氨基酸序列在用于使昆虫Dip3蛋白失活中的应用的制作方法
本申请涉及氨基酸序列在用于使昆虫Dip3蛋白失活中的应用。
背景技术:
斜纹夜蛾属于鳞翅目夜蛾科,广泛分布在世界各地,在我国各地均有发生,是典型的杂食性昆虫。已知可以危害的植物达99科290多种,包括棉花、烟草和蔬菜等农作物。由于斜纹夜蛾繁殖能力强,且易对农药产生抗药性,近年来,对蔬菜和花卉的危害越来越严重,防治难度也越来越大。目前,主要依靠化学农药来防治,但化学农药大量和长期使用,一方面害虫已经产生了耐药性;另一方面,带来了环境污染、水质污染及人体健康等一系列问题。需求有效的替代防治技术迫在眉睫。
伴随着分子生物学的高速发展,将分子生物学技术应用到害虫防治中成为可能。
分子防治具有如下优点:减少农药的使用,避免环境污染,维持生态平衡,提高农作物产品质量,并且不会产生抗药性。
但是,如何将分子生物学技术应用到害虫防治中,则需要确定具体可应用的蛋白和基因;以及进一步的能够以其作为靶标并通过与其结合等行为来失活该靶标蛋白或基因的物质。
dip3作为斜纹夜蛾天然免疫信号通路NF-κB的辅助因子之一,参与调控了斜纹夜蛾抗菌肽等下游免疫基因,是防治斜纹夜蛾的一个重要靶标蛋白。双斑侧沟茧蜂是斜纹夜蛾的自然宿主,它带有的双斑侧沟茧蜂病毒具有高丰度的Vank基因,特别是Vank86、Vank92和 Vank101,其表达产物能与斜纹夜蛾的dip3蛋白结合形成异源二聚体,使dip3蛋白失活,从而使斜纹夜蛾产生免疫抑制,最终使斜纹夜蛾死亡,达到生物防治的目的。然而dip3受到抑制后,斜纹夜蛾个体水平上的头壳宽度和体长无明显变化,抑制效应只能通过观察斜纹夜蛾血淋巴粒细胞凋亡来检测。
技术实现要素:
本申请之一提供了一种氨基酸序列在用于使昆虫Dip3蛋白失活中的应用,其中,所述氨基酸序列为如(I)至(III)中的至少一种;
(I)所述氨基酸序列如1)或2)所示:
1)SEQ ID No.10所示的氨基酸序列;
2)与1)中的氨基酸序列的一致性在95%以上,且与1)中的氨基酸序列具有相同功能的氨基酸序列;优选其氨基酸序列为与1)中的氨基酸序列的一致性在99%以上,且与1) 中的氨基酸序列具有相同功能的氨基酸序列;
(II)所述氨基酸序列如3)或4)所示:
3)SEQ ID No.12所示的氨基酸序列;
4)与3)中的氨基酸序列的一致性在95%以上,且与3)中的氨基酸序列具有相同功能的氨基酸序列;优选其氨基酸序列为与3)中的氨基酸序列的一致性在99%以上,且与3) 中的氨基酸序列具有相同功能的氨基酸序列;
(III)所述氨基酸序列如5)或6)所示:
5)SEQ ID No.14所示的氨基酸序列;
6)与5)中的氨基酸序列的一致性在95%以上,且与5)中的氨基酸序列具有相同功能的氨基酸序列;优选其氨基酸序列为与5)中的氨基酸序列的一致性在99%以上,且与5) 中的氨基酸序列具有相同功能的氨基酸序列。
本申请之二提供了一种能够编码如本申请之一所述的氨基酸序列的核苷酸序列在用于使昆虫Dip3蛋白失活中的应用。
在一个具体实施方式中,编码如SEQ ID No.10所示的氨基酸序列的核苷酸序列如SEQ ID No.9所示;和/或编码如SEQ ID No.12所示的氨基酸序列的核苷酸序列如SEQ ID No.11 所示;和/或编码如SEQ ID No.14所示的氨基酸序列的核苷酸序列如SEQ ID No.13所示。
本申请之三提供了一种转基因微生物,其含有如本申请之二所述的应用中的所述核苷酸序列,且能产生编码如本申请之一所述的应用中的所述氨基酸序列。
在一个具体实施方式中,所述转基因微生物包括芽胞杆菌(Bacillus)、假单胞菌 (Pseudomonas)、肠杆菌(Escherichia)和酵母(Saccharomyces)中的至少一种。
在一个具体实施方式中,优选所述芽胞杆菌包括苏云金芽胞杆菌(Bacillus thuringiensis)、枯草芽胞杆菌(Bacillus subtilis)、萎缩芽胞杆菌(Bacillus atrophaeus)和蜡样芽胞杆菌(Bacillus cereus)中的至少一种;所述假单胞菌包括荧光假单胞杆菌 (Pseudomonas fluorescens);所述肠杆菌包括大肠杆菌(Escherichia coli)。
本申请之四提供了根据本申请之三所述的转基因微生物在用于使昆虫Dip3蛋白失活中的应用。
在一个具体实施方式中,所述应用用于害虫防治。
在一个具体实施方式中,优选所述应用用于斜纹夜蛾属(Spodoptera)害虫的防治。
在一个具体实施方式中,更优选所述应用用于斜纹夜蛾(Spodoptera litura Fabricius)害虫的防治。
本申请之五提供了根据本申请之二所述的应用中的所述核苷酸序列在制备转基因植物中的应用,并且制备得到的所述转基因植物能够产生编码如本申请之一所述的应用中的所述氨基酸序列。
在一个具体实施方式中,所述转基因植物包括禾本科(Liliaceae)、豆科 (Leguminosae)、藜科(Chenopodiaceae)、菊科(Compositae)、和锦葵科(Malvaceae) 中的至少一种。
在一个具体实施方式中,优选所述转基因植物包括蜀黍属(Zea)、高梁属 (Sorghum)、狗尾巴草属(Setaria)、稻属(Oryza)、甘蔗属(Saccharum)、小麦属 (Triticum)、大豆属(Glycine)、苜蓿属(Medicago)、甜菜属(Beta)、向日葵属 (Helianthus)和棉属(Gossypium)中的至少一种。
在一个具体实施方式中,优选所述转基因植物包括玉米(Zea mays L.)、高粱 (Sorghum bicolor(L.)Moench)、谷子(Setaria italica)、水稻(Oryza sativa L.)、甘蔗 (Saccharum officinarum)、小麦(Triticum aestivum)、大豆(Glycine max(L.)Merr)、苜蓿(Medicago sativa Linn)、甜菜(Beta vulgaris)、向日葵(Helianthus)、亚洲棉 (Gossypium arboreum L.)、非洲棉(Gossypium herbaceum L.)、陆地棉(Gossypium hirsutum L.)和海岛棉(Gossypium barbadense L.)中的至少一种。
在一个具体实施方式中,所述应用用于害虫防治。
在一个具体实施方式中,优选所述应用用于斜纹夜蛾属(Spodoptera)害虫的防治。
在一个具体实施方式中,更优选所述应用用于斜纹夜蛾(Spodoptera litura Fabricius)害虫的防治。
本申请的有益效果:
一方面,发明人在研究过程中首次发现了本申请的dip3转录水平的高低直接影响着 Innexin2的转录,从而确定了dip3是NF-κB信号通路中的一个重要辅助因子,同时也确定了Innexin2是dip3下游的靶基因之一。在此基础之上,还可以通过类似的方法研究NF-κB 信号通路中其他基因的转录表达与dip3转录的关系。因此,本申请为研究dip3调节其相关下游基因的关系奠定了基础。
另一方面,发明人在研究过程中首次发现了缺失或干扰例如斜纹夜蛾幼虫体内的Dip3 会导致血淋巴细胞凋亡。可见Dip3可以作为斜纹夜蛾在分子水平上的攻击靶标位点。如果将这一惊奇的发现与害虫的防治相结合,即将这一发现应用于害虫防治中,那么无疑会减少农药的使用,避免环境污染,维持生态平衡,提高农作物产品质量,并且害虫也不会产生抗药性。这将是一种非常有应用前景的害虫防治技术。
幸运的是,发明人还找到了通过与Dip3蛋白结合而使Dip3失活的三种蛋白,即 Vank86、Vank92、Vank101,这三种蛋白来源于与斜纹夜蛾的天敌双斑侧沟茧蜂共生的茧蜂病毒。由于Dip3的缺失会导致血淋巴细胞凋亡,因此,Dip蛋白的失活,可以使昆虫细胞凋亡。这也是首次从分子水平上确定了双斑侧沟茧蜂病毒可使斜纹夜蛾致死的机理,即 Vank86、Vank92、Vank101三个蛋白是对斜纹夜蛾致死最为有效的蛋白之一。因此,通过使用这些蛋白中的至少一种可以导致如斜纹夜蛾幼虫的血淋巴细胞凋亡,达到生物或分子防治的目的。
附图说明
图1显示了对斜纹夜蛾幼虫进行dsRNA Dip3RNAi饲喂六天后,对Dip3沉默效果检测和血淋巴细胞凋亡检测及早晚期凋亡细胞数目的统计。
图1A为RT-PCR检测Dip3dsRNA饲喂后干扰效果。M为DNA Marker,Control(1泳道)为阴性对照,即不做任何处理,阳性对照为含有HT115菌株(2泳道)和转化了L4440 载体的HT115菌株(3泳道),实验组为含有连接成功的干扰载体L4440-dip3转化的 HT115菌株(4泳道)。根据RNAi饲喂方法,在幼虫一经孵出,喂养特制饲料6天,提取虫体RNA,进行RT-PCR,结果如图所示。由图可知:Dip3基因被有效沉默;
图1B为Annexin V-FITC/PI凋亡检测试剂盒检测Dip3dsRNA(ds Dip3)饲喂后斜纹夜蛾血淋巴细胞的凋亡情况。Annexin V-FITC检测早期凋亡细胞,呈浅灰色,例如图中的细胞 a和细胞c;PI检测晚期凋亡细胞,用斜网格线表示,例如细胞b和细胞d(Bar=10μm)。图中,根据t检验分析的结果,***表示在p<0.001时,差异显著。由图可知:比起阴性对照组(Ctrl RNAi),饲喂实验组(Dip3RNAi)中,斜纹夜蛾血淋巴凋亡细胞显著增加,包括早期凋亡细胞及晚期凋亡细胞。
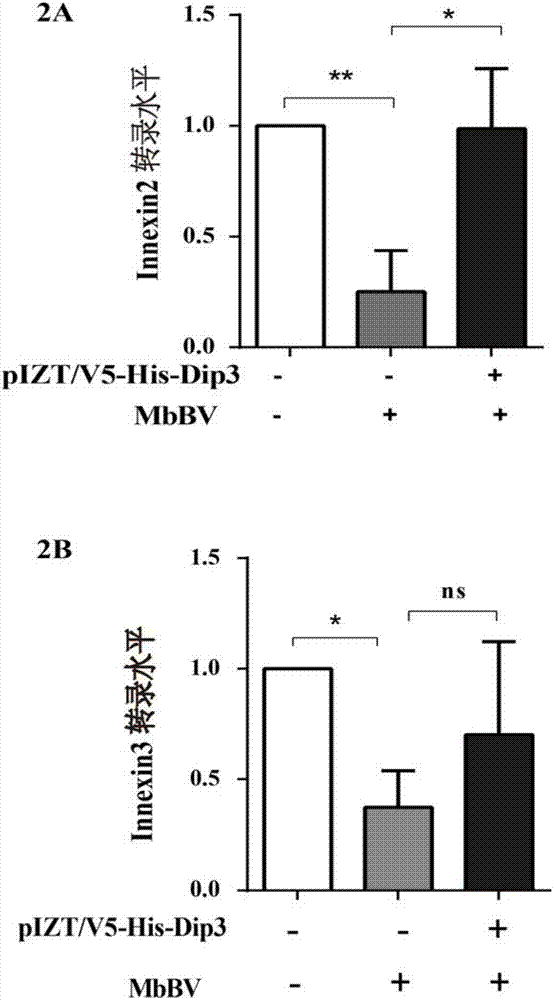
图2显示了qRT-PCR检测Innexin转录水平的结果。
图2A为qRT-PCR检测Innexin2转录情况,根据t检验分析的结果,*表示p<0.05时,差异显著,**表示p<0.01时,差异非常显著。由图可知,在斜纹夜蛾的本体细胞Spli221 中,在受到MbBV病毒刺激时,Innexin2转录受到显著抑制,而Dip3的过表达可以挽救 Innexin2的转录,即提高其转录效率。
图2B为qRT-PCR检测Innexin3转录情况,根据t检验分析的结果,*表示p<0.05时,差异显著,ns表示无差异。由图可知,在斜纹夜蛾的本体细胞Spli221中,在受到MbBV病毒刺激时,Innexin3转录受到显著抑制,而Dip3的过表达不能挽救Innexin3的转录。
图3为Western Blot检测pIZT/V5-His-Vank86、pIZT/V5-His-Vank92和pIZT/V5-His- Vank101分别导入斜纹夜蛾细胞后Vank86,Vank92和Vank101基因的表达情况。M为蛋白 Marker,Ctrl为阴性对照,V5为导入pIZT/His-V5空载体对照,V5-Vank86为导入pIZT/V5- His-Vank86的实验组,V5-Vank92为导入pIZT/V5-His-Vank92的实验组,V5-Vank101导入 pIZT/V5-His-Vank101的实验组,Anti-V5为所使用的抗体,Tubulin为蛋白内参。由图3A 可知:比起阴性对照组,实验组中,Vank86和Vank92蛋白在斜纹夜蛾细胞中表达;同理,图3B显示Vank101在斜纹夜蛾细胞中表达。
图4为斜纹夜蛾Dip3蛋白受到Vank86,Vank92和Vank101蛋白结合的影响。图中, M为蛋白Marker,pIZT/His-V5为使用的空白载体名称,Anti-V5和Flag(碧云天)为所使用的抗体,Tubulin为蛋白内参。由图可知:斜纹夜蛾Dip3蛋白被Vank86(图4A)和 Vank92(图4B)以及Vank101(图4C)结合形成异源复合体。
具体实施方式
下面结合具体实施例对本发明作进一步说明,但以下实施例并不够成对本申请的限制,凡包括在本申请的精神范围内的技术方案,均在本申请的保护范围内。
实施例1
1.Dip3特异引物设计
从斜纹夜蛾转录组数据(Li M,Pang Z,Xiao W,Liu X,Zhang Y,et al.(2014)A Transcriptome Analysis Suggests Apoptosis-Related Signaling Pathways in Hemocytes of Spodoptera litura After Parasitization by Microplitis bicoloratus.PLoS ONE 9(10):e110967. doi:10.1371/journal.pone.0110967)中选取Dip3基因序列,Dip3基因序列(如SEQ ID No.2 所示)大小为750bp,共编码249个氨基酸(SEQ ID No.1)。采用引物设计软件Primer Premier 5设计特异引物:上游引物pIZT/V5-His-Dip3-F(SEQ ID No.3)和下游引物 pIZT/V5-His-Dip3-R(SEQ ID No.4)。
2.斜纹夜蛾幼虫总RNA的提取及cDNA的合成
本实验采用实验室方案提取斜纹夜蛾幼虫总RNA,具体步骤如下:
1)取10头3日龄幼虫,放置于加入少量RNAisoTM Plus裂解液的EP管中,充分捣碎研磨,继续加入RNAisoTM Plus裂解液至总体积达到1ml,涡旋混匀后,室温静置6min;
2)将上述处理好的样品,12000rpm,4℃离心5min;
3)取上清,转移至新的EP管中,加入氯仿200μL,剧烈振荡10-15s,充分混匀后,室温静置5min;
4)12000rpm,4℃离心15min;离心后的溶液分为三层:无色的上清液、中间的白色蛋白层及带有颜色的下层有机相,吸取上清液转移至另一新的离心管中;
5)向上清中加入等体积的异丙醇,混匀后室温静置10min;12000rpm,4℃离心10 min;轻轻弃去上清,缓慢地沿离心管壁加入75%的乙醇l ml(0.1%DEPC水配制);12000 rpm,4℃离心5min;
6)弃去乙醇,室温干燥5min,加入50μL的RNase-free H2O溶解沉淀,标记好后,- 80℃保存,取5μL马上进行琼脂糖凝胶电泳检测。
本实验用PrimeScriptTM RT reagent Kit反转录试剂盒(货号6210A)来反转录,具体步骤如下:
1)在离心管(EP管)中配置下列反应混合液(体系10μL),其中,Oligo dT primer 1 μL,dNTP mixture 1μL,模板RNA 8μL,65℃,5min,冰上骤冷;
2)在上述EP管中加入下列反转录反应液:5×PrimeScrip Buffer 4μL,PrimeScrip Reverse Transcriptase(200U/μL)4μL,Ribonuclease Inhibitor(40U/μL)2μL,总体积15 μL,45℃保温45min。然后,95℃反应5min,冰上骤冷;-80℃保存。
3.Dip3基因片段的克隆
1)以上述所得的cDNA为模板,用Dip3特异引物进行PCR扩增。
2)将上述PCR产物进行电泳得到Dip3的基因片段为750bp。将PCR产物回收,并连接到克隆载体pMD19-T载体上,经测序确定正确,得到含有靶标片段的pMD19-Dip3质粒。
4.干扰载体L4440-dip3的构建
取测序成功的pMD19-Dip3菌液,加A+过夜培养,用普通质粒提取试剂盒提取质粒,根据的相应酶切位点对质粒进行双酶切(XbaI/HindIII),用同样的限制性内切酶对干扰载体L4440进设计行双酶切。将Dip3基因片段和L4440载体片段进行回收,T4连接酶连接。转化HT115菌株,鉴定成功,得到L4440-Dip3。
5.Dip3 dsRNA饲喂
(1)L4440-Dip3干扰菌株在含A+浓度为100μg/mL和T+浓度为10μg/mL的液体培养基160ml中摇菌至OD值为0.4,加IPTG至终浓度为0.4mM/mL,离心获取含有Dip3 dsRNA的菌液沉淀,加入5ml蒸馏水悬浮;同时阳性对照只含有HT115菌株的菌液和只含有转化了L4440的HT115菌株的菌液也做相同的处理。
(2)配制饲料
A.营养物质(1份的量):黄豆粉5.0g、麦麸5.0g、酵母提取物2.0g、琼脂粉1.0g、干酪素1.0g。准确称量上述营养物,然后装入200mL的保鲜盒中,在本次实验中制备4 份。
B.混合粉末(1份的量):尼泊金甲酯0.125g、山梨酸0.125g、抗坏血酸0.2g、胆固醇0.5g、氯化胆碱0.0425g、六合维生素丸(碾碎)半粒。准确称量上述试剂,然后装于 50ml的离心管中混合均匀,最后分装于1.5mL的离心管中(每管0.68g),在本次实验中制备4份。
将每一份A中饲料经过120℃30分钟高温灭菌后,降温至50℃后分别加入一份B中的混合粉末和上述菌液,搅拌混匀后便得到需要的饲料了。以不加入任何菌液的A和B的混合物——饲料作为阴性对照。
(3)取适量已消毒的卵片,放入标记好的培养器具中,置于温度为27℃,湿度为60%的养虫室。设置对照组和处理组,其中阴性对照组为Control,用不添加任何菌液的饲料饲喂;阳性对照组为HT115和HT115-L4440,即加入HT115菌液,和转化了L4440的HT115 的菌液经过dsRNA诱导的菌液;处理组为RNAi Dip3,即加入含有Dip3dsRNA的HT115 菌液。幼虫一经孵出,便用对应的饲料饲喂,每天进行新饲料与旧饲料的更换。
(4)饲喂六天后,提取幼虫血淋巴中的RNA,逆转录cDNA,方法如实施例1中的2 部分。进行RT-PCR。其中:
1)PCR体系:上游引物0.5μl、下游引物0.5μl、cDNA 1.0μl、Ex Taq Mix 7.5μl、最后加入dd H2O至终体积15μl。
2)PCR程序:94℃ 5min;94℃ 30s;58℃ 30s;30个循环;72℃ 1min;72℃ 10min;
结果如图1A所示,与对照组相比,Dip3基因被有效沉默。
(5)饲喂六天后,取一部份幼虫用来凋亡细胞检测,具体操作:取10μl(约1×104个细胞)血淋巴,加入100μl 1×结合缓冲液悬浮细胞;加入5μl Annexin V-FITC和5μl PI的染色液,轻轻混匀;避光、室温反应10min;加入400μl 1×结合缓冲液,混匀,加入24孔细胞培养板贴壁约15min,样品在1h内用荧光显微镜检测。结果如图1B所示,由图可以看出:比起对照组(Control)Dip3dsRNA饲喂实验组中,斜纹夜蛾血淋巴凋亡细胞显著增加,包括早期凋亡细胞及晚期凋亡细胞。
6.真核表达质粒pIZT/V5-His-Dip3质粒构建
1)真核表达质粒pIZT/V5-His-Dip3的获取
取测序成功的pMD19-Dip3菌液,加A+过夜培养,用普通质粒提取试剂盒提取质粒,根据设计的相应酶切位点对质粒进行双酶切(Sal I/EcoRV),用同样的限制性内切酶对载体pIZT/V5-His进行双酶切。将Dip3基因片段和pIZT/V5-His载体片段进行回收,T4连接酶连接。转化真核表达菌株Trans5α,鉴定成功,得到含有Dip3核酸序列的pIZT/V5-His-Dip3真核菌株。
2)过表达Dip3
准备三组生长状态良好的Spli221细胞,两组用MbBV病毒刺激,1×106个细胞/一只病毒当量,24h后,选取一组病毒刺激过的细胞转染pIZT/V5-His-Dip3质粒,转染方法如下:
(1)当细胞生长密度达到80%-90%时,移除旧培养基,加入新的培养基,用枪头轻轻吹打悬浮细胞,台盼蓝计数;
(2)将计数后的细胞27℃贴壁培养2h以上直至细胞贴壁、状态良好;
(3)配制转染试剂复合物,吸取100μL的双无培养基置于500μL EP管中,加入1μg 的质粒,取100μL的双无培养基加入5ul转染试剂Cell fectionⅡ,震荡混匀,两者混匀后室温静置45min,每15分钟震荡几下,用双无培养基清洗细胞30min,将加入800ul双无培养基的转染混合物逐滴加入到相应孔中,放到摇床上轻轻晃动,使转染复合物在12孔板中混合均匀,27℃继续培养,4h-5h吸出转染混合液,加入有血清培养基。
7.提取RNA和qRT-PCR
转染pIZT/V5-His-Dip3质粒到Spli221细胞72h后,分别提取未处理Spli221细胞、感染 MbBV的Spli221细胞以及在感染MbBV的基础上转染pIZT/V5-His-Dip3质粒的Spli221细胞的RNA,反转录cDNA(方法同上),进行qRT-PCR检测Innexin2的转录和Innexin3的转录。其中Innexin2的qRT-PCR检测使用引物Q-Inx2-F(SEQ ID No.5)和Q-Inx2-R (SEQ ID No.6);Innexin3的qRT-PCR检测使用引物Q-Inx3-F(SEQ ID No.7)和Q-Inx3- R(SEQ ID No.8)。方法如下:
将通过上述反转录得到的cDNA按照试剂盒Premix Ex TaqTM II(Tli RNaseH Plus)配制qRT-PCR反应体系(15μL),按相应的顺序放入板中,每个样三个重复,于 qRT-PCR仪(Biosystems 7500Real-Time PCR System)中反应。其中,SYBR Premix Ex Taq (2×)7.5μL,上游引物(10μM)0.2μL,下游引物(10μM)0.2μL,Rox Reference DyeII 0.3μL,cDNA 1.0μL,无RNase dH2O至终体积15μl。
Innexin2的qRT-PCR检测结果见图2A。由图可知,MbBV病毒感染Spli221细胞24小时后,结果发现Innexin2的转录水平降低(阴性对照组)说明MbBV病毒感染会在一定程度上影响Innexin2的转录水平,使Innexin2的转录水平降低;而当在MbBV感染Spli221细胞24小时后的基础上过表达Dip3(实验组)时,发现Innexin2的转录水平显著上升,由此可知,Innexin2的转录依赖于Dip3。Innexin3的qRT-PCR检测结果见图2B,由图可知, MbBV病毒感染Spli221细胞24小时后,结果发现Innexin3的转录水平降低(阴性对照组) 说明MbBV病毒感染会在一定程度上影响Innexin3的转录水平,使Innexin3的转录水平降低;然而当在MbBV感染Spli221细胞24小时后的基础上过表达Dip3(实验组)时, Innexin3的转录水平与阴性对照组相比,没有显著变化,所以Innexin3的转录并不依赖于 Dip3。
8.重组质粒Bacmid-flag3-dip3的构建
取测序成功的pMD19-Dip3菌液,加A+过夜培养,用普通质粒提取试剂盒提取质粒,根据的相应酶切位点对质粒进行双酶切(XbaI/HindIII),用同样的限制性内切酶对干扰载体L4440进设计行双酶切。将Dip3基因片段和L4440载体片段进行回收,T4连接酶连接。转化HT115菌株,鉴定成功,得到L4440-Dip3。用同样的限制性内切酶对载体pFastBac- flag3和L4440-Dip3进行双酶切。将Dip3基因片段和pFastBac-flag3载体片段进行回收,T4 连接酶连接。转化真核表达菌株Top10,鉴定成功,得到含有Dip3核酸序列的pFastBac- flag3-Dip3捐献菌株,转化DH10Bac感受态细胞,得到重组表达质粒Bacmid-flag3-Dip3。
实施例2
1.Vank86,Vank92,Vank101特异引物设计
从双斑侧沟茧蜂病毒基因组数据(Yu,D.S.,Chen,Y.B.,Li,M.,Yang,M.J.,Yang,Y.,& Hu,J.S.,et al.(2016).A polydnaviral genome of microplitis bicoloratus bracovirus and molecular interactions between the host and virus involved in nf-κb signaling.Archives of Virology,161(11), 3095.)中选取Vank86、Vank92和Vank101基因序列,Vank86基因序列(如SEQ ID No.9 所示)大小为516bp,共编码171个氨基酸(SEQ ID No.10);Vank92基因序列(如SEQ ID No.11所示)大小为501bp,共编码166个氨基酸(SEQ ID No.12);Vank101基因序列 (SEQ ID No.13)大小为558bp,共编码186个氨基酸(SEQ ID No.14)。采用引物设计软件Primer Premier 5设计特异引物,上游引物Vank86-F(SEQ ID No.15,含EcoR I酶切位点)和下游引物Vank86-R(SEQ ID No.16,含Not I酶切位点)用于扩增Vank86; Vank92-F(SEQ ID No.17,含EcoR I酶切位点)和下游引物Vank92-R(SEQ ID No.18,含 Not I酶切位点)用于扩增Vank92;上游引物Vank101-F(SEQ ID No.19,含EcoR I酶切位点)和下游引物Vank101-R(SEQ ID No.20,含Not I酶切位点)用于扩增Vank101。
2.双斑侧沟茧蜂病毒总DNA的提取
本实验采用实验室方案提取双斑侧沟茧蜂病毒总DNA,具体步骤如下:
1)在显微镜下选取羽化后3~4天的寄生蜂雌蜂,尾部略显黄色,在显微镜下取出50- 100只卵巢;放置于已冰浴的EP管中(含有灭菌的1×PBS1mL),用5mL注射器上下吹吸数次,直至卵巢破碎;
2)将上述处理好的样品,3000rpm,4℃离心3min;
3)取上清,转移至新的EP管中,将0.45um的滤膜放置新的EP管中,吸取相应的 1×PBS溶液放入滤膜中,用注射器将溶液吹打过膜,直至将滤膜完全润湿;将已经完全吹打破碎的卵巢用注射器吸起后缓慢放入滤膜,吹打到EP管中收集所得即为所需病毒;
4)15000rpm,4℃离心15min,弃上清,离心后的沉淀再加入200μL的PDV-DNA裂解液;
5)加入25μL的蛋白酶K(终浓度为:2.5mg/ml),再加入8μL SarcosyI,于55℃, 3h;
6)加入40μL RNaseA(终浓度为1mg/ml),于37℃,1h。用等量(300μL酚+300μL 氯仿)抽提一次,10000rpm,10min,收集上清;
7)用等量的氯仿抽提一次,10000rpm,10min,收集上清,重复一次;
8)加入等体积的(100%乙醇和1/10的乙酸钠混合物(9:1)),混匀后于-70℃,沉淀 10min,离心12000g,10min,弃上清;
9)加入75%乙醇清洗5min,真空干燥10min。
用上述所得的双斑侧沟茧蜂病毒基因组DNA为模板,用Vank86、Vank92和Vank101 特异性引物进行PCR扩增。得到Vank86的基因片段为516bp,Vank92的基因片段为 501bp,Vank101的基因片段为558bp。将PCR产物回收,并连接到克隆载体pMD19-T载体上,经测序确定正确,分别得到含有靶标片段的pMD19-Vank86、pMD19-Vank92、 pMD19-Vank101质粒。
3.Vank86、Vank92和Vank101基因片段的表达载体构建
1)上述所得的含有靶标片段的pMD19-Vank86和pMD19-Vank92以及pMD19-Vank101 分别利用EcoR I和Not I进行双酶切,得到带有酶切位点粘性末端的Vank86基因片段、 Vank92基因片段和Vank101基因片段。
2)将上述所得的带有酶切位点粘性末端的Vank86基因片段、Vank92基因片段和 Vank101基因片段分别连接到pIZT/V5-His表达载体(可商业购买获得)上。
经过分别对上述重组质粒产物进行双酶切、PCR检测以及测序分析,得到阳性质粒 pIZT/V5-His-Vank86、pIZT/V5-His-Vank92和pIZT/V5-His-Vank101。
4.表达载体pIZT/V5-His-Vank86、pIZT/V5-His-Vank92和pIZT/V5-His-Vank101的转染和表达
1)pIZT/V5-His-Vank86、pIZT/V5-His-Vank92和pIZT/V5-His-Vank101的转染
(1)当细胞培养瓶里细胞的生长密度达到80%-90%时,弃去旧培养基,加入新培养基并将细胞轻吹起,台盼蓝计数个;
(2)将细胞传代到十二孔板中,每孔1ml,数量控制在2.5-3.0×105个细胞/孔,27℃贴壁培养40min以上直到细胞贴壁且状态良好;
(3)配制转染试剂复合物,取3个100μL的无血清培养基置于500μL的离心管中,分别加入1μg的pIZT/V5-His-Vank86、pIZT/V5-His-Vank92和pIZT/V5-His-Vank101质粒,再分别加入4μL的X-tremegene HP DNA Transfection Reagent,混匀后静置15min,将转染试剂混合物逐滴加入到相应孔,轻晃动使转染复合物在12孔板中混合均匀,27℃继续培养。
注:根据实验需要可相应扩大或者缩小体系。
2)表达载体pIZT/V5-His-Vank86和pIZT/V5-His-Vank92以及pIZT/V5-His-Vank101的转染和表达
A.细胞总蛋白的提取
(1)弃去培养基,1×PBS清洗1次,用1×PBS轻将贴壁的细胞吹起,移至轻将贴壁的细胞吹起,移至1.5ml EP管中,3000g,4℃离心5min;
(2)弃去上清,加入50-70μL高效RIPA组织/细胞裂解液,RIPA中需添加中需添加 PMSF(10μL/ml),现用现加,根据需要还可添加其他蛋白酶抑制剂,如磷酸酶抑制剂;
(3)轻轻吹吸沉淀,使细胞均匀混合与RIPA中,震荡30s,冰上放置5-10min使细胞充分裂解;13000g,4℃离心10min,取上清即为总蛋白。
蛋白浓度测定及变性
使用BCA蛋白浓度测定试剂盒进行蛋浓度测定:
(1)将上述得到的总蛋白加入96孔板中,每个孔中加样1μL样品,并添加9μL灭菌双蒸水;组内三次平行以减小误差;
(2)用试剂盒内配套的标准蛋白溶液BSA绘制标准曲线,将蛋白标准液按照0、2、 4、6、8、10微升加到96孔板中,灭菌双蒸水补足10μL;
(3)将试剂盒内A/B溶液按照比例混合后(50个体积A、1个体积B),每孔加入每孔加入200μL,37℃反应30min;
(4)酶标仪562nm波长下测定其吸光值,通过标准曲线计算蛋白浓度;
(5)将蛋白与5×SDS-PAGE上样缓冲液按比例混合后,90℃热处理15min;
(6)立即冷却进行SDS-PAGE凝胶电泳或存放于-80℃备用。
B.SDS-PAGE蛋白凝胶电泳
参照分子克隆配制SDS-PAGE胶:
从制胶器上取下已经凝结的,加入1×电泳缓冲液后拔出梳子,蛋白上样电泳。先用0V 进行跑胶,待条带过层积之后用进行跑胶,待条带过层积之后用100V电压跑大约电压跑大约100min。
C.转膜及后续操作
(1)先把PVDF膜在甲醇中浸湿在甲醇中浸湿1min。并按顺序将PVDF膜覆盖在胶上。在冰水浴中以100V电压跑120min;(2)转膜完成后取出PVDF膜,5%脱脂奶粉室温封闭膜1h;(3)1×PBST清洗已封闭的膜两次,每5min;(4)一抗过夜孵育(4℃); (5)回收一抗,1×PBST清洗以封闭的膜三次,每次5min;(6)二抗(用3%的牛奶以 1:5000-10000稀释)室温孵育膜1h;(7)用1×PBST清洗膜三次,每次10min;(8) Pierce ECL Western Blotting Substrate试剂盒A、B液等体积混合后,滴加在PVDF膜上; FluorChem EFE0511中曝光成像;利用Image J软件进行灰度分析。
根据上述步骤,得到如图3所示的结果,M为蛋白Marker,Anti-V5(Thermo, R96025)为使用的抗体,Tubulin为内参。由图可以看出,pIZT/V5-His-Vank86,pIZT/V5- His-Vank92和pIZT/V5-His-Vank101在昆虫细胞系中成功表达,得到24KDa左右的Vank86 和23kDa大小的Vank92和25kDa大小的Vank101蛋白。
5.Vank蛋白与Dip3共转
将pIZT/V5-His-Vank86和Bacmid-flag3-Dip3按照上述转染操作共转到昆虫细胞Sf9 中,将pIZT/V5-His-Vank92和Bacmid-flag3-Dip3按照上述转染操作共转到昆虫细胞Sf9 中,将pIZT/V5-His-Vank101和Bacmid-flag3-Dip3按照上述转染操作共转到昆虫细胞Sf9 中。通过Western Blot检测两个质粒共同表达的情况,得到图4所示结果,M为蛋白 Marker,Anti-V5和Anti-Flag(购自碧云天)为使用的抗体,Tubulin(购自索莱宝)和 Actin(购自索莱宝)为内参。由图可以看出,pIZT/V5-His-Vank86、pIZT/V5-His- Vank92、pIZT/V5-His-Vank101分别与Bacmid-flag3-Dip3共同表达时,Vank86、Vank92和 Vank101都会分别与Dip3结合形成二聚体,最后只得到一条约为57kDa的蛋白条带(图 4A,4B,4C)。在图4中,分别用两种标签抗体Anti-V5和Anti-Flag孵育,都得到了二聚体的条带。
综上所述,说明:Vank86和Vank92以及Vank101导入斜纹夜蛾体内会能正常表达,并能与斜纹夜蛾Dip3蛋白相结合,引起斜纹夜蛾的血淋巴细胞产生大量而显著的凋亡。因此,利用Dip3、Vank86、Vank92和Vank101可用于防治斜纹夜蛾。
序列表
<110> 云南大学
<120> 氨基酸序列及其在用于使昆虫Dip3蛋白失活中的应用
<130> LHA1760718
<160> 20
<170> SIPOSequenceListing 1.0
<210> 1
<211> 249
<212> PRT
<213> 斜纹夜蛾(Spodoptera litura Fabricius)
<400> 1
Met Gln Phe Glu Tyr Asp Pro Glu Arg Leu Ile Glu Glu Val Arg Lys
1 5 10 15
Arg Pro Gly Ile Trp Asn Tyr Glu Asp Met Glu Tyr Arg Ala Lys Thr
20 25 30
Met Arg Tyr Lys Leu Trp Asn Glu Val Val Thr Glu Leu Met Asn Pro
35 40 45
Glu Ala Lys Val Ser Lys Ser Glu Met Arg Glu Leu Glu Ile Gln Leu
50 55 60
Gln Lys Lys Trp Lys Ser Ile Arg Asp Cys Phe Gln Lys Tyr Ile Ala
65 70 75 80
Asn Pro Asn Arg Thr Lys Lys Pro Tyr Ile Tyr Ser Arg Gln Leu Gln
85 90 95
Phe Leu Leu Lys Asp Gln Glu Glu Leu Pro Arg Lys Glu Asp Gly Gly
100 105 110
Gly Thr Thr Glu Ser Asp Asp Asp Lys Pro Ile Lys Lys Lys Lys Val
115 120 125
Trp Arg Ser Lys Arg Lys Leu Lys Leu Val Lys Glu Glu Thr Ser Glu
130 135 140
Glu Asp Asn Asp Asn Ser Ile Asn Tyr Glu Ala Asn Asp Asp Ser Arg
145 150 155 160
Asp Tyr Thr Met Glu Thr Ser Glu Thr Gly Arg Ser Ala Lys Val Pro
165 170 175
Lys Thr Ser Lys Pro Gln Pro Gln Val Asp Glu Phe Val Phe Ala Ser
180 185 190
Val Asp Thr Gln Ile Lys Asn Glu Ser Asp Asp Pro Asp Arg Met Phe
195 200 205
Leu Leu Ser Leu Leu Pro His Leu Lys Met Ile Pro Glu Glu Phe Arg
210 215 220
Leu Asn Val Lys Met Asp Met Met His Val Leu Arg Asn Ala Ser Tyr
225 230 235 240
Asn Thr Ala Arg Glu His Lys Leu Ile
245
<210> 2
<211> 750
<212> DNA
<213> 斜纹夜蛾(Spodoptera litura Fabricius)
<400> 2
atgcagttcg aatacgatcc cgaaaggctt atagaagaag tgaggaaaag acctggcata 60
tggaattatg aagatatgga atatcgtgct aaaactatga ggtacaaact ctggaacgaa 120
gtcgtcacgg aacttatgaa tcccgaggcc aaggtctcta aaagtgaaat gagagaactc 180
gaaatccaac tacaaaagaa atggaaaagc attcgggact gctttcaaaa atacattgca 240
aatcctaata gaacgaagaa accgtatatt tattccagac agttacaatt tctattgaaa 300
gatcaggagg aattaccacg caaagaggac ggaggaggaa caactgaatc agatgatgac 360
aaaccaatta aaaagaagaa agtttggaga tccaaaagaa agttgaaatt agtgaaagag 420
gagacttcag aagaggacaa tgacaatagt attaattacg aagctaatga tgattccaga 480
gattatacta tggagacgtc tgaaactgga cgatcagcga aggtgccaaa gacaagcaag 540
ccacagccac aagtcgacga gtttgtattt gcaagtgttg atacacaaat aaaaaatgaa 600
tcggatgatc cagatagaat gtttttactg tcacttttgc cacatttgaa gatgatacca 660
gaagaatttc gtttgaacgt gaaaatggat atgatgcacg tactgcgaaa cgcaagttat 720
aacaccgcta gagaacacaa gttgatttaa 750
<210> 3
<211> 19
<212> DNA
<213> 人工序列(non)
<400> 3
gagctcatgc agttcgaat 19
<210> 4
<211> 24
<212> DNA
<213> 人工序列(non)
<400> 4
gatatcaatc aacttgtgtt ctct 24
<210> 5
<211> 19
<212> DNA
<213> 人工序列(non)
<400> 5
cgttccgttt ctttatctg 19
<210> 6
<211> 17
<212> DNA
<213> 人工序列(non)
<400> 6
acacgctcct ctggctc 17
<210> 7
<211> 17
<212> DNA
<213> 人工序列(non)
<400> 7
atcgcatcac atcagcc 17
<210> 8
<211> 18
<212> DNA
<213> 人工序列(non)
<400> 8
aggtaatcca gcaatagg 18
<210> 9
<211> 516
<212> DNA
<213> 双斑侧沟茧蜂病毒(Microplitis bicoloratus bracovirus)
<400> 9
atgggtgttt tagatcttct agttctgctt aagataaact ggcgcgcaca tctcgcgtat 60
agtggagaga acgtttttca cgtgattgtg aaaaatgggt ggtataattc gttatgtgaa 120
ctcgaaccgt tgttggatga caagttcaag acgcggctta atcagaagaa cgatcaagga 180
caaacttgcc ttcacgttgc agccagggta catcgaggcc agcgagcaat aaagatgatg 240
gaaaagttgg tgaattatgg agcggatctt aatgtacgag aaggtagtag tggtgatact 300
gtacttcaca ttgctgtggc ataccaagac tacgaactag ctgcatggtt gtgtcaacat 360
ccaaacatgg atttagaagc agaaaactac tacactcaga cggcgttcaa gaaggctttg 420
aaaaataagg acaaagagat gacaaagata ctcaaaactt atggtgcgat ttctcggcga 480
gagtgtgtct ggctactgcg aagaaagttt ttctga 516
<210> 10
<211> 171
<212> PRT
<213> 双斑侧沟茧蜂病毒(Microplitis bicoloratus bracovirus)
<400> 10
Met Gly Val Leu Asp Leu Leu Val Leu Leu Lys Ile Asn Trp Arg Ala
1 5 10 15
His Leu Ala Tyr Ser Gly Glu Asn Val Phe His Val Ile Val Lys Asn
20 25 30
Gly Trp Tyr Asn Ser Leu Cys Glu Leu Glu Pro Leu Leu Asp Asp Lys
35 40 45
Phe Lys Thr Arg Leu Asn Gln Lys Asn Asp Gln Gly Gln Thr Cys Leu
50 55 60
His Val Ala Ala Arg Val His Arg Gly Gln Arg Ala Ile Lys Met Met
65 70 75 80
Glu Lys Leu Val Asn Tyr Gly Ala Asp Leu Asn Val Arg Glu Gly Ser
85 90 95
Ser Gly Asp Thr Val Leu His Ile Ala Val Ala Tyr Gln Asp Tyr Glu
100 105 110
Leu Ala Ala Trp Leu Cys Gln His Pro Asn Met Asp Leu Glu Ala Glu
115 120 125
Asn Tyr Tyr Thr Gln Thr Ala Phe Lys Lys Ala Leu Lys Asn Lys Asp
130 135 140
Lys Glu Met Thr Lys Ile Leu Lys Thr Tyr Gly Ala Ile Ser Arg Arg
145 150 155 160
Glu Cys Val Trp Leu Leu Arg Arg Lys Phe Phe
165 170
<210> 11
<211> 501
<212> DNA
<213> 双斑侧沟茧蜂病毒(Microplitis bicoloratus bracovirus)
<400> 11
atggcgcctc cggaagaaat tgcatctgtt agtaatcctg acattgaagg tgagaatata 60
ctacatttcc tctgccgtga tggcgatatc actgatttga tggcgttcaa gaatgtaatc 120
aatgatgcca atcggcatct ggtgcagcaa ttcaatcgac atggaaaaca gtgtgtccac 180
attgtctcca acccaggcat tgctgaccct caggaaaaac tgaaacttct gatggagtgg 240
ggtgctgaca tcaatggcca agagcgagtt ttcggtaata caccgctgca tattgcggca 300
tatacacgaa accataagct agcaacctgg ctgtgcaatc agccgggaat taacattgaa 360
atttataatt atctatacaa gacgccgtat tatatcgcgg gtgagcgtag tgattggaag 420
atgataaata tattacgtgc aaaaagaggg caatgtagaa tctaccggtg caacgaagct 480
tggttattta ctagccatta a 501
<210> 12
<211> 166
<212> PRT
<213> 双斑侧沟茧蜂病毒(Microplitis bicoloratus bracovirus)
<400> 12
Met Ala Pro Pro Glu Glu Ile Ala Ser Val Ser Asn Pro Asp Ile Glu
1 5 10 15
Gly Glu Asn Ile Leu His Phe Leu Cys Arg Asp Gly Asp Ile Thr Asp
20 25 30
Leu Met Ala Phe Lys Asn Val Ile Asn Asp Ala Asn Arg His Leu Val
35 40 45
Gln Gln Phe Asn Arg His Gly Lys Gln Cys Val His Ile Val Ser Asn
50 55 60
Pro Gly Ile Ala Asp Pro Gln Glu Lys Leu Lys Leu Leu Met Glu Trp
65 70 75 80
Gly Ala Asp Ile Asn Gly Gln Glu Arg Val Phe Gly Asn Thr Pro Leu
85 90 95
His Ile Ala Ala Tyr Thr Arg Asn His Lys Leu Ala Thr Trp Leu Cys
100 105 110
Asn Gln Pro Gly Ile Asn Ile Glu Ile Tyr Asn Tyr Leu Tyr Lys Thr
115 120 125
Pro Tyr Tyr Ile Ala Gly Glu Arg Ser Asp Trp Lys Met Ile Asn Ile
130 135 140
Leu Arg Ala Lys Arg Gly Gln Cys Arg Ile Tyr Arg Cys Asn Glu Ala
145 150 155 160
Trp Leu Phe Thr Ser His
165
<210> 13
<211> 558
<212> DNA
<213> 双斑侧沟茧蜂病毒(Microplitis bicoloratus bracovirus)
<400> 13
atggatagct cagatagttc agatcttgaa ctgctgctag ccttagactg ggagcgacat 60
cgcgcgcgta ctggaaataa tatttttcac gtcattgcta aaaaaggatg gttgaagatg 120
ctatatgcac tcgaggggtt cgtaaatgaa aatatcaagc catggctgag agaaaggaac 180
aaaaaaggca atacttgcct ccacgaagca gcgtggagaa ataaaggact gcaagcaata 240
cagatgatgg aaaagttagt ggagtatggg gcagacctga atttaagaag ttcttgtaag 300
aaacctgtac ttcacgtggc agtagaaaaa gaagatcatg aagtagttgc atggatatgt 360
cagcaaccag aaatcgattt ggaagccgaa aatttctcca aattcaccgc gtacaaatgg 420
gcatcgaaaa actgtctcaa gagtgaccaa aaaatgatga agatacttaa aactcatggt 480
gcaatttctc gggaagatag cactagtgaa gaagaagaag aagaagagag tgatttagat 540
cagtacctcg aagtttga 558
<210> 14
<211> 185
<212> PRT
<213> 双斑侧沟茧蜂病毒(Microplitis bicoloratus bracovirus)
<400> 14
Met Asp Ser Ser Asp Ser Ser Asp Leu Glu Leu Leu Leu Ala Leu Asp
1 5 10 15
Trp Glu Arg His Arg Ala Arg Thr Gly Asn Asn Ile Phe His Val Ile
20 25 30
Ala Lys Lys Gly Trp Leu Lys Met Leu Tyr Ala Leu Glu Gly Phe Val
35 40 45
Asn Glu Asn Ile Lys Pro Trp Leu Arg Glu Arg Asn Lys Lys Gly Asn
50 55 60
Thr Cys Leu His Glu Ala Ala Trp Arg Asn Lys Gly Leu Gln Ala Ile
65 70 75 80
Gln Met Met Glu Lys Leu Val Glu Tyr Gly Ala Asp Leu Asn Leu Arg
85 90 95
Ser Ser Cys Lys Lys Pro Val Leu His Val Ala Val Glu Lys Glu Asp
100 105 110
His Glu Val Val Ala Trp Ile Cys Gln Gln Pro Glu Ile Asp Leu Glu
115 120 125
Ala Glu Asn Phe Ser Lys Phe Thr Ala Tyr Lys Trp Ala Ser Lys Asn
130 135 140
Cys Leu Lys Ser Asp Gln Lys Met Met Lys Ile Leu Lys Thr His Gly
145 150 155 160
Ala Ile Ser Arg Glu Asp Ser Thr Ser Glu Glu Glu Glu Glu Glu Glu
165 170 175
Ser Asp Leu Asp Gln Tyr Leu Glu Val
180 185
<210> 15
<211> 25
<212> DNA
<213> 人工序列(non)
<400> 15
gaattcatgg gtgttttaga tcttc 25
<210> 16
<211> 18
<212> DNA
<213> 人工序列(non)
<400> 16
gcggccgcag aaaaactt 18
<210> 17
<211> 25
<212> DNA
<213> 人工序列(non)
<400> 17
gaattcatgg gtgttttaga tcttc 25
<210> 18
<211> 18
<212> DNA
<213> 人工序列(non)
<400> 18
gcggccgcaa tggctagt 18
<210> 19
<211> 23
<212> DNA
<213> 人工序列(non)
<400> 19
ggcgaattca tggatagctc aga 23
<210> 20
<211> 21
<212> DNA
<213> 人工序列(non)
<400> 20
gcggccgcaa taaacttcga g 21
- 还没有人留言评论。精彩留言会获得点赞!