生物样品中降解酶和非降解酶的检测、鉴定与纯化的制作方法
本申请要求2016年10月11日提交的美国临时申请系列号62/406,781的优先权,其通过引用全文纳入本文。
本公开一般涉及检测复杂生物样品中蛋白酶活性的方法,具体是在无需样品处理的情况下,检测复杂生物样品(例如血液、血浆或血清)中降解和非降解酶活性的方法。
背景技术:
已经鉴定了超过300种蛋白质翻译后修饰,它们大幅影响蛋白质功能,定位,活性和结构。常见的翻译后修饰包括磷酸化,糖基化,小蛋白质如泛素和sumo(小泛素相关修饰物)的偶联,以及n末端和赖氨酸残基的乙酰化,但蛋白质的蛋白水解修饰的重要性和普遍性往往被忽视。实际上,许多疾病的生物标志物是生物体液中的稳定蛋白水解片段。
蛋白酶是在蛋白质和多肽内进行肽键水解的酶。除了它们在消化和蛋白质转换中的作用外,蛋白酶在严格调控的级联和信号转导事件中起作用。通过精确的蛋白水解加工,蛋白酶参与每个生物过程,包括dna复制和修复,细胞周期进程,细胞增殖,分化和迁移,形态发生和组织重塑,神经元突触生长,止血,伤口愈合,免疫,血管生成和细胞凋亡。实际上,人们已经认识到超过53种特定的蛋白水解遗传性疾病,因此蛋白酶与许多病症有关并不奇怪。此外,许多病原体在侵入其宿主期间利用蛋白酶,在宿主内复制,或调节宿主的免疫应答。
由于这些原因,认为蛋白酶是治疗介入的重要靶标。蛋白酶占药物靶标的5-10%。有六类蛋白酶占任何生物基因的约2%(参见肽酶数据库merops获取有用信息),次于泛素连接酶,蛋白酶构成人类第二大酶家族,总共有超过567种蛋白酶。已经开发了几种蛋白酶抑制剂用于抗hiv感染,多发性骨髓瘤和高血压等疾病的药物。尽管取得了这些成功,但由于对靶向的蛋白酶功能作用缺乏选择性或有错误认识,许多蛋白酶抑制剂在临床试验中失败。
这些例子强调了在作为早期药物发现过程、生命科学研究和诊断开发的一部分的更复杂和生物学相关的环境中更好地理解蛋白酶活性的重要性。
技术实现要素:
本文提供了检测复杂生物样品中的蛋白酶活性的方法,尤其是在无需样品处理的情况下,检测复杂生物样品中降解和非降解酶活性的方法。在一些实施方式中,复杂生物样品选自血液、血浆和血清。
本文提供了鉴定复杂生物样品中蛋白酶活性的方法,包括:a)获得复杂生物样品;b)将该复杂样品与底物文库和标签孵育,以产生裂解产物;c)从该复杂样品分离裂解产物;和d)产生复杂样品的蛋白水解特征。
在所述方法的一些实施方式中,复杂生物样品是全血。
在所述方法的一些实施方式中,底物文库是合成肽文库。在一些实施方式中,合成肽文库包含以下部分的组合:氨基酸对,带正电和带负电的残基,或沿合成肽长度的不同位置处的修饰残基。
在所述方法的一些实施方式中,通过凝胶电泳,毛细管电泳或dc和ac动电学技术的组合将裂解产物与复杂样品分离。
在所述方法的一些实施方式中,通过根据迁移检测和对肽混合物中的个体底物进行分级来产生底物特征。在一些实施方式中,个体底物通过标签检测。在一些实施方式中,标签是荧光染料,放射性探针或亲和标签。
在所述方法的一些实施方式中,蛋白水解特征包含多类蛋白酶活性。在一些实施方式中,多类蛋白酶活性选自丝氨酸,半胱氨酸,苏氨酸,天冬氨酰和金属肽酶活性。在一些实施方式中,通过将复杂样品与活性抑制剂分开孵育来检测多类蛋白酶活性。在一些实施方式中,活性抑制剂选自金属螯合剂、半胱氨酸肽酶抑制剂和弹性蛋白酶特异性抑制剂。在一些实施方式中,活性抑制剂选自edta,e-64,cao74和氯甲基酮抑制剂。
本文提供了纯化生物样品中的蛋白酶的方法,包括:a)获得生物样品;b)使样品与基于活性的探针(abp)接触,所述探针包含与带正电的部分连接的化学反应性基团,其中abp结合生物样品中蛋白酶的活性位点,并且abp的带正电的部分保持暴露在蛋白酶活性位点外;和c)分离已结合abp的蛋白酶。
在所述方法的一些实施方式中,化学反应性基团通过间隔分子与带正电的部分连接。在所述方法的一些实施方式中,通过在体外将abp与样品一起孵育来进行接触。在一些实施方式中,化学反应性基团与活性位点亲核残基不可逆地反应。在一些实施方式中,通过电泳分离将已结合abp的蛋白酶与样品分离。
本文提供了观察生物样品中的蛋白酶的方法,包括:a)获得生物样品;b)使样品与基于活性的探针(abp)接触,所述探针包含与带正电的部分连接的化学反应性基团,其中abp结合生物样品中蛋白酶的活性位点,并且abp的带正电的部分保持暴露在蛋白酶活性位点外;和c)观察已结合abp的蛋白酶。
在所述方法的一些实施方式中,接触通过体内注射abp进行。在一些实施方式中,化学反应性基团通过间隔分子与带正电的部分连接。在一些实施方式中,化学反应性基团与活性位点亲核残基不可逆地反应。在一些实施方式中,abp还包含标签。在一些实施方式中,标签是荧光染料或放射性探针。
在一些实施方式中,该方法还包括在观察位置进行活检。在一些实施方式中,观察的位置是肿瘤。
在一些实施方式中,该方法还包括通过电泳分离法分离已结合abp的蛋白酶。
本文提供了观察体内酸性微环境中蛋白酶活性的方法,该方法包括,将带负电的荧光标记的肽或纳米颗粒注入生物体中,其中所述肽或纳米颗粒进入酸性微环境并在微环境中被蛋白酶裂解,以产生带正电的荧光裂解产物;和,观察裂解产物。
在所述方法的一些实施方式中,酸性微环境在肿瘤中。在一些实施方式中,裂解产物粘附于肿瘤的细胞表面。
本文提供了通过接触角测量来检测复杂生物样品中的两亲性酶底物的方法,包括:a)将复杂生物样品的液滴与两亲性底物一起孵育,所述两亲性底物平衡蛋白酶裂解位点周围的疏水性和亲水性,其中亲水和疏水产物从两亲性底物释放;和b)测量当疏水裂解产物迁移到液滴表面处的液滴-空气界面表面时形成的接触角,其中接触角的减小指示蛋白酶活性。
本文提供了通过荧光检测复杂生物样品中的两亲性酶底物的方法,包括:a)将复杂生物样品的液滴与荧光标记的两亲性底物一起孵育,所述两亲性底物平衡蛋白酶裂解位点周围的疏水性和亲水性,其中亲水和疏水产物从两亲性底物释放;和b)测量迁移到液滴表面处的液滴-空气界面表面的疏水裂解产物的荧光,荧光指示蛋白酶活性。
本文提供了检测生物样品中非降解性电荷转移酶的方法,包括,将带正电的底物与复杂样品孵育,其经设计以在通过样品中的激酶磷酸化之后带负电,和,测量电泳凝胶中的荧光。
在所述方法的一些实施方式中,底物包含选自丝氨酸、苏氨酸、酪氨酸及其组合的残基。
附图说明
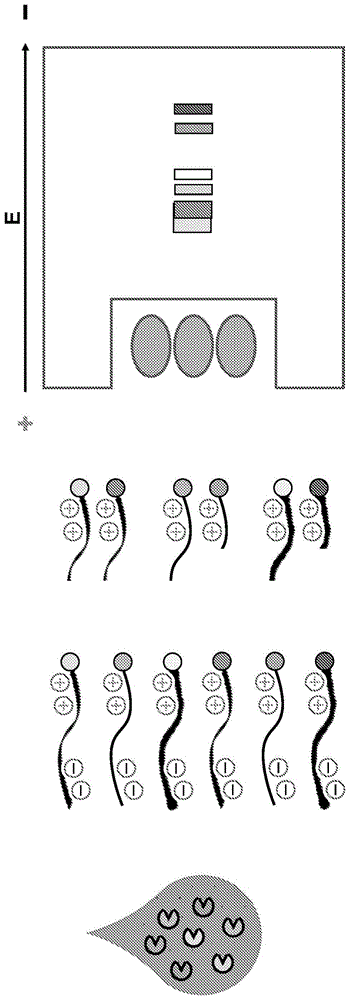
图1a-1b显示底物文库的使用和所得的底物裂解模式以鉴定样品中的酶。图1a说明了非靶向的底物文库的使用和所得的底物裂解模式,以鉴定样品中的多种酶。图1b说明了底物组和所得的底物裂解模式,以区分具有重叠裂解特异性的酶,例如mmp-2(上图)和mmp-9(下图)。
图2a-2d显示用于蛋白酶纯化的具有带电部分的基于活性的探针。图2a显示具有带负电的抑制区和带正电的尾部的蛋白酶抑制剂分子。图2b显示蛋白酶结合抑制剂分子并掩蔽带负电的抑制区,而尾部保持暴露于正电荷,但不裂解分子。图2c显示对样品施加电场以将带负电的血液组分与带正电的蛋白酶/抑制剂分离。图2d显示该方法用于通过分离所有抑制的蛋白酶并表征脱靶蛋白酶来筛选复杂样品中的抑制剂分子。
图3a-3d显示蛋白酶活性的体内成像和使用电荷改变型底物的定位。图3a显示将fret肽注射到动物中,其带负电荷以提供血流中更好的转运。图3b显示在到达肿瘤微环境时,肿瘤微环境的酸度使组氨酸质子化,使肽中性并能够渗透到肿瘤中。图3c显示设计为被与肿瘤微环境相关的蛋白酶裂解的肽被裂解且未淬灭(unquenched)。图3d显示带正电的裂解产物对细胞表面是“粘性的”。荧光信号指示肿瘤蛋白酶活性和肿瘤蛋白酶定位。
图4a-4e显示3步电荷改变型底物,其增强肿瘤的酸性微环境中的蛋白酶检测、成像和有效负载递送。图4a显示经设计以包含癌细胞表面展示的与癌症相关的蛋白酶的裂解位点的肽。该肽经设计以通过肿瘤相关的ph变化和癌细胞表面相关的蛋白水解裂解来改变电荷,以增强纳米颗粒(np)递送。图4b显示血流中带负电的np用于循环中的隐形(stealth)。图4c显示np在酸性肿瘤微环境中是中性的并且保持光滑(slippery)以深入扩散到肿瘤中。图4d显示癌细胞表面的蛋白酶裂解np表面的肽,使np带正电荷以供粘性细胞结合。图4e显示带正电的np粘附于癌细胞膜并被静电吸附,引发细胞摄取。
图5a-5b说明了两亲性酶底物的检测。图5a显示样品液滴的接触角指示产生疏水产物的蛋白酶活性。(1)将样品与底物一起孵育,所述底物平衡蛋白酶裂解位点周围的疏水性和亲水性;(2)蛋白酶裂解底物,产生亲水和疏水产物;和(3)疏水产物迁移至空气/液体界面并减小样品液滴的接触角,指示蛋白水解活性。图5b显示荧光疏水裂解产物在液滴表面组装,其中荧光没有被液滴组分淬灭。(1)将样品与底物孵育,所述底物平衡蛋白酶裂解位点周围的疏水性和亲水性;(2)蛋白酶裂解底物,产生亲水和疏水产物;和(3)疏水产物迁移到空气/液体界面,荧光显示在液滴表面,指示蛋白水解活性。
图6是电荷改变型底物的侧流试验/薄层色谱,显示(1)用于样品与底物孵育的中性表面和(2)结合血液的负性组分(negativecomponent)并造成正性裂解产物(positivecleavageproduct)的阳离子表面,所述正性裂解产物与液相的关联性高于固相。
图7a-7b显示检测非降解酶如磷酸化酶和激酶,所述检测采用电荷改变型底物,和从未处理样品的电泳分离。图7a显示(1)磷酸化酶从底物移除带负电的磷酸基团,例如磷酸化丝氨酸基团,和(2)带正电的去磷酸化底物的电泳分离。图7b显示(1)向底物添加带负电的磷酸基团的激酶,和(2)具有减少荧光的带负电的磷酸化底物的激酶活性的样品之间的比较。
图8a-8b显示电泳聚焦试验。
图9显示选择性靶向蛋白酶的电荷改变型肽序列,所述蛋白酶包括,凝血相关蛋白酶(凝血酶)、金属蛋白酶(mmp1、mmp3和mmp8)、组织蛋白酶(组织蛋白酶-s和组织蛋白酶-d)、细菌蛋白酶(omp-t和sspb)、激肽释放酶(激肽释放酶2和激肽释放酶3)和钙蛋白酶(钙蛋白酶1和钙蛋白酶2)。
图10a-10d显示采用凝血酶特异性电荷改变型肽底物的凝血酶检测。图10a显示凝血酶和肽底物在1xpbs中的孵育。图10b的图显示在1xpbs中孵育后的凝血酶浓度。图10c显示在柠檬酸化的血液中凝血酶和肽底物的孵育。图10d的图显示在柠檬酸化的血液中孵育后的凝血酶浓度。
图11a-11b显示采用凝血酶特异性电荷改变型肽底物的全血凝血酶检测。图11a说明不同孵育时间点的荧光信号。与阴性对照相比。图11b的图显示不同孵育时间点的荧光强度。
图12显示采用组织蛋白酶-s特异性电荷改变型荧光底物的缓冲液(1xpbs)检测中的组织蛋白酶-s。
图13a-13b说明服用抗凝血剂(阿司匹林)对比对照的对象的血液样品中的凝血酶活性。图13a显示对照对象(上图)对比抗凝血剂对象(下图)一定时间后的荧光。图13b的图显示随时间变化的凝血酶活性。
图14a-14c说明用于实时凝固监测的原型。将样品置于具有肽底物(例如,电荷改变型肽底物)的样品孔中(图14a),在t0测量荧光(图14b,上图)并且在稍后的时间点再次测量荧光(图14b,下图)。图14c显示可实时监测凝血酶活性。
图15a-15b显示小型化阵列原型。图15a显示小型化阵列原型。图15b显示电泳聚焦试验的结果,其中测量蛋白酶(胰蛋白酶)活性(作为荧光强度),对比浓度。
图16a-16b显示小型化阵列原型。图16a显示小型化阵列原型。图16b显示电泳聚焦试验的结果,其中测量一周后的蛋白酶(胰蛋白酶)活性(作为荧光强度),对比浓度。
图17a-17b显示小型化阵列原型。图17a显示小型化阵列原型。图17b显示电泳聚焦试验的结果,其中测量血液样品中的蛋白酶(胰蛋白酶)活性(作为荧光强度),对比浓度。
图18a-d显示小型化阵列原型和电泳聚焦试验的结果。
图19显示来自慢性淋巴细胞白血病对象的血浆样品中mmp2和mmp活性的检测,对比血浆样品中的对照(白色条)和经介电泳(dep)分离的血浆样品中的对照(黑色条)。
具体实施方式
本文提供了在无需样品处理的情况下,检测复杂生物样品中降解和非降解酶活性的方法。在一些实施方式中,基于它们在酶促作用(例如,裂解或部分添加/减少)时的性质变化,将性质变化(例如,荷电、疏水性)的酶底物的靶向或非靶向文库从生物样品分离。在一些实施方式中,性质变化(例如荷电,疏水性)的酶底物的分离通过电动力学方法、色谱技术和疏水-亲水的(例如,空气-液体)界面处的原位组装进行。在一些实施方式中,使用荧光/比色染料、阻抗/电容检测器、接触角和浊度/光吸收测量来检测电荷改变型和疏水性改变型酶底物的裂解产物,以指示复杂样品中的酶活性。在一些实施方式中,复杂生物样品选自血液、血浆和血清。在一些实施方式中,使用多种形式,包括非靶向文库平台、96孔板、侧流试验、盒、皮肤贴片和适于基于电荷/疏水性变化进行分离的微阵列。
本文还提供了能够在无需样品处理的情况下,从复杂样品纯化酶(例如蛋白酶)的方法。在一些实施方式中,复杂生物样品选自血液、血浆和血清。在一些实施方式中,使用通过其带负电的部分抑制/结合酶的裂解/变构位点的电荷平衡探针。在一些实施方式中,酶掩蔽探针的这种结合的带负电的部分,而探针的带正电的部分未被掩蔽且用于在电场的作用下从复杂样品中分离出酶。在一些实施方式中,复杂生物样品选自血液、血浆和血清。
在一些实施方式中,试验技术用于在几微升全血中直接检测和测量蛋白酶。在一些实施方式中,蛋白酶选自固有(indigenous)和非固有蛋白酶,包括但不限于胰蛋白酶、胰凝乳蛋白酶、mmp及其组合。在一些实施方式中,蛋白酶是凝血酶。许多固有的蛋白酶如凝血酶对血液凝固活性和相关的心脏疾病(dvt、动脉粥样硬化等)而言是重要的。某些非固有蛋白酶如胰蛋白酶、胰凝乳蛋白酶和mmp的出现通常是休克、癌症和其它疾病的生物标志物。因此,测量全血中蛋白酶活性的能力将使研究人员能够进一步阐明循环蛋白酶水平与许多重要疾病之间的关系。
本文提供了合成电荷改变型荧光肽底物,其对蛋白酶具有特异性。在一些实施方式中,蛋白酶选自胰蛋白酶、胰凝乳蛋白酶、mmp2、mmp9、弹性蛋白酶和凝血酶。在一些实施方式中,肽底物允许直接检测复杂生物样品中的蛋白酶活性并消除对样品准备的需要。在一些实施方式中,复杂生物样品选自血液、血浆和血清。在一些实施方式中,将底物添加至几微升的复杂样品中用于反应,然后以简单的电泳微凝胶形式快速分离裂解的产物。在一些实施方式中,复杂样品是血液。
在一些实施方式中,本发明的靶标是催化酶,包括但不限于蛋白酶、脂肪酶、核酸酶和非催化酶。在一些实施方式中,靶标是蛋白酶。
在本文提供的试验和方法的一些实施方式中,待分析的样品是生物样品。在一些实施方式中,输入材料是生物样品,例如血液、唾液、脑脊液或尿液。在一些实施方式中,生物样品是血液。在一些实施方式中,样品在置入该过程之前不需要特殊处理。
在一些实施方式中,本文提供的试验和方法可在实验室器皿中进行。在一些实施方式中,实验室器皿包括96孔板或培养瓶、反应容器例如盒,或类似于妊娠测试试剂盒的侧流装置等。在一些实施方式中,还考虑了用于放置在人体上的、允许连续监测的贴片型装置。
在一些实施方式中,反应要素是化学物质。在一些实施方式中,反应要素包括但不限于肽、dna和rna的合成序列。在一些实施方式中,化学物质带有“标签”以促进该过程。在一些实施方式中,化学物质被改造成带有亲水和疏水区域,或带正电和带负电的区域,但是也可选择其它区别特征。在一些实施方式中,在该方法中使用单一化学物质。在一些实施方式中,存在多种化学物质,其中可以选择多种化学物质以供反应输出的独特组合。这些多种化学物质将包含可用分子的“文库”以供使用。
在一些实施方式中,标签是荧光标签。在一些实施方式中,标签是发光的或放射性的。在一些实施方式中,标签包括由具有特定物理化学性质(例如电荷或疏水性/亲水性)的实体主导的序列。
在本文提供的试验和方法的一些实施方式中,发生的反应是化学反应。在一些实施方式中,化学反应是存在于生物样品中的酶消化/降解带标签化学物质的蛋白水解活性。在一些实施方式中,化学反应是结合或另一反应。带标签的化学物质作为酶的底物或蛋白酶特异性肽起作用。在一些实施方式中,反应裂解肽,释放疏水性或高度带电或以其他方式“极化”的实体。出于有用目的,这样的实体具有可以分离和测量的特性。或者,在一些实施方式中,带标签的化学物质用于结合样品中酶的区域或口袋,其中这种结合仅暴露化学物质的一部分,其中这种暴露允许进行分离和测量的方法。
在一些实施方式中,在反应之后,可以进行处理步骤。在一些实施方式中,处理步骤是电泳法、色谱法、侧流或移动到液体表面的疏水部分等。这可以是单个步骤或几个步骤。在一些实施方式中,该处理还可包括纯化或提取感兴趣的酶。
在一些实施方式中,在处理之后,可以进行测量。在一些实施方式中,测量为电、电导率、阻抗或比色。在一些实施方式中,测量结果由操作员或软件解释,从而能够呈现有用的结果,例如诊断性输出。
采用电荷改变型肽文库的蛋白酶活性分析
生物系统中肽酶的功能表征通常涉及鉴定候选蛋白质,确定底物特异性并产生特异性探针或抑制剂。尽管候选方法具有可靠的记录,但仍需要使用无偏全局方法来表征蛋白水解。最近开发了两种dna微阵列技术,hu/muprotin芯片和
为了研究这一点,引入了“反向降解(reversedegradomics)”的概念,该概念旨在测量复杂生物样品对特定底物的总体影响。将荧光标记的底物与生物样品孵育,并且在不同的时间点以高通量方式评估降解程度。这使得人们能够在与体内底物环境相近的环境中预测和比较复杂样品对例如关键疾病相关底物的总体影响。然而,这些反向技术,包括酶谱法和质谱法,依赖于耗时的方案、高度专业化的设备以及通过处理来操作样品。
鉴定复杂样品的蛋白水解特征
本文提供了鉴定复杂样品的蛋白水解特征的方法。在一些实施方式中,复杂生物样品选自血液、血浆和血清。在一些实施方式中,本文提供了产生复杂样品中酶类的综合底物特征的方法。在一些实施方式中,底物文库和所得的底物裂解模式用于鉴定样品中的酶。例如,可以制备合成肽文库,其包含氨基酸对的组合,带正电和带负电残基或沿合成肽长度的不同位置处的修饰残基的组合,以及标签如荧光染料。在一些实施方式中,在将复杂样品与底物文库孵育,并通过凝胶电泳、毛细管电泳或dc和ac动电学技术的组合从孵育的底物文库分离裂解产物后,可产生底物特征(图1a-1b)。在一些实施方式中,肽混合物中的个体底物根据迁移进行分级并通过荧光或其它标签来检测。在一些实施方式中,可以区分具有重叠裂解特异性的酶。例如,在一些实施方式中,可区分mmp-2和mmp-9。
在一些实施方式中,为了剖析多类蛋白酶活性(例如丝氨酸、半胱氨酸和金属肽酶)的贡献,样品在与不同活性抑制剂分开孵育后进行测试。在一些实施方式中,活性抑制剂是金属螯合剂edta,半胱氨酸肽酶抑制剂e-64和cao74,或弹性蛋白酶特异性氯甲基酮抑制剂。在一些实施方式中,将来自抑制的样品的裂解位点特异性数据组合以产生每种肽酶类别的底物特征。在一些实施方式中,产生二级合成肽文库以进一步确定复杂样品中肽酶的裂解偏好。在一些实施方式中,复杂生物样品选自血液、血浆和血清。
采用电荷改变型基于活性的探针的蛋白酶纯化
通过结合使用基于活性的探针(abp)和质谱分析,可以实现生物样品中存在的活性蛋白酶的鉴定和定量。abp是源自基于机制的抑制剂的分子。已经开发了几种酶类别的abp,但已证明对蛋白酶的研究特别有用。已经开发了靶向丝氨酸蛋白酶、半胱氨酸蛋白酶、苏氨酸蛋白酶、天冬氨酰蛋白酶和金属蛋白酶的abp。abp不可逆地与活性蛋白酶结合,但不与无活性的酶原或抑制的酶结合。abp包含化学反应性基团,其通常称为“弹头(warhead)”,通过间隔分子与标签部分连接。通过添加特异性增强模块可以增加探针的选择性。在abp与蛋白酶活性位点结合后,化学反应性基团与活性位点亲核残基不可逆地反应。abp的标签允许abp·蛋白酶复合物的观察(荧光团或放射性探针)或分离(亲和标签)。
然而,abp方法依赖于耗时的方案、高度专业化的设备以及通过处理来操作样品。需要能够对蛋白酶活性进行非放大研究的技术,如采用abp实现的(与可裂解底物法不同),其也在复杂样品中起作用但不需要专门的设备,例如质谱。在一些实施方式中,复杂生物样品选自血液、血浆和血清。
通过基于活性的探针进行体外/体内纯化
在一些实施方式中,本文提供了具有用于蛋白酶纯化的带电部分的基于活性的探针。在一些实施方式中,提供了abp设计,其包含通过间隔分子与带正电的部分连接的化学反应性基团(图2a-2d)。在一些实施方式中,蛋白酶抑制剂分子具有带负电的抑制区和带正电的尾部。在一些实施方式中,abp与样品在体外孵育或在体内注射。在一些实施方式中,abp结合蛋白酶活性位点,并且化学反应性基团与活性位点亲核残基不可逆地反应。在一些实施方式中,蛋白酶结合抑制剂分子但不裂解该分子。在一些实施方式中,蛋白酶结合抑制剂分子并掩蔽带负电的抑制区。在一些实施方式中,abp的带正电的部分(例如尾部)保持暴露在蛋白酶的活性位点外。
在一些实施方式中,体外电泳分离用于分离已结合abp的蛋白酶,因为带正电的部分将与复杂生物样品的带负电的大多数组分分离。在一些实施方式中,复杂生物样品选自血液、血浆和血清。在一些实施方式中,将电场施加至样品。在一些实施方式中,带负电的血液组分与带正电的蛋白酶/抑制剂分离。
在一些实施方式中,在体内注射具有带正电的部分的abp和识别标签(例如近红外或荧光染料)之后,可观察蛋白酶活性。在一些实施方式中,可以观察肿瘤部位的蛋白酶活性。在一些实施方式中,对肿瘤进行活检。在一些实施方式中,在活检后,通过电泳分离法将abp结合的蛋白酶与复杂样品分离。在一些实施方式中,已经从复杂样品分离的abp结合的蛋白酶可用于进一步的下游应用和翻译后修饰、突变等的表征。在一些实施方式中,复杂生物样品选自血液、血浆和血清。
蛋白酶检测和体内酸性微环境的成像
小动物全身成像技术的引入使体内酶谱(ivz)成为可能。该技术依赖于蛋白酶活化的荧光探针的使用。已经开发了含有不同荧光团的特异性探针,其允许同时检测各种酶。ivz允许在完整的生物体中映射mmp活性模式。然而,ivz探针不设计为优先定位于身体的某些微环境中,这可提供对难以进入的微观环境(例如肿瘤的酸性微环境)的更准确的成像。
通过肽进行体内蛋白酶检测
在一些实施方式中,提供了使用电荷改变型底物对蛋白酶活性成像和定位的方法。在一些实施方式中,成像是体内的。在一些实施方式中,蛋白酶检测技术能够基于电荷改变型荧光探针在酸性微环境中对局部蛋白酶活性进行体内成像(图3a-3d)。在一些实施方式中,将带负电的荧光标记的肽注射到生物体中。在一些实施方式中,肽是fret肽。在一些实施方式中,肽的负电荷在生物体的血流中提供隐形。在一些实施方式中,在遇到肿瘤的酸性微环境时,肽的组氨酸残基变为带正电的且肽是整体中性的。在一些实施方式中,肽呈中性并且能够穿透肿瘤。在一些实施方式中,中性电荷允许肽穿透肿瘤。在一些实施方式中,肽被蛋白酶裂解,其序列经设计。在一些实施方式中,肽被设计为被与肿瘤微环境相关的蛋白酶裂解。在一些实施方式中,肽被裂解且未淬灭。在一些实施方式中,裂解产物带正电并且带荧光或近红外标记,这使其粘附于细胞表面并提供蛋白酶活性和定位的成像。在一些实施方式中,荧光信号指示肿瘤蛋白酶活性和肿瘤蛋白酶定位。
纳米运载体的表面电荷影响它们向肿瘤组织中的渗透。胞外基质以有效的静电带通(electrostaticbandpass)形式呈现,抑制带正电和带负电物体的扩散运动,这允许不带电的粒子容易地扩散通过,同时有效地捕获带电粒子。用于递送至肿瘤的最佳颗粒在离开血管后应该是中性的(参见例如,sunq,radoszm,sheny,第3章:平移纳米运载体的合理设计(chapter3:rationaldesignoftranslationalnanocarriers).《用于纳米医药的功能性聚合物》(functionalpolymersfornanomedicine),younqingshen编)。
为向缺乏血管灌注的肿瘤中心区域(其中存在最具侵袭性和抗性的细胞)提供足够的药物浓度,纳米运载体在外渗后不应释放携带的药物,而是应进一步扩散深入肿瘤。这要求纳米运载体保持光滑并且具有尽可能小的尺寸。因此,纳米运载体最好是中性的,并且在到达肿瘤中心之前不呈现任何结合基团(包括靶向基团)。
在到达靶向的区域后,纳米运载体应高效地进入细胞以释放药物。此时,赋予纳米运载体隐形的相同特性可防止肿瘤细胞对它的细胞摄取。由于静电排斥,带负电的纳米运载体将被细胞膜排斥。因此,一旦进入肿瘤,运载体必须结合或粘附至靶向肿瘤细胞,以供快速细胞摄取。面临的挑战是如何协调这两个相反的需求,隐形循环和扩散,以及,粘附供于靶向。例如,众所周知,由于静电吸附引起快速细胞摄取,带正电的运载体可靠地粘附到细胞膜上,但带正电的运载体不适合体内应用,因为它们具有系统毒性且循环时间短(参见例如,sunq,radoszm,sheny,第3章:平移纳米运载体的合理设计(chapter3:rationaldesignoftranslationalnanocarriers).《用于纳米医药的功能性聚合物》(functionalpolymersfornanomedicine),younqingshen编)。
正电荷可以促进运载体在带负电的膜上的吸附,从而引发吸附介导的内吞作用。因此,另一种方法是通过“电荷反转”技术利用肿瘤胞外酸度使运载体具有正电荷。
通过肽修饰的纳米颗粒进行体内蛋白酶检测
在一些实施方式中,提供了使用肽修饰的纳米颗粒检测体内蛋白酶活性的方法。在一些实施方式中,提供了蛋白酶检测技术,其能够在酸性微环境中对局部蛋白酶活性进行体内成像。在一些实施方式中,该方法使用修饰有电荷改变型探针的纳米颗粒(图4a-4e)。在一些实施方式中,肽经设计以含有与癌症相关的蛋白酶的裂解位点。在一些实施方式中,与癌症相关的蛋白酶展示在癌细胞表面。在一些实施方式中,肽经设计以随肿瘤相关的ph变化改变电荷。在一些实施方式中,肽经设计以随癌细胞表面相关蛋白水解裂解改变电荷。在一些实施方式中,电荷变化和裂解增强纳米颗粒递送。
在本文提供方法的一些实施方式中,将用带负电的肽修饰的纳米颗粒注射到生物体中。在一些实施方式中,肽的负电荷在生物体的血流中提供隐形。在一些实施方式中,在遇到肿瘤的酸性微环境时,肽的组氨酸残基变为带正电且肽是整体中性的,这允许修饰的纳米颗粒穿透肿瘤。在一些实施方式中,纳米颗粒表面上的肽被蛋白酶裂解,其序列经设计。在一些实施方式中,裂解产物带正电,这使得修饰的纳米颗粒粘附至细胞表面。在一些实施方式中,带正电的纳米颗粒被静电吸附并引发细胞摄取。在一些实施方式中,纳米颗粒将其负载的成像剂和/或药物分子释放到肿瘤细胞中。
在本文提供方法的一些实施方式中,电荷改变型底物增强蛋白酶检测。在本文提供方法的一些实施方式中,电荷改变型底物增强蛋白酶成像。在本文提供方法的一些实施方式中,电荷改变型底物增强肿瘤酸性微环境中的负载物递送。
在本文提供方法的一些实施方式中,电荷改变型肽序列选择性地靶向蛋白酶,所述蛋白酶选自凝血相关蛋白酶、丝氨酸蛋白酶、金属蛋白酶、组织蛋白酶、细菌蛋白酶、激肽释放酶和钙蛋白酶(图9)。在一些实施方式中,凝血相关蛋白酶是凝血酶且电荷改变型肽序列具有序列ac-n-d--d--nle-t-p-r+//g-s-a-g-a-g-a-g-二氨基-乙基-bfl。在一些实施方式中,丝氨酸蛋白酶是胰蛋白酶。在一些实施方式中,金属蛋白酶选自mmp1、mmp2、mmp3、mmp8和mmp9。在一些实施方式中,金属蛋白酶是mmp1且电荷改变型肽序列具有序列ac-n-d--g-p-q-a//i-a-g-q-g-a-g-二氨基-乙基-bfl。在一些实施方式中,金属蛋白酶是mmp2。在一些实施方式中,金属蛋白酶是mmp3且电荷改变型肽序列具有序列ac-d--g-p-k+-p-v-e-//nva-y-n-k(ε-bfl)-nh2。在一些实施方式中,金属蛋白酶是mmp8且电荷改变型肽序列具有序列ac-n-d--g-p-q-g//y-a-g-q-g-a-g-二氨基-乙基-bfl。在一些实施方式中,金属蛋白酶是mmp9。在一些实施方式中,组织蛋白酶选自组织蛋白酶-s和组织蛋白酶-d。在一些实施方式中,组织蛋白酶是组织蛋白酶-s且电荷改变型肽序列具有序列n-suc--e--g-r+-w-h-t-v-g//l-r+-w-e--c(cy5-)-r+-co-nh2。在一些实施方式中,组织蛋白酶是组织蛋白酶-d且电荷改变型肽序列具有n-suc--d--d--l-v-v-l//f-v-k+-k+-c(cy5-)-a-co-nh2所示的序列。在一些实施方式中,细菌蛋白酶选自omp-t和sspb。在一些实施方式中,细菌蛋白酶是omp-t且电荷改变型肽序列具有序列ac-suc--d--g-d--k+-y-r+//r+-a-w-g-d-t-i-二氨基-乙基-bfl。在一些实施方式中,细菌蛋白酶是sspb且电荷改变型肽序列具有序列ac-d--g-d--a-f-s//k+-a-l-p-k(ε-bfl)-nh2。在一些实施方式中,激肽释放酶选自激肽释放酶2和激肽释放酶3。在一些实施方式中,激肽释放酶是激肽释放酶2且电荷改变型肽序列具有序列ac-n-d--g-d--t-f-r+//s-a-a-g-k(ε-bfl)-nh2。在一些实施方式中,激肽释放酶是激肽释放酶3且电荷改变型肽序列具有序列ac-n-d--g-s-s-i-y//q-s-s-t-g-二氨基-乙基-bfl。在一些实施方式中,钙蛋白酶选自钙蛋白酶1和钙蛋白酶2。在一些实施方式中,钙蛋白酶是钙蛋白酶1或钙蛋白酶2且电荷改变型肽序列具有序列n-suc--e--p-l-f//a-a-r+-k(ε-bfl)-nh2。
两亲性酶底物的检测
接触角测量
本文提供了检测两亲性酶底物的方法。在一些实施方式中,该方法包括测量样品液滴的接触角。在一些实施方式中,提供了检测复杂样品的液滴中的蛋白酶活性的技术。在一些实施方式中,复杂生物样品选自血液、血浆和血清。在一些实施方式中,蛋白酶活性通过接触角测量检测(图5a)。在一些实施方式中,样品液滴的接触角指示蛋白酶活性。在一些实施方式中,蛋白酶活性产生疏水产物。在一些实施方式中,两亲性底物经设计以在感兴趣的蛋白酶的裂解位点周围含有疏水性和亲水性部分。在一些实施方式中,底物与复杂样品的液滴孵育。在一些实施方式中,复杂生物样品选自血液、血浆和血清。在一些实施方式中,发生蛋白水解切割。在一些实施方式中,蛋白水解切割产生亲水和疏水产物。在一些实施方式中,当发生蛋白水解裂解时,从两亲性底物释放亲水和疏水产物。在一些实施方式中,疏水裂解产物优先在样品液滴表面的液滴-空气界面处组装。在一些实施方式中,随着更多疏水裂解产物迁移到液滴表面,液滴的接触角减小。在一些实施方式中,测量接触角作为蛋白酶活性的指标。
在液滴表面上检测
本文提供了检测两亲性酶底物的方法。在一些实施方式中,该方法包括测量样品液滴的荧光。在一些实施方式中,提供了检测复杂样品的液滴中的蛋白酶活性的技术。在一些实施方式中,复杂生物样品选自血液、血浆和血清。在一些实施方式中,通过液滴表面处的荧光检测来检测蛋白酶活性(图5b)。在一些实施方式中,荧光标记的两亲性底物经设计以在感兴趣的蛋白酶的裂解位点周围含有疏水和亲水部分。在一些实施方式中,底物与复杂样品的液滴孵育。在一些实施方式中,复杂生物样品选自血液、血浆和血清。在一些实施方式中,发生蛋白水解切割。在一些实施方式中,蛋白水解切割产生亲水和疏水产物。在一些实施方式中,当发生蛋白水解裂解时,从两亲性底物释放亲水和疏水产物。在一些实施方式中,荧光标记的疏水裂解产物优先在样品液滴表面的液滴-空气界面处组装。在一些实施方式中,检测液滴表面处的荧光。在一些实施方式中,液滴表面处的荧光是蛋白酶活性的指标,其中由样品所致的猝灭最小。
用于蛋白酶检测试验的侧流试验形式
本文提供了基于电荷改变型底物的蛋白酶检测试验的侧流试验形式(图6)。在一些实施方式中,将中性或带负电的底物与复杂样品在侧流表面的中性部分上孵育。在一些实施方式中,复杂生物样品选自血液、血浆和血清。在一些实施方式中,中性孵育部分与侧流表面的阳离子部分相邻。在一些实施方式中,样品组分和带负电的裂解产物与带正电的固体表面相关联。在一些实施方式中,带正电的裂解产物是荧光或比色标记的。在一些实施方式中,荧光或比色标记的、带正电的裂解产物与侧流试验的液相相关联。在一些实施方式中,阳离子表面结合血液的负性组分。在一些实施方式中,阳离子表面结合血液的负性组分,并使正性裂解产物与液相的关联性高于固相。在一些实施方式中,裂解产物比样品组分迁移得更远。在一些实施方式中,裂解产物比带负电的裂解产物迁移得更远。在一些实施方式中,裂解产物比样品组分和带负电的裂解产物迁移得更远。在一些实施方式中,迁移的带正电的裂解产物通过其荧光标记检测。在一些实施方式中,迁移的带正电的裂解产物通过其比色标记检测。在一些实施方式中,荧光或比色标记被检测为远离猝灭样品的条带。在一些实施方式中,条带指示蛋白酶活性。
非降解性电荷转移酶的检测
本文提供了检测未处理样品中的非降解性电荷转移酶的方法。在一些实施方式中,方法使用电荷改变型底物和来自未处理样品的电泳分离。在一些实施方式中,非降解酶选自磷酸化酶和激酶。在一些实施方式中,将具有磷酸化残基的底物与复杂样品孵育(图7a)。在一些实施方式中,复杂生物样品选自血液、血浆和血清。在一些实施方式中,具有磷酸化残基的底物选自丝氨酸、苏氨酸和酪氨酸。在一些实施方式中,具有磷酸化残基的底物被设计成在磷酸基团裂解时变为带正电。在一些实施方式中,磷酸基团的裂解通过样品中存在的磷酸化酶发生。在一些实施方式中,将带正电的反应产物与复杂样品分离。在一些实施方式中,通过施加电场将带正电的反应产物与复杂样品分离。
在一些实施方式中,将带正电的底物与复杂样品孵育。在一些实施方式中,复杂生物样品选自血液、血浆和血清。在一些实施方式中,带正电的底物含有选自丝氨酸、苏氨酸和酪氨酸的残基。在一些实施方式中,底物经设计以在样品中的激酶磷酸化后变为带负电。在一些实施方式中,可以在从复杂样品电泳分离磷酸化产物后比较样品间的相对激酶活性(图7b)。在一些实施方式中,激酶将带负电的磷酸基团添加至底物。在一些实施方式中,与具有较低激酶活性的样品相比,更高的激酶活性导致荧光减少,因为磷酸化的底物变为带负电并且在复杂样品(而非带正电的非磷酸化底物)的方向上分离。在一些实施方式中,激酶活性通过测量电泳凝胶中非磷酸化底物条带处的荧光减少来推断。
用于血液中蛋白酶检测试验的ac/dc动电学微阵列装置
使用ac/dc动电学微阵列装置进行蛋白酶和其他疾病相关酶检测的能力对于开发护理点(poc)和其他形式的高灵敏度和特异度试验而言具有特殊优势。然而,电子微阵列装置的一般问题是可施加到装置的相对低水平的ac和dc电压和电流降低了它们在高电导(>0.1s/m)缓冲液和生物样品(血液、血浆、血清)中的有效性。
ac/dc动电学微阵列装置
在一个实施方式中,本文提供了可以在ac和dc模式下操作的稳健的微阵列装置。在一些实施方式中,每个微电极可通过直接微布线或嵌入式cmos或其他半导体控制元件独立寻址。在一些实施方式中,此类器件通过光刻和其他众所周知的用于制造微电子器件的技术制造。在一些实施方式中,微阵列在硅、玻璃、陶瓷或塑料基础基底材料上制造。在一些实施方式中,使用二氧化硅和硝酸硅材料进一步绝缘微阵列。在一些实施方式中,微电极和较大电极结构由铂、钯、金、碳、其组合或其他耐电化学降解的导体材料制成。在一些实施方式中,使用微电子领域公知的溅射、电子束或其他沉积技术将这种微电极和电极制造到支持结构上。在一些实施方式中,关键要求是微电极结构坚固并且耐电化学降解。在一些实施方式中,微电极较厚(100nm至10微米)并且在减少缺陷的条件下沉积且允许微电极/电极用作真固体金属。在一些实施方式中,阵列装置上的微电极具有各种形状,包括但不限于圆形、椭圆形、环形、正方形、矩形和细长线状结构。在一些实施方式中,阵列装置上的微电极具有10nm至900微米的各种尺寸。在一些实施方式中,电极结构的尺寸范围为1mm至10cm。在一些实施方式中,阵列上微电极的数量范围为两至两万,间隔范围为100nm至1mm。在一些实施方式中,微电极阵列结构覆盖有多孔结构或渗透层。在一些实施方式中,这允许样品溶液(电解质、阳离子和阴离子)接触金属电极表面,但防止较大的生物分子到达表面,在那里它们可被电解产物(h+、oh-、o2、h2、自由基、热量、气泡等)破坏。这种渗透层可包括但不限于水凝胶(琼脂糖、聚丙烯酰胺等)、玻璃、有机物、聚合物、尼龙网状或多孔结构;以及实现所需渗透层特性的纳米结构的平面结构(lawn)。在一些实施方式中,渗透层的厚度范围为10nm至500微米。在一些实施方式中,渗透层用共价和非共价结合剂浸渍。在一些实施方式中,结合剂允许其他结合剂的附着。此类结合剂包括但不限于胺、硫醇基、羧基、生物素、链霉亲和素、抗体、适体、寡核苷酸等。在一些实施方式中,结合剂位于微阵列的特定区域中(在特定微电极上)。在一些实施方式中,不同的结合剂位于微阵列的不同区域。在一些实施方式中,将动电学微阵列(芯片)封闭在具有相关微流体的样品室结构中,所述微流体允许样品、洗涤缓冲液和试剂以受控方式递送。在一些实施方式中,样品室结构具有固定微阵列(芯片),防止与电解质接触并提供与微阵列的外部电连接以提供进行ac介电电泳(dep)和ac电泳过程所需的ac和dc电压和电流的组件。在一些实施方式中,样品室结构还在微阵列(芯片)上方具有玻璃、二氧化硅或塑料窗口,其允许视觉监测以及色彩和/或落射荧光检测。在一些实施方式中,封闭在流体样品室中的微阵列(芯片)连接至ac/dc电源和功能发生器;用于递送样品、洗涤缓冲液和试剂且用于接受洗脱样品和流体废液的相关流体组分;视觉监测和落射荧光显微镜及相关计算机,用于整体控制、芯片监测、检测和分析。在一些实施方式中,所有组分构成完整的蛋白酶试验检测系统。
ac模式下进行的微阵列蛋白酶试验
在ac模式下的甚至稳健微阵列装置用于血液中介电泳(dep)分离的操作情况中,在侵入性电化学开始破坏该过程之前,有效工作电压被限制为<25伏ac。在这些条件下(<25伏ac),红色和白色细胞被浓集至dep高场区域(微电极之间),纳米级实体(20nm-800nm)被浓集至dep高场区域(在微电极上),而较小的生物分子保持相对不受dep场的影响。在分离之后,可以利用流体洗涤从微阵列装置去除细胞和其他低分子量血浆组分,同时纳米级实体保持在dep高场区域中的浓集。
在一些实施方式中,在以ac模式操作的微阵列装置上的蛋白酶试验利用特殊纳米颗粒实体以及对电荷改变肽产物具有亲和性的电荷改变型肽底物。在一些实施方式中,当肽底物被血液中的蛋白酶裂解时,电荷改变型肽产物变得与特定纳米颗粒实体结合。在一些实施方式中,这些纳米颗粒实体包括但不限于尺寸范围为20nm至800nm的荧光和非荧光纳米颗粒;聚合物、脂质、金属、半导体和陶瓷纳米粒子;具有负电荷或正电荷的纳米颗粒、用配体结合剂修饰的纳米颗粒,用于共价和非共价结合,包括抗体、适体和凝集素。在一些实施方式中,在蛋白酶反应完成后,施加ac场且将具有结合的肽产物的纳米颗粒浓集至微电极上的dep高场区域中,同时将血液细胞浓集至低场区域中。在一些实施方式中,然后施加流体洗涤。在一些实施方式中,流体洗涤去除血液细胞和血浆组分以及任何未反应的电荷改变型肽底物。在一些实施方式中,纳米颗粒-肽产物保持在检测得到它们的dep高场区域中浓集。
在一些实施方式中,特定蛋白酶试验使用带负电的荧光电荷改变型肽底物进行,所述带负电的荧光电荷改变型肽底物产生带正电的荧光肽产物和带负电的非荧光纳米颗粒。在一些实施方式中,特定蛋白酶在血液中进行裂解后,带正电的荧光肽产物与带负电的非荧光纳米颗粒结合。在一些实施方式中,施加ac场并且将具有结合的荧光电荷改变型肽产物的纳米颗粒分离到dep高场区域中。在一些实施方式中,然后使用流体洗涤来去除细胞、血浆材料和任何未反应的带负电的荧光电荷改变型肽底物。在一些实施方式中,纳米颗粒-荧光肽产物在微电极上保持分离,用于落射荧光检测和分析。该实例涉及带负电的荧光电荷改变型肽和带负电的非荧光纳米颗粒,如图7所示。许多其它此类ac/dc微阵列试验形式也在本发明的范围内,另一个例子包括使用非荧光纳米颗粒,其用对电荷改变型肽产物具有特异性的抗体标记。可采用许多其他ac模式试验形式。
dc模式下进行的微阵列蛋白酶试验
在dc模式下的甚至稳健微阵列装置用于血液中基于电泳的分离的操作情况中,在侵入性电化学开始破坏该过程之前,有效工作电压被限制为<10伏dc。dc模式操作的另一个问题是在高电导(>0.5s/m)缓冲液和生物样品(血液、血浆、血清)中,带电实体如肽底物和产物的电泳吸引力大大降低。
在一些实施方式中,提供了新的方法和操作,其允许通过以dc模式操作的微阵列装置进行蛋白酶试验和电荷改变型肽产物的检测。仅通过许多实例中的一个,将具有特定带负电的荧光电荷改变型肽底物的血液样品应用于微阵列芯片装置。在适当的反应时间之后,将dc场施加到芯片,其中靠近流体输入端口的一组微电极偏置为负,并且靠近出口端口的一组微电极偏置为正。同时,含有低电导两性离子缓冲液的洗涤溶液将通过入口缓慢地递送至芯片。当低电导缓冲液在芯片上移动时,带正电的荧光电荷改变型肽产物将通过电泳被更大幅地吸引至芯片器件一侧的带负电的微电极,而所有其他组分、细胞、血浆蛋白和带负电的荧光电荷改变型肽通过电泳被吸引至微阵列的对侧。最终,洗涤液的流动将去除大部分细胞和血浆蛋白,而带正电的荧光电荷改变型肽产物仍然集中在输入端口附近的一组微电极上。此时,可以通过落射荧光检测和分析荧光肽产物。在该操作中,低电导缓冲液可包括但不限于1mm至100mm范围内的两性离子组氨酸缓冲液。许多其他dc模式分离形式和检测方案均在本发明的范围内。
实施例
实施例1:鉴定复杂样品的蛋白水解特征
制备合成肽文库,其包含氨基酸对的组合,带正电和带负电残基或沿合成肽长度的不同位置处的修饰残基的组合,以及标签如荧光染料(图1a)。在将复杂样品(例如,血液、血浆、血清)与底物文库孵育并通过凝胶电泳、毛细管电泳或dc和ac动电学技术的组合从复杂样品分离孵育的底物文库的裂解产物之后,产生底物特征。
根据迁移对肽混合物中的个体底物进行分级,并通过荧光或其他标签检测。为了剖析多类蛋白酶活性(如丝氨酸、半胱氨酸和金属肽酶)的贡献,样品在与不同活性抑制剂(如金属螯合剂edta、半胱氨酸肽酶抑制剂e-64和cao74,或弹性蛋白酶特异性氯甲基酮抑制剂)分别孵育后测试。将来自抑制的样品的裂解位点特异性数据组合以产生各个肽酶类的底物特征。产生二级合成肽文库以进一步确定复杂样品中肽酶的裂解偏好。
实施例2:通过基于活性的探针进行的体外/体内纯化
设计了包含通过间隔分子与带正电的部分连接的化学反应性基团的abp(图2a-2d)。将abp与样品一起在体外孵育或在体内注射。在abp与蛋白酶活性位点结合后,化学反应性基团与活性位点亲核残基不可逆地反应。abp的带正电的部分保持暴露在蛋白酶的活性位点之外。体外电泳分离用于分离已结合abp的蛋白酶,因为带正电的部分与复杂生物样品的大部分带负电的组分分离。在体内注射具有带正电的部分和识别标签(例如近红外或荧光染料)的abp后,可观察蛋白酶活性,例如在活检的肿瘤部位处。活检后,通过电泳分离将abp结合的蛋白酶与复杂样品分离。从复杂样品分离的abp结合的蛋白酶可用于进一步的下游应用和翻译后修饰、突变等的表征。
实施例3:通过肽进行的体内蛋白酶检测
将带负电的荧光标记的肽注射到小鼠中。在遇到肿瘤的酸性微环境时,肽的组氨酸残基变为带正电,并且肽是整体中性的,这允许肽穿透肿瘤。肽被蛋白酶裂解,其序列经设计。带正电、荧光或近红外标记的裂解产物粘附于细胞表面并允许对蛋白酶活性和定位的体内成像(图3a-3d)。
实施例4:通过肽修饰的纳米颗粒进行的体内蛋白酶检测
将修饰有带负电的肽的纳米颗粒注射到小鼠中。在遇到肿瘤的酸性微环境时,肽的组氨酸残基变为带正电,并且肽是整体中性的,这允许经修饰的纳米颗粒穿透肿瘤。纳米颗粒表面上的肽被蛋白酶裂解,其序列经设计。裂解产物带正电,这使得修饰的纳米颗粒粘附到细胞表面并将其负载的成像剂和/或药物分子的释放到肿瘤细胞中(图4a-4e)。
实施例5:利用接触角测量进行的两亲酶底物检测
开发了通过接触角测量来检测复杂样品的液滴中的蛋白酶活性的技术(图5a)。设计两亲性底物,其在感兴趣的蛋白酶的裂解位点周围含有疏水和亲水部分。将底物与复杂样品(例如血液)的液滴孵育,当发生蛋白水解裂解时,亲水和疏水产物从两亲性底物释放。疏水裂解产物优先在样品液滴表面的液滴-空气界面处组装。随着更多疏水裂解产物迁移到液滴表面,液滴的接触角减小并且测量为蛋白酶活性的指标。
实施例6:液滴表面上两亲性酶底物的检测
开发了通过在液滴表面进行荧光检测来检测复杂样品液滴中蛋白酶活性的技术(图5b)。荧光标记的两亲性底物经设计以在感兴趣的蛋白酶的裂解位点周围含有疏水和亲水部分。将底物与复杂样品(例如血液)的液滴孵育,当发生蛋白水解裂解时,亲水和疏水产物从两亲性底物释放。荧光标记的疏水裂解产物优先在样品液滴表面的液滴-空气界面处组装。液滴表面的荧光被检测为蛋白酶活性的指标,其中样品造成的猝灭最小。
实施例7:电荷改变型底物的侧流试验
开发了基于电荷改变型底物的蛋白酶检测试验的侧流试验形式(图6)。将中性或带负电的底物与复杂样品在侧流表面的中性部分上孵育。中性孵育部分与侧流表面的阳离子部分相邻,这导致样品组分和带负电的裂解产物与带正电的固体表面相关联。荧光或比色标记的带正电的裂解产物与侧流试验的液相相关并且比样品组分或带负电的裂解产物迁移得更远。迁移的带正电的裂解产物通过它们的荧光或比色标记物被检测为远离猝灭样品的条带,其指示蛋白酶活性。
实施例8:非降解性电荷转移酶的检测
开发了一种方法,通过该方法基于电荷改变型底物和复杂样品的电泳分离来测量非降解性电荷转移酶如激酶和磷酸化酶。具有磷酸化残基(如丝氨酸、苏氨酸和酪氨酸)的底物与复杂样品孵育,并设计以在样品中存在的磷酸化酶裂解磷酸基团后带正电(图7a)。然后通过施加电场将带正电的反应产物与复杂样品分离。
将含有残基(如丝氨酸、苏氨酸和酪氨酸)的带正电的底物与复杂样品一起孵育,并设计以在样品中的激酶磷酸化后带负电(图7b)。在从复杂样品电泳分离磷酸化产物后比较样品间的相对激酶活性。与具有较低激酶活性的样品相比,更高的激酶活性导致荧光减少,因为磷酸化的底物变为带负电并且在复杂样品(而非带正电的非磷酸化底物)的方向上分离。通过测量电泳凝胶中非磷酸化底物条带处荧光的减少来推断激酶活性。
实施例9:采用电荷改变型肽的凝血酶检测
不同浓度(0nm、0.5nm、5nm、10nm、20nm或100nm)的凝血酶靶向的电荷改变型肽如图9所示(并且具有序列ac-n-d--d--nle-t-p-r+//g-s-a-g-a-g-a-g-二氨基-乙基-bfl)在1xpbs(图10a)或柠檬酸化的血液(图10b)中孵育,两者都含有凝血酶。孵育后,测量荧光,指示肽与样品中凝血酶的特异性结合。荧光随着凝血酶浓度的增加而增加。
实施例10:全血凝血酶检测
将图9中所示的凝血酶靶向的电荷改变型肽(以及序列ac-n-d--d--nle-t-p-r+//g-s-a-g-a-g-a-g-二氨基-乙基-bfl)在未处理的全血中孵育长达33分钟。还制备了阴性对照,参见图11a。在孵育期间以3分钟的间隔测量荧光。指示肽与全血样品中凝血酶的特异性结合的荧光信号随时间增加,在约18分钟后观察到峰值(图11b)。
实施例11:缓冲液中的组织蛋白酶-s检测
如图9所示的组织蛋白酶-s靶向的电荷改变型肽(并且序列为n-suc--e--g-r+-w-h-t-v-g//l-r+-w-e--c(cy5-)-r+-co-nh2)在具有不同浓度的组织蛋白酶-s的1xpbs缓冲液中孵育。孵育后,测量荧光,指示肽与样品中组织蛋白酶-s的特异性结合。荧光随组织蛋白酶-s浓度的增加而增加(图12)。
实施例12:凝血监测
为了检测凝血酶活性并监测凝血,将图9中所示的凝血酶靶向的电荷改变型肽(且具有序列ac-n-d--d--nle-t-p-r+//g-s-a-g-a-g-a-g-二氨基-乙基-bfl)在来自采用抗凝血剂(阿司匹林)的对象和来自未采用抗凝血剂的对照对象的全血的液滴中孵育。在长达30分钟的不同时间点测量荧光信号,其指示肽与全血样品中凝血酶的特异性结合。图13a的上图显示对照组中的荧光,而下图显示抗凝血剂组中的荧光。两个对象组的荧光随时间增加,但在对照组中更快(图13b)。
实施例13:采用电泳聚焦试验和小型化阵列原型进行的蛋白酶(胰蛋白酶)活性测量
使用电泳聚焦试验,采用小型化阵列原型来测量胰蛋白酶活性(图15a)。将具有荧光标签的电荷改变型胰蛋白酶特异性肽与含有不同浓度的胰蛋白酶的样品孵育。利用电泳分离将胰蛋白酶键与电荷改变型肽底物分离。具有结合的底物的胰蛋白酶带正电并与带负电的复杂生物样品的大部分组分分离。随着胰蛋白酶浓度的增加,观察到荧光增加(图15b)。
使用小型阵列原型在一周后测量胰蛋白酶活性(图16a)。随着胰蛋白酶浓度的增加,观察到荧光增加(图16b)。
还使用小型阵列原型测量血液样品中的胰蛋白酶活性(图17a)。随着胰蛋白酶浓度的增加,观察到荧光增加(图17b)。
实施例14:采用dep进行蛋白酶的浓缩和检测
使用介电泳(dep)分离来检测和确定来自慢性淋巴细胞白血病对象的血浆样品中mmp2和mmp9的浓度,并与对照对象进行比较。将血浆样品与对mmp2/9具有特异性的电荷改变型肽底物孵育。将ac场施加到一些血浆样品上,并将具有结合的荧光电荷改变型肽产物的蛋白酶分离到dep高场区域中。如图19所示,与未经dep(白色条)的血浆样品相比,dep血浆样品(黑色条)中的所有对象的荧光强度更大。
其他实施方式
应理解虽然本发明已结合其详述进行描述,但以上描述意在说明而不是限制本发明的范围,该范围由所附权利要求的范围限定。其他方面、优势和修改在所附权利要求的范围内。
参考文献
蛋白水解和翻译后修饰的生物特征
barrett,a.j.(2004)merops数据库中蛋白酶的生物信息学(bioinformaticsofproteasesinthemeropsdatabase).curr.opin.drug.discov.dev.7,334–341.
cohen,p.(2002)蛋白质磷酸化的起源(theoriginsofproteinphosphorylation).nat.cellbiol.4,e127–e130.
daviet,l.和colland,f.(2008)靶向泛素特异性蛋白酶进行药物发现(targetingubiquitinspecificproteasesfordrugdiscovery).biochimie(paris)90,270–283.
geiss-friedlander,r.和melchior,f.(2007)类泛素化的概念:十年之久(conceptsinsumoylation:adecadeon).nat.rev.mol.cellbiol.8,947–956.
nandi,d.,tahiliani,p.,kumar,a.和chandu,d.(2006)泛素-蛋白酶体系统(theubiquitin-proteasomesystem).j.biosci.31,137–155.
polevoda,b.和sherman,f.(2003)n-末端乙酰转移酶和真核蛋白n-末端乙酰化的序列要求(n-terminalacetyltransferasesandsequencerequirementsforn-terminalacetylationofeukaryoticproteins).j.mol.biol.325,595–622.
puente,x.s.,sanchez,l.m.,overall,c.m.和lópez-otín,c.(2003)人和小鼠蛋白酶:比较基因组方法(humanandmouseproteases:acomparativegenomicapproach).nat.rev.genet.4,544–558.
sadoul,k.,boyault,c.,pabion,m.和khochbin,s.(2008)通过乙酰转移酶和脱乙酰酶调节蛋白质周转(regulationofproteinturnoverbyacetyltransferasesanddeacetylases).biochimie(paris)90,306–312.
walsh,g.和jefferis,r.(2006)在治疗性蛋白质的背景下的翻译后修饰(post-translationalmodificationsinthecontextoftherapeuticproteins).nat.biotechnol.24,1241–1252.
疾病与蛋白酶和翻译后酶的治疗靶向
coussens,l.m.,fingleton,b.,matrisian,l.m.(2002)基质金属蛋白酶抑制剂和癌症:试验和磨难(matrixmetalloproteinaseinhibitorsandcancer:trialsandtribulations).science295,2387–2392.
drag,m.和salvesen,g.s.(2010)基于蛋白酶的药物发现的新兴原则(emergingprinciplesinprotease-baseddrugdiscovery).nat.rev.drugdiscovery9,690–701.
gutierrez-fernandez,a.,fueyo,a.,folgueras,a.r.,garabaya,c.,pennington,c.j.,pilgrim,s.,edwards,d.r.,holliday,d.l.,jones,j.l.,span,p.n.,sweep,f.c.,puente,x.s.和lópez-otín,c.(2008)基质金属蛋白酶-8通过调节肿瘤细胞粘附和侵袭而起到转移抑制剂的作用(matrixmetalloproteinase-8functionsasametastasissuppressorthroughmodulationoftumorcelladhesionandinvasion).cancerres.68,2755–2763.
hegedus,l.,cho,h.,xie,x.和eliceiri,g.l.(2008)额外mda-mb-231乳腺癌细胞基质金属蛋白酶促进侵袭性(additionalmda-mb-231breastcancercellmatrixmetalloproteinasespromoteinvasiveness).j.cell.physiol.216,480–485.
mcquibban,g.a.,gong,j.h.,tam,e.m.,mcculloch,c.a.,clark-lewis,i.和overall,c.m.(2000)通过明胶酶a裂解单核细胞趋化蛋白-3抑制的炎症(inflammationdampenedbygelatinaseacleavageofmonocytechemoattractantprotein-3).science289,1202–1206.
mcquibban,g.a.,gong,j.h.,wong,j.p.,wallace,j.l.,clark-lewis,i.和overall,c.m.(2002)单核细胞趋化蛋白的基质金属蛋白酶加工产生具有体内抗炎特性的cc趋化因子受体拮抗剂(matrixmetalloproteinaseprocessingofmonocytechemoattractantproteinsgeneratesccchemokinereceptorantagonistswithanti-inflammatorypropertiesinvivo).blood100,1160–1167.
overall,c.m.和kleifeld,o.(2006)迈向用于癌症治疗的第三代基质金属蛋白酶抑制剂(towardsthirdgenerationmatrixmetalloproteinaseinhibitorsforcancertherapy).br.j.cancer94,941–946.
overall,c.m.和kleifeld,o.(2006)肿瘤微环境-意见:验证基质金属蛋白酶作为药物靶点和癌症治疗的抗靶标(tumourmicroenvironment-opinion:validatingmatrixmetalloproteinasesasdrugtargetsandanti-targetsforcancertherapy).nat.rev.cancer6,227–239.
stoch,s.a.和wagner,j.a.(2008)组织蛋白酶k抑制剂:骨质疏松症治疗的新靶点(cathepsinkinhibitors:anoveltargetforosteoporosistherapy).clin.pharmacol.ther.83,172–176.
tremblay,g.m.,janelle,m.f.和bourbonnais,y.(2003)中性粒细胞弹性蛋白酶抑制剂的抗炎活性(anti-inflammatoryactivityofneutrophilelastaseinhibitors).curr.opin.investig.drugs4,556–565.
turk,b.(2006)靶向蛋白酶:成功、失败和未来前景(targetingproteases:successes,failuresandfutureprospects).nat.rev.drugdiscov.5,785–799.
yang,x.,dong,y.,zhao,j.,sun,h.,deng,y.,fan,j.和yan,q.(2007)人巨噬细胞金属弹性蛋白酶(mmp-12)表达增加与子宫内膜腺癌的侵袭有关(increasedexpressionofhumanmacrophagemetalloelastase(mmp-12)isassociatedwiththeinvasionofendometrialadenocarcinoma).pathol.res.pract.203,499–505.
用于降解组学的微阵列技术
chen,g.y.,uttamchandani,m.,zhu,q.,wang,g.,yao,s.q.(2003)开发基于活性的蛋白质微阵列中酶检测策略(developingastrategyforactivity-baseddetectionofenzymesinaproteinmicroarray).chembiochem4,336–339.
eppinger,j.,funeriu,d.p.,miyake,m.,denizot,l.,miyake,j.(2004)酶微阵列:基于酶活性的亲和标记检测的抑制常数的芯片上测定(enzymemicroarrays:on-chipdeterminationofinhibitionconstantsbasedonaffinity-labeldetectionofenzymaticactivity).angew.chem.116,3894–3898.
funeriu,d.p.,eppinger,j.,denizot,l.,miyake,m.,miyake,j.(2005)使用酶微阵列对化学文库进行酶家族特异性和基于活性的筛选(enzymefamily-specificandactivity-basedscreeningofchemicallibrariesusingenzymemicroarrays).nat.biotechnol.23,622–627.
overall,c.m.,tam,e.m.,kappelhoff,r.,connor,a.,ewart,t.,morrison,c.j.,puente,x.,lópez-otín,c.和seth,a.(2004)蛋白酶降解组学:蛋白酶底物的质谱发现和clip-chip,一种所有人类蛋白酶和抑制剂的专用dna微阵列(proteasedegradomics:massspectrometrydiscoveryofproteasesubstratesandtheclip-chip,adedicateddnamicroarrayofallhumanproteasesandinhibitors).biol.chem.385,493–504.
pease,a.c.,solas,d.,sullivan,e.j.,cronin,m.t.,holmes,c.p.,fodor,s.p.(1994)光生成的寡核苷酸阵列,用于快速dna序列分析(light-generatedoligonucleotidearraysforrapiddnasequenceanalysis).proc.natl.acad.sci.usa91,5022–5026.
sieber,s.a.,mondala,t.s.,head,s.r.,cravatt,b.f.(2004)用于分析复杂蛋白质组中酶活性的微阵列平台(microarrayplatformforprofilingenzymeactivitiesincomplexproteomes).j.am.chem.soc.126,15640–15641.
uttamchandani,m.,liu,k.,panicker,r.c.,yao,s.q.(2007)基于活性的指纹识别和微阵列中半胱氨酸蛋白酶的抑制剂发现(activity-basedfingerprintingandinhibitordiscoveryofcysteineproteasesinamicroarray).chem.commun.1518–1520.
winssinger,n.,damoiseaux,r.,tully,d.c.,geierstanger,b.h.,burdick,k.,harris,j.l.(2004)pna编码的蛋白酶底物微阵列(pna-encodedproteasesubstratemicroarrays).chem.biol.11,1351–1360.
wu,h.,ge,j.,yang,p.-y.,wang,j.,uttamchandani,m.,yao,s.q.(2011)肽醛微阵列,用于细胞事件的高通量分析(apeptidealdehydemicroarrayforhigh-throughputprofilingofcellularevents).j.am.chem.soc.133,1946–1954.
用于降解组学的质谱技术
aebersold,r.和mann,m.(2003)基于质谱的蛋白质组学(massspectrometry-basedproteomics).nature422,198–207.
dean,r.a.和overall,c.m.(2007)通过itraq标记在细胞环境中蛋白质组学发现金属蛋白酶底物揭示了多种mmp-2底物降解组(proteomicsdiscoveryofmetalloproteinasesubstratesinthecellularcontextbyitraqlabelingrevealsadiversemmp-2substratedegradome).mol.cellproteomics6,611–623.
domon,b.和aebersold,r.(2006)质谱和蛋白质分析(massspectrometryandproteinanalysis).science312,212–217.
lópez-otín,c.和overall,c.m.(2002)蛋白酶降解组学:蛋白质组学的新挑战(proteasedegradomics:anewchallengeforproteomics).nat.rev.mol.cellbiol.3,509–519.
mann,m.,hendrickson,r.c.和pandey,a.(2001)通过质谱分析蛋白质和蛋白质组(analysisofproteinsandproteomesbymassspectrometry).annu.rev.biochem.70,437–473.
o’donoghue,a.j.,eroy-reveles,a.a.,knudsen,g.m.,ingram,j.,zhou,m.,statnekov,j.b.,…craik,c.s.(2012)通过多重底物谱分析全面鉴定肽酶特异性(globalidentificationofpeptidasespecificitybymultiplexsubstrateprofiling).naturemethods,9(11),1095–1100.
overall,c.m.,tam,e.m.,kappelhoff,r.,connor,a.,ewart,t.,morrison,c.j.,puente,x.,lópez-otín,c.和seth,a.(2004)蛋白酶降解组学:蛋白酶底物的质谱发现和clip-chip,一种所有人类蛋白酶和抑制剂的专用dna微阵列(proteasedegradomics:massspectrometrydiscoveryofproteasesubstratesandtheclip-chip,adedicateddnamicroarrayofallhumanproteasesandinhibitors).biol.chem.385,493–504.
schwartz,d.r.,moin,k.,yao,b.,matrisian,l.m.,coussens,l.m.,bugge,t.h.,fingleton,b.,acuff,h.b.,sinnamon,m.,nassar,h.,platts,a.e.,krawetz,s.a.,linebaugh,b.e.和sloane,b.f.(2007)hu/muprotin寡核苷酸微阵列:用于分析肿瘤及其微环境中蛋白酶和蛋白酶抑制剂基因表达的双物种阵列(hu/muprotinoligonucleotidemicroarray:dual-speciesarrayforprofilingproteaseandproteaseinhibitorgeneexpressionintumorsandtheirmicroenvironment).mol.cancerres.5,443–54.
用于降解组学的基于活性的探针和质谱分析
blum,g.,vondegenfeld,g.,merchant,m.j.,blau,h.m.和bogyo,m.(2007)使用基于荧光猝灭活性的探针的半胱氨酸蛋白酶活性的无创光学成像(noninvasiveopticalimagingofcysteineproteaseactivityusingfluorescentlyquenchedactivity-basedprobes).nat.chem.biol.3,668–677
chan,e.w.,chattopadhaya,s.,panicker,r.c.,huang,x.和yao,s.q.(2004)开发用于蛋白质组学分析的光活性亲和探针:用于金属蛋白酶的基于异羟肟酸的探针(developingphotoactiveaffinityprobesforproteomicprofiling:hydroxamate-basedprobesformetalloproteases).j.am.chem.soc.126,14435–14446.
chun,j.,yin,y.i.,yang,g.,tarassishin,l.和li,y.m.(2004)立体选择性合成光反应性拟肽γ-分泌酶抑制剂(stereoselectivesynthesisofphotoreactivepeptidomimeticγ-secretaseinhibitors).j.org.chem.69,7344–7347.
grabarek,j.和darzynkiewicz,z.(2002)通过用荧光染料标记的抑制剂亲和标记其酶活性中心,检测细胞凋亡过程中半胱天冬酶和丝氨酸蛋白酶的原位激活(insituactivationofcaspasesandserineproteasesduringapoptosisdetectedbyaffinitylabelingtheirenzymeactivecenterswithfluorochrome-taggedinhibitors).exp.hematol.30,982–989.
greenbaum,d.,medzihradszky,k.f.,burlingame,a.和bogyo,m.(2000)环氧化物亲电子作为活性依赖性半胱氨酸蛋白酶分析和发现工具(epoxideelectrophilesasactivity-dependentcysteineproteaseprofilinganddiscoverytools).chem.biol.7,569–581.
greenbaum,d.,baruch,a.,hayrapetian,l.,darula,z.,burlingame,a.,medzihradszky,k.f.和bogyo,m.(2002)用于功能性探测蛋白质组的化学方法(chemicalapproachesforfunctionallyprobingtheproteome).mol.cell.proteomics1,60–68.
kessler,b.m.,tortorella,d.,altun,m.,kisselev,a.f.,fiebiger,e.,hekking,b.g.,ploegh,h.l.和overkleeft,h.s.(2001)扩展的基于肽的抑制剂有效地靶向蛋白酶体并揭示催化β-亚基的重叠特异性(extendedpeptide-basedinhibitorsefficientlytargettheproteasomeandrevealoverlappingspecificitiesofthecatalyticbeta-subunits).chem.biol.8,913–929.
kidd,d.,liu,y.和cravatt,b.f.(2001)分析复杂蛋白质组中的丝氨酸水解酶活性(profilingserinehydrolaseactivitiesincomplexproteomes).biochemistry40,4005–4015.
liu,y.,patricelli,m.p.和cravatt,b.f.(1999)基于活性的蛋白质分析:丝氨酸水解酶(activity-basedproteinprofiling:theserinehydrolases).proc.natl.acad.sci.u.s.a.96,14694–14699.
saghatelian,a.,jessani,n.,joseph,a.,humphrey,m.和cravatt,b.f.(2004)用于金属蛋白酶蛋白质组学分析的基于活性的探针(activity-basedprobesfortheproteomicprofilingofmetalloproteases).proc.natl.acad.sci.u.s.a.101,10000–10005.
schmidinger,h.,hermetter,a.和birner-gruenberger,r.(2006)基于活性的蛋白质组学:复杂蛋白质组中的酶活性谱(activity-basedproteomics:enzymaticactivityprofilingincomplexproteomes).aminoacids30,333–350.
thornberry,n.a.,peterson,e.p.,zhao,j.j.,howard,a.d.,griffin,p.r.和chapman,k.t.(1994)肽(酰氧基)甲基酮对白细胞介素-1β转化酶的灭活作用(inactivationofinterleukin-1βconvertingenzymebypeptide(acyloxy)methylketones).biochemistry33,3934–3940.
wang,c.c.,bozdech,z.,liu,c.l.,shipway,a.,backes,b.j.,harris,j.l.和bogyo,m.(2003)布氏锥虫20s蛋白酶体的生化分析(biochemicalanalysisofthe20sproteasomeoftrypanosomabrucei).j.biol.chem.278,15800–15808.
williams,e.b.,krishnaswamy,s.和mann,k.g.(1989)使用肽氯甲基酮的酶原/酶鉴别(zymogen/enzymediscriminationusingpeptidechloromethylketones).j.biol.chem.264,7536–7545.
区分酶活性与非活性形式的挑战
moldoveanu,t.,hosfield,c.m.,lim,d.,elce,j.s.,jia,z.c.,davies,p.l.(2002)ca(2+)开关对准钙蛋白酶的活性位点(aca(2+)switchalignstheactivesiteofcalpain).cell108,649–660.
turk,v.,turk,b.,turk,d.(2001)溶酶体半胱氨酸蛋白酶:事实和机会(lysosomalcysteineproteases:factsandopportunities).emboj.20,4629–4633.
酶谱和其他反向降解组学技术
baragi,v.m.等.(2000)使用琥珀酰化明胶的明胶酶的通用测定法(aversatileassayforgelatinasesusingsuccinylatedgelatin).matrixbiol.19,267–273.
kaberdin,v.r.andmcdowall,k.j.(2003)通过将确定的小底物与凝胶基质化学连接,扩大酶谱的使用(expandingtheuseofzymographybythechemicallinkageofsmall,definedsubstratestothegelmatrix).genomeres.13,1961–1965.
kleiner,d.e.和stetler-stevenson,w.g.(1994)定量酶谱法:检测皮克量的明胶酶(quantitativezymography:detectionofpicogramquantitiesofgelatinases).anal.biochem.218,325–329.
kupai,k.等.(2010)基质金属蛋白酶活性测定:酶谱的重要性(matrixmetalloproteinaseactivityassays:importanceofzymography).j.pharmacol.toxicol.methods61,205–209.
mckerrow,j.h.,pino-heiss,s.,lindquist,r.和werb,z.(1985)曼氏血吸虫尾蚴分泌的弹性蛋白酶的纯化和鉴定(purificationandcharacterizationofanelastinolyticproteinasesecretedbycercariaeofschistosomamansoni).j.biol.chem.260,3703–3707.
paemen,l.等.(1996)四环素和化学修饰的四环素类似物的明胶酶抑制活性,通过新型微量滴定法测定抑制剂(thegelatinaseinhibitoryactivityoftetracyclinesandchemicallymodifiedtetracyclineanaloguesasmeasuredbyanovelmicrotiterassayforinhibitors).biochem.pharmacol.52,105–111.
piccard,h.等.(2009)“反向降解组学”,通过多ce和荧光底物和反应产物的共聚焦检测监测蛋白水解修剪(“reversedegradomics”,monitoringofproteolytictrimmingbymulti-ceandconfocaldetectionoffluorescentsubstratesandreactionproducts).electrophoresis30,2366–2377.
vandooren,j.等.(2011)明胶降解测定揭示了mmp-9抑制剂和o-糖基化结构域的功能(gelatindegradationassayrevealsmmp-9inhibitorsandfunctionofo-glycosylateddomain).worldj.biol.chem.2,14–24.
wilkesman,j.和kurz,l.(2009)通过酶谱法进行蛋白酶分析:对技术和专利的评论(proteaseanalysisbyzymography:areviewontechniquesandpatents).recentpat.biotechnol.3,175–184.
酶活性的体内成像
bremer,c.,tung,c.h.和weissleder,r.体内分子靶点评估基质金属蛋白酶的抑制作用(invivomoleculartargetassessmentofmatrixmetalloproteinaseinhibition).nat.med.7,743–748(2001).
crawford,b.d.和pilgrim,d.b.通过体外和体内酶谱法研究斑马鱼胚胎中基质金属蛋白酶活性的发生和调节(ontogenyandregulationofmatrixmetalloproteinaseactivityinthezebrafishembryobyinvitroandinvivozymography).dev.biol.286,405–414(2005).
kaijzel,e.l.,vanderpluijm,g.和lowik,c.w.用于评估癌症发展和进展的动物模型中的全身光学成像(whole-bodyopticalimaginginanimalmodelstoassesscancerdevelopmentandprogression).clin.cancerres.13,3490–3497(2007).
keow,j.y.,pond,e.d.,cisar,j.s.,cravatt,b.f.和crawford,b.d.活体脊椎动物胚胎中基质金属蛋白酶的基于活性的标记(activity-basedlabelingofmatrixmetalloproteinasesinlivingvertebrateembryos).plosone7,e43434(2012).
saghatelian,a.,jessani,n.,joseph,a.,humphrey,m.和cravatt,b.f.用于金属蛋白酶蛋白质组学分析的基于活性的探针(activity-basedprobesfortheproteomicprofilingofmetalloproteases).proc.natl.acad.sci.usa101,10000–10005(2004).
scherer,r.l.,vansaun,m.n.,mcintyre,j.o.和matrisian,l.m.使用蛋白水解纳米信标在体内对基质金属蛋白酶-7活性进行光学成像(opticalimagingofmatrixmetalloproteinase-7activityinvivousingaproteolyticnanobeacon).mol.imaging7,118–131(2008).
weissleder,r.,tung,c.h.,mahmood,u.和bogdanov,a.jr.用蛋白酶激活的近红外荧光探针对肿瘤进行体内成像(invivoimagingoftumorswithprotease-activatednear-infraredfluorescentprobes).nat.biotechnol.17,375–378(1999).
wyatt,r.a.等.斑马鱼胚胎:一种用于研究基质重塑的强大模型系统(thezebrafishembryo:apowerfulmodelsystemforinvestigatingmatrixremodeling).zebrafish6,347–354(2009).
- 还没有人留言评论。精彩留言会获得点赞!