatagtggaaaaggaaggtggctc180
ctacaaatgccatcattgcgataaaggaaaggccatcgttgaagatgcctctgccgacag240
tggtcccaaagatggacccccacccacgaggagcatcgtggaaaaagaagacgttccaac300
cacgtcttcaaagcaagtggattgatgtgatgaattcaaccgtctcttcgtgagaataac360
cgtggcctaaaaataagccgatgaggataaataaaatgtggtggtacagtacttcaagag420
gtttactcatcaagaggatgcttttccgatgagctctagtagtacatcggacctcacata480
cctccattgtggtgaaatattttgtgctcatttagtgatgggtaaattttgtttatgtca540
ctctaggttttgacatttcagttttgccactcttaggttttgacaaataatttccattcc600
gcggcaaaagcaaaacaattttattttacttttaccactcttagctttcacaatgtatca660
caaatgccactctagaaattctgtttatgccacagaatgtgaaaaaaaacactcacttat720
ttgaagccaaggtgttcatggcatggaaatgtgacataaagtaacgttcgtgtataagaa780
aaaattgtactcctcgtaacaagagacggaaacatcatgagacaatcgcgtttggaaggc840
tttgcatcacctttggatgatgcgcatgaatggagtcgtctgcttgctagccttcgccta900
ccgcccactgagtccgggcggcaactaccatcggcgaacgacccagctgacctctaccga960
ccggacttgaatgcgctaccttcgtcagcgacgatggccgcgtacgctggcgacgtgccc1020
ccgcatgcatggcggcacatggcgagctcagaccgtgcgtggctggctacaaatacgtac1080
cccgtgagtgccctagctagaaacttacacctgcaactgcgagagcgagcgtgtgagtgt1140
agccgagta1149
<210>11
<211>310
<212>dna
<213>马铃薯(solanumtuberosum)
<400>11
ctagacttgtccatcttctggattggccaacttaattaatgtatgaaataaaaggatgca60
cacatagtgacatgctaatcactataatgtgggcatcaaagttgtgtgttatgtgtaatt120
actagttatctgaataaaagagaaagagatcatccatatttcttatcctaaatgaatgtc180
acgtgtctttataattctttgatgaaccagatgcatttcattaaccaaatccatatacat240
ataaatattaatcatatataattaatatcaattgggttagcaaaacaaatctagtctagg300
tgtgttttgc310
- 还没有人留言评论。精彩留言会获得点赞!