抗-IgE抗体的制作方法
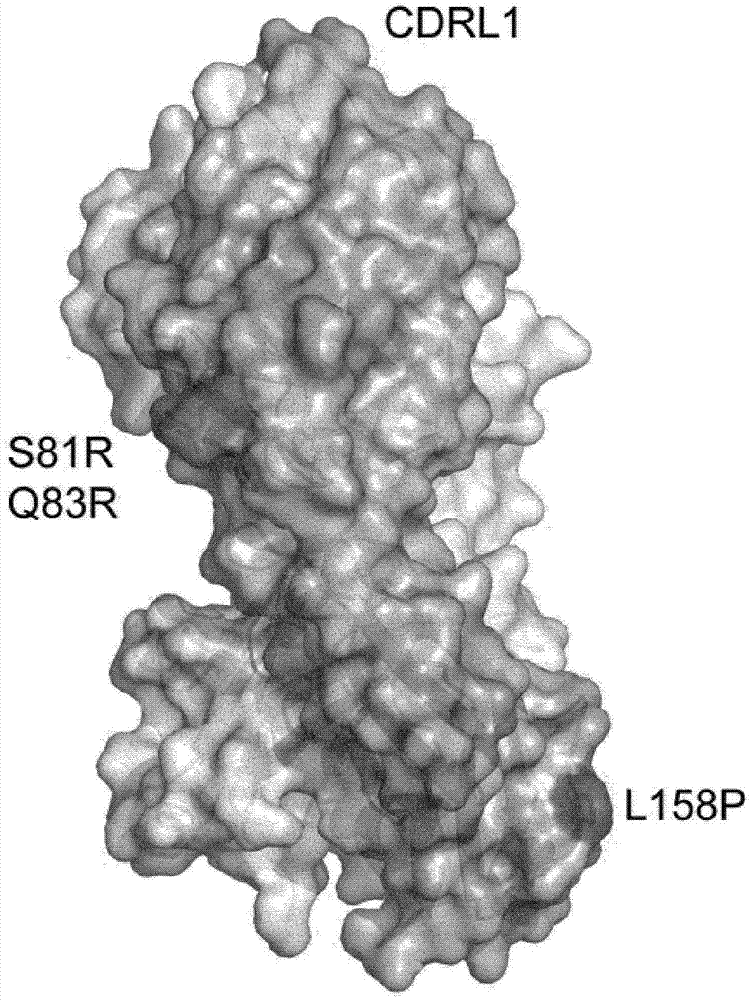
本发明在靶向ige的改善的抗-ige抗体和抗原结合剂和其组合物、例如用于治疗由ige引起的病症(诸如过敏反应,或某些自身免疫性反应),且特别是由ige与fcεri受体的相互作用引起的病症的领域中。具体而言,本发明涉及与奥马珠单抗的新型突变体有关的改善的抗-ige抗体和抗原结合剂。本发明的改善的抗-ige抗体和抗原结合剂可具有对ige的改善的亲和力和/或与ige的cε2结构域和/或ige上的改善的修饰表位(例如进一步涉及ige的cε2结构域)的改善的相互作用和/或在药学相关浓度下使ige自fcεri受体解离的能力。在一个方面,公开了用于ige介导的病症的改善或新型的治疗,其中靶向ige(例如游离ige和/或与fcεri受体复合的ige)。发明背景ige是介导过敏反应诸如哮喘、食物过敏、1型超敏反应和基于众多原因的常见鼻窦炎的免疫球蛋白质家族的成员。ige由b细胞分泌并在b细胞的表面上表达。b细胞合成的ige通过经短膜结合区连接至成熟ige序列的跨膜结构域锚定于b细胞膜中。ige还通过其fc区连接至低亲和力ige受体(fcεrii)而结合至b细胞(以及单核细胞、嗜酸性细胞和血小板)。在哺乳动物暴露于过敏原后,b细胞无性扩增,其合成结合该过敏原的ige。该ige进而由b细胞释放至循环中,在循环中其被b细胞结合(经由fcεrii)且通过在肥大细胞和嗜碱性细胞表面上发现的所谓高亲和力受体(fcεri)而被肥大细胞和嗜碱性细胞所结合。由此,此类肥大细胞和嗜碱性细胞针对过敏原敏化。接下来暴露于过敏原将交联这些细胞上的fcεri且因此活化其组胺和负责临床超敏反应和过敏反应的其他因子的释放。奥马珠单抗是选择性结合人类免疫球蛋白e(ige)[cε3结构域]的一种重组dna衍生的人源化igg1κ单克隆抗体。所述抗体具有约149kd的分子量。由中国仓鼠卵巢细胞悬浮液培养物在含有抗生素庆大霉素(gentamicin)的培养基中产生。是在一次性使用小瓶中含有的用usp无菌注射用水(swfi)复原的无菌、白色、不含防腐剂的冻干粉末(或替代地,作为无菌注射器中的液体制剂)且作为皮下(sc)注射液施用[参见ep602126(和基于其的spc/gb06/005);wo93/04173;us6267958(和基于该专利的pte);wo97/04807;wo97/04801;presta等人(1993)j.immunol.151:2623-2632]。奥马珠单抗目前适用于治疗皮肤测试呈阳性或在体外对长期气源性致敏原具有反应性且具有吸入性皮质类固醇不充分控制的症状的患者中的中度至重度持久性哮喘(来自处方信息)。用奥马珠单抗的问题在于:1)其在药学相关剂量下靶向游离ige但不靶向(或不能有效靶向)ige/fcεri复合物的致病性种类;2)可能由于ige/fcεri复合物的的致病性种类不被靶向,“xolair治疗花费至少12-16周来显示有效性”(150mg溶液-summaryofproductcharacteristics2014)-或实际上确定是否会对特定患者起作用或是否需要不同治疗;3)其不应当用于具有高ige水平的患者(例如,因为ige/fcεri复合物的致病性种类不被靶向且鉴于患者中的高游离ige水平而不会随时间耗散);4)“当服用奥马珠单抗时可以发生i型局部或全身反应,包括过敏反应和过敏性休克”150mg溶液-summaryofproductcharacteristics2014);5)其对ige的亲和力并不是特别好(约2nm)。本发明的一个目的是鉴定改善这些问题中的一种或多种的新型抗体。另一目的是鉴定针对新型表位的抗体(相比于奥马珠单抗,具有增加的igecε2相互作用),和/或具有改善的亲和力和/或改善的解离ige/fcεri复合物的能力的基于新型奥马珠单抗突变体的抗体。本发明的又一目的是鉴定用于治疗与ige有关的病症,特别是与ige/fcεri复合物有关的病症,例如过敏性病症的新化合物、方法和组合物。发明概述在本发明的一个方面,提供了抗-ige抗体或抗原结合剂,其接触包含人类ige的cε3结构域的残基t373、w374、s375、r376、a377、s378、g379、p381、q417、c418、r419、t421、p426、r427、a428以及cε2结构域的残基d278和t281的表位。在进一步实施方案中,所述表位可以进一步包含人类ige的cε3结构域的残基k380和/或m430中的一个或多个和/或人类ige的cε2结构域的残基d276、v277、l279、s280、a282和/或t298中的一个或多个。本发明基于实施例1的晶体结构的观察结果,其首次显示改善的抗体(基于奥马珠单抗)与ige-fc的相互作用,其中在igecε2结构域的突变区域中观察到显著相互作用。这可以导致抗-ige抗体或抗原结合剂的功能特征相对于奥马珠单抗和/或奥马珠单抗fab改善。例如,抗-ige抗体或抗原结合剂能够在小于7、3、1、0.66、0.5或0.3μm的浓度(或峰值血清浓度)下将人类ige从fcεri解离(例如,如通过实施例2中所述的方法进行)。例如,所述抗-ige抗体或抗原结合剂可以具有相对于奥马珠单抗和/或奥马珠单抗fab改善/增强的针对人类ige(例如使用ige-fc)的亲和力(较低kd)(例如,如通过实施例6中所述的方法进行);和/或相对于奥马珠单抗和/或奥马珠单抗fab改善的解离ige/fcεri复合物的能力(例如,如通过实施例2中所述的方法测定);和/或在低于奥马珠单抗和/或奥马珠单抗fab的浓度(或峰值血清浓度)下将人类ige与fcεri解离的性能(例如,如通过实施例2中所述的方法测定)。改善的kd意指比奥马珠单抗和/或奥马珠单抗fab的kd低至少5%、10%、20%、30%、40%或50%。本发明的抗-ige抗体或抗原结合剂的kd可以小于2、1.9、1.8、1.7、1.6、1.5、1.4、1.3、1.2、1.1、1.0、0.9、0.8、0.7、0.6、0.5、0.4或0.3nm。改善的将人类ige与fcεri解离的能力或性能意指相对于奥马珠单抗和/或奥马珠单抗fab改善至少5%、10%、20%、30%、40%、50%或100%(例如,当如实施例2和7中所述测量ige/fcεri复合物的%解离和/或表观解离速率时),和/或在奥马珠单抗和/或奥马珠单抗fab没有实现解离的浓度下实现解离。为了避免疑问,本发明的抗-ige抗体或抗原结合剂不是奥马珠单抗或奥马珠单抗fab。在一个实施方案中,通过在复合的ige-fc/抗-ige抗体或抗原结合剂的晶体结构中测定所述抗-ige抗体或抗原结合剂的或内的ige残基来结晶学测定表位(例如,如实施例1中所述)。所用ige-fc可以是seq.idno.108的ige-fc(具有额外n265q和n371q突变)。在一个实施方案中,所述抗-ige抗体或抗原结合剂在特定结合位点接触所述表位,其中所述表位的cε3结构域和cε2结构域部分在人类ige的不同链上。ige在fc结构域中具有两条链,各自具有cε3结构域和cε2结构域。在一个实施方案中,所述抗-ige抗体或抗原结合剂在特定结合位点接触所述表位,其中所述表位的cε3结构域和cε2结构域部分在人类ige的相同链上。为了避免疑问,本发明的两种抗-ige抗体或抗原结合剂可以结合人类ige,但仅需要这些之一与包含cε3和cε2结构域的本发明的表位相互作用(另一个可以例如仅与另一cε3结构域相互作用)。在一个实施方案(任选地进一步采用本发明的第一个方面的特征的实施方案)中,所述抗-ige抗体或抗原结合剂对包含人类ige的cε3结构域的残基t373、w374、s375、r376、a377、s378、g379、p381、q417、c418、r419、t421、p426、r427、a428以及cε2结构域的残基d278和t281的表位是特异性的。任选地,所述表位可以进一步包含人类ige的cε3结构域的残基k380和/或m430中的一个或多个和/或人类ige的cε2结构域的残基d276、v277、l279、s280、a282和/或t298中的一个或多个。为了避免疑问,如果所述抗-ige抗体或抗原结合剂识别且结合包含所述表位的特定人类ige结构而非一般性地结合人类ige,则其对所述表位是特异性的。在一个进一步的方面(任选地进一步采用本发明的第一个方面的特征的方面)中,提供了抗-ige抗体或抗原结合剂,其含有包含氨基酸序列为seq.idno.18的互补决定区cdr-h3的重链可变区,和包含氨基酸序列为seq.idno.29的互补决定区cdr-l1的轻链可变区,其中所述轻链可变区进一步包含具有选自seq.idno.32的氨基酸序列的构架区fr-l3,所述氨基酸序列具有一、二、三、四、五、六、七个或更多个氨基酸取代以加强所述抗-ige抗体或抗原结合剂与人类ige的cε2结构域的相互作用。在一个进一步的方面(任选地进一步采用本发明的前述方面的特征的方面)中,提供了抗-ige抗体或抗原结合剂,其含有包含氨基酸序列为seq.idno.18的互补决定区cdr-h3的重链可变区,和包含氨基酸序列为seq.idno.29的互补决定区cdr-l1的轻链可变区,其中所述轻链可变区进一步包含具有作为seq.idno.28的氨基酸序列的构架区fr-l1,所述氨基酸序列具有一、二、三、四、五、六、七个或更多个氨基酸取代以加强所述抗-ige抗体或抗原结合剂与人类ige的cε2结构域的相互作用。在cdr-h3和cdr-l1区域将抗-ige抗体或抗原结合剂锚定和定向于igecε3区域(按照奥马珠单抗)上时,对fr-l3和/或fr-l1序列的变化允许与人类ige的cε2结构域的较强相互作用。所述突变体相对于奥马珠单抗或奥马珠单抗fab的较强相互作用可以通过亲和力测量[较低kd](例如,如通过实施例6中所述的方法进行)和/或改善的ige/fcεri复合物解离的特征(例如,如通过实施例2中所述的方法测定)进行评估。抗-ige抗体或抗原结合剂与人类ige的cε2结构域的较强相互作用可以通过抗-ige抗体或抗原结合剂的功能特征相对于奥马珠单抗和/或奥马珠单抗fab的改善来表征。例如,抗-ige抗体或抗原结合剂能够在小于7、3、1、0.66、0.5或0.3μm的浓度(或峰值血清浓度)下将人类ige从fcεri解离(例如,如通过实施例2中所述的方法进行)。例如,所述抗-ige抗体或抗原结合剂可以具有相对于奥马珠单抗和/或奥马珠单抗fab改善/增强的针对人类ige(例如使用ige-fc)的亲和力(较低kd)(例如,如通过实施例6中所述的方法进行);和/或相对于奥马珠单抗和/或奥马珠单抗fab改善的解离ige/fcεri复合物的能力(例如,如通过实施例2中所述的方法测定);和/或在低于奥马珠单抗和/或奥马珠单抗fab的浓度(或峰值血清浓度)下将人类ige与fcεri解离的性能(例如,如通过实施例2中所述的方法测定)。改善的kd意指比奥马珠单抗和/或奥马珠单抗fab的kd低至少5%、10%、20%、30%、40%或50%。本发明的抗-ige抗体或抗原结合剂的kd可以小于2、1.9、1.8、1.7、1.6、1.5、1.4、1.3、1.2、1.1、1.0、0.9、0.8、0.7、0.6、0.5、0.4或0.3nm。改善的将人类ige与fcεri解离的能力或性能意指相对于奥马珠单抗和/或奥马珠单抗fab改善至少5%、10%、20%、30%、40%、50%或100%(例如,当如实施例2和7中所述测量ige/fcεri复合物的%解离和/或表观解离速率时),和/或在奥马珠单抗和/或奥马珠单抗fab没有实现解离的浓度下实现解离。为了避免疑问,本发明的抗-ige抗体或抗原结合剂不是奥马珠单抗或奥马珠单抗fab。在一个实施方案中,参照seqidno;129,fr-l3区域在位置s60、s63、s76、s77和/或q79(kabat)中的一个或多个处突变成其他天然氨基酸之一。例如,fr-l3区域可以在位置s60(kabat)处突变成其他天然氨基酸之一,例如,突变成m、r、k、n、q或t,特别是m。例如,fr-l3区域可以在位置s63(kabat)处突变成其他天然氨基酸之一,例如,突变成w或y,特别是y。例如,fr-l3区域可以在位置s76(kabat)处突变成其他天然氨基酸之一,特别是n。例如,fr-l3区域可以在位置s77(kabat)处突变成其他天然氨基酸之一,例如,突变成r或k,特别是r。例如,fr-l3区域可以在位置q79(kabat)处突变成其他天然氨基酸之一,例如,突变成r或k,特别是r。例如,参照seqidno:20,fr-l1区可以在g16和/或r18(kabat)上突变成其他天然氨基酸之一。在某些实施方案中,所述抗-ige抗体或抗原结合剂的突变的fr-l3区的氨基酸序列选自seq.idno.43-49、60-83、131或138。在一个进一步实施方案中,参照seqidno:129,fr-l3区进一步在位置s67(kabat)处突变成其他天然氨基酸之一以改善其对人类ige的亲和力(较低kd)。在该情况下,所述突变可以加强抗-ige抗体或抗原结合剂与ige的cε3结构域的相互作用。例如,fr-l3区域可以在位置s67(kabat)处突变成m(特别地)、e或d。在某些实施方案中,所述抗-ige抗体或抗原结合剂的突变的fr-l3区域的氨基酸序列选自seq.idno.53-59、84-107、131或138。本发明的抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.31的互补决定区cdr-l2的轻链可变区。在一个实施方案中,cdr-l2区域在位置s52(kabat)处突变成其他天然氨基酸之一以改善其对人类ige的亲和力(较低kd)。在该情况下,所述突变可以加强抗-ige抗体或抗原结合剂与ige的cε3结构域的相互作用。例如,参照seqidno:129,cdr-l2区域可以在位置s52(kabat)处突变成d(特别地)、e、q或r。在某些实施方案中,突变的cdr-l2区域的氨基酸序列选自seq.idno.50或seq.idno.51。本发明的抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.14的互补决定区cdr-h1的重链可变区。本发明的抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.16的互补决定区cdr-h2的重链可变区。本发明的抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.33的互补决定区cdr-l3的轻链可变区。本发明的抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.13的构架区fr-h1的重链可变区。本发明的抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.15的构架区fr-h2的重链可变区。本发明的抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.17的构架区fr-h3的重链可变区。本发明的抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.19的构架区fr-h4的重链可变区。本发明的抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.30的构架区fr-l2的轻链可变区。本发明的抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.34的构架区fr-l4的轻链可变区。本发明的抗-ige抗体或抗原结合剂可以具有轻链可变区vl,其具有选自seq.idno.35、seq.idno.132或seqidno.134或seq.idno.141或seqidno.144或seqidno.145或seqidno.158或seqidno.159的氨基酸序列。本发明的抗-ige抗体或抗原结合剂可以具有氨基酸序列为seq.idno.1的重链可变区vh。本发明的抗-ige抗体或抗原结合剂可以进一步包含轻链恒定区。本发明的抗-ige抗体或抗原结合剂可以具有作为κ恒定区的轻链恒定区。本发明的抗-ige抗体或抗原结合剂可以具有轻链恒定区,所述轻链恒定区具有突变l154p(kabat)。本发明的抗-ige抗体或抗原结合剂可以具有轻链可变区和轻链恒定区vl-cl,其具有选自seq.idno.39或seq.idno.41或seq.idno.117或seq.idno.119或seq.idno.125或seq.idno.127或seq.idno.136或seq.idno.143的氨基酸序列,其任选地包含氨基酸序列为seqidno.160的信号序列。本发明的抗-ige抗体或抗原结合剂可以进一步包含重链恒定区ch1。本发明的抗-ige抗体或抗原结合剂可以具有重链可变区和重链恒定区vh-ch1,其具有作为seq.idno.5的氨基酸序列。本发明的抗-ige抗体或抗原结合剂可以进一步包含重链fc区fc。本发明的抗-ige抗体或抗原结合剂可以具有来自人类igg1或人类igg4的fc。本发明的抗-ige抗体或抗原结合剂可以具有重链可变区、重链恒定区和重链fc区vh-ch1-fc,其具有作为seq.idno.9的氨基酸序列。在本发明的一个进一步的方面,提供了抗-ige抗体或抗原结合剂,其含有包含氨基酸序列为seq.idno.18的互补决定区cdr-h3的重链可变区,和包含氨基酸序列为seq.idno.29的互补决定区cdr-l1的轻链可变区,其中:a.所述轻链可变区进一步包含氨基酸序列为seq.idno.32的构架区fr-l3,其中参照seqidno:129,所述fr-l3区在位置s67(kabat)处突变成其他天然氨基酸之一以改善所述抗-ige抗体或抗原结合剂对人类ige的亲和力(较低kd);和/或b.所述轻链可变区进一步包含氨基酸序列为seq.idno.31的互补决定区cdr-l2,其中参照seqidno:129,所述cdr-l2区在位置s52(kabat)处突变成其他天然氨基酸之一以改善所述抗-ige抗体或抗原结合剂对人类ige的亲和力(较低kd)。本发明人在本文已经发现,这些突变中的任一个或两个可以令人惊讶地基于奥马珠单抗或奥马珠单抗fab改善抗-ige抗体或抗原结合剂对人类ige(例如使用ige-fc)的亲和力(改善或较低kd)(例如,如通过实施例6中所述的方法进行)。具体而言,亲和力的改善相对于奥马珠单抗和/或奥马珠单抗fab。所述突变可以改善与ige的cε3结构域的相互作用。改善或较低的kd意指比奥马珠单抗和/或奥马珠单抗fab的kd低至少5%、10%、20%、30%、40%或50%。本发明的抗-ige抗体或抗原结合剂的kd可以小于2、1.9、1.8、1.7、1.6、1.5、1.4、1.3、1.2、1.1、1.0、0.9、0.8、0.7、0.6、0.5、0.4或0.3nm。例如,参照seqidno:129,fr-l3区域可以在位置s67(kabat)处突变成m(特别地)、e或d。在某些实施方案中,所述抗-ige抗体或抗原结合剂的突变的fr-l3区域的氨基酸序列选自seq.idno.52-59、84-107、131或138。例如,参照seqidno:129,cdr-l2区域可以在位置s52(kabat)处突变成d(特别地)、e、q或r。在某些实施方案中,所述抗-ige抗体或抗原结合剂的突变的cdr-l2区域的氨基酸序列选自seq.idno.50(特别地)或seq.idno.51。所述抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.14的互补决定区cdr-h1的重链可变区。所述抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.16的互补决定区cdr-h2的重链可变区。所述抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.33的互补决定区cdr-l3的轻链可变区。所述抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.13的构架区fr-h1的重链可变区。所述抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.15的构架区fr-h2的重链可变区。所述抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.17的构架区fr-h3的重链可变区。所述抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.19的构架区fr-h4的重链可变区。所述抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.30的构架区fr-l2的轻链可变区。所述抗-ige抗体或抗原结合剂可以具有进一步包含氨基酸序列为seq.idno.34的构架区fr-l4的轻链可变区。所述抗-ige抗体或抗原结合剂可以具有轻链可变区vl,其包含连续fr-l1、cdr-l1、fr-l2、cdr-l2、fr-l3、cdr-l3和fr-l4区域,且具有作为seq.idno.20的氨基酸序列,除了所述cdr-l2区域具有选自seq.idno.50(特别地)或seq.idno.51的氨基酸序列。所述抗-ige抗体或抗原结合剂可以具有轻链可变区vl,其包含连续fr-l1、cdr-l1、fr-l2、cdr-l2、fr-l3、cdr-l3和fr-l4区域,且具有作为seq.idno.20的氨基酸序列,除了所述fr-l3区域具有作为seq.idno.52的氨基酸序列。所述抗-ige抗体或抗原结合剂可以具有轻链可变区vl,其包含连续fr-l1、cdr-l1、fr-l2、cdr-l2、fr-l3、cdr-l3和fr-l4区域,且具有作为seq.idno.20的氨基酸序列,除了所述cdr-l2区域具有选自seq.idno.50(特别地)或seq.idno.51的氨基酸序列且所述fr-l3区域具有选自seq.idno.52、131或138的氨基酸序列。本发明的抗-ige抗体或抗原结合剂可以具有氨基酸序列为seq.idno.1的重链可变区vh。所述抗-ige抗体或抗原结合剂可以进一步包含轻链恒定区。所述抗-ige抗体或抗原结合剂可以具有作为κ恒定区的轻链恒定区。所述抗-ige抗体或抗原结合剂可以具有轻链可变区和轻链恒定区vl-cl,其具有作为seq.idno.24的氨基酸序列,除了所述cdr-l2区域具有选自seq.idno.50(特别地)或seq.idno.51的氨基酸序列。所述抗-ige抗体或抗原结合剂可以具有轻链可变区和轻链恒定区vl-cl,其具有作为seq.idno.24的氨基酸序列,除了所述fr-l3区域具有作为seq.idno.52的氨基酸序列。所述抗-ige抗体或抗原结合剂可以具有轻链可变区和轻链恒定区vl-cl,其具有作为seq.idno.24的氨基酸序列,除了所述cdr-l2区域具有选自seq.idno.50(特别地)或seq.idno.51的氨基酸序列且所述fr-l3区域具有选自seq.idno.52、131或138的氨基酸序列。在一个进一步的方面,本发明提供了抗-ige抗体或抗原结合剂,其包含重链可变区和轻链可变区,其中:a.所述重链可变区包含氨基酸序列为seqidno.14的cdr-h1、氨基酸序列为seqidno.16的cdr-h2和氨基酸序列为seqidno.18的cdr-h3且所述轻链可变区包含氨基酸序列为seqidno.29的cdr-l1、氨基酸序列为seqidno.50的cdr-l2、氨基酸序列为seqidno.33的cdr-l3和氨基酸序列为seqidno.131或138的构架区fw-l3;或b.所述重链可变区包含作为seqidno.1的氨基酸序列且所述轻链可变区包含选自seqidno.132或139的氨基酸序列。在一个实施方案中,所述抗-ige抗体或抗原结合剂可以进一步包含轻链恒定区,其中轻链可变区和轻链恒定区vl-cl具有选自seqidno.136或143的氨基酸序列,任选地包含具有作为seqidno.160的氨基酸序列的信号序列。如本文所述的所有实施方案中的抗-ige抗体或抗原结合剂可以进一步包含重链恒定区ch1。所述抗-ige抗体或抗原结合剂可以具有重链可变区和重链恒定区vh-ch1,其具有作为seq.idno.5的氨基酸序列。所述抗-ige抗体或抗原结合剂可以进一步包含重链fc区,fc。所述抗-ige抗体或抗原结合剂可以具有来自人类igg1或人类igg4的fc。所述抗-ige抗体或抗原结合剂可以具有重链可变区、重链恒定区和重链fc区vh-ch1-fc,其具有作为seq.idno.9的氨基酸序列。本发明的所有方面的抗-ige抗体或抗原结合剂可以选自:具有全长重链和轻链的完整抗体分子,或其片段。本发明的抗-ige抗体或抗原结合剂可以选自:fab片段、修饰的fab’片段、fab’片段、f(ab’)2片段、fv、scfv、scab、双功能抗体、双特异性抗体、三链抗体、fabfv、fab-fv-fv、三功能抗体或(fab-fv)2-fc。不受理论束缚,如果本发明的抗-ige抗体或抗原结合剂仅具有一个而非多个抗-ige抗原结合位点,则其可以具有较低的与其相关的过敏反应风险。在一个实施方案中,所述抗-ige抗体是直接或经由接头连接至scfv的fab片段,所述scfv结合血清载体蛋白质,诸如人类血清白蛋白。在一个实施方案中,所述scfv可以包含优选经由具有seq.idno.151的接头连接的重链可变区和轻链可变区,其中所述重链可变区包含氨基酸序列为seqidno.152的cdr-h1、氨基酸序列为seqidno.153的cdr-h2和氨基酸序列为seqidno.154的cdr-h3且所述轻链可变区包含氨基酸序列为seqidno.155的cdr-l1、氨基酸序列为seqidno.156的cdr-l2、氨基酸序列为seqidno.157的cdr-l3。在一个实施方案中,所述scfv具有作为seqidno.150的氨基酸序列。在一个优选实施方案中,所述fab片段包含重链可变区和轻链可变区,其中:a.所述重链可变区包含氨基酸序列为seqidno.14的cdr-h1、氨基酸序列为seqidno.16的cdr-h2和氨基酸序列为seqidno.18的cdr-h3且所述轻链可变区包含氨基酸序列为seqidno.29的cdr-l1、氨基酸序列为seqidno.50的cdr-l2、氨基酸序列为seqidno.33的cdr-l3和氨基酸序列为seqidno.131或138的构架区fw-l3;或b.所述重链可变区包含作为seqidno.1的氨基酸序列且所述轻链可变区包含选自seqidno.132或139的氨基酸序列。在另一个实施方案中,所述fab片段进一步包含重链恒定区和轻链恒定区,其中重链可变区和重链恒定区vl-ch1具有作为seqidno.5的氨基酸序列且其中轻链可变区和轻链恒定区vl-cl具有选自seqidno.136或143的氨基酸序列,任选地包含具有作为seqidno.160的氨基酸序列的信号序列。在另一个实施方案中,scfv经由具有作为seqidno.149的氨基酸序列的接头连接至fab片段的ch1。在一个实施方案中,所述重链可变区和重链恒定区、接头和scfv具有作为seqidno.147的氨基酸序列,任选地包含具有作为seqidno.160的氨基酸序列的信号序列。在一个其他实施方案中,连接至具有seq.idno.147的scfv的fab片段的重链与具有seqidno.136或143的轻链可变区和恒定区配对。本发明的抗-ige抗体或抗原结合剂可以具有与之连接的效应子或报告子分子。本发明的抗-ige抗体或抗原结合剂可以被糖基化(例如在fc结构域内)和/或可以缀合至选自淀粉、白蛋白和聚乙二醇(peg)的聚合物。在一个实施方案中,缀合的peg可以具有范围5至50kda内的分子量。本发明的抗-ige抗体或抗原结合剂可以被人源化。本发明的一个进一步的方面涉及分离的dna序列,其编码本发明的抗-ige抗体或抗原结合剂的重链和/或轻链。进一步提供了克隆或表达载体,其包含一个或多个本发明的dna序列。例如,克隆或表达载体可以包含一个或多个选自以下的dna序列:seq.idno.36、seq.idno.38、seq.idno.40或seq.idno.42或seq.idno.133或seqidno.135或seqidno.137或seqidno.140或seq.idno.142或seqidno.144,且任选地可以进一步包含一个或多个选自以下的dna序列:seq.idno.2、seq.idno.4、seq.idno.6、seq.idno.8、seq.idno.10或seq.idno.12或seq.idno.148。本发明的一个进一步的方面是宿主细胞,其包含一种或多种本发明的克隆或表达载体。本发明的宿主细胞可以任选地进一步包含一种或多种克隆或表达载体,其包含一种或多种选自以下的dna序列:seq.idno.2、seq.idno.4、seq.idno.6、seq.idno.8、seq.idno.10或seq.idno.12或seqidno.148。还提供了用于产生本发明的抗-ige抗体或抗原结合剂的方法,其包括培养本发明的宿主细胞和分离所述抗-ige抗体或抗原结合剂。一个进一步的方面涉及药物组合物,其包含本发明的抗-ige抗体或抗原结合剂,以及药学上可接受的赋形剂、稀释剂或载体中的一种或多种。合适地,本发明的抗-ige抗体或抗原结合剂以每ml稀释剂50至200mg、优选约150mg或精确地150mg的剂量存在。在某些实施方案中,所述赋形剂包含l-精氨酸、l-组氨酸中的一种或两种。所述赋形剂可以独立地或组合地包含聚山梨醇酯20。所述稀释剂可以是水或水性等渗溶液。本发明的药物组合物可以作为在皮下施用之前复原的粉末装载于无菌小瓶内,或装载于无菌注射器内用于其实时皮下施用。本发明的药物组合物可以含有75至600mg,例如约或精确地100或150mg的抗-ige抗体或抗原结合剂的总剂量。本发明的药物组合物可以另外包含与抗-ige抗体或抗原结合剂一起包含或与抗-ige抗体或抗原结合剂分开共施用的其他活性成分。例如,本发明的药物组合物可以用于基于过敏的特异性免疫疗法的背景下,其中本发明的抗-ige抗体或抗原结合剂与过敏原分开共施用(但可以共包装)。因此,本发明的药物组合物可以用于基于过敏的特异性免疫疗法中,其中所述患者在接受治疗性过敏原之前7天、6天、5天、4天、3天、2天或1天(或同一天)接受本发明的药物组合物。本发明的抗-ige抗体或抗原结合剂或组合物可以用作药物。本发明的抗-ige抗体或抗原结合剂或组合物可以用于治疗或预防疾病。本发明的抗-ige抗体或抗原结合剂或组合物可以用于治疗或预防与人类ige和fcεri的复合物相关的病症。本发明的抗-ige抗体或抗原结合剂或组合物可以通过解离人类ige和fcεri的复合物和所述抗-ige抗体或抗原结合剂对人类ige的结合而用于治疗或预防病症。本发明的抗-ige抗体或抗原结合剂或组合物可以用于治疗或预防以下中的一种或多种:过敏;过敏性哮喘;重度哮喘;中度哮喘;慢性自发性荨麻疹;慢性特发性荨麻疹;常年性过敏性鼻炎;季节性过敏性鼻炎;急性哮喘恶化;急性支气管痉挛;哮喘持续状态;高ige综合征;过敏性支气管肺曲霉病;过敏性反应的预防;食物过敏;特应性皮炎;过敏性鼻炎;蜂毒敏感;特发性过敏反应;花生过敏;乳胶过敏;炎性皮肤病;荨麻疹(日光、冷诱导、局部热诱导和/或长期压力诱导);皮肤肥大细胞增多症;全身肥大细胞增多症;嗜伊红细胞相关胃肠病症;大疱性类天疱疮;间质性膀胱炎;鼻息肉;特发性血管性水肿;或非过敏性哮喘。进一步提供了用于治疗或预防人类受试者中的疾病的方法,所述方法包括向所述受试者施用有效量的本发明的抗-ige抗体或抗原结合剂或组合物。所述方法可以用于治疗或预防与人类ige和fcεri的复合物有关的病症。本发明的方法可以通过解离人类ige和fcεri的复合物和本发明的抗-ige抗体或抗原结合剂对人类ige的结合来治疗或预防疾病。本发明的方法可以用于治疗或预防以下中的一种或多种:过敏;过敏性哮喘;重度哮喘;中度哮喘;慢性自发性荨麻疹;慢性特发性荨麻疹;常年性过敏性鼻炎;季节性过敏性鼻炎;急性哮喘恶化;急性支气管痉挛;哮喘持续状态;高ige综合征;过敏性支气管肺曲霉病;过敏性反应的预防;食物过敏;特应性皮炎;过敏性鼻炎;蜂毒敏感;特发性过敏反应;花生过敏;乳胶过敏;炎性皮肤病;荨麻疹(日光、冷诱导、局部热诱导和/或长期压力诱导);皮肤肥大细胞增多症;全身肥大细胞增多症;嗜伊红细胞相关胃肠病症;大疱性类天疱疮;间质性膀胱炎;鼻息肉;特发性血管性水肿;或非过敏性哮喘。在本发明中,已阐明针对第一多肽的抗体或抗原结合剂能够结合游离和结合的第一多肽两种,稳定此类第一多肽的构型,所述多肽凭借结合第二多肽(诸如受体)来引发其生理反应。此类稳定的构型对第二多肽的结合亲和力比在所述抗体或抗原结合剂不存在的情况下更弱,因此,触发第一多肽自第二多肽的更快解离。在这方面,本发明提供了一个进一步的方面,其涉及能够结合游离和fcεri结合的ige且稳定ige的构型的抗体或抗原结合剂。当所述ige呈此类构型时,其对fcεri的结合亲和力比在所述抗体或抗原结合剂不存在的情况下更弱并且其中fcεri结合的人类ige自fcεri解离。任选地,当ige呈此类构型时,ige对奥马珠单抗或其片段的结合亲和力低于本发明的抗体或抗原结合剂。例如,所述抗体或抗原结合剂是如本文所述的抗体。在一个进一步的方面,本发明涉及用于选择如本文所述的此类抗体或抗原结合剂的方法。所述方法包括:a.使测试抗体或抗原结合剂与包含结合至人类fcεri的人类ige的样品接触;b.测量所述测试抗体或抗原结合剂将人类ige自人类fcεri解离的解离常数;c.将如步骤b)中测量的解离常数与奥马珠单抗或其片段将人类ige自人类fcεri解离的解离常数进行比较;d.如果抗体或抗原结合剂将ige自fcεri解离比奥马珠单抗或其片段更快,则选择所述抗体或抗原结合剂。或者,根据本发明的用于选择抗体或抗原结合剂的方法包括:a.使测试抗体或抗原结合剂与包含结合至人类fcεri的人类ige的样品接触;b.测量所述测试抗体或抗原结合剂对来自人类fcεri的人类ige的结合亲和力;c.将如步骤b)中测量的结合亲和力与人类ige对fcεri的结合亲和力进行比较;d.如果抗体或抗原结合剂对ige的结合亲和力高于ige对fcεri的结合亲和力,则选择所述抗体或抗原结合剂。任选地,所选抗体或抗原结合剂使仍结合至fcεri的ige采取这样的构型,其中所述稳定构型的ige可以比在奥马珠单抗或其片段存在的情况下结合至fcεri的ige更快自fcεri解离;和/或对所述抗体或抗原结合剂的结合亲和力可以高于对fcεri的结合亲和力。在最后一个方面,本发明涉及特定抗体或抗原结合剂,其包含:a.包含seq.idno.:1的重链可变区和包含以下的轻链可变区:i.seq.idno.:109;或ii.seq.idno.:113;或iii.seq.idno.:121;或iv.seq.idno.:132;或v.seq.idno.:139;或b.seq.idno.:5和i.seq.idno.:24,其中s77和s79被q置换;ii.seq.idno.:117或iii.seq.idno.:125;或iv.seq.idno.:136;或v.seq.idno.:143。在本发明的该最后一个方面的一个实施方案中,所述抗-ige抗体或抗原结合剂接触参照seqidno:108包含人类ige的cε3结构域的残基t373、w374、s375、r376、a377、s378、g379、p381、q417、c418、r419、t421、p426、r427、a428和cε2结构域的残基d278和t281的表位,或接触所述表位且对其是特异性的。附图简述参照物和seqid见于参照附图的实施例中。图1.奥马珠单抗fab3含有三个点突变。奥马珠单抗fab3衍生自奥马珠单抗,并在抗原结合cdr的远端含有三个点突变,两个在vl结构域构架区中(ser81arg、gln83arg)且一个在cκ结构域中(leu158pro)。重链和轻链分别呈白色和蓝色。突变的残基呈红色,且crdl1呈绿色,以指示fab的取向。图2.ige-fc与奥马珠单抗fab3的复合物的整体结构。(a)奥马珠单抗fab3以2:1化学计算量结合ige-fc。fab1(绿色)仅仅通过cε3结构域接合ige-fcb链(粉红色)。fab2(蓝色)通过cε3结构域与ige-fca链(黄色)相互作用并与来自ige-fcb链(粉红色)的cε2结构域形成微小相互作用。(b)两个fab关于fcε3-4区的二重轴形成伪对称复合物。为清楚起见,未显示cε2结构域。(c)ige-fc在奥马珠单抗fab3复合物中不对称地弯曲。来自b链(粉红色)的cε2结构域接触fab2(蓝色)。图3.奥马珠单抗fab3和ige-fc之间的界面。显示奥马珠单抗fab3fab2(重链和轻链分别呈绿色和黄色)和来自ige-fc的cε3(粉红色)之间的界面。奥马珠单抗fab3和cε3结构域残基标记分别呈蓝色和黑色。该界面包含氢键和范德华相互作用。该界面的一个值得注意的特征是arg419(cε3结构域)和phe103(奥马珠单抗fab3cdrh3)之间的阳离子-π相互作用。phe103侧链大多数埋于由thr373、trp374、ser375、gln417和arg419(cε3结构域)产生的口袋中。图4.奥马珠单抗fab3和darpine2_79结合重叠界面。奥马珠单抗fab3和darpine2_7920两者都结合cε3结构域。仅形成奥马珠单抗fab3界面的一部分的ige-fc残基呈橙色,而仅形成darpine2_79界面的一部分且包括cε3-cε4接头的一部分的残基呈蓝色。呈粉红色的ige-fc残基,包括arg419和met430,是奥马珠单抗fab3和darpine2_79界面共有的。图5.ige-fc的构型柔性。(a)游离ige-fc8的侧视图,显示其尖锐的不对称弯曲。(b)游离ige-fc的正视图(由(a)中显示的视图逆时针旋转90°)。(c)来自奥马珠单抗fab3复合物中ige-fc的侧视图,其揭示部分弯曲构型。(d)奥马珠单抗fab3复合物中ige-fc的正视图(由(c)中显示的视图逆时针旋转90°)。(e)由抗-ige-fcfab(奥马珠单抗fab3)16捕获的完全伸展的ige-fc的侧视图。(f)伸展的ige-fc的正视图(由(e)中显示的视图逆时针旋转90°)。图6.ige-fc的构型柔性。先前已通过分子动力学16研究ige-fc的柔性,且从弯曲形式伸直成完全伸展的构型。如先前所述16,ige-fc伸直表示为自由能表面。(a)ige-fc的伸展构型捕获于奥马珠单抗fab3/ige-fc复合物的晶体结构中16。(b)在奥马珠单抗fab3/ige-fc复合物的晶体结构中观察到部分弯曲的ige-fc构型。(c)游离ige-fc的弯曲构型7,8。ige-fc的弯曲构型占据最低能量池,而在奥马珠单抗fab3/ige-fc复合物中观察到的部分弯曲的构型占据明显不同的能量池(b)。图7.ige-fc和fcεri之间的相互作用的破坏。在奥马珠单抗fab3复合物中,cε3结构域采用迄今为止关于ige-fc所报导的最开放构型,其妨碍与fcεriα的接合。在与fcεriα8的复合物中ige-fc的结构呈黄色,且指出受体接合的两个子位点。在与ige-fc的复合物中奥马珠单抗fab3的结构重叠于cε4结构域上,且cε3结构域呈蓝色。指出该两个结构中his424和pro426的位置,以突出显示cε3结构域所采取的不同位置。图8.ige-fc和cd23之间的相互作用的破坏。(a)奥马珠单抗fab3和cd23界面两者共有的cε3结构域残基呈粉红色。(b)来自奥马珠单抗fab3/ige-fc复合物和cd23/fcε3-4复合物11的cε3结构域(呈深灰色)的叠加揭示cd23(黄色)和奥马珠单抗fab3(粉红色)之间的碰撞。图9.奥马珠单抗fab3与ige-fc的相互作用研究。(a)经由c末端his标签捕获的奥马珠单抗fab3与ige-fc的结合;奥马珠单抗fab3在以下浓度下流过ige-fc:100nm(黑色)、50nm(红色)、25nm(绿色)、12.5nm(蓝色)、6.2nm(青色)、3.1nm(紫色)、1.6nm(洋红色)和0.8nm(深红色)。采用标准双参照法36;每种浓度均一式两份运行。(b)第二奥马珠单抗fab3结合位点的结合使用spr夹心结合实验表征。将ige-fc捕获于奥马珠单抗fab3表面上,且然后将以下浓度的第二奥马珠单抗fab3分子添加至ige-fc/奥马珠单抗fab3复合物:1000nm(黑色)、500nm(红色)、250nm(绿色)、125nm(蓝色)、62.5nm(青色)、31.2nm(紫色)、15.6nm(洋红色)、7.8nm(深红色)和0nm(深蓝色)。(c)由c末端his标签(红色)捕获的奥马珠单抗fab3结合ige-fc的能力和通过结合fcεriα(蓝色)捕获的ige-fc的比较;测试其各自的两倍连续稀释液,其中最高浓度为1000nm。插图显示奥马珠单抗fab3仍可以结合ige-fc/fcεriα复合物,但具有低bmax值。(d)通过增加奥马珠单抗fab3的浓度介导的ige-fc/fcεriα复合物的加速解离。首先通过将ige-fc捕获在固定的fcεriα上且然后结合以下浓度的奥马珠单抗fab3来确定1:1ige-fc/fcεriα复合物:5000nm(洋红色)、1000nm(紫色)、200nm(青色)、40nm(蓝色)、8nm(绿色)、1.6nm(红色)和0nm(黑色)。插图显示加速解离过程的放大图。所有浓度均一式两份运行。所有结合实验均在25℃下进行,除了表征第二奥马珠单抗fab3结合位点的实验(图4b),所述实验在5℃下进行以使两个位点之间的变构通信最小化。图10.直接结合、竞争实验和加速解离的分析。测量ige-fc与固定的奥马珠单抗fab3(a)、奥马珠单抗fab(b)和完整奥马珠单抗(c)的直接结合。使用胺偶联试剂盒(gehealthcare)将fab或完整抗体以低密度共价固定;使用最高浓度为100nm的两倍连续稀释液,使各种浓度的ige-fc流过这些表面。所有浓度均一式两份运行。(d)奥马珠单抗fab3和αγ-融合蛋白质之间关于ige-fc的tr-fret竞争结合实验。用递增浓度的未标记奥马珠单抗fab3作为抑制剂测量铽标记的αγ-融合蛋白质和alexafluor647标记的ige-fc之间的结合:0μm(黑色)、2.5nm(蓝色)、5nm(绿色)、10nm(洋红色)、20nm(红色)。作为抑制剂,奥马珠单抗fab同时影响ige-fc和αγ-融合蛋白质之间的表观kd和bmax,指示一些变构抑制特性。(e)由浓度各自为5μm的完整奥马珠单抗(黑色)、奥马珠单抗fab(红色)或奥马珠单抗fab3(蓝色)介导的ige-fc/sfcεriα复合物的加速解离的比较。图11.代表性电子密度图。显示a链cε3结构域的一部分和在asn394处共价n连接的寡糖部分在1.1σ处形成波状外形的2fo-fc电子密度图的立体视图。图12.ige-fc自固定的sfcεriα解离的biacore传感图。监测在运行缓冲液(实线)或ige结合配偶体(所有其他传感图)存在的情况下的解离。如测定方法(1)实施例2中所述进行测定。图13.ige-fc自固定的sfcεriα解离的biacore传感图。监测在对照上清液(实线)或ige结合配偶体(所有其他传感图)存在的情况下的解离。如测定方法(2)实施例2中所述进行测定。图14.alexa488标记的ige-fc自rbl-sx38细胞的表面解离的分析。测量的结合数据针对在t=0时的100%归一化且解离数据绘制为仍然结合的ige-fc的比例随时间的变化。图15.野生型奥马珠单抗fab和奥马珠单抗fab3的治疗性给药对72小时pca模型的影响的分析。图16.野生型人类ige-fc序列(如seqidno:108中所示)的残基224至547且残基224和547以粗体显示。编号根据dorringtonandbennich(1978)immunol.rev.41:3-25,其中在l253之后的l(leu;亮氨酸)编号为l235a(加框)且随后的残基为c254。剩余残基连续编号且无进一步添加。表位残基以星号(*)显示。发明详述本文的抗体氨基酸编号将来自抗体的连续氨基酸序列(例如,奥马珠单抗包含seq.idno.1的vh序列和seq.idno.20或seqidno:129的vl序列)所谓的“pdb”编号或者可以利用常用的kabat编号方案。在描述vh或vl序列的常见免疫球蛋白部分(cdr-互补决定区,或fr-构架区)时,其以标准顺序连接(vh=fr-h1.cdr-h1.fr-h2.cdr-h2.fr-h3.cdr-h3.fr-h4;vl=fr-l1.cdr-l1.fr-l2.cdr-l2.fr-l3.cdr-l3.fr-l4)。对于奥马珠单抗,vh(seq.idno.1)部分的“pdb”编号为:fr-h1(氨基酸1-25)、cdr-h1(26-36)、fr-h2(37-50)、cdr-h2(51-66)、fr-h3(67-98)、cdr-h3(99-110)、fr-h4(111-121);而kabat编号为:fr-h1(氨基酸1-25)、cdr-h1(26-35)、fr-h2(36-49)、cdr-h2(50-65)、fr-h3(66-94)、cdr-h3(95-102)、fr-h4(103-113)。对于奥马珠单抗,vl(seq.idno.20)部分的“pdb”编号为:fr-l1(氨基酸1-23)、cdr-l1(24-38)、fr-l2(39-53)、cdr-l2(54-60)、fr-l3(61-92)、cdr-l3(93-101)、fr-l4(102-111);而kabat编号为:fr-l1(氨基酸1-23)、cdr-l1(24-34)、fr-l2(35-49)、cdr-l2(50-56)、fr-l3(57-88)、cdr-l3(89-97)、fr-l4(98-107)。ige抗体编号如dorrington&bennich(1978)immunol.rev.41:3-25)所报导。因此,本发明中使用的ige-fc多肽(参见seq.idno.108)来自v224-k547(包括c225a突变)。如图16中所示,遵循的编号根据dorrington和bennich(1978)immunol.rev.41:3-25,其中在位置253之后的l(leu,亮氨酸)编号为l253a且剩余残基从l253a连续编号为c254等。在结晶学实验中,还将以下突变插入ige-fc中以简化糖基化模式:n265q和n371q。一般公认ige-fc的cε2区占据序列s226-d330。本文中提及ige可以参照人类ige(且反之亦然),并在本文所描述的测定和方法的上下文中,也可以构成对ige-fc的参考。全长人类ige抗体的fab臂序列不包括在本说明书中,因为其不存在于晶体结构中。本文中提及的“奥马珠单抗”是参照商售的产品;或含有包含作为seq.idno.1的vh氨基酸序列的重链和包含作为seq.idno.20的vl氨基酸序列的轻链的igg全长抗体;或含有包含作为seq.idno.5的vh-ch1氨基酸序列的重链和包含作为seq.idno.24的vl-cl氨基酸序列的轻链的igg全长抗体;或含有包含作为seq.idno.9的vh-ch1-fc氨基酸序列的重链和包含作为seq.idno.24的vl-cl氨基酸序列的轻链的igg全长抗体。提及“奥马珠单抗fab”参照含有包含作为seq.idno.1的vh氨基酸序列的重链和包含作为seq.idno.20的vl氨基酸序列的轻链的fab片段;或(特别是)含有包含作为seq.idno.5的vh-ch1氨基酸序列的重链和包含作为seq.idno.24的vl-cl氨基酸序列的轻链的fab片段。一般定义除非另外规定,否则本文中所用的所有技术及科学术语均具有与本发明所属领域的本领域普通技术人员通常所理解相同的含义。尽管相似或等效于本文所描述的那些方法及材料的方法及材料可用于实践或测试本发明,但本文描述合适方法及材料。所有公开、专利申请、专利及本文提及的其他参考文献以其整体通过引用并入本文中。此外,材料、方法以及实例仅为说明性的和并不欲限制。除非另外为情形所需,否则单数术语应包括复数和复数术语应包括单数。在本申请中,除非另外规定,否则“或”的使用意指“和/或”。此外,使用术语“包括”以及其他形式(诸如“包含”及“含有”)不具限制性。此外,除非另外具体规定,否则诸如“元件”或“组件”的术语涵盖包含一个单元的元件及组件以及包含一个以上子单元的元件及组件两者。一般而言,本文中所述的与细胞及组织培养、分子生物学、免疫学、微生物学、遗传学及蛋白质与核酸化学及杂交结合使用的命名法及其技术为此项技术中众所周知和常用的命名法及技术。除非另外指明,否则本发明的方法及技术一般根据本领域中众所周知的习知方法和如本说明书通篇所引用及讨论的各种一般及更特定文献中所述来进行。酶促反应及纯化技术可根据制造商的说明书或如通常在本领域中所实现或如本文所描述来进行。本文中所述的与分析化学、合成有机化学及药物与医药化学结合使用的命名法及其实验室程序与技术为在本领域中众所周知和常用的命名法及实验室程序与技术。标准技术用于化学合成、化学分析、医药制备、调配及递送及治疗患者。为更容易地理解本发明,下文定义所选术语。如本文所用,术语“宿主”通常是指人受试者,和尤其在人或人源化构架用作受体结构的情况下。在治疗另一宿主的情况下,本领域技术人员应了解抗体或抗原结合剂可需要针对宿主调整以避免排斥或使其更相容性。已知针对一系列宿主的所需递送及功能,如何在本发明中使用cdr和将其工程改造至适当构架或肽序列中。其他宿主可包括其他哺乳动物或脊椎动物物种。因此,术语“宿主”或者可指诸如小鼠、猴、狗、猪、兔、家养猪(猪及肉猪)、反刍动物、马类动物、家禽、猫科动物、鼠类动物、牛类动物、犬科动物及其相似动物的动物,其中必要时抗体或抗原结合剂经适当设计以与宿主相容。如本文所用,术语“多肽”是指氨基酸的任何聚合链。术语“肽”及“蛋白质”可与术语多肽互换使用且还指氨基酸的聚合链。术语“多肽”涵盖天然或人工蛋白质、蛋白质片段及蛋白质序列的多肽相似物。多肽可为单体或聚合物。如本文所用,术语“回收”是指通过分离(例如使用本领域中众所周知的蛋白质纯化技术)使得诸如多肽的化学物质基本上不含天然相关组分的过程。如本文所用,在提及抗体、蛋白质或肽与第二化学物质的相互作用时,术语“特异性结合(specificbinding)”或“特异性地结合(specificallybinding)”意指相互作用视化学物质上的特定结构(例如如下所定义的“抗原决定簇”或“表位”)存在而定;例如,抗体识别和结合特定蛋白质结构而非一般蛋白质。如果抗体对表位“a”具有特异性,则在含经标记的“a”及该抗体的反应中,含表位a(或未经标记的游离a)的分子的存在将减少结合该抗体的经标记的a的量。在本文中提到本发明的表位时,本发明的抗-ige抗体或抗原结合剂对所述表位是特异性的。如本文所用,术语“抗体”泛指由四条多肽链(两条重链(h)链及两条轻(l)链)构成的任何免疫球蛋白(ig)分子或保留ig分子的表位结合特征的至少一部分以允许特异性结合ige的其任何功能片段、突变体、变体或衍生物。此类突变体、变体或衍生物抗体形式为本领域中已知且在下文描述。下文讨论了其非限制性实施例。如果抗体能够特异性地与分子(或表位)反应使得将该分子(或表位)结合抗体,则称该抗体“能够结合”分子(或表位)。如本文所用,“单克隆抗体”意欲指相比于含有不同抗体的混合物的“多株”抗体制剂,共享通用重链及通用轻链氨基酸序列的抗体分子制剂,或至少保留ig分子的轻链表位结合特征的其任何功能片段、突变体、变体或衍生物。单克隆抗体可通过几种已知技术产生,所述技术如噬菌体、细菌、酵母或核糖体展示以及经典方法,例如杂交瘤来源的抗体(例如通过杂交瘤技术,诸如标准kohler及milstein杂交瘤方法((1975)nature256:495-497)制备的杂交瘤所分泌的抗体)。在全长抗体中,每一重链由重链可变区(本文中缩写为hcvr或vh)和重链恒定区(ch)构成。重链恒定区由四个结构域-ch1、铰链、ch2和ch3(重链γ、α和δ)或ch1、ch2、ch3及ch4(重链μ和ε)构成。每个轻链由轻链可变区(本文中缩写为lcvr或vl)和轻链恒定区(cl)构成。轻链恒定区由一个结构域cl构成。vh和vl区可进一步再分成高变区,称为互补决定区(cdr),穿插有称为构架区(fr)的更保守区。每个vh及vl由自氨基端至羧基端按以下顺序排列的三个cdr及四个fr构成:fr1、cdr1、fr2、cdr2、fr3、cdr3、fr4。免疫球蛋白分子可具有任何同种型/类别(例如igg、ige、igm、igd、iga和igy)、类别(例如igg1、igg2、igg3、igg4、iga1和iga2)或子类。如本文所用,术语“抗原结合剂”是指保留特异性结合抗原(例如ige)的能力的抗体的一个或多个片段或部分,或保留与抗原的所需结合能力的抗体片段的合成变体。已展示抗体的抗原结合功能可通过全长抗体的片段或某些部分或其变体进行。实例包括可特异性结合两种或更多种不同抗原或抗原的几个表位或不连续表位区域的双特异性、双重特异性及多特异性形式。抗原结合剂的非限制性实例包括(i)fab片段,由vl、vh、cl和ch1结构域组成的单价片段;(ii)f(ab’)2片段,包含在铰链区通过二硫桥键连接的两个fab片段的二价片段;(iii)由vh和ch1结构域组成的fd片段;(iv)由抗体单臂的vl和vh结构域组成的fv片段;(v)dab片段(ward等人,(1989)nature341:544-546;winter等人,pct公开号wo90/05144a1,通过引用并入本文中),其包含单一可变结构域;(vi)分离的互补决定区(cdr);(vii)抗体片段的融合物,诸如特征为免疫球蛋白的抗体片段融合物,例如双功能抗体、scab、双特异性、triabody、fab-fv、fab-fv-fv、tribody、(fab-fv)2-fc;和(viii)移植至诸如纤连蛋白或亮氨酸拉链的非免疫球蛋白构架上的抗体部分,诸如cdr或抗体环(参见binz等人(2005)nat.biotech.23:1257-1268,并入本文中)。此外,虽然fv片段的两个结构域vl和vh通过分开基因编码,但其可使用重组或其他方法,通过合成或天然存在的接头连接,所述接头使其制备成其中vl和vh区配对形成单价分子的单一蛋白质链(称为单链fv(scfv);参见例如bird等人(1988)science242:423-426;以及huston等人(1988)proc.natl.acad.sci.usa85:5879-5883)。还希望此类单链抗体涵盖于术语抗原结合剂内。还涵盖单链抗体的其他形式,诸如双功能抗体。双功能抗体为二价双特异性抗体,其中vh及vl结构域展示于单一多肽链上,但使用太短以致不允许两个结构域之间在同一链上配对的接头,从而迫使结构域与另一链的互补结构域配对和产生两个抗原结合位点(参见例如holliger,p.等人(1993)proc.natl.acad.sci.usa90:6444-6448;poljak,r.j.等人(1994)structure.2:1121-1123)。此类抗体结合部分为本领域中已知(kontermann及dubel编辑,antibodyengineering(2001)springer-verlag.newyork.790页(isbn3-540-41354-5)。如本文所用,术语“抗体构建体”是指包含一个或多个本发明的抗原结合部分连接于接头多肽或免疫球蛋白恒定结构域的多肽。连接多肽包含两个或更多个由肽键连接的氨基酸残基,并且用于连接一个或多个抗原结合部分。此类接头多肽为本领域中众所周知(参见例如holliger,p.等人,(1993)proc.natl.acad.sci.usa90:6444-6448;poljak,r.j.等人,(1994)structure2:1121-1123)。免疫球蛋白恒定结构域是指重链或轻链恒定结构域,例如人iga、igd、ige、igg或igm恒定结构域。重链和轻链恒定结构域氨基酸序列为本领域中已知。ig重链γ1恒定区和ig轻链λ和κ链的非限制性实例分别提供了于表8和6。此外,抗体或其抗原结合部分可为较大免疫粘附分子的一部分,所述免疫粘附分子通过抗体或抗体部分与一种或多种其他蛋白质或肽的共价或非共价缔合而形成。此类免疫粘附分子的实例包括使用链霉抗生物素蛋白核心区域制备四聚体scfv分子(kipriyanov,s.m.等人(1995)humanantibodiesandhybridomas6:93-101)和使用半胱氨酸残基、标记物肽和c端聚组氨酸标签制备二价和生物素标记的scfv分子(kipriyanov,s.m.等人(1994)mol.immunol.31:1047-1058)。诸如fab及f(ab’)2片段的抗体部分可由全抗体,使用众所周知技术,诸如对全抗体分别进行木瓜蛋白酶或胃蛋白酶消化来制备。此外,抗体、抗体部分和免疫粘附分子可使用如本文中所述的标准重组dna技术获得。如本文所用,“分离的抗体”意欲指这样的抗体,其实质上不含其他具有不同抗原特异性的抗体(例如特异性结合ige的分离的抗体实质上不含特异性结合除ige以外的抗原的抗体)。然而,特异性结合例如人ige的分离抗体可与诸如来自其他物种的ige分子的其他抗原具有交叉反应性。另外,分离的抗体可实质上不含其他细胞物质和/或化学物质。术语“cdr移植抗体”是指包含来自一种物种的重链及轻链可变区序列,但其中vh和/或vl的一个或多个cdr区的序列经另一物种的cdr序列置换的抗体,诸如具有其中一个或多个人cdr(例如cdr3)已经被鼠类cdr序列置换的人重链和轻链可变区的抗体。术语“kabat编号”、“kabat定义”及“kabat标记”在本文中可互换使用。本领域中公认的这些术语是指抗体或其抗原结合部分的重链及轻链可变区中比其他氨基酸残基更可变(即高变)的氨基酸残基的编号系统(kabat等人(1971)ann.nyacad,sci.190:382-391及kabat,e.a等人(1991)sequencesofproteinsofimmunologicalinterest,fifthedition,u.s.departmentofhealthandhumanservices,nih公开号91-3242)。对于重链可变区,根据kabat编号系统,高变区在氨基酸位置31-35(cdr-h1)、残基50-65(cdr-h2)及残基95-102(cdr-h3)范围内。然而,根据chothia(chothia等人,(1987)j.mol.biol.,196,901-917(1987)),等同于cdr-h1的环自残基26延伸至残基32。因此,除非另外规定,否则如本文所采用的“cdr-h1”意欲指如kabat编号系统与chothia拓扑环定义的组合所描述的,残基26至35。对于轻链可变区,高变区范围就cdrl1而言自氨基酸位置24至34,就cdrl2而言自氨基酸位置50至56,和就cdrl3而言自氨基酸位置89至97。如本文所用,术语“受体”和“受体抗体”是指提供或编码一个或多个构架区的氨基酸序列的至少80%、至少85%、至少90%、至少95%、至少98%或100%的抗体或核酸序列。在一些实施方案中,术语“受体”是指提供或编码恒定区的抗体氨基酸或核酸序列。在另一个实施方案中,术语“受体”是指提供或编码构架区及恒定区中的一个或多个的抗体氨基酸或核酸序列。在一个特定实施方案中,术语“受体”是指提供或编码一个或多个构架区的氨基酸序列的至少80%、优选至少85%、至少90%、至少95%、至少98%或100%的人抗体氨基酸或核酸序列。根据此实施方案,受体可含有至少1个、至少2个、至少3个、至少4个、至少5个或至少10个不出现于人抗体的一个或多个特定位置的氨基酸残基。受体构架区和/或一个或多个受体恒定区可例如源自或获自种系抗体基因、成熟抗体基因、功能性抗体(例如本领域中众所周知的抗体、处于开发中的抗体或市售抗体)。如本文所用,术语“cdr”是指在抗体可变序列内的互补决定区。重链和轻链的各可变区中存在三个cdr,对于重链cdr而言其称为cdrh1、cdrh2和cdrh3,和对于轻链cdr而言其称为cdrl1、cdrl2和cdrl3。如本文所用,术语“cdr组”是指存在于能够结合抗原的单一可变区中的一组三个cdr。这些cdr的准确边界已根据不同系统不同地加以界定。由kabat(kabat等人,sequencesofproteinsofimmunologicalinterest(nationalinstitutesofhealth,bethesda,md.(1987)和(1991))描述的系统不仅提供了适用于抗体的任何可变区的明确的残基编号系统,且还提供了界定三个cdr的精确残基边界。这些cdr可称为kabatcdr。chothia及同事(chothia及lesk,j.mol.biol.196:901-917(1987)和chothia等人,nature342:877-883(1989))发现,kabatcdr内的某些子部分采用几乎同一的肽主链构型,尽管在氨基酸序列水平上具有极大差异。这些子部分称为l1、l2和l3或h1、h2和h3,其中“l”和“h”分别表示轻链及重链区。这些区域可称为chothiacdr,其具有与kabatcdr重叠的边界。界定与kabatcdr重叠的cdr的其他边界已由padlan(fasebj.9:133-139(1995))及maccallum(jmolbiol262(5):732-45(1996))描述。其他cdr边界界定可不严格遵循以上系统中的一个,但仍然将与kabatcdr重叠,除了其可根据特定残基或残基组或甚至全部cdr不显著影响抗原结合的预测或实验发现进行缩短或延长。本文中使用的方法可利用根据这些系统任一个所界定的cdr,除了优选实施方案使用kabat或chothia或其混合物界定的cdr。如本文所用,术语“典型”残基是指cdr或构架中界定如chothia等人(j.mol.biol.196:901-907(1987);chothia等人,j.mol.biol.227:799(1992),均通过引用并入本文中)所定义的特定典型cdr结构的残基。根据chothia等人,多种抗体的cdr的重要部分具有几乎同一的肽主链构型,尽管在氨基酸序列水平上具有极大差异。各典型结构主要给形成环的氨基酸残基的相邻区段指定一组肽主链扭转角。如本文所用,术语“供体”及“供体抗体”是指提供了一个或多个cdr的抗体。在一个优选实施方案中,供体抗体为来自不同于获得或产生构架区的抗体的物种的抗体。在人源化抗体的情况下,术语“供体抗体”是指提供了一个或多个cdr的非人抗体。如本文所用,术语“构架”或“构架序列”是指可变区减去cdr的剩余序列。因为cdr序列的准确定义可通过不同系统确定,所以构架序列的含义相对应地经历不同解释。六个cdr(轻链的cdr-l1、cdr-l2和cdr-l3以及重链的cdr-h1、cdr-h2和cdr-h3)还将轻链和重链上的构架区分成各链上四个子区(fr1、fr2、fr3和fr4),其中cdr1位于fr1和fr2之间,cdr2位于fr2和fr3之间,且cdr3位于fr3和fr4之间。在不指定特定子区为fr1、fr2、fr3或fr4的情况下,如他人所提及,构架区表示单一天然存在的免疫球蛋白链的可变区内的组合fr。如本文所用,一个fr表示四个子区之一,且多个fr表示构成构架区的四个子区中的两个或更多个。如本文所用,术语“种系抗体基因”或“基因片段”是指由未经历导致遗传重排及突变以供特定免疫球蛋白表达的成熟过程的非淋巴细胞编码的免疫球蛋白序列。参见例如shapiro等人,crit.rev.immunol.22(3):183-200(2002);marchalonis等人,advexpmedbiol484:13-302001)。本发明的各个实施方案提供的优点之一利用以下认识:种系抗体基因比成熟抗体基因更可能保存作为个体在物种中的特征的基础氨基酸序列结构,因此当在治疗学上用于该物种中时不大可能识别为来自外来来源。如本文所用,术语“关键”残基是指可变区内对抗体、尤其人源化抗体的结合特异性和/或亲和力更具影响的某些残基。关键残基包括但不限于以下中的一个或多个:与cdr相邻的残基、潜在糖基化位点(可为n-或o-糖基化位点)、稀少残基、能够与抗原相互作用的残基、能够与cdr相互作用的残基、典型残基、重链可变区与轻链可变区之间的接触残基、维尼尔区(vernierzone)内的残基和在可变重链cdr1的chothia定义与第一重链构架的kabat定义之间重叠的区域中的残基。术语“人源化抗体”一般是指包含来自非人物种(例如兔、小鼠等)的重链和轻链可变区序列,但其中vh和/或vl序列的至少一部分已改变成更“人样”,即更相似于人种系可变序列的抗体。一种类型人源化抗体为其中人cdr序列引入非人vh及vl序列中以替换对应非人cdr序列的cdr移植抗体。另一类型人源化抗体为其中至少一个非人cdr插入人构架中的cdr移植抗体。后者通常为本发明的焦点。具体地,如本文所用,术语“人源化抗体”为免疫特异性结合相关抗原和包含具有实质上人抗体的氨基酸序列的构架(fr)区及具有实质上非人抗体的氨基酸序列的互补决定区(cdr)的抗体或其变体、衍生物、相似物或片段。如本文所用,在cdr的情况下术语“实质上”是指氨基酸序列与非人抗体cdr的氨基酸序列至少50%、55%、60%、65%、70%、75%或80%、优选至少85%、至少90%、至少95%、至少98%或至少99%同一的cdr。在一个实施方案中,人源化抗体具有与非人抗体cdr相比,具有一个或多个(例如1、2、3或4个)氨基酸取代、添加和/或缺失的cdr区。此外,非人cdr可使用已知技术进行工程改造以更“人样”或与人体相容。人源化抗体包含实质上所有的至少一个和通常两个可变结构域(fab,fab′,f(ab’)2,f(ab’)c,fv),其中所有或实质上所有cdr区对应于非人免疫球蛋白(即供体抗体)的cdr区和所有或实质上所有构架区为人免疫球蛋白共同序列的构架区。优选地,人源化抗体还包含至少一部分免疫球蛋白恒定区(fc),通常人免疫球蛋白的恒定区。在一些实施方案中,人源化抗体含有轻链以及至少重链的可变结构域。抗体还可包括重链的ch1、铰链、ch2和ch3,或ch1、ch2、ch3和ch4。在一些实施方案中,人源化抗体仅仅含有人源化轻链。在一些实施方案中,人源化抗体仅仅含有人源化重链。在特定实施方案中,人源化抗体仅仅含有轻链和/或人源化重链的人源化可变结构域。尽管本文所讨论的突变中的一些可能通常不是“人类”的,但这些不足以使本发明的抗-ige抗体或抗原结合剂不是“人源化”的。人源化抗体可选自任何类别免疫球蛋白,包括igy、igm、igg、igd、iga和ige和任何同种型,包括但不限于igal、iga2、igg1、igg2、igg3及igg4。人源化抗体可包含来自超过一种类别或同种型的序列,且可使用本领域中众所周知的技术选择特定恒定结构域以优化所需效应功能。人源化抗体的构架和cdr区无需准确对应于亲本序列,例如供体抗体cdr或共同构架可通过至少一个氨基酸残基取代、插入和/或缺失而诱变,使得该位点的cdr或构架残基不准确对应于供体抗体或共同构架。然而,在一个优选实施方案中,此类突变将不为大规模的。通常,人源化抗体残基的至少50%、55%、60%、65%、70%、75%或80%、优选至少85%、更优选至少90%和最优选至少95%、98%或99%将对应于亲本fr及cdr序列的残基。在一个实施方案中,与亲本fr和cdr序列相比(例如与奥马珠单抗或奥马珠单抗fab序列相比),一个或多个(例如1、2、3、4、5、6、7、8、9或10个)氨基酸取代、添加和/或缺失可存在于人源化抗体中。如本文所用,术语“共同构架”是指共同免疫球蛋白序列中的构架区。如本文所用,术语“共同免疫球蛋白序列”是指相关免疫球蛋白序列家族中由最频繁出现的氨基酸(或核苷酸)形成的序列(参见例如winnaker,fromgenestoclones(verlagsgesellschaft,weinheim,germany1987)。在免疫球蛋白家族中,共同序列中的每个位置由该家族中最频繁出现于该位置的氨基酸占据。如果两个氨基酸同等频繁地出现,则共同序列中可包括任一个。如本文所用,“维尼尔”区是指如foote及winter(1992,j.mol.biol.224:487-499,其通过引用并入本文中)所描述,可调节cdr结构和微调与抗原的合适度的构架残基子集。维尼尔区残基形成在cdr下面的层并且可影响cdr结构及抗体亲和力。如本文所用,术语“中和”是指当本文中描述的本发明的抗ige抗体或抗原结合剂特异性结合ige蛋白质时中和ige的生物活性。中和可为该抗体与ige的不同结合方式的结果。优选地,中和抗体为结合ige,中和ige生物活性的抗体。优选地,中和结合蛋白质结合ige和将ige生物活性降低至少约5%、10%、15%、20%、25%、30%、35%、40%、50%、60%、80%、85%或更多。中和抗体对ige生物活性的中和可通过测量本文中描述的ige生物活性的一个或多个指示物来评估。如本文所用的“中和单克隆抗体”意欲指在结合ige后能够部分或完全抑制或降低ige生物活性的抗体分子制剂。如本文所用,术语“减弱(attenuation)”、“减弱(attenuate)”及其相似术语是指由血清ige水平升高所引起的症状或病况的严重程度减轻或降低。术语“表位”或“抗原决定簇”包括能够特异性结合免疫球蛋白或t细胞受体的任何多肽决定簇。在某些实施方案中,表位决定簇包括分子的化学活性表面分组(诸如氨基酸、糖侧链、磷酰基或磺酰基),且在某些实施方案中,可具有特定三维结构特征和/或特定电荷特征。表位为抗体所结合的抗原区域。在某些实施方案中,在蛋白质和/或大分子的复杂混合物中,当抗体优先识别其标靶抗原时,称该抗体特异性结合抗原。如本文所用,术语“kon”意欲指抗体与抗原缔合以形成如本领域中已知的抗体/抗原复合物的缔合速率常数。如本文所用,术语“koff”意欲指抗体自如本领域中已知的抗体/抗原复合物解离的解离速率常数。如本文所用,术语“kd”或“kd”意欲指如本领域中已知的特定抗体-抗原相互作用的解离常数。免疫结合相互作用的强度或亲和力可根据相互作用的解离常数(kd或kd)表示,其中kd越小,代表亲和力越大或越高。所选多肽的免疫结合特性可使用本领域中众所周知的方法定量。一种此类方法包括测量抗原结合位点/抗原复合物形成及解离的速率,其中这些速率视复合配偶体的浓度、相互作用的亲和力及同等影响两个方向上速率的几何结构参数而定。因此,“缔合速率常数”(“kon”)及“解离速率常数”(“koff”)可通过计算浓度及缔合与解离的实际速率来确定。(nature361:186-87(1993))。koff/kon的比率能够抵消不与亲和力相关的所有参数,且等于解离常数kd。davies等人(1990)annualrevbiochem59:439-473。术语“抗体缀合物”是指化学上连接至第二化学部分诸如治疗剂或细胞毒性剂的结合蛋白质,诸如抗体或抗体片段或其结合部分。术语“药剂”在本文中用以表示化合物、化合物的混合物、生物大分子或由生物材料制成的提取物。如本文所用,术语“晶体”及“结晶”是指呈晶体形式存在的抗体或其抗原结合部分。晶体为物质的一种固态形式,其不同于诸如非晶形固态或液晶态的其他形式。晶体由原子、离子、分子(例如蛋白质,诸如抗体)或分子集合体(例如,抗原/抗体复合物)的规则、重复、三维阵列构成。这些三维阵列根据本领域中充分了解的特定数学关系排列。在晶体中重复的基本单元或建构嵌段称为不对称单元。不对称单元以符合给定的良好定义的晶体学对称性的排列重复提供了晶体的“单位晶胞”。单位晶胞在所有三个维度上通过规则变换进行重复提供了晶体。参见giege,r.及ducruix,a.barrett,crystallizationofnucleicacidsandproteins,apracticalapproach,第2版,第20页1-16,oxforduniversitypress,newyork,n.y.,(1999)”。如本文中所提及,术语“多核苷酸”意指两种或更多种核苷酸,核糖核苷酸或脱氧核苷酸或任一核苷酸类型的修饰形式的聚合物形式。术语包括dna的单链和双链形式,但优选为双链dna。如本文所用,术语“分离的多核苷酸”意指一种多核苷酸(例如基因组、cdna或合成来源,或其某种组合),根据其来源,该“分离的多核苷酸”与在自然界中与“分离的多核苷酸”一起发现的多核苷酸的全部或一部分不相关;与在自然界中不与其连接的多核苷酸可操作地连接;或不作为较大序列的一部分存在于自然界中。如本文所用,术语“载体”意欲指能够运输其已连接的另一种核酸的核酸分子。一种载体类型为“质粒”,其是指其中可连接其他dna区段的环形双链dna环。另一类型的载体为病毒载体,其中其他dna片段可连接至病毒基因组。某些载体能够在引入其的宿主细胞中自发复制(例如具有细菌复制起点的细菌载体及游离型哺乳动物载体)。其他载体(例如非游离型哺乳动物载体)当引入至宿主细胞中时可整合至宿主细胞的基因组中,且进而与宿主基因组一起复制。此外,某些载体能够引导其以可操作方式连接的基因表达。此类载体在本文中称作“重组表达载体”(或简称为“表达载体”)。一般而言,用于重组dna技术中的表达载体常常呈质粒形式。在本说明书中,由于质粒为最常用的载体形式,因此“质粒”与“载体”可互换使用。然而,本发明意欲包括提供了同等功能的此类其他形式表达载体,诸如病毒载体(例如复制缺陷型反转录病毒、腺病毒及腺相关病毒)。术语“可操作地连接”是指所描述的组分处于允许其以其预期方式起作用的关系中的并置。“可操作地连接”至编码序列的控制序列是以使编码序列的表达在与控制序列相容的条件下实现的方式连接。“可操作地连接”的序列包括与相关基因相邻的表达控制序列及以反式作用或以一定距离作用以控制相关基因的表达控制序列。如本文所用,术语“表达控制序列”是指实现所连接的编码序列的表达及加工所需的多核苷酸序列。表达控制序列包括适当转录起始、终止、启动子及增强子序列;有效rna加工信号,诸如剪接及多腺苷酸化信号;使细胞质mrna稳定的序列;增强翻译功效的序列(即科扎克共同序列(kozakconsensussequence));增强蛋白质稳定性的序列;及必要时增强蛋白质分泌的序列。此类控制序列的性质视宿主生物体而不同;在原核生物中,此类控制序列一般包括启动子、核糖体结合位点及转录终止序列;在真核生物中,此类控制序列一般包括启动子及转录终止序列。术语“控制序列”意欲包括其存在对表达及加工而言至关重要的组件,且还可包括其存在有利的其他组件,例如前导序列及融合配偶体序列。如本文所定义的“转化”是指外源性dna进人宿主细胞的任何过程。转化可在天然或人工条件下使用本领域中众所周知的各种方法进行。转化可依赖于将外来核酸序列插入原核或真核宿主细胞中的任何已知方法。方法基于转化的宿主细胞选择和可包括但不限于病毒感染、电穿孔、脂质体转染和粒子轰击。此类“转化”细胞包括插入的dna能够作为自主复制质粒或作为宿主染色体的一部分复制的稳定转化细胞。其还包括短暂表达插入的dna或rna持续有限时间段的细胞。如本文所用,术语“重组宿主细胞”(或简称为“宿主细胞”)意欲指已引入外源性dna的细胞。应了解,此类术语不仅意指特定受试者细胞,而是指此类细胞的后代。因为某些修饰可能因突变或环境影响而出现在后代中,所以此类后代可能实际上不与亲本细胞同一,但仍包括于如本文所使用的术语“宿主细胞”范畴内。优选地,宿主细胞包括选自任一生命界的原核及真核细胞。优选真核细胞包括原生生物、真菌、植物和动物细胞。最优选地,宿主细胞包括但不限于原核细胞系大肠杆菌;哺乳动物细胞系cho、hek293及cos;昆虫细胞系sf9;以及真菌细胞酿酒酵母(saccharomycescerevisiae)。对重组dna、寡核苷酸合成和组织培养和转化可使用标准技术(例如电穿孔、脂质体转染)。酶促反应和纯化技术可根据制造商的说明书或如通常在本领域中所实现或如本文所描述来进行。前述技术及程序一般可根据本领域中众所周知及如在本说明书通篇中所引用及讨论的各种一般及更特定参考文献中所描述的众所周知方法进行。参见例如sambrook等人molecularcloning:alaboratorymanual(第2版,coldspringharborlaboratorypress,coldspringharbor,n.y,(1989)),其通过引用并入本文中以实现任何目的。如本文所用,术语“有效量”是指疗法足以降低或改善病症或其一种或多种症状的严重程度和/或持续时间;阻止病症进展;引起病症消退;防止与病症相关的一种或多种症状复发、出现、发作或进展;检测病症;或增强或改善另一疗法(例如预防剂或治疗剂)的预防或治疗效果的量。本文提供的人ige蛋白质的特定区域或表位可通过本领域中已知的任何适合的表位定位方法以及本发明提供的任一抗体来鉴定。此类方法的实例包括针对与本发明的抗体的结合,筛选衍生自ige的变化长度的肽,其中可特异性结合抗体的最小片段含有抗体所识别的表位的序列。ige肽可以以合成方式或通过蛋白分解消化ige蛋白质产生。结合抗体的肽可通过例如质谱分析来鉴定。在另一个实施方案中,nmr光谱分析或x射线结晶学可用于鉴定本发明的抗体所结合的表位。优选结晶及x射线结晶学技术用于确定ige的结构及本发明的抗ige抗体或抗原结合剂结合的ige上的表位。用于本发明的抗体还可使用单一淋巴细胞抗体方法,通过克隆及表达免疫球蛋白可变区cdna产生,免疫球蛋白可变区cdna自针对特异性抗体产生所选择的单一淋巴细胞,通过例如babcook,j.等人,1996,proc.natl.acad.sci.usa93(15):7843-78481;wo92/02551;wo2004/051268及国际专利申请号wo2004/106377描述的方法产生。可使用测量与人ige结合的测定法和/或测量阻断ige与其天然受体结合的能力的测定法筛选抗体。结合测定的一个实例为elisa。人源化抗体(其包括cdr移植抗体)为具有一个或多个来自非人物种(例如兔或小鼠)的互补决定区(cdr)和来自人免疫球蛋白分子的构架区的抗体分子(参见例如us5,585,089;wo91/09967)。应了解可能仅仅需要转变cdr的特异性决定残基而非整个cdr(参见例如kashmiri等人,2005,methods,36,25-34)。人源化抗体可任选地进一步包含一个或多个来源于cdr所来源的非人物种的构架残基。后者常常称为供体残基。本发明的抗体分子合适地具有小于2nm的结合亲和力(kd)。亲和力可使用本领域中已知的任何合适方法,包括如本文中的实施例中所述的biacore(参见实施例6),使用分离的天然或重组ige或适合融合蛋白质/多肽来测量。本发明的抗体或抗原结合剂的亲和力以及结合剂(诸如抗体)抑制结合的程度可通过本领域普通技术人员,使用常规技术,例如scatchard等人描述的技术(ann.ky.acad.sci.51:660-672(1949))或通过表面等离子共振(spr),使用诸如biacore的系统测定。对于表面等离子共振,标靶分子被固定于固相上和在沿流槽流动的移动相中暴露于配体。如果发生配体结合固定标靶,则局部折射率改变,导致spr角变化,其可通过检测反射光强度的变化而进行实时监测。可分析spr信号的变化速率以针对结合反应的缔合和解离相产生表观速率常数。这些值的比率产生表观平衡常数(亲和力)(参见例如wolff等人,cancerres.53:2560-65(1993))。应了解本发明提供的抗体的亲和力可使用本领域中已知的任何合适方法改变。因此,本发明还关于本发明的抗体分子的变体,其具有改善的对ige的亲和力。此类变体可通过许多亲和力成熟方案获得,包括使cdr突变(yang等人,j.mol.biol.,254,392-403,1995)、链改组(marks等人,bio/technology,10,779-783,1992)、使用大肠杆菌的增变基因菌株(low等人,j.mol.biol.,250,359-368,1996)、dna改组(patten等人,curr.opin.biotechnol.,8,724-733,1997)、噬菌体展示(thompson等人,j.mol.biol.,256,77-88,1996)和性pcr(crameri等人,nature,391,288-291,1998)。vaughan等人(上文)讨论这些亲和力成熟方法。人源化抗体和抗原结合剂在本发明的一个方面,本文提供了人源化抗-ige单克隆抗体和抗原结合剂。人源化抗体为其中重链和/或轻链含有一个或多个来自供体抗体(例如非人抗体,诸如鼠类或兔单克隆抗体)的cdr(必要时包括一个或多个修饰的cdr)移植至受体抗体(例如人抗体)的重链和/或轻链可变区构架中的抗体。关于综述,参见vaughan等人,naturebiotechnology,16,535-539,1998。在一个实施方案中,不是转移整个cdr,而是仅仅来自以上本文所述的任一cdr的一个或多个特异性决定残基被转移至人抗体构架(参见例如kashmiri等人,2005,methods,36,25-34)。在一个实施方案中,仅仅来自本文中描述的一个或多个cdr的特异性决定残基被转移至人抗体构架。在另一个实施方案中,仅仅来自本文中描述的各cdr的特异性决定残基被转移至人抗体构架。当移植cdr或特异性决定残基时,关于cdr衍生自的供体抗体的类别/类型,可使用任何适当受体可变区构架序列,包括小鼠、兔、灵长类动物及人构架区。合适地,根据本发明的人源化抗体具有包含人受体构架区以及一个或多个本文中特定提供的cdr的可变结构域。因此,一个实施方案中提供了一种结合人ige的人源化单克隆抗体,其中可变结构域包含人受体构架区(具有如本文所述的任选的突变)和非人供体cdr。cdr移植抗体的构建一般描述于欧洲专利申请ep-a-0239400中,该专利申请公开了一种方法,其中通过定点突变诱发,使用长寡核苷酸将小鼠单克隆抗体的cdr移植至人免疫球蛋白的可变结构域的构架区上,将该申请并入本文中。cdr决定抗体的抗原结合特异性并且是可变结构域的构架区上携带的相对较短的肽序列。关于通过cdr移植将单克隆抗体人源化的最早工作在识别诸如np的合成抗原的单克隆抗体上进行。然而,verhoeyen等人(science,239,1534-1536,1988)和riechmann等人(nature,332,323-324,1988)已分别描述其中识别溶菌酶的小鼠单克隆抗体和识别人t细胞上抗原的大鼠单克隆抗体通过cdr移植入源化的实例。抗体人源化通过非人抗体诸如小鼠、大鼠、山羊或兔抗体的cdr移植至“相似”人构架(受体)上和选择最少数目的人工选自供体单克隆抗体和并入人受体构架中的关键构架残基(回复突变)以维持原始cdr构型来实现。此类方法为本领域中已知,且包括以下中描述的这些方法:jones等人,nature321:522(1986);verhoeyen等人,science239:1534(1988)),sims等人,j.immunol.151:2296(1993);chothiaandlesk,j.mol.biol.196:901(1987),carter等人,proc.natl.acad.sci.u.s.a.89:4285(1992);presta等人,j.immunol.151:2623(1993),padlan,molecularimmunology28(4/5):489-498(1991);studnicka等人,proteinengineering7(6):805-814(1994);roguska.等人,pnas91:969-973(1994);pct申请wo91/09967,pct/:us98/16280,us96/18978,us91/09630,us91/05939,us94/01234,gb89/01334,gb91/01134,gb92/01755;wo90/14443,wo90/14424,wo90/14430,ep229246,ep592,106;ep519,596,ep239,400,u.s.专利号5,565,332,5,723,323,5,976,862,5,824,514,5,817,483,5,814,476,5,763,192,5,723,323,5,766,886,5,714,352,6,204,023,6,180,370,5,693,762,5,530,101,5,585,089,5,225,539;4,816,567,其均并入本文中。人可变重链和轻链种系子家族分类可衍生自kabat种系亚群名称:特定vh序列的vh1、vh2、vh3、vh4、vh5、vh6或vh7,以及构架4的特定可变重链连接组的jh1、jh2、jh3、jh4、jh5及jh6;构架1、2及3的特定vlκ序列的vk1、vk2、vk3、vk4、vk5或vk6,以及构架4的特定κ连接组的jk1、jk2、jk3、jk4或jk5;或构架1、2及3的特定vlλ序列的vl1、vl2、vl3、vl4、vl5、vl6、vl7、vl8、vl9或vl10,以及构架4的特定λ连接组的jl1、jl2、jl3或jl7。本发明的抗体分子的恒定区结构域如果存在则可根据抗体分子的所提出功能及尤其可能需要的效应功能进行选择。例如,恒定区结构域可为人iga、igd、ige、igg或igm结构域。在特定实施方案中,当抗体分子意欲用于治疗性用途和需要抗体效应功能时,可使用人igg恒定区结构域,尤其igg1和igg3同种型。或者,当抗体分子意欲用于实现治疗目的和不需要抗体效应功能时可使用igg2和igg4同种型。应了解还可使用这些恒定区结构域的序列变体。例如,可使用如angal等人,molecularimmunology,1993,30(1),105-108中所述,其中位置241的丝氨酸已改变成脯氨酸的igg4分子。本领域技术人员还了解抗体可进行多种翻译后修饰。这些修饰的类型及程度常常视用于表达抗体的宿主细胞系以及培养条件而定。此类修饰可包括糖基化变化、甲硫氨酸氧化、二酮哌嗪形成、天冬氨酸盐异构化及天冬酰胺去酰胺。常见修饰为由于羧基肽酶作用而丧失羧基端基础残基(诸如赖氨酸或精氨酸)(如harris,rj.journalofchromatography705:129-134,1995中所述)。因此,可缺乏抗体重链的c端赖氨酸。cdr及人构架修饰riechmann等人发现单独cdr的转移(如kabat(kabat等人(上文))和wu等人,j.exp.med.,132,211-250,1970)所定义)不足以在cdr移植产物中提供了令人满意的抗原结合活性。发现许多构架残基必须作出改变,使得其对应于供体构架区的残基。用于选择需要作出改变的构架残基的所提出的标准描述于国际专利申请wo90/07861中,该专利申请并入本文中。如果人可变结构域构架采用与cdr所来源的非人可变构架相同或相似的构型,则非人cdr至人可变结构域构架中的取代最可能保留cdr正确空间取向。此通过自构架序列展现与cdr所来源的非人可变构架结构域高度序列同一性的人抗体获得人可变结构域来实现。如上所述,重链及轻链可变构架区可衍生自相同或不同人抗体序列。人抗体序列可为天然存在的人抗体的序列或可为几种人抗体的共同序列。参见kettleborough等人,proteinengineering4:773(1991);kolbinger等人,proteinengineering6:971(1993)及carter等人,wo92/22653。已鉴定了非人供体免疫球蛋白及适当人受体免疫球蛋白的互补决定区,下一步是确定来自这些组件的哪些残基(如果存在)应经取代以优化所得人源化抗体的特性。一般而言,人氨基酸残基经非人氨基酸残基的取代应减至最少,因为非人残基的引入增加在人中抗体引发人-抗-供体-抗体(hada)反应的风险。可进行本领域中公认的确定免疫反应的方法以监测特定宿主中或临床试验期间的hada反应。在该疗法施用开始及整个过程中,可对施用人源化抗体的宿主进行免疫原性评估。例如通过使用本领域技术人员已知的方法,包括表面等离子共振技术(biacore)和/或固相elisa分析,检测来自宿主的血清样品中人源化治疗试剂的抗体来测量hada反应。部分通过计算机建模来选择用于取代(还有本文的“突变”)的氨基酸残基。在本文中描述了用于产生免疫球蛋白分子的三维图像的计算机硬件及软件。一般而言,分子模型自免疫球蛋白链或其结构域的解析结构开始产生。比较待建模的链与解析的三维结构的链或结构域的氨基酸序列相似性,且选择显示最大序列相似性的链或结构域作为构建分子模型的起点。选择共享至少50%序列同一性的链或结构域用于建模,和优选选择共享至少60%、70%、80%、90%序列同一性或更大序列同一性的链或结构域用于建模。解析的起始结构经修饰以允许建模的免疫球蛋白链或结构域中实际氨基酸与起始结构中的氨基酸之间的差异。然后经修饰的结构组装成复合免疫球蛋白。最终,模型通过能量最小化及通过验证所有原子在彼此适当距离内和键长及键角在化学上可接受的限度内来改进。选择氨基酸残基用于取代还可部分通过检查特定位置的氨基酸的特征,或特定氨基酸的取代或突变诱发的作用的实验观测结果来确定。例如,当供体可变区构架残基和所选人可变区构架残基之间的氨基酸不同时,当合理地预期氨基酸如下时人构架氨基酸通常应被来自供体抗体的同等构架氨基酸取代:(1)直接非共价结合抗原,(2)与cdr区相邻,(3)另外与cdr区相互作用(例如如通过计算机建模所测定,在cdr区的约3-6埃内),或(4)参与vl-vh界面。“直接非共价结合抗原”的残基包括构架区中根据已确立的化学力,例如通过氢键、范德华力(vanderwaalforce)、疏水相互作用及其相似作用,很可能直接与抗原上氨基酸相互作用的位置中的氨基酸。cdr和构架区如通过上文kabat等人或chothia等人所定义。当如通过上文kabat等人所定义的构架残基构成如上文chothia等人所定义的结构环残基时,可选择存在于供体抗体中的氨基酸用于取代至人源化抗体中。“与cdr区相邻”的残基包括紧邻人源化免疫球蛋白链的一级序列中一个或多个cdr的位置中,例如紧邻如kabat所定义的cdr或如chothia所定义的cdr的位置中的氨基酸残基(参见例如chothia及lesk1mb196:901(1987))。这些氨基酸尤其可能与cdr中的氨基酸相互作用,且如果选自受体,则使供体cdr变形和降低亲和力。此外,相邻氨基酸可直接与抗原相互作用(amit等人,science,233:747(1986),其通过引用并入本文中)且可希望自供体选择这些氨基酸保持提供了原始抗体中亲和力的所有抗原接触。如本文所述,fr序列也可被取代/突变以改善本发明的抗-ige抗体或抗原结合剂对ige的亲和力(和/或扩展其相互作用或ige的cε2上的表位)。“另外与cdr区相互作用”的残基包括通过二级结构分析确定呈足够实现cdr区的空间取向的残基。在一个实施方案中,“另外与cdr区相互作用”的残基通过分析供体免疫球蛋白的三维模型(例如计算机产生的模型)鉴定。通常原始供体抗体的三维模型显示在cdr外的某些氨基酸接近cdr和很可能与cdr中的氨基酸通过氢键、范德华力、疏水相互作用等相互作用。在那些氨基酸位置,可选择供体免疫球蛋白氨基酸而非受体免疫球蛋白氨基酸。根据此标准的氨基酸一般将在cdr中一些原子的约3埃单位(a)内具有侧链原子,并且必须含有可与cdr原子根据已确立的化学力、诸如上列化学力相互作用的原子。在可形成氢键的原子的情况下,在其核之间测量但对于未形成键的原子,在其范德华表面之间测量因此,在后一情况下,对于认为能够相互作用的原子,核必须在约内(加范德华半径总和)。在多数情况下,核将相隔4或5至在确定氨基酸是否可与cdr相互作用时,优选不考虑重链cdr2的最后8个氨基酸作为cdr的一部分,因为出于结构的观点,此8个氨基酸更多表达为构架的一部分。能够与cdr(或fr)中氨基酸相互作用的氨基酸可用另一种方式鉴定。各构架氨基酸的溶剂可及表面积以两种方式计算:(1)完整抗体中,和(2)由移除cdr的抗体组成的假设分子中。这些数目之间约10平方埃或更多的显著差异显示溶剂对构架氨基酸的接近至少部分由cdr阻断,且因此氨基酸与cdr接触。氨基酸的溶剂可及表面积可基于抗体的三维模型,使用本领域中已知的算法计算(例如connolly,j.appl.cryst.16:548(1983)及lee及richards,j.mol.biol.(1971),均通过引用并入本文中)。构架氨基酸偶尔还可与cdr,通过影响又接触cdr的另一构架氨基酸的构型而间接相互作用。已知许多抗体中构架中几个位置的特定氨基酸能够与cdr相互作用(chothia及lesk,上文;chothia等人,上文及tramontano)等人,j.mol.biol.215:175(1990),均通过引用并入本文中)。特别地,已知许多抗体中轻链的位置2、48、64和71的氨基酸和重链的位置71及94的氨基酸(根据kabat编号)能够与cdr相互作用。轻链中位置35和重链中93和103的氨基酸还可能与cdr相互作用。在所有这些编号位置,在人源化免疫球蛋白中选择供体氨基酸而非受体氨基酸(当其不同时)为优选的。另一方面,能够与cdr区相互作用的某些残基,诸如轻链的头5个氨基酸有时可在人源化免疫球蛋白中不丧失亲和力的情况下选自受体免疫球蛋白。“参与vl-vh界面”的残基或“填充残基”包括在如例如novotny及haber,proc.natl.acad.sci.usa,82:4592-66(1985)或chothia等人(同上)所定义的vl和vh之间的界面的这些残基。一般而言,如果异常填充残基不同于人构架中的填充残基,则其应保留在人源化抗体中。一般而言,满足以上标准的一个或多个氨基酸被取代。在一些实施方案中,所有或大部分满足以上标准的氨基酸被取代。有时,关于是否特定氨基酸符合以上标准存在一些不明确性,并且产生替代的变体免疫球蛋白,其中之一具有特定取代,而其他则不具有。可在本文中关于所需活性描述的任一测定中测试由此产生的替代变异免疫球蛋白,且选择优选的免疫球蛋白。通常人源化抗体中cdr区与供体抗体的对应cdr区实质上相同,且更通常相同。虽然通常是不希望的,但有时可在不明显影响所得人源化免疫球蛋白的结合亲和力下对cdr残基进行一个或多个保守氨基酸取代。保守或相似取代是预期的组合,诸如亮氨酸取代异亮氨酸或缬氨酸。经常可彼此取代的其他氨基酸包括但不限于:苯丙氨酸、酪氨酸和色氨酸(具有芳族侧链的氨基酸);赖氨酸、精氨酸和组氨酸(具有碱性侧链的氨基酸);天冬氨酸盐和谷氨酸盐(具有酸性侧链的氨基酸);天冬酰胺和谷氨酰胺(具有酰胺侧链的氨基酸);以及半胱氨酸和甲硫氨酸(具有含硫侧链的氨基酸)。用于取代的其他候选者为人免疫球蛋白在该位置不常见或“稀少”的受体人构架氨基酸。这些氨基酸可被来自供体抗体的同等位置的氨基酸或来自更典型人免疫球蛋白的同等位置的氨基酸取代。例如,当受体免疫球蛋白的人构架区中氨基酸为该位置稀少和供体免疫球蛋白中对应氨基酸为人免疫球蛋白序列中该位置常用时;或当相对于其他人序列,受体免疫球蛋白中氨基酸为该位置稀少和供体免疫球蛋白中对应氨基酸也稀少时,取代可以是期望的。这些标准帮助确保人构架中的非典型氨基酸不破坏抗体结构。此外,通过用来自供体抗体的人抗体典型的氨基酸替换非常见人受体氨基酸,可使人源化抗体免疫原性较低。如本文所用,术语“稀少”指示在序列的代表性样品中氨基酸在小于序列的约20%,但通常小于约10%中出现在该位置,且如本文所用,术语“常用”指示在序列的代表性样品中氨基酸在小于序列的约25%,但通常超过约50%中出现。例如,所有人轻链和重链可变区序列分别被分组成尤其彼此同源和在某些关键位置具有相同氨基酸的序列“亚组”(kabat等人,上文)。当确定人受体序列中的氨基酸在人序列之间为“稀少”还是“常用”时,常常优选仅仅考虑相同亚组中的这些人序列为受体序列。用于取代的其他候选残基为对应于稀少或不常见供体构架残基的受体构架残基。稀少或不常见供体构架残基为供体抗体在该位置稀少或不常见(如本文所定义)的残基。对于供体抗体,可根据kabat确定亚组和鉴定不同于共同序列的残基位置。这些供体特定的差异可指出供体序列中增强活性的体细胞突变。保留预测影响结合的不常见残基,而预测对于结合而言不重要的残基可经取代。用于取代的其他候选者为在受体构架区域中出现的非种系残基。例如,当受体抗体链(即与供体抗体链共享显著序列同一性的人抗体链)与种系抗体链(同样与供体链共享显著序列同一性)对准时,在受体链构架与种系链构架之间不匹配的残基可被来自种系序列的对应残基取代。除如上文所讨论的特定氨基酸取代外,人源化免疫球蛋白的构架区通常与其所来源的人抗体的构架区实质上同一且更通常同一(除了如本文出于本发明的目的而描述)。当然,构架区中许多氨基酸对抗体的特异性或亲和力几乎无直接作用。因此,可在不明显改变所得人源化免疫球蛋白的特异性或亲和力下容许构架残基的许多个别保守取代。因此,在一个实施方案中,人源化免疫球蛋白的可变构架区与此类序列的人可变构架区序列或共同序列共享至少65、75或85%序列相似性或同一性。在另一个实施方案中,人源化免疫球蛋白的可变构架区与此类序列的人可变构架区序列或共同序列共享至少90%、优选95%、更优选96%、97%、98%或99%序列相似性或同一性。然而,一般而言,此类取代是不期望的(除了如本文所述的那些)。如本文所用,同一性和相似性的程度容易例如如以下中所述计算:computationalmolecularbiology,lesk,a.m.,编,oxforduniversitypress,newyork,1988;biocomputing.informaticsandgenomeprojects,smith,d.w.,编,academicpress,newyork,1993;computeranalysisofsequencedata,part1,griffin,a.m.,andgriffin,h.g.,编,humanapress,newjersey,1994;sequenceanalysisinmolecularbiology,vonheinje,g.,academicpress,1987,sequenceanalysisprimer,gribskov,m.anddevereux,j.,编,mstocktonpress,newyork,1991,theblastsoftwareavailablefromncbi(altschul,s.f.等人,1990,j.mol.biol.215:403-410;gish,w.&states,d.j.1993,naturegenet.3:266-272.madden,t.l.等人,1996,meth.enzymol.266:131-141;altschul,s.f.等人,1997,nucleicacidsres.25:3389-3402;zhang,j.&madden,t.l.1997,genomeres.7:649-656,其通过引用并入本文中。已公开了讨论cdr移植抗体的许多评述,包括vaughan等人(naturebiotechnology,16,535-539,1998),其通过引用并入本文中。本发明的抗-ige抗体可包括其他结合结构域,例如根据如wo2007/024715中公开的分子dvd-ig,或wo2011/030107中描述的所谓(fabfv)2fc。因此,如本文所采用的抗体包括二价、三价或四价全长抗体。抗原结合剂抗原结合剂包括单链抗体(即全长重链和轻链);fab、修饰的fab、fab’、修饰的fab’、f(ab’)2、fv、fab-fv、fab-dsfv、单结构域抗体(例如vh或vl或vhh)(例如如wo2001090190中所述)、scfv、二价、三价或四价抗体、双-scfv、双功能抗体、tribody、triabody、四功能抗体及以上任一个的表位-抗原结合剂(参见例如holliger及hudson,2005,naturebiotech.23(9):1126-1136;adair及lawson,2005,drugdesignreviews-online2(3),209-217)。用于产生和制造这些抗体片段的方法为本领域中所众所周知(参见例如verma等人,1998,journalofimmunologicalmethods,216,165-181)。fab-fv形式首先在wo2009/040562中公开并且其二硫化物稳定形式fab-dsfv首先在wo2010/035012中公开。用于本发明的其他抗体片段包括国际专利申请wo2005/003169、wo2005/003170和wo2005/003171中描述的fab和fab’片段。多价抗体可包括多特异性,例如双特异性,或可为单特异性(参见例如wo92/22583及wo05/113605)。后者的一种此类实例为如wo92/22583中所述的tri-fab(或tfm)。典型的fab’分子包含重链及轻链对,其中重链包含可变区vh、恒定结构域ch1及天然或修饰的铰链区和轻链包含可变区vl和恒定结构域cl。在一个实施方案中,提供了根据本发明的fab’的二聚体,以产生f(ab’)2,例如二聚化可通过本文中描述的天然铰链序列或其衍生物或合成的铰链序列。抗体结合结构域一般将包含6个cdr,三个来自重链和三个来自轻链。在一个实施方案中,cdr在构架中并且一起形成可变区。因此,在一个实施方案中,抗原结合剂包括对ige具有特异性的包含轻链可变区和重链可变区的结合结构域。应理解在不显著改变抗体结合ige的能力下,如以上或以下所述,可对本发明提供的cdr或其他序列(例如可变结构域)进行一个或多个(例如1、2、3或4个)氨基酸取代、添加和/或缺失。本领域技术人员例如通过使用本文、尤其实施例中所述的方法,容易测试任何氨基酸取代、添加和/或缺失的作用。在一个实施方案中,可对本发明提供的抗体或片段中采用的cdr或构架区进行一个或多个(例如1、2、3或4个)氨基酸取代、添加和/或缺失,使得本发明的抗-ige抗体或抗原结合剂对ige的结合亲和力(kd)小于2、1.9、1.8、1.7、1.6、1.5、1.4、1.3、1.2、1.1、1.0、0.9、0.8、0.7、0.6、0.5、0.4或0.3nm。在一个实施方案中,提供了一种修饰的人源化抗体,其中对cdr、构架区或两者进行修饰,以降低kd例如至小于2、1.9、1.8、1.7、1.6、1.5、1.4、1.3、1.2、1.1、1.0、0.9、0.8、0.7、0.6、0.5、0.4或0.3nm。本发明的抗体片段包括fab、fab’、f(ab’)2、scfv、双功能抗体、scfab、dfv、单结构域轻链抗体、dsfv、包含cdr的肽等。fab为具有约50,000的分子量和抗原结合活性的抗体片段,其中在通过用蛋白酶木瓜蛋白酶(在h链的位置224的氨基酸残基切割)处理igg获得的片段之间,h链的n端侧的约一半与整个l链经由二硫键结合在一起。本发明的fab可通过用蛋白酶木瓜蛋白酶处理与ige特异性反应的本发明的人源化cdr移植抗体来获得。此外,fab可通过将编码抗体的fab的dna插入原核生物的表达载体或真核生物的表达载体中和将载体引人原核生物或真核生物中以表达fab来产生。f(ab’)2为具有约100,000的分子量和抗原结合活性的抗体片段,在通过用蛋白酶胃蛋白酶处理igg获得的片段中,其略微大于经由铰链区的二硫键结合的fab。本发明的f(ab’)2可通过用蛋白酶胃蛋白酶处理与ige特异性反应的人cdr移植抗体来获得。此外,f(ab’)2可通过经由硫醚键或二硫键结合下文所述的fab’产生。fab’为具有约50,000的分子量和抗原结合活性的抗体片段,其通过切割f(ab’)2的铰链区的二硫键获得。本发明的fab’可通过用还原剂二硫苏糖醇处理与ige特异性反应的f(ab’)2获得。此外,本发明的fab’可通过将编码与ige特异性反应的本发明的人cdr移植抗体的fab’的dna插入原核生物的表达载体或真核生物的表达载体中和将载体引入至原核生物或真核生物中以表达fab’来产生。scfv为一种vh-p-vl或vl-p-vh多肽,其中一条链vh和一条链vl使用12或更多个残基的适当肽接头(p)连接,和具有抗原结合活性。本发明的scfv可通过获得编码与ige特异性反应的本发明的人cdr移植抗体的vh和vl的cdna,构建编码scfv的dna、将dna插入原核生物的表达载体或真核生物的表达载体中和然后将表达载体引入原核生物或真核生物中以表达scfv来产生。本发明的fab片段可以直接或经由接头连接至scfv。如本文所采用的“单链可变片段”或“scfv”是指通过vh和vl可变结构域之间的肽接头,例如具有氨基酸序列seqidno.151的肽接头稳定的单链可变片段。与fab片段的连接可以是化学缀合,但最优选为翻译融合,即基因融合,其中各自的序列由表达载体按顺序编码。因此,接头通常是如本文所述的氨基酸接头。连接至fab片段的本发明的scfv可以结合血清载体蛋白质以便延长抗体融合蛋白质在体内的半衰期。以此方式延长半衰期不取决于ige结合且可以是有利的。如本文所采用的“血清载体蛋白质”是指可以scfv结合的任何合适的血浆载体蛋白质,在一个实例中,血清载体蛋白质选自甲状腺素结合蛋白、甲状腺素运载蛋白、α1-酸糖蛋白、转铁蛋白、血纤维蛋白原和白蛋白,或其中任一种的片段。通常,该scfv结合白蛋白,优选人类血清白蛋白。任何合适白蛋白结合scfv均可以并入本发明的抗体融合蛋白质中。本领域中先前已描述合适的白蛋白结合结构域。双功能抗体为一种抗体片段,其中具有相同或不同抗原结合特异性的scfv形成二聚体,和具有与相同抗原的二价抗原结合活性或与不同抗原的两种特定抗原结合活性。本发明的双功能抗体,例如与ige特异性反应的二价双功能抗体,可通过获得编码与ige特异性反应的抗体的vh及vl的cdna,构建编码具有3至10个残基的多肽接头的scfv的dna,将dna插入原核生物的表达载体或真核生物的表达载体中和然后将表达载体引入原核生物或真核生物中以表达双功能抗体来产生。dsfv通过其中vh和vl各自一个氨基酸残基经半胱氨酸残基取代的多肽经由半胱氨酸残基之间的二硫键结合来获得。经半胱氨酸残基取代的氨基酸残基可基于抗体的三维结构估计根据reiter等人(proteinengineering,7,697(1994))显示的方法选择。本发明的dsfv可通过获得编码与ige特异性反应的本发明的人cdr移植抗体的vh和vl的cdna,构建编码dsfv的dna,将dna插入原核生物的表达载体或真核生物的表达载体中和然后将表达载体引入原核生物或真核生物中以表达dsfv来产生。包含cdr的肽由包括h链及l链cdr的至少一个区域构成。复数个cdr可直接或经由适当肽接头结合。本发明的包含cdr的肽可通过获得编码与ige特异性反应的人cdr移植抗体的vh和vl的cdr的cdna,构建编码cdr的dna,将dna插入原核生物的表达载体或真核生物的表达载体中和然后通过将表达载体引入原核生物或真核生物中以表达该肽来产生。此外,包含cdr的肽还可通过化学合成方法,诸如fmoc方法(芴基甲氧羰基法)、tboc方法(叔丁氧基羰基法)或其相似方法来产生。本发明的抗体包括其中放射性同位素、蛋白质、试剂或其相似物与本发明的抗体化学或基因上缀合的抗体衍生物。本发明的抗体衍生物可通过将放射性同位素、蛋白质或试剂化学缀合至与ige特异性反应的抗体或抗体片段的h链或l链的n端侧或c端侧、抗体或抗体片段的适当取代基或侧链或者抗体或抗体片段中的糖链来产生(antibodyengineeringhandbook,osamukanemitsu编辑,chijinshokan公开(1994))。此外,其可通过将编码与ige特异性反应的本发明的抗体或抗体片段的dna与编码待结合的蛋白质的其他dna连接,将dna插入表达载体中和将表达载体引入宿主细胞中而遗传上产生。放射性同位素包括131i、125i等,和其可通过例如氯胺t方法缀合至抗体。药剂优选为低分子量化合物。实例包括抗癌剂,诸如烷基化剂(例如氮芥(nitrogenmustard)、环磷酰胺(cyclophosphamide))、代谢拮抗剂(例如5-氟尿嘧啶(5-fluorouracil)、甲氨蝶呤(methotrexate))、抗生素(例如道诺霉素(daunomycin)、博莱霉素(bleomycin)、丝裂霉素c(mitomycinc)、道诺比星、多柔比星(doxorubicin))、植物碱(例如长春新碱(vincristine)、长春碱(vinblastine)、长春地辛(vindesine))、激素药物(例如他莫昔芬(tamoxifen)、地塞米松(dexamethasone))等(clinicaloncology,japanesesocietyofclinicaloncology编辑,cancerandchemotherapy公开(1996));消炎剂,诸如类固醇剂(例如皮质酮(hydrocortisone)、泼尼松(prednisone))、非类固醇药(例如阿司匹林(aspirin)、吲哚美辛(indomethacin))、免疫调节剂(例如硫金苹果酸盐、青霉胺(penicillamine))、免疫抑制剂(例如环磷酰胺、硫唑嘌呤(azathioprine))和抗组胺剂(例如氯芬尼拉明顺丁烯二酸盐(chlorpheniraminemaleate)、氯马斯汀(clemastine))(inflammationandanti-inflammatorytherapy,ishiyakushuppan(1982));等。将道诺霉素缀合至抗体的方法包括其中道诺霉素与抗体的氨基经由戊二醛缀合的方法、其中道诺霉素的氨基与抗体的羧基经由水溶性碳化二亚胺缀合的方法及其相似方法。此外,为了直接抑制癌细胞,可使用诸如蓖麻毒素、白喉毒素和其相似物的毒素。例如,与蛋白质的融合抗体可通过将编码抗体或抗体片段的cdna连接于编码蛋白质的其他cdna,构建编码融合抗体的dna,将dna插入原核生物的表达载体或真核生物的表达载体中且然后将其引入原核生物或真核生物中以表达融合抗体来产生。本文中进一步涵盖抗体片段或抗原结合剂,包括结合剂的融合物,例如免疫球蛋白样片段和试剂,诸如双功能抗体、scab、双特异性片段、triabody、fab-fv-fv、fab-fv、tribody、(fab-fv)2-fc及抗体片段或部分,诸如cdr或包括cdr的抗体环,其移植至诸如纤连蛋白或亮氨酸拉链的非ig构架上,如全文并入本文中的binz等人,(2005)nat.biotech.23:1257-1268中所述。缀合的抗-ige单克隆抗体和抗原结合剂必要时,用于本发明的抗体或抗原结合剂可缀合至一个或多个效应分子。应了解效应分子可包含单个效应分子或两个或更多个经连接以形成可附接于本发明抗体的单一部分的此类分子。在需要获得连接于效应分子的抗体片段下,这可通过其中抗体片段直接或经由偶合剂连接于效应分子的标准化学或重组dna程序来制备。用于将此类效应分子缀合至抗体的技术为本领域中所众所周知(参见hellstrom等人,controlleddrugdelivery,第2版,robinson等人编辑,1987,第623-53页;thorpe等人,1982,immunol.rev.,62:119-58及dubowchik等人,1999,pharmacologyandtherapeutics,83,67-123)。特定化学程序包括例如wo93/06231、wo92/22583、wo89/00195、wo89/01476和wo03/031581中描述的程序。或者,在效应分子为蛋白质或多肽的情况下,连接可使用重组dna程序,例如如wo86/01533和ep0392745中所述来实现。如本文所用,术语效应分子包括例如抗肿瘤剂、药物、毒素、生物活性蛋白质(例如酶)、其他抗体或抗体片段、抗原结合剂、合成(包括peg)或天然存在的聚合物、核酸及其片段(例如dna、rna及其片段)、放射性核素、尤其放射性碘化物、放射性同位素、螯合金属、纳米粒子及报导基团,诸如荧光化合物或可通过nmr或esr光谱分析检测到的化合物。效应分子的实例可包括细胞毒素或细胞毒性剂,包括任何对细胞不利(例如杀死)的试剂。实例包括康普瑞汀(combrestatin)、多拉司他汀(dolastatin)、埃博霉素(epothilone)、星形孢菌素(staurosporin)、类美登素(maytansinoid)、海绵素(spongistatin)、根瘤菌素(rhizoxin)、软海绵素(halichondrin)、杆孢菌素(roridin)、海米斯林(hemiasterlin)、紫杉醇(taxol)、松胞菌素b(cytochalasinb)、短杆菌素d(gramicidind)、溴化乙锭(ethidiumbromide)、吐根素(emetine)、丝裂霉素(mitomycin)、依托泊苷(etoposide)、特诺波赛(tenoposide)、长春新碱(vincristine)、长春碱(vinblastine)、秋水仙碱(colchicin)、多柔比星(doxorubicin)、道诺比星、二羟基炭疽菌素二酮(dihydroxyanthracindione)、米托蒽醌(mitoxantrone)、光神霉素(mithramycin)、放线菌素d(actinomycind)、1-去氢睾酮、糖皮质激素、普鲁卡因(procaine)、丁卡因(tetracaine)、利多卡因(lidocaine)、普萘洛尔(propranolol)和嘌呤霉素(puromycin)及其相似物或同源物。效应分子还包括但不限于抗代谢物(例如甲氨蝶呤、6-巯基嘌呤(6-mercaptopurine)、6-硫代鸟嘌呤(6-thioguanine)、阿糖胞苷(cytarabine)、5-氟尿嘧啶、达卡巴嗪(decarbazine))、烷基化剂(例如甲氮芥(mechlorethamine)、噻替派(thioepa)、苯丁酸氮芥(chlorambucil)、美法仑(melphalan)、卡莫司汀(carmustine,bsnu)及洛莫司汀(lomustine,ccnu)、环硫酰胺(cyclothosphamide)、白消安(busulfan)、二溴甘露醇(dibromomannitol)、链脲菌素(streptozotocin)、丝裂霉素c和顺式-二氯二胺铂(ii)(ddp)、顺铂(cisplatin))、蒽环霉素(anthracycline)(例如道诺比星(以前为道诺霉素)及多柔比星)、抗生素(例如放线菌素d(以前为放射菌素)、博莱霉素(bleomycin)、光神霉素、安曲霉素(anthramycin,amc)、卡奇霉素或多卡米星(duocarmycin))和抗有丝分裂剂(例如长春新碱及长春碱)。其他效应分子可包括螯合放射性核素,诸如111in和90y、lul77、铋213、锎252、铱192及钨188/铼188;或药物,诸如(但不限于)烷基磷酸胆碱、拓扑异构酶i抑制剂、紫杉烷类及苏拉明(suramin)。其他效应分子包括蛋白质、肽和酶。目标酶包括但不限于蛋白水解酶、水解酶、裂解酶、异构酶、转移酶。相关蛋白质、多肽和肽包括但不限于免疫球蛋白、毒素(诸如相思豆毒素、蓖麻毒素a、假单胞菌外毒素或白喉毒素)、蛋白质(诸如胰岛素、肿瘤坏死因子、α-干扰素、β-干扰素、神经生长因子、血小板衍生生长因子或组织纤维蛋白溶酶原活化因子)、血栓性剂或抗血管生成剂(例如血管生长抑素或内皮生长抑素)或生物反应调节剂(诸如淋巴因子、白介素-1(il-1)、白介素-2(il-2)、粒细胞巨噬细胞集落刺激因子(gm-csf)、粒细胞集落刺激因子(g-csf)、神经生长因子(ngf)或其他生长因子及免疫球蛋白)。其他效应分子可包括可用于例如诊断中的可检测物质。可检测物质的实例包括各种酶、辅基、荧光物质、发光物质、生物发光物质、放射性核素、正电子发射金属(用于正电子发射断层摄影法)及非放射性顺磁性金属离子。关于金属离子,一般参见美国专利号4,741,900,其可缀合至抗体用于诊断学。合适酶包括辣根过氧化酶、碱性磷酸酶、β-半乳糖苷酶或乙酰胆碱酯酶;适合辅基包括链霉抗生物素蛋白、抗生物素蛋白及生物素;合适荧光物质包括伞酮、荧光素、异硫氰酸荧光素、若丹明、二氯三嗪基胺荧光素、丹磺酰氯及藻红素;合适发光材料包括鲁米诺(luminol);合适生物发光材料包括荧光素酶、荧光素及水母素;和合适放射性核素包括125i、131i、111in及99tc。在另一个实施方案中,效应分子可增加抗体在体内的半衰期,和/或降低抗体的免疫原性和/或增强抗体跨越上皮障壁递送至免疫系统。此类型的合适效应分子的实例包括聚合物、白蛋白、白蛋白结合蛋白质或白蛋白结合化合物,诸如wo05/117984中描述的那些。在一个实施方案中,通过与ige或抗人ige抗体无关的效应分子提供的半衰期为有利的。在效应分子为聚合物的情况下,其一般可为合成或天然存在的聚合物,例如任选地取代的直链或分支链聚亚烷基、聚亚烯基或聚氧化烯聚合物或分支链或未分支多糖(例如均多糖或杂多糖)。可存在于以上提及的合成聚合物上的任选的特定取代基包括一个或多个羟基、甲基或甲氧基。合成聚合物的特定实例包括任选地取代的直链或分支链聚(乙二醇)、聚(丙二醇)、聚(乙烯醇)或其衍生物,尤其任选地取代的聚(乙二醇),诸如甲氧基聚(乙二醇)或其衍生物。特定天然存在的聚合物包括乳糖、直链淀粉、右旋糖酐、肝糖或其衍生物。在一个实施方案中,聚合物为白蛋白或其片段,诸如人血清白蛋白或其片段。在一个实施方案中,聚合物为peg分子。如本文关于缀合物所用,“衍生物”意欲包括反应性衍生物,例如硫醇选择性反应基,诸如顺丁烯二酰亚胺等。反应性基团可直接或经由接头区段连接于聚合物。应了解此类基团的残基在一些情况下将作为抗体片段与聚合物之间的连接基团形成产物的一部分。天然或合成聚合物的尺寸可按需要变化,但一般将在500da至50000da、例如5000至40000da,诸如20000至40000da的平均分子量范围内。聚合物尺寸可尤其基于产物的意欲用途,例如定位至某些组织,诸如肿瘤或延长循环半衰期的能力而选择(评述参见chapman,2002,advanceddrugdeliveryreviews,54,531-545)。因此,例如,在产物意欲离开循环和渗透组织,例如用于治疗肿瘤的情况下,合适地使用例如分子量为大约5000da的小分子量聚合物。对于产物保持在循环中的应用,合适地使用较高分子量聚合物,例如分子量在20000da至40000da范围内的聚合物。合适聚合物包括聚亚烷基聚合物,诸如聚(乙二醇)或尤其甲氧基聚(乙二醇)或其衍生物,和尤其分子量在约15000da至约40000da范围内。在一个实例中,用于本发明的抗体附接于聚(乙二醇)(peg)部分。在一个特定实例中,抗体为抗体片段和peg分子可经由位于抗体片段中的任何可用的氨基酸侧链或末端氨基酸官能基例如任何游离氨基、亚氨基、硫醇、羟基或羧基附接。此类氨基酸可天然存在于抗体片段中或可使用重组dna方法经工程改造至片段中(参见例如us5,219,996;us5,667,425;wo98/25971、wo2008/038024)。在一个实例中,本发明的抗体分子为经修饰的fab片段,其中该修饰为一个或多个氨基酸添加至其重链的c端以允许效应分子附接。合适地,附加的氨基酸形成含有一个或多个效应分子可附接的半胱氨酸残基的经修饰的铰链区。多个位点可用于附接两个或更多个peg分子。合适地,peg分子通过位于抗体片段中的至少一个半胱氨酸残基的硫醇基共价连接。各附接于修饰的抗体片段的聚合物分子可共价连接于位于片段中的半胱氨酸残基的硫原子。共价键一般将为二硫键或尤其硫-碳键。在硫醇基用作附接点的情况下,可使用适当活化的效应分子,例如硫醇选择性衍生物,诸如顺丁烯二酰亚胺及半胱氨酸衍生物。活化聚合物可用作制备如上所述的经聚合物修饰的抗体片段中的起始物质。活化聚合物可为含有硫醇反应性基团的任何聚合物,诸如α-卤基羧酸或酯,例如碘乙酰胺、酰亚胺(例如顺丁烯二酰亚胺)、乙烯基砜或二硫化物。此类起始物质可购得(例如自nektar,以前为shearwaterpolymersinc.,huntsville,al,usa)或可使用众所周知化学程序由市售起始物质制备。特定peg分子包括20k甲氧基-peg-胺(可获自nektar,以前为shearwater;rapppolymere;以及sunbio)及m-peg-spa(可获自nektar,以前为shearwater)。在一个实施方案中,抗体为修饰的fab片段、fab’片段或difab,其例如根据ep0948544或ep1090037中公开的方法聚乙二醇化,即peg(聚(乙二醇))共价附接[还参见“poly(ethyleneglycol)chemistry,biotechnicalandbiomedicalapplications”,1992,j.miltonharris(编辑),plenumpress,newyork,“poly(ethyleneglycol)chemistryandbiologicalapplications”,1997,j.miltonharris及s.zalipsky(编辑),americanchemicalsociety,washingtondcand“bioconjugationproteincouplingtechniquesforthebiomedicalsciences”,1998,m.aslam及a.dent,grovepublishers,newyork;chapman,a.2002,advanceddrugdeliveryreviews2002,54:531-545]。在一个实例中,peg附接于铰链区中的半胱氨酸。在一个实例中,peg修饰的fab片段具有共价连接于经修饰的铰链区中单一硫醇基的马来酰亚胺基团。赖氨酸残基可共价连接于马来酰亚胺基和赖氨酸残基上的各氨基可附接分子量为大约20,000da的甲氧基聚(乙二醇)聚合物。因此附接于fab片段的peg的总分子量可为大约40,000da。特定peg分子包括经n,n’-双(甲氧基聚(乙二醇)mw20,000)修饰的赖氨酸的2-[3-(n-马来酰亚胺基)丙酰胺基]乙基酰胺,还称为peg2mal40k(可获自nektar,以前为shearwater)。peg接头的替代来源包括nof,其供应gl2-400ma3(其中以下结构中的m为5)和gl2-400ma(其中m为2)和n大约为450:换言之,各peg为约20,000da。因此,在一个实施方案中,peg为2,3-双(甲基聚氧乙烯-氧基)-l-{[3-(6-马来酰亚胺基-1-氧基己基)氨基]丙基氧基}己烷(2臂分支peg,-ch2)3nhco(ch2)5-mal,mw40,000,称为sunbrightgl2-400ma3。以下类型的其他替代peg效应分子获自drreddy、nof及jenkem:在一个实施方案中,提供了一种聚乙二醇化的本发明的抗体(例如具有本文中描述的peg),其在或约在链中氨基酸226、例如重链的氨基酸226(通过依序编号)经由半胱氨酸氨基酸残基附接。在一个实施方案中,本发明提供了一种fab’peg分子,其包含一个或多个peg聚合物,例如1或2个聚合物,诸如40kda聚合物或聚合物。根据本发明的fab’-peg分子尤其有利,因为其具有与fc片段无关的半衰期。在一个实例中,本发明提供了一种治疗通过调节人ige生物活性而改善的疾病的方法,其包括施用治疗有效量的抗-ige抗体或其抗原结合剂,其中抗体或其抗原结合剂具有与fc结合ige无关的半衰期。在一个实施方案中,提供了缀合至聚合物诸如peg分子、淀粉分子或白蛋白分子的fab’。在一个实施方案中,提供了缀合至聚合物诸如peg分子、淀粉分子或白蛋白分子的scfv。在一个实施方案中,抗体或片段缀合至淀粉分子,例如以增加半衰期。将淀粉缀合至蛋白质的方法如us8,017,739中所述,通过引用并入本文中。多核苷酸本发明还提供了一种分离的dna序列,其编码本发明的抗体分子的重链和/或轻链。合适地,dna序列编码本发明的抗体分子的重链或轻链。本发明的dna序列可包含合成dna,例如通过化学加工、cdna、基因组dna或其任何组合产生。编码本发明的抗体分子的dna序列可通过本领域技术人员众所周知的方法获得。例如,编码抗体重链和轻链部分或所有的dna序列可按需要从测定的dna序列或基于对应氨基酸序列合成。编码受体构架序列的dna为本领域技术人员广泛可获得和容易基于其已知的氨基酸序列合成。分子生物学的标准技术可用于制备编码本发明的抗体分子的dna序列。所需dna序列可使用寡核苷酸合成技术完全或部分合成。适当时可使用定点突变诱发及聚合酶链反应(pcr)技术。本发明还涉及一种克隆或表达载体,其包含本发明的一个或多个dna序列。因此,提供了一种克隆或表达载体,其包含编码本发明的抗体的一个或多个dna序列。合适地,克隆或表达载体包含分别编码本发明的抗体分子的轻链及重链的两个dna序列,及适合信号序列。在一个实例中,载体在重链与轻链之间包含基因间序列(参见wo03/048208)。可构建载体的一般方法、转染方法及培养方法为本领域技术人员所众所周知。在这方面,参考“currentprotocolsinmolecularbiology”,1999,f.m.ausubel(编辑),wileyinterscience,coldspringharbourpublishing出版的newyorkandthemaniatismanual。表达抗-ige抗体或其片段的宿主细胞还提供了一种宿主细胞,其包含一种或多种克隆或表达载体,所述克隆或表达载体包含编码本发明的抗体的一种或多种dna序列。任何合适的宿主细胞/载体系统可用于表达编码本发明的抗体分子的dna序列。可使用细菌,例如大肠杆菌及其他微生物系统,或还可使用真核,例如哺乳动物、宿主细胞表达系统。合适哺乳动物宿主细胞包括cho、骨髓瘤或杂交瘤细胞。用于本发明的中国仓鼠卵巢(cho细胞)的合适类型可包括cho和cho-k1细胞,包括dhfr-cho,诸如可用于dhfr可选标记物的cho-dg44细胞和cho-dxb11细胞或可用于谷氨酰胺合成酶可选标记物的chok1-sv细胞。用于表达抗体的其他细胞类型包括淋巴细胞性细胞系,例如nso骨髓瘤细胞和sp2细胞、cos细胞。其他合适的细胞可包括本领域中已知的人胚肾(hek)纤维母细胞,例如hek293f和expihek细胞。对于本发明的全长ab,cho是优选的,因为这是产生奥马珠单抗的标准宿主(在一个实施方案中,为本发明抗体给予奥马珠单抗的标准糖基化模式)[还参见wo2013/181577]。抗-ige抗体或其片段的产生本发明还提供了一种用于产生根据本发明的抗体分子的方法,其包括在适合于从编码本发明的抗体分子的dna表达蛋白质的条件下培养含有本发明的载体的宿主细胞和分离抗体分子。抗体分子可仅仅包含重链或轻链多肽,在此情况下仅仅重链或轻链多状编码序列需要用于转染宿主细胞。为产生包含重链和轻链的产物,细胞系可用两种载体转染,第一载体编码轻链多肽且第二载体编码重链多肽。或者,可使用单一载体,该载体包括编码轻链和重链多肽的序列。提供了一种培养宿主细胞和表达抗体或其片段、分离后者和任选地纯化其以提供了分离的抗体或片段的方法。在一个实施方案中,该方法进一步包括将效应分子缀合至分离的抗体或片段,例如缀合至尤其如本文所述的peg聚合物的步骤。在一个实施方案中,提供了一种用于纯化抗体(尤其根据本发明的抗体或片段)的方法,其包括以下步骤:以非结合模式进行阴离子交换层析法,使得杂质保留在柱上和洗脱抗体。在一个实施方案中,纯化采用蛋白质a柱上的亲和力捕获和随后滴定。在一个实施方案中,纯化采用蛋白质g柱上的亲和力捕获,和随后hplc滴定。在一个实施方案中,纯化采用ige柱上的亲和力捕获,和随后滴定。在一个实施方案中,纯化采用辛巴蓝或相似者,用于纯化白蛋白融合物或缀合物分子。适用于该方法的离子交换树脂包括q.ff树脂(由ge-healthcare供应)。步骤可例如在约ph8下进行。该方法可进一步包括采用阳离子交换层析的初始捕获步骤,例如在约ph4至5、诸如4.5下进行。阳离子交换层析可例如采用树脂,诸如captos树脂或sp琼脂糖凝胶ff(ge-healthcare供应)。随后抗体或片段可自树脂洗脱,采用诸如氯化钠的离子盐溶液,例如浓度为200mm。因此,适当时,层析步骤可包括一个或多个洗涤步骤。纯化方法还可包括一个或多个过滤步骤,诸如透滤步骤或hplc过滤步骤。因此,在一个实施方案中,提供了一种纯化的抗-ige抗体或片段,例如人源化抗体或片段,尤其根据本发明的抗体或片段,其实质上自内毒素和/或宿主细胞蛋白质或dna纯化、尤其不含或实质上不含内毒素和/或宿主细胞蛋白质或dna。如上文使用的纯化意指至少90%纯,诸如91、92、93、94、95、96、97、98、99%w/w或更纯。实质上不含内毒素一般意指每毫克抗体产物1eu的内毒素含量或更低,诸如每毫克产物0.5或0.1eu。实质上不含宿主细胞蛋白质或dna一般意指每毫克抗体产物400μg宿主细胞蛋白质和/或dna含量或更低,诸如每毫克100μg或更低,尤其适当时每毫克20μg。药物组合物因为本发明的抗体适用于治疗和/或预防病理性病况,所以本发明还提供了一种药物或诊断组合物,其包含本发明的抗体或抗原结合剂与一种或多种药学上可接受的赋形剂、稀释剂或载体组合。因此,提供了本发明的抗体或抗原结合剂用于制造药剂的用途。组合物通常将作为通常包括药学上可接受的载体的无菌药物组合物的一部分供应。本发明的药物组合物可另外包含药学上可接受赋形剂。本发明还提供了一种用于制备药物或诊断组合物的方法,其包括添加和混合本发明的抗体或抗原结合剂连同一种或多种药学上可接受的赋形剂、稀释剂或载体。抗体或抗原结合剂可为药物或诊断组合物中的唯一活性成分或可伴随有其他活性成分,包括其他抗体成分或非抗体成分,诸如类固醇或其他药物分子,尤其半衰期与ige结合无关的药物分子。药物组合物合适地包含治疗有效量的本发明的抗体或抗原结合剂。如本文所用,术语“治疗有效量”是指治疗、改善或预防靶向疾病或病况或展现可检测的治疗或预防效果所需要的治疗剂的量。对于任何公开的抗体或抗原结合剂,可最初在细胞培养测定中或在动物模型中,通常在啮齿动物、兔、狗、猪或灵长类动物中估计治疗有效量。动物模型还可用于确定施用的适当浓度范围及途径。所述信息可随后用于确定适用于施用人的剂量及途径。人受试者的精确治疗有效量将视疾病病况的严重程度、受试者的整体健康、受试者的年龄、重量及性别、饮食、施用的时间及频率、药物组合、反应敏感性及对疗法的耐受性/反应而定。此量可通过常规实验确定和在临床医师的判断内。一般而言,治疗有效量为0.01mg/kg至500mg/kg,例如0.1mg/kg至200mg/kg,诸如100mg/kg。药物组合物可以以每剂含有预定量的本发明的活性剂的单位剂型呈现。根据本发明的抗体或抗原结合剂的治疗剂量未显示体内明显毒理学作用。有利地,通过施用连续剂量的根据本发明的抗体或结合剂,体内ige活性的水平可维持在适当减少的水平。组合物可个别施用于患者或可与其他药剂、药物或激素组合施用(例如同时、连续或分开)。药物组合物还可含有药学上可接受的载体以施用抗体或抗原结合剂。载体本身不应诱发对接收组合物的个体有害的抗体产生和不应具有毒性。合适载体可为大的缓慢代谢的大分子,诸如蛋白质、多肽、脂质体、多糖、聚乳酸、聚乙醇酸、聚合氨基酸、氨基酸共聚物和失活病毒粒子。可使用药学上可接受的盐,例如无机酸盐,诸如盐酸盐、氢溴酸盐、磷酸盐和硫酸盐,或有机酸的盐,诸如乙酸盐、丙酸盐、丙二酸盐和苯甲酸盐。治疗组合物中药学上可接受的载体可另外含有液体,诸如水、盐水、甘油和乙醇。另外,辅助物质诸如润湿剂或乳化剂或ph缓冲物质可存在于此类组合物中。此类载体使得药物组合物能够调配成锭剂、丸剂、糖衣药丸、胶囊、液体、凝胶、糖浆、浆液和悬浮液,以便患者摄入。优选施用形式包括适合于肠胃外施用的形式,例如通过注射或输注,例如通过快速注射或连续输注。在产物用于注射或输注下,其可呈于油性或水性媒介物中的悬浮液、溶液或乳液的形式和其可含有配制试剂,诸如悬浮剂、防腐剂、稳定化剂和/或分散剂。或者,抗体分子可呈干燥形式,在使用之前用适当无菌液体复原。一旦配制,本发明的组合物可直接施用于受试者。待治疗的受试者可为动物。然而,组合物优选适合于施用于人受试者。本发明的药物组合物可通过多种途径施用,包括但不限于经口、静脉内、肌肉内、动脉内、髓内、鞘内、室内、透皮、经皮(例如参见wo98/20734)、皮下、腹膜内、鼻内、经肠、局部、舌下、阴道内或直肠途径。还可使用无针注射器施用本发明的药物组合物。通常,治疗组合物可制备成作为液体溶液或悬浮液的可注射剂。还可制备适合于在注射前溶解或悬浮于液体媒介物中的固体形式。组合物的直接递送将一般通过皮下(具体地)、腹膜内、经静脉内或肌肉内注射实现,或递送至组织的胞间隙。组合物还可施用于病灶中。剂量治疗可为单剂量方案或多剂量方案。应了解组合物中的活性成分将为抗体分子。因此,将易在胃肠道中降解。因此,如果组合物通过使用胃肠道的途径施用,则组合物将需要含有保护抗体免于降解但一旦自胃肠道吸收则释放抗体的试剂。药学上可接受的载体的充分讨论可得自remington'spharmaceuticalsciences(mackpublishingcompany,n.j.1991)。结构-功能特性在本发明的一个方面,所述抗体或抗原结合剂结合游离和fcεri结合的人类ige。当本发明的抗体(或抗原结合剂)结合fcεri结合的人类ige时,其稳定ige的构型。在此类稳定构型中,ige对fcεri或奥马珠单抗(或其片段)的结合亲和力比在本发明的抗体或抗原结合剂不存在的情况下更弱并且其中fcεri结合的人类ige自fcεri解离。优选地,ige在自fcεri解离后仍结合至如本文所述的抗体或抗原结合剂。如下文(例如实施例1和图2中)所示,本发明的抗体或抗原结合剂结合构型不同于当结合至奥马珠单抗ige所具有的构型的ige。不希望受理论束缚,本发明抗体引起ige采用部分弯曲的构型(图2c以及5c和5d),因此自游离或fcεri结合的ige结构伸直。此类伸直损害ige结合fcεri或保持结合fcεri的能力。如实施例部分中所示,由于本发明抗体与ige上的结合位点竞争结合fcεri,据信所述抗体能够与结合至fcεri的ige形成复合物,从而改变ige在结合至fcεri时的结构并将其自fcεri解离。具有这些特性的根据本发明的抗体是本文所述的抗体,诸如包含以下的抗体或抗原结合剂:a.包含seqidno.:1的重链可变区和包含以下的轻链可变区:i.seqidno.:109;或ii.seqidno.:113;或iii.seqidno.:121;或iv.seqidno.:132;或v.seqidno.139;或b.seqidno.:5和i.seqidno.,其中s77和s79被q置换;ii.seqidno.:117或iii.seqidno.:125;或iv.seqidno.:136;或v.seqidno.:143。具有此类特性的抗-ige抗体或抗原结合剂接触参照seqidno:108包含人类ige的cε3结构域的残基t373、w374、s375、r376、a377、s378、g379、p381、q417、c418、r419、p426、r427、a428和cε2结构域的残基d278和t281的表位,或接触所述表位且对其是特异性的。本发明的抗体在几个位置处具有甲硫氨酸残基。甲硫氨酸残基的氧化是最常见的蛋白质降解途径。参照seqidno.20在位置s64和s71处已引入甲硫氨酸残基的本发明的抗体可以在不影响所述抗体加速ige-fc:sfcriα复合物解离的能力的情况下,经历完整氧化。因此,本发明还提供了抗-ige抗体或抗原结合剂,其包含具有作为seqidno.1的氨基酸序列的重链可变区和包含cdr-l2和fw-l3的轻链可变区,其中所述轻链可变区具有作为seqidno.20的氨基酸序列,除了所述cdr-l2具有作为seqidno.50的氨基酸序列且所述fw-l3具有作为seqidno.131或138的氨基酸序列,其中参照seqidno.20,在位置64和/或71的甲硫氨酸残基被氧化。本发明还提供了抗-ige抗体或抗原结合剂,其包含具有作为seqidno.1的氨基酸序列的重链可变区和具有作为seqidno.132或139的氨基酸序列的轻链可变区,其中参照seqidno.132或139,在位置64和/或71的甲硫氨酸残基被氧化。本发明的抗体中的其他甲硫氨酸残基可以在不影响该抗体加速ige-fc:sfcriα复合物解离的能力的情况下被氧化。现在将参照附图中示出的实施方案,凭借实施例进一步描述本发明。实施例实施例1:结合至ige-fc的治疗性抗-ige抗体奥马珠单抗突变体的结构揭示其作用机制摘要免疫球蛋白e和其与受体fcεri和cd23的相互作用在过敏性疾病起到关键作用。奥马珠单抗,一种临床上批准的治疗性抗体,抑制ige与fcεri之间的相互作用,防止肥大细胞和嗜碱性细胞活化,且阻断ige结合cd23。我们解析出奥马珠单抗衍生的fab和ige-fc之间的2:1复合物的晶体结构,其中一个fab结合至各cε3结构域(且仅一个fab结合至cε2结构域)。尽管游离ige-fc在溶液中主要尖锐地弯曲,但在复合物中,其仅部分弯曲,妨碍与fcεri的相互作用;cd23结合由于在各cε3结构域上的结合位点重叠而在空间上受抑制。溶液态相互作用分析表明抑制两种受体相互作用的正构和变构基础,并与该结构一起揭示奥马珠单抗(且尤其是所述的奥马珠单抗突变体)可以如何利用ige的固有动力学和变构潜能加速受体结合的ige自fcεri的解离。引言免疫球蛋白e(ige)抗体通过其fab臂结合过敏原且通过结合fc区的受体表达其效应功能,在过敏性疾病中起到重要作用1。两种主要的ige受体是fcεri和cd23/fcεrii,通常分别称为高亲和力受体和低亲和力受体。在肥大细胞和嗜碱性细胞上,ige紧密地结合fcεri(kd≈10-10m-1),使得此类细胞仅需要存在过敏原交联ige/fcεri复合物用预先结合的ige敏化,且引发立即反应。cd23是同三聚体且因此可以通过在结合免疫复合物中聚集的ige时的亲合力效应来增强每个ige结合的c型样凝集素“头部”结构域固有地较低亲和力(kd≈10-7m-1),近似地匹配fcεri对ige的亲和力2。b细胞上表达的cd23参与ige调控,并在气管和肠上皮细胞上的表达介导ige/过敏原复合物的转胞吞作用1,2。fcεri和cd23也均在一系列抗原呈递细胞上表达。因此,ige-受体相互作用参与过敏性反应的多个方面且ige是治疗性干预的长期靶标3。ige的fc区包含二硫化物连接的以下三个结构域的二聚体:cε2、cε3和cε4。嵌合ige的早期fret研究4,5和ige-fc的x射线溶液散射研究6指示一种紧凑、弯曲的结构,且后来的ige-fc的晶体结构揭示一种尖锐且不对称的弯曲构型,其中(cε2)2结构域对向后折叠至cε3和cε4结构域上7。发现该弯曲(定义为(cε2)2结构域对的局部双重轴和fcε3-4(仅包含cε3和cε4结构域的区域)的局部双重轴之间的角度)在结合至sfcεriα(受体的ige结合α链的可溶性细胞外结构域)的ige-fc的晶体结构中变得甚至更尖锐(62°至54°)8。近期用n末端和c末端标记的ige-fc进行的fret研究证实这在sfcεriα结合后增大的弯曲9。fcεri结合位点跨越cε2邻近区域中的两个cε3结构域8,10,尽管不直接涉及cε2结构域;两条链的接合说明1:1的结合化学计算量。相比之下,两个cd23分子结合ige-fc,一个在每条链中,且另一个在cε3结构域的cε4近端11,12,13。cd23结合也引起ige-fc中的构型变化14,但并非显著影响弯曲的构型9。然而,相比于游离ige-fc,在与cd23的可溶头部结构域(scd23)的复合物中cε3结构域相对“封闭”的安置与fcεri结合所需的这些结构域的较“开放”布置不相容。这部分解释了fcεri和cd23结合的相互排斥11,12,尽管其他因素诸如局部构型变化和构型动力学改变15也可能促进该两个受体结合位点之间的变构通信2。近来通过研究ige-fc与抗-ige-fcfab(称为aεfab)的复合物发现ige-fc具有极大的柔性程度16。两个aεfab分子以对称方式结合至ige-fc,一个在每个cε3结构域上,俘获完全伸展的构型,其中(cε2)2结构域与fcε3-4区域的局部双重轴几乎一致。溶液中形成复合物的分析,以及对游离ige-fc的分子动力学模拟表明,((cε2)2结构域对可以自fcε3-4区域的一侧“翻转”至另一侧16。通过该抗-ige抗体稳定的ige-fc构型与fcεri结合不相容,由此解释其抑制活性。奥马珠单抗是批准用于治疗应用的抗-ige单克隆igg1抗体(novartis)17。其结合游离ige且抑制fcεri和cd23结合两者;结合位点已通过肽抑制和分子建模而定位至cε3结构域18,19,但其作用机制未知。然而,结合fret标记的ige-fc指示微小的伸直程度9和因此变构而非直接抑制的可能。近来发现活跃地破坏预先形成的ige/fcεri复合物的一类抑制剂:发现设计的锚蛋白重复蛋白(designedankyrinrepeatprotein(darpin))结合受体结合的ige的cε3结构域且加速其自fcεri解离20。受工程改造的二硫键限制的darpine2_79与fcε3-4分子的2:1复合物的晶体结构揭示结合位点的性质和位置,但其作用机制仍不清楚。随后报导,奥马珠单抗可以类似地促进fcεri结合的ige的解离,尽管仅仅在基本上大于治疗应用中所实现的浓度的非常高浓度下实现21,22。此处报导了ige-fc和新抗体片段(衍生自奥马珠单抗的fab(奥马珠单抗fab3)之间的复合物的晶体结构,所述fab在抗原(ige-fc)结合互补决定区(cdr)的远端含有三个点突变。参照seqidno.125,所述突变是s81r、q83r和l158p(或参照seqidno.129,s77r、q79r和l154p)。该复合物的结构揭示奥马珠单抗的作用机制,且溶液研究展示该机制利用ige的固有动力学。结果尽管付出大量努力,但对于ige-fc与奥马珠单抗fab的复合物的结晶试验仅导致fab片段的选择性结晶。其他试验已类似地报导无法使该复合物结晶23。我们因此设计一种新抗体,即衍生自奥马珠单抗的fab,其具有三个点突变,两个在vl结构域构架区中(ser81arg、gln83arg)且一个在cκ结构域中(leu158pro)(seq.idno125,pdb编号)(图1),目的在于破坏在奥马珠单抗fab晶体结构中所观察到的有利晶体接触(结果在别处报导)。我们将该奥马珠单抗衍生的fab称为“奥马珠单抗fab3”。ige-fc/奥马珠单抗fab3复合物的整体结构我们以分辨率确定ige-fc和奥马珠单抗fab3之间的复合物的晶体结构(图2a)。两个奥马珠单抗fab3分子(fab1和fab2)结合不对称的部分弯曲的ige-fc分子,每个fab接合一个cε3结构域(图2b和2c)。fab1接合ige-fcb链的cε3结构域,而fab2接合ige-fca链的cε3结构域。由于复合物中的ige-fc的部分弯曲的构型,所以fab2的轻链也与来自ige-fcb链的cε2结构域形成微小相互作用(对于该相互作用的详情,参见本实施例下文)。ige-fc和奥马珠单抗fab3之间的界面每个奥马珠单抗fab3分子接合cε3结构域暴露面的一个边缘(fcεri受体结合的fg环的c、c'、f和g股,以及碱基)。涉及奥马珠单抗fab3的重链和轻链,前者构成的界面面积的约60%(图2和3)。在两个界面之间略有不同的奥马珠单抗fab3重链(seq.idno.5)接触可以如下概述:gly32和tyr33(cdrh1)与ala377和ser378(cε3)(ige-fc序列如seq.idno108和图16中所示)形成范德华相互作用,而tyr54(cdrh2)接触gly379-pro381(cε3)。当与未结合的fab结构相比时,cdrh3残基贡献最大接触面积,并在形成复合物后经历相当大的构型变化(未公开的结果,19,23)。cdrh3残基ser100、his101、tyr102和trp106均与cε3结构域残基形成范德华相互作用,所述cε3结构域残基包括ser375-gly379、gln417和arg419(cε3)。然而,该界面部分的最显著特征是与phe103(cdrh3)的相互作用。phe103大部分埋于由thr373、trp374主链、ser375、gln417和arg419(cε3)产生的袋中,并与arg419形成阳离子/π堆叠相互作用(图3)。arg419(cε3)还在与奥马珠单抗fab3轻链(seq.idno.125)的相互作用中起关键作用(图3)。arg419(cε3)除接触asp32、asp34和tyr36侧链(与tyr36羟基形成氢键)外,还在tyr31(cdrl1)和asp32(cdrl1)主链羰基氧原子的氢键合距离内。asp32还与thr373和thr421(cε3)形成范德华相互作用。相比之下,仅两个cdrl2残基构成界面:tyr53(cdrl2)接触gln417(cε3),且tyr53和tyr57两者与met430(cε3)形成范德华相互作用;tyr57还与met430主链形成氢键。对于重链相互作用,fab1和fab2的轻链接触存在微小差异。奥马珠单抗fab3上cdr接触残基编号-呈(pdb/kabat/chothia)形式。重链序列:seq.idno5;轻链序列:seq.idno125cdrh1:ser(31/31/31)、gly(32/32/31a)、tyr(33/33/32)cdrh2:tyr(54/53/53)cdrh3:ser(100/96/96)、his(101/97/97)、tyr(102/98/98)、phe(103/99/99)、trp(106/101b/101b)cdrl1:asp(30/27c/30)、tyr(31/27d/30a)、asp(32/28/30b)、gly(33/29/30c)、asp(34/30/30d)、tyr(36/32/32)cdrl2:tyr(53/49/49)、ser(56/52/52)、tyr(57/53/53)、ser(60/56/56)cdrl1和cdrh3具有参与相互作用的大部分残基,且因此表征奥马珠单抗如何结合ige-fc和其自身相对于ige-fc如何定向。cdrl3不参与结合ige-fc。奥马珠单抗fab3界面与其他抗-ige复合物的比较cε3结构域上用于奥马珠单抗fab3与近来描述的darpine2_7920的结合位点重叠(图4),且具有类似大小,分别为和两个界面之间共有的cε3结构域残基包括ser375-gly379、gln417、arg419、arg427和met430,但奥马珠单抗fab3与受体结合的cε3fg环形成较紧密的接触,而darpine2_79界面在相反方向上延伸以包括cε3-4结构域接头。奥马珠单抗fab3和darpine2_79的重叠结合位点明显不同于近来关于奥马珠单抗fab3所述的界面,其捕获呈完全伸展的构型的ige-fc16(图5)。不仅奥马珠单抗fab3界面面积是奥马珠单抗fab3和darpine2_79的界面面积的约两倍,为但奥马珠单抗fab3在cε3中以arg393为中心的位点处接合ige-fc,并且还接触cε2结构域中的残基和cε2-cε3接头16。另一抗-ige抗体fab(medi4212)与fcε3-4的2:1复合物的晶体结构揭示cε3结构域内供抗体接合的又另一个位点,这涉及n-连接的寡糖部分的位点在asn394处24。ige-fc当结合至奥马珠单抗fab3时采取部分弯曲的构型ige-fc在溶液中是主要弯曲的5,6,9,25,26,27,28,且游离ige-fc的晶体结构揭示一种尖锐地弯曲(62°)、不对称构型,其中(cε2)2结构域对向后折叠至cε3和cε4结构域上(图5a和b),一条链(b链)的cε2结构域接触另一条链(a链)的cε4结构域7,8。ige-fc在接合fcεriα后变成甚至更尖锐地弯曲(54°)8,9,且相关构型变化涉及a链的cε3结构域与(cε2)2结构域对一起作为一个刚性单元远离b链的cε3结构域旋转8。相比于ige-fc采取完全伸展的线性构型的奥马珠单抗fab3复合物16,ige-fc在奥马珠单抗fab3复合物中采取部分弯曲的构型(图2c和5c&d),与先前揭示奥马珠单抗引起ige-fc伸直的fret研究一致9。fab1所结合的位点在游离、尖锐弯曲的ige-fc中暴露,但使fab2占据的位点可及需要将ige-fc进一步伸直至刚好超过90°。这种在奥马珠单抗fab3复合物中ige-fc的伸直与两个cε3结构域的打开相关,以产生几乎对称的fcε3-4区域(图2b)。(cε2)2结构域对位于来自每条链的cε3和cε4结构域之间且不再与来自a链的cε3结构域如此紧密地相关。在近期探究ige-fc伸直成伸展结构的分子动力学模拟中,发现在游离ige-fc的晶体结构中观察到的尖锐弯曲的构型占据最低能量池,同时还观察到对应于部分弯曲的ige-fc构型的另一不同且明确定义的能量池16。奥马珠单抗fab3/ige-fc复合物中ige-fc所采取的部分弯曲的构型占据该特殊能量池(图6)。cε3结构域在奥马珠单抗fab3/ige-fc复合物中采取显著开放的构型在ige-fc和fcε3-4子片段的晶体结构中,cε3结构域采取一系列不同的取向7,8,10,11,13,14,16,24,29,一种与ige结合其两种主要受体fcεri和cd23的变构调控有关的特性8,11,12,14。已使用各cε3结构域之间的距离以及其相对于cε4结构域的位置来描述对于fcε3-4区域所观察的构型的种类29(对于这些测量的完整描述在本实例中稍后提供)。在奥马珠单抗fab3/ige-fc复合物中,cε3结构域彼此间和与cε4结构域比含有ige-fc或fcε3-4的任何其他晶体结构中相隔更远地安置,且因此采取迄今为止所观察的最开放构型(图5);该构型比fcεri结合的ige-fc的构型显著更开放(图7)。奥马珠单抗fab3对fcεri和cd23受体结合的影响奥马珠单抗不仅抑制ige-fc和fcεri之间的相互作用,而且还抑制ige-fc和cd23之间的相互作用30。与后者一致,奥马珠单抗fab3/ige-fc和cd23/fcε3-4复合物的比较11揭示在fcε3-4上的两个cd23接合位点处奥马珠单抗fab3和cd23之间的碰撞。另外,cε3结构域残基arg376、ser378和lys380参与奥马珠单抗fab3和cd23结合11,31。相比于cd23结合ige,fcεriα跨越两个cε3结构域结合。然而,在奥马珠单抗fab3/ige-fc复合物中,cε3结构域采取的构型太开放而不允许同时接合两条链(图8)。另外,如果进而将奥马珠单抗fab3/ige-fc和sfcεriα/ige-fc复合物叠加于各cε3结构域上,则可能在每种情况下发生空间碰撞。如果叠加于a链的cε3结构域上,则奥马珠单抗fab3fab2将与fcεriα复合物中的(cε2)2结构域对碰撞,且来自奥马珠单抗fab3复合物的cε2-cε3接头将可能与fcεriα碰撞。如果叠加于b链的cε3结构域上,则奥马珠单抗fab3fab1和cε2-cε3接头(奥马珠单抗fab3复合物的接头)均可能与fcεriα之间碰撞,尽管fab1和fcεriα的结合位点实际上不重叠。然而,奥马珠单抗fab3cdrl1残基紧邻fcεriα结合的cε3结构域fg环安置。该环在b链中促进疏水性“脯氨酸夹心”相互作用,其中cε3中的pro426填充在fcεriα的两个色氨酸残基之间。asp32(cdrl1)接触thr421,gly33(cdrl1)接触pro426、arg427和ala428,且asp34(cdrl1)接触arg427和ala428。这些相互作用改变cε3结构域fg环的位置并将进一步损害ige与fcεri的结合。近来,已报导奥马珠单抗与fcεriα结合的ige的结合21,32,尽管基于ige-fc与sfcεriα8和奥马珠单抗fab3的复合物的静态晶体结构,难以看出奥马珠单抗如何能接合fcεri结合的ige。因此,我们研究了奥马珠单抗fab3与ige-fc的结合,且表征奥马珠单抗fab3和ige-fc/fcεri复合物之间的相互作用。我们的结果提供了对于奥马珠单抗的作用机制的理解。奥马珠单抗fab3与ige-fc在溶液中的相互作用我们以两种不同方式,即通过将奥马珠单抗fab3直接固定于表面上且结合ige-fc,或使奥马珠单抗fab3结合spr传感器表面上捕获的his标记的ige-fc,来表征ige-fc/奥马珠单抗fab3相互作用。使用抗-his标签抗体(gehealthcare)捕获c末端his标记的ige-fc构建体,且比较奥马珠单抗fab3、完整奥马珠单抗和奥马珠单抗fab的结合特征。不令人惊讶,在竞争结合实验中,全部三个分子均竞争相同结合位点且显示大致类似的结合亲和力(数据未显示)。奥马珠单抗fab3构建体展现的亲和力略高于奥马珠单抗fab3和完整奥马珠单抗(图8a-c)。与晶体结构一致,两个奥马珠单抗fab3分子结合ige-fc:该结合为明显双相的,在低配体浓度下观察到的高亲和力(~1nm)相互作用,和在更高浓度下观察到的第二(较弱)结合位点(~30nm)(图9a)。夹心spr实验允许分别表征该两个ige-fc/奥马珠单抗fab3结合位点。使用该方法,将奥马珠单抗fab3共价固定于传感器表面上,且使ige-fc流过该表面。在低浓度下,在这些条件下,高亲和力位点主导该相互作用,且结合曲线可以由单相相互作用动力学描述(kd~1nm,kon~1.2x106m-1s-1,koff~8x10-4s-1)。然后可以使用这种捕获于spr生物传感器表面上的1:1ige-fc/奥马珠单抗fab3复合物来测量第二奥马珠单抗fab3分子的结合,其结合明显比第一个分子更弱(kd~30nm,kon~2x105m-1s-1,koff~6x10-3s-1)(图9b)。奥马珠单抗fab3和fcεriα结合位点之间的竞争和奥马珠单抗fab3/ige-fc/fcεriα复合物的形成我们接下来研究奥马珠单抗fab3影响ige-fc和fcεriα之间的相互作用的能力。在溶液竞争结合实验中,递增浓度的奥马珠单抗fab3抑制ige-fc与fcεriα的结合(图8d)。在机理上,奥马珠单抗fab3影响可用结合位点的数量(bmax)和ige-fc/fcεriα相互作用的表观kd两者;这是混合抑制机制特征性的33。bmax值减小指示变构抑制过程,且该相互作用的表观亲和力降低最常与共有结合位点的直接竞争有关(即,正构抑制),但也可见于一些变构抑制剂。考虑到在晶体结构中观察到的结合位点,奥马珠单抗fab3可能使用正构和变构机制两者抑制ige-fc与fcεri的结合。在许多出版物中已描述奥马珠单抗和fcεriα结合位点之间的竞争,但始终将其解释为关于相同(或重叠)结合位点的直接竞争。这种解释通常用于解释奥马珠单抗为何不能结合细胞上的ige-fcεri复合物。然而,我们观察到,奥马珠单抗fab3可以高亲和力与预先结合至fcεriα的ige-fc结合(图9c,插图)。其他研究已暗示奥马珠单抗/ige-fc/fcεri复合物的存在21,32,但之前尚未实验表征该复合物。数据表明,尽管ige-fc与fcεriα的结合没有显著改变奥马珠单抗fab3对ige-fc的亲和力,但其明显地改变fcεriα结合的ige-fc分子群中奥马珠单抗fab3可用结合位点的数量。我们比较经由抗-his标签抗体捕获的ige-fc分子与由sfcεriα捕获的ige-fc分子的kd和bmax结合值,且发现相比于his标签捕获的ige-fc,fcεriα结合的ige-fc具有小于10%的奥马珠单抗fab3结合位点,如预期,其显示与2:1化学计算量一致的结合水平(图9c)。因此,与一般假设的不同,由于fcεriα和奥马珠单抗结合位点重叠,所以奥马珠单抗不结合经肥大细胞结合的ige。反而,fcεriα变构地作用于ige-fc,改变不同ige-fc构型的动态平衡16,导致fcεriα结合的ige-fc分子群中奥马珠单抗结合位点的数量实质性减少。奥马珠单抗fab3介导ige-fc/fcεriα复合物加速解离的机制kim等人20报导,darpine2_79可以加速预先形成的ige-fcεri复合物的分解。遵循该观察结果,eggel等人21随后显示,奥马珠单抗也可促进ige自fcεri解离。与这些观察结果类似,我们发现当奥马珠单抗fab3结合至ige-fc/fcεriα复合物时,其可以加速ige-fc自fcεriα解离(图9d),且奥马珠单抗fab3在这方面的确比奥马珠单抗fab3更有效,且比完整奥马珠单抗有效得多(图10e)。一个fab接合ige-fc/fcεriα复合物,但不加速ige-fc自fcεriα解离。引人注目的是,看起来加速解离仅在占据第二结合位点(即,低亲和力位点)之后发生。(奥马珠单抗fab3)2/ige-fc/fcεriα四分子复合物必须以此方式改变ige-fc的能量图谱以明显地降低ige-fc/fcεriα解离的能量屏障,导致快速解离这种另外非常稳定的复合物。奥马珠单抗fab3轻链和cε2结构域之间相互作用的详情在奥马珠单抗fab3/ige-fc复合物中,一个cε2结构域与两个突变残基(ser81arg和gln83arg)形成约的微小相互作用(相比于奥马珠单抗fab3和cε3结构域之间的平均相互作用面积)。pro158和ige-fc之间不存在接触。来自fab2轻链中的arg81侧链(奥马珠单抗fab3中的突变残基之一;(seq.idno125,pdb编号))靠来自b链的cε2结构域(seq.idno.108)的val277和asp278填充。ser80(奥马珠单抗fab3)靠asp278、leu279和thr281(cε2结构域)填充,而ser64(奥马珠单抗fab3)靠asp276和asp278填充。ser64和ser80在奥马珠单抗和奥马珠单抗fab3中相同。在奥马珠单抗fab3/ige-fc复合物中,由于asp278(cε2结构域)侧链中的无序,arg83(奥马珠单抗fab3中的突变残基之一)不明显接触cε2结构域。然而,如果asp278侧链是有序的,则可能在arg83与asp278之间形成氢键或盐桥。结晶学确定的奥马珠单抗fab3/ige-fc复合物之间的接触在晶体结构中在内抗体和抗原之间的接触通常指示表位/互补位界面。在奥马珠单抗fab3重链和轻链cdr的内的ige-fc残基确定以下表位:t373、w374、s375、r376、a377、3s78、g379、p381、q417、c418、r419、p426、r427、a428(在a链上)。另外,在奥马珠单抗fab3轻链fr1和fr3残基的内的ige-fc残基将表位延伸至:d278、t281(在b链-cε2结构域上)[且在如实施例5中所述的分子动力学模拟期间揭示,与抗体的r18、s64、s80和r81接触-和进一步与r83接触]。在晶体结构中内抗体和抗原之间的接触还提供定义抗体/抗原界面的信息。在奥马珠单抗fab3重链和轻链cdr的内的额外ige-fc残基是:k380、m430(在a链上)。另外,在奥马珠单抗fab3轻链fr1和fr3残基的内的额外ige-fc残基是:d276、v277、l279、s280、a282(在b链-cε2结构域上)[且进一步与fr1的g16和fr3的r65接触]。奥马珠单抗fab3/ige-fc复合物中cε3结构域取向的分析在分析cε3结构域关于cε4结构域的位置的一种方法中,使用来自一条链的cε3结构域的asn394cα原子和来自另一条链的cε4结构域中lys497cα原子之间的原子间距离来描述cε3结构域的“开放度”29。val336cα之间的原子间距离用于描述“摆动”,或cε3结构域彼此如何接近29。对于cε3结构域采取开放构型的fcεri结合的ige-fc和fcεri结合的fcε3-4,“开放度”值范围为而“摆动”值为的平均值8,10。奥马珠单抗fab3/ige-fc复合物的相应值对于“开放度”为的平均值,且对于“摆动”为的平均值。在奥马珠单抗fab3复合物中,cε3结构域采取迄今为止所述的最开放构型(彼此相距最远)。讨论我们报导在分辨率下ige-fc和衍生自治疗性抗-ige抗体奥马珠单抗的fab片段之间的复合物的结构;我们将在构架区中远离抗原结合位点处含有三个点突变的fab片段称为奥马珠单抗fab3。结构揭示两个奥马珠单抗fab3分子与ige-fc的复合物,一个结合至两个cε3结构域中的每一个(但仅一个fab结合至cε2结构域),并阐明奥马珠单抗抑制ige与fcεri和cd23两者的结合的能力。还发现在奥马珠单抗fab3复合物中ige-fc采取部分弯曲的构型,与先前使用fret标记的ige-fc进行的研究一致,该研究指出相对于游离ige-fc略微伸直9。ige-fc在溶液中是主要弯曲的5,6,9,25,26,27,28,并在游离ige-fc的晶体结构中,(cε2)2结构域对靠cε3和cε4结构域向后折叠7,8。近来,当我们解析在ige-fc与抗-ige-fcfab(aεfab)的复合物中捕获的完全伸展构型的结构时,明显增强ige-fc的构型柔性的理解16。探究ige-fc自尖锐地弯曲伸直成伸展构型的分子动力学模拟揭示对应于部分弯曲的构型的能量池(图6)。此处报导的其中(cε2)2结构域对和fcε3-4结构域之间的弯曲为~90°的奥马珠单抗fab3/ige-fc复合物对应于该模拟中的不同能量池16。有趣的是,奥马珠单抗fab3结合位点的位置将不妨碍进一步伸直成完全伸展的构型且因此ige-fc可能经历相当大的构型变化,甚至在与奥马珠单抗复合时。除(cε2)2结构域对相对于cε3和cε4结构域弯曲外,各种ige-fc、fcε3-4和受体复合物结构已展示,cε3结构域可以从封闭至开放采取一系列相对取向7,8,10,11,13,14,16,24,29。cε3结构域的打开和闭合促进ige-fc中受体结合的变构调控11,12:在cd23复合物中,其为相对封闭的11,13,14,而在fcεri复合物中,其更开放8,10。cd23/fcε3-4和奥马珠单抗fab3/ige-fc复合物的结构的比较显示,cd23和奥马珠单抗位点重叠,且竞争结合实验指示,奥马珠单抗对ige与cd23的结合的抑制是直接正构的。然而,fcεri结合的抑制是更复杂的。在奥马珠单抗fab3复合物中,cε3结构域采取比在任何先前结构中所见的更开放的构型,开放度如此高以致ige-fc与fcεri之间相互作用的两个子位点(一个涉及每个cε3结构域)不能同时接合。引起该抑制的另一个原因可归于奥马珠单抗fab3(fab1)分子与cε3中受体结合的fg环的邻近位置,其可以直接影响fcεriα的接触残基的构型。最后,即使在ige-fc上的奥马珠单抗fab3和fcεriα结合表位不严格地重叠,但如果两者同时结合,则存在空间碰撞的可能性。因此,晶体结构表明,奥马珠单抗的抑制机制主要是变构抑制,但也可能存在正构成分。spr研究使得我们能够评估该两个奥马珠单抗fab3结合位点的动力学和亲和力。两者亲和力明显地不同,其中kd值为~1nm和~30nm,前者与较快缔合速率常数(kon~2x106m-1s-1,与~2x105m-1s-1相比)和略微较慢的解离速率常数(koff~2x10-3s-1,与~6x10-3s-1相比)相关。可推断,较高的亲和力相互作用对应于fab1的结合,其将不受阻碍地接近弯曲的ige-fc分子,而较低的亲和力和较慢缔合速率对应于fab2,但我们对此并不确定。研究奥马珠单抗fab3抑制ige-fc结合fcεriα的机制的进一步spr实验揭示当与fcεriα复合时ige-fc上奥马珠单抗fab3可利用的位点的数量减少(bmax降低)。奥马珠单抗抑制ige结合fcεri常常以关于重叠位点的直接竞争来解释,但报导表明,奥马珠单抗可以结合受体结合的ige21,32。我们此处直接展示当ige-fc已结合至fcεriα时奥马珠单抗fab3结合ige-fc以形成三分子复合物的能力。ige-fc预先结合fcεriα的影响将ige-fc上奥马珠单抗fab3结合位点的数量减少至小于游离ige-fc中可用结合位点数量的10%;该影响只能由变构调节引起。奥马珠单抗fab3与ige-fc/fcεri复合物相互作用的性质提供了对于加速解离的机制的理解。该现象最先对于darpin报导且随后对于奥马珠单抗报导20,21,后者的浓度实质上高于在治疗上使用的浓度22,且此处现在针对奥马珠单抗fab片段显示该现象。我们进一步展示解离仅在首先结合第二(较低亲和力)奥马珠单抗fab3分子之后发生。换言之,必须形成四分子复合物-(奥马珠单抗fab3)2/ige-fc/fcεriα,才会发生显著加速解离。基于我们用奥马珠单抗fab3、ige-fc和sfcεriα的观察结果,我们设想奥马珠单抗、ige和fcεri发生以下机制:ige结合fcεri,且在这些条件下,一小群这些结合的ige分子采取可以结合奥马珠单抗分子的构型;当第二奥马珠单抗分子结合以形成四聚复合物时,ige的能量图谱改变,使得与fcεri的相互作用不稳定,且ige迅速自fcεri解离。理解该机制的关键在于理解ige能量图谱的复杂性,和动态平衡中存在的不同构型状态。奥马珠单抗的抑制活性看起来利用ige的固有柔性,和至少对于加速解离过程而言,ige/fcεri复合物的动力学。相比于其他抗体同种型,ige具有多种不常见的结构特征,包括cε2结构域的存在和独特的构型动力学,即cε3结构域的熔球样特征34。总体而言,这些特性产生一种变构通信途径,其防止cd23和fcεri的同时接合;这对于避免通过三聚cd23分子交联fcεri结合的ige的不依赖于过敏原的肥大细胞活化是必不可少的12。对于膜结合的igeb细胞受体,已提出与ige的动力学有关的其他功能益处16。奥马珠单抗不利用预期的正构机制用于抑制ige/fcεri相互作用的观察结果指示,其还利用ige的不常见动力学特性,其作为阻断抗体的能力和其避免肥大细胞结合的ige交联的能力。最后,奥马珠单抗可以通过利用ige甚至在与其受体形成复合物时也存在的变构性和固有柔性有效地将ige自fcεri解离,即使在高于治疗上使用的浓度下21。方法克隆、蛋白质表达和纯化.如16中所述克隆、表达和纯化奥马珠单抗人类igg1fab和奥马珠单抗fab3。如先前所述35产生ige-fc。ige-fc根据seq.idno.108(根据dorrington&bennich(1978)immunol.rev.41:3-25的v224-k547,但在ige-fc中插入以下突变以简化糖基化模式:n265q和n371q)。奥马珠单抗购自novartiseuropharmlimited。通过尺寸排阻色谱纯化2:1奥马珠单抗fab3/ige-fc复合物,将其洗脱至25mmtris-hclph7.5、20mmnacl和0.05%(w/v)nan3中,并浓缩至23mg/ml。表面等离振子共振.在biacoret200仪器(gehealthcare)上进行spr实验。通过使用胺偶联方案(gehealthcare),以<300个共振单位的偶联密度共价偶联蛋白质,或使用抗-his传感器表面捕获his标记的蛋白质,来制备特定表面。对于捕获his标记的配体,采用抗-his标签单克隆抗体且根据制造商的说明书(biacorehis捕获试剂盒,gehealthcare)固定。在结合实验中,通常使用180-240秒的缔合时间,且监测解离组分至少500秒。用20mmhepesph7.4、150mmnacl和0.005%(v/v)表面活性剂p-20(gehealthcare)的运行缓冲液以25μlmin-1的流动速率进行注射。大多数实验测量在25℃下进行;一些夹心结合实验在5℃下进行以便使加速解离现象最小化。使用标准双参照数据减法(standarddoublereferencingdatasubtractionmethods)36且动力学拟合使用origin软件(originlab)进行。tr-fret.ige-fc通过使100mm碳酸氢钠、50mmnacl(ph9.3)中的4mg/ml蛋白质与5倍摩尔过量的铽螯合剂异硫氰酸盐(invitrogen)反应来用供体荧光团标记。在搅动下,在室温下孵育3小时之后,通过透析至pbs(20mm磷酸盐缓冲盐水、150mmnacl,ph7.4)中来去除过量未反应的荧光团。sfcεriα-igg4-fc融合蛋白质(α-γ)37通过使3mg/ml蛋白质与2.5倍摩尔过量的alexafluor647琥珀酰亚胺基酯(invitrogen)在室温下反应1小时来用受体荧光团标记。通过透析至pbs中来去除过量荧光团。通过使1nm铽标记的ige-fc和0-20nmalexafluor647标记的sfcεriα-igg4-fc与一系列浓度的奥马珠单抗fab3竞争来进行tr-fret抑制测定。在384孔hi-base白色板(greinerbioone)中,使用lanthascreen缓冲液(invitrogen)作为稀释剂进行测定。将该板在室温下孵育过夜且通过artemis读板器(bertholdtechnologies)读取。然后将每个孔的tr-fret比率计算为在665nm下受体的发射除以在620nm下供体的发射并乘以10,000。结晶.使用沉滴式蒸气扩散法,在18℃下生长具有矩形形态的晶体,其长度为至多400μm。储层含有50μl4%(w/v)peg8000和0.03m氟化钠,且液滴含有100nl蛋白质和300nl储层。尽管在优化方面付出大量努力,但晶体的衍射质量无法进一步改善超出本研究使用的质量。晶体通常在数天之后开始生长,并且通常溶解于其液滴中,但可以在4mtmao(三甲胺n-氧化物)中稳定,所述tmao成功地用作低温保护剂。数据收集和处理.在金刚石光源(harwell,uk)处,在光束线i02和i03下收集数据。如xia2套装中所实施,使用xds38进行积分39。所述晶体各向异性地衍射,且合并来自多个晶体的数据。用来自ccp4套件的aimless将数据按比例调整至分辨率40,41,且然后使用ucla衍射各向异性服务器截断至和的分辨率限值42。对于不对称单元中的单一2:1奥马珠单抗fab3/ige-fc复合物(~170kda的分子量),matthews系数的计算指示~62%的溶剂含量。结构测定、模型构建和精化.通过使用来自pdb条目2wqr8的蛋白质原子且以分辨率奥马珠单抗fab结构(未公开的结果)作为搜索模型,用来自ccp4套件40的phaser43和molrep44进行分子替代来解析结构。最初用refmac45且随后用phenix46进行精化,且与在coot47中手动模型构建交替进行。用molprobity48、polygon49和phenix图形界面50内的其他验证工具评估模型的质量。数据处理和精化统计学呈现于表1中。电子密度图的区域显示于图11中。用pisa51分析界面,并用pymol52制图。表1.数据处理和精化统计学a圆括号中的值针对最高分辨率壳层b通过数据截断至和的分辨率限值来进行精化crfree集合包含5%反射参考文献1.gould,h.j.,sutton,b.j.igeinallergyandasthmatoday.nat.rev.immunol.8,205-217(2008).2.sutton,b.j.,davies,a.m.structureanddynamicsofige–receptorinteractions:fcεriandcd23/fcεrii.immunol.rev.268,222-235(2015).3.holgate,s.t.newstrategieswithanti-igeinallergicdiseases.worldallergyorgan.j.7,17(2014).4.holowka,d.,baird,b.structuralstudiesonthemembrane-boundimmunoglobuline(ige)-receptorcomplex.2.mappingofdistancesbetweensitesonigeandthemembranesurface.biochemistry22,3475-3484(1983).5.zheng,y.,shopes,b.,holowka,d.,baird,b.conformationsofigeboundtoitsreceptorfcepsilonriandinsolution.biochemistry30,9125-9132(1991).6.beavil,a.j.,young,r.j.,sutton,b.j.,perkins,s.j.bentdomainstructureofrecombinanthumanige-fcinsolutionbyx-rayandneutronscatteringinconjunctionwithanautomatedcurvefittingprocedure.biochemistry34,1449-1461(1995).7.wan,t.等人,thecrystalstructureofigefcrevealsanasymmetricallybentconformation.nat.immunol.3,681-686(2002).8.holdom,m.d.等人,conformationalchangesinigecontributetoitsuniquelyslowdissociationratefromreceptornat.struct.mol.biol.18,571-576(2011).9.hunt,j.等人,afluorescentbiosensorrevealsconformationalchangesinhumanimmunoglobulinefc:implicationsformechanismsofreceptorbinding,inhibition,andallergenrecognition.j.biol.chem.287,17459-17470(2012).10.garman,s.c.,wurzburg,b.a.,tarchevskaya,s.s.,kinet,j.p.,jardetzky,t.s.structureofthefcfragmentofhumanigeboundtoitshigh-affinityreceptorfcεriα.nature406,259-266(2000).11.dhaliwal,b.等人,crystalstructureofigeboundtoitsb-cellreceptorcd23revealsamechanismofreciprocalallostericinhibitionwithhighaffinityreceptorfcεri.proc.natl.acad.sciusa109,12686-12691(2012).12.borthakur,s.等人,mappingofthecd23bindingsiteonimmunoglobuline(ige)andallostericcontroloftheige-fcεriinteraction.j.biol.chem.287,31457-31461(2012).13.yuan,d.等人,ca2+-dependentstructuralchangesintheb-cellreceptorcd23increaseitsaffinityforhumanimmunoglobuline.j.biol.chem.288,21667-21677(2013).14.dhaliwal,b.,pang,m.o.,yuan,d.,beavil,a.j.,sutton,b.j.arangeofcε3-cε4interdomainanglesinigefcaccommodatebindingtoitsreceptorcd23.actacrystallogr.f70,305-309(2014).15.cooper,a.,dryden,d.t.allosterywithoutconformationalchange.aplausiblemodel.eur.biophys.j.11,103-109(1984).16.drinkwater,n.等人,humanimmunoglobulineflexesbetweenacutelybentandextendedconformations.nat.struct.mol.biol.21,397-404(2014).17.holgate,s.,casale,t.,wenzel,s.,bousquet,j.,deniz,y.,reisner,c.theanti-inflammatoryeffectsofomalizumabconfirmthecentralroleofigeinallergicinflammation.j.allergyclin.immunol.115,459-465(2005).18.zheng,l.等人,fineepitopemappingofhumanizedanti-igemonoclonalantibodyomalizumab.biochem.biophys.res.comm.375,619-622(2008).19.wright,j.d.等人,structuralandphysicalbasisforanti-igetherapy.sci.rep.5,11581(2015).20.kim,b.等人,accelerateddisassemblyofige-receptorcomplexesbyadisruptivemacromolecularinhibitor.nature491,613-617(2012).21.eggel,a.等人,accelerateddissociationofige-fcεricomplexesbydisruptiveinhibitorsactivelydesensitizesallergiceffectorcells.j.allergyclin.immunol.133,1709-1719(2014).22.lowe,p.j.,tannenbaum,s.,gautier,a.,jimenez,p.relationshipbetweenomalizumabpharmacokinetics,igepharmacodynamicsandsymptomsinpatientswithseverepersistentallergic(ige-mediated)asthma.br.j.clin.pharmacol.68,61-76(2009).23.jensen,r.k.等人,structureoftheomalizumabfab.actacrystallogr.f71,419-426(2015).24.cohen,e.s.等人,anovelige-neutralizingantibodyforthetreatmentofsevereuncontrolledasthma.mabs6,755-763(2014).25.zheng,y.,shopes,b.,holowka,d.,baird,b.dynamicconformationscomparedforigeandigg1insolutionandboundtoreceptors.biochemistry31,7446–7456(1992).26.holowka,d.,baird,b.structuralstudiesonthemembrane-boundimmunoglobuline-receptorcomplex.2.mappingofdistancesbetweensitesonigeandthemembrane-surface.biochemistry22,3475–3484(1983).27.holowka,d.,conrad,d.h.,baird,b.structuralmappingofmembrane-boundimmunoglobulin-ereceptorcomplexes:useofmonoclonalanti-igeantibodiestoprobetheconformationofreceptor-boundige.biochemistry24,6260–6267(1985).28.davis,k.g.,glennie,m.,harding,s.e.&burton,d.r.amodelforthesolutionconformationofratige.biochem.soc.trans.18,935–936(1990).29.wurzburg,b.a.,jardetsky,t.s.conformationalflexibilityinimmunoglobuline-fc3–4revealedinmultiplecrystalforms.j.mol.biol.393,176-190(2009).30.shiung,l.l.等人,ananti-igemonoclonalantibodythatbindstoigeoncd23butnotonhigh-affinityige.fcreceptors.immunobiology217,676-683(2012).31.borthakur,s.,andrejeva,g.,mcdonnell,j.m.basisoftheintrinsicflexibilityofthecε3domainofige.biochemistry50,4608-4614(2011).32.serrano-candelas,e.等人,comparableactionsofomalizumabonmastcellsandbasophils.clin.exp.allergy.46,92-102(2016).33.fersht,a.instructureandmechanisminproteinscience.w.h.freeman,newyork,pp.103–131(1999).34.price,n.e.,price,n.c.,kelly,s.m.,mcdonnell,j.m.thekeyroleofproteinflexibilityinmodulatingigeinteractions.j.biol.chem.280,2324-2330(2005).35.young,r.j.等人,secretionofrecombinanthumanige-fcbymammaliancellsandbiologicalactivityofglycosylationsitemutants.proteineng.8,193-199(1995).36.myszyka,d.g.improvingbiosensoranalysis.journalofmolecularrecognition12,279-284(1999).37.shi,j.等人,interactionofthelow-affinityreceptorcd23/fcεriilectindomainwiththefcε3-4fragmentofhumanimmunoglobuline.biochemistry36,2112-2122(1997).38.kabsch,w.xds.actacrystallogr.d66,125-132(2010).39.winter,g.xia2:anexpertsystemformacromolecularcrystallographydatareduction.j.appl.cryst.43,186-190(2010).40.winn,m.d.等人,overviewoftheccp4suiteandcurrentdevelopments.actacrystallogr.d67,235-242(2011).41.evans,p.r.,murshudov,g.n.howgoodaremydataandwhatistheresolution?actacrystallogr.d69,1204–1214(2013).42.strong,m.,sawaya,m.r.,wang,s.,phillips,m.,cascio,d.,eisenberg,d.towardthestructuralgenomicsofcomplexes:crystalstructureofape/ppeproteincomplexfrommycobacteriumtuberculosis.proc.natl.acad.sci.usa.103,8060-8065(2006).43.mccoy,a.j.等人,,phasercrystallographicsoftware.j.appl.cryst.40,658-674(2007).44.vagin,a.,teplyakov,a.molecularreplacementwithmolrep.acta.cryst.d66,22-25(2010).45.murshudov,g.n.等人,refmac5fortherefinementofmacromolecularcrystalstructures.actacrystallogr.d67,355-367(2011).46.p.v.afonine等人,towardsautomatedcrystallographicstructurerefinementwithphenix.refine.actacrystallogr.d68,352-367(2012).47.emsley,p.,lohkamp,b.,scott,w.g.,cowtan,k.featuresanddevelopmentofcoot.actacrystallogr.d66,486-501(2010).48.chen,v.b.等人,molprobity:all-atomstructurevalidationformacromolecularcrystallography.actacrystallogr.d66,12-21(2010).49.urhumsteva,l.,afonine,p.v.,adams,p.d.,urhumstev,a.crystallographicmodelqualityataglance.actacrystallogr.d65,297-300(2009).50.echols,n.等人,graphicaltoolsformacromolecularcrystallographyinphenix.j.appl.cryst.45,581-586(2012).51.krissinel,e.,henrick,k.inferenceofmacromolecularassembliesfromcrystallinestate.j.mol.biol.372,774-797(2007).52.thepymolmoleculargraphicssystem,version1.1r1llc。实施例2:通过biacore测量ige-fc自固定的sfcεriα的加速解离biacore技术测量生物分子之间的相互作用,而不需要标记。将相互作用物之一,称为配体,直接固定于或捕获至传感器表面上,而另一相互作用物,称为分析物,在溶液中流过捕获的表面。传感器检测当分析物结合配体时和当分析物自配体解离时在传感器表面处的质量变化。这些对应于缔合和解离过程。在加速解离测定中,sfcεriα是配体且固定至传感器表面。ige-fc是分析物且由sfcεriα捕获。通过缓冲液流过传感器表面或通过ige结合配偶体溶液流过传感器表面来监测ige-fc自sfcεriα的解离。该方法的详情如下:仪器:biacore3000,gehealthcareab,uppsala,sweden传感器芯片:cm5。目录号br100399bia归一化溶液:70%(w/w)甘油。bia维持试剂盒的一部分。目录号br100651。该bia维持试剂盒在4℃下储存。胺偶联试剂盒:目录号br100633。将乙基-3-(3-二甲基氨基丙基)碳化二亚胺盐酸盐(edc)于蒸馏水中制成75mg/ml且以200μl等分样品在-70℃下储存。将n-羟基琥珀酰亚胺(nhs)于蒸馏水中制成11.5mg/ml且以200μl等分样品在-70℃下储存。乙醇胺盐酸盐-naoh(ph8.5)在4℃下储存。用于氧化sfcεriα的试剂.碳酰肼(sigmaaldrich,目录号c11006)在蒸馏水中制成5mm。氰基硼氢化钠(sigmaaldrich,目录号156159)于乙酸钠(bdh,目录号s1104-500gm),100mm,ph=4中制成100mm。偏高碘酸钠(sigmaaldrich,目录号s-1878),100mm,ph=5.5于乙酸钠(bdh,目录号s1104-500gm)制成50mm。将sfcεriα在ph5.5,0.1m乙酸钠中稀释至1mg/ml。然后,将4μl高碘酸钠(50mm),1/50稀释)添加至200μl的1mg/mlsfcεriα溶液中。使混合物在冰上保持20分钟。在固定之前,用10mm乙酸钠(gehealthcare,目录号br100669),ph=4.0将sfcεriα溶液稀释至7μg/ml。缓冲液:运行缓冲液是hbs-ep(为10mmhepesph7.4、150mmnacl、3mmedta、0.05%表面活性剂p20,从10×储备溶液复原):目录号br100669。固定缓冲液为乙酸盐4.0(为10mm乙酸钠ph4.0)。目录号br100349。缓冲液在4℃下储存。配体:sfcεriα,为人类高亲和力ige受体α链的细胞外部分。在cho细胞中表达为重组蛋白质形式并纯化。分析物:ige-fc,为人类ige的fc部分,在cho细胞中表达为重组蛋白质并纯化。使用携带c225a突变的野生型人类ige-fc(根据dorrington&bennich(1978)immunol.rev.41:3-25的编号v224-k547的cε2-cε4结构域)(seq.idno.108)。突变体的命名:·奥马珠单抗fab1:s81r、q83r[根据kabat编号,s77r、q79r](作为seq.idno.24的可变区轻链+κ恒定区,其中s77和s79被q置换;作为seqidno:5的可变区重链+ch1恒定区)·奥马珠单抗fab2:l158p[根据kabat编号,l154p](作为seq.idno113的可变区轻链+κ恒定区;作为seqidno:5的可变区重链+ch1恒定区)·奥马珠单抗fab3:s81r、q83r、l158p[根据kabat编号,s77r、q79r、l154p](可变区轻链seq.idno121;作为seqidno:5的可变区重链+ch1恒定区)ige结合配偶体(1):全长奥马珠单抗(novartis);在cho细胞中表达奥马珠单抗的重组fab片段并纯化。ige结合配偶体(2):奥马珠单抗的重组fab片段,和其突变,在hek-293细胞中表达并测定培养物上清液。在分析之前对培养物上清液进行十倍浓缩。测定方法(1):通过醛偶联将sfcεriα偶联至传感器表面,达到~500个反应单位(ru)的水平。使用hbs-ep缓冲液作为运行缓冲液,流动速率为30μl/min。将ige-fc在hbs-ep中稀释至10nm且注射至固定的sfcεriα上,持续290秒,随后运行缓冲液或在运行缓冲液中稀释的ige结合配偶体注射3次,各自持续690秒。ige-fc的捕获水平为~90ru。通过两次60秒的10mm盐酸甘氨酸ph2.5注射使表面再生。将ige-fc自固定的sfcεriα解离的量计算为初始结合量的函数并将解离速率计算为ige-fc自固定的sfcεriα损失的量,其针对初始结合量随经过时间的变化归一化。测定方法(2):将sfcεriα通过醛偶联偶联至传感器表面,达到~2000个反应单位(ru)的水平。使用hbs-ep缓冲液作为运行缓冲液,流动速率为30μl/min。将ige-fc在hbs-ep中稀释至10nm且注射至固定的sfcεriα上,持续180秒,随后运行缓冲液或在运行缓冲液中稀释的ige结合配偶体注射1次,持续180秒。ige-fc的捕获水平为~275ru。通过两次60秒的10mm甘氨酸-hcl,ph2.5注射使表面再生。将ige-fc自固定的sfcεriα解离的量计算为初始结合量的函数且将解离速率计算为ige-fc自固定的sfcεriα损失的量,针对初始结合量随经过时间的变化归一化。表2:ige-fc自固定的sfcεriα解离的量的计算。ige与sfcεriα的初始结合归一化为100%且解离相对于其计算。表观解离速率基于假定的单一速率方法。进行额外突变诱导以确定是否可以鉴定出超出fab3中所述的突变的加速ige-fc自sfcεriα解离的进一步突变。这产生突变s64m(参照seq.idno.20),在fab3的情况下,该突变能够进一步增加ige自sfcεriα的解离。这些数据描述于图13和表3中。表3:ige-fc自固定的sfcεriα解离的量的计算。ige与sfcεriα的初始结合归一化为100%且解离相对于其计算。表观解离速率基于假定的单一速率方法。突变体s64m:轻链seq.idno39;奥马珠单抗fabwt:轻链seq.idno20;奥马珠单抗fab3:轻链seq.idno125。对于全部三种,重链seqidno:5。结论:综合而言,这些数据表明,奥马珠单抗fab的突变形式可以加速ige自高亲和力ige受体fcεri的固定形式解离。实现这一点的轻链中的突变包括但不必限于参照seqidno:24,s64m、s81r、q83r和l158p,且得到seqidno:39。实施例3:通过流式细胞术测量ige-fc自fcεri的加速解离。仪器:facscantoii流式细胞仪(bectondickinson)细胞系:在补充有10%胎牛血清、2mmglutamax和500μg/ml遗传霉素(lifetechnologies)的最低必需培养基中培养表达人类fcεri的rbl-sx38细胞。在测定时,在pbs中洗涤细胞且在accutase中孵育,直至脱离,然后将其以1x106个细胞/ml再悬浮于培养基中。所有子序列孵育步骤均在培养基中进行。测定方法:在37℃下,将1x106个细胞/ml的rbl-sx38与5nmalexa-488标记的ige-fc一起孵育1小时。在培养基中洗涤细胞两次以移除未结合的ige-488,然后将其以1x106个细胞/ml悬浮于培养基中或在培养基中稀释的100μg/mlige-结合剂中。然后在37℃下,在恒定旋转下孵育细胞。在每一时间点,取出0.5x105个细胞,在冰冷的pbs中洗涤且然后在4℃下,通过再悬浮于pbs中的1%三聚甲醛中来固定16小时。使用facscantoii流式细胞仪测定细胞结合的alexa-488荧光的量。流式细胞术:将固定的细胞在facs缓冲液(0.1%w/vbsa、0.01%w/vnan3/pbs,ph7.4)中洗涤两次,并将其再悬浮于200μlfacs缓冲液中。在facscantoii细胞计数器上使用标准方法执行流式细胞术,并使用flowjo软件计算结合至完整细胞的alexa-488的几何平均荧光强度。将alexa-488标记的ige-fc的解离速率计算为以过量未标记的ige-fc或在ige结合剂存在的情况下在培养基中孵育细胞引起的几何平均荧光强度随时间的变化。突变体的命名:·奥马珠单抗fab1:s81r、q83r[根据kabat编号,s77r、q79r](作为seq.idno.24的可变区轻链+κ恒定区,其中s77和s79被q置换;作为seqidno:5的可变区重链+ch1恒定区)·奥马珠单抗fab2:l158p[根据kabat编号,l154p](作为seq.idno113的可变区轻链+κ恒定区;作为seqidno:5的可变区重链+ch1恒定区)·奥马珠单抗fab3:s81r、q83r、l158p[根据kabat编号,s77r、q79r、l154p](作为seq.idno121的可变区轻链+κ恒定区;作为seqidno:5的可变区重链+ch1恒定区)。表4:alexa488标记的ige-fc自rbl-sx38细胞的表面解离的解离速率。对于所有可用时间点,由-ln(r1/r0)随时间的线性回归拟合测定表观解离速率。结论:综合而言,这些数据表明,奥马珠单抗fab的突变形式可以加速ige自细胞表面上表达的高亲和力ige受体fcεri的解离。实现这一点的轻链突变包括,但不必限于,s81r、q83r和l158p。实施例4:展示当在72小时pca模型中治疗性给药时奥马珠单抗fab3具有优于野生型奥马珠单抗fab的功效。通过将7.5μl6.68mg/ml储备液添加至1992.5μlpbs中来制备25μg/ml人类抗dnp-ige溶液。20μl该溶液的注射液将得到500ng剂量的ige。刮去动物(higertg小鼠)侧腹上的毛且然后在第0天在2pm对每一侧腹皮内注射。将20μlpbs注射至每只动物的左侧腹中作为阴性对照。将抗dnp-ige(20μl)注射至右侧腹中。对总共40只小鼠进行注射。在ige后18小时(8am)开始用野生型奥马珠单抗fab或奥马珠单抗fab3(具有s81r、q83r、l158p突变[根据kabat编号,s77r、q79r、l154p])治疗。两组小鼠(n=8只/组)皮下接受100mg/kg野生型奥马珠单抗fab或奥马珠单抗fab3。另一组8只小鼠s.c.接受pbs。10小时后(6pm),如上再次对小鼠给药。此时,在ige后28小时,还皮下给药另外两组(n=8只/组)100mg/kg野生型奥马珠单抗fab或奥马珠单抗fab3。所有组均在8am、6pm和再次在实验最后一天的8am再给药。皮内给药后72小时(2pm),对所有动物静脉内注射100μl在100iu/ml肝素中制备的1mg/mldnp-hsa、2.5%w/v伊文思蓝(evansblue)。1小时后,根据方案1方法杀死动物。移除侧腹中皮内注射部位周围的皮肤并取得穿孔活检体。将皮肤样品放置于700μl甲酰胺中并在55℃下消化过夜。在消化之后,从各样品取出100μl×2流体且放置于96孔elisa板中。然后在620nm下测量吸光度。结论:这些数据展示,当与野生型奥马珠单抗fab相比时,可以加速ige自高亲和力ige受体fcεri解离的奥马珠单抗fab的突变形式也能够以统计学显著方式减少被动皮肤过敏反应(如通过伊文思蓝染料自反应部位渗漏的抑制所示)。实现这一点的轻链突变包括,但不必限于,参照seqidno:24,s81r、q83r和l158p,且得到seqidno:39。实施例5:奥马珠单抗fab3与igefc的复合物的分子动力学模拟方法:在molecularoperatingenvironment(moe)2014.0901(1)中,通过在使用amber14(2)进行分子动力学(md)模拟之前,使cε2和cε3结构域之间的一些残基的缺失侧链和缺失环完整来制备奥马珠单抗fab3与igefc区的复合物的晶体结构(参见实施例1)。将该复合物结构加氢并在自箱的任何边缘延伸至蛋白质原子的截断的八面体箱中,使用tip3p明确水模型,用0.15mnacl盐溶质进行溶剂化。该系统使用amberff12sb和寡糖glycam_06j-1(3)力场设立并在针对库仑和范德华相互作用以及基于栅格的邻区列表设定的截止值下,通过进行50,000步的偶联物梯度算法来最小化。随后,在对所有溶质重原子具有限制(谐和力约束为5.0)的npt整体中,在恒定体积下,在125ps内将该系统从0k逐渐加热至300k,随后平衡2.25ns。对于静电,在的库仑相互作用截止值以及默认傅里叶间距和容差设置下,使用第四级pme。通过分别对蛋白质和溶剂应用弱偶联算法来控制温度且时间常数为1.0ps,且用应用于完整系统的各向同性berenson气压调节器控制压力,且时间常数为1.0ps且压缩性为巴-1。最后,在无任何限制下,使用与平衡相同的参数进行1000ns生产模拟。为了在gpu基础结构上实现4fs时间步长,使用parmed(4)将蛋白质和糖的氢质量再分配至3.024道尔顿,同时通过使总质量保持不变所需的量来调整将其所键合的原子的质量。通过使抗体轻链中的r81和r83分别虚拟突变成野生型丝氨酸和天冬酰胺,由奥马珠单抗-突变体3与igefc的复合物的晶体结构建立野生型奥马珠单抗fab与igefc区的复合物的结构的模型。使用与突变体3相同的设置方案进行md模拟。对于与igefc复合的奥马珠单抗fab3和奥马珠单抗fab,使用ambertoolscpptraj模块(7)对md轨迹进行聚类分析。采用分层聚合算法(hierarchicalagglomerativealgorithm)且各聚类之间的平均连接距离低于经由抗体轻链v区残基中各cα-原子之间的最佳拟合坐标rmsd计算各框之间的距离。仅对每10个框进行聚类分析且基于在聚类后其与聚类质心如何接近,将所有其他框添加至聚类中。参考文献1.molecularoperatingenvironment(moe),2014.09;chemicalcomputinggroupinc.,1010sherbookest.west,suite#910,montreal,qc,canada,h3a2r7,2015.2.case,d.a.,等人,amber2014,2014,universityofcalifornia,sanfrancisco.3.kirschner,k.等人,glycam06:ageneralizablebiomolecularforcefield.carbohydrates.j.comput.chem.,2008,29,622–655.4.http://parmed.github.io/parmed/html/index.html结果:为了研究奥马珠单抗fab3的轻链中的s81r和q83r突变如何影响与ige的相互作用,分别对奥马珠单抗fab和奥马珠单抗fab3与igefc结构的复合物进行一微秒分子动力学模拟。将轨迹快照聚类起来且分析前2个填入的聚类中心结构,其清楚地展示,在该两个聚类中,突变体3中的两个精氨酸突变分别与相邻igecε2结构域中的残基d278和s280形成强氢键网络。目测检查该轨迹证实,相对于如晶体结构中所公开的更接近的cε3结构域,r81-s280和r83-d278成对相互作用在模拟期间保守和稳定得多,因此经由抗体桥连冻结伸直的cε2构型。有趣的是,尽管双精氨酸突变在空间上邻近于奥马珠单抗fab3的晶体结构中的cε2d278和s280,但如md模拟中所示,不存在直接氢键合。对于奥马珠单抗fab,正如预期,如在突变体3中所见的氢键网络在该前2个聚类中心结构中缺失。目测轨迹检查显示,cε2结构域变得较少系栓至抗体轻链构架且因此其与更接近的cε3结构域的相对位置变化比奥马珠单抗fab3的相对位置变化更大。除此之外,还对其复合物中奥马珠单抗fab3和ige-fc之间的界面进行iota分析(iota是用于测定在蛋白质界面或结合位点处给定接触原子的概率的统计潜能工具),其预测奥马珠单抗fab3(参照seqidno:113)中的轻链位置s56、s64和s71可以被突变以增加奥马珠单抗对ige-fc的亲和力且因此增强预测会在以上详述的s81r和q38r突变所见的影响。结论:总体而言,md模拟提出以下假设:s81r和q38r突变通过与cε2中的相邻d278和s280直接静电相互作用和氢键相互作用来促进igecε2结构域定位至抗体,且通过将cε2锁定在伸直构型来影响ige可塑性。这与统计方法组合,所述统计方法预测s64m突变和s56(变成d、e、q或r)和s71(变成d、e或m)将具有类似影响。实施例6:通过biacore测量抗-igefab对ige-fc的亲和力。biacore技术测量生物分子之间的相互作用,而不需要标记。将相互作用物之一,称为配体,直接固定于或捕获至传感器表面上,而另一相互作用物,称为分析物,在溶液中流过捕获的表面。传感器检测当分析物结合配体时和当分析物自配体解离时在传感器表面处的质量变化。这些对应于缔合和解离过程。在动力学测定中,抗-igefab是配体且捕获至传感器表面上。ige-fc是分析物且由抗-igefab捕获。通过使ige-fc流过传感器表面(缔合相)或缓冲液流过传感器表面(解离相)来监测ige-fc与捕获的抗-igefab的缔合和解离。该方法的详情如下:仪器:biacore3000,gehealthcareab,uppsala,sweden传感器芯片:cm5。目录号br100399bia归一化溶液:70%(w/w)甘油。bia维持试剂盒的一部分。目录号br100651。bia维持试剂盒在4℃下储存。胺偶联试剂盒:目录号br100633。将乙基-3-(3-二甲基氨基丙基)碳化二亚胺盐酸盐(edc)于蒸馏水中制成75mg/ml且以200μl等分样品在-70℃下储存。将n-羟基琥珀酰亚胺(nhs)于蒸馏水中制成11.5mg/ml且以200μl等分样品在-70℃下储存。乙醇胺盐酸盐-naohph8.5在4℃下储存。缓冲液:运行缓冲液是hbs-ep(为10mmhepesph7.4、150mmnacl、3mmedta、0.05%表面活性剂p20,自10×储备溶液复原):目录号br100669。固定缓冲液为乙酸盐4.0(为10mm乙酸钠ph4.0)。目录号br100349。缓冲液在4℃下储存。配体:抗-igefab为该配体。这些在hek-293细胞中瞬时表达为重组蛋白质且不经进一步纯化即使用。分析物:ige-fc,为人类ige的fc部分,在cho细胞中表达为重组蛋白质且纯化。使用携带c225a突变的野生型人类ige-fc(根据dorrington&bennich(1978)immunol.rev.41:3-25,编号v224-k547的cε2-cε4结构域)(seq.idno.108)。使用标准方法,通过胺偶联将山羊抗人类igg1(fab2片段特异性)(jacksonimmunolabs,目录号109-006-097)的fab2片段固定至传感器表面。在运行缓冲液(hbs-ep)中稀释抗-igefab并将约200个共振单位捕获至该表面。在运行缓冲液中稀释ige-fc且使2nm至125pm的连续稀释系列通过捕获的抗-igefab。缔合相持续180秒且解离相持续300秒。通过暴露于40mmhcl60秒,随后暴露于10mmnaoh60秒且然后再暴露于40mmhcl60秒,使传感器表面再生。使用biaevaluation软件,遵循标准程序用双参照处理所有结合数据。表5:如在biacore动力学测定中测量的奥马珠单抗fab和突变体对ige-fc的亲和力。解离常数(kd)计算为kd/ka。样品ka(x10^6)1/mskd(x10^-3)1/skd(pm)wt2.861.42496s56d3.260.713219s56e2.860.717251s71e2.570.583226s71d2.510.696278表6:如在biacore动力学测定中测量的奥马珠单抗fab(wt)和突变体对ige-fc的亲和力。解离常数(kd)计算为kd/ka。四个突变体在野生型序列背景中(轻链seq.idno20)。样品ka(x10^6)1/mskd(x10^-3)1/skd(pm)wt2.2516.2721s71e1.76.25368s56d2.177.09327s71d,s56d2.138.11380s71e,s56d1.858.17443s71m,s56d2.325.45238s71d,s56e1.968.33426s71e,s56e1.946.66343s71m,s56e1.764.54258s71d,s56q1.856.12331s71e,s56q1.875.03268s71m,s56q1.444.11286s71d,s56r1.54.86325s71e,s56r1.654.63281s71m,s56r1.055.58534表7a:如在biacore动力学测定中测量的奥马珠单抗fab(wt)和突变体对ige-fc的亲和力。解离常数(kd)计算为kd/ka。所述突变参照轻链seq.idno20)。表7b:如在biacore动力学测定中所测量的奥马珠单抗fab对ige-fc的亲和力。解离常数(kd)计算为kd/ka。所述突变在轻链中(参照seqidno:20)。fab突变体的轻链可变区序列示于左边一栏(vlseq.idno.)中。表7c:如在biacore动力学测定中所测量的奥马珠单抗fab突变体对ige-fc的亲和力。解离常数(kd)计算为kd/ka。所述突变在轻链中(参照seq.idno20)。样品已经历强制氧化研究且通过质谱法测定在每个时间点时甲硫氨酸氧化的水平。结论:这些数据展示,奥马珠单抗fab的突变形式可以增加奥马珠单抗fab对ige-fc的亲和力。就改善亲和力而言,最佳突变组合是s71m与s56d的组合。该亲和力增加主要由fab自ige-fc解离的速率降低引起。鉴于相比于未突变的奥马珠单抗fab,抗体与ige-fccε2的相互作用改善,奥马珠单抗fab1(具有s81r、q83r突变[根据kabat编号,s77r、q79r])和fab3(具有s81r、q83r、l158p突变[根据kabat编号,s77r、q79r、l154p]参照seqidno;125)的亲和力也改善。实施例7:通过biacore测量ige-fc自固定的sfcεriα的加速解离使用实施例2中所概述的方法(测定方法2)测量抗-igefab对ige-fc自sfcεriα解离的影响。所有抗-igefab都在hek-29sfab中表达,通过标准方法纯化并使用计算的摩尔消光系数,通过在280nm下的吸光度进行定量。在该测定中,ige-fc的浓度为2nm且解离时间为200秒。将ige-fc自固定的sfcεriα解离的量计算为初始结合量的函数且将解离速率计算为ige-fc自固定的sfcεriα损失的量,针对初始结合量随经过时间归一化。s80n(pdb编号)被认为与cε2ige的d278相互作用,且s67w/y(pdb编号)被认为与cε2ige的t298相互作用。表8:ige-fc自固定的sfcεriα解离的量的计算。ige与sfcεriα的初始结合归一化为100%且解离相对于其计算。表观解离速率基于假定的单一速率方法。结论:这些数据展示,奥马珠单抗fab的突变形式可以加速ige自高亲和力ige受体fcεri的固定形式解离。实现这一点的轻链(seq.idno20)中的突变包括,但不必限于,在位置s56、s64、s67、s71、s80、s81、q83和l158(根据kabat编号,分别为s52、s60、s63、s67、s76、s77、q79和l154)处的突变。实施例8:突变奥马珠单抗fab的强制氧化使抗-igefab样品经历强制氧化方案以确定氧化轻链可变区中的甲硫氨酸对fab对ige-fc的亲和力的影响和加速ige-fc:sfcεriα复合物解离的能力。在室温下,将突变奥马珠单抗fab与0.1%和1%(v/v)过氧化氢一起孵育长达14天。孵育之后,将样品缓冲液交换回pbsph7.4并使用计算的消光系数,通过在280nm下的吸光度测定浓度。通过在变性条件下还原和烷基化材料,随后胰蛋白酶消化(在37℃下50μg/ml胰蛋白酶消化180分钟,随后tfa淬灭)且然后使用thermoorbitrapqexactiveplus质谱仪,通过lc-ms分析,来进行质谱分析以测定轻链可变区甲硫氨酸的氧化的量。相对于假设在合成时没有甲硫氨酸氧化来计算氧化的甲硫氨酸的百分比且与保持在4℃的参考材料进行比较。表9:突变奥马珠单抗fab的轻链可变区中氧化的甲硫氨酸的百分比。该奥马珠单抗fab突变体含有以下突变:s64m_s81r_q83r_s56d_s71m(参照seqidno:20)。结论:这些数据指示,有可能产生关键轻链甲硫氨酸(m64和m71)基本上完全氧化的突变奥马珠单抗fab。基于这些数据,将来自第1天、第3天和第7天样品的材料合并并用于确定甲硫氨酸氧化对突变奥马珠单抗fab对ige-fc的亲和力的影响和加速ige-fc:sfcεriα复合物解离的能力。实施例9:通过fret测量ige-fc自sfcεriα的加速解离在均质fret测定中测量抗-igefab对ige-fc自sfcεriα解离的影响。所有抗-igefab都在hek-293细胞中表达,且通过标准方法纯化并使用计算的摩尔消光法,通过在280nm下的吸光度进行定量。fret测定使用tb标记的ige-fc作为供体并使用alexa488标记的sfcεriα作为受体。混合两种试剂并在室温下平衡60分钟,最终测定浓度为1nm。将抗-igefab添加至混合物中,最终测定浓度为500nm,且每20分钟读取荧光(在330nm下激发,在495和520nm下发射),持续800分钟。将荧光发射绘制为时间的函数且ige-fc自sfcεriα解离的解离速率计算为复合物的半衰期。这些数据报导于表10和11中。表10:在均质fret测定中抗-igefab(wt和突变体)对ige-fc自sfcεriα解离的影响。ige-fc:sfcεriα复合物的解离为一种两相衰减。较快的初始相是大多数ige-fc:sfcεriα复合物解离且由此测定该复合物的半衰期的相。所述突变在轻链中(参照seq.idno20)。fab突变的轻链可变区序列示于左边一栏(vlseq.idno.)中。样品复合物的半衰期(min)缓冲液1240s64m_s81r_q83r_s56d_s71m10.8s64m_s81r_q83r_s56d_q71m强制氧化17.3表11:在均质fret测定中甲硫氨酸氧化对抗-igefab使ige-fc自sfcεriα解离的能力的影响。ige-fc:sfcεriα复合物的解离为一种两相衰减。较快的初始相是大多数ige-fc:sfcεriα复合物解离且由此测定该复合物的半衰期的相。结论:这些数据展示,奥马珠单抗的突变形式能够以比野生型序列更快的速率使ige-fc自sfcεriα解离。具体而言,实现这一点的轻链(参照seq.idno20)中的突变包括,但不必限于,在位置s64和s67处的突变。另外,氧化甲硫氨酸(m64和m71,seq.idno20)对fab加速ige-fc:sfcεriα复合物解离的能力无显著影响。本说明书已参考本发明的实施方案描述。然而,本领域普通技术人员了解,可在不偏离如所附权利要求中示出的本发明范围的情况下进行各种修改及变化。因此,本说明书应以说明性而非限制性意义来看待,并且所有此类修改意欲包括在本发明范围内。序列重链seq.idno.1v-区域seq.idno.2v-区域seq.idno.3具有加下划线和斜体的信号序列的v-区域seq.idno.4具有加下划线和斜体的信号序列的v-区域seq.idno.5v-区域+γ1ch1恒定区seq.idno.6v-区域+γ1ch1恒定区seq.idno.7具有加下划线和斜体的信号序列的v-区域+γ1ch1恒定区seq.idno.8具有加下划线和斜体的信号序列的v-区域+γ1ch1恒定区seq.idno.9v-区域+γ1全长恒定区seq.idno.10v-区域+γ1全长恒定区seq.idno.11具有加下划线和斜体的信号序列的v-区域+γ1全长恒定区seq.idno.12具有加下划线和斜体的信号序列的v-区域+γ1全长恒定区seq.idno.13frh1evqlvesggglvqpggslrlscavsseq.idno.14cdrh1gysitsgyswnseq.idno.15frh2wirqapgkglewvaseq.idno.16cdrh2sitydgstnynpsvkgseq.idno.17frh3ritisrddskntfylqmnslraedtavyycarseq.idno.18cdrh3gshyfghwhfavseq.idno.19frh4wgqgtlvtvss轻链seq.idno.20奥马珠单抗_v-区域seq.idno.21奥马珠单抗_v-区域seq.idno.22具有加下划线和斜体的信号序列的奥马珠单抗v-区域seq.idno.23具有加下划线和斜体的信号序列的奥马珠单抗v-区域seq.idno.24奥马珠单抗_v-区域+κ恒定区seq.idno.25奥马珠单抗_v-区域+κ恒定区seq.idno.26具有加下划线和斜体的信号序列的奥马珠单抗v-区域+κ恒定区seq.idno.27具有加下划线和斜体的信号序列的奥马珠单抗v-区域+κ恒定区seq.idno.28frl1diqltqspsslsasvgdrvtitcseq.idno.29cdrl1rasqsvdydgdsymnseq.idno.30frl2wyqqkpgkapklliyseq.idno.31cdrl2aasylesseq.idno.32frl3gvpsrfsgsgsgtdftltisslqpedfatyycseq.idno.33cdrl3qqshedpytseq.idno.34frl4fgqgtkveik突变轻链seq.idno.35s60m_s77r_q79r_v-区域(kabat)seq.idno.36s60m_s77r_q79r_v-区域(kabat)seq.idno.37具有加下划线和斜体的信号序列的s60m_s77r_q79r_v-区域(kabat)seq.idno.38具有加下划线和斜体的信号序列的s60m_s77r_q79r_v-区域(kabat)seq.idno.39s60m_s77r_q79r_v-区域+包括l154p的κ恒定区(kabat)seq.idno.40s60m_s77r_q79r_v-区域+包括l154p的κ恒定区(kabat)seq.idno.41具有加下划线和斜体的信号序列的s60m_s77r_q79r_v-区域+包括l154p的κ恒定区(kabat)seq.idno.42具有加下划线和斜体的信号序列的s60m_s77r_q79r_v-区域+包括l154p的κ恒定区(kabat)seq.idno.43frl3s64m(pdb)s60m(kabat)gvpmrfsgsgsgtdftltisslqpedfatyycseq.idno.44frl3s81r(pdb)s77r(kabat)gvpsrfsgsgsgtdftltisrlqpedfatyycseq.idno.45frl3q83r(pdb)q79r(kabat)gvpsrfsgsgsgtdftltisslrpedfatyycseq.idno.46frl3s64ms81r(pdb)s60ms77r(kabat)gvpmrfsgsgsgtdftltisrlqpedfatyycseq.idno.47frl3s64mq83r(pdb)s60mq79r(kabat)gvpmrfsgsgsgtdftltisslrpedfatyycseq.idno.48frl3s81rq83r(pdb)s77rq79r(kabat)gvpsrfsgsgsgtdftltisrlrpedfatyycseq.idno.49frl3s64ms81rq83r(pdb)s60ms77rq79r(kabat)gvpmrfsgsgsgtdftltisrlrpedfatyycseq.idno.50cdrl2s56d(pdb)s52d(kabat)aadylesseq.idno.51cdrl2s56e(pdb)s52e(kabat)aaeylesseq.idno.52frl3s71m(pdb)s67m(kabat)gvpsrfsgsgsgtdftltisslqpedfatyycseq.idno.53frl3s64ms71m(pdb)s60ms67m(kabat)gvpmrfsgsgmgtdftltisslqpedfatyycseq.idno.54frl3s81rs71m(pdb)s77rs67m(kabat)gvpsrfsgsgmgtdftltisrlqpedfatyycseq.idno.55frl3q83rs71m(pdb)q79rs67m(kabat)gvpsrfsgsgmgtdftltisslrpedfatyycseq.idno.56frl3s64ms81rs71m(pdb)s60ms77rs67m(kabat)gvpmrfsgsgmgtdftltisrlqpedfatyycseq.idno.57frl3s64mq83rs71m(pdb)s60mq79rs67m(kabat)gvpmrfsgsgmgtdftltisslrpedfatyycseq.idno.58frl3s81rq83rs71m(pdb)s77rq79rs67m(kabat)gvpsrfsgsgmgtdftltisrlrpedfatyycseq.idno.59frl3s64ms81rq83rs71m(pdb)s60ms77rq79rs67m(kabat)gvpmrfsgsgmgtdftltisrlrpedfatyycseq.idno.60frl3s67y(pdb)s63y(kabat)gvpsrfygsgsgtdftltisslqpedfatyycseq.idno.61frl3s64ms67y(pdb)s60ms63y(kabat)gvpmrfygsgsgtdftltisslqpedfatyycseq.idno.62frl3s81rs67y(pdb)s77rs63y(kabat)gvpsrfygsgsgtdftltisrlqpedfatyycseq.idno.63frl3q83rs67y(pdb)q79rs63y(kabat)gvpsrfygsgsgtdftltisslrpedfatyycseq.idno.64frl3s64ms81rs67y(pdb)s60ms77rs63y(kabat)gvpmrfygsgsgtdftltisrlqpedfatyycseq.idno.65frl3s64mq83rs67y(pdb)s60mq79rs63y(kabat)gvpmrfygsgsgtdftltisslrpedfatyycseq.idno.66frl3s81rq83rs67y(pdb)s77rq79rs63y(kabat)gvpsrfygsgsgtdftltisrlrpedfatyycseq.idno.67frl3s64ms81rq83rs67y(pdb)s60ms77rq79rs63y(kabat)gvpmrfygsgsgtdftltisrlrpedfatyycseq.idno.68frl3s80n(pdb)s76n(kabat)gvpsrfsgsgsgtdftltinslqpedfatyycseq.idno.69frl3s64ms80n(pdb)s60ms76n(kabat)gvpmrfsgsgsgtdftltinslqpedfatyycseq.idno.70frl3s81rs80n(pdb)s77rs76n(kabat)gvpsrfsgsgsgtdftltinrlqpedfatyycseq.idno.71frl3q83rs80n(pdb)q79rs76n(kabat)gvpsrfsgsgsgtdftltinslrpedfatyycseq.idno.72frl3s64ms81rs80n(pdb)s60ms77rs76n(kabat)gvpmrfsgsgsgtdftltinrlqpedfatyycseq.idno.73frl3s64mq83rs80n(pdb)s60mq79rs76n(kabat)gvpmrfsgsgsgtdftltinslrpedfatyycseq.idno.74frl3s81rq83rs80n(pdb)s77rq79rs76n(kabat)gvpsrfsgsgsgtdftltinrlrpedfatyycseq.idno.75frl3s64ms81rq83rs80n(pdb)s60ms77rq79rs76n(kabat)gvpmrfsgsgsgtdftltinrlrpedfatyycseq.idno.76frl3s80ns67y(pdb)s76ns63y(kabat)gvpsrfygsgsgtdftltinslqpedfatyycseq.idno.77frl3s64ms80ns67y(pdb)s60ms76ns63y(kabat)gvpmrfygsgsgtdftltinslqpedfatyycseq.idno.78frl3s81rs80ns67y(pdb)s77rs76ns63y(kabat)gvpsrfygsgsgtdftltinrlqpedfatyycseq.idno.79frl3q83rs80ns67y(pdb)q79rs76ns63y(kabat)gvpsrfygsgsgtdftltinslrpedfatyycseq.idno.80frl3s64ms81rs80ns67y(pdb)s60ms77rs76ns63y(kabat)gvpmrfygsgsgtdftltinrlqpedfatyycseq.idno.81frl3s64mq83rs80ns67y(pdb)s60mq79rs76ns63y(kabat)gvpmrfygsgsgtdftltinslrpedfatyycseq.idno.82frl3s81rq83rs80ns67y(pdb)s77rq79rs76ns63y(kabat)gvpsrfygsgsgtdftltinrlrpedfatyycseq.idno.83frl3s64ms81rq83rs80ns67y(pdb)s60ms77rq79rs76ns63y(kabat)gvpmrfygsgsgtdftltinrlrpedfatyycseq.idno.84frl3s67ys71m(pdb)s63ys67m(kabat)gvpsrfygsgmgtdftltisslqpedfatyycseq.idno.85frl3s64ms67ys71m(pdb)s60ms63ys67m(kabat)gvpmrfygsgmgtdftltisslqpedfatyycseq.idno.86frl3s81rs67ys71m(pdb)s77rs63ys67m(kabat)gvpsrfygsgmgtdftltisrlqpedfatyycseq.idno.87frl3q83rs67ys71m(pdb)q79rs63ys67m(kabat)gvpsrfygsgmgtdftltisslrpedfatyycseq.idno.88frl3s64ms81rs67ys71m(pdb)s60ms77rs63ys67m(kabat)gvpmrfygsgmgtdftltisrlqpedfatyycseq.idno.89frl3s64mq83rs67ys71m(pdb)s60mq79rs63ys67m(kabat)gvpmrfygsgmgtdftltisslrpedfatyycseq.idno.90frl3s81rq83rs67ys71m(pdb)s77rq79rs63ys67m(kabat)gvpsrfygsgmgtdftltisrlrpedfatyycseq.idno.91frl3s64ms81rq83rs67ys71m(pdb)s60ms77rq79rs63ys67m(kabat)gvpmrfygsgmgtdftltisrlrpedfatyycseq.idno.92frl3s80ns71m(pdb)s76ns67m(kabat)gvpsrfsgsgmgtdftltinslqpedfatyycseq.idno.93frl3s64ms80ns71m(pdb)s60ms76ns67m(kabat)gvpmrfsgsgmgtdftltinslqpedfatyycseq.idno.94frl3s81rs80ns71m(pdb)s77rs76ns67m(kabat)gvpsrfsgsgmgtdftltinrlqpedfatyycseq.idno.95frl3q83rs80ns71m(pdb)q79rs76ns67m(kabat)gvpsrfsgsgmgtdftltinslrpedfatyycseq.idno.96frl3s64ms81rs80ns71m(pdb)s60ms77rs76ns67m(kabat)gvpmrfsgsgmgtdftltinrlqpedfatyycseq.idno.97frl3s64mq83rs80ns71m(pdb)s60mq79rs76ns67m(kabat)gvpmrfsgsgmgtdftltinslrpedfatyycseq.idno.98frl3s81rq83rs80ns71m(pdb)s77rq79rs76ns67m(kabat)gvpsrfsgsgmgtdftltinrlrpedfatyycseq.idno.99frl3s64ms81rq83rs80ns71m(pdb)s60ms77rq79rs76ns67m(kabat)gvpmrfsgsgmgtdftltinrlrpedfatyycseq.idno.100frl3s80ns67ys71m(pdb)s76ns63ys67m(kabat)gvpsrfygsgmgtdftltinslqpedfatyycseq.idno.101frl3s64ms80ns67ys71m(pdb)s60ms76ns63ys67m(kabat)gvpmrfygsgmgtdftltinslqpedfatyycseq.idno.102frl3s81rs80ns67ys71m(pdb)s77rs76ns63ys67m(kabat)gvpsrfygsgmgtdftltinrlqpedfatyycseq.idno.103frl3q83rs80ns67ys71m(pdb)q79rs76ns63ys67m(kabat)gvpsrfygsgmgtdftltinslrpedfatyycseq.idno.104frl3s64ms81rs80ns67ys71m(pdb)s60ms77rs76ns63ys67m(kabat)gvpmrfygsgmgtdftltinrlqpedfatyycseq.idno.105frl3s64mq83rs80ns67ys71m(pdb)s60mq79rs76ns63ys67m(kabat)gvpmrfygsgmgtdftltinslrpedfatyycseq.idno.106frl3s81rq83rs80ns67ys71m(pdb)s77rq79rs76ns63ys67m(kabat)gvpsrfygsgmgtdftltinrlrpedfatyycseq.idno.107frl3s64ms81rq83rs80ns67ys71m(pdb)s60ms77rq79rs76ns63ys67m(kabat)gvpmrfygsgmgtdftltinrlrpedfatyycseq.idno.108携带c225a突变的野生型人类ige-fc(具有根据dorrington&bennich(1978)immunol.rev.41:3-25的编号v224-k547的cε2-cε4结构域);nb,在结晶学实验中,以下突变也插入ige-fc中以简化糖基化模式:n265q&n371q(再次根据dorrington&bennich编号)轻链fab1seq.idno.109s77r_q79r_v-区域fab1)seq.idno.110s77r_q79r_v-区域(fab1)seq.idno.111具有加下划线和斜体的信号序列的s77r_q79r_v-区域(fab1)seq.idno.112具有加下划线和斜体的信号序列的s77r_q79r_v-区域(fab1)fab2奥马珠单抗v-区域+包括l154p的κ恒定区seq.idno.113fab2_v-区域seq.idno.114fab2_v-区域seq.idno.115具有加下划线和斜体的信号序列的fab2_v-区域seq.idno.116具有加下划线和斜体的信号序列的fab2_v-区域seq.idno.117fab2_v-区域+包括l154p的κ恒定区seq.idno.118fab2_v-区域+包括l154p的κ恒定区seq.idno.119具有加下划线和斜体的信号序列的fab2_v-区域+包括l154p的κ恒定区seq.idno.120具有加下划线和斜体的信号序列的fab2_v-区域+包括l154p的κ恒定区fab3s77r_q79r_v-区域(来自fab1)+包括l154p的κ恒定区(来自fab2)seq.idno.121s77r_q79r_v-区域seq.idno.122s77r_q79r_v-区域seq.idno.123具有加下划线和斜体的信号序列的s77r_q79r_v-区域seq.idno.124具有加下划线和斜体的信号序列的s77r_q79r_v-区域seq.idno.125s77r_q79r_v-区域+包括l154p的κ恒定区seq.idno.126s77r_q79r_v-区域+包括l154p的κ恒定区seq.idno.127具有加下划线和斜体的信号序列的s77r_q79r_v-区域+包括l154p的κ恒定区seq.idno.128具有加下划线和斜体的信号序列的s77r_q79r_v-区域+包括l154p的κ恒定区seq.idno.129奥马珠单抗_v-区域seq.idno.130奥马珠单抗_v-区域seqidno:131fr3gvpmrfsgsgmgtdftltisrlrpedfatyycseqidno:132s60m_s52d_s67m_s77r_q79r_v-区域seqidno:133s60m_s52d_s67m_s77r_q79r_v-区域gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcagaagcccggaaaagccccaaagttgctgatctacgccgccgactaccttgaaagcggcgtgcctatgcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcgagactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgtttggccagggaaccaaggtcgaaatcaagseqidno:134具有加下划线和斜体的信号序列的s60m_s52d_s67m_s77r_q79r_v-区域seqidno:135具有加下划线和斜体的信号序列的s60m_s52d_s67m_s77r_q79r_v-区域atgaagtgggtcaccttcatctccctgctgtttctgttctcctccgcctactccgatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcagaagcccggaaaagccccaaagttgctgatctacgccgccgactaccttgaaagcggcgtgcctatgcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcgagactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgtttggccagggaaccaaggtcgaaatcaagseqidno:136s60m_s52d_s67m_s77r_q79r_v-区域+κ恒定区seqidno:137s60m_s52d_s67m_s77r_q79r_v-区域+κ恒定区gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcagaagcccggaaaagccccaaagttgctgatctacgccgccgactaccttgaaagcggcgtgcctatgcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcgagactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgtttggccagggaaccaaggtcgaaatcaagcgtacggtagcggccccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtseqidno:138fr3gvprrfsgsgmgtdftltisrlrpedfatyycseqidno:139s60r_s52d_s67m_s77r_q79r_v-区域seqidno:140s60r_s52d_s67m_s77r_q79r_v-区域gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcagaagcccggaaaagccccaaagttgctgatctacgccgccgattaccttgaaagcggcgtgcctcgtcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcgagactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgtttggccagggaaccaaggtcgaaatcaagseqidno:141具有加下划线和斜体的信号序列的s60r_s52d_s67m_s77r_q79r_v-区域seqidno:142具有加下划线和斜体的信号序列的s60r_s52d_s67m_s77r_q79r_v-区域atgaagtgggtcaccttcatctccctgctgtttctgttctcctccgcctactccgatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcagaagcccggaaaagccccaaagttgctgatctacgccgccgattaccttgaaagcggcgtgcctcgtcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcgagactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgtttggccagggaaccaaggtcgaaatcaagseqidno:143s60r_s52d_s67m_s77r_q79r_v-区域+κ恒定区seqidno:144s60r_s52d_s67m_s77r_q79r_v-区域+κ恒定区gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcagaagcccggaaaagccccaaagttgctgatctacgccgccgattaccttgaaagcggcgtgcctcgtcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcgagactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgtttggccagggaaccaaggtcgaaatcaagcgtacggtagcggccccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgtseqidno:145s60k_s52d_s67m_s77r_q79r_v-区域(kabat)seqidno:146s60q_s52d_s67m_s77r_q79r_v-区域(kabat)seqidno:147具有加下划线和斜体的信号序列的v-区域+γ1ch1恒定区加ca645gl4gh5scfvevqlvesggglvqpggslrlscavsgysitsgyswnwirqapgkglewvasitydgstnynpsvkgritisrddskntfylqmnslraedtavyycargshyfghwhfavwgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscsggggtggggsevqllesggglvqpggslrlscavsgidlsnyainwvrqapgkclewigiiwasgttfyatwakgrftisrdnskntvylqmnslraedtavyycartvpgystapyfdlwgqgtlvtvssggggsggggsggggsggggsdiqmtqspssvsasvgdrvtitcqsspsvwsnflswyqqkpgkapklliyeaskltsgvpsrfsgsgsgtdftltisslqpedfatyycgggyssisdttfgcgtkveikrtseqidno:148具有加下划线和斜体的信号序列的v-区域+γ1ch1恒定区加ca645gl4gh5scfvgaagtgcagttggtggagtcgggtggagggctggtgcagcctggcggtagcctgaggctgtcctgtgccgtgtccggatactccattacctccggctactcgtggaactggatcagacaggctcccggaaagggacttgagtgggtggcgtccatcacctacgacggctcaaccaactataacccgtccgtgaagggccgcatcaccatttcgcgcgacgacagcaagaatactttttacctccaaatgaacagcctgcgggccgaagatactgccgtgtactactgcgcgcggggatcacattacttcgggcactggcacttcgccgtctggggacagggcaccctcgtcactgtctcgagcgcttctacaaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccagtgacggtgtcgtggaactcaggtgccctgaccagcggcgttcacaccttcccggctgtcctacagtcttcaggactctactccctgagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtcgataagaaagttgagcccaaatcttgtagtggaggtgggggcaccggtggaggtggcagcgaggttcaactgcttgagtctggaggaggcctagtccagcctggagggagcctgcgtctctcttgtgcagtaagcggcatcgacctgagcaattacgccatcaactgggtgagacaagctccggggaagtgtttagaatggatcggtataatatgggccagtgggacgaccttttatgctacatgggcgaaaggaaggtttacaattagccgggacaatagcaaaaacaccgtgtatctccaaatgaactccttgcgagcagaggacacggcggtgtactattgtgctcgcactgtcccaggttatagcactgcaccctacttcgatctgtggggacaagggaccctggtgactgtttcaagtggcggagggggtagtggagggggtggctctgggggtggcggaagcggtggcgggggttctgacatacaaatgactcagtctccttcatcggtatccgcgtccgttggcgatagggtgactattacatgtcaaagctctcctagcgtctggagcaattttctatcctggtatcaacagaaaccggggaaggctccaaaacttctgatttatgaagcctcgaaactcaccagtggagttccgtcaagattcagtggctctggatcagggacagacttcacgttgacaatcagttcgctgcaaccagaggactttgcgacctactattgtggtggaggttacagtagcataagtgatacgacatttgggtgcggtactaaggtggaaatcaaacgtaccseqidno:149ch1和ca645gl4gh5scfv之间的接头sggggtggggsseqidno:150ca645gl4gh5scfvevqllesggglvqpggslrlscavsgidlsnyainwvrqapgkclewigiiwasgttfyatwakgrftisrdnskntvylqmnslraedtavyycartvpgystapyfdlwgqgtlvtvssggggsggggsggggsggggsdiqmtqspssvsasvgdrvtitcqsspsvwsnflswyqqkpgkapklliyeaskltsgvpsrfsgsgsgtdftltisslqpedfatyycgggyssisdttfgcgtkveikrtseqidno:151scfv内ca645gh5和ca645gl4之间的接头ggggsggggsggggsggggsseqidno:152ca645cdrh1gidlsnyainseqidno:153ca645cdrh2iiwasgttfyatwakgseqidno:154ca645cdrh3tvpgystapyfdlseqidno:155ca645cdrl1qsspsvwsnflsseqidno:156ca645cdrl2easkltsseqidno:157ca645cdrl3gggyssisdttseqidno:158s60m_s52d_s67m_s77r_q79r_s63w_s76n_v-区域(kabat)seqidno:159s60m_s52d_s67m_s77r_q79r_s63y_s76n_v-区域(kabat)seqidno:160信号序列mkwvtfisllflfssays序列表<110>ucb生物制药私人有限公司<120>抗-ige抗体<130>pf0087-wo<150>gb1610198.2<151>2016-06-10<160>160<170>patentin3.5版<210>1<211>121<212>prt<213>人工序列<220><223>v-区域<400>1gluvalglnleuvalgluserglyglyglyleuvalglnproglygly151015serleuargleusercysalavalserglytyrserilethrsergly202530tyrsertrpasntrpileargglnalaproglylysglyleuglutrp354045valalaserilethrtyraspglyserthrasntyrasnproserval505560lysglyargilethrileserargaspaspserlysasnthrphetyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaargglyserhistyrpheglyhistrphisphealavaltrpgly100105110glnglythrleuvalthrvalserser115120<210>2<211>363<212>dna<213>人工序列<220><223>v-区域<400>2gaagtgcagttggtggagtcgggtggagggctggtgcagcctggcggtagcctgaggctg60tcctgtgccgtgtccggatactccattacctccggctactcgtggaactggatcagacag120gctcccggaaagggacttgagtgggtggcgtccatcacctacgacggctcaaccaactat180aacccgtccgtgaagggccgcatcaccatttcgcgcgacgacagcaagaatactttttac240ctccaaatgaacagcctgcgggccgaagatactgccgtgtactactgcgcgcggggatca300cattacttcgggcactggcacttcgccgtctggggacagggcaccctcgtcactgtctcg360agc363<210>3<211>139<212>prt<213>人工序列<220><223>具有信号序列的v-区域<400>3metlystrpvalthrpheileserleuleupheleupheserserala151015tyrsergluvalglnleuvalgluserglyglyglyleuvalglnpro202530glyglyserleuargleusercysalavalserglytyrserilethr354045serglytyrsertrpasntrpileargglnalaproglylysglyleu505560glutrpvalalaserilethrtyraspglyserthrasntyrasnpro65707580servallysglyargilethrileserargaspaspserlysasnthr859095phetyrleuglnmetasnserleuargalagluaspthralavaltyr100105110tyrcysalaargglyserhistyrpheglyhistrphisphealaval115120125trpglyglnglythrleuvalthrvalserser130135<210>4<211>417<212>dna<213>人工序列<220><223>具有信号序列的v-区域<400>4atgaagtgggtcaccttcatctccctgctgtttctgttctccagcgcctactccgaagtg60cagttggtggagtcgggtggagggctggtgcagcctggcggtagcctgaggctgtcctgt120gccgtgtccggatactccattacctccggctactcgtggaactggatcagacaggctccc180ggaaagggacttgagtgggtggcgtccatcacctacgacggctcaaccaactataacccg240tccgtgaagggccgcatcaccatttcgcgcgacgacagcaagaatactttttacctccaa300atgaacagcctgcgggccgaagatactgccgtgtactactgcgcgcggggatcacattac360ttcgggcactggcacttcgccgtctggggacagggcaccctcgtcactgtctcgagc417<210>5<211>224<212>prt<213>人工序列<220><223>v-区域+γ1ch1恒定区<400>5gluvalglnleuvalgluserglyglyglyleuvalglnproglygly151015serleuargleusercysalavalserglytyrserilethrsergly202530tyrsertrpasntrpileargglnalaproglylysglyleuglutrp354045valalaserilethrtyraspglyserthrasntyrasnproserval505560lysglyargilethrileserargaspaspserlysasnthrphetyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaargglyserhistyrpheglyhistrphisphealavaltrpgly100105110glnglythrleuvalthrvalserseralaserthrlysglyproser115120125valpheproleualaproserserlysserthrserglyglythrala130135140alaleuglycysleuvallysasptyrpheprogluprovalthrval145150155160sertrpasnserglyalaleuthrserglyvalhisthrpheproala165170175valleuglnserserglyleutyrserleuserservalvalthrval180185190proserserserleuglythrglnthrtyrilecysasnvalasnhis195200205lysproserasnthrlysvalasplyslysvalgluprolyssercys210215220<210>6<211>672<212>dna<213>人工序列<220><223>v-区域+γ1ch1恒定区<400>6gaagtgcagttggtggagtcgggtggagggctggtgcagcctggcggtagcctgaggctg60tcctgtgccgtgtccggatactccattacctccggctactcgtggaactggatcagacag120gctcccggaaagggacttgagtgggtggcgtccatcacctacgacggctcaaccaactat180aacccgtccgtgaagggccgcatcaccatttcgcgcgacgacagcaagaatactttttac240ctccaaatgaacagcctgcgggccgaagatactgccgtgtactactgcgcgcggggatca300cattacttcgggcactggcacttcgccgtctggggacagggcaccctcgtcactgtctcg360agcgcttctacaaagggcccatcggtcttccccctggcaccctcctccaagagcacctct420gggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtg480tcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcc540tcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccag600acctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgag660cccaaatcttgt672<210>7<211>242<212>prt<213>人工序列<220><223>具有信号序列的v-区域+γ1ch1恒定区<400>7metlystrpvalthrpheileserleuleupheleupheserserala151015tyrsergluvalglnleuvalgluserglyglyglyleuvalglnpro202530glyglyserleuargleusercysalavalserglytyrserilethr354045serglytyrsertrpasntrpileargglnalaproglylysglyleu505560glutrpvalalaserilethrtyraspglyserthrasntyrasnpro65707580servallysglyargilethrileserargaspaspserlysasnthr859095phetyrleuglnmetasnserleuargalagluaspthralavaltyr100105110tyrcysalaargglyserhistyrpheglyhistrphisphealaval115120125trpglyglnglythrleuvalthrvalserseralaserthrlysgly130135140proservalpheproleualaproserserlysserthrserglygly145150155160thralaalaleuglycysleuvallysasptyrpheprogluproval165170175thrvalsertrpasnserglyalaleuthrserglyvalhisthrphe180185190proalavalleuglnserserglyleutyrserleuserservalval195200205thrvalproserserserleuglythrglnthrtyrilecysasnval210215220asnhislysproserasnthrlysvalasplyslysvalgluprolys225230235240sercys<210>8<211>726<212>dna<213>人工序列<220><223>具有信号序列的v-区域+γ1ch1恒定区<400>8atgaagtgggtcaccttcatctccctgctgtttctgttctccagcgcctactccgaagtg60cagttggtggagtcgggtggagggctggtgcagcctggcggtagcctgaggctgtcctgt120gccgtgtccggatactccattacctccggctactcgtggaactggatcagacaggctccc180ggaaagggacttgagtgggtggcgtccatcacctacgacggctcaaccaactataacccg240tccgtgaagggccgcatcaccatttcgcgcgacgacagcaagaatactttttacctccaa300atgaacagcctgcgggccgaagatactgccgtgtactactgcgcgcggggatcacattac360ttcgggcactggcacttcgccgtctggggacagggcaccctcgtcactgtctcgagcgct420tctacaaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggc480acagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtgg540aactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcagga600ctctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctac660atctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaa720tcttgt726<210>9<211>451<212>prt<213>人工序列<220><223>v-区域+γ1全长恒定区<400>9gluvalglnleuvalgluserglyglyglyleuvalglnproglygly151015serleuargleusercysalavalserglytyrserilethrsergly202530tyrsertrpasntrpileargglnalaproglylysglyleuglutrp354045valalaserilethrtyraspglyserthrasntyrasnproserval505560lysglyargilethrileserargaspaspserlysasnthrphetyr65707580leuglnmetasnserleuargalagluaspthralavaltyrtyrcys859095alaargglyserhistyrpheglyhistrphisphealavaltrpgly100105110glnglythrleuvalthrvalserseralaserthrlysglyproser115120125valpheproleualaproserserlysserthrserglyglythrala130135140alaleuglycysleuvallysasptyrpheprogluprovalthrval145150155160sertrpasnserglyalaleuthrserglyvalhisthrpheproala165170175valleuglnserserglyleutyrserleuserservalvalthrval180185190proserserserleuglythrglnthrtyrilecysasnvalasnhis195200205lysproserasnthrlysvalasplyslysvalgluprolyssercys210215220asplysthrhisthrcysproprocysproalaprogluleuleugly225230235240glyproservalpheleupheproprolysprolysaspthrleumet245250255ileserargthrprogluvalthrcysvalvalvalaspvalserhis260265270gluaspprogluvallyspheasntrptyrvalaspglyvalgluval275280285hisasnalalysthrlysproargglugluglntyrasnserthrtyr290295300argvalvalservalleuthrvalleuhisglnasptrpleuasngly305310315320lysglutyrlyscyslysvalserasnlysalaleuproalaproile325330335glulysthrileserlysalalysglyglnproarggluproglnval340345350tyrthrleuproproserarggluglumetthrlysasnglnvalser355360365leuthrcysleuvallysglyphetyrproseraspilealavalglu370375380trpgluserasnglyglnprogluasnasntyrlysthrthrpropro385390395400valleuaspseraspglyserphepheleutyrserlysleuthrval405410415asplysserargtrpglnglnglyasnvalphesercysservalmet420425430hisglualaleuhisasnhistyrthrglnlysserleuserleuser435440445proglylys450<210>10<211>1353<212>dna<213>人工序列<220><223>v-区域+γ1全长恒定区<400>10gaagtgcagttggtggagtcgggtggagggctggtgcagcctggcggtagcctgaggctg60tcctgtgccgtgtccggatactccattacctccggctactcgtggaactggatcagacag120gctcccggaaagggacttgagtgggtggcgtccatcacctacgacggctcaaccaactat180aacccgtccgtgaagggccgcatcaccatttcgcgcgacgacagcaagaatactttttac240ctccaaatgaacagcctgcgggccgaagatactgccgtgtactactgcgcgcggggatca300cattacttcgggcactggcacttcgccgtctggggacagggcaccctcgtcactgtctcg360agcgcttctacaaagggcccctccgtgttcccgctcgctccatcatcgaagtctaccagc420ggaggcactgcggctctcggttgcctcgtgaaggactacttcccggagccggtgaccgtg480tcgtggaacagcggagccctgaccagcggggtgcacacctttccggccgtcttgcagtca540agcggcctttactccctgtcatcagtggtgactgtcccgtccagctcattgggaacccaa600acctacatctgcaatgtgaatcacaaacctagcaacaccaaggttgacaagaaagtcgag660cccaaatcgtgtgacaagactcacacttgtccgccgtgcccggcacccgaactgctggga720ggtcccagcgtctttctgttccctccaaagccgaaagacacgctgatgatctcccgcacc780ccggaggtcacttgcgtggtcgtggacgtgtcacatgaggacccagaggtgaagttcaat840tggtacgtggatggcgtcgaagtccacaatgccaaaactaagcccagagaagaacagtac900aattcgacctaccgcgtcgtgtccgtgctcacggtgttgcatcaggattggctgaacggg960aaggaatacaagtgcaaagtgtccaacaaggcgctgccggcaccgatcgagaaaactatc1020tccaaagcgaagggacagcctagggaacctcaagtctacacgctgccaccatcacgggaa1080gaaatgactaagaatcaagtctcactgacttgtctggtgaaggggttttaccctagcgac1140attgccgtggagtgggaatccaacggccagccagagaacaactacaagactacccctcca1200gtgctcgactcggatggatcgttcttcctttactcgaagctcaccgtggataagtcccgg1260tggcagcagggaaacgtgttctcctgctcggtgatgcatgaagccctccataaccactat1320acccaaaagtcgctgtccctgtcgccgggaaag1353<210>11<211>469<212>prt<213>人工序列<220><223>具有信号序列的v-区域+γ1全长恒定区<400>11metlystrpvalthrpheileserleuleupheleupheserserala151015tyrsergluvalglnleuvalgluserglyglyglyleuvalglnpro202530glyglyserleuargleusercysalavalserglytyrserilethr354045serglytyrsertrpasntrpileargglnalaproglylysglyleu505560glutrpvalalaserilethrtyraspglyserthrasntyrasnpro65707580servallysglyargilethrileserargaspaspserlysasnthr859095phetyrleuglnmetasnserleuargalagluaspthralavaltyr100105110tyrcysalaargglyserhistyrpheglyhistrphisphealaval115120125trpglyglnglythrleuvalthrvalserseralaserthrlysgly130135140proservalpheproleualaproserserlysserthrserglygly145150155160thralaalaleuglycysleuvallysasptyrpheprogluproval165170175thrvalsertrpasnserglyalaleuthrserglyvalhisthrphe180185190proalavalleuglnserserglyleutyrserleuserservalval195200205thrvalproserserserleuglythrglnthrtyrilecysasnval210215220asnhislysproserasnthrlysvalasplyslysvalgluprolys225230235240sercysasplysthrhisthrcysproprocysproalaprogluleu245250255leuglyglyproservalpheleupheproprolysprolysaspthr260265270leumetileserargthrprogluvalthrcysvalvalvalaspval275280285serhisgluaspprogluvallyspheasntrptyrvalaspglyval290295300gluvalhisasnalalysthrlysproargglugluglntyrasnser305310315320thrtyrargvalvalservalleuthrvalleuhisglnasptrpleu325330335asnglylysglutyrlyscyslysvalserasnlysalaleuproala340345350proileglulysthrileserlysalalysglyglnproargglupro355360365glnvaltyrthrleuproproserarggluglumetthrlysasngln370375380valserleuthrcysleuvallysglyphetyrproseraspileala385390395400valglutrpgluserasnglyglnprogluasnasntyrlysthrthr405410415proprovalleuaspseraspglyserphepheleutyrserlysleu420425430thrvalasplysserargtrpglnglnglyasnvalphesercysser435440445valmethisglualaleuhisasnhistyrthrglnlysserleuser450455460leuserproglylys465<210>12<211>1407<212>dna<213>人工序列<220><223>具有信号序列的v-区域+γ1全长恒定区<400>12atgaagtgggtcaccttcatctccctgctgtttctgttctccagcgcctactccgaagtg60cagttggtggagtcgggtggagggctggtgcagcctggcggtagcctgaggctgtcctgt120gccgtgtccggatactccattacctccggctactcgtggaactggatcagacaggctccc180ggaaagggacttgagtgggtggcgtccatcacctacgacggctcaaccaactataacccg240tccgtgaagggccgcatcaccatttcgcgcgacgacagcaagaatactttttacctccaa300atgaacagcctgcgggccgaagatactgccgtgtactactgcgcgcggggatcacattac360ttcgggcactggcacttcgccgtctggggacagggcaccctcgtcactgtctcgagcgct420tctacaaagggcccctccgtgttcccgctcgctccatcatcgaagtctaccagcggaggc480actgcggctctcggttgcctcgtgaaggactacttcccggagccggtgaccgtgtcgtgg540aacagcggagccctgaccagcggggtgcacacctttccggccgtcttgcagtcaagcggc600ctttactccctgtcatcagtggtgactgtcccgtccagctcattgggaacccaaacctac660atctgcaatgtgaatcacaaacctagcaacaccaaggttgacaagaaagtcgagcccaaa720tcgtgtgacaagactcacacttgtccgccgtgcccggcacccgaactgctgggaggtccc780agcgtctttctgttccctccaaagccgaaagacacgctgatgatctcccgcaccccggag840gtcacttgcgtggtcgtggacgtgtcacatgaggacccagaggtgaagttcaattggtac900gtggatggcgtcgaagtccacaatgccaaaactaagcccagagaagaacagtacaattcg960acctaccgcgtcgtgtccgtgctcacggtgttgcatcaggattggctgaacgggaaggaa1020tacaagtgcaaagtgtccaacaaggcgctgccggcaccgatcgagaaaactatctccaaa1080gcgaagggacagcctagggaacctcaagtctacacgctgccaccatcacgggaagaaatg1140actaagaatcaagtctcactgacttgtctggtgaaggggttttaccctagcgacattgcc1200gtggagtgggaatccaacggccagccagagaacaactacaagactacccctccagtgctc1260gactcggatggatcgttcttcctttactcgaagctcaccgtggataagtcccggtggcag1320cagggaaacgtgttctcctgctcggtgatgcatgaagccctccataaccactatacccaa1380aagtcgctgtccctgtcgccgggaaag1407<210>13<211>25<212>prt<213>人工序列<220><223>frh1<400>13gluvalglnleuvalgluserglyglyglyleuvalglnproglygly151015serleuargleusercysalavalser2025<210>14<211>11<212>prt<213>人工序列<220><223>cdrh1<400>14glytyrserilethrserglytyrsertrpasn1510<210>15<211>14<212>prt<213>人工序列<220><223>frh2<400>15trpileargglnalaproglylysglyleuglutrpvalala1510<210>16<211>16<212>prt<213>人工序列<220><223>cdrh2<400>16serilethrtyraspglyserthrasntyrasnproservallysgly151015<210>17<211>32<212>prt<213>人工序列<220><223>frh3<400>17argilethrileserargaspaspserlysasnthrphetyrleugln151015metasnserleuargalagluaspthralavaltyrtyrcysalaarg202530<210>18<211>12<212>prt<213>人工序列<220><223>cdrh3<400>18glyserhistyrpheglyhistrphisphealaval1510<210>19<211>11<212>prt<213>人工序列<220><223>frh4<400>19trpglyglnglythrleuvalthrvalserser1510<210>20<211>111<212>prt<213>人工序列<220><223>轻链v-区域<400>20aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalasertyrleugluserglyvalproser505560argpheserglyserglyserglythraspphethrleuthrileser65707580serleuglnprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelys100105110<210>21<211>333<212>dna<213>人工序列<220><223>轻链v-区域<400>21gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgacc60atcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtat120cagcagaagcccggaaaagccccaaagttgctgatctacgccgcctcataccttgaaagc180ggcgtgccttcgcgcttctcgggaagcgggtcgggcaccgatttcaccctgaccatttcg240tcgctgcagccggaggacttcgcgacttactactgccaacagtcccacgaggacccctat300acgtttggccagggaaccaaggtcgaaatcaag333<210>22<211>129<212>prt<213>人工序列<220><223>具有信号序列的轻链v-区域<400>22metlystrpvalthrpheileserleuleupheleupheserserala151015tyrseraspileglnleuthrglnserproserserleuseralaser202530valglyaspargvalthrilethrcysargalaserglnservalasp354045tyraspglyaspsertyrmetasntrptyrglnglnlysproglylys505560alaprolysleuleuiletyralaalasertyrleugluserglyval65707580proserargpheserglyserglyserglythraspphethrleuthr859095ileserserleuglnprogluaspphealathrtyrtyrcysglngln100105110serhisgluaspprotyrthrpheglyglnglythrlysvalgluile115120125lys<210>23<211>387<212>dna<213>人工序列<220><223>具有信号序列的轻链v-区域<400>23atgaagtgggtcaccttcatctccctgctgtttctgttctcctccgcctactccgatatt60cagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcact120tgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcag180aagcccggaaaagccccaaagttgctgatctacgccgcctcataccttgaaagcggcgtg240ccttcgcgcttctcgggaagcgggtcgggcaccgatttcaccctgaccatttcgtcgctg300cagccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgttt360ggccagggaaccaaggtcgaaatcaag387<210>24<211>218<212>prt<213>人工序列<220><223>轻链v-区域+κ恒定区<400>24aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalasertyrleugluserglyvalproser505560argpheserglyserglyserglythraspphethrleuthrileser65707580serleuglnprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelysarg100105110thrvalalaalaproservalpheilepheproproseraspglugln115120125leulysserglythralaservalvalcysleuleuasnasnphetyr130135140proargglualalysvalglntrplysvalaspasnalaleuglnser145150155160glyasnserglngluservalthrgluglnaspserlysaspserthr165170175tyrserleuserserthrleuthrleuserlysalaasptyrglulys180185190hislysvaltyralacysgluvalthrhisglnglyleuserserpro195200205valthrlysserpheasnargglyglucys210215<210>25<211>654<212>dna<213>人工序列<220><223>v-区域+κ恒定区<400>25gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgacc60atcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtat120cagcagaagcccggaaaagccccaaagttgctgatctacgccgcctcataccttgaaagc180ggcgtgccttcgcgcttctcgggaagcgggtcgggcaccgatttcaccctgaccatttcg240tcgctgcagccggaggacttcgcgacttactactgccaacagtcccacgaggacccctat300acgtttggccagggaaccaaggtcgaaatcaagcgtacggtagcggccccatctgtcttc360atcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctg420aataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcg480ggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagc540agcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtc600acccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt654<210>26<211>236<212>prt<213>人工序列<220><223>具有信号序列的v-区域+κ恒定区<400>26metlystrpvalthrpheileserleuleupheleupheserserala151015tyrseraspileglnleuthrglnserproserserleuseralaser202530valglyaspargvalthrilethrcysargalaserglnservalasp354045tyraspglyaspsertyrmetasntrptyrglnglnlysproglylys505560alaprolysleuleuiletyralaalasertyrleugluserglyval65707580proserargpheserglyserglyserglythraspphethrleuthr859095ileserserleuglnprogluaspphealathrtyrtyrcysglngln100105110serhisgluaspprotyrthrpheglyglnglythrlysvalgluile115120125lysargthrvalalaalaproservalpheilepheproproserasp130135140gluglnleulysserglythralaservalvalcysleuleuasnasn145150155160phetyrproargglualalysvalglntrplysvalaspasnalaleu165170175glnserglyasnserglngluservalthrgluglnaspserlysasp180185190serthrtyrserleuserserthrleuthrleuserlysalaasptyr195200205glulyshislysvaltyralacysgluvalthrhisglnglyleuser210215220serprovalthrlysserpheasnargglyglucys225230235<210>27<211>708<212>dna<213>人工序列<220><223>具有信号序列的v-区域+κ恒定区<400>27atgaagtgggtcaccttcatctccctgctgtttctgttctcctccgcctactccgatatt60cagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcact120tgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcag180aagcccggaaaagccccaaagttgctgatctacgccgcctcataccttgaaagcggcgtg240ccttcgcgcttctcgggaagcgggtcgggcaccgatttcaccctgaccatttcgtcgctg300cagccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgttt360ggccagggaaccaaggtcgaaatcaagcgtacggtagcggccccatctgtcttcatcttc420ccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataac480ttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaac540tcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcacc600ctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccat660cagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt708<210>28<211>23<212>prt<213>人工序列<220><223>frl1<400>28aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcys20<210>29<211>15<212>prt<213>人工序列<220><223>cdrl1<400>29argalaserglnservalasptyraspglyaspsertyrmetasn151015<210>30<211>15<212>prt<213>人工序列<220><223>frl2<400>30trptyrglnglnlysproglylysalaprolysleuleuiletyr151015<210>31<211>7<212>prt<213>人工序列<220><223>cdrl2<400>31alaalasertyrleugluser15<210>32<211>32<212>prt<213>人工序列<220><223>frl3<400>32glyvalproserargpheserglyserglyserglythraspphethr151015leuthrileserserleuglnprogluaspphealathrtyrtyrcys202530<210>33<211>9<212>prt<213>人工序列<220><223>cdrl3<400>33glnglnserhisgluaspprotyrthr15<210>34<211>10<212>prt<213>人工序列<220><223>frl4<400>34pheglyglnglythrlysvalgluilelys1510<210>35<211>111<212>prt<213>人工序列<220><223>s60m_s77r_q79r_v-区域<400>35aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalasertyrleugluserglyvalpromet505560argpheserglyserglyserglythraspphethrleuthrileser65707580argleuargprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelys100105110<210>36<211>333<212>dna<213>人工序列<220><223>s60m_s77r_q79r_v-区域<400>36gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgacc60atcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtat120cagcagaagcccggaaaagccccaaagttgctgatctacgccgcctcataccttgaaagc180ggcgtgcctatgcgcttctcgggaagcgggtcgggcaccgatttcaccctgaccatttcg240agactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctat300acgtttggccagggaaccaaggtcgaaatcaag333<210>37<211>129<212>prt<213>人工序列<220><223>具有信号序列的s60m_s77r_q79r_v-区域<400>37metlystrpvalthrpheileserleuleupheleupheserserala151015tyrseraspileglnleuthrglnserproserserleuseralaser202530valglyaspargvalthrilethrcysargalaserglnservalasp354045tyraspglyaspsertyrmetasntrptyrglnglnlysproglylys505560alaprolysleuleuiletyralaalasertyrleugluserglyval65707580prometargpheserglyserglyserglythraspphethrleuthr859095ileserargleuargprogluaspphealathrtyrtyrcysglngln100105110serhisgluaspprotyrthrpheglyglnglythrlysvalgluile115120125lys<210>38<211>387<212>dna<213>人工序列<220><223>具有信号序列的s60m_s77r_q79r_v-区域<400>38atgaagtgggtcaccttcatctccctgctgtttctgttctcctccgcctactccgatatt60cagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcact120tgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcag180aagcccggaaaagccccaaagttgctgatctacgccgcctcataccttgaaagcggcgtg240cctatgcgcttctcgggaagcgggtcgggcaccgatttcaccctgaccatttcgagactg300aggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgttt360ggccagggaaccaaggtcgaaatcaag387<210>39<211>218<212>prt<213>人工序列<220><223>s60m_s77r_q79r_v-区域+包括l154p的κ恒定区<400>39aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalasertyrleugluserglyvalpromet505560argpheserglyserglyserglythraspphethrleuthrileser65707580argleuargprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelysarg100105110thrvalalaalaproservalpheilepheproproseraspglugln115120125leulysserglythralaservalvalcysleuleuasnasnphetyr130135140proargglualalysvalglntrplysvalaspasnalaproglnser145150155160glyasnserglngluservalthrgluglnaspserlysaspserthr165170175tyrserleuserserthrleuthrleuserlysalaasptyrglulys180185190hislysvaltyralacysgluvalthrhisglnglyleuserserpro195200205valthrlysserpheasnargglyglucys210215<210>40<211>654<212>dna<213>人工序列<220><223>s60m_s77r_q79r_v-区域+包括l154p的κ恒定区<400>40gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgacc60atcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtat120cagcagaagcccggaaaagccccaaagttgctgatctacgccgcctcataccttgaaagc180ggcgtgcctatgcgcttctcgggaagcgggtcgggcaccgatttcaccctgaccatttcg240agactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctat300acgtttggccagggaaccaaggtcgaaatcaagcgtacggtagcggccccatctgtcttc360atcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctg420aataacttctatcccagagaggccaaagtacagtggaaggtggataacgccccgcaatcg480ggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagc540agcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtc600acccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt654<210>41<211>236<212>prt<213>人工序列<220><223>具有信号序列的s60m_s77r_q79r_v-区域+包括l154p的κ恒定区<400>41metlystrpvalthrpheileserleuleupheleupheserserala151015tyrseraspileglnleuthrglnserproserserleuseralaser202530valglyaspargvalthrilethrcysargalaserglnservalasp354045tyraspglyaspsertyrmetasntrptyrglnglnlysproglylys505560alaprolysleuleuiletyralaalasertyrleugluserglyval65707580prometargpheserglyserglyserglythraspphethrleuthr859095ileserargleuargprogluaspphealathrtyrtyrcysglngln100105110serhisgluaspprotyrthrpheglyglnglythrlysvalgluile115120125lysargthrvalalaalaproservalpheilepheproproserasp130135140gluglnleulysserglythralaservalvalcysleuleuasnasn145150155160phetyrproargglualalysvalglntrplysvalaspasnalapro165170175glnserglyasnserglngluservalthrgluglnaspserlysasp180185190serthrtyrserleuserserthrleuthrleuserlysalaasptyr195200205glulyshislysvaltyralacysgluvalthrhisglnglyleuser210215220serprovalthrlysserpheasnargglyglucys225230235<210>42<211>708<212>dna<213>人工序列<220><223>具有信号序列的s60m_s77r_q79r_v-区域+包括l154p的κ恒定区<400>42atgaagtgggtcaccttcatctccctgctgtttctgttctcctccgcctactccgatatt60cagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcact120tgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcag180aagcccggaaaagccccaaagttgctgatctacgccgcctcataccttgaaagcggcgtg240cctatgcgcttctcgggaagcgggtcgggcaccgatttcaccctgaccatttcgagactg300aggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgttt360ggccagggaaccaaggtcgaaatcaagcgtacggtagcggccccatctgtcttcatcttc420ccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataac480ttctatcccagagaggccaaagtacagtggaaggtggataacgccccgcaatcgggtaac540tcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcacc600ctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccat660cagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt708<210>43<211>32<212>prt<213>人工序列<220><223>frl3s64ms60m<400>43glyvalprometargpheserglyserglyserglythraspphethr151015leuthrileserserleuglnprogluaspphealathrtyrtyrcys202530<210>44<211>32<212>prt<213>人工序列<220><223>frl3s81rs77r<400>44glyvalproserargpheserglyserglyserglythraspphethr151015leuthrileserargleuglnprogluaspphealathrtyrtyrcys202530<210>45<211>32<212>prt<213>人工序列<220><223>frl3q83rq79r<400>45glyvalproserargpheserglyserglyserglythraspphethr151015leuthrileserserleuargprogluaspphealathrtyrtyrcys202530<210>46<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rs60ms77r<400>46glyvalprometargpheserglyserglyserglythraspphethr151015leuthrileserargleuglnprogluaspphealathrtyrtyrcys202530<210>47<211>32<212>prt<213>人工序列<220><223>frl3s64mq83rs60mq79r<400>47glyvalprometargpheserglyserglyserglythraspphethr151015leuthrileserserleuargprogluaspphealathrtyrtyrcys202530<210>48<211>32<212>prt<213>人工序列<220><223>frl3s81rq83rs77rq79r<400>48glyvalproserargpheserglyserglyserglythraspphethr151015leuthrileserargleuargprogluaspphealathrtyrtyrcys202530<210>49<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rq83rs60ms77rq79r<400>49glyvalprometargpheserglyserglyserglythraspphethr151015leuthrileserargleuargprogluaspphealathrtyrtyrcys202530<210>50<211>7<212>prt<213>人工序列<220><223>cdrl2s56ds52d<400>50alaalaasptyrleugluser15<210>51<211>7<212>prt<213>人工序列<220><223>cdrl2s56es52e<400>51alaalaglutyrleugluser15<210>52<211>32<212>prt<213>人工序列<220><223>frl3s71ms67m<400>52glyvalproserargpheserglyserglyserglythraspphethr151015leuthrileserserleuglnprogluaspphealathrtyrtyrcys202530<210>53<211>32<212>prt<213>人工序列<220><223>frl3s64ms71ms60ms67m<400>53glyvalprometargpheserglyserglymetglythraspphethr151015leuthrileserserleuglnprogluaspphealathrtyrtyrcys202530<210>54<211>32<212>prt<213>人工序列<220><223>frl3s81rs71ms77rs67m<400>54glyvalproserargpheserglyserglymetglythraspphethr151015leuthrileserargleuglnprogluaspphealathrtyrtyrcys202530<210>55<211>32<212>prt<213>人工序列<220><223>frl3q83rs71mq79rs67m<400>55glyvalproserargpheserglyserglymetglythraspphethr151015leuthrileserserleuargprogluaspphealathrtyrtyrcys202530<210>56<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rs71ms60ms77rs67m<400>56glyvalprometargpheserglyserglymetglythraspphethr151015leuthrileserargleuglnprogluaspphealathrtyrtyrcys202530<210>57<211>32<212>prt<213>人工序列<220><223>frl3s64mq83rs71ms60mq79rs67m<400>57glyvalprometargpheserglyserglymetglythraspphethr151015leuthrileserserleuargprogluaspphealathrtyrtyrcys202530<210>58<211>32<212>prt<213>人工序列<220><223>frl3s81rq83rs71ms77rq79rs67m<400>58glyvalproserargpheserglyserglymetglythraspphethr151015leuthrileserargleuargprogluaspphealathrtyrtyrcys202530<210>59<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rq83rs71ms60ms77rq79rs67m<400>59glyvalprometargpheserglyserglymetglythraspphethr151015leuthrileserargleuargprogluaspphealathrtyrtyrcys202530<210>60<211>32<212>prt<213>人工序列<220><223>frl3s67ys63y<400>60glyvalproserargphetyrglyserglyserglythraspphethr151015leuthrileserserleuglnprogluaspphealathrtyrtyrcys202530<210>61<211>32<212>prt<213>人工序列<220><223>frl3s64ms67ys60ms63y<400>61glyvalprometargphetyrglyserglyserglythraspphethr151015leuthrileserserleuglnprogluaspphealathrtyrtyrcys202530<210>62<211>32<212>prt<213>人工序列<220><223>frl3s81rs67ys77rs63y<400>62glyvalproserargphetyrglyserglyserglythraspphethr151015leuthrileserargleuglnprogluaspphealathrtyrtyrcys202530<210>63<211>32<212>prt<213>人工序列<220><223>frl3q83rs67yq79rs63y<400>63glyvalproserargphetyrglyserglyserglythraspphethr151015leuthrileserserleuargprogluaspphealathrtyrtyrcys202530<210>64<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rs67ys60ms77rs63y<400>64glyvalprometargphetyrglyserglyserglythraspphethr151015leuthrileserargleuglnprogluaspphealathrtyrtyrcys202530<210>65<211>32<212>prt<213>人工序列<220><223>frl3s64mq83rs67ys60mq79rs63y<400>65glyvalprometargphetyrglyserglyserglythraspphethr151015leuthrileserserleuargprogluaspphealathrtyrtyrcys202530<210>66<211>32<212>prt<213>人工序列<220><223>frl3s81rq83rs67ys77rq79rs63y<400>66glyvalproserargphetyrglyserglyserglythraspphethr151015leuthrileserargleuargprogluaspphealathrtyrtyrcys202530<210>67<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rq83rs67ys60ms77rq79rs63y<400>67glyvalprometargphetyrglyserglyserglythraspphethr151015leuthrileserargleuargprogluaspphealathrtyrtyrcys202530<210>68<211>32<212>prt<213>人工序列<220><223>frl3s80ns76n<400>68glyvalproserargpheserglyserglyserglythraspphethr151015leuthrileasnserleuglnprogluaspphealathrtyrtyrcys202530<210>69<211>32<212>prt<213>人工序列<220><223>frl3s64ms80ns60ms76n<400>69glyvalprometargpheserglyserglyserglythraspphethr151015leuthrileasnserleuglnprogluaspphealathrtyrtyrcys202530<210>70<211>32<212>prt<213>人工序列<220><223>frl3s81rs80ns77rs76<400>70glyvalproserargpheserglyserglyserglythraspphethr151015leuthrileasnargleuglnprogluaspphealathrtyrtyrcys202530<210>71<211>32<212>prt<213>人工序列<220><223>frl3q83rs80nq79rs76n<400>71glyvalproserargpheserglyserglyserglythraspphethr151015leuthrileasnserleuargprogluaspphealathrtyrtyrcys202530<210>72<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rs80ns60ms77rs76n<400>72glyvalprometargpheserglyserglyserglythraspphethr151015leuthrileasnargleuglnprogluaspphealathrtyrtyrcys202530<210>73<211>32<212>prt<213>人工序列<220><223>frl3s64mq83rs80ns60mq79rs76n<400>73glyvalprometargpheserglyserglyserglythraspphethr151015leuthrileasnserleuargprogluaspphealathrtyrtyrcys202530<210>74<211>32<212>prt<213>人工序列<220><223>frl3s81rq83rs80ns77rq79rs76n<400>74glyvalproserargpheserglyserglyserglythraspphethr151015leuthrileasnargleuargprogluaspphealathrtyrtyrcys202530<210>75<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rq83rs80ns60ms77rq79rs76n<400>75glyvalprometargpheserglyserglyserglythraspphethr151015leuthrileasnargleuargprogluaspphealathrtyrtyrcys202530<210>76<211>32<212>prt<213>人工序列<220><223>frl3s80ns67ys76ns63y<400>76glyvalproserargphetyrglyserglyserglythraspphethr151015leuthrileasnserleuglnprogluaspphealathrtyrtyrcys202530<210>77<211>32<212>prt<213>人工序列<220><223>frl3s64ms80ns67ys60ms76ns63y<400>77glyvalprometargphetyrglyserglyserglythraspphethr151015leuthrileasnserleuglnprogluaspphealathrtyrtyrcys202530<210>78<211>32<212>prt<213>人工序列<220><223>frl3s81rs80ns67ys77rs76ns63y<400>78glyvalproserargphetyrglyserglyserglythraspphethr151015leuthrileasnargleuglnprogluaspphealathrtyrtyrcys202530<210>79<211>32<212>prt<213>人工序列<220><223>frl3q83rs80ns67yq79rs76ns63y<400>79glyvalproserargphetyrglyserglyserglythraspphethr151015leuthrileasnserleuargprogluaspphealathrtyrtyrcys202530<210>80<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rs80ns67ys60ms77rs76ns63y<400>80glyvalprometargphetyrglyserglyserglythraspphethr151015leuthrileasnargleuglnprogluaspphealathrtyrtyrcys202530<210>81<211>32<212>prt<213>人工序列<220><223>frl3s64mq83rs80ns67ys60mq79rs76ns63y<400>81glyvalprometargphetyrglyserglyserglythraspphethr151015leuthrileasnserleuargprogluaspphealathrtyrtyrcys202530<210>82<211>32<212>prt<213>人工序列<220><223>frl3s81rq83rs80ns67ys77rq79rs76ns63y<400>82glyvalproserargphetyrglyserglyserglythraspphethr151015leuthrileasnargleuargprogluaspphealathrtyrtyrcys202530<210>83<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rq83rs80ns67ys60ms77rq79rs76ns63y<400>83glyvalprometargphetyrglyserglyserglythraspphethr151015leuthrileasnargleuargprogluaspphealathrtyrtyrcys202530<210>84<211>32<212>prt<213>人工序列<220><223>frl3s67ys71ms63ys67m<400>84glyvalproserargphetyrglyserglymetglythraspphethr151015leuthrileserserleuglnprogluaspphealathrtyrtyrcys202530<210>85<211>32<212>prt<213>人工序列<220><223>frl3s64ms67ys71ms60ms63ys67m<400>85glyvalprometargphetyrglyserglymetglythraspphethr151015leuthrileserserleuglnprogluaspphealathrtyrtyrcys202530<210>86<211>32<212>prt<213>人工序列<220><223>frl3s81rs67ys71ms77rs63ys67m<400>86glyvalproserargphetyrglyserglymetglythraspphethr151015leuthrileserargleuglnprogluaspphealathrtyrtyrcys202530<210>87<211>32<212>prt<213>人工序列<220><223>frl3q83rs67ys71mq79rs63ys67m<400>87glyvalproserargphetyrglyserglymetglythraspphethr151015leuthrileserserleuargprogluaspphealathrtyrtyrcys202530<210>88<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rs67ys71ms60ms77rs63ys67m<400>88glyvalprometargphetyrglyserglymetglythraspphethr151015leuthrileserargleuglnprogluaspphealathrtyrtyrcys202530<210>89<211>32<212>prt<213>人工序列<220><223>frl3s64mq83rs67ys71ms60mq79rs63ys67m<400>89glyvalprometargphetyrglyserglymetglythraspphethr151015leuthrileserserleuargprogluaspphealathrtyrtyrcys202530<210>90<211>32<212>prt<213>人工序列<220><223>frl3s81rq83rs67ys71ms77rq79rs63ys67m<400>90glyvalproserargphetyrglyserglymetglythraspphethr151015leuthrileserargleuargprogluaspphealathrtyrtyrcys202530<210>91<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rq83rs67ys71ms60ms77rq79rs63ys67m<400>91glyvalprometargphetyrglyserglymetglythraspphethr151015leuthrileserargleuargprogluaspphealathrtyrtyrcys202530<210>92<211>32<212>prt<213>人工序列<220><223>frl3s80ns71ms76ns67m<400>92glyvalproserargpheserglyserglymetglythraspphethr151015leuthrileasnserleuglnprogluaspphealathrtyrtyrcys202530<210>93<211>32<212>prt<213>人工序列<220><223>frl3s64ms80ns71ms60ms76ns67m(<400>93glyvalprometargpheserglyserglymetglythraspphethr151015leuthrileasnserleuglnprogluaspphealathrtyrtyrcys202530<210>94<211>32<212>prt<213>人工序列<220><223>frl3s81rs80ns71ms77rs76ns67m<400>94glyvalproserargpheserglyserglymetglythraspphethr151015leuthrileasnargleuglnprogluaspphealathrtyrtyrcys202530<210>95<211>32<212>prt<213>人工序列<220><223>frl3q83rs80ns71mq79rs76ns67m<400>95glyvalproserargpheserglyserglymetglythraspphethr151015leuthrileasnserleuargprogluaspphealathrtyrtyrcys202530<210>96<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rs80ns71ms60ms77rs76ns67m<400>96glyvalprometargpheserglyserglymetglythraspphethr151015leuthrileasnargleuglnprogluaspphealathrtyrtyrcys202530<210>97<211>32<212>prt<213>人工序列<220><223>frl3s64mq83rs80ns71ms60mq79rs76ns67m<400>97glyvalprometargpheserglyserglymetglythraspphethr151015leuthrileasnserleuargprogluaspphealathrtyrtyrcys202530<210>98<211>32<212>prt<213>人工序列<220><223>frl3s81rq83rs80ns71ms77rq79rs76ns67m<400>98glyvalproserargpheserglyserglymetglythraspphethr151015leuthrileasnargleuargprogluaspphealathrtyrtyrcys202530<210>99<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rq83rs80ns71ms60ms77rq79rs76ns67m<400>99glyvalprometargpheserglyserglymetglythraspphethr151015leuthrileasnargleuargprogluaspphealathrtyrtyrcys202530<210>100<211>32<212>prt<213>人工序列<220><223>frl3s80ns67ys71ms76ns63ys67m<400>100glyvalproserargphetyrglyserglymetglythraspphethr151015leuthrileasnserleuglnprogluaspphealathrtyrtyrcys202530<210>101<211>32<212>prt<213>人工序列<220><223>frl3s64ms80ns67ys71ms60ms76ns63ys67m<400>101glyvalprometargphetyrglyserglymetglythraspphethr151015leuthrileasnserleuglnprogluaspphealathrtyrtyrcys202530<210>102<211>32<212>prt<213>人工序列<220><223>frl3s81rs80ns67ys71ms77rs76ns63ys67m<400>102glyvalproserargphetyrglyserglymetglythraspphethr151015leuthrileasnargleuglnprogluaspphealathrtyrtyrcys202530<210>103<211>32<212>prt<213>人工序列<220><223>frl3q83rs80ns67ys71mq79rs76ns63ys67m<400>103glyvalproserargphetyrglyserglymetglythraspphethr151015leuthrileasnserleuargprogluaspphealathrtyrtyrcys202530<210>104<211>32<212>prt<213>人工序列<220><223>frl3s64ms81rs80ns67ys71ms60ms77rs76ns63ys67m<400>104glyvalprometargphetyrglyserglymetglythraspphethr151015leuthrileasnargleuglnprogluaspphealathrtyrtyrcys202530<210>105<211>32<212>prt<213>人工序列<220><223>frl3s64mq83rs80ns67ys71ms60mq79rs76ns63ys67m<400>105glyvalprometargphetyrglyserglymetglythraspphethr151015leuthrileasnserleuargprogluaspphealathrtyrtyrcys202530<210>106<211>32<212>prt<213>人工序列<220><223>frl3s81rq83rs80ns67ys71ms77rq79rs76ns63ys67m<400>106glyvalproserargphetyrglyserglymetglythraspphethr151015leuthrileasnargleuargprogluaspphealathrtyrtyrcys202530<210>107<211>30<212>prt<213>人工序列<220><223>frl3s64ms81rq83rs80ns67ys71ms60ms77rq79rs76ns63ys67m<400>107prometargphetyrglyserglymetglythraspphethrleuthr151015ileasnargleuargprogluaspphealathrtyrtyrcys202530<210>108<211>325<212>prt<213>人工序列<220><223>具有根据dorrington&bennich(1978)编号v224-k547的野生型人类ige-fc(ce2-ce4结构域)<400>108valalaserargaspphethrproprothrvallysileleuglnser151015sercysaspglyglyglyhispheproprothrileglnleuleucys202530leuvalserglytyrthrproglythrileasnilethrtrpleuglu354045aspglyglnvalmetaspvalaspleuserthralaserthrthrgln505560gluglygluleualaserthrglnsergluleuthrleuserglnlys65707580histrpleuseraspargthrtyrthrcysglnvalthrtyrglngly859095histhrphegluaspserthrlyslyscysalaaspserasnproarg100105110glyvalseralatyrleuserargproserpropheaspleupheile115120125arglysserprothrilethrcysleuvalvalaspleualaproser130135140lysglythrvalasnleuthrtrpserargalaserglylysproval145150155160asnhisserthrarglysgluglulysglnargasnglythrleuthr165170175valthrserthrleuprovalglythrargasptrpilegluglyglu180185190thrtyrglncysargvalthrhisprohisleuproargalaleumet195200205argserthrthrlysthrserglyproargalaalaprogluvaltyr210215220alaphealathrproglutrpproglyserargasplysargthrleu225230235240alacysleuileglnasnphemetprogluaspileservalglntrp245250255leuhisasngluvalglnleuproaspalaarghisserthrthrgln260265270proarglysthrlysglyserglyphephevalpheserargleuglu275280285valthrargalaglutrpgluglnlysaspglupheilecysargala290295300valhisglualaalaserproserglnthrvalglnargalavalser305310315320valasnproglylys325<210>109<211>111<212>prt<213>人工序列<220><223>s77r_q79r_v-区域(fab1)<400>109aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalasertyrleugluserglyvalproser505560argpheserglyserglyserglythraspphethrleuthrileser65707580argleuargprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelys100105110<210>110<211>333<212>dna<213>人工序列<220><223>77r_q79r_v-区域(fab1)<400>110gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgacc60atcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtac120cagcagaagcccggaaaagccccaaagttgctgatctacgccgcctcctaccttgaaagc180ggcgtgccttcacgcttctcgggaagcgggtctggcaccgatttcaccctgaccatttcg240agactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctat300acgtttggccagggtaccaaggtcgaaatcaag333<210>111<211>129<212>prt<213>人工序列<220><223>具有信号序列的s77r_q79r_v-区域(fab1)<400>111metlystrpvalthrpheileserleuleupheleupheserserala151015tyrseraspileglnleuthrglnserproserserleuseralaser202530valglyaspargvalthrilethrcysargalaserglnservalasp354045tyraspglyaspsertyrmetasntrptyrglnglnlysproglylys505560alaprolysleuleuiletyralaalasertyrleugluserglyval65707580proserargpheserglyserglyserglythraspphethrleuthr859095ileserargleuargprogluaspphealathrtyrtyrcysglngln100105110serhisgluaspprotyrthrpheglyglnglythrlysvalgluile115120125lys<210>112<211>387<212>dna<213>人工序列<220><223>具有信号序列的s77r_q79r_v-区域(fab1)<400>112atgaagtgggtcaccttcatctccctgctgtttctgttctcctccgcctactccgatatt60cagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcact120tgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtaccagcag180aagcccggaaaagccccaaagttgctgatctacgccgcctcctaccttgaaagcggcgtg240ccttcacgcttctcgggaagcgggtctggcaccgatttcaccctgaccatttcgagactg300aggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgttt360ggccagggtaccaaggtcgaaatcaag387<210>113<211>111<212>prt<213>人工序列<220><223>fab2_v-区域<400>113aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalasertyrleugluserglyvalproser505560argpheserglyserglyserglythraspphethrleuthrileser65707580serleuglnprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelys100105110<210>114<211>333<212>dna<213>人工序列<220><223>fab2_v-区域<400>114gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgacc60atcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtat120cagcagaagcccggaaaagccccaaagttgctgatctacgccgcctcataccttgaaagc180ggcgtgccttcgcgcttctcgggaagcgggtcgggcaccgatttcaccctgaccatttcg240tcgctgcagccggaggacttcgcgacttactactgccaacagtcccacgaggacccctat300acgtttggccagggaaccaaggtcgaaatcaag333<210>115<211>129<212>prt<213>人工序列<220><223>具有信号序列的fab2_v-区域<400>115metlystrpvalthrpheileserleuleupheleupheserserala151015tyrseraspileglnleuthrglnserproserserleuseralaser202530valglyaspargvalthrilethrcysargalaserglnservalasp354045tyraspglyaspsertyrmetasntrptyrglnglnlysproglylys505560alaprolysleuleuiletyralaalasertyrleugluserglyval65707580proserargpheserglyserglyserglythraspphethrleuthr859095ileserserleuglnprogluaspphealathrtyrtyrcysglngln100105110serhisgluaspprotyrthrpheglyglnglythrlysvalgluile115120125lys<210>116<211>387<212>dna<213>人工序列<220><223>具有信号序列的fab2_v-区域<400>116atgaagtgggtcaccttcatctccctgctgtttctgttctcctccgcctactccgatatt60cagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcact120tgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcag180aagcccggaaaagccccaaagttgctgatctacgccgcctcataccttgaaagcggcgtg240ccttcgcgcttctcgggaagcgggtcgggcaccgatttcaccctgaccatttcgtcgctg300cagccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgttt360ggccagggaaccaaggtcgaaatcaag387<210>117<211>218<212>prt<213>人工序列<220><223>fab2_v-区域+包括l154p的κ恒定区<400>117aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalasertyrleugluserglyvalproser505560argpheserglyserglyserglythraspphethrleuthrileser65707580serleuglnprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelysarg100105110thrvalalaalaproservalpheilepheproproseraspglugln115120125leulysserglythralaservalvalcysleuleuasnasnphetyr130135140proargglualalysvalglntrplysvalaspasnalaproglnser145150155160glyasnserglngluservalthrgluglnaspserlysaspserthr165170175tyrserleuserserthrleuthrleuserlysalaasptyrglulys180185190hislysvaltyralacysgluvalthrhisglnglyleuserserpro195200205valthrlysserpheasnargglyglucys210215<210>118<211>654<212>dna<213>人工序列<220><223>fab2_v-区域+包括l154p的κ恒定区<400>118gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgacc60atcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtat120cagcagaagcccggaaaagccccaaagttgctgatctacgccgcctcataccttgaaagc180ggcgtgccttcgcgcttctcgggaagcgggtcgggcaccgatttcaccctgaccatttcg240tcgctgcagccggaggacttcgcgacttactactgccaacagtcccacgaggacccctat300acgtttggccagggaaccaaggtcgaaatcaagcgtacggtagcggccccatctgtcttc360atcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctg420aataacttctatcccagagaggccaaagtacagtggaaggtggataacgccccgcaatcg480ggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagc540agcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtc600acccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt654<210>119<211>236<212>prt<213>人工序列<220><223>具有信号序列的fab2_v-区域+包括l154p的κ恒定区<400>119metlystrpvalthrpheileserleuleupheleupheserserala151015tyrseraspileglnleuthrglnserproserserleuseralaser202530valglyaspargvalthrilethrcysargalaserglnservalasp354045tyraspglyaspsertyrmetasntrptyrglnglnlysproglylys505560alaprolysleuleuiletyralaalasertyrleugluserglyval65707580proserargpheserglyserglyserglythraspphethrleuthr859095ileserserleuglnprogluaspphealathrtyrtyrcysglngln100105110serhisgluaspprotyrthrpheglyglnglythrlysvalgluile115120125lysargthrvalalaalaproservalpheilepheproproserasp130135140gluglnleulysserglythralaservalvalcysleuleuasnasn145150155160phetyrproargglualalysvalglntrplysvalaspasnalapro165170175glnserglyasnserglngluservalthrgluglnaspserlysasp180185190serthrtyrserleuserserthrleuthrleuserlysalaasptyr195200205glulyshislysvaltyralacysgluvalthrhisglnglyleuser210215220serprovalthrlysserpheasnargglyglucys225230235<210>120<211>708<212>dna<213>人工序列<220><223>具有信号序列的fab2_v-区域+包括l154p的κ恒定区<400>120atgaagtgggtcaccttcatctccctgctgtttctgttctcctccgcctactccgatatt60cagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcact120tgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcag180aagcccggaaaagccccaaagttgctgatctacgccgcctcataccttgaaagcggcgtg240ccttcgcgcttctcgggaagcgggtcgggcaccgatttcaccctgaccatttcgtcgctg300cagccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgttt360ggccagggaaccaaggtcgaaatcaagcgtacggtagcggccccatctgtcttcatcttc420ccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataac480ttctatcccagagaggccaaagtacagtggaaggtggataacgccccgcaatcgggtaac540tcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcacc600ctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccat660cagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt708<210>121<211>111<212>prt<213>人工序列<220><223>fab3_s77r_q79r_v-区域<400>121aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalasertyrleugluserglyvalproser505560argpheserglyserglyserglythraspphethrleuthrileser65707580argleuargprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelys100105110<210>122<211>333<212>dna<213>人工序列<220><223>fab3_s77r_q79r_v-区域<400>122gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgacc60atcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtac120cagcagaagcccggaaaagccccaaagttgctgatctacgccgcctcctaccttgaaagc180ggcgtgccttcacgcttctcgggaagcgggtctggcaccgatttcaccctgaccatttcg240agactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctat300acgtttggccagggtaccaaggtcgaaatcaag333<210>123<211>129<212>prt<213>人工序列<220><223>具有信号序列的fab3_s77r_q79r_v-区域<400>123metlystrpvalthrpheileserleuleupheleupheserserala151015tyrseraspileglnleuthrglnserproserserleuseralaser202530valglyaspargvalthrilethrcysargalaserglnservalasp354045tyraspglyaspsertyrmetasntrptyrglnglnlysproglylys505560alaprolysleuleuiletyralaalasertyrleugluserglyval65707580proserargpheserglyserglyserglythraspphethrleuthr859095ileserargleuargprogluaspphealathrtyrtyrcysglngln100105110serhisgluaspprotyrthrpheglyglnglythrlysvalgluile115120125lys<210>124<211>387<212>dna<213>人工序列<220><223>具有信号序列的fab3_s77r_q79r_v-区域<400>124atgaagtgggtcaccttcatctccctgctgtttctgttctcctccgcctactccgatatt60cagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcact120tgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtaccagcag180aagcccggaaaagccccaaagttgctgatctacgccgcctcctaccttgaaagcggcgtg240ccttcacgcttctcgggaagcgggtctggcaccgatttcaccctgaccatttcgagactg300aggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgttt360ggccagggtaccaaggtcgaaatcaag387<210>125<211>218<212>prt<213>人工序列<220><223>fab3_s77r_q79r_v-区域+包括l154p的κ恒定区<400>125aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalasertyrleugluserglyvalproser505560argpheserglyserglyserglythraspphethrleuthrileser65707580argleuargprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelysarg100105110thrvalalaalaproservalpheilepheproproseraspglugln115120125leulysserglythralaservalvalcysleuleuasnasnphetyr130135140proargglualalysvalglntrplysvalaspasnalaproglnser145150155160glyasnserglngluservalthrgluglnaspserlysaspserthr165170175tyrserleuserserthrleuthrleuserlysalaasptyrglulys180185190hislysvaltyralacysgluvalthrhisglnglyleuserserpro195200205valthrlysserpheasnargglyglucys210215<210>126<211>654<212>dna<213>人工序列<220><223>fab3_s77r_q79r_v-区域+包括l154p的κ恒定区<400>126gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgacc60atcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtac120cagcagaagcccggaaaagccccaaagttgctgatctacgccgcctcctaccttgaaagc180ggcgtgccttcacgcttctcgggaagcgggtctggcaccgatttcaccctgaccatttcg240agactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctat300acgtttggccagggtaccaaggtcgaaatcaagcgtacggtagcggccccatctgtcttc360atcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctg420aataacttctatcccagagaggccaaagtacagtggaaggtggataacgccccgcaatcg480ggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagc540agcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtc600acccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt654<210>127<211>236<212>prt<213>人工序列<220><223>具有信号序列的fab3_s77r_q79r_v-区域+包括l154p的κ恒定区<400>127metlystrpvalthrpheileserleuleupheleupheserserala151015tyrseraspileglnleuthrglnserproserserleuseralaser202530valglyaspargvalthrilethrcysargalaserglnservalasp354045tyraspglyaspsertyrmetasntrptyrglnglnlysproglylys505560alaprolysleuleuiletyralaalasertyrleugluserglyval65707580proserargpheserglyserglyserglythraspphethrleuthr859095ileserargleuargprogluaspphealathrtyrtyrcysglngln100105110serhisgluaspprotyrthrpheglyglnglythrlysvalgluile115120125lysargthrvalalaalaproservalpheilepheproproserasp130135140gluglnleulysserglythralaservalvalcysleuleuasnasn145150155160phetyrproargglualalysvalglntrplysvalaspasnalapro165170175glnserglyasnserglngluservalthrgluglnaspserlysasp180185190serthrtyrserleuserserthrleuthrleuserlysalaasptyr195200205glulyshislysvaltyralacysgluvalthrhisglnglyleuser210215220serprovalthrlysserpheasnargglyglucys225230235<210>128<211>708<212>dna<213>人工序列<220><223>具有信号序列的fab3_s77r_q79r_v-区域+包括l154p的κ恒定区<400>128atgaagtgggtcaccttcatctccctgctgtttctgttctcctccgcctactccgatatt60cagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcact120tgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtaccagcag180aagcccggaaaagccccaaagttgctgatctacgccgcctcctaccttgaaagcggcgtg240ccttcacgcttctcgggaagcgggtctggcaccgatttcaccctgaccatttcgagactg300aggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgttt360ggccagggtaccaaggtcgaaatcaagcgtacggtagcggccccatctgtcttcatcttc420ccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataac480ttctatcccagagaggccaaagtacagtggaaggtggataacgccccgcaatcgggtaac540tcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcacc600ctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccat660cagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt708<210>129<211>107<212>prt<213>人工序列<220><223>奥马珠单抗_v-区域<400>129thrglnserproserserleuseralaservalglyaspargvalthr151015ilethrcysargalaserglnservalasptyraspglyaspsertyr202530metasntrptyrglnglnlysproglylysalaprolysleuleuile354045tyralaalasertyrleugluserglyvalproserargphesergly505560serglyserglythraspphethrleuthrileserserleuglnpro65707580gluaspphealathrtyrtyrcysglnglnserhisgluaspprotyr859095thrpheglyglnglythrlysvalgluilelys100105<210>130<211>321<212>dna<213>人工序列<220><223>奥马珠单抗_v-区域<400>130actcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcacttgccgg60gcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcagaagccc120ggaaaagccccaaagttgctgatctacgccgcctcataccttgaaagcggcgtgccttcg180cgcttctcgggaagcgggtcgggcaccgatttcaccctgaccatttcgtcgctgcagccg240gaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgtttggccag300ggaaccaaggtcgaaatcaag321<210>131<211>32<212>prt<213>人工序列<220><223>fr3<400>131glyvalprometargpheserglyserglymetglythraspphethr151015leuthrileserargleuargprogluaspphealathrtyrtyrcys202530<210>132<211>111<212>prt<213>人工序列<220><223>s60m_s52d_s67m_s77r_q79r_v-区域<400>132aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalaasptyrleugluserglyvalpromet505560argpheserglyserglymetglythraspphethrleuthrileser65707580argleuargprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelys100105110<210>133<211>333<212>dna<213>人工序列<220><223>s60m_s52d_s67m_s77r_q79r_v-区域<400>133gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgacc60atcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtat120cagcagaagcccggaaaagccccaaagttgctgatctacgccgccgactaccttgaaagc180ggcgtgcctatgcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcg240agactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctat300acgtttggccagggaaccaaggtcgaaatcaag333<210>134<211>129<212>prt<213>人工序列<220><223>具有信号序列的s60m_s52d_s67m_s77r_q79r_v-区域<400>134metlystrpvalthrpheileserleuleupheleupheserserala151015tyrseraspileglnleuthrglnserproserserleuseralaser202530valglyaspargvalthrilethrcysargalaserglnservalasp354045tyraspglyaspsertyrmetasntrptyrglnglnlysproglylys505560alaprolysleuleuiletyralaalaasptyrleugluserglyval65707580prometargpheserglyserglymetglythraspphethrleuthr859095ileserargleuargprogluaspphealathrtyrtyrcysglngln100105110serhisgluaspprotyrthrpheglyglnglythrlysvalgluile115120125lys<210>135<211>387<212>dna<213>人工序列<220><223>具有信号序列的s60m_s52d_s67m_s77r_q79r_v-区域<400>135atgaagtgggtcaccttcatctccctgctgtttctgttctcctccgcctactccgatatt60cagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcact120tgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcag180aagcccggaaaagccccaaagttgctgatctacgccgccgactaccttgaaagcggcgtg240cctatgcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcgagactg300aggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgttt360ggccagggaaccaaggtcgaaatcaag387<210>136<211>218<212>prt<213>人工序列<220><223>s60m_s52d_s67m_s77r_q79r_v-区域+κ恒定区<400>136aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalaasptyrleugluserglyvalpromet505560argpheserglyserglymetglythraspphethrleuthrileser65707580argleuargprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelysarg100105110thrvalalaalaproservalpheilepheproproseraspglugln115120125leulysserglythralaservalvalcysleuleuasnasnphetyr130135140proargglualalysvalglntrplysvalaspasnalaleuglnser145150155160glyasnserglngluservalthrgluglnaspserlysaspserthr165170175tyrserleuserserthrleuthrleuserlysalaasptyrglulys180185190hislysvaltyralacysgluvalthrhisglnglyleuserserpro195200205valthrlysserpheasnargglyglucys210215<210>137<211>654<212>dna<213>人工序列<220><223>s60m_s52d_s67m_s77r_q79r_v-区域+κ恒定区<400>137gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgacc60atcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtat120cagcagaagcccggaaaagccccaaagttgctgatctacgccgccgactaccttgaaagc180ggcgtgcctatgcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcg240agactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctat300acgtttggccagggaaccaaggtcgaaatcaagcgtacggtagcggccccatctgtcttc360atcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctg420aataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcg480ggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagc540agcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtc600acccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt654<210>138<211>32<212>prt<213>人工序列<220><223>fr3<400>138glyvalproargargpheserglyserglymetglythraspphethr151015leuthrileserargleuargprogluaspphealathrtyrtyrcys202530<210>139<211>111<212>prt<213>人工序列<220><223>s60r_s52d_s67m_s77r_q79r_v-区域<400>139aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalaasptyrleugluserglyvalproarg505560argpheserglyserglymetglythraspphethrleuthrileser65707580argleuargprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelys100105110<210>140<211>333<212>dna<213>人工序列<220><223>s60r_s52d_s67m_s77r_q79r_v-区域<400>140gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgacc60atcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtat120cagcagaagcccggaaaagccccaaagttgctgatctacgccgccgattaccttgaaagc180ggcgtgcctcgtcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcg240agactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctat300acgtttggccagggaaccaaggtcgaaatcaag333<210>141<211>129<212>prt<213>人工序列<220><223>具有信号序列的s60r_s52d_s67m_s77r_q79r_v-区域<400>141metlystrpvalthrpheileserleuleupheleupheserserala151015tyrseraspileglnleuthrglnserproserserleuseralaser202530valglyaspargvalthrilethrcysargalaserglnservalasp354045tyraspglyaspsertyrmetasntrptyrglnglnlysproglylys505560alaprolysleuleuiletyralaalaasptyrleugluserglyval65707580proargargpheserglyserglymetglythraspphethrleuthr859095ileserargleuargprogluaspphealathrtyrtyrcysglngln100105110serhisgluaspprotyrthrpheglyglnglythrlysvalgluile115120125lys<210>142<211>387<212>dna<213>人工序列<220><223>具有信号序列的s60r_s52d_s67m_s77r_q79r_v-区域<400>142atgaagtgggtcaccttcatctccctgctgtttctgttctcctccgcctactccgatatt60cagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgaccatcact120tgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtatcagcag180aagcccggaaaagccccaaagttgctgatctacgccgccgattaccttgaaagcggcgtg240cctcgtcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcgagactg300aggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctatacgttt360ggccagggaaccaaggtcgaaatcaag387<210>143<211>218<212>prt<213>人工序列<220><223>s60r_s52d_s67m_s77r_q79r_v-区域+κ恒定区<400>143aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalaasptyrleugluserglyvalproarg505560argpheserglyserglymetglythraspphethrleuthrileser65707580argleuargprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelysarg100105110thrvalalaalaproservalpheilepheproproseraspglugln115120125leulysserglythralaservalvalcysleuleuasnasnphetyr130135140proargglualalysvalglntrplysvalaspasnalaleuglnser145150155160glyasnserglngluservalthrgluglnaspserlysaspserthr165170175tyrserleuserserthrleuthrleuserlysalaasptyrglulys180185190hislysvaltyralacysgluvalthrhisglnglyleuserserpro195200205valthrlysserpheasnargglyglucys210215<210>144<211>654<212>dna<213>人工序列<220><223>s60r_s52d_s67m_s77r_q79r_v-区域+κ恒定区<400>144gatattcagctgactcagagcccgagctcactctccgcttccgtgggggatagagtgacc60atcacttgccgggcatcccagtcggtggactacgacggagactcctacatgaactggtat120cagcagaagcccggaaaagccccaaagttgctgatctacgccgccgattaccttgaaagc180ggcgtgcctcgtcgcttctcgggaagcgggatgggcaccgatttcaccctgaccatttcg240agactgaggccggaggacttcgcgacttactactgccaacagtcccacgaggacccctat300acgtttggccagggaaccaaggtcgaaatcaagcgtacggtagcggccccatctgtcttc360atcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctg420aataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcg480ggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagc540agcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtc600acccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt654<210>145<211>111<212>prt<213>人工序列<220><223>s60k_s52d_s67m_s77r_q79r_v-区域<400>145aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalaasptyrleugluserglyvalprolys505560argpheserglyserglymetglythraspphethrleuthrileser65707580argleuargprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelys100105110<210>146<211>111<212>prt<213>人工序列<220><223>s60q_s52d_s67m_s77r_q79r_v-区域<400>146aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalaasptyrleugluserglyvalprogln505560argpheserglyserglymetglythraspphethrleuthrileser65707580argleuargprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelys100105110<210>147<211>506<212>prt<213>人工序列<220><223>具有信号序列的v-区域+γ1ch1恒定区加ca645gl4gh5scfv<400>147metlystrpvalthrpheileserleuleupheleupheserserala151015tyrsergluvalglnleuvalgluserglyglyglyleuvalglnpro202530glyglyserleuargleusercysalavalserglytyrserilethr354045serglytyrsertrpasntrpileargglnalaproglylysglyleu505560glutrpvalalaserilethrtyraspglyserthrasntyrasnpro65707580servallysglyargilethrileserargaspaspserlysasnthr859095phetyrleuglnmetasnserleuargalagluaspthralavaltyr100105110tyrcysalaargglyserhistyrpheglyhistrphisphealaval115120125trpglyglnglythrleuvalthrvalserseralaserthrlysgly130135140proservalpheproleualaproserserlysserthrserglygly145150155160thralaalaleuglycysleuvallysasptyrpheprogluproval165170175thrvalsertrpasnserglyalaleuthrserglyvalhisthrphe180185190proalavalleuglnserserglyleutyrserleuserservalval195200205thrvalproserserserleuglythrglnthrtyrilecysasnval210215220asnhislysproserasnthrlysvalasplyslysvalgluprolys225230235240sercysserglyglyglyglythrglyglyglyglysergluvalgln245250255leuleugluserglyglyglyleuvalglnproglyglyserleuarg260265270leusercysalavalserglyileaspleuserasntyralaileasn275280285trpvalargglnalaproglylyscysleuglutrpileglyileile290295300trpalaserglythrthrphetyralathrtrpalalysglyargphe305310315320thrileserargaspasnserlysasnthrvaltyrleuglnmetasn325330335serleuargalagluaspthralavaltyrtyrcysalaargthrval340345350proglytyrserthralaprotyrpheaspleutrpglyglnglythr355360365leuvalthrvalserserglyglyglyglyserglyglyglyglyser370375380glyglyglyglyserglyglyglyglyseraspileglnmetthrgln385390395400serproserservalseralaservalglyaspargvalthrilethr405410415cysglnserserproservaltrpserasnpheleusertrptyrgln420425430glnlysproglylysalaprolysleuleuiletyrglualaserlys435440445leuthrserglyvalproserargpheserglyserglyserglythr450455460aspphethrleuthrileserserleuglnprogluaspphealathr465470475480tyrtyrcysglyglyglytyrserserileseraspthrthrphegly485490495cysglythrlysvalgluilelysargthr500505<210>148<211>1518<212>dna<213>人工序列<220><223>具有信号序列的v-区域+γ1ch1恒定区加接头加ca645gl4gh5scfv<400>148atgaagtgggtcaccttcatctccctgctgtttctgttctccagcgcctactccgaagtg60cagttggtggagtcgggtggagggctggtgcagcctggcggtagcctgaggctgtcctgt120gccgtgtccggatactccattacctccggctactcgtggaactggatcagacaggctccc180ggaaagggacttgagtgggtggcgtccatcacctacgacggctcaaccaactataacccg240tccgtgaagggccgcatcaccatttcgcgcgacgacagcaagaatactttttacctccaa300atgaacagcctgcgggccgaagatactgccgtgtactactgcgcgcggggatcacattac360ttcgggcactggcacttcgccgtctggggacagggcaccctcgtcactgtctcgagcgct420tctacaaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggc480acagcggccctgggctgcctggtcaaggactacttccccgaaccagtgacggtgtcgtgg540aactcaggtgccctgaccagcggcgttcacaccttcccggctgtcctacagtcttcagga600ctctactccctgagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctac660atctgcaacgtgaatcacaagcccagcaacaccaaggtcgataagaaagttgagcccaaa720tcttgtagtggaggtgggggcaccggtggaggtggcagcgaggttcaactgcttgagtct780ggaggaggcctagtccagcctggagggagcctgcgtctctcttgtgcagtaagcggcatc840gacctgagcaattacgccatcaactgggtgagacaagctccggggaagtgtttagaatgg900atcggtataatatgggccagtgggacgaccttttatgctacatgggcgaaaggaaggttt960acaattagccgggacaatagcaaaaacaccgtgtatctccaaatgaactccttgcgagca1020gaggacacggcggtgtactattgtgctcgcactgtcccaggttatagcactgcaccctac1080ttcgatctgtggggacaagggaccctggtgactgtttcaagtggcggagggggtagtgga1140gggggtggctctgggggtggcggaagcggtggcgggggttctgacatacaaatgactcag1200tctccttcatcggtatccgcgtccgttggcgatagggtgactattacatgtcaaagctct1260cctagcgtctggagcaattttctatcctggtatcaacagaaaccggggaaggctccaaaa1320cttctgatttatgaagcctcgaaactcaccagtggagttccgtcaagattcagtggctct1380ggatcagggacagacttcacgttgacaatcagttcgctgcaaccagaggactttgcgacc1440tactattgtggtggaggttacagtagcataagtgatacgacatttgggtgcggtactaag1500gtggaaatcaaacgtacc1518<210>149<211>11<212>prt<213>人工序列<220><223>ch1和ca645gl4gh5scfv之间的接头<400>149serglyglyglyglythrglyglyglyglyser1510<210>150<211>253<212>prt<213>人工序列<220><223>ca645gl4gh5scfv<400>150gluvalglnleuleugluserglyglyglyleuvalglnproglygly151015serleuargleusercysalavalserglyileaspleuserasntyr202530alaileasntrpvalargglnalaproglylyscysleuglutrpile354045glyileiletrpalaserglythrthrphetyralathrtrpalalys505560glyargphethrileserargaspasnserlysasnthrvaltyrleu65707580glnmetasnserleuargalagluaspthralavaltyrtyrcysala859095argthrvalproglytyrserthralaprotyrpheaspleutrpgly100105110glnglythrleuvalthrvalserserglyglyglyglyserglygly115120125glyglyserglyglyglyglyserglyglyglyglyseraspilegln130135140metthrglnserproserservalseralaservalglyaspargval145150155160thrilethrcysglnserserproservaltrpserasnpheleuser165170175trptyrglnglnlysproglylysalaprolysleuleuiletyrglu180185190alaserlysleuthrserglyvalproserargpheserglysergly195200205serglythraspphethrleuthrileserserleuglnprogluasp210215220phealathrtyrtyrcysglyglyglytyrserserileseraspthr225230235240thrpheglycysglythrlysvalgluilelysargthr245250<210>151<211>20<212>prt<213>人工序列<220><223>scfv内ca645gh5和ca645gl4之间的接头<400>151glyglyglyglyserglyglyglyglyserglyglyglyglysergly151015glyglyglyser20<210>152<211>10<212>prt<213>人工序列<220><223>ca645cdrh1<400>152glyileaspleuserasntyralaileasn1510<210>153<211>16<212>prt<213>人工序列<220><223>ca645cdrh2<400>153ileiletrpalaserglythrthrphetyralathrtrpalalysgly151015<210>154<211>13<212>prt<213>人工序列<220><223>ca645cdrh3<400>154thrvalproglytyrserthralaprotyrpheaspleu1510<210>155<211>12<212>prt<213>人工序列<220><223>ca645cdrl1<400>155glnserserproservaltrpserasnpheleuser1510<210>156<211>7<212>prt<213>人工序列<220><223>ca645cdrl2<400>156glualaserlysleuthrser15<210>157<211>10<212>prt<213>人工序列<220><223>ca645cdrl3<400>157glyglytyrserserileseraspthrthr1510<210>158<211>111<212>prt<213>人工序列<220><223>s60m_s52d_s67m_s77r_q79r_s63w_s76n_v-区域<400>158aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalaasptyrleugluserglyvalpromet505560argphetrpglyserglymetglythraspphethrleuthrileasn65707580argleuargprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelys100105110<210>159<211>111<212>prt<213>人工序列<220><223>s60m_s52d_s67m_s77r_q79r_s63y_s76n_v-区域<400>159aspileglnleuthrglnserproserserleuseralaservalgly151015aspargvalthrilethrcysargalaserglnservalasptyrasp202530glyaspsertyrmetasntrptyrglnglnlysproglylysalapro354045lysleuleuiletyralaalaasptyrleugluserglyvalpromet505560argphetyrglyserglymetglythraspphethrleuthrileasn65707580argleuargprogluaspphealathrtyrtyrcysglnglnserhis859095gluaspprotyrthrpheglyglnglythrlysvalgluilelys100105110<210>160<211>18<212>prt<213>人工序列<220><223>信号序列<400>160metlystrpvalthrpheileserleuleupheleupheserserala151015tyrser当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1