一种用于预防、延缓胰腺癌及癌前病变的代餐组合物及其应用的制作方法
本发明涉及食品领域,尤其涉及一种用于预防、延缓胰腺癌及癌前病变的代餐组合物及其应用。
背景技术:
胰腺腺管内上皮瘤(pancreaticintraepithelialneoplasia,panin)是近几年提出的新术语,目前对其缺乏足够认识。
代谢重编程是由ottowarburg首先提出的肿瘤代谢特征之一,主要指肿瘤细胞即使在氧含量充足时也偏好于糖酵解。随着对肿瘤代谢的不断研究,warburg效应的概念也不断扩充。除了糖酵解,在其他代谢途径如脂肪酸代谢,氨基酸代谢和一碳单位循环等(hanahanandweinberg,2011)也不断地涵盖到广义的warburg效应。
支链氨基酸(bcaa)是必需氨基酸,包括亮氨酸、异亮氨酸和缬氨酸。支链氨基酸转氨酶(bcat)和支链氨基酸酮酸脱氢酶复合物(bckdc)是bcaa分解代谢的两个关键酶(shimomuraetal.,2001)。bcat参与催化bcaa的第一步反应,主要有两种亚型,一种是主要位于细胞胞浆中的bcat1或bcatc,另一个是主要位于线粒体中的bcat2或bcatm。尽管它们在细胞内分布不同,但均催化相同的化学反应,即将支链氨基酸上的氨基转给阿尔法-酮戊二酸(α-kg),产生对应的支链酮酸(bcka)和谷氨酸,该催化过程中以磷酸吡哆醛(plp)为辅酶(ichiharaandkoyama,1966;taylorandjenkins,1966)。进一步bcka在bckdc等一系列相关酶的催化下最终生成乙酰辅酶a(ace-coa)和琥珀酰辅酶a(suc-coa),进入tca循环。
约有90%的胰腺癌存在kras突变(halbrookandlyssiotis,2017)。在kras突变的pdac动物模型中,血浆中bcaa浓度在pdac早期已显著升高,这提示bcaa代谢可能与pdac的发生发展相关联(mayersetal.,2014)。有意思的是,研究表明在kras突变引起的癌变模型中,非小细胞肺癌的发展是bcat依赖的,而胰腺癌的发展是bcat非依赖的(mayersetal.,2016)。而最新的研究表明,在苹果酸酶缺失的pdac中,过表达bcat2显著促进肿瘤细胞的增殖(deyetal.,2017)。显而易见,这些相互矛盾的报道,表明仍需深入的研究以阐明bcaa与pdac发生发展之间的复杂关系。
综上所述,现有研究并没有阐明bcaa与胰腺癌的关系,也没有公开通过bcaa预防胰腺癌的有效措施。
技术实现要素:
针对上述技术问题,本发明提供了一种用于预防、延缓胰腺癌及癌前病变的代餐组合物及其应用,
用于预防、延缓胰腺癌及癌前病变的代餐组合物,包括蛋白质、碳水化合物和脂类;bcaa重量百分比少于1%。
优选地,蛋白质、碳水化合物和脂类按照重量百分比分别为10%-20%、50%-70%、5%-10%。
优选地,代餐组合物还包括维生素和盐;维生素和盐重量百分比分别为0.5%-1.5%、1%-10%。
优选地,所述bcaa重量百分比为0.1%-0.6%。
优选地,所述bcaa包括异亮氨酸、亮氨酸和缬氨酸中的一种或多种。
优选地,所述异亮氨酸、亮氨酸和缬氨酸比例为(0-2):(0-1):(0-1);进一步优选地,所述异亮氨酸、亮氨酸和缬氨酸比例为(0-1.5):(0-1):(0-1)。
优选地,所述蛋白质还包括丙氨酸、左旋精氨酸、胱氨酸、甘氨酸、组氨酸、苏氨酸、左旋色氨酸、苯丙氨酸、甲硫氨酸、赖氨酸中的一种或多种。
本发明的代餐组合物在制备用于预防、延缓胰腺癌及癌前病变的代餐食品中的应用。
优选地,所述的胰腺癌前病变为胰腺上皮内瘤变(panin)。
优选地,所述的胰腺癌为胰腺导管腺癌(pdac)。
与现有技术相比,本发明的技术方案具有以下优点:本发明首次公开了低bcaa饮食可以预防、延缓panin。本发明还进一步公开了能够预防、延缓panin、pdac及癌前病变的低bcaa代餐组合物。
附图说明
为了更清楚地说明本发明实施例或现有技术中的技术方案,下面将对实施例或现有技术描述中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图仅仅是本发明的一些实施例,对于本领域普通技术人员来讲,在不付出创造性劳动性的前提下,还可以根据这些附图获得其他的附图。
图1是本发明实施例4的4组小鼠胰腺的h&e染色结果示意图;
图2是本发明实施例4的4组小鼠panin1、2和3期分别占导管总含量的比例;
图3是本发明实施例4的4组小鼠胰腺重量结果;
图4是本发明实施例4的4组小鼠ck19,bcat2和ki67表达水平;
图5是本发明实施例4的4组小鼠血浆中支链氨基酸的浓度;
图6是本发明实施例4的4组小鼠食量结果;
图7是本发明实施例5的7个样本bcat2的westernblot结果;
图8是本发明实施例5的7个样本细胞消耗培养基中bcaa含量培养基中bcaa浓度;
图9是本发明实施例5的敲减bcat2对细胞产酸速率和耗氧率的影响;
图10是本发明实施例5的bcat2抑制剂对耗氧率的影响;
图11是本发明实施例5的bcat2抑制剂对细胞内nadh/nad+的比值,及乙酰辅酶a(ace-coa)和琥珀酰辅酶a的水平的影响;
图12是本发明实施例5的胰腺细胞中bcat1蛋白水平;
图13是本发明实施例5的bxpc3细胞bcat2的敲减效率;
图14是本发明实施例5的sw1990细胞bcat2的敲减效率;
图15是本发明实施例6的三株细胞系中过表达kras活化突变和三株细胞系中敲减kras的结果;
图16是本发明实施例6的sw1990、panc1和aspc1细胞中敲减kras对bcat2和trim21相互作用的影响;
图17是本发明实施例6的处理后bcat2含量变化;
图18是本发明实施例6的krasg12v对bcaa的代谢的影响;
图19是本发明实施例6的将bcat2-flag与ha-ub的质粒共转染hek293t细胞中的结果;
图20是本发明实施例6的在mg132处理的、敲减trim1的hek293t细胞中共转染bcat2-flag与ha-ub质粒的结果;
图21是本发明实施例6的敲减trim1的细胞内bcat2的蛋白水平;
图22是本发明实施例6的bxpc3细胞中过表达kras活化突变对bcat2和trim21相互作用的影响;
图23是本发明实施例7的过表达bcat2和敲减bcat2对细胞消耗培养基中bcaas的含量的影响;
图24是本发明实施例7的细胞生长曲线;
图25是本发明实施例7的细胞的克隆形成能力检测;
图26是本发明实施例7的对照组和kc组小鼠胰腺组织bcat2的免疫组织化学染色镜下图;;
图27是本发明实施例7的kc小鼠胰腺组织连续切片中ck19、bcat2和ki67的免疫组织化学染色镜下图;
图28是本发明实施例7的wb鉴定bcat2在在h6c7和hpne细胞中过表达bcat2-flag质粒后bcat2的蛋白表达水平;
图29是本发明实施例7的bcat2抑制剂处理对细胞消耗培养基中bcaas的含量的影响;
图30是本发明实施例7的h6c7和hpne细胞分别过表达bcat2的细胞克隆形成结果镜下图;
图31是本发明实施例7的h6c7和hpne细胞分别过表达bcat2对细胞迁移影响的检测;
图32是本发明实施例7的在sw1990和bxpc3细胞敲减bcat2的细胞克隆形成结果镜下图;
图33是本发明实施例7的在sw1990和bxpc3细胞敲减bcat2的细胞迁移结果检测。
具体实施方式
以下将结合附图对本发明的构思、具体结构及产生的技术效果作进一步说明,以充分地了解本发明的目的、特征和效果。
实施例1
按照以下配方制备本发明的用于预防、延缓胰腺癌及癌前病变的代餐组合物。
实施例2
按照以下配方制备本发明的用于预防、延缓胰腺癌及癌前病变的代餐组合物。
实施例3
按照以下配方制备本发明的用于预防、延缓胰腺癌及癌前病变的代餐组合物。
实施例4
1、制备小鼠模型
制备了条件性敲除bcat2的转基因小鼠模型,并通过不同转基因小鼠杂交分别获得了pdx1-cre;lsl-krasg12d(简称kc),pdx1-cre;bcat2flox/flox(简称cb)和lsl-krasg12d;bcat2flox/folx;pdx1-cre(简称kcb)的小鼠模型。kc小鼠模型能够很好的模拟胰腺癌前病变panin期的发生发展的进程及其病理表现(hingoranietal.,2003),而kcb小鼠模型能观察bcat2对panin发生发展的影响。
2、将20只1月龄正常小鼠和kc小鼠分为四组,分别为:
a组:7只正常小鼠;
b组:7只正常小鼠;
c组:7只kc小鼠;
d组:7只kc小鼠。
3、将a组和c组小鼠给予正常饲料饮食,将b组和d组给予实施例3的代餐组合物饮食,持续5个月。
4、将四组小鼠解剖,取出胰腺,用h&e染色,并置于比例尺为50μm镜下观察。
5、将四组h&e染色的小鼠的胰腺中panin各时期定量分析,分别检测4组小鼠panin1、2和3期分别占导管总含量的比例。
6、将取出的胰腺进行称重并分析。
7、用免疫组织化学染色鉴定ck19,bcat2和ki67在小鼠胰腺中的表达水平。比例尺为50μm。
7、检测4组小鼠血浆中支链氨基酸的浓度。
8、分析记录的4组小鼠食量。
上述实验结果具体分析如下:
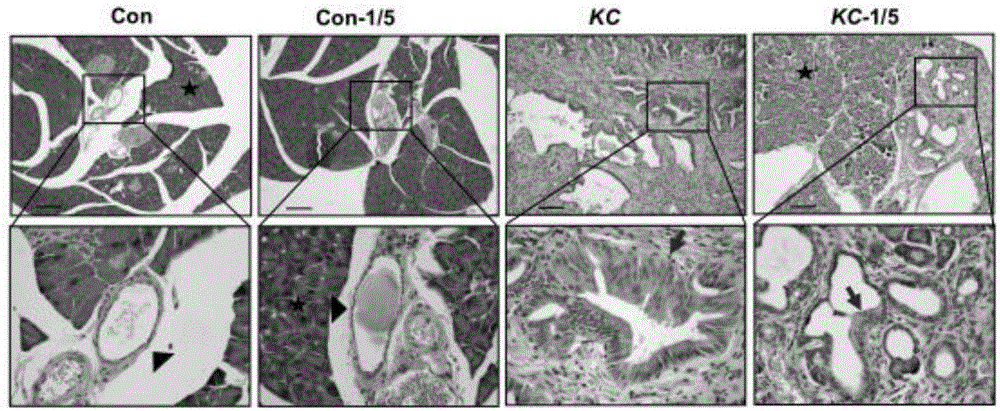
图1为小鼠胰腺的h&e染色结果示意图,结果显示:实施例3的代餐组合物显著减弱kc小鼠胰腺panin进展。
图2为小鼠panin1、2和3期分别占导管总含量的比例,结果显示:在正常饮食的kc小鼠中,panin1,2,和3期分别占导管总含量的54%,18%和5%。而实施例3的代餐组合物的kc小鼠中,panin1,2,和3期分别占导管总含量的23%,3%和0%。
图3为小鼠胰腺重量分析,结果显示:实施例3的代餐组合物显著减弱kc小鼠病变胰腺的重量;kc小鼠胰腺重量是正常小鼠胰腺重量的3倍。
图4为小鼠ck19,bcat2和ki67表达水平,结果显示:箭头分别指示ck19、bcat2和ki67染色。实施例3的代餐组合物显著减弱kc小鼠panin导管中细胞的增殖。
图5为小鼠血浆中支链氨基酸的浓度,结果显示:实施例3的代餐组合物显著下调血浆中bcaa浓度而对小鼠摄食量无影响。
图6是4组小鼠食量分析,结果显示:实验过程中动物饮食量并无显著变化通过上述多项实验证明了本发明实施例3的代餐组合物可用于预防、延缓panin,成为预防和临床早期pdac治疗的方案。本发明的代餐组合物为低bcaa饮食,以上结果表明,bcaa代谢在pdac早期发生中起着重要作用。
以下详细介绍本发明提供的低bcaa饮食产品预防、延缓panin的原理。
实施例5胰腺癌中bcat2促进bcaa代谢
为研究支链氨基酸代谢与胰腺癌的关系,我们重点关注了参与支链氨基酸分解代谢的第一个关键酶——支链氨基酸转氨酶(bcat)。它包括两种亚型,位于胞浆中的bcat1和位于线粒体的bcat2。通过以下大量实验分析支链氨基酸代谢与胰腺癌的关系。
1、在永生化的正常胰腺导管上皮细胞和胰腺导管癌细胞中分别检测了bcat1和bcat2的蛋白水平。选取2株永生化的正常胰腺导管上皮细胞htert-hpne及hpde6c7(以下简称hpne和h6c7),和5株胰腺导管癌细胞作为研究对象。利用westernblot分别检测胰腺细胞中bcat2的蛋白水平。hpne和h6c7为永生化的正常胰腺导管上皮细胞,panc1、bxpc3、aspc1、capan1和sw1990为pdac细胞株,以β-actin为内参进行定量。westernbolt的数据结果如图7和12所示:胰腺癌细胞中bcat2的蛋白水平较永生化的正常细胞有显著上调(如图7所示);而bcat1蛋白仅在panc1细胞中上调,在其他胰腺导管癌细胞中未发生显著变化(如图12所示)。
2、进一步,为验证胰腺导管细胞中bcat2上调是否促进bcaa的代谢。将细胞培养24h后,检测各细胞消耗培养基中bcaas的量,发现胰腺导管癌细胞消耗bcaa的量是正常胰腺导管细胞hpne的1.5-2.5倍,与bcat2的表达呈正相关,pdac消耗bcaas能力增强(如图8所示)。
3、为研究bcat2对bcaa代谢流的调控,首先用seahorse技术分别检测了bcat2对细胞糖酵解和有氧呼吸的影响,其中细胞产酸速率(ecar)代表细胞糖酵解能力,耗氧率(ocr)代表细胞有氧呼吸能力。敲减bcat2引起细胞ecar升高和ocr降低。如图9上图所示,对照或敲减bcat2的bxpc3细胞中,在指定的时间点依次加入葡萄糖(glucose),寡霉素(oligomycin)和2-脱氧-d葡萄糖(2-dg)。绘制ecar曲线;如图9下图所示,对照或敲减bcat2的bxpc3细胞中,在指定的时间点依次加入寡霉素(oligomycin),三氟氰苯腙(trifluoromethoxycarbonylcyanidephenylhydrazone,fccp)和鱼藤酮(rotenone)。绘制ocr曲线。*p<0.05;**p<0.01;***p<0.001。bcat2抑制剂下调细胞ocr水平。bcat2抑制剂处理bxpc3细胞,在指定的时间点依次加入寡霉素(oligomycin),三氟氰苯腙(trifluoromethoxycarbonylcyanidephenylhydrazone,fccp)和鱼藤酮(rotenone)。绘制ocr曲线。*p<0.05;**p<0.01。结果显示:在bxpc3细胞中,敲减bcat2能够明显增强细胞的ecar水平,同时减弱ocr水平(如图9和图13所示)。与之一致的是bcat2抑制剂处理bxpc3细胞后,细胞中ocr水平也显著下调(如图10所示)
4、检测了细胞内nadh/nad+的比值,及bcaa下游代谢产物乙酰辅酶a(ace-coa)和琥珀酰辅酶a(suc-coa)的水平。结果表明,在sw1990细胞中敲减bcat2后,nadh/nad+的比值明显上调,而细胞内ace-coa和suc-coa浓度没有明显变化(图11和图14)。如图11所示,敲减bcat2上调细胞内nadh/nad的比值但不影响ace-coa和suc-coa。对照或敲减bcat2的sw1990细胞中,试剂盒检测nadh/nad+的比值,lc-ms检测乙酰辅酶a(ace-coa)和琥珀酰辅酶a(suc-coa)。**p<0.01;n.s.代表无显著差异。
实施例6kras突变稳定bcat2蛋白促进bcaa代谢
约有90%的胰腺导管癌存在kras突变(kandaetal.,2012)。因此,我们进一步研究了kras突变是否调控bcat2的蛋白水平。
1、选取三株kras野生型的细胞系,即h6c7、hpne胰腺导管上皮细胞和bxpc3细胞,并在这三株细胞系中过表达kras活化突变体(krasg12v),发现kras突变上调bcat2蛋白水平,而对bcat2的mrna水平无影响。h6c7,hpne和bxpc3细胞中分别过表达kras活化突变体krasg12v,sw1990,panc1和aspc1细胞分别敲减kras,bcat2和kras的蛋白水平通过wb检测。β-actin作为内进行定量。q-pcr检测各细胞系中bcat2的mrna表达水平。n.s.代表无显著差异。krasg12v分别上调bcat2的蛋白水平为1.9,1.7和1.2倍,而bcat2的mrna水平并无明显变化(图15,左图)。相应的,敲减sw1990、panc1和aspc1细胞中kras,bcat2的蛋白水平明显下调,而bcat2的mrna水平并无明显变化(如图15,右图)。以上结果提示kras可能通过翻译后修饰调控bcat2的蛋白水平。
2、将bcat2-flag与ha-ub的质粒共转染hek293t细胞中,发现bcat2受到显著的泛素化修饰(如图19)。
3、泛素化的可逆调控主要由4种酶组成,e1,e2,e3泛素连接酶和去泛素化酶(dub)(leestemakerandovaa,2017)。其中,e3泛素连接酶是泛素化介导降解途径的底物特异性酶(hershkoandciechanover,1998)。通过串联亲和纯化和质谱技术,即tap-ms,鉴定结果显示三基序蛋白21(trim21)获得高得分。提示trim21可能是调节bcat2蛋白降解的e3连接酶。将trim21敲减后,显著下调bcat2的泛素化水平(如图20)。此外,在h6c7、sw1990、aspc1和panc1细胞中分别将内源性trim21敲减后,细胞内bcat2的蛋白水平均出现不同程度的显著上升(如图21)。这些结果表明,trim21是bcat2的e3泛素连接酶。
4、为进一步阐明krasg12v对bcat2蛋白水平调控的分子机制。如图16所示,在sw1990、panc1和aspc1细胞中,敲减kras可以显著上调二者的相互作用,敲减kras增强bcat2与trim21的相互作用。在sw1990、panc1和aspc1细胞中分别敲减kras,内源ip和wb检测bcat2和trim21的相互作用。bcat2作为ip内参进行定量。相应的,如图22所示,在bxpc3细胞中过表达krasg12v,发现krasg12v能明显阻断trim21和bcat2的结合,在bxpc3细胞中过表达krasg12v阻断bcat2与trim21的相互作用。过表达krasg12v的bxpc3细胞结合mg132处理6h,内源ip和wb检测bcat2和trim21的相互作用。trim21作为ip内参进行定量。如图17所示,过表达krasg12v显著延长bcat2的半衰期。(h6c7细胞过表达krasg12v,chx处理不同时间,wb检测bcat2的蛋白量(如图17,左图)和定量(如图17,右图)。*p<0.05)。
这些数据表明,kras突变能够阻断bcat2和e3泛素连接酶trim21的结合,降低bcat2的泛素化水平,稳定bcat2的蛋白水平。
5、通过gc-ms进一步检测了krasg12v对bcaa的代谢的影响,发现过表达krasg12v可以上调h6c7和bxpc3细胞内bcaa和α-酮异己酸(isovaleryl-coa,kic)水平;反之,敲减kras引起panc1细胞内bcaa及kic的水平下降(图18)。kras突变促进bcaas分解代谢。如图18、左图所示,krasg12v促进bcaas分解代谢。在h6c7细胞中过表达krasg12v,用lc-ms检测bcaas相关代谢物的含量;如图18、右图所示,kras敲减抑制bcaas分解代谢,在panc1细胞中敲减kras,lc-ms检测bcaas相关代谢物的含量。*p<0.05;**p<0.01;***p<0.001。
以上研究结果表明kras突变促进bcaa的分解代谢,而bcat2是bcaa分解代谢中的关键酶。
实施例7bcat2促进胰腺导管癌细胞的增殖
1、检测bcat2对胰腺导管上皮细胞和pdac细胞吸收bcaa的影响。结果如图23和28所示,稳定性过表达bcat2明显增加h6c7和hpne细胞吸收bcaa的能力;而稳定性敲减bcat2造成sw1990和bxpc3细胞吸收bcaa的能力明显减弱。其中,图23为bcat2促进细胞对bcaas的吸收。过表达bcat2的h6c7和hpne细胞中,检细胞消耗培养基中bcaas的含量(图23,左图);敲减bcat2的sw1990和bxpc3细胞中,检细胞消耗培养基中bcaas的含量(图23,右图)。*p<0.05。图28为过表达bcat2的稳定细胞株鉴定。wb鉴定bcat2在h6c7(图28,左图)和hpne(图28,右图)细胞中的稳定性表达。此外,随着bcat2抑制剂处理时间延长和处理浓度的增加,细胞消耗bcaa的能力也逐渐减弱(如图29所示)。图29为bcat2抑制剂下调细胞对bcaas的吸收。不同浓度的bcat2抑制剂处理bxpc3细胞24h(图29,左图),48h(图29,右图),检测细胞消耗培养基中bcaas的含量。*p<0.05;**p<0.01;***p<0.001。
2、计数不同时间点的细胞数绘制生长曲线检测bcat2对胰腺导管上皮细胞和pdac细胞增殖的影响。结果如图24所示,在永生化的正常胰腺细胞中过表达bcat2显著促进h6c7和hpne的细胞增殖。同时过表达bcat2显著促进上述细胞的克隆形成能力和细胞迁移能力(图25,左图;图30-31)。反之,在癌变细胞中敲减bcat2显著抑制sw1990和bxpc3细胞的增殖(图24,下图),并且减弱细胞的克隆形成能力和细胞迁移能力(图25,右图;图32-33)。其中,如图24所示,bcat2促进pdac细胞生长。h6c7和hpne细胞分别稳定过表达bcat2(图24,上图),sw1990和bxpc3细胞中分别稳定敲减bcat2(图24,下图),在不同时间点计数细胞,绘制生长曲线。*p<0.05;**p<0.01;***p<0.001。如图25所示,bcat2促进胰腺导管上皮细胞克隆形成。在h6c7和hpne细胞分别过表达bcat2促进细胞克隆形成(图25,左图),在sw1990和bxpc3细胞中分别稳定敲减bcat2抑制pdac细胞克隆形成(图25,右图)。*p<0.05;**p<0.01。图30为过表达bcat2促进胰腺导管上皮细胞克隆形成。h6c7和hpne细胞分别稳定过表达bcat2,两周后显微镜下拍照记录克隆形成。图31为bcat2促进细胞迁移。h6c7和hpne细胞分别稳定过表达bcat2,transwell实验检测细胞迁移(图31,左图)并定量(图31,右图)。比例尺为200μm。*p<0.05;**p<0.01。图32为敲减bcat2抑制pdac细胞克隆形成。sw1990和bxpc3细胞中分别稳定敲减bcat2,两周后显微镜下拍照记录克隆形成(图32,左)并定量(图32,右)。比例尺为200μm。**p<0.01。图33为敲减bcat2抑制pdac细胞迁移。sw1990和bxpc3细胞中分别稳定敲减bcat2,transwell穿膜实验检测细胞迁移(图33,左)并定量(图33,右)。比例尺为200μm。*p<0.05;**p<0.01。
以上数据表明,bcat2促进胰腺导管上皮细胞和pdac细胞对bcaa的吸收,并调控胰腺导管上皮细胞和pdac细胞的增殖和迁移。
3、为深入研究bcat2在体内生理和病理条件下的功能,收集了正常小鼠和kc小鼠以及kcb胰腺组织标本,通过免疫组织化学染色(immunohistochemistry,ihc)的方法,发现bcat2在正常小鼠(包括wt,pdx1-cre和lsl-krasg12d)和kc小鼠的胰腺腺泡细胞中均有一定表达,但在正常对照组中的胰腺导管上皮细胞中无表达(图26)。然而,在kc组panin期的导管上皮细胞中bcat2呈现高表达,甚至高于胰腺细胞的表达(图26)。进一步选取同一组织块的连续切片,用ck19(导管标志分子),ki67(细胞增殖标志分子)和bcat2抗体,对kc组小鼠的动物胰腺组织分别进行ihc染色。我们发现ck19,ki67和bcat2的表达在发生panin病变的胰腺导管组织内共定位(图27)。如图26所示,bcat2在kc动物模型的panin期的胰腺导管组织内呈现高表达。用免疫组织化学染色(ihc)鉴定bcat2在小鼠胰腺中的表达水平(16周龄,6只)。三角指示为正常导管组织,五星指示为腺泡组织,箭头指示为panin期导管。比例尺为50μm。如图27所示,bcat2与ki67在panin期胰腺导管组织内共定位。用免疫组织化学染色(ihc)鉴定ck19,bcat2和ki67在小鼠胰腺中的表达水平(16周龄,5只)。箭头分别指示ck19、bcat2和ki67染色。比例尺为12.5μm。
以上数据显示bcat2在胰腺癌发生的早期panin期的导管上皮细胞中高表达,提示bcat2在panin发生发展过程中发挥重要作用。
以上详细描述了本发明的较佳具体实施例。应当理解,本领域的普通技术人员无需创造性劳动就可以根据本发明的构思作出诸多修改和变化。因此,凡本技术领域中技术人员依本发明的构思在现有技术的基础上通过逻辑分析、推理或者有限的实验可以得到的技术方案,皆应在由权利要求书所确定的保护范围内。
- 还没有人留言评论。精彩留言会获得点赞!