用于经由定向分化将前体细胞转化为胃组织的方法和系统与流程
本发明是在由国立卫生研究院授予的DK080823,DK092456和GM063483下的政府支持下进行的。政府对本发明有一定的权利。
优先权要求
本申请要求于2014年5月28日提交的题为“Methods and Systems for Converting Precursor Cells into Gastric Tissues through Directed Differentiation”的Wells等人的美国临时专利申请No.62/003,719的优先权和权益用于所有目的。
技术领域
本文中公开了涉及通过定向分化将干细胞转化成特定组织或器官的方法和系统。特别地,公开了用于从人多能干细胞促进定形内胚层形成的方法和系统。还公开了用于从分化的定形内胚层促进胃类器官或组织形成的方法和系统。
发明背景
胃功能和构造在哺乳动物物种之间广泛变化,以适应极其多种栖息地和饮食。因此,非人类胃发育和疾病模型具有相当大的局限性。例如,细菌幽门螺杆菌(Helicobacter Pylori)感染世界人口的50%,其中10%发展为消化性溃疡病,1-2%1-3发展为胃癌。胃疾病,包括消化性溃疡病和胃癌,影响世界人口的10%,并且主要是由于慢性幽门螺杆菌感染。幽门螺杆菌诱导的疾病的当前模型依赖于不表现出与人对感染的响应相同的病理生理特征的动物模型4,并且胃细胞系缺乏体内胃上皮的细胞和构造复杂性。因此,没有足够的模型来研究幽门螺杆菌感染的作用,就像它发生在人类中一样。虽然最近使用成体胃干细胞的进展允许体外啮齿类胃上皮的生长5,但是从人患者获得这些细胞将需要手术。此外,这种方法不能用于对人胃的胚胎发育或基质-上皮相互作用建模。胚胎发育和成体胃的构造的物种差异使鼠模型对于器官发生和该器官的发病机理的研究变得不是最佳的。因此,需要强大的体外系统来阐明人类胃发育和疾病的根本机制,以及鉴定用于这些疾病的人类治疗的新型治疗。
本领域中需要用于精确控制前体细胞,如人多能干细胞的目的地的方法和系统,以便创建期望的特定类型的组织或生物体,特别是胃组织,其可以用于一个或多个上述目的。
发明概述
公开了诱导胃细胞和/或胃组织形成的方法,如以胃类器官的形式。可以通过激活和/或抑制前体细胞内的一种或多种信号传导途径来进行胃细胞和/或组织的形成。还公开了使用所公开的胃细胞,胃组织和/或源自前体细胞的胃类器官的方法。
附图简述
本领域技术人员将理解,下面描述的附图仅用于说明的目的。附图不旨在以任何方式限制本教导的范围。
专利或申请文件包含至少一个彩色附图。根据请求并支付必要的费用后,由官方提供具有彩色附图的本专利或专利申请公开文本的副本。
图1描绘了Sox2/Cdx2/B-连环蛋白在胃球体中的表达和RA的作用。
图2A-2E描述了用于定向将hPSC分化为三维胃类器官的体外培养系统的示意图(图2A),通过用Sox2,Pdx1和Cdx2对小鼠E10.5胚胎的整装免疫荧光染色的发育中的后部前肠器官的限定标志物(图2B),在存在和不存在RA的情况下的PDX1表达(图2C),显示后部前肠球体生长成hGO期间的形态学变化的立体显微照片(图2D),在E14.5和E18.5和hGO发育的相当阶段时的发育中的小鼠窦的比较(图2E)。
图3A-3D描述了P12窦,E18.5窦和d34类器官中的Muc5AC,TFF2,GSII UEAI和CHGA表达(图3A),hGO发育期间在生长,形态发生和细胞类型特化中EGF的不同作用的示意图(图3B),在具有和没有DOX的胃类器官中胃泌素,生长素释放肽,5-HT和ChrA的表达(图3C),以及在多种浓度的EGF下的NEUROG3的相对表达(图3D)。
图4A-4D描绘了在d34类器官,E18.5窦和P12窦中的SOX9Ki67表达(图4A),使用亮视野显微术和免疫荧光染色显现的类器官的幽门螺杆菌感染(图4B),癌基因c-Met的免疫沉淀(图4C),和通过EdU掺入测量的hGO上皮中的细胞增殖(图4D)。
图5A-5D描绘了在存在GSK3β抑制剂CHIR99021的情况下的Sox2和Cdx2表达和在存在和不存在头蛋白的情况下的重组WNT3A(图5A),使用明视野显微术显现的CHIR诱导的肠管形态发生和球体产生(图5B),单层培养物的免疫荧光染色以评价在CHIR/FGF处理的内胚层中的CDX2诱导和在头蛋白和CHIR/FGF/头蛋白处理的内胚层中的SOX2诱导(图5C),BMP靶基因MSX1/2的qPCR分析(图5D),以及在存在和不存在BMP2的情况下的SOX2和CDX2表达。
图6A-6G描绘了比较两个hESC系(H1和H9)和一个iPSC系(72.3)之间的球体形成和特征的表(图6A),源自H1和iPSC 72.3细胞系的第34天hGO的免疫荧光染色(图6B),第34天hGO中的器官上皮细胞类型定量(图6C),诱导的多能干细胞系iPSC 72.3的表征(图6D-G)。
图7A-7D描绘了显示前肠模式化(patterning)实验的示意图(图7A),显示RA增加了从前肠单层培养物产生的球体数目的明场图像(图7B),图1d的较低功率图像,其显示具有位于前肠后部部分的Hnf1β蛋白的14个体节期胚胎的免疫荧光图像(图7C),用RA处理的前肠球体中的基因表达的qPCR分析(图7D)。
图8描述了hGO分化的晚期阶段的明场图像和免疫染色。
图9描述了在体内窦发育的四个胚胎阶段(E12.5,E14.5,E16.5和E18.5)和一个出生后阶段(P12)期间在小鼠窦和人胃类器官发育期间的转录因子表达。
图10描述了E12.5窦和第13天hGO中的pHH3/E-Cad/DAPI表达和aPCC/E-CAD/DAPI表达。
图11A-11C描述了窦间质转录因子BAPX1的表达(图11A)和间质细胞类型标志物的染色(图11C)。
图12描绘了体内胃窦内分泌细胞发育。
图13A-13B描述了泛内分泌标志物CHGA的染色(图13A)和内分泌标志物CHGA,胃泌素,生长素释放肽和促生长素抑制素的表达(图13B)。
图14显示了胃类器官的定向分化的方法的概述。指示分化过程中的每个步骤以及代表性的立体显微照片。
图15描绘了小鼠胃的示意图和在前胃,胃底,窦和十二指肠中的已知区域标志物的测量。
图16描绘了在前胃,胃底,窦和十二指肠中的新的区域标志物的测量。
图17描绘了胃底特化方案和在对照,Wnt100,Wnt500和CHIR处理的细胞中的GAPDH,Gata4,Axin2,Sox2,Pdx1和Cdx2的测量。y轴代表相对基因表达。
图18描绘了胃底方案中Axin2,IRX2,IRX3,Pitx1和IRX4的测量。y轴代表相对基因表达。
图19是描述来自定形内胚层的肠组织,胃底组织和窦组织形成的示意图。
发明详述
除非另有说明,术语应根据相关领域的普通技术人员的常规用法来理解。
如本文中使用,术语“全能干细胞”(也称为全能干细胞)是可以分化成胚胎和胚外细胞类型的干细胞。此类细胞可以构建完整的,活的生物体。这些细胞由卵和精细胞的融合产生。受精卵的前几次分裂产生的细胞也是全能的。
如本文中使用,术语“多能干细胞(PSC)”包括可以分化为几乎所有身体细胞类型的任何细胞,即源自三个胚层(生殖上皮)中的任一个的细胞,包括内胚层(内部胃粘膜(interior stomach lining),胃肠道,肺),中胚层(肌肉,骨,血液,泌尿生殖器)和外胚层(表皮组织和神经系统)。PSC可以是前植入囊胚(primplantation blastocyst)的内细胞团细胞的后代,或通过强制某些基因的表达通过诱导非多能细胞(例如成体体细胞)获得。多能干细胞可以源自任何合适的来源,如本领域技术人员容易理解的。多能干细胞来源的实例包括哺乳动物来源,包括人,啮齿类,猪,牛,但不限于此。
如本文中使用,术语“诱导多能干细胞(iPSC)”,通常也缩写为iPS细胞,是指通过诱导某些基因的“强制”表达人工源自正常非多能细胞(如成体体细胞)的多能干细胞类型。
如本文中使用,术语“胚胎干细胞(ESC)”,通常也缩写为ES细胞,是指多能性并源自胚泡(早期胚胎)的内部细胞团的细胞。为了本发明的目的,术语“ESC”有时也广泛用于涵盖胚胎生殖细胞。
如本文中使用,术语“前体细胞”包括可用于本文所述的方法中的任何细胞,经由所述方法,一种或多种前体细胞获得更新自身或分化成一种或多种特化细胞类型的能力。在一些实施方案中,前体细胞是多能的或具有变成多能性的能力。在一些实施方案中,对前体细胞进行外部因子(例如生长因子)的处理以获得多能性。在一些实施方案中,前体细胞可以是全能(或全能)干细胞;多能干细胞(诱导或非诱导);多能干细胞;寡能干细胞和单能干细胞。在一些实施方案中,前体细胞可来自胚胎,婴儿,儿童或成人。在一些实施方案中,前体细胞可以是经受治疗的体细胞,使得通过遗传操作或蛋白质/肽治疗赋予多能性。
在发育生物学中,细胞分化是不太特化的细胞变为更特化的细胞类型的过程。如本文中使用,术语“定向分化”描述了不太特化的细胞变为特定的特化靶细胞类型的过程。可通过可用于定义或改变初始细胞的命运的任何可应用的方法来确定特化靶细胞类型的特殊性。示例性方法包括但不限于遗传操作,化学处理,蛋白质处理和核酸处理。
如本文中使用,术语“细胞成分”是单个基因,蛋白质,mRNA表达基因和/或任何其它可变的细胞组分或蛋白质活性,例如蛋白质修饰(例如磷酸化)的程度,例如,其通常由本领域技术人员在生物实验(例如,通过微阵列或免疫组织化学)中测量。与存活系统,常见的人类疾病,基因发现和结构测定根本的生物化学过程的复杂网络有关的重大发现现在可归因于作为研究过程的一部分的细胞组分丰度数据的应用。细胞组分丰度数据可以帮助鉴定生物标志物,区分疾病亚型和鉴定毒性的机制。
在所有多细胞生物体中发现干细胞。它们的特征在于通过有丝分裂细胞分裂自我更新并分化成多种多样的一批特化细胞类型的能力。两种广泛类型的哺乳动物干细胞是1)从胚泡的内部细胞团分离的胚胎干细胞,和2)在成体组织中发现的成体干细胞。在发育中的胚胎中,干细胞可以分化成所有特化的胚胎组织。在成年生物体中,干细胞和祖细胞充当身体的修复系统,补充特化的细胞,但也保持再生器官(如血液,皮肤或胃组织)的正常更新。
现在可以通过细胞培养使干细胞生长并转化成具有与各种组织(如肌肉或神经)的细胞一致的特征的特化细胞。来自多种来源的高度塑性成体干细胞,包括脐带血和骨髓,常规用于医疗疗法。通过治疗性克隆产生的胚胎细胞系和自体胚胎干细胞也已被提出作为未来治疗的有希望的候选者。
干细胞的经典定义通常指示两种性质:自我更新,经历多次细胞分裂周期同时保持未分化状态的能力,以及效力,分化成特化细胞类型的能力。在一些实施方案中,干细胞是全能的或多能的,即它们能够产生任何成熟细胞类型,尽管多能或单能祖细胞有时可以称为干细胞。
效力指定干细胞的分化潜能(分化成不同细胞类型的潜力)。全能干细胞(也称为全能性干细胞)可以分化成胚胎和胚外细胞类型。这些细胞可以构建完整的,活的生物体。细胞由卵和精细胞的融合产生。受精卵的前几次分裂产生的细胞也是全能的。多能干细胞(PSC)是全能细胞的后代,并且可以分化成几乎所有细胞,即源自三个胚层中的任一个的细胞,包括内胚层(内部胃粘膜,胃肠道,肺),中胚层(肌肉,骨,血,泌尿生殖器)和外胚层(表皮组织和神经系统)。多能干细胞可以分化成许多细胞,但是仅分化为密切相关的细胞家族的细胞。寡能干细胞可以分化成仅少数细胞,如淋巴样或髓样干细胞。单能细胞只能产生一种它们自己的细胞类型,但具有将其与非干细胞(例如肌肉干细胞)区分开的自我更新的性质。
胚胎和诱导的多能干细胞对研究人类疾病和产生在动物模型中治疗有效的替代组织的能力已经具有前所未有的影响。
在发育生物学中,细胞分化是不太特化的细胞变成更特化的细胞类型的过程。定向将人PSC分化为治疗性细胞类型的最成功的努力是基于胚胎器官发育的研究。实例包括肝的肝细胞和胰腺内分泌细胞的产生,其在肝病和糖尿病的动物模型中显示出功能潜力。类似地,PSC分化为肠可以为疾病,如坏死性小肠结肠炎(necrotizing enterocolitis),炎性肠病和短肠综合征提供治疗益处。
如上讨论,多能干细胞具有分化成三种胚层中的任一种的潜能:内胚层(内部胃粘膜,胃肠道,肺),中胚层(肌肉,骨,血液,泌尿生殖器)和外胚层(表皮组织和神经系统)。因此,多能干细胞可以产生任何胎儿或成人细胞类型。然而,特定多能干细胞的命运由许多细胞信号传导途径和许多因素控制。此外,多能干细胞单独不能发育成胎儿或成年动物,因为它们缺乏有助于胚胎外组织,如胎盘的潜力。
迄今为止,尚未从人多能干细胞(hPSC)产生胃组织。将PSC分化为肺,肝,胰腺和肠细胞的成功努力已经取决于对这些器官的胚胎发育的良好的分子理解。6-10不幸的是,本领域中的一个问题是在内胚层形成后的胃发育的理解中的许多缺口。因此,为了指导hPSC分化为胃组织,申请人鉴定了调节早期胃发育的几个关键阶段的信号传导途径,包括前肠特化和模式化,胃特化,以及最后胃上皮生长和分化。此外,为了产生更功能性的,复杂的三维组织,申请人旨在诱导在胃发育期间发生的几种形态发生过程,包括前肠管的形态发生和胃上皮结构(包括腺体和凹陷(pit))的形成。
如本文中所述,使用生长因子操作的时间系列来建立方法和系统,以模拟培养物中的胚胎胃组织发育。特别地,建立了在体外指导PSC(人胚胎干细胞(hESC)和诱导多能干细胞(iPSC)二者)分化成胃组织的方法和系统。这些因素在体外在接近胎儿肠发育的阶段中指导人类肠发育:激活蛋白诱导的定形内胚层(DE)形成;FGF/Wnt/BMP诱导后部前肠模式化,和最后通过调节视黄酸和EFG信号转导获得的前胃培养系统,所述信号转导促进胃组织生长,形态发生和细胞分化成功能性胃细胞类型和形态,包括胃腺体和凹陷,增殖区,表面和窦粘液细胞和表达胃泌素,生长素释放肽和促生长素抑制素的内分泌细胞。
申请人已经鉴定了新的胚胎信号传导途径,其允许人PSC有效地逐步分化成具有复杂构造和细胞组成的胃细胞,胃组织和/或三维胃组织(hGO)。申请人进一步发现了,发育中的hGO经历分子和形态学分化阶段,其与小鼠发育中的窦几乎相同,并且所得的胃类器官可含有构成与胎儿/出生后胃相当的正常窦上皮和三维构造的粘液,内分泌和祖细胞的排列(array)。
所公开的人胃细胞,胃组织和/或胃类器官(hGO)可以用作体外系统以鉴定人胃发育,生理学的新机制,并且可以用作胃上皮对幽门螺杆菌的病理生理应答的模型。所公开的胃细胞,胃组织和/或胃hGO和方法为药物发现和胃癌的早期阶段的建模呈现了新的机会。此外,本文中公开了人胚胎前肠的第一次三维产生,其是产生其它前肠器官组织,包括肺和胰腺的有前景的起点。
在一个方面,公开了诱导从前体细胞形成胃细胞,胃组织和/或胃hGO的方法。所述方法可以包括以下步骤:a)激活前体细胞内的一种或多种信号传导途径,其中所述一种或多种信号传导途径选自WNT信号传导途径,WNT/FGF信号传导途径和FGF信号传导途径,以获得从前体细胞转变的胃细胞,胃组织和/或胃hGO。该方法可以进一步包括抑制前体细胞内的一种或多种信号传导途径的步骤b)。受到抑制的一种或多种信号传导途径可以包含BMP信号传导途径。
该方法还可以包括使前体细胞与视黄酸接触的步骤。前体细胞与视黄酸的接触可以在上述激活和抑制步骤之后发生。
该方法可以进一步包括以足以将胃类器官的直径增加到直径大于约1mm或直径大于约2mm,或直径大于约3mm,或直径大于约4mm的浓度和/或时间长度使胃类器官与EGF接触的步骤。
在一个方面,所述一种或多种信号传导途径可以选自Wnt信号传导途径,Wnt/β-连环蛋白信号传导,Wnt/APC信号传导和Wnt/PCP途径信号传导。
在一个方面,激活Wnt信号途径的步骤可包括使前体细胞与选自Wnt1,Wnt2,Wnt2b,Wnt3,Wnt3a,Wnt4,Wnt5a,Wnt5b,Wnt6,Wnt7a,Wnt7b,Wnt8a,Wnt8b,Wnt9a,Wnt9b,Wnt10a,Wnt10b,Wnt11和Wnt16的一种或多种分子接触。
在一个方面,激活FGF信号传导途径的步骤可包括使前体细胞与选自FGF1,FGF2,FGF3,FGF4,FGF5,FGF6,FGF7,FGF8,FGF9,FGF10,FGF11,FGF12,FGF13,FGF14,FGF16,FGF17,FGF18,FGF19,FGF20,FGF21,FGF22和FGF23的一种或多种分子接触。
在一个方面,抑制BMP信号传导途径的步骤可以包括使前体细胞与BMP抑制剂接触。在一个方面,BMP抑制剂可以选自Dorsomorphin,LDN189,DMH-1,头蛋白及其组合。在一个方面,BMP抑制剂可以是头蛋白。
在一个方面,激活步骤可以包括在称为温育期的规定期里使前体细胞与Wnt3a,FGF4和BMP抑制剂接触。接触步骤可以同时进行,或者在其它方面,接触步骤可以随后进行。
在一个方面,在第一温育期可以通过信号传导剂接触可包含定形内胚层的前体细胞,所述信号传导剂可以包含1)Wnt3a或GSK-抑制剂(例如CHIRON)与2)FGF4的组合。第一温育期可以进一步包含BMP抑制剂。在第一温育期后,可将前体细胞进行第二温育期,其中使前体细胞与视黄酸(RA)接触。在一个方面,第一温育期和第二温育期重叠。在一些实施方案中,第一温育期和第二温育期不重叠。
在一个方面,第一和/或第二温育期和/或第一和第二温育期的总体可以为24至120小时,或约36至约108小时,或约48至约110小时,约96小时,或约60至约84小时。在一个方面,第一温育期可以是至少约24小时。
一方面,第二温育期(其中前体细胞可与RA接触)在第一温育期后约72小时开始。在另一方面,第二温育期在培养物已经从前体细胞形成前肠球体之后开始。然后,可将前肠球体在适于形成胃类器官的生长条件下转移至3-维基质,例如通过将前肠球应用于基质胶TM(Corning,BD Bioscience)进行。在转移到基质胶后,在第三温育期期间使前肠球体与RA接触,在第三温育期中可发生持续的3D生长。然后,可以在第四温育期期间使球体与EGF接触,所述第四温育期可与第三温育期重叠。第三温育期可以是约24小时。
在一个方面,前体细胞可以与50-1500ng/ml,或约100至约1200ng/ml,或约200至约1000ng/ml,或约300至约900ng/ml,或约400至约800ng/ml,或约500至约700ng/ml的浓度的Wnt3a接触。
在一个方面,前体细胞可以选自胚胎干细胞,胚胎生殖细胞,诱导的多能干细胞,中胚层细胞,定形内胚层细胞,后内胚层细胞和后肠细胞。
在一个方面,前体细胞可以是源自多能干细胞的定形内胚层细胞。
在一个方面,前体细胞可以是多能干细胞,如胚胎干细胞,胚胎干细胞或诱导的多能干细胞。
在一个方面,可通过使多能干细胞与选自激活蛋白,TGF-β生长因子的超家族的BMP亚类;Nodal,激活蛋白A,激活蛋白B,BMP4,Wnt3a及其组合的一种或多种分子接触衍生定形内胚层细胞。
在一个方面,可以从一种或多种前体细胞体外产生胃组织。
在一个方面,一种或多种前体细胞可以选自胚胎干细胞,中胚层细胞,定形内胚层细胞,后内胚层细胞,前内胚层细胞,前肠细胞和后肠细胞。
在一个方面,多能干细胞可以是哺乳动物多能干细胞,包括但不限于人多能干细胞或小鼠多能干细胞。
在一个方面,人多能干细胞可以选自人胚胎干细胞,人胚胎生殖细胞和诱导的人多能干细胞。
在一个方面,提供了包含在体外从一种或多种前体细胞产生的胃细胞,组织或类器官的试剂盒。
在一个方面,提供了用于鉴定胃细胞或组织的吸收效应的方法。该方法可以包括以下步骤:使源自前体细胞的胃细胞,组织或类器官与化合物接触;并且检测所述胃细胞或组织对化合物的吸收水平。
在一个方面,提供了用于鉴定化合物对胃细胞或组织的毒性的方法。该方法可以包括以下步骤:使源自前体细胞的胃细胞,组织或类器官与化合物接触;并且检测所述胃细胞或组织对化合物的吸收水平。
在一个方面,公开了包含从头产生的三维人胃类组织(hGO)的组合物,以及通过人多能干细胞(hPSC)的定向分化来制备它们的方法。此类hGO可用于对胃发育以及幽门螺杆菌感染期间发生的早期事件建模。
在一个方面,公开了通过人多能干细胞(hPSC)的定向分化体外产生hGO的方法。该人胃组织可用于对人胃发育和疾病建模。还公开了诱导定形内胚层(DE)以形成3维肠管结构的方法。在一个方面,这可以通过激活FGF和WNT信号传导来进行,而前肠命运可以通过同时抑制BMP信号传导来促进。然后,可以通过操纵视黄酸和EGF信号传导将前肠球体定向为后部前肠和胃命运,产生hGO。
形成中的hGO可以经历与发育中的小鼠窦几乎相同的分子和形态发生变化,形成胃腺和凹陷,增殖区,表面和窦粘液细胞和表达胃泌素,生长素释放肽和促生长素抑制素的内分泌细胞。使用hGO对人胃发育建模,已经确定EGF信号传导抑制转录因子NEUROGENIN 3上游的内分泌细胞发育。申请人进一步发现hGO忠实地重演由幽门螺杆菌引发的胃疾病的早期阶段,包括快速激活c-Met信号传导和上皮增殖。这些研究一起描述一个新颖和强力的体外系统以阐明人类胃发育和疾病的根本机制。
源自胚胎细胞的多能干细胞
在一个方面,方法可包括获得多能性或可被诱导变成多能性的干细胞的步骤。在一些实施方案中,多能干细胞源自胚胎干细胞,其又源自早期哺乳动物胚胎的全能细胞,并且能够在体外具有无限的未分化增殖。胚胎干细胞是源自胚泡(早期胚胎)的内细胞团的多能干细胞。用于从胚细胞衍生胚胎干细胞的方法是本领域熟知的。例如,虽然本文中举例说明了某些细胞类型,但是本领域技术人员应当理解,本文中所述的方法和系统适用于任何干细胞。
可以在根据本发明的实施方案中使用的另外的干细胞包括但不限于以下提供的或由以下主办的数据库;和由普林斯顿大学和宾夕法尼亚大学主办的干细胞数据库中描述的那些干细胞:国家干细胞库(National Stem Cell Bank,NSCB),人类胚胎干细胞研究中心(NSCB),在加利福尼亚大学旧金山分校(UCSF);WISC细胞库,在Wi细胞研究所;威斯康星大学干细胞和再生医学中心(UW-SCRMC);Novocell,Inc.(San Diego,Calif.);Cellartis AB(Goteborg,Sweden);ES Cell International Pte Ltd(新加坡);Technion,以色列理工学院(以色列海法)。可用于根据本发明的实施方案中的示例性胚胎干细胞包括但不限于SA01(SA001);SA02(SA002);ES01(HES-1);ES02(HES-2);ES03(HES-3);ES04(HES-4);ES05(HES-5);ES06(HES-6);BG01(BGN-01);BG02(BGN-02);BG03(BGN-03);TE03(13);TE04(14);TE06(16);UC01(HSF1);UC06(HSF6);WA01(H1);WA07(H7);WA09(H9);WA13(H13);WA14(H14)。
在一些实施方案中,干细胞可以进一步修饰以掺入另外的性质。示例性修饰的细胞系包括但不限于H1 OCT4-EGFP;H9 Cre-LoxP;H9 hNanog-pGZ;H9 hOct4-pGZ;GFPhES中的H9;以及H9 Syn-GFP。
关于胚胎干细胞的更多细节可以参见例如Thomson et al.,1998,“Embryonic Stem Cell Lines Derived from Human Blastocysts,”Science 282(5391):1145-1147;Andrews et al.,2005,“Embryonic stem(ES)cells and embryonal carcinoma(EC)cells:opposite sides of the same coin,”Biochem Soc Trans 33:1526-1530;Martin 1980,“Teratocarcinomas and mammalian embryogenesis,”.Science 209(4458):768-776;Evans and Kaufman,1981,“Establishment in culture of pluripotent cells from小鼠embryos,”Nature 292(5819):154-156;Klimanskaya et al.,2005,“Human embryonic stem cells derived without feeder cells,”Lancet 365(9471):1636-1641;每篇在此以其整体并入本文。
或者,多能干细胞可源自胚胎生殖细胞(EGC),其是产生有性繁殖的生物体的配子的细胞。EGC源自在晚期胚胎的性腺脊中发现的原始生殖细胞,具有胚胎干细胞的许多性质。胚胎中的原始生殖细胞发育成干细胞,其在成年人中产生繁殖配子(精子或卵)。在小鼠和人中,可以在合适的条件下在组织培养物中培养胚胎生殖细胞。EGC和ESC二者都是多能的。为了本发明的目的,术语“ESC”有时广泛地用于涵盖EGC。
诱导的多能干细胞(iPSC)
在一些实施方案中,通过将某些干细胞相关基因转染到非多能细胞(如成体成纤维细胞)中衍生iPSC。转染通常通过病毒载体,如逆转录病毒实现。转染的基因包括主转录调节子Oct-3/4(Pouf51)和Sox2,虽然提示了其它基因增强诱导的效率。3-4周后,少量转染的细胞开始在形态和生物化学上类似于多能干细胞,并且通常通过形态选择,倍增时间或通过报告基因和抗生素选择来分离。如本文中使用,iPSC可包括但不限于第一代iPSC,小鼠中的第二代iPSC和人诱导的多能干细胞。在一些实施方案中,逆转录病毒系统可用于使用四种关键基因:Oct3/4,Sox2,Klf4和c-Myc将人成纤维细胞转化为多能干细胞。在备选实施方案中,慢病毒系统用于用OCT4,SOX2,NANOG和LIN28转化体细胞。其表达可在iPSC中诱导的基因包括但不限于Oct-3/4(例如Pou5fl);Sox基因家族的某些成员(例如Sox1,Sox2,Sox3和Sox15);Klf家族的某些成员(例如Klf1,Klf2,Klf4和Klf5),Myc家族的某些成员(例如C-myc,L-myc和N-myc),Nanog和LIN28。
在一些实施方案中,可以采用基于非病毒的技术来产生iPSC。在一些实施方案中,腺病毒可用于将必需的四种基因转运到小鼠的皮肤和肝细胞的DNA中,产生与胚胎干细胞相同的细胞。由于腺病毒不将其自身基因中的任一种与靶向宿主组合,消除了创建肿瘤的危险。在一些实施方案中,重编程可以通过根本没有任何病毒转染系统的质粒完成,尽管效率非常低。在其它实施方案中,蛋白质的直接递送用于产生iPSC,因此消除对病毒或遗传修饰的需要。在一些实施方案中,使用类似的方法可以产生小鼠iPSC:用通过聚精氨酸锚定物引导入细胞中的某些蛋白质重复处理细胞足以诱导多能性。在一些实施方案中,多能性诱导基因的表达也可以通过在低氧条件下用FGF2处理体细胞来增加。
关于胚胎干细胞的更多细节可以参见例如Kaji et al.,2009,“Virus free induction of pluripotency and subsequent excision of reprogramming factors,”Nature 458:771-775;Woltjen et al.,2009,“piggyBac transposition reprograms fibroblasts to induced pluripotent stem cells,”Nature 458:766-770;Okita et al.,2008,“Generation of小鼠Induced Pluripotent Stem Cells Without Viral Vectors,”Science 322(5903):949-953;Stadtfeld et al.,2008,“Induced Pluripotent Stem Cells Generated without Viral Integration,”Science 322(5903):945-949;and Zhou et al.,2009,“Generation of Induced Pluripotent Stem Cells Using Recombinant Proteins,”Cell Stem Cell 4(5):381-384;每篇在此以其整体并入本文。
在一些实施方案中,示例性iPS细胞系包括但不限于iPS-DF19-9;iPS-DF19-9;iPS-DF4-3;iPS-DF6-9;iPS(包皮);iPS(IMR90);和iPS(IMR90)。
已经显示iPSC能够以类似于ESC的方式分化成完全分化的组织。例如,将iPSC分化成表达βIII-微管蛋白,酪氨酸羟化酶,AADC,DAT,ChAT,LMX1B和MAP2的神经元。儿茶酚胺相关酶的存在可能表明iPSCs,像hESC一样,可以是可分化成多巴胺能神经元。显示干细胞相关基因在分化后下调。还已经显示iPSC可以分化成自发开始跳动的心肌细胞。心肌细胞表达TnTc,MEF2C,MYL2A,MYHCβ和NKX2.5。干细胞相关基因在分化后下调。
胃器官和发育
在申请人的发明之前,没有系统可用于将前体细胞,如胚胎干细胞和/或iPSC转化为胃组织。
在一些实施方案中,PSC,如ESC和iPSC以逐步方式首先进行定向分化成内胚层(DE),然后分化成三维肠管结构(前肠球体),然后通过后部前肠/胃组织的形成分化成三维胃类器官组织(hGO)。
在一些实施方案中,PSC,如ESC和iPSC以非步进方式进行定向分化,其中用于促进DE形成的分子(例如生长因子,配体)和用于随后组织形成的分子在相同的时间添加。
定形内胚层
胃的上皮源自称为定形内胚层(DE)的简单的细胞片层。前部DE形成前肠及其相关器官,包括肺,食道,胃,肝和胰腺,并且后部DE形成中肠和后肠,其形成小肠和大肠以及泌尿生殖系统的部分。DE在体内产生胃肠道和呼吸道的上皮。使用小鼠,鸡和青蛙胚胎的研究表明在原肠胚阶段在DE中建立前部-后部模式是随后的前肠和后肠发育的先决条件。在一些实施方案中,PSC,如ESC和iPSC以逐步的方式首先定向分化成定形内胚层(DE),然后分化成前部/前肠上皮(例如,前肠球),然后分化成胃组织。认为BMP,Wnt和FGF信号传导途径对于该过程是关键的。WNT和FGF的激活作用为促进肠管形态发生,并且抑制BMP信号传导促进前肠命运。前肠的简单立方上皮首先发育成假复层柱状上皮,然后发育成腺体和凹陷,它们含有胃上皮和在绒毛基部的增殖区,其对应于假定的祖先域。
建立了强力和有效的过程,以在体外指导DE分化成胃组织。在一些实施方案中,通过选择性激活iPSC和/或DE细胞中的某些信号传导途径来实现定向分化。在一些实施方案中,信号传导途径是在胃组织发育中有活性的那些,包括但不限于Wnt信号传导途径,Wnt/APC信号传导途径,FGF信号传导途径,TGF-β信号传导途径,BMP信号传导途径;EGF信号途径和视黄酸信号途径。
关于与DE发育和/或肠发育相关的信号传导途径的功能的更多细节一般可以参见例如Zorn and Wells,2009,“Vertebrate endoderm development and organ formation,”Annu Rev Cell Dev Biol 25:221-251;Dessimoz et al.,2006,“FGF signaling is necessary for establishing gut tube domains along the anterior-posterior axis in vivo,”Mech Dev 123:42-55;McLin et al.,2007,“Repression of Wnt/{beta}-catenin signaling in the anterior endoderm is essential for liver and pancreas development.Development,”134:2207-2217;Wells and Melton,2000,Development 127:1563-1572;de Santa Barbara et al.,2003,“Development and differentiation of the intestinal epithelium,”Cell Mol Life Sci 60(7):1322-1332;Sancho et al.,2004,“Signaling Pathways in Intestinal Development and Cancer,”Annual Review of Cell and Developmental Biology 20:695-723;Logan and Nusse,2004,“The Wnt Signaling Pathway in Development and Disease,”Annual Review of Cell and Developmental Biology 20:781-810;Taipalel and Beachyl,2001,“The Hedgehog and Wnt signalling pathways in cancer,”Nature 411:349-354;Gregorieff and Clevers,2005,“Wnt signaling in the intestinal epithelium:from endoderm to cancer,”Genes&Dev.19:877-890;每篇在此以其整体并入本文。
用于从多能细胞(例如iPSC或ESC)产生定形内胚层的任何方法适用于本文所述的方法。在一些实施方案中,多能细胞源自桑椹胚。在一些实施方案中,多能干细胞是干细胞。在这些方法中使用的干细胞可以包括但不限于胚胎干细胞。胚胎干细胞可以源自胚胎内细胞团或胚胎性腺脊。胚胎干细胞或生殖细胞可以源自多种动物物种,包括但不限于各种哺乳动物物种,包括人。在一些实施方案中,人胚胎干细胞用于产生定形内胚层。在一些实施方案中,人胚胎生殖细胞用于产生定形内胚层。在一些实施方案中,iPSC用于产生定形内胚层。
在一些实施方案中,在从多能干细胞到DE细胞的分化过程中使用一种或多种生长因子。在分化过程中使用的一种或多种生长因子可以包括来自TGF-β超家族的生长因子。在此类实施方案中,一种或多种生长因子包含TGF-β生长因子超家族的Nodal/激活蛋白和/或BMP亚群。在一些实施方案中,所述一种或多种生长因子选自Nodal,激活蛋白A,激活蛋白B,BMP4,Wnt3a或任何这些生长因子的组合。
在一些实施方案中,用一种或多种生长因子将胚胎干细胞或诱导的多能细胞和iPSC处理6小时以上;12小时以上;18小时以上;24小时以上;36小时以上;48小时以上;60小时以上;72小时以上;84小时以上;96小时以上;120小时以上;150小时以上;180小时以上;或240小时以上。
在一些实施方案中,用浓度为10ng/ml或更高;20ng/ml或更高;50ng/ml或更高;75ng/ml或更高;100ng/ml或更高;120ng/ml或更高;150ng/ml或更高;200ng/ml或更高;500ng/ml或更高;1,000ng/ml或更高;1,200ng/ml或更高;1,500ng/ml或更高;2,000ng/ml或更高;5,000ng/ml或更高;7,000ng/ml或更高;10,000ng/ml或更高;或15,000ng/ml或更高的一种或多种生长因子处理胚胎干细胞或生殖细胞和iPSC。在一些实施方案中,在整个治疗中生长因子的浓度维持在恒定水平。在其它实施方案中,生长因子的浓度在治疗过程中变化。在一些实施方案中,生长因子悬浮在包括具有不同HyClone浓度的胎牛丝氨酸(FBS)的培养基中。本领域技术人员将理解,本文所述的方案适用于单独或组合的任何已知的生长因子。当使用两种或更多种生长因子时,每种生长因子的浓度可以独立地变化。
在一些实施方案中,使用富含定形内胚层细胞的细胞群体。在一些实施方案中,定形内胚层细胞是分离的或基本纯化的。在一些实施方案中,分离的或基本上纯化的定形内胚层细胞以比OCT4,AFP,TM,SPARC和/或SOX7标志物更大的程度表达SOX17,FOXA2和/或CXRC4标志物。
也涵盖了用定形内胚层富集细胞群体的方法。在一些实施方案中,可以如下从混合的细胞群体分离或基本上纯化定形内胚层细胞:使细胞与结合存在于定形内胚层细胞表面上但不存在于混合细胞群体中的其它细胞表面上的分子的试剂接触,然后分离与试剂结合的细胞。在某些实施方案中,存在于定形内胚层细胞表面上的细胞成分是CXCR4。
本发明的其它实施方案涉及CXCR4抗体,SDF-1配体或CXCR4的其它配体可用于获得富集,分离或基本纯化形式的定形内胚层细胞。例如,CXCR4抗体,SDF-1配体或CXCR4的另一种配体可以在诸如基于亲和力的分离或基于磁性的分离的方法中用作试剂,以富集,分离或基本上纯化结合试剂的定形内胚层的制备物。
在一些实施方案中,用一种或多种生长因子处理定形内胚层细胞和hESC。此类生长因子可以包括来自TGF-β超家族的生长因子。在此类实施方案中,一种或多种生长因子包含TGF-β生长因子超家族的Nodal/激活蛋白和/或BMP亚组。在一些实施方案中,所述一种或多种生长因子选自Nodal,激活蛋白A,激活蛋白B,BMP4,Wnt3a或任何这些生长因子的组合。
用于获得或创建可用于本发明的DE细胞的其它方法包括但不限于描述于D'Amour等人的美国专利号7,510,876;Fisk等人的美国专利号7,326,572;Kubol et al.,2004,“Development of definitive endoderm from embryonic stem cells in culture,”Development 131:1651-1662;D'Amour et al.,2005,“Efficient differentiation of human embryonic stem cells to definitive endoderm,”Nature Biotechnology 23:1534-1541;and Ang et al.,1993,“The formation and maintenance of the definitive endoderm lineage in the小鼠:involvement of HNF3/forkhead proteins,”Development 119:1301-1315;其各自通过引用整体并入本文。
后化DE(posteriorized DE)的定向分化
在一些实施方案中,激活蛋白诱导的定形内胚层(DE)可进一步经历FGF/Wnt/头蛋白诱导的前内胚层模式化,前肠特化和形态发生,以及最后前胃培养系统,以促进胃组织生长,形态发生和细胞分化成功能性胃细胞类型,包括表面粘液细胞,粘液腺细胞,内分泌和祖细胞。在一些实施方案中,人PSC被有效地导向在体外分化成包括粘液,内分泌和祖细胞类型的胃上皮。应当理解,诸如生长因子等分子可以添加到发育的任何阶段以促进特定类型的胃组织形成。
在一些实施方案中,DE的前化(anteriorized)内胚层细胞进一步发育成一种或多种特化的细胞类型。
在一些实施方案中,可溶性FGF和Wnt配体和BMP拮抗剂用于模拟培养物中的早期前肠规定,以通过定向分化将从iPSC或ESC发育的DE转化成前肠上皮,其有效地产生所有主要胃窦胃细胞类型。在人类中,通过选择性激活对胃发育重要的某些信号传导途径来实现DE的定向分化。
人胃/胃发育在体外发生在近似胎儿肠发育的阶段;内胚层形成,前内胚层模式化,前肠形态发生,胎儿胃,窦和胃底发育,上皮形态发生,推定的祖先域的形成和分化为胃的功能性细胞类型。
本领域技术人员应当理解,根据本发明改变任何Wnt信号传导蛋白与任何FGF配体的组合的表达可以产生定向分化。在一些实施方案中,所述改变是Wnt3,特别是Wnt3a的过表达。在一些实施方案中,所述改变是Wnt1或其它Wnt配体的过表达。
本领域技术人员应当理解,根据本发明,改变Wnt信号传导途径的信号传导活性与改变FGF信号传导途径的信号传导活性相组合可以产生定向分化。在一些实施方案中,所述改变是通过使用激活上述途径的小分子调节剂。例如,Wnt途径的小分子调节剂包括但不限于氯化锂;2-氨基-4,6-二取代的嘧啶(杂)芳基嘧啶;IQ1;QS11;NSC668036;DCAβ-连环蛋白;2-氨基-4-[3,4-(亚甲基二氧基)-苄基-氨基]-6-(3-甲氧基苯基)嘧啶。
在可选的实施方案中,可以抑制与Wnt和/或FGF信号传导途径相关的细胞成分,例如,途径的天然抑制剂或拮抗剂,导致Wnt和/或FGF信号传导途径的激活。
在一些实施方案中,细胞成分被其它细胞成分或外在分子抑制。Wnt信号传导的示例性天然抑制剂包括但不限于Dkk1,SFRP蛋白和FrzB。在一些实施方案中,外在分子可以包括但不限于小分子,例如WAY-316606;SB-216763;或BIO(6-溴靛玉红-3'-肟(6-bromoindirubin-3′-oxime))。
更多细节参见例如Liu et al.,“A small-molecule agonist of the Wnt signaling pathway,”Angew Chem Int Ed Engl.44(13):1987-1990(2005);Miyabayashi et al.,“Wnt/beta-catenin/CBP signaling maintains long-term murine embryonic stem cell pluripotency,”Proc Natl Acad Sci USA.104(13):5668-5673(2007);Zhang et al.,“Small-molecule synergist of the Wnt/beta-catenin signaling pathway,”Proc Natl Acad Sci U S A.104(18):7444-7448(2007);Neiiendam et al.,“An NCAM-derived FGF-receptor agonist,the FGL-peptide,induces neurite outgrowth and neuronal survival in primary rat neurons,”J.Neurochem.91(4):920-935(2004);Shan et al.,“Identification of a specific inhibitor of the dishevelled PDZ domain,”Biochemistry 44(47):15495-15503(2005);Coghlan et al.,“Selective small molecule inhibitors of glycogen synthase kinase-3 modulate glycogen metabolism and gene transcription,”Chem.Biol.7(10):793-803(2000);Coghlan et al.,“Selective small molecule inhibitors of glycogen synthase kinase-3 modulate glycogen metabolism and gene transcription,”Chemistry&Biology 7(10):793-803;和Pai et al.,“Deoxycholic acid activates beta-catenin signaling pathway and increases colon cell cancer growth and invasiveness,”Mol Biol Cell.15(5):2156-2163(2004);每篇通过引用整体并入本文。
在一些实施方案中,使用靶向与Wnt和/或FGF信号传导途径相关的细胞成分的siRNA和/或shRNA来激活这些途径。本领域技术人员应当理解,靶细胞组分可以包括但不限于SFRP蛋白;GSK3,Dkk1和FrzB。
关于基于RNAi的技术的更多细节可以参见例如Couzin,2002,Science 298:2296-2297;McManus et al.,2002,Nat.Rev.Genet.3,737-747;Hannon,G.J.,2002,Nature 418,244-251;Paddison et al.,2002,Cancer Cell 2,17-23;Elbashir et al.,2001.EMBO J.20:6877-6888;Tuschl et al.,1999,Genes Dev.13:3191-3197;Hutvagner et al.,Sciencexpress 297:2056-2060;每篇通过引用整体并入本文。
成纤维细胞生长因子(FGF)是涉及血管生成,伤口愈合和胚胎发育的生长因子家族。FGF是肝素结合蛋白,并且已经显示与细胞表面相关的硫酸乙酰肝素蛋白聚糖的相互作用对于FGF信号转导是必需的。FGF是极其多种细胞和组织的增殖和分化过程中的关键参与者。在人类中,已鉴定了FGF家族的22个成员,所有这些都是结构相关的信号传导分子。成员FGF1至FGF10都结合成纤维细胞生长因子受体(FGFR)。FGF1也称为酸性,并且FGF2也称为碱性成纤维细胞生长因子。成员FGF11,FGF12,FGF13和FGF14,也称为FGF同源因子1-4(FHF1-FHF4),已显示与FGF相比具有独特的功能差异。虽然这些因子具有显著相似的序列同源性,但它们不结合FGFR并且参与与FGF无关的胞内过程。该组也称为“iFGF”。成员FGF16至FGF23是较新的,并且没有被良好表征。FGF15是人FGF19的小鼠直向同源物(因此不存在人FGF15)。人FGF20基于其与蟾蜍(Xenopus)FGF-20(XFGF-20)的同源性鉴定。与其它FGF的局部活性相反,FGF15/FGF19,FGF21和FGF23具有更多的全身作用。
在一些实施方案中,本领域技术人员将理解,任何FGF可以与来自Wnt信号传导途径的蛋白质联合使用。在一些实施方案中,可溶性FGF可包括但不限于FGF4,FGF2和FGF3。
在一些实施方案中,FGF信号传导途径的细胞组分受其它细胞成分或外在分子抑制。FGF信号传导的示例性天然抑制剂可以包括但不限于Sprouty蛋白家族和Spred蛋白家族。如上所讨论,蛋白质,小分子,核酸可用于激活FGF信号传导途径。
本领域技术人员将理解,通过实例提供了本文所述的与Wnt和FGF信号传导途径有关的方法和组合物。类似的方法和组合物适用于本文中公开的其它信号传导途径。
在一些实施方案中,可用本文所述的信号传导途径的一个或多个分子处理DE培养物达6小时以上;12小时以上;18小时以上;24小时以上;36小时以上;48小时以上;60小时以上;72小时以上;84小时以上;96小时以上;120小时以上;150小时以上;180小时以上;200小时或更多小时,240小时或更多小时;270小时以上;300小时以上;350小时以上;400小时以上;500小时以上;600小时以上;700小时以上;800小时以上;900小时以上;1,000小时以上;1,200小时以上;或1,500小时以上。
在一些实施方案中,以10ng/ml或更高;20ng/ml或更高;50ng/ml或更高;75ng/ml或更高;100ng/ml或更高;120ng/ml或更高;150ng/ml或更高;200ng/ml或更高;500ng/ml或更高;1,000ng/ml或更高;1,200ng/ml或更高;1,500ng/ml或更高;2,000ng/ml或更高;5,000ng/ml或更高;7,000ng/ml或更高;10,000ng/ml或更高;或15,000ng/ml或更高的浓度用本文所述的一种或多种信号传导途径分子处理DE培养物。在一些实施方案中,信号传导分子的浓度在整个治疗中保持恒定。在其它实施方案中,信号传导途径分子的浓度在治疗过程中变化。在一些实施方案中,将根据本发明的信号传导分子悬浮在包含DMEM和胎牛丝氨酸(FBS)的培养基中。FBS可以为2%以上;5%以上;10%以上;15%以上;20%以上;30%以上;或50%以上的浓度。本领域技术人员应当理解,本文所述的方案可单独或组合地应用于本文所述的信号传导途径的任何已知分子,包括但不限于Wnt和FGF信号传导途径中的任何分子。
在两种或更多的信号分子用于处理DE培养物的实施方案中,可以同时或分别添加信号传导分子。当使用两种或更多种分子时,每种分子的浓度可以独立地变化。
PSC分化成DE培养物并随后分化成各种中间成熟胃细胞类型可以通过阶段特异性细胞标志物的存在来确定。在一些实施方案中,代表性细胞成分的表达用于测定DE形成。代表性的细胞成分可包括但不限于,CMKOR1,CXCR4,GPR37,RTN4RL1,SLC5A9,SLC40A1,TRPA1,AGPAT3,APOA2,C20orf56,C21orf129,CALCR,CCL2,CER1,CMKOR1,CRIP1,CXCR4,CXorf1,DIO3,DIO30S,EB-1,EHHADH,ELOVL2,EPSTI1,FGF17,FLJ10970,FLJ21195,FLJ22471,FLJ23514,FOXA2,FOXQ1,GATA4,GPR37,GSC,LOC283537,MYL7,NPPB,NTN4,PRSS2,RTN4RL1,SEMA3E,SIAT8D,SLC5A9,SLC40A1,SOX17,SPOCK3,TMOD1,TRPA1,TTN,AW166727,AI821586,BF941609,AI916532,BC034407,N63706和AW772192。
适合于检测DE形成的另外的细胞组分可以参见例如2005年6月23日提交的美国专利申请流水号11/165,305;2005年12月22日提交的美国专利申请流水号11/317,387;2004年12月23日提交的美国专利申请流水号11/021,618;2005年4月26日提交的美国专利申请流水号11/021,618,11/115,868;2005年12月22日提交的美国专利申请流水号11/317,387;2006年6月23日提交的美国专利申请流水号11/474,211;2005年6月23日提交的美国专利申请流水号11/165,305;2008年8月29日提交的美国专利申请流水号11/587,735;2008年2月28日提交的美国专利申请流水号12/039,701;2009年3月30日提交的美国专利申请流水号12/414,482;2009年6月2日提交的美国专利申请流水号12/476,570;2008年7月21日提交的美国专利申请流水号12/093,590;2009年10月20日提交的美国专利申请流水号12/582,600;其各自通过引用整体并入本文。
在一些实施方案中,使用SOX2的表达揭示在DE已经与FGF4和Wnt3a加头蛋白温育一段时间,例如12小时以上;18小时以上;24小时以上;36小时以上;48小时以上;60小时以上;或90小时以上后的前肠形成倾向。在一些实施方案中,需要较长的温育期以实现稳定前内胚层表型,如通过长期表达CDX2测量。在此类实施方案中,温育时段可以是60小时以上;72小时以上;84小时以上;96小时以上;108小时或更长;120小时以上;140小时以上;160小时以上;180小时以上;200小时以上;240小时或更长;或300小时以上。
或者,在一些实施方案中,细胞组分(如后肠标志物例如CDX2)的不存在可用于揭示定向前肠形成。在一些实施方案中,胃转录因子PDX1,KLF5和SOX9可用于代表胃发育。在一些实施方案中,GATA4和/或GATA6蛋白表达可用于代表胃发育。在这些实施方案中,温育时段可以持续12小时以上;18小时以上;24小时以上;36小时以上;48小时以上;60小时以上;或90小时以上。或者,温育时段可以持续60小时以上;72小时以上;84小时以上;96小时以上;108小时以上;120小时以上;140小时以上;160小时或更长;180小时以上;200小时以上;240小时以上;或300小时以上。
在一些实施方案中,通过使用靶向相关信号传导途径中的分子的一抗和/或二抗的免疫组织化学测定细胞组分的丰度数据,例如蛋白质和/或基因表达水平。在其它实施方案中,通过微阵列分析测定细胞组分的丰度数据,例如蛋白质和/或基因表达水平。
或者,形态学变化可用于表示定向分化的进展。在一些实施方案中,前肠球体可以进一步经受用于进一步成熟的3维培养条件。此外,可以在6天以上;7天以上;9天以上;10天以上;12天以上;15天以上;20天以上;25天以上;28天以上;32天以上;36天以上;40天以上;45天以上;50天以上;或60天以上中观察胃类器官。
多能干细胞的定向分化
在一些实施方案中,通过“一步”方法将多能干细胞转化为胃细胞类型。例如,可以将多能干细胞分化为DE培养物的一种或多种分子(例如,激活蛋白A)与可以促进DE培养物的定向分化的另外的分子(例如Wnt3a/FGF4激活剂和BMP抑制剂)组合,以直接处理多能干细胞。
效用和试剂盒实施方案
在一些实施方案中,本文所述的胃组织或相关细胞类型可用于对药物筛选胃摄取和/或运输机制和/或幽门螺杆菌的治疗。例如,这可以以高通量方式进行,以筛选最容易吸收或有效的药物,并且可以增加为了研究药物胃吸收和胃毒性进行的1期临床试验。这可以包括小分子,肽,代谢物,盐的胞周和胞内转运机制。本文中公开的胃组织可以进一步用于评估与旨在与胃组织接触以评价生物相容性的任何药剂和/或装置的相容性。
在一些实施方案中,本文所述的胃细胞,胃组织和/或胃hGO可用于鉴定正常人胃发育的分子基础。
在一些实施方案中,本文所述的胃细胞,胃组织和/或胃hGO可用于鉴定影响人胃发育的先天缺陷的分子基础。
在一些实施方案中,本文所述的胃细胞,胃组织和/或胃hGO可用于校正由遗传突变引起的先天性胃遗传缺陷。特别地,影响人胃发育的突变可以使用本文所述的iPSC技术和遗传正常的胃组织或相关细胞类型来校正。在一些实施方案中,本文所述的胃组织或相关细胞类型可用于产生替换组织。遗传性疾病的实例包括但不限于Neurog3突变和肠道内分泌腺病,PTF1a突变和新生儿糖尿病,实现胃的肠内分泌细胞的PDX1突变。
在一些实施方案中,本文所述的胃细胞,胃组织和/或胃hGO可用于产生用于疾病或病症(如消化性溃疡病,梅内特里耶病(Ménétrier's disease))或胃癌患者的替换胃组织。
在一些实施方案中,本文所述的胃细胞,胃组织和/或胃hGO可用于研究与人宿主上皮和宿主免疫的微生物相互作用。
在一些实施方案中,本文所述的胃组织或相关细胞类型,特别是肠内分泌细胞可用于研究由胃内分泌介导的摄食行为,代谢的激素调节。
在一些实施方案中,本文所述的胃细胞,胃组织和/或胃hGO,特别是产生激素胃泌素或生长素释放肽的肠内分泌细胞,可用于研究和改善例如具有肥胖,代谢综合征或2型糖尿病的患者中的代谢控制。
在一些实施方案中,本文所述的胃细胞,胃组织和/或胃hGO可用于代替有需要的受试者中的任何损伤或移除的胃组织。
在一些实施方案中,本文所述的胃细胞,胃组织和/或胃hGO可用于筛选作用于胃组织的任何药物的毒性和功效。
在其中本文所述的胃细胞,胃组织和/或胃hGO用于测定化合物的吸收水平的一些实施方案中,化合物将与胃细胞,胃组织和/或胃hGO接触;并且可以量化由胃细胞,胃组织和/或胃hGO对化合物的吸收水平。在一些实施方案中,化合物可以用放射性同位素,荧光标记物和/或初级或次级可见标志物标记。
在一些实施方案中,开发诊断试剂盒或包装以包括本文所述并且基于一种或多种上述效用的胃细胞,胃组织和/或胃hGO。
已经详细描述了本发明,显而易见的是,在不脱离所附权利要求书中限定的本发明的范围的情况下,修改,变化和等同实施方案是可能的。此外,应当理解,本公开中的所有示例都作为非限制性示例提供。
实施例
提供以下非限制性实施例以进一步说明本文公开的本发明的实施方案。本领域技术人员应当理解,以下实施例中公开的技术代表已经发现在本发明的实践中良好地起作用的方法,因此可以被认为构成其实践的模式的实例。然而,根据本公开内容,本领域技术人员应当理解,在不脱离本发明的精神和范围的情况下,可以对所公开的具体实施方案进行许多改变,并且仍然获得相同或相似的结果。
多能干细胞培养物
从WiCell获得人胚胎干细胞系WA01(H1)和WA09(H9)。ESC和iPSC系在无滋养层条件下在mTesR1培养基(Stem Cell Technologies)中的HESC量化基质胶(BD Biosciences)上维持为集落。每四天用分散酶(Invitrogen)常规传代细胞。
DE诱导。
(图14中总结)在经基质胶(BD Biosciences)包被的24孔板中以每孔150,000个细胞将人ES和iPS细胞作为单细胞铺在加有ROCK抑制剂Y27632(10μM;Stemgent)的mTesR1培养基中。ROCK抑制剂在用于分化的铺板后增强干细胞的存活。从第二天开始,在含有渐增浓度的0%,0.2%和2.0%的限定胎牛血清(dFBS;Invitrogen)的RPMI 1640(Invitrogen)中用激活蛋白A(100ng ml-1;Cell Guidance Systems)处理细胞三天。
定形内胚层(DE)的分化
为了分化,使用accutase(Stem Cell Technologies),在具有ROCK抑制剂Y-27632(10μM;Stemgent)的mTesR1中以150,000个细胞/孔的密度将PSC作为单细胞铺板在基质胶包被的24孔板。次日,如前所述11,35,将PSC分化为DE。在RPMI 1640培养基(Invitrogen)中将细胞暴露于激活蛋白A(100ng ml-1;Cell Guidance Systems)三天,所述培养基含有渐增浓度的0%0.2%和2.0%限定胎牛血清(dFBS;Invitrogen)。此外,在DE诱导的第一天添加BMP4(50ngml-1;R&D Systems)。
内胚层模式化和肠管形态发生。
在DE诱导后,在具有2.0%dFBS的RPMI 1640中用生长因子/拮抗剂处理细胞三天。为了产生后部前肠球体,用头蛋白(200ng ml-1;R&D Systems),FGF4(500ng ml-1;R&D Systems)和WNT3A(500ng ml-1;R&D Systems)或CHIR99021(2μM;Stemgent)处理DE三天。CHIR99021是刺激Wnt信号传导途径的小分子。在最后一天添加RA(2μM;Sigma Aldrich)。三维生长和窦特化。如前所述10,12,将后部前肠球体包埋入的基质胶(BD Biosciences)中,随后在补充有N2(Invitrogen),B27(Invitrogen),L-谷氨酰胺,10μM HEPES,青霉素/链霉素,和EGF(100ng ml-1;R&D Systems)的Advanced DMEM/F12(Invitrogen)中培养。对于窦特化,在三维生长的前三天添加RA和头蛋白。对于内分泌细胞特化,在第30天将EGF浓度降低至10ng ml-1。
内胚层模式化和前肠球体产生
DE诱导后,将细胞在含有2.0%dFBS和生长因子:WNT3A(500ng ml-1;R&D Systems),CHIR99021(2μM;Stemgent);FGF4(500ng ml-1;R&D Systems)和头蛋白(200ng ml-1;R&D Systems)的RPMI 1640培养基中培养。每天更换培养基。三天后,WNT3A(或CHIR99021),FGF4和头蛋白的组合在培养孔中导致浮动的前肠球体。为了前化前肠内胚层,在WNT/FGF/头蛋白处理的第三天加入RA(2μM;Sigma Aldrich)。
胃类器官的三维培养
将球体转移到三维体外培养系统,如先前所述的5,10,12。简言之,收集球体,重悬于50μl基质胶(BD Biosciences)中,并铺板在三维液滴中。在允许基质胶在组织培养温育箱中固化10-15分钟后,用肠培养基覆盖球形体:具有N2(Invitrogen),B27(Invitrogen),L-谷氨酰胺,10μM HEPES,青霉素/链霉素,和EGF(100ng ml-1;R&D Systems)的Advanced DMEM/F12。对于前三天,将RA和头蛋白加入到肠培养基中。在必要时,每3-4天更换培养基。在第20天,收集类器官,并在约1:12的稀释度的新鲜基质胶中重新铺板。
dox诱导型hNEUROG3hESC系的产生
为了产生过表达构建体,使用Gateway Cloning(Invitrogen)方法将hNEUROG3 cDNA(Dana-Farber/Harvard Cancer Center DNA Resource Core;克隆HsCD00345898)克隆到pInducer20慢病毒载体(来自T.Westbrook36的赠品)中。高滴度慢病毒颗粒由CCHMC Viral Vector Core产生。将H1hESC用Accutase解离,作为单细胞悬液在具有10μMY-27632的mTesR1中铺板,并暴露于慢病毒4小时。每天更换mTesR1,两天后,向培养基中加入G418(200μg ml-1)以选择整合的克隆。G418抗性细胞无限期地保持在抗生素中,但是以其它方式培养并正常传代。
iPSC系的产生和表征
原代人包皮成纤维细胞(HFF)从新生儿人包皮组织培养并通过辛迪纳大学皮肤病学系(Department of Dermatology,University of Cincinnati)从2个供体获得,并且是来自Susanne Wells博士的赠品。将HFF在由补充有10%FCS(Hyclone)的DMEM(Invitrogen)组成的成纤维细胞培养基中培养,并用于在第5代和第8代之间重编程。先前描述了用于此研究的基于EBNA1/OriP的附加型质粒pCLXE-hOct3/4-shp53,pCLXE-hSox2-Klf4,pCLXE-hLmyc-Lin28,和pCLXE-GFP37,并从Addgene(分别为ID#:27077,27078,27080和27082)获得。优化的人真皮成纤维细胞Nucleofector试剂盒(VPD-1001;Lonza)用于用附加体质粒转染HFF。简言之,对于每次转染,通过在室温下以200×g离心10分钟使1×10 6个HFF沉淀,重悬于100μl室温Nucleofector溶液中,并用1.25μg每种附加体质粒(程序U20)核转染。将来自2次转染(2x10 6总细胞)的细胞在成纤维细胞培养基中的10cm组织培养板中重新铺板,并在37℃/5%CO2下培养。转染后6天,将4.5×10 5个HFF在成纤维细胞培养基中在明胶包被的含有1.07x106个照射的小鼠胚胎成纤维细胞(MEF)的10cm皿中重新铺板。从转染后第7天开始,每天用补充有20%敲除血清替换,1mM L-谷氨酰胺,0.1mMβ-巯基乙醇,0.1mM非必需氨基酸和4ng ml-1碱性FGF(均来自Invitrogen)的DMEM/F12培养基中喂养细胞。大约2周后,手动切出具有hESC样形态的离散集落,并在用hESC量化的基质胶(Becton Dickinson)包被的组织培养皿中在mTeSR1培养基(Stem Cell Technologies)中重新铺板。在适应于mTeSR1/基质胶培养后,扩增保持强力增殖和具有最小自发分化的hESC样形态的iPSC,用于冷冻保存和表征。
通过CCHMC细胞遗传学实验室测定标准中期扩展和G带化核型。对于畸胎瘤形成,将来自6孔皿的3个孔的iPSC组合,并轻轻地重悬浮于冰冷的DMEM/F12中。在即将注射之前,加入基质胶至终浓度约33%,并将细胞皮下注射到免疫受损的NOD/SCID GAMMA C-/-小鼠中。肿瘤在6-12周内形成。固定切出的畸胎瘤,包埋在石蜡中,用苏木精和曙红染色切片用于组织学检查。
用于胃类器官的示例性方案
下表说明了用于从前体细胞形成胃类器官的示例性处理方案。**基础肠培养基=Advanced DMEM/F12+B27+N2+1-谷氨酰胺+HEPES;mTesR1可获自StemCell Technologies。
胃底特化方案
申请人首先寻求在胚胎阶段鉴定在胃底中但不在窦中特异性表达的基因。显微切割E14.5小鼠胚胎的消化道,并分成四个区域:前胃(包括食管),胃底,窦和十二指肠。参见图15。然后,通过qPCR对这些区域分析区域化的标志物。图15显示了已知在不同区域中表达的对照基因的表达。可以通过Sox2和Gata4的其高表达,以及不存在P63和Cdx2,将胃底和窦与前胃和十二指肠区分开。重要的是,Pdx1(窦的标志物)以在窦组织中比在胃底中高得多的水平表示,表明准确的解剖。
使用来自胚胎小鼠内胚层和成年人胃组织的公开的微阵列数据集的生物信息学分析来产生可以优先在胃底而不是窦中表达的候选基因的列表。通过qPCR检测这些推定的标志物在E14.5小鼠节段中的表达。Irx1,Irx2,Irx3,Irx5和Pitx1确实在胃底中比在窦中以更高的水平表达。因此,这些标志物可以用作hPSC衍生的前肠培养物中的胃底特化的指标。参见图16。
接下来,测试Wnt信号传导在胃类器官分化方案的6-9天的胃底-窦模式化的调节中的功能。添加Wnt3a(在100ng/mL和500ng/mL)对球体基因表达没有影响,但它也的确诱导Wnt靶基因Axin2的表达。因此,测试了小分子CHIR99021(CHIR;2uM)的作用。CHIR99021以受体非依赖性方式刺激Wnt信号传导。暴露于CHIR导致Pdx1表达水平的强力抑制,与胃底特化一致。CHIR不诱导肠标志物Cdx2的表达。参见图17。图18显示,与Pdx1的阻抑一致,暴露于CHIR诱导了胃底特异性标志物IRX3和IRX5的高水平表达。
幽门螺杆菌感染
在由Columbia Agar Base(Fisher Scientific),5%马血液(Colorado Serum Company),5μg ml-1,万古霉素和10μgml-1甲氧苄啶组成的血液琼脂板上培养幽门螺杆菌菌株G2738和缺少CagA(ΔCagA)39的突变体G27菌株,如先前描述40。对于类组织注射,将幽门螺杆菌以1x10 9细菌ml-1的浓度重悬于布鲁氏肉汤(brucella broth)中,并装载到Nanoject II(Drummond)微量注射器装置上。将大约200nl(含有2x10 5个细菌)直接注射到每种类器官的内腔中,并将注射的类器官培养24小时。注射布鲁氏菌肉汤作为阴性对照。
材料和方法
免疫荧光染色
将所有组织在4%多聚甲醛中在室温下固定1小时用于冷冻处理或在4℃下过夜用于石蜡处理。对于冷冻切片,将组织在30%蔗糖中在4℃下保护过夜,然后包埋在OCT(Tissue-Tek)中,并切成10μm。对于石蜡切片,通过分级乙醇系列,然后二甲苯处理组织,然后包埋在石蜡中并在7μm处切割。将组织培养细胞在室温下固定15分钟并直接染色。为了染色,将冻结的载玻片解冻至室温并在PBS中再水合,而石蜡载玻片脱蜡并进行抗原修复。将载玻片在室温下在含有0.5%Triton-X的PBS中的5%正常驴血清(Jackson Immuno Research)中封闭30分钟。将一抗(在方法表1中列出)在封闭缓冲液中稀释,并在4℃温育过夜。将载玻片在PBS中洗涤,并在室温下与二抗温育1小时,并使用Fluoromount-G(Southern Biotech)安装盖玻片。在Nikon A1Rsi倒置共焦显微镜上捕获共焦图像。
RNA分离和qPCR
使用Nucleospin RNA II试剂盒(Machery-Nagel)从组织中分离总RNA。使用Superscript VILO cDNA Synthesis Kit(Invitrogen)根据制造商的方案从100ng RNA进行逆转录。使用Quantitect SybrGreen Master Mix(Qiagen)在CFX-96实时PCR检测系统(BioRad)上进行qPCR。使用ΔΔCT方法进行分析。使用来自qPrimerDepot(http://primerdepot.nci.nih.gov)的序列设计PCR引物,并列于表2中。
免疫沉淀和Western印迹分析
在冰冷的PBS中从基质胶收获幽门螺杆菌感染的类器官,并在150g下离心5分钟。将组织在补充有蛋白酶抑制剂(Roche)的M-PER哺乳动物蛋白提取试剂(Thermo Scientific)中裂解。来自细胞裂解物的10μg总蛋白用抗c-Met抗体(2μg;Cell Signaling 4560)在4℃下免疫沉淀16小时。然后加入蛋白A/G琼脂糖珠(20μl;Santa Cruz Biotechnology),将样品在4℃下温育16小时。将免疫沉淀物在PBS中洗涤3次,然后重悬于含有β-巯基乙醇(40μl;BioRad)的Laemmli上样缓冲液中。样品在4-20%Tris-甘氨酸梯度凝胶(Invitrogen)上运行并在80V下运行3.5小时。将凝胶在105V下转移到硝酸纤维素膜(Whatman Protran,0.45μm)上1.5小时。将膜在KPL检测器封闭溶液(Kirkeaard&Perry Laboratories)中在室温下封闭1小时,然后在4℃与一抗温育过夜。使用的一抗:抗磷酸酪氨酸(Santa Cruz,sc-7020;1:100),抗c-Met(Abcam,ab59884;1:100)和抗幽门螺杆菌CagA(Abcam,ab90490;1:100)。将膜洗涤并在Alexa Fluor抗小鼠680(Invitrogen;1:100)二抗中温育。使用Odyssey红外成像软件系统(Licor)对印迹进行成像。
讨论
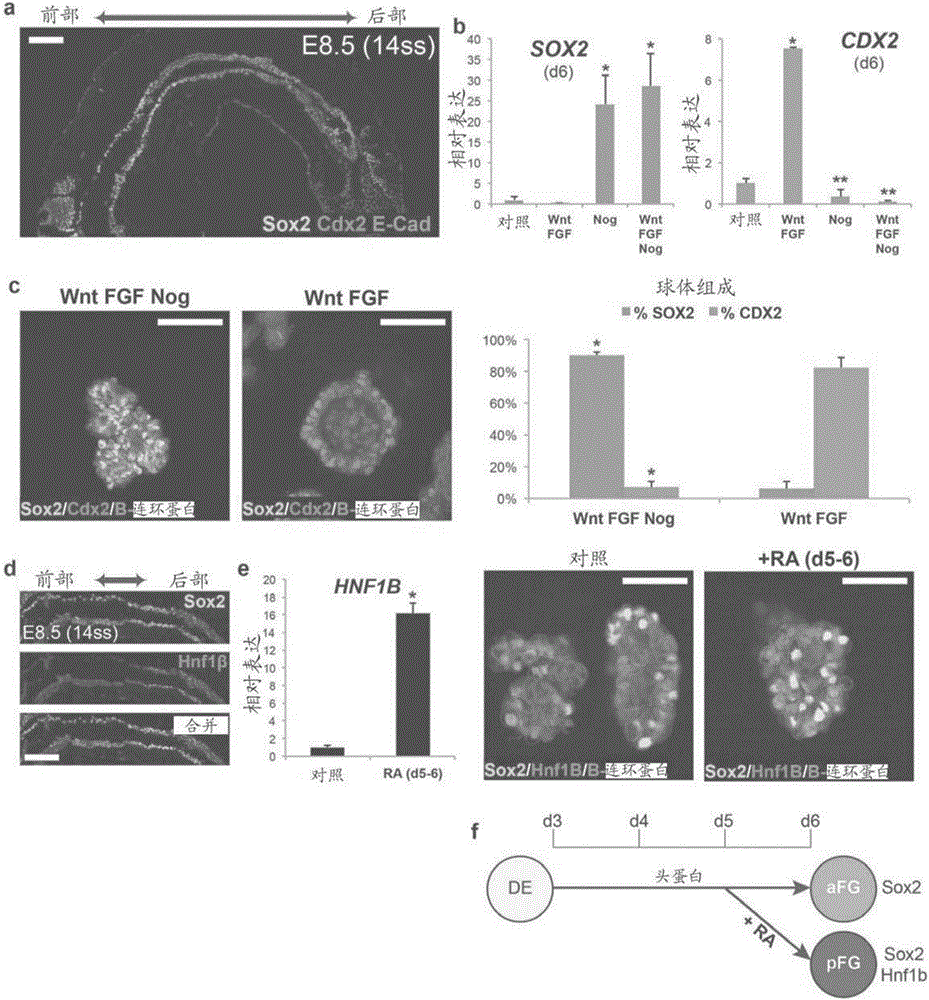
将hPSC分化成定形内胚层(DE)11,其在体内产生胃肠道和呼吸道的上皮。所有内胚层器官的发育中的下两个基本事件是沿着前部-至-后部(A-P)轴和肠管形态发生的DE模式化,导致在前部中的Sox2+前肠和在后部中的Cdx2+中-后肠的形成(如在E8.5,14体节阶段小鼠胚胎中突出显示的,图1A)。这种形态发生和内胚层和中胚层之间的组织相互作用似乎对于体内和体外二者的正确器官发生是关键的。WNT3A和FGF4先前已经证明协同作用做三件事:后化hPSC衍生的DE,促进间质的扩张,并且诱导表达中后肠标志物CDX2的肠管样结构的组装10,12。图1A显示Sox2蛋白标记前肠内胚层,并且Cdx2蛋白标记e8.5(14体节期)小鼠胚胎中的中/后肠内胚层。图1B显示抑制BMP抑制中/后肠的命运并促进前肠标志物SOX2的表达。在单独(对照)或具有指定的生长因子/拮抗剂的培养基中暴露于三天的hPSC-DE培养物中的模式化标志物的PCR分析。如先前报道的10,WNT和FGF的组合活性诱导CDX2表达,而BMP拮抗剂头蛋白抑制CDX2表达,并且足以诱导高水平的前肠标志物SOX2。*,与对照相比p<0.05。**,与WNT/FGF相比p<0.005。图1C描述了与单独用Wnt和FGF产生的具有高水平CDX2的球体相比,根据整装免疫荧光染色和mRNA,用Wnt/FGF/头蛋白产生的前肠球体具有高水平的SOX2蛋白。*,p<1.0×10-6。图1D描述了e8.5,14-体细胞阶段小鼠胚胎中的后部前肠产生胃和胰腺并且具有高水平的Hnf1β蛋白。图1E描述了在球体产生步骤的最后一天将培养物暴露于RA诱导在SOX2表达上皮中HNF1β的表达,导致后部前肠球体的形成。*,p<0.005。图1F描绘了总结在前部和后部前肠内胚层两者的形成中头蛋白和RA的模式化效应的谱系图。比例尺,100μm。误差棒表示标偏差。
表1:一抗。
表2:qPCR引物序列
为了促进hPSC衍生的DE中前肠结构的形成,申请人试图分离WNT/FGF刺激肠管形态发生的能力与其在促进后部内胚层命运中的作用。基于来自发育模型生物的体内研究13,14,测试BMP信号传导在调节A-P模式化中的功能,申请人确定WNT/FGF需要BMP活性来启动后肠程序。具体地,即使在存在WNT/FGF的情况下,用拮抗剂头蛋白抑制BMP信号传导抑制CDX2并在三天后在DE培养物中诱导前肠标志物SOX2(图1B-C和图5)。重要的是,对BMP信号传导的抑制对WNT/FGF促进间质扩展和肠管结构的装配的能力没有影响,因此导致SOX2+前肠球体的形成。
图5显示与WNT和FGF的激活平行地需要BMP信号传导以促进后部命运。图5A显示GSK3β抑制剂CHIR99021(CHIR;2μM)诱导与重组WNT3A相同的后化作用,并且这些可以被BMP抑制阻断。图5B显示CHIR诱导的肠管形态发生和球体产生以与WNT3A类似的方式发生。图5C描绘了单层培养物的免疫荧光染色,其证实了在CHIR/FGF处理的内胚层中的CDX2诱导和头蛋白和CHIR/FGF/头蛋白处理的内胚层中的SOX2诱导的高效率。图5D显示BMP靶基因MSX1/2的qPCR分析,其指示BMP活性不响应于Wnt/FGF而增加,但是靶基因响应头蛋白而抑制,表明存在内源性BMP信号传导。图5E显示加入BMP2(100ng mL-1)不代替或提升Wnt/FGF使内胚层后化的能力。这些数据指示Wnt/FGF的后化作用不是由BMP信号传导的上调介导的,但是确实需要内源性BMP活性。比例尺,图5B中的1mm;图5C中的100μm。误差棒表示标准偏差。
球体形态发生在hESC和hiPSC系二者中是强力的过程(图6A),并且>90%的球体细胞表达SOX2(图1C),指示有效规定成前肠谱系。因此,申请人已经鉴定了WNT,FGF和BMP之间的新的上位关系,其中所有三种途径协作以促进中后肠命运,而WNT和FGF与BMP分开作用以驱动内胚层和中胚层组装成肠管结构。
图2A-2G描绘了胃类器官分化是有效的和细胞系非依赖性过程。图2A,比较两个hESC系(H1和H9)和一个iPSC系(72.3)之间的球体形成和特征的表。图2B,源自H1和iPSC 72.3细胞系的第34天hGO的免疫荧光染色。iPSC衍生的类器官表现出与源自hESC的那些相同的形态和分子特征。图2C。在第34天hGO中的器官上皮细胞类型定量。大于90%的上皮是窦性的,由PDX1表达和缺乏PTF1A表达指示,而小于5%表达与源自内胚层的其它器官相关的标志物,包括CDX2(肠),白蛋白(肝)和p63(鳞状上皮)。d-g,诱导的多能干细胞系iPSC 72.3的表征。图2D,iPSC 72.3表现出与H1hESC系相比的多能干细胞集落的正常形态学特征,并且图2E具有正常的46;XY核型。图2F,iPSC 72.3表达多能标志物OCT3/4和NANOG,并且图2G,通过在体内畸胎瘤测定法中分化成内胚层,中胚层和外胚层谱系来证明多能性。比例尺,100μm。误差棒表示标准偏差。
在体内,胃的基底和窦域与胰腺,肝和十二指肠一起都源自Sox2+前肠内胚层的后段。为了将SOX2+前肠球体定向成胃谱系,申请人试图鉴定促进后部前肠命运的信号传导途径。申请人聚焦于视黄酸(RA)信号传导,鉴于其在后部前肠衍生的器官的发育中的作用。15-17在体内,后部前肠由Hnf1β的表达标记(图1D)。申请人鉴定了在模式化/球体生成阶段(FGF4/WNT3A/头蛋白)的最后一天(第5-6天)的24小时暴露于RA导致后部前肠标志物的强烈激活和SOX2/HNF1β+后部前肠球体的形成(图1E和图7)。因此,RA,WNT,FGF和BMP信号传导途径的精确时间和组合操作允许产生三维后部前肠球体。
图7A-7D显示视黄酸使前肠内胚层后化。图7A描述了前肠模式化实验的示意图。DE培养物用Wnt(CHIR)/FGF/头蛋白处理三天以产生Sox2阳性前肠球体,并且在模式化的第三天添加RA达24小时。图7B描述了显示RA增加从前肠单层培养物产生的球体数目的明视野图像。图7C描绘了图1D的低倍图像,显示了具有位于前肠后部的Hnf1β蛋白的14个体节期胚胎的免疫荧光图像。胚胎的框示区域显示在图1D中。图7D显示用RA处理的前肠球体中基因表达的qPCR分析。后部前肠标志物HNF1β和HNF6通过24小时暴露于RA而被强烈诱导。*,p<0.05。比例尺,图7B中的1mm;图7C中的100μm。误差棒表示标准偏差。
将后部前肠定向为不同器官谱系的分子机制了解甚少。在发育早期,推定的器官域由不同的基因表达模式标记:Sox2+基底,Sox2/Pdx1+窦,Pdx1/Ptf1α+胰腺和Pdx1/Cdx2+十二指肠(图2B)。申请人使用这些分子标志物来鉴定将后部前肠球体培养物定向为胃谱系的信号传导途径。在将球体转移到三维培养条件后,用RA进一步处理72小时(6-9天)导致PDX1mRNA水平>100倍的增加,同时保持高SOX2表达(图2C)。重要的是,RA处理不促进其它人9观察到的胰腺命运,因为没有诱导胰腺特异性标志物PTF1α的表达。这些数据证明RA信号传导与三维生长的组合有效地将后部前肠球体定向成指示早期窦性命运的SOX2/PDX1+上皮。
图2一般地描述了人窦胃类器官的特化和生长。误差棒表示标准偏差。图2A描绘了用于指导hPSC分化为三维胃类器官的体外培养系统的示意图,图2B描述了通过用Sox2,Pdx1和Cdx2对小鼠E10.5胚胎进行整装疫荧光染色来限定发育中的后部前肠器官的标志物。Sox2和Pdx1的共表达是对于胃上皮的远端部分是独特的,推定的窦(a),Sox2表达标记胃底(f),Pdx1(和Ptf1a)表达标记背侧(dp)和腹侧(vp)胰腺,并且Pdx1/Cdx2共表达标记十二指肠(d)。图2C显示在RA(2μM)存在下在三维基质中培养三天的后部前肠球体共表达高水平的PDX1和SOX2,并且不表达胰腺标志物PTF1α,类似于发展中的窦*p<0.05。图2D描绘了显示后部前肠球体生长成胃类器官期间形态学变化的立体显微照片。到四周,hGO的上皮展现出复杂的腺体构造,比例尺,500μm。图2E描述了E14.5和E18.5和相当的hGO发育阶段时的发育中的小鼠窦的比较。Sox2和Pdx1在小鼠窦和hGO二者中的早期假复层上皮中共表达。在后期阶段,Sox2被下调,因为上皮转变成更成熟的腺体结构。Pdx1在体内贯穿成人期和在hGO中检查的所有阶段保持在窦中,图2E中的比例尺100μm。
申请人使用SOX2/PDX1+球体来鉴定促进早期胃上皮的生长和形态发生的途径,并且发现高浓度的EGF(100ng mL-1)足以促进人窦胃类器官(hGO)的强力生长。在3-4周的过程中,直径<100μm的球体生长成直径为2-4mm的类器官。在培养的后期(约第27天),hGO上皮经历了一系列形态发生变化,使人想起胚胎胃发育的晚期,在此期间,简单的平坦的假复层上皮转变成精细的,旋绕的腺上皮(图2D)。前肠球体的初始生长依赖于EGF(数据未显示);此外,当在第27天从培养基中除去EGF时,不发生上皮扩张和形态发生成腺体(图8)。这些结果支持已公布的发现,其指示EGF在促进胃粘膜的适当生长中的重要作用19,20。
图8显示EGF是胃类器官中腺体形态发生所需的。明视野图像和免疫染色证明了在hGO分化的晚期阶段上皮形态发生和腺形成需要EGF。当在第27天从生长培养基中除去EGF时,在腺形态发生之前,hGO上皮保持了不能形成腺体的简单的立方结构。比例尺,100μm。
hGO生长与胚胎小鼠胃发育的比较揭示了hGO发育与体内胃器官发生惊人地相似。在早期阶段(小鼠中的E12-14和13天hGO),这两种上皮都是假复层的,并且包含集中朝向腔面的有丝分裂细胞(图9和图10),指示相互动力核迁移过程(interkinetic nuclear migration process)21。早期hGO是适当地极化的,并且包含由顶端标志物aPKC22的表达描绘轮廓的次级腔(secondary lumina)(图10)。
在E16.5和产后早期阶段之间,窦转化成表现出由腺体和凹陷组成的高度结构化构造的简单柱状上皮(图2E和图9)。在体外13和34天之间,hGO上皮经历类似的转变以形成具有类似于晚期胎儿窦的腺状结构的高柱状上皮(图2E)。对转录因子Sox2,Pdx1,Gata4和Klf5的表达的分析揭示在体内和体外伴随这些形态发生过程的定型(stereotypic)时空表达模式(图9)。在早期阶段,这些因子都在未成熟的假复层上皮中共表达。然而,在后期阶段,Sox2表达下调,因为上皮形成早期腺体和凹陷,而其它因素的表达无限期地维持。基于这些数据,估计13天hGO代表类似于E12-14小鼠窦的发育阶段,而34天hGO更相当于晚期胎儿早期产后窦。此外,推断hGO重演正常胚胎发育,并且发生在窦发育期间的分子和形态发生过程在啮齿类和人类之间是保守的。
图9显示在小鼠窦和人胃类器官发育期间转录因子表达的比较。分析了体内窦发育的四个胚胎期(E12.5,E14.5,E16.5和E18.5)和一个出生后阶段(P12)的转录因子表达:Sox2,Pdx1,Gata4,Klf5和FoxF1。在体外hGO发育的两个阶段(第13天和第34天)分析相同的标志物,并且揭示类器官发育与在体内发生的事情平行。在窦发育的早期阶段,上皮标志物Sox2遍在表达,但在后期阶段时下调,而其它上皮转录因子Pdx1,Gata4和Klf5在整个发育中表现出持续表达。早期和晚期hGO二者都包含围绕上皮的FoxF1阳性间质细胞。比例尺,100μm。图10显示早期阶段人胃类器官表现出定型结构和核行为。在13天,hGO含有显示由顶端标志物aPKC和基底外侧标志物E-钙粘蛋白标记的顶基部(apicobasal)极性的假复层上皮细胞,类似于E12.5小鼠窦。此外,在类器官上皮内看到衬里为顶膜的次级腔(白色箭头)。E12.5小鼠窦和第7天hGO两者似乎仅在细胞的顶端部分中经历相互动力学核迁移,其由有丝分裂核,pHH3的存在指示。比例尺,50μm。
前肠球体含有类似于先前描述10的中后肠球体的间质组分。在分化为胃类器官组织期间,间质扩展并表达与窦间质发育相关的关键转录因子,包括FOXF1和BAPX1(图10和图11)。在后期阶段,hGO间质主要由VIMENTIN+粘膜下成纤维细胞和较少数量的ACTA2+上皮下肌纤维母细胞(指示未成熟的胃间质)组成(图11)。hGO不形成如在体内发生的平滑肌的分化层。鉴于在除了EGF之外没有任何外源因子的情况下存在这种强力的上皮形态发生,似乎可能的是间质在上皮发育中起作用。因此,令人惊讶的是,上皮不表现出促进间质的强力分化。这提示其它刺激物(可能是机械的)在胃间质分化中发挥作用。
图11显示胃类器官中的间质分化。图11A显示窦间质转录因子BAPX1的时间表达分析。类似于其已知的胚胎表达模式,BAPX1是在hGO分化的早期阶段期间上调,然后下调,与功能细胞类型标志物表达一致。图11B显示间质细胞类型标志物的染色揭示第34天hGO含有FOXF1/波形蛋白阳性粘膜下成纤维细胞和少量表达波形蛋白/ALPHA-SM-ACTIN(SMA)的上皮下成纤维细胞。hGO缺乏稳定的平滑肌层,其由体内窦中的SMA/结蛋白阳性细胞表示。比例尺,100μm。误差棒表示标准偏差。
在窦中发现的主要功能细胞类型是粘液细胞,其分泌作为胃上皮衬里的保护性粘液层和分泌激素以调节胃肠生理学和代谢稳态的内分泌细胞24。到第34天,hGO含有将粘液分泌到管腔中并具有与其体内对应物相同的高柱状形态的表面粘液细胞(MUC5AC/UEAI+)。hGO还含有TFF2/GSII+窦腺细胞,指示在窦粘液谱系中的适当分化(图3A)。此外,hGO形成由基本定位的增殖和SOX9表达区域指示的祖细胞小生境(图4A),尽管上皮的增殖指数是可变的并且在1-10%之间变化。因此,体外hGO含有包含祖细胞和分化细胞类型二者的生理性胃上皮。
图4显示人胃类器官表现出对幽门螺杆菌感染的急性响应。图4A显示第28天hGO含有以Ki67标记的增殖细胞和限制到早期腺体底部的SOX9+祖细胞,类似于晚期胚胎和出生后小鼠窦。图4B显示hGO用于对幽门螺杆菌感染的人特异性疾病过程建模。将细菌显微注射到hGO的腔中,并且通过明视野显微术(黑色箭头)和免疫荧光染色在注射后24小时在腔中显现细菌。图4C描述了癌基因c-Met的免疫沉淀,并且证明幽门螺杆菌诱导c-Met的强力激活(酪氨酸磷酸化),并且这是CagA依赖性过程。此外,CagA直接与人类胃上皮细胞中的c-Met相互作用。图4D显示在24小时内,幽门螺杆菌感染导致hGO上皮中的增殖细胞数量增加两倍,其通过EdU掺入测量。*,p<0.05。比例尺,a中100μm;b中25μm。误差棒表示。
图3证明人胃类器官组织含有正常分化的窦细胞类型,并且可用于对人胃发育建模。图3A证明hGO含有所有主要的窦细胞谱系。34天hGO具有表面粘液细胞(Muc5AC)和粘液腺细胞(TFF2),以及凝集素染色区分表面粘液,UEAI和粘液腺细胞GSII。hGO还包含如由染色颗粒素(Chromogranin)A(CHGA)标记的内分泌细胞。图3B是在hGO发育期间EGF在生长,形态发生和细胞类型特化中的不同作用的示意图。在早期发育阶段需要高水平的EGF以形成腺体,然而其在发育的晚期抑制内分泌分化;因此,到第30天,EGF浓度降低以允许内分泌细胞发育。图3C显示,在撤去包括胃泌素,生长素释放肽和血清素(5-HT)的EGF时,所有主要内分泌激素在hGO中表达。图3D显示高水平的EGF抑制NEUROG3表达。在第30天EGF浓度的降低导致在第34天通过qPCR测量的NEUROG3表达的显著增加,指示EGF在内分泌特化中作用于NEUROG3的上游。*,p<0.05。图3E显示NEUROG3在EGF下游作用以诱导内分泌细胞命运。使用dox诱导性系统的NEUROG3的强制表达足以超越高EGF(100ng mL-1)的内分泌抑制作用。hGO在第30天暴露于dox(1μg/mL)24小时,并在第34天进行分析。经Dox处理的类器官表现出对表达ChrA的内分泌细胞的强烈诱导。比例尺,100μm。误差棒表示标准偏差。
在34天hGO中还存在丰富的染色颗粒素-A(CHGA)+内分泌细胞,包括表达胃泌素,生长素释放肽,生长抑素和血清素的窦中的四种主要内分泌细胞类型(图3C和图12)。有趣的是,我们观察到高水平的EGF抑制内分泌细胞形成,使得100ng ml-1导致每个类器官的<1个内分泌细胞。相反,从第30-34天在较低水平的EGF(10ng ml-1)中培养的hGO产生大量的内分泌细胞(图13)。此外,高EGF还抑制促内分泌转录因子NEUROG3的表达(图3D),其在胰腺和肠中广泛研究25-28并且是大多数胃内分泌谱系的形成中所需要的29,30。这些数据提示EGFR信号传导在NEUROG3上游的胃内分泌细胞特化中的新的抑制作用。为了测试该模型,申请人使用多西环素诱导型hNEUROG3过表达hESC系,并且发现NEUROG3表达足以克服高EGF(100ng ml-1)的内分泌抑制作用,导致CHGA+内分泌细胞的强力形成(图3E和图13)。从这些发现,我们得出结论,EGF通过抑制NEUROG3抑制内分泌祖细胞的形成,并且NEUROG3足以用于人胃内分泌细胞的特化。
图12显示了体内胃窦内分泌细胞发育。窦中的内分泌细胞分化首先在E18.5明显,但在出生后阶段更明确(显示的P12)。在早期阶段,所有预期的胃内分泌亚型是明显的,包括胃泌素,生长素释放肽,生长抑素和血清素(5-HT)。比例尺,100μm。图13显示EGF信号传导抑制NEUROG3依赖性胃内分泌特化程序。图13A显示维持在高浓度的EGF(100ng mL-1)中的hGO在第34天具有非常少的内分泌细胞,其通过泛内分泌标志物CHGA的染色显示。在第24天EGF浓度(10ng mL-1)的减少导致胃上皮中内分泌细胞的更多生理数目。图13B显示从用dox诱导型NEUROG3过表达转基因稳定转染的hESC系产生hGO,以测试NEUROG3上游是否发生内分泌分化的EGF阻遏。hGO维持在高EGF(100ng mL-1)中,然后在第30天用多西环素(1μg/mL-1)处理24小时,然后在第34天进行分析。经Dox处理的hGO显示内分泌标志物CHGA,胃泌素,生长素释放肽和促生长素抑制素,并且它们含有具有内分泌形态的CHGA-(图3A),生长素释放肽-和促生长素抑制素阳性细胞。*,p<0.05。比例尺,100μm。误差棒表示标准偏差。
临床证据指示窦的主要定植在幽门螺杆菌介导的疾病中具有重要作用31,32。因此,申请人测试了hGO是否可用于对人胃对病原体幽门螺杆菌的病理生理响应建模。为了模拟正常的宿主-病原体界面,我们通过显微注射到类器官的腔中直接将幽门螺杆菌引入上皮的腔表面并测量上皮信号传导和增殖(图4)。通过免疫荧光观察细菌与hGO上皮紧密结合(图4B)。在24小时内,申请人观察到对幽门螺杆菌的显著上皮应答,包括胃癌基因c-Met33的强力激活和上皮细胞增殖的2倍增加。幽门螺杆菌毒力因子CagA在疾病的病因中起关键作用。与已发表的研究34一致,申请人证明CagA移位到类器官上皮细胞中并与c-Met形成复合物(图4C)。此外,当用缺乏CagA的幽门螺杆菌的非致病性菌株注射hGO时,上皮应答被消除,增强了该因子在幽门螺杆菌介导的人类发病机理中的重要性。因此,hGO对幽门螺杆菌的病理生理应答使它们成为阐明由幽门螺杆菌介导的人胃疾病的起始事件的前所未有的模型。
其它参考文献
1.Wen,S.&Moss,S.F.Helicobacter pylori virulence factors in gastric carcinogenesis.Cancer Lett.282,1–8(2009).
2.Yuan,Y.,Padol,I.T.&Hunt,R.H.Peptic ulcer disease today.Nat Clin Pract Gastroenterol Hepatol 3,80–89(2006).
3.Parkin,D.M.The global health burden of infection-associated cancers in the year 2002.Int.J.Cancer 118,3030–3044(2006).
4.Peek,R.M.Helicobacter pylori infection and disease:from humans to animal models.Dis Model Mech 1,50–55(2008).
5.Barker,N.et al.Lgr5(+ve)stem cells drive self-renewal in the stomach and build long-lived gastric units in vitro.Cell Stem Cell 6,25–36(2010).
6.Longmire,T.A.et al.Efficient Derivation of Purified Lungand Thyroid Progenitors from Embryonic Stem Cells.Stem Cell 10,398–411(2012).
7.Mou,H.et al.Generation of Multipotent Lung and Airway Progenitors from Mouse ESCs and Patient-Specific Cystic Fibrosis iPSCs.Stem Cell 10,385–397(2012).
8.Si-Tayeb,K.et al.Highly efficient generation of human hepatocyte-like cells from induced pluripotent stem cells.Hepatology 51,297–305(2010).
9.D'Amour,K.A.et al.Production of pancreatic hormone-expressing endocrine cells from human embryonic stem cells.Nat Biotechnol 24,1392–1401(2006).
10.Spence,J.R.et al.Directed differentiation of human pluripotent stem cells into intestinal tissue in vitro.Nature 470,105–109(2011).
11.D'Amour,K.A.et al.Efficient differentiation of human embryonic stem cells to definitive endoderm.Nat Biotechnol 23,1534–1541(2005).
12.McCracken,K.W.,Howell,J.C.,Spence,J.R.&Wells,J.M.Generating human intestinal tissue from pluripotent stem cells in vitro.Nature Protocols 6,1920–1928(2011).
13.Kumar,M.,Jordan,N.,Melton,D.&Grapin-Botton,A.Signals from lateral plate mesoderm instruct endoderm toward a pancreatic fate.Dev Biol 259,109–122(2003).
14.Tiso,N.,Filippi,A.,Pauls,S.,Bortolussi,M.&Argenton,F.BMP signalling regulates anteroposterior endoderm patterning in zebrafish.Mech Dev 118,29–37(2002).
15.Wang,Z.,Dollé,P.,Cardoso,W.V.&Niederreither,K.Retinoic acid regulates morphogenesis and patterning of posterior foregut derivatives.Dev Biol 297,433–445(2006).
16.Martín,M.et al.Dorsal pancreas agenesis in retinoic acid-deficient Raldh2mutant mice.Dev Biol 284,399–411(2005).
17.Molotkov,A.,Molotkova,N.&Duester,G.Retinoic acid generated by Raldh2 in mesoderm is required for小鼠dorsal endodermal pancreas development.Dev Dyn 232,950–957(2005).
18.Kawaguchi,Y.et al.The role of the transcriptional regulator Ptf1a in converting intestinal to pancreatic progenitors.Nat Genet 32,128–134(2002).
19.Johnson,L.R.&Guthrie,P.D.Stimulation of rat oxyntic gland mucosal growth by epidermal growth factor.Am.J.Physiol.238,G45–9(1980).
20.Majumdar,A.P.Postnatal undernutrition:effect of epidermal growth factor on growth and function of the gastrointestinal tract in rats.J.Pediatr.Gastroenterol.Nutr.3,618–625(1984).
21.Spear,P.C.&Erickson,C.A.Interkinetic nuclear migration:A mysterious process in search of a function.Develop.Growth Differ.54,306–316(2012).
22.Grosse,A.S.et al.Cell dynamics in fetal intestinal epithelium:implications for intestinal growth and morphogenesis.Development 138,4423–4432(2011).
23.Verzi,M.P.et al.Role of the homeodomain transcription factor Bapx1 in小鼠distal stomach development.Gastroenterology 136,1701–1710(2009).
24.Mills,J.C.&Shivdasani,R.A.Gastric Epithelial Stem Cells.Gastroenterology 140,412–424(2011).
25.Gradwohl,G.,Dierich,A.,LeMeur,M.&Guillemot,F.neurogenin3 is required for the development of the four endocrine cell lineages of the pancreas.Proc Natl Acad Sci USA 97,1607–1611(2000).
26.Jenny,M.et al.Neurogenin3is differentially required for endocrine cell fate specification in the intestinal and gastric epithelium.EMBO J 21,6338–6347(2002).
27.Johansson,K.A.et al.Temporal control of neurogenin3 activity in pancreas progenitors reveals competence windows for the generation of different endocrine cell types.Dev Cell 12,457–465(2007).
28.López-Díaz,L.et al.Intestinal Neurogenin 3 directs differentiation of a bipotential secretory progenitor to endocrine cell rather than goblet cell fate.Dev Biol 309,298–305(2007).
29.Schonhoff,S.E.,Giel-Moloney,M.&Leiter,A.B.Neurogenin 3-expressing progenitor cells in the gastrointestinal tract differentiate into both endocrine and non-endocrine cell types.Dev Biol 270,443–454(2004).
30.Lee,C.S.,Perreault,N.,Brestelli,J.E.&Kaestner,K.H.Neurogenin 3 is essential for the proper specification of gastric enteroendocrine cells and the maintenance of gastric epithelial cell identity.Genes Dev 16,1488–1497(2002).
31.Olbe,L.,Hamlet,A.,J.&L.A mechanism by which Helicobacter pylori infection of the antrum contributes to the development of duodenal ulcer.Gastroenterology 110,1386–1394(2001).
32.Xia,H.H.et al.Antral-type mucosa in the gastric incisura,body,and fundus(antralization):a link between Helicobacter pylori infection and intestinal metaplasia?Am.J.Gastroenterol.95,114–121(2000).
33.Churin,Y.et al.Helicobacter pylori CagA protein targets the c-Met receptor and enhances the motogenic response.J.Cell Biol.161,249–255(2003).
34.Peek,R.M.et al.Helicobacter pylori cagA+strains and dissociation of gastric epithelial cell proliferation from apoptosis.J.Natl.Cancer Inst.89,863–868(1997).
35.Teo,A.K.K.et al.Activin and BMP4 Synergistically Promote Formation of Definitive Endoderm in Human Embryonic Stem Cells.Stem Cells 30,631–642(2012).
36.Meerbrey,K.L.et al.The pINDUCER lentiviral toolkit for inducible RNA interference in vitro and in vivo.Proc Natl Acad Sci USA 108,3665–3670(2011).
37.Okita,K.et al.An efficient nonviral method to generate integration-free human-induced pluripotent stem cells from cord blood and peripheral blood cells.Stem Cells 31,458–466(2013).
38.Covacci,A.et al.Molecular characterization of the 128-kDa immunodominant antigen of Helicobacter pylori associated with cytotoxicity and duodenal ulcer.Proc Natl Acad Sci USA 90,5791–5795(1993).
39.Amieva,M.R.,Salama,N.R.,Tompkins,L.S.&Falkow,S.Helicobacter pylori enter and survive within multivesicular vacuoles of epithelial cells.Cell.Microbiol.4,677–690(2002).
40.Schumacher,M.A.et al.Gastric Sonic Hedgehog acts as a macrophage chemoattractant during the immune response to Helicobacter pylori.Gastroenterology 142,1150–1159.e6(2012).
- 还没有人留言评论。精彩留言会获得点赞!
- 一种用于外阴鳞状细胞癌中药制剂及其制备方法
- miR-455-3p在食管鳞状细胞癌中的诊断、治疗和预后的应用
- Eif5a2在制备食管鳞状细胞癌预后诊断试剂中的应用的制作方法
- 粘纤网球食管脱落细胞采集器的制作方法
- 一种新的多肽——人细胞分化转录因子13.7和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——人细胞分化转录因子10和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——人细胞分化转录因子11和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——人细胞分化转录因子14和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——人细胞分化转录因子11.1和编码这种多肽的多核苷酸的制作方法
- 一种新的多肽——人细胞分化转录因子58和编码这种多肽的多核苷酸的制作方法