使用工程化全细胞催化剂生产无热量甜味剂的制作方法
支持提交序列表的声明
本文提供了序列表的纸质复印件和含有命名为“32559_63_1.txt”,大小为36,234字节(正如在microsoft
公开背景
本公开总体上涉及天然甜味剂。更具体地,本公开涉及无热量甜味剂及合成无热量甜味剂的方法。
甜菊醇糖苷是从甜叶菊(steviarebaudiana)叶片分离的天然产物。甜菊醇糖苷广泛地用作高强度、低热量的甜味剂并且明显比蔗糖更甜。作为天然甜味剂,不同的甜菊醇葡糖苷具有不同的甜度和后味。甜菊醇糖苷的甜度明显高于蔗糖的甜度。例如,甜菊苷比蔗糖甜100-150倍,具有苦的后味。莱鲍迪苷c(rebaudiosidec)比蔗糖甜40-60倍。杜尔可苷a(dulcosidea)比蔗糖甜约30倍。
天然存在的甜菊醇糖苷共有相同的基本甜菊醇结构,但是在c13和c19位置的碳水化合物残基(例如,葡萄糖、鼠李糖和木糖残基)的含量上不同。具有已知结构的甜菊醇糖苷包括,甜菊醇、甜菊苷、莱鲍迪苷a、莱鲍迪苷b、莱鲍迪苷c、莱鲍迪苷d、莱鲍迪苷e、莱鲍迪苷f和杜尔可苷a(参见例如,表1)。其它甜菊醇糖苷为莱鲍迪苷m、莱鲍迪苷n和莱鲍迪苷o。
表1.甜菊醇糖苷。
按干重计,甜菊苷、莱鲍迪苷a、莱鲍迪苷c和杜尔可苷a分别占叶中甜菊醇糖苷总重量的9.1、3.8、0.6和0.3%,而其它甜菊醇糖苷以低得多的量存在。来自甜叶菊植株的提取物可商购,通常含有作为主要化合物的甜菊苷和莱鲍迪苷a。其它甜菊醇糖苷在甜叶菊提取物中作为微量组分存在。例如,商用制剂中莱鲍迪苷a的量可以在总甜菊醇糖苷含量的约20%至大于90%变化,而莱鲍迪苷b的量可为总甜菊醇糖苷的约1-2%,莱鲍迪苷c的量可为总甜菊醇糖苷的约7-15%,且莱鲍迪苷d的量可为总甜菊醇糖苷的约2%。
大多数甜菊醇糖苷是使用尿苷5’-二磷酸葡萄糖(udp-葡萄糖)作为糖部分的供体,通过通常由udp-糖基转移酶(ugt)催化的甜菊醇的几个糖基化反应形成的,如图1a、图1b、图1c、图1d和图1e中所示。植物中的ugt构成了酶的非常不同的类,所述酶将葡萄糖残基从udp-葡萄糖转移到甜菊醇。例如,甜菊苷的c-13-o-葡萄糖的c-3’的糖基化产生莱鲍迪苷a;且甜菊苷的19-o-葡萄糖的c-2’的糖基化产生莱鲍迪苷e。进一步地莱鲍迪苷a(在c-2’-19-o-葡萄糖处)或莱鲍迪苷e(在c-3’-13-o-葡萄糖处)的糖基化生成莱鲍迪苷d。莱鲍迪苷d(在c-3’-19-o-葡萄糖处)的糖基化生成莱鲍迪苷m。
替代性甜味剂正受到越来越多的关注,因为意识到许多疾病与高糖食品和饮料的消耗相关。虽然人造甜味剂可用,但许多人造甜味剂如甘素(dulcin)、环己氨基磺酸钠和糖精由于对其安全性的考虑已被一些国家禁止或限制。因此,天然来源的无热量甜味剂变得越来越受欢迎。广泛使用甜叶菊甜味剂的主要障碍之一是其不良的口味属性。因此,需要开发替代性甜味剂及其生产方法以提供甜味潜能和风味性质的最佳组合。
发明概述
本公开总体上涉及天然甜味剂。更具体地,本公开涉及无热量甜味剂和合成无热量天然甜味剂的方法。
全细胞催化剂。一方面,本公开涉及至少一种全细胞催化剂。在这个方面中,所述至少一种全细胞催化剂是包括至少一个编码至少一种酶的核苷酸序列的转化宿主细胞,所述至少一种酶包括但不限于尿苷二磷酸糖基转移酶(udp-糖基转移酶)和蔗糖合成酶(sus)。合适的尿苷二磷酸糖基转移酶的非限制性实例包括:ugt76g1、hv1、eugt11及其任何组合。
在这个方面中,转化宿主细胞可以是通过包括至少一个编码至少一种酶的核苷酸序列的至少一个表达盒转化的任何合适宿主细胞。合适的宿主细胞的非限制性实例包括细菌、酵母、丝状真菌、蓝细菌藻和植物细胞。一方面,选择所述至少一个表达盒以与宿主细胞类型相容,并且编码所述至少一种酶的所述至少一个核苷酸序列内的密码子可以使用已知方法进行密码子优化以便增强所述至少一种酶在选定的宿主细胞类型中的表达。
合成甜菊醇糖苷的方法。另一方面,所述至少一种全细胞催化剂可用于合成如本文所述的至少一种甜菊醇糖苷的方法中。所述方法包括在包括底物的培养基中体外培养所述至少一种全细胞催化剂。不受任何特定理论的限制,由转化宿主细胞内的所述至少一个表达盒生成的所述至少一种酶酶促糖基化培养基内的底物以合成所需的至少一种甜菊醇糖苷。
全细胞催化剂巴斯德毕赤酵母(pichiapastoris)菌株。另一方面,本公开涉及一种全细胞催化剂,其包括通过含至少一个核苷酸序列的至少一个表达盒转化的巴斯德毕赤酵母宿主细胞,所述至少一个核苷酸序列包括:编码融合蛋白的第一核苷酸序列,其包括编码毕赤酵母(pichia)细胞壁蛋白的第一核苷酸片段,第一核苷酸片段插入编码udp-糖基转移酶的第二核苷酸片段c端的框内;和编码蔗糖合成酶的第二核苷酸序列。转化巴斯德毕赤酵母细胞的所述至少一个核苷酸序列可表达以生成全细胞催化剂,其特征在于一定量展示在巴斯德毕赤酵母细胞的细胞表面上的udp-糖基转移酶和一定量的细胞内蔗糖合成酶。
转化巴斯德毕赤酵母一方面可包括所述至少一个核苷酸序列中的每一个的一个或多个拷贝。第一巴斯德毕赤酵母菌株(g/k4s2)包括:四个拷贝的含第一核苷酸序列的表达盒,第一核苷酸序列包括插入编码udp-糖基转移酶(ugt76g1)的第二核苷酸片段c端的框内的毕赤酵母细胞壁蛋白(gcw61);和两个拷贝的含编码蔗糖合成酶(mbsus1)的第二核苷酸序列的表达盒。第二巴斯德毕赤酵母菌株(g/h5s2)包括:五个拷贝的含第一核苷酸序列的表达盒,第一核苷酸序列包括插入编码udp-糖基转移酶(hv1)的第二核苷酸片段c端的框内的毕赤酵母细胞壁蛋白(gcw61);和两个拷贝的含编码蔗糖合成酶(mbsus1)的第二核苷酸序列的表达盒。
由甜菊苷生成莱鲍迪苷a的方法。另一方面,本公开涉及一种由甜菊苷合成莱鲍迪苷a的方法。所述方法包括在包括甜菊苷底物的培养基中培养全细胞催化剂足够的时间以生成莱鲍迪苷a,其中葡萄糖与甜菊苷共价偶联以生成莱鲍迪苷a。全细胞催化剂是表达一定量的udp-糖基转移酶(ugt76g1)和一定量的蔗糖合成酶(mbsus1)的转化宿主细胞。一方面,全细胞催化剂可以是巴斯德毕赤酵母的g/k4s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp-葡萄糖、udp及其任何组合。
由莱鲍迪苷d生成莱鲍迪苷m的方法。一方面,本公开涉及一种由莱鲍迪苷d合成莱鲍迪苷m的方法。所述方法包括用包括莱鲍迪苷d底物的培养基培养全细胞催化剂足够的时间以生成莱鲍迪苷m,其中葡萄糖与莱鲍迪苷d共价偶联以生成莱鲍迪苷m。全细胞催化剂是表达一定量的udp-糖基转移酶(ugt76g1)和一定量的蔗糖合成酶(mbsus1)的转化宿主细胞。一方面,全细胞催化剂可以是巴斯德毕赤酵母的g/k4s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp-葡萄糖、udp及其任何组合。
由莱鲍迪苷e生成莱鲍迪苷d的方法。另一方面,本公开涉及一种由莱鲍迪苷e合成莱鲍迪苷d的方法。所述方法包括用包括莱鲍迪苷e底物的培养基培养全细胞催化剂足够的时间以生成莱鲍迪苷d,其中葡萄糖与莱鲍迪苷e共价偶联以生成莱鲍迪苷d。全细胞催化剂是表达一定量的udp-糖基转移酶(ugt76g1)和一定量的蔗糖合成酶(mbsus1)的转化宿主细胞。一方面,全细胞催化剂可以是巴斯德毕赤酵母的g/k4s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp-葡萄糖、udp及其任何组合。
由莱鲍迪苷a生成莱鲍迪苷d的方法。另一方面,本公开涉及一种由莱鲍迪苷a合成莱鲍迪苷d的方法。所述方法包括用包括莱鲍迪苷a底物的培养基培养全细胞催化剂足够的时间以生成莱鲍迪苷d,其中葡萄糖与莱鲍迪苷a共价偶联以生成莱鲍迪苷d。全细胞催化剂是表达一定量的udp-糖基转移酶(hv1-gcw61)和一定量的蔗糖合成酶(mbsus1)的转化宿主细胞。一方面,全细胞催化剂可以是巴斯德毕赤酵母的g/h5s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp-葡萄糖、udp及其任何组合。
由甜菊苷生成莱鲍迪苷e的方法。另一方面,本公开涉及一种由甜菊苷底物合成莱鲍迪苷e的方法。所述方法包括用包括甜菊苷底物的培养基培养全细胞催化剂足够的时间以生成莱鲍迪苷e,其中葡萄糖与甜菊苷共价偶联以生成莱鲍迪苷e。全细胞催化剂是表达一定量的udp-糖基转移酶(hv1-gcw61)和一定量的蔗糖合成酶(mbsus1)的转化宿主细胞。一方面,全细胞催化剂可以是巴斯德毕赤酵母的g/h5s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp-葡萄糖、udp及其任何组合。
由甜菊苷和/或莱鲍迪苷a生成莱鲍迪苷m的方法。另一方面,本公开涉及一种由甜菊苷和/或莱鲍迪苷a合成莱鲍迪苷m的方法。所述方法包括用包括甜菊苷和/或莱鲍迪苷a底物的培养基培养全细胞催化剂足够的时间以生成莱鲍迪苷m。全细胞催化剂包括:表达一定量的udp-糖基转移酶(ugt76g1)和一定量的蔗糖合成酶(mbsus1)的第一转化宿主细胞;和表达一定量的udp-糖基转移酶(hv1-gcw61)和一定量的蔗糖合成酶(mbsus1)的第二转化宿主细胞。一方面,全细胞催化剂可以是巴斯德毕赤酵母的g/k4s2和g/h5s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp-葡萄糖、udp及其任何组合。
由莱鲍迪苷ka生成莱鲍迪苷e的方法。另一方面,本公开涉及一种由莱鲍迪苷ka合成莱鲍迪苷e的方法。所述方法包括用包括莱鲍迪苷ka底物的培养基培养全细胞催化剂足够的时间以生成莱鲍迪苷e,其中葡萄糖与莱鲍迪苷ka共价偶联以生成莱鲍迪苷e。全细胞催化剂是表达一定量的udp-糖基转移酶(hv1-gcw61)和一定量的蔗糖合成酶(mbsus1)的转化宿主细胞。一方面,全细胞催化剂可以是巴斯德毕赤酵母的g/h5s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp-葡萄糖、udp及其任何组合。
由甜茶苷生成莱鲍迪苷ka和莱鲍迪苷e的方法。另一方面,本公开涉及一种由甜茶苷合成莱鲍迪苷ka的方法。所述方法包括用包括甜茶苷底物的培养基培养全细胞催化剂足够的时间以生成莱鲍迪苷ka,其中葡萄糖与甜茶苷共价偶联以生成莱鲍迪苷ka,葡萄糖继续与莱鲍迪苷ka共价偶联以生成莱鲍迪苷e。全细胞催化剂是表达一定量的udp-糖基转移酶(hv1-gcw61)和一定量的蔗糖合成酶(mbsus1)的转化宿主细胞。一方面,全细胞催化剂可以是巴斯德毕赤酵母的g/h5s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp-葡萄糖、udp及其任何组合。
附图简述
本公开在考虑到以下对其的详述时将更好理解,并且优于以上所提到的那些的特征、方面和优势将变得显而易见。此详述参考以下附图,其中:
图1a、图1b、图1c、图1d和图1e描绘自甜菊醇开始的甜菊醇糖苷生物合成途径。
图2a、图2b和图2c汇总了使用培养的g/k4s2细胞由甜菊苷生成莱鲍迪苷a(“reba”)。图2a和图2b分别汇总了甜菊苷(图2a)和莱鲍迪苷a(图2b)标准品的hplc保留时间。图2c汇总了在将诱导的g/k4s2细胞引入体外反应体系后24小时从g/k4s2培养基中取样,由诱导的g/k4s2细胞酶促生成的莱鲍迪苷a的hplc保留时间。
图3a、图3b和图3c汇总了使用培养的g/k4s2细胞由莱鲍迪苷d(“rebd”)生成莱鲍迪苷m(“rebm”)。图3a和图3b分别汇总了莱鲍迪苷d(图3a)和莱鲍迪苷m(图3b)标准品的hplc保留时间。图3c汇总了在将诱导的g/k4s2细胞引入体外反应体系后24小时从g/k4s2培养基中取样,由诱导的g/k4s2细胞酶促生成的莱鲍迪苷m和剩余莱鲍迪苷d的hplc保留时间。
图4a、图4b和图4c汇总了使用培养的g/k4s2细胞由莱鲍迪苷e(“rebe”)生成莱鲍迪苷d(“rebd”)。图4a和图4b分别汇总了莱鲍迪苷e(图4a)和莱鲍迪苷d(图4b)标准品的hplc保留时间。图4c汇总了在将诱导的g/k4s2细胞引入体外反应体系后24小时从g/k4s2培养基中取样,由诱导的g/k4s2细胞酶促生成的莱鲍迪苷d和剩余莱鲍迪苷e的hplc保留时间。
图5a、图5b、图5c、图5d和图5e汇总了使用培养的g/h5s2细胞由莱鲍迪苷a(“reba”)生成莱鲍迪苷d(“rebd”)。图5a和图5b分别汇总了莱鲍迪苷a(图5a)和莱鲍迪苷d(图5b)标准品的hplc保留时间。图5c汇总了在将g/h5s2细胞引入体外反应体系后24小时从培养基中取样,由培养的g/h5s2细胞酶促生成的莱鲍迪苷d和剩余莱鲍迪苷a的hplc保留时间。图5d汇总了在将诱导的h5细胞引入体外反应体系后24小时从h5培养基中取样,由培养的其中引入了仅含hv1-gcw61序列的载体的巴斯德毕赤酵母菌株(h5)酶促生成的莱鲍迪苷d,以及剩余莱鲍迪苷a的hplc保留时间;h5培养基中莱鲍迪苷d的量减少表明hv1-gcw61和mbsus1两种基因均参与reba通过g/h5s2细胞向rebd的转化。图5e汇总了在将诱导的phka细胞引入体外反应体系后24小时从phka培养基中取样,由培养的其中并入了缺乏hv1-gcw61和mbsus1序列两者的空载体的巴斯德毕赤酵母菌株(phka)酶促生成的莱鲍迪苷d,以及剩余莱鲍迪苷a的hplc保留时间;phka培养基中缺乏莱鲍迪苷d进一步表明hv1-gcw61和mbsus1序列两者均参与reba通过g/h5s2细胞向rebd的转化。
图6a、图6b、图6c和图6d汇总了使用培养的g/h5s2细胞由甜菊苷(“ste”)生成莱鲍迪苷e(“rebe”)。图6a和图6b分别汇总了甜菊苷(“ste”)和莱鲍迪苷e(“rebe”)标准品的hplc保留时间。图6c汇总了在将诱导的g/h5s2细胞引入体外反应体系后24小时从培养基中取样,由培养的g/h5s2细胞酶促生成的莱鲍迪苷e和剩余甜菊苷的hplc保留时间。图6d汇总了在将诱导的phka细胞引入体外反应体系后24小时从反应中取样,由培养的其中并入了缺乏hv1-gcw61和mbsus1序列两者的空载体的巴斯德毕赤酵母菌株(phka)酶促生成的莱鲍迪苷e,以及剩余甜菊苷的hplc保留时间;反应中缺乏莱鲍迪苷e进一步表明hv1-gcw61和mbsus1序列两者均参与reba通过g/h5s2细胞向rebd的转化。
图7a、图7b、图7c、图7d和图7e汇总了使用共同培养的g/k4s2和g/h5s2细胞由莱鲍迪苷a(“reba”)生成莱鲍迪苷m(“rebm”)。图7a、图7b和图7c分别汇总了莱鲍迪苷a(“reba”)、莱鲍迪苷d(“rebd”)和莱鲍迪苷m(“rebm”)标准品的hplc保留时间。图7d和图7e汇总了在将诱导的g/k4s2和g/h5s2细胞引入体外反应体系24小时(图7d)和48小时(图7e)后从培养基中取样,由共同培养的g/k4s2和g/h5s2细胞酶促生成的莱鲍迪苷m和莱鲍迪苷d及剩余莱鲍迪苷a的hplc保留时间。
图8a汇总了在将诱导的g/k4s2和g/h5s2细胞引入体外反应体系后,在各种g/h5s2:g/k4s2细胞密度比率下使用共同培养的g/k4s2和g/h5s2细胞由莱鲍迪苷a(“reba”)几次生成莱鲍迪苷d中间产物(“rebd”)。图8b汇总了在将诱导的g/k4s2和g/h5s2细胞引入体外反应体系后16、24和48小时,在各种g/h5s2:g/k4s2细胞密度比率下使用共同培养的g/k4s2和g/h5s2细胞由莱鲍迪苷a(“reba”)生成莱鲍迪苷m(“rebm”)。
图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h是描绘甜菊醇糖苷生物合成途径的示意图。
图10是udp-糖基转移酶(“ugt”)和蔗糖合成酶(“sus”)的偶联反应体系的示意图。反应1显示ugt催化的使莱鲍迪苷a(“reba”)转化为莱鲍迪苷d(“rebd”)的反应,其使用udp-葡萄糖作为葡萄糖供体并且引起udp的生成。反应2显示sus催化的使udp转化为udp-葡萄糖的反应,其使用蔗糖作为葡萄糖供体。反应2还显示sus催化的反应可以与ugt催化的反应偶联。
图11是说明使用全细胞催化剂生成甜菊醇葡糖苷化合物的方法的方框图。
图12a、图12b、图12c和图12d汇总了使用共同培养的g/k4s2和g/h5s2细胞由甜菊苷(“ste”)生成莱鲍迪苷m(“rebm”)。图12a和图12b分别汇总了莱鲍迪苷a(“reba”)、莱鲍迪苷d(“rebd”)、莱鲍迪苷m(“rebm”)和甜菊苷(“ste”)标准品的hplc保留时间。图12c和图12d汇总了在将诱导的g/k4s2和g/h5s2细胞引入体外反应体系10小时(图12c)和24小时(图12d)后从培养基中取样,由共同培养的g/k4s2和g/h5s2细胞酶促生成的莱鲍迪苷m、莱鲍迪苷d和莱鲍迪苷a及剩余甜菊苷的hplc保留时间。
图13a和13b汇总了使用培养的g/h5s2细胞由莱鲍迪苷ka(“rebka”)生成莱鲍迪苷e(“rebe”)。图13a分别汇总了莱鲍迪苷e(“rebe”)、莱鲍迪苷ka(“rebka”)、甜茶苷(“rub”)标准品的hplc保留时间。图13b汇总了在将g/h5s2细胞引入体外反应体系后24小时从培养基中取样,由培养的g/h5s2细胞酶促生成的莱鲍迪苷e及剩余莱鲍迪苷ka的hplc保留时间。
图14a、图14b和图14c汇总了使用g/h5s2细胞由甜茶苷(“rub”)生成莱鲍迪苷ka(“rebka”)和莱鲍迪苷e(“rebe”)。图14a分别汇总了莱鲍迪苷e(“rebe”)、莱鲍迪苷ka(“rebka”)和甜茶苷(“rub”)标准品的hplc保留时间。图14b和图14c汇总了在将诱导的g/h5s2细胞引入体外反应体系14小时(图14b)和24小时(图14c)后从培养基中取样,由g/h5s2细胞酶促生成的莱鲍迪苷ka和莱鲍迪苷e的hplc保留时间。
虽然本公开易受各种修改和替代性形式影响,但其具体实施方案已经在附图中以举例的方式示出并且本文在下面有详细描述。然而,应理解对具体实施方案的描述并非旨在限制本公开而包括属于所附权利要求书所定义的本公开的精神和范围内的所有修改、等效方案和替代方案。
详述
除非另有定义,否则本文使用的所有技术和科学术语具有本公开所属领域中普通技术人员通常所理解的相同含义。虽然在本公开的实践或试验中可使用与本文描述的那些类似或等效的任何方法和材料,但下面描述的是优选材料和方法。
术语“互补”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于描述能够相互杂交的核苷酸碱基之间的关系。例如,就dna而言,腺苷与胸腺嘧啶互补且胞嘧啶与鸟嘌呤互补。因此,主题技术还包括与所附序列表中所报告的完整序列互补的分离的核酸片段以及那些基本上相似的核酸序列。
术语“核酸”和“核苷酸”根据本领域中普通技术人员所理解的其各自的普通和惯常含义使用,并且不限于用于指呈单链或双链形式的脱氧核糖核苷酸或核糖核苷酸及其聚合物。除非特别限制,该术语涵盖含有天然核苷酸的已知类似物,具有与参考核酸相似的结合性质并且以类似于天然存在的核苷酸的方式代谢的核酸。除非另有说明,否则特定核酸序列还涵盖其经保守性修饰的或简并性变体(例如简并密码子取代)和互补序列,以及明确指示的序列。
术语“分离的”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且用于分离的核酸或分离的多肽的上下文中时,不限于用于指经人工,远离其天然环境存在且因此不是天然产物的核酸或多肽。分离的核酸或多肽可以呈纯化形式存在或者可以存在于非天然环境中,例如转基因宿主细胞中。
如本文中所用的术语“温育”是指混合两种或更多种化学或生物实体(如化学化合物和酶)并允许其在利于生成甜菊醇糖苷组合物的条件下相互作用的过程。
术语“简并性变体”是指具有与参考核酸序列的不同之处在于一个或多个简并密码子取代的残基序列的核酸序列。简并密码子取代可通过产生其中一个或多个选定(或所有)密码子的第三位被混合碱基和/或脱氧肌苷残基取代的序列来实现。核酸序列及其所有简并性变体将表达相同的氨基酸或多肽。
术语“多肽”、“蛋白质”和“肽”根据本领域中普通技术人员所理解的其各自的普通和惯常含义使用;所述三个术语有时可互换使用,并且不限于用于指氨基酸聚合物或氨基酸类似物,而不管其大小或功能如何。虽然“蛋白质”常常在提到相对较大的多肽时使用,而“肽”常常在提到较小多肽时使用,但这些术语在本领域中的使用有重叠和变化。除非另外指出,否则如本文中所用的术语“多肽”是指肽、多肽和蛋白质。术语“蛋白质”、“多肽”和“肽”在提到多核苷酸产物时可互换使用。因此,示例性多肽包括多核苷酸产物、天然存在的蛋白质、同源物、直系同源物、旁系同源物、片段和其它等效物、前述物质的变体和类似物。
术语“多肽片段”和“片段”在提到参考多肽使用时,根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指与参考多肽本身相比其中有氨基酸残基缺失,但是其中剩余的氨基酸序列通常与参考多肽中的相应位置相同的多肽。此类缺失可以出现在参考多肽的氨基末端或羧基末端,或可选地出现在两处。
术语多肽或蛋白质的“功能片段”是指为全长多肽或蛋白质的一部分,并且具有与全长多肽或蛋白质基本上相同的生物活性,或执行基本上相同的功能(例如,执行相同的酶促反应)的肽片段。
术语“变体多肽”、“经修饰的氨基酸序列”或“经修饰的多肽”,可互换使用,是指与参考多肽的不同之处在于一个或多个氨基酸,例如一个或多个氨基酸取代、缺失和/或添加的氨基酸序列。一方面,变体是保持参考多肽的一些或所有能力的“功能变体”。
术语“功能变体”还包括经保守性取代的变体。术语“经保守性取代的变体”是指具有与参考肽的不同之处在于一个或多个保守性氨基酸取代并且保持参考肽的一些或所有活性的氨基酸序列的肽。“保守性氨基酸取代”是氨基酸残基被功能相似的残基取代。保守性取代的实例包括一个非极性(疏水性)残基如异亮氨酸、缬氨酸、亮氨酸或甲硫氨酸对另一个的取代;一个带电的或极性(亲水性)残基对另一个的取代如精氨酸和赖氨酸之间,谷氨酰胺和天冬酰胺之间,苏氨酸和丝氨酸之间;一个碱性残基如赖氨酸或精氨酸对另一个的取代;或一个酸性残基如天冬氨酸或谷氨酸对另一个的取代;或一个芳香族残基,如苯丙氨酸、酪氨酸或色氨酸对另一个的取代。预计此类取代对蛋白质或多肽的表观分子量或等电点影响很小或没有影响。短语“经保守性取代的变体”还包括其中残基被化学衍生化残基置换的肽,条件是所得肽保持如本文所述的参考肽的一些或所有活性。
与主题技术的多肽有关的术语“变体”,还包括具有与参考多肽的氨基酸序列至少75%、至少76%、至少77%、至少78%、至少79%、至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%相同的氨基酸序列的功能活性多肽。
术语“同源”在其所有语法形式和拼写变型中是指具有“共同进化起源”的多核苷酸或多肽,包括来自超家族的多核苷酸或多肽及来自不同物种的同源多核苷酸或蛋白质之间的关系(reeck等,cell50:667,1987)。此类多核苷酸或多肽,不管是在百分比同一性还是保守性位置的特定氨基酸或基序的存在上,都具有其序列相似性所反映的序列同源性。例如,两个同源多肽可具有至少75%、至少76%、至少77%、至少78%、至少79%、至少80%、至少81%、至少82%、至少83%、至少84%、至少85%、至少86%、至少87%、至少88%、至少89%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%相同的氨基酸序列。
关于主题技术的变体多肽序列的“百分比(%)氨基酸序列同一性”是指在比对序列并且在必要时引入空位以实现最大百分比序列同一性之后,而未将任何保守性取代视为序列同一性的一部分,候选序列中的氨基酸残基与参考多肽的氨基酸残基相同的百分比。
为了测定百分比氨基酸序列同一性的比对可以按本领域技术范围内的各种方式实现,例如,使用可公用的计算机软件如blast、blast-2、align、align-2或megalign(dnastar)软件。本领域的技术人员可以测定适当的参数用于测量比对,包括在所比较的序列全长上实现最大限度比对所需的任何算法。例如,可使用序列比较程序ncbi-blast2测定氨基酸序列同一性%。ncbi-blast2序列比较程序可从ncbi.nlm.nih.gov下载。ncbiblast2使用几个搜索参数,其中那些搜索参数全部设为缺省值,包括例如,未遮掩是,链=全部,期望发生10,最小低复杂性长度=15/5,多程e值=0.01,多程常数=25,最终缺口比对的降低=25和评分矩阵=blosum62。在采用ncbi-blast2进行氨基酸序列比较的情况下,给定氨基酸序列a对、与或针对给定氨基酸序列b的氨基酸序列同一性%(可以可选地表达为给定氨基酸序列a对、与或针对给定氨基酸序列b具有或包含某一%氨基酸序列同一性)计算如下:100×分数x/y,其中x是通过序列比对程序ncbi-blast2在该程序对a和b的比对中评分为相同匹配的氨基酸残基的数量,并且其中y为b中的氨基酸残基的总数。将认识到氨基酸序列a的长度不等于氨基酸序列b的长度时,a对b的%氨基酸序列同一性将不等于b对a的%氨基酸序列同一性。
在这个意义上,测定氨基酸序列“相似性”的技术是本领域公知的。一般而言,“相似性”是指在氨基酸相同或具有相似化学和/或物理性质如电荷或疏水性的适当位置对两个或更多个多肽的氨基酸与氨基酸的精确比较。然后可测定所比较的多肽序列之间的所谓“百分比相似性”。测定核酸和氨基酸序列同一性的技术也是本领域公知的并且包括测定对于该基因而言mrna的核苷酸序列(通常经由cdna中间体)并测定其中编码的氨基酸序列,并且将这与第二氨基酸序列进行比较。一般而言,“同一性”是指分别两个多核苷酸或多肽序列的核苷酸与核苷酸或氨基酸与氨基酸的精确一致性。正如两个或更多个氨基酸序列一样,可以通过测定其“百分比同一性”来比较两个或更多个多核苷酸序列。wisconsin序列分析包第8版(可从geneticscomputergroup,madison,wis.获得)中可用的程序,例如gap程序,能够分别计算两个多核苷酸之间的同一性及两个多肽序列之间的同一性和相似性。计算序列之间的同一性或相似性的其它程序为本领域技术人员已知。
与参考位置“对应”的氨基酸位置是指通过比对氨基酸序列鉴定,与参考序列匹配的位置。此类比对可通过人工或通过使用公知的序列比对程序如clustalw2、blast2等进行。
除非另有说明,否则两个多肽或多核苷酸序列的百分比同一性是指在两个序列中较短序列的整个长度上相同氨基酸残基或核苷酸的百分比。
“编码序列”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指编码特定氨基酸序列的dna序列。
“合适调控序列”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指位于编码序列上游(5’非编码序列)、内部或下游(3’非编码序列)的核苷酸序列,并且其影响相关编码序列的转录、rna加工或稳定性或翻译。调控序列可包括启动子、翻译前导序列、内含子和聚腺苷酸化识别序列。
“启动子”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指能够控制编码序列或功能rna的表达的dna序列。一般而言,编码序列位于启动子序列的3’。启动子可整个源自天然基因,或由源自自然界发现的不同启动子的不同元件组成,或甚至包含合成dna片段。本领域的技术人员应理解,不同启动子可指导不同细胞类型中或不同发育阶段或响应于不同环境条件的基因的表达。使基因大多数时间在大多数细胞类型中表达的启动子,通常称为“组成型启动子”。还进一步认识到,因为在大多数情况下调控序列的确切边界尚未完全定义,所以不同长度的dna片段可具有相同的启动子活性。
术语“可操作连接的”是指单个核酸片段上的核酸序列的缔合,使得一个核酸序列的功能受另一个影响。例如,启动子在能够影响编码序列的表达(即编码序列处于启动子的转录控制下)时,启动子与该编码序列可操作连接。编码序列可以在有义或反义方向上与调控序列可操作连接。
如本文中所用的术语“表达”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指源自主题技术的核酸片段的有义(mrna)或反义rna的转录和稳定积聚。“过表达”是指转基因或重组生物体中基因产物的生成超过正常或非转化生物体中的生成水平。
“转化”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指多核苷酸向靶细胞中的转移。转移的多核苷酸可以并入到靶细胞的基因组或染色体dna中,产生基因稳定性遗传,或其可以独立于宿主染色体而复制。含有转化核酸片段的宿主生物体称为“转基因”或“重组”或“转化”生物体。
术语“转化”、“转基因”和“重组”,当在本文中连同宿主细胞使用时,根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指已经向其中引入了异源核酸分子的宿主生物体的细胞,如植物或微生物细胞。核酸分子可以稳定整合到宿主细胞的基因组中,或核酸分子可以作为染色体外分子存在。此类染色体外分子可以自主复制。转化细胞、组织或受试者被理解为不但涵盖转化过程的最终产物,而且涵盖其转基因后代。
术语“重组”、“异源”和“外源”,当在本文中连同多核苷酸使用时,根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指对特定宿主细胞来说外来来源的,或者若来自相同来源,则由其原始形式修饰的多核苷酸(例如,dna序列或基因)。因此,宿主细胞中的异源基因包括特定宿主细胞内源的但已经通过例如使用定点诱变或其它重组技术修饰的基因。所述术语还包括天然存在的dna序列的非天然存在的多个拷贝。因此,所述术语是指对细胞来说外来的或异源的,或与细胞同源但处于宿主细胞内一般未发现该元件的位置或形式的dna片段。
类似地,术语“重组”、“异源”和“外源”,当在本文中连同多肽或氨基酸序列使用时,意指来源于对特定宿主细胞来说外来来源的或者,若来自相同来源,则由其原始形式修饰的多肽或氨基酸序列。因此,重组dna片段可以在宿主细胞中表达以生成重组多肽。
术语“质粒”、“载体”和“盒”根据本领域中普通技术人员所理解的其普通和惯常含义使用,并且不限于用于指常常携带并非细胞中心代谢的一部分且通常呈环状双链dna分子形式的基因的染色体外元件。此类元件可为源自任何来源的单链或双链dna或rna的线性或环状自主复制序列、基因组整合序列、噬菌体或核苷酸序列,其中许多核苷酸序列已经连接或重组到能够连同适当的3’非翻译序列一起将选定基因产物的启动子片段和dna序列引入细胞中的独特构造中。“转化盒”是指含有外来基因且除该外来基因外还具有利于特定宿主细胞转化的元件的特定载体。“表达盒”是指含有外来基因且除该外来基因外还具有允许该基因在外来宿主中增强表达的元件的特定载体。
本文中所用的标准重组dna和分子克隆技术是本领域公知的并且,例如,由sambrook,j.,fritsch,e.f.和maniatis,t.molecularcloning:alaboratorymanual,第2版;coldspringharborlaboratory:coldspringharbor,n.y.,1989(下文称为“maniatis”);和由silhavy,t.j.、bennan,m.l.和enquist,l.w.experimentswithgenefusions;coldspringharborlaboratory:coldspringharbor,n.y.,1984;和由ausubel,f.m.等,在1987年由greenepublishing和wiley-interscience出版的currentprotocolsinmolecularbiology中有描述,其各自整体据此通过引用在与之一致的程度上并入本文。
如本文中所用,“合成”或“有机合成”或“化学合成”用于指通过一系列化学反应制备化合物;这不包括提取化合物,例如,从天然来源提取。
如本文中所用的术语“可口服消耗产品”是指与人或动物口腔接触的任何饮料、食物产品、膳食补充剂、营养制剂、药物组合物、口腔卫生组合物和美容产品,包括摄入口腔并随后由口腔排出的物质及饮用、嚼食、吞咽或以其它方式消化的物质;及在通常可接受的浓度范围内使用时对人或动物消耗而言是安全的物质。
如本文中所用的术语“食物产品”是指水果、蔬菜、果汁、肉制品如火腿、培根(bacon)、香肠;蛋制品、水果浓缩物、明胶和明胶样产品如果酱、果冻、蜜饯等;乳制品如冰激凌、酸奶油、酸奶和冻果汁露;糖霜、糖浆,包括糖蜜;玉米、小麦、黑麦、大豆、燕麦、水稻和大麦制品、谷类制品、坚果仁和坚果制品、蛋糕、曲奇、糖食(如糖果)、树胶、水果味硬糖(fruitflavoreddrop)和巧克力、口香糖、薄荷、奶油、糖霜、冰淇淋、馅饼和面包。“食物产品”还指调味品如药草、香料和佐料、增香剂,如谷氨酸一钠。“食物产品”进一步指还包括制备的包装产品,如饮食甜味剂、液体甜味剂、桌面调味剂、与水冲调得到非碳酸饮料的颗粒调味混合物、速食布丁混合物、速溶咖啡和茶、咖啡伴侣、麦乳精混合物、宠物食品、牲畜饲料、烟草,以及用于烘焙应用的材料,例如用于制备面包、曲奇、蛋糕、薄煎饼、甜甜圈等的粉状烘焙混合物。“食物产品”还指含有少量或不含蔗糖的减肥或低热量食品和饮料。
如本文中所用,术语“立体异构体”是仅在其原子在空间中的取向上不同的独立分子的所有异构体的统称。“立体异构体”包括对映异构体和彼此非镜像的具有多于一个手性中心的化合物的异构体(非对映异构体)。
如本文中所用,术语“甜味强度”是指由个体(例如人)所观察到的或经历的甜味感觉的相对强度,或由尝味者例如基于布里标度(brixscale)检测的甜味的程度或量。
如本文中所用,术语“提高甜度”是指与不含莱鲍迪苷m和/或莱鲍迪苷d的相应可口服消耗产品相比,莱鲍迪苷m和/或莱鲍迪苷d增加、增大、增强、加重、放大和/或加强本公开的饮料产品或可消耗产品的一种或多种甜度特性的感官知觉而不改变其性质或质量的作用。
如本文中所用,术语“异味”是指非典型或不常存在于本公开的饮料产品或可消耗产品中的味道的量或程度。例如,异味是对于消费者来说增甜消耗品不理想的味道,例如,苦味、类甘草味、金属味、令人反感的味道、涩味、延迟的甜味产生、绵长的甜回味及类似味道,等等。
如本文中所用,术语“w/v-%”是指对于每100ml含化合物如糖的本公开的液体可口服消耗产品而言此类化合物的重量(按克计)。如本文中所用,术语“w/w-%”是指对于每克含化合物如糖的本公开的可口服消耗产品而言此类化合物的重量(按克计)。
如本文中所用,术语“ppm”是指按重量计的百万分率,例如每千克含化合物的本公开的可口服消耗产品中此类化合物,如莱鲍迪苷m和/或莱鲍迪苷d的重量(按毫克计)(即,mg/kg)或每升含化合物的本公开的可口服消耗产品中此类化合物,如莱鲍迪苷m和/或莱鲍迪苷d的重量(按毫克计)(即,mg/l);或按体积计的百万分率,例如每升含化合物的本公开的可口服消耗产品中此类化合物,如莱鲍迪苷m和/或莱鲍迪苷d的重量(按毫升计)(即,ml/l)。
根据本公开,公开了甜菊醇糖苷无热量甜味剂和合成甜菊醇糖苷的方法。同样依照本公开,公开了包括生成一种或多种酶的一种或多种工程化细胞的全细胞催化剂,生成所述全细胞催化剂的方法,及使用所述全细胞催化剂制备甜菊醇糖苷的方法。
全细胞催化剂
一方面,提供了至少一种全细胞催化剂。在这个方面中,所述至少一种全细胞催化剂是表达至少一种酶的转化宿主细胞,所述至少一种酶包括但不限于:尿苷二磷酸糖基转移酶(udp-糖基转移酶或ugt)和蔗糖合成酶(sus)。在下文描述的各个方面中,所述至少一种全细胞催化剂可用于合成无热量甜味剂的方法中,所述方法包括将所述至少一种全细胞催化剂与合适的底物混合;所述至少一种全细胞催化剂产生的酶催化所述底物和任何所得中间产物的至少一个糖基化反应以生成所需的无热量甜味剂。
另一方面,由转化宿主细胞表达的ugt酶可展示在宿主细胞的表面。在这个方面中,表达的ugt酶可以与展示多肽融合以利于表达的ugt酶在转化宿主细胞表面的展示。可以根据至少几个因素中的一个或多个选择与表达的酶融合的展示多肽,所述因素包括但不限于:对展示的ugt酶的酶促功能无干扰并且与宿主细胞类型相容。
在各个方面中,全细胞催化剂可包括表达附接于细胞表面的ugt酶并且还在细胞内表达蔗糖合成酶(sus)的转化宿主细胞。在这些各个方面中,通过并入包括至少一个编码所述至少一种酶的核苷酸序列的至少一个表达盒来转化宿主细胞,所述至少一种酶包括但不限于尿苷二磷酸糖基转移酶(udp-糖基转移酶或ugt)和蔗糖合成酶(sus)。另外,编码ugt酶的核苷酸序列还可包括编码展示多肽的额外核苷酸片段,使得宿主细胞表达包括与展示蛋白融合的ugt酶的融合多肽。
宿主细胞
在这个方面中,转化宿主细胞可以是通过至少一个表达盒转化的任何合适的宿主细胞,表达盒包括至少一个编码由每种全细胞催化剂所表达的至少一种酶的核苷酸序列。用于转化为全细胞催化剂的合适的宿主细胞的非限制性实例包括:细菌、酵母、丝状真菌、蓝细菌藻和植物细胞。
一方面,宿主细胞可以是细菌,包括但不限于:肠细菌如埃希氏杆菌属(escherichia)、沙门氏菌属(salmonella)和克雷白杆菌属(klebsiella);杆菌属(bacillus);不动杆菌属(acinetobacter);泛生菌属(pantoea);放线菌类(actinomycetes)如链霉菌属(streptomyces)和棒状杆菌属(corynebacterium);甲烷氧化菌类(methanotrophs)如甲基弯曲菌属(methylosinus)、甲基单胞菌属(methylomonas)、红球菌属(rhodococcus)和假单胞菌属(pseudomona);梭状芽胞杆菌属(clostridium)如丙酮丁醇梭状芽孢杆菌(clostridiumacetobutylicum);和蓝细菌(cyanobacteria)如红细菌属(rhodobacter)和集胞藻属(synechocystis)。另一方面,宿主细胞可为酵母,包括但不限于:arxula属、假丝酵母属(candidaspp.)、德巴利酵母属(debaryomycesspp.)、汉森酵母属(hansenulaspp.)、克鲁维酵母属(kluyveromycesspp.)、毛霉菌属(mucorspp.)、管囊酵母属(pachysolenspp.)、法夫酵母属(phaffiaspp.)、毕赤酵母属(pichiaspp.)、红冬孢酵母属(rhodosporidiumspp.)、酵母属(saccharomycesspp.)、复膜孢酵母属(saccharomycopsisspp.)、许旺酵母属(scwanniomycesspp.)、丝孢酵母属(tricosporonspp.)、球拟酵母属(torulopsisspp.)、耶氏酵母属(yarrowiaspp.)、和耶氏酵母属(zygosaccharomycesspp.)。再一方面,宿主细胞可为酵母,包括但不限于:arxulaadeninivorans、白假丝酵母(candidaalbicans)、博伊丁假丝酵母(candidaboidinii)、无名假丝酵母(candidafamata)、麦芽糖假丝酵母(candidamaltosa)、热带假丝酵母(candidatropicalis)、产朊假丝酵母(candidautilis)、休哈塔假丝酵母(candidashehatae)、多形汉森酵母(hansenulapolymorpha)、马克斯克鲁维酵母(kluyveromycesmarxianus)、乳酸克鲁维酵母(kluyveromyceslactis)、嗜单宁管囊酵母(pachysolentannophilus)、红法夫酵母(phaffiarhodozyma)、季也蒙毕赤酵母(pichiaguillermondii)、甲醇毕赤酵母(pichiamethanolica)、巴斯德毕赤酵母(pichiapastoris)、圆红冬孢酵母(rhodosporidiumtoruloides)、酿酒酵母(saccharomycescerevisiae)、酿酒酵母糖化变种(saccharomycescerevisiaevar.diastaticus)、布拉酵母(saccharomycesboulardii)、梨形酵母(saccharomycespyriformis)、贝酵母(saccharomycesbayanis)、扣囊复膜孢酵母(saccharomycopsisfibuligera)、卡斯坦氏许旺酵母(scwanniomycescastellii)、西方许旺酵母(scwanniomycesoccidentalis)、皮状丝孢酵母(tricosporoncutaneum)、解脂耶氏酵母(yarrowialipolytica)和鲁氏接合酵母(zygosaccharomycesrouxii)。另一方面,宿主细胞可为丝状真菌,包括但不限于:曲霉属(aspergillus)和节丛孢属(arthrobotrys)。再另一方面,宿主细胞可为藻类,包括但不限于:螺旋藻属(spirulina)、红球藻属(haemotacoccus)和杜氏藻属(dunaliella)。
再另一方面,宿主细胞可为植物细胞,包括但不限于源自单子叶植物或双子叶植物并且能够构成未分化组织如愈伤组织、分化组织如胚、单子叶植物的部分、单子叶植物或种子的任何细胞。术语“植物”理解为意指能够进行光合作用的任何分化的多细胞生物体,包括单子叶植物和双子叶植物。在一些实施方案中,植物细胞可为拟南芥(arabidopsis)植物细胞、烟草植物细胞、大豆植物细胞、矮牵牛植物细胞或来自另一油料作物的细胞,包括但不限于芥花(canola)植物细胞、油菜植物细胞、棕榈植物细胞、向日葵植物细胞、棉花植物细胞、玉米植物细胞、花生植物细胞、亚麻植物细胞和芝麻植物细胞。有用的植物宿主可包括支持主题技术的重组多肽生成的任何植物。用作宿主的合适绿色植物包括但不限于大豆、油菜(甘蓝型油菜(brassicanapus)、白菜型油菜(b.campestris))、向日葵(向日葵(helianthusannus))、棉花(陆地棉(gossypiumhirsutum))、玉米、烟草(烟草(nicotianatabacum))、苜蓿(紫花苜蓿(medicagosativa))、小麦(小麦属(triticumspp.))、大麦(大麦(hordeumvulgare))、燕麦(燕麦(avenasativa))、高粱(高粱(sorghumbicolor))、水稻(水稻(oryzasativa))、拟南芥、十字花科蔬菜(花茎甘蓝、花椰菜、卷心菜、欧洲防风草等)、瓜、胡萝卜、芹菜、欧芹、西红柿、马铃薯、草莓、花生、葡萄、牧草种子作物、甜菜、甘蔗、豆类、豌豆、黑麦、亚麻、阔叶树、针叶树和饲草。一方面,宿主细胞可以是源自植物的植物细胞,包括但不限于:阿拉伯芥(arabidopsisthaliana)、水稻(水稻(oryzasativa))、大麦(hordeumvulgare)、柳枝稷(柳枝稷(panicumvigratum))、短柄草属(brachypodiumspp.)、芸苔属(brassicaspp.)和深海两节荠(crambeabyssinica)。
在各个方面中,包括至少一个编码至少一种酶的核苷酸序列的表达盒可以并入这些和其它宿主细胞中以制备大的、商业上有用量的甜菊醇糖苷。
酶
在各个方面中,将含有至少一个编码至少一种酶的核苷酸序列的至少一个表达盒并入宿主细胞中以将宿主细胞转化为全细胞催化剂。通常,选择所述至少一个表达盒以与宿主细胞类型相容,并且编码所述至少一种酶的所述至少一个核苷酸序列内的密码子可以使用已知方法进行密码子优化以便增强所述至少一种酶在选定的宿主细胞类型中的表达。表达盒的设计取决于诸如待转化的宿主细胞的选择、所需酶的表达水平等因素。可将表达盒引入宿主细胞内,从而产生重组多肽酶,包括但不限于尿苷二磷酸糖基转移酶(udp-糖基转移酶)和蔗糖合成酶(sus)。
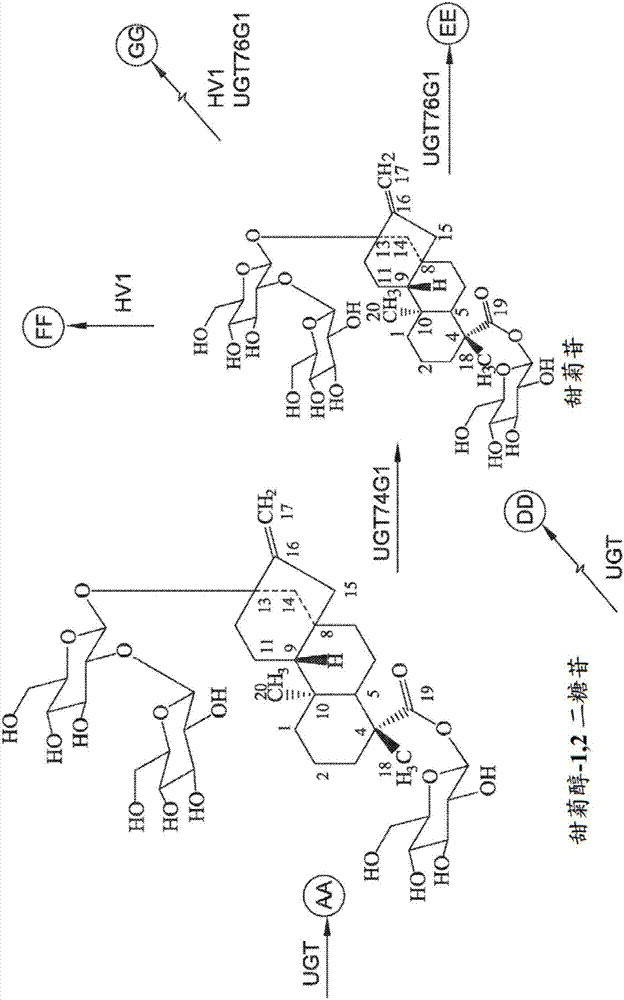
一方面,并入所述至少一个表达盒内的核苷酸序列包括编码尿苷二磷酸糖基转移酶(udp-糖基转移酶)的至少一个核苷酸序列。udp-糖基转移酶(ugt)是指将糖残基从活化供体分子(通常为udp-葡萄糖)转移到受体分子的酶。在各个方面中,ugt酶可具有与甜菊醇糖苷合成相关的任何一种或多种活性,包括但不限于:1,2-19-o-葡萄糖糖基化活性、1,2-13-o-葡萄糖糖基化活性、1,3-13-o-葡萄糖糖基化活性和1,3-19-o-葡萄糖糖基化。1,2-19-o-葡萄糖糖基化活性是指将糖部分转移到底物的19-o-葡萄糖部分的c-2’的酶活性,底物包括但不限于:甜茶苷、甜菊苷、莱鲍迪苷a或莱鲍迪苷e(参见图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h)。1,2-13-o-葡萄糖糖基化活性是指将糖部分转移到底物的13-o-葡萄糖部分的c-2’的酶活性,底物包括但不限于:莱鲍迪苷ka和莱鲍迪苷e(参见图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h)。任何已知的ugt均可并入所述至少一个表达盒内,包括但不限于:ugt76g1、hv1、eugt11及其任何组合。
ugt76g1是具有1,3-13-o-葡萄糖糖基化活性以分别由甜菊苷和莱鲍迪苷e生成相关的莱鲍迪苷a和d的ugt。另外,ugt76g1还具有1,3-19-o-葡萄糖糖基化活性以由莱鲍迪苷d生成莱鲍迪苷m。ugt76g1也可以将莱鲍迪苷ka转化为rebv并继续形成rebw(参见图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h)。一方面,ugt76g1酶具有seqidno:9的氨基酸序列。
在这样的一个方面,所述至少一个表达盒可包含编码ugt76g1酶的核苷酸序列,所述核苷酸序列包含与seqidno:10中列出的核酸序列具有至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同源性的核苷酸序列。适当地,编码ugt76g1酶的核苷酸序列可具有与seqidno:9中列出的氨基酸序列具有至少80%序列同一性的氨基酸序列。更适当地,核苷酸序列可编码ugt76g1酶,所述酶具有与seqidno:9中列出的氨基酸序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的氨基酸序列。编码ugt76g1酶的核苷酸序列因此包括编码seqidno:9的功能片段、seqidno:9的功能变体或与seqidno:9具有(例如)至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的其它同源多肽的那些核苷酸序列。正如本领域技术人员所知,编码ugt76g1酶的核酸序列可以经密码子优化以在合适的宿主生物体,例如细菌或酵母中表达。
hv1是具有分别由甜菊苷、莱鲍迪苷a和莱鲍迪苷e生成相关莱鲍迪苷e、d和z的1,2-19-o-葡萄糖糖基化活性的ugt。hv1还具有由甜茶苷生成莱鲍迪苷ka的1,2-19-o-葡萄糖糖基化活性。hv1还可将rebg转化成rebv且将rebka转化成rebe(参见图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h)。hv1还具有由莱鲍迪苷e生成莱鲍迪苷z1的1,2-13-o-葡萄糖糖基化活性。一方面,hv1酶具有seqidno:7的氨基酸序列。
在这样的一个方面,所述至少一个表达盒可包含编码hv1酶的核苷酸序列,所述核苷酸序列包含与seqidno:8中列出的核酸序列具有至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同源性的核苷酸序列。适当地,编码hv1酶的核苷酸序列可具有与seqidno:7中列出的氨基酸序列具有至少80%序列同一性的氨基酸序列。更适当地,核苷酸序列可编码hv1酶,所述酶具有与seqidno:7中列出的氨基酸序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的氨基酸序列。编码hv1酶的核苷酸序列因此包括编码seqidno:7的功能片段、seqidno:7的功能变体或与seqidno:7具有(例如)至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的其它同源多肽的那些核苷酸序列。正如本领域技术人员所知,编码hv1酶的核酸序列可以经密码子优化以在合适的宿主生物体,例如细菌或酵母中表达。
eugt11是至少具有1,2-19-o-葡萄糖糖基化活性和1,2-13-o-葡萄糖糖基化活性的ugt。eugt11可催化甜菊苷糖基化为莱鲍迪苷e和莱鲍迪苷a糖基化为莱鲍迪苷d(参见图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h)。eugt11也可以在体外用于通过另一种酶活性,β1,6-13-o-葡萄糖糖基化活性由莱鲍迪苷e合成莱鲍迪苷d2。eugt11还具有由甜茶苷生成莱鲍迪苷ka的1,2-19-o-葡萄糖糖基化活性。
在各个方面中,ugt可以与上文所述的展示多肽融合,使得ugt可以展示在转化宿主细胞的表面。一方面,所述至少一个表达盒可包括至少一个核苷酸序列,其包括含编码本文先前所述的ugt酶的序列的第一核苷酸部分和编码展示多肽的第二核苷酸部分。在这一方面,所述至少一个核苷酸序列可表达包括附接于展示多肽的ugt酶的融合蛋白,使得ugt酶经由展示多肽附接于宿主细胞的细胞表面。
在各个方面中,可选择展示多肽以与经转化生成全细胞催化剂的宿主细胞的类型相容。一方面,如果宿主细胞为细菌细胞,则ugt酶可以与细菌细胞表面蛋白融合,包括但不限于:细菌外膜蛋白或菌毛蛋白或鞭毛蛋白。另一方面,如果宿主细胞为酵母细胞,则ugt酶可以与酵母细胞表面蛋白融合,包括但不限于:酵母细胞壁蛋白如gcw61(seqidno:5)、酵母交配粘附蛋白的亚基如凝集素或生物素结合肽。在这其它方面,如果ugt酶与酵母交配粘附蛋白的亚基如凝集素融合,则酵母宿主细胞可进一步转化以表达酵母交配粘附蛋白附接于酵母细胞壁的相应亚基,使得表达的ugt-凝集素融合蛋白可以与附接于酵母细胞壁的相应亚基结合,导致ugt酶在细胞壁表面上展示。在这其它方面,如果ugt酶与生物素结合肽融合,则酵母细胞可进一步修饰以在酵母细胞表面展示生物素并且在分泌之前使融合蛋白生物素化;酵母细胞与抗生物素蛋白接触,抗生物素蛋白与酵母细胞壁内的生物素以及与生物素化融合蛋白结合。
一方面,如果宿主细胞为巴斯德毕赤酵母细胞,则与ugt酶融合的展示多肽可为具有seqidno:5的氨基酸序列的毕赤酵母细胞壁蛋白(gcw61)。
在这样的一个方面,所述至少一个表达盒可包含编码毕赤酵母细胞壁蛋白(gcw61)的核苷酸序列,所述核苷酸序列包含与seqidno:6中列出的核酸序列具有至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同源性的核苷酸序列。适当地,编码毕赤酵母细胞壁蛋白(gcw61)的核苷酸序列可具有与seqidno:5中列出的氨基酸序列具有至少80%序列同一性的氨基酸序列。更适当地,核苷酸序列可编码毕赤酵母细胞壁蛋白(gcw61),所述毕赤酵母细胞壁蛋白具有与seqidno:5中列出的氨基酸序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的氨基酸序列。编码毕赤酵母细胞壁蛋白(gcw61)的核苷酸序列因此包括编码seqidno:5的功能片段、seqidno:5的功能变体或与seqidno:5具有(例如)至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的其它同源多肽的那些核苷酸序列。正如本领域技术人员所知,编码毕赤酵母细胞壁蛋白(gcw61)的核酸序列可以经密码子优化以在合适的宿主生物体,例如细菌或酵母中表达。
另一方面,编码毕赤酵母细胞壁蛋白(gcw61)的核苷酸序列可插入编码ugt酶的核苷酸序列c端的框内。在这样的其他方面,编码ugt酶的核苷酸序列可包括上文所述的任何核苷酸序列,包括但不限于:编码ugt76g1酶的seqidno:10和编码hv1酶的seqidno:8。在这样的其他方面,包括插入ugt76g1核苷酸序列(seqidno:10)或hv1核苷酸序列(seqidno:8)的c端框内的gcw61序列(seqidno:6)的融合蛋白序列分别表达为融合蛋白ugt76g1-gcw61(seqidno:3)或hv1-gcw61(seqidno:1)中的一种。
在这样的其他方面,所述至少一个表达盒可包含编码融合的ugt酶-细胞壁多肽(ugt76g1-gcw61)的核苷酸序列,所述核苷酸序列包含与seqidno:4中列出的核酸序列具有至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同源性的核苷酸序列。适当地,编码融合的ugt酶-细胞壁多肽(ugt76g1-gcw61)的核苷酸序列可具有与seqidno:3中列出的氨基酸序列具有至少80%序列同一性的氨基酸序列。更适当地,核苷酸序列可编码融合的ugt酶-细胞壁多肽(ugt76g1-gcw61),所述多肽具有与seqidno:3中列出的氨基酸序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的氨基酸序列。编码融合的ugt酶-细胞壁多肽(ugt76g1-gcw61)的核苷酸序列因此包括编码seqidno:3的功能片段、seqidno:3的功能变体或与seqidno:3具有(例如)至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的其它同源多肽的那些核苷酸序列。正如本领域技术人员所知,编码融合的ugt酶-细胞壁多肽(ugt76g1-gcw61)的核酸序列可以经密码子优化以在合适的宿主生物体,例如细菌和酵母中表达。
在这样的其他方面,所述至少一个表达盒还可以包含编码融合的ugt酶-细胞壁多肽(hv1-gcw61)的核苷酸序列,所述核苷酸序列包含与seqidno:2中列出的核酸序列具有至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同源性的核苷酸序列。适当地,编码融合的ugt酶-细胞壁多肽(hv1-gcw61)的核苷酸序列可具有与seqidno:1中列出的氨基酸序列具有至少80%序列同一性的氨基酸序列。更适当地,核苷酸序列可编码融合的ugt酶-细胞壁多肽(hv1-gcw61),所述多肽具有与seqidno:1中列出的氨基酸序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的氨基酸序列。编码融合的ugt酶-细胞壁多肽(hv1-gcw61)的核苷酸序列因此包括编码seqidno:1的功能片段、seqidno:1的功能变体或与seqidno:1具有(例如)至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的其它同源多肽的那些核苷酸序列。正如本领域技术人员所知,编码融合的ugt酶-细胞壁多肽(hv1-gcw61)的核酸序列可以经密码子优化以在合适的宿主生物体,例如细菌和酵母中表达。
另一方面,并入所述至少一个表达盒内的核苷酸序列包括编码蔗糖合成酶(sus)的至少一个核苷酸序列。蔗糖合成酶(sus)催化在蔗糖的存在下udp转化为udp-葡萄糖(参见图10)。因此,对于利用udp-葡萄糖的糖基化反应(如ugt催化的那些反应),sus可用于由udp再生udp-葡萄糖,增强了此类反应的效率。
编码任何合适的蔗糖合成酶的核苷酸序列均可并入到所述至少一个表达盒中,且无限制。一般而言,蔗糖合成酶被归类为糖基转移酶,系统名称为ndp-葡萄糖:d-果糖2-α-d-葡萄糖基转移酶。用于描述蔗糖合成酶的其它名称的非限制性实例包括:udp葡萄糖-果糖葡糖基转移酶、蔗糖合成酶、蔗糖-udp葡糖基转移酶、蔗糖-尿苷二磷酸葡糖基转移酶和尿苷二磷酸葡萄糖-果糖葡糖基转移酶。
一方面,合适的蔗糖合成酶包括但不限于来源于以下的那些:阿拉伯芥和绿豆sus基因,或编码由阿拉伯芥和绿豆sus1序列编码的蔗糖合成酶的功能同源物的任何基因,或其功能同源物。合适的蔗糖合成酶的非限制实例包括:拟南芥(arabidopsis)蔗糖合成酶1;拟南芥蔗糖合成酶3;和绿豆(vignaradiate)蔗糖合成酶(mbsus1)。一方面,蔗糖合成酶可以是具有seqidno:11的氨基酸序列的mbsus1酶。
在这样的一个方面,所述至少一个表达盒可包含编码mbsus1酶的核苷酸序列,所述核苷酸序列包含与seqidno:12中列出的核酸序列具有至少70%、至少75%、至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同源性的核苷酸序列。适当地,编码mbsus1酶的核苷酸序列可具有与seqidno:11中列出的氨基酸序列具有至少80%序列同一性的氨基酸序列。更适当地,核苷酸序列可编码mbsus1酶,所述酶具有与seqidno:11中列出的氨基酸序列具有至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的氨基酸序列。编码mbsus1酶的核苷酸序列因此包括编码seqidno:11的功能片段、seqidno:11的功能变体或与seqidno:11具有(例如)至少80%、至少85%、至少90%、至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%和甚至100%序列同一性的其它同源多肽的那些核苷酸序列。正如本领域技术人员所知,编码mbsus1酶的核酸序列可以经密码子优化以在合适的宿主生物体,例如细菌或酵母中表达。
表达盒
在各个方面中,如上所述编码至少一种酶的所述至少一个核苷酸并入到至少一个表达盒中,将表达盒引入宿主细胞中以便产生用作全细胞催化剂的转化宿主细胞。可以选择任何表达盒、表达质粒、表达载体或向宿主细胞中引入外源遗传物质的任何其它已知工具用作全细胞催化剂的所述至少一个表达盒。一方面,可以根据至少几个因素中的任一个或多个来选择表达盒,所述因素包括但不限于与宿主细胞类型的相容性、表达效率、易用性和任何其它相关因素。
对于微生物宿主细胞,包括但不限于细菌细胞和酵母细胞,指导外来蛋白质高水平表达的微生物宿主细胞表达系统和表达载体是本领域技术人员公知的。这些中的任一种均可用于构建在微生物宿主细胞中表达主题技术的重组多肽的载体。然后可经由转化将这些载体引入适当的微生物中以允许主题技术的重组多肽高水平表达。
用于转化合适的微生物宿主细胞的载体或盒是本领域公知的。通常,所述载体或盒含有指导相关多肽转录和翻译的序列、可选标记和允许自主复制或染色体整合的序列。合适的载体包含具有转录起始控制的多核苷酸的5’区和控制转录终止的dna片段的3’区。虽然应理解此类控制区不需要源自选作宿主的特定物种天然的基因,但优选控制区源自与转化宿主细胞同源的基因。
用于驱动重组多肽在预期微生物宿主细胞内的表达的起始控制区或启动子有许多并且为本领域的技术人员所熟悉。几乎任何能够驱动这些基因的启动子都适于主题技术,包括但不限于cyc1、his3、gal1、gal10、adh1、pgk、pho5、gapdh、adc1、trp1、ura3、leu2、eno、tpi(用于在酵母属中表达);aox1(用于在毕赤酵母属中表达);及lac、trp、ipl、ipr、t7、tac和trc(用于在大肠杆菌(escherichiacoli)中表达)。
终止控制区也可源自微生物宿主天然的各个基因。对于本文描述的微生物宿主而言任选地可包括终止位点。
在植物细胞中,主题技术的表达载体可包括与能够指导主题技术的重组多肽在预期发育阶段在预期组织内的表达的启动子可操作地连接的编码区。为了方便起见,待表达的多核苷酸可包含源自相同多核苷酸的启动子序列和翻译前导序列。还应存在编码转录终止信号的3’非编码序列。表达载体还可包含一个或多个内含子以便促进多核苷酸表达。
对于植物宿主细胞而言,能够诱导编码区表达的任何启动子和任何终止子的任何组合均可用于主题技术的载体序列中。启动子和终止子的一些合适的实例包括来自胭脂碱合成酶(nos)、真蛸碱合成酶(ocs)和花椰菜花叶病毒(camv)基因的那些。可以使用的一类有效植物启动子是高水平植物启动子。此类启动子,与主题技术的表达载体呈可操作连接,应该能够启动载体的表达。可用于主题技术中的高水平植物启动子包括例如来自大豆的1,5-二磷酸核酮糖羧化酶的小亚基(ss)的启动子(berry-lowe等人,j.molecularandapp.gen.,1:483498(1982),其整体据此并入本文达到与之一致的程度)和叶绿素a/b结合蛋白的启动子。已知这两种启动子在植物细胞中受光诱导(参见,例如,geneticengineeringofplants,anagriculturalperspective,a.cashmore,plenum,n.y.(1983),第2938页;coruzzi,g.等人,thejournalofbiologicalchemistry,258:1399(1983),及dunsmuir,p.等人,journalofmolecularandappliedgenetics,2:285(1983),其各自据此以引用的方式并入本文达到与之一致的程度)。
对质粒载体的选择取决于将用于转化宿主植物的方法。技术人员充分意识到在质粒载体上必须存在遗传元件以便成功地转化、选择和增殖含嵌合多核苷酸的宿主细胞。技术人员还会认识到不同的独立转化事件将引起不同水平和模式的表达(jones等人,emboj.4:24112418(1985);dealmeida等人,mol.gen.genetics218:7886(1989),其各自据此以引用的方式并入本文达到与之一致的程度),因此多个事件必须筛选以便获得展示出预期表达水平和模式的细胞系。此类筛选可以通过对dna印迹的southern分析、对mrna表达的northern分析、蛋白质表达的western分析或表型分析实现。
将主题技术的表达载体引入植物细胞内可以通过本领域普通技术人员已知的各种方法进行,包括将目标核酸序列插入发根土壤杆菌(agrobacteriumrhizogenes)ri或根癌土壤杆菌(agrobacteriumtumefaciens)ti质粒中,显微注射,电穿孔或直接沉淀法。通过提供实例的方式,在一些实施方案中,目标多核苷酸的瞬时表达可通过农杆菌浸润法进行。就这一点而言,含目标多核苷酸的根癌土壤杆菌的悬浮液可在培养物中生长,然后通过将注射器的尖端放置在叶片的下面,同时向叶片的另一面施加轻微的反向压力而注入植物中。然后土壤杆菌属溶液通过气孔注入叶片内部的空间。一旦进入叶片内部,土壤杆菌属就将目标基因转化到植物的一部分,然后基因在这里瞬时表达。
再如,目标质粒向植物细胞的转化可以通过粒子枪轰击技术(即,基因枪)进行。就这一点而言,植物胚胎的悬浮液可以在液体培养物中生长,然后用附接于金粒子的质粒或多核苷酸轰击,其中与目标质粒或核酸结合的金粒子可以被推动通过植物组织如胚性组织的膜。轰击后,可以使用适当的抗生素来选择转化胚以产生新的、克隆繁殖的、转化胚性悬浮培养物。
宿主细胞可以是未修饰的细胞或细胞系,或已经基因修饰的细胞系。在一些实施方案中,宿主细胞是已经修饰以允许在预期条件下,例如在较低温度下生长的细胞系。
标准重组dna方法可用于获得编码本文描述的重组多肽的核酸,将该核酸并入表达载体中,并且将载体引入宿主细胞,如sambrook等人(编辑),molecularcloning;alaboratorymanual,第三版,coldspringharbor,(2001);及ausubel,f.m.等人(编辑)currentprotocolsinmolecularbiology,johnwiley&sons(1995)中描述的那些。可将编码多肽的核酸插入一个表达载体或多个载体中,使得核酸与转录和翻译控制序列(如启动子序列、转录终止序列等)可操作地连接。通常将表达载体和表达控制序列选择为与所用表达宿主细胞相容。
可通过密码子优化进一步提高多肽在本文所述宿主中的表达。例如,以更常见的密码子修饰不太常见的密码子可以影响mrna的半衰期或通过引入干扰信息翻译的二级结构而改变mrna结构。可优化整个或一部分编码区。在一些情况下对表达的预期调节可通过基本上优化整个基因来实现。在其它情况下,预期调节将通过优化基因的部分但非整个序列来实现。
可调节任何编码序列的密码子使用以获得预期性质,例如在特定细胞类型中的高水平表达。此类优化的起始点可以是具有100%常见密码子的编码序列,或含有常见和非常见密码子的混合物的编码序列。
可以生成在密码子使用上不同的两个或更多个候选序列并测试以确定它们是否具有预期性质。可通过使用计算机搜索调控元件,如沉默子或增强子的存在,并且搜索可以通过密码子使用的改变而转化为此类调控元件的编码序列区域的存在来评估候选序列。额外标准可包括特定核苷酸,例如a、c、g或u的富集,特定氨基酸的密码子偏好,或特定mrna二级或三级结构的存在或不存在。对候选序列的调节可基于许多此类标准进行。
在某些实施方案中,密码子优化的核酸序列可表达其蛋白质,水平是由尚未经密码子优化的核酸序列表达的水平的约110%、约150%、约200%、约250%、约300%、约350%、约400%、约450%或约500%。
除了编码主题技术的重组多肽的核酸外,主题技术的表达载体可另外携带控制蛋白质在宿主细胞内的表达的调控序列,如启动子、增强子或控制核酸的转录或翻译的其它表达控制元件。此类调控序列是本领域已知的。本领域的技术人员将认识到,表达载体的设计,包括调控序列的选择,可取决于诸如待转化宿主细胞的选择,预期蛋白质的表达水平等因素。另外,主题技术的重组表达载体可携带额外序列,如调控载体在宿主细胞内的复制的序列(例如,复制起点)和可选标记基因。
在各个方面中,所述至少一个表达盒可包括编码至少一种酶的至少一个核苷酸序列,所述至少一种酶包括但不限于如上文所述的尿苷二磷酸糖基转移酶(udp-糖基转移酶或ugt)和蔗糖合成酶(sus)。一方面,所述至少一个表达盒可包括第一表达盒,第一表达盒包括至少一个ugt序列,和第二表达盒,第二表达盒包括至少一个sus序列。另一方面,所述至少一个表达盒可包括一个包括所述至少一个ugt序列和所述至少一个usu序列两者的表达盒。
在各个其它方面中,ugt和/或sus序列的表达程度可通过并入多个拷贝的ugt序列和/或多个拷贝的sus序列来调节。一方面,用于转化宿主细胞的所述至少一个表达盒可包括单表达盒,单表达盒包括多个拷贝的ugt序列和/或多个拷贝的sus序列。另一方面,所述至少一个表达盒可包括多个拷贝的第一盒,第一盒包括单个拷贝的ugt序列,和多个拷贝的第二盒,第二盒包括单个拷贝的sus序列。
一方面,所述至少一个表达盒可包括一个拷贝的编码ugt76g1酶(seqidno:9)的ugt76g1核苷酸序列(seqidno:10)。在其它方面中,所述至少一个表达盒可包括两个拷贝的ugt76g1核苷酸序列、三个拷贝的ugt76g1核苷酸序列、四个拷贝的ugt76g1核苷酸序列、五个拷贝的ugt76g1核苷酸序列、六个拷贝的ugt76g1核苷酸序列、七个拷贝的ugt76g1核苷酸序列、八个拷贝的ugt76g1核苷酸序列、九个拷贝的ugt76g1核苷酸序列或十个拷贝的ugt76g1核苷酸序列。
一方面,所述至少一个表达盒可包括一个拷贝的编码hv1酶(seqidno:7)的hv1核苷酸序列(seqidno:8)。在其它方面中,所述至少一个表达盒可包括两个拷贝的hv1核苷酸序列、三个拷贝的hv1核苷酸序列、四个拷贝的hv1核苷酸序列、五个拷贝的hv1核苷酸序列、六个拷贝的hv1核苷酸序列、七个拷贝的hv1核苷酸序列、八个拷贝的hv1核苷酸序列、九个拷贝的hv1核苷酸序列或十个拷贝的hv1核苷酸序列。
一方面,所述至少一个表达盒可包括一个拷贝的编码与gcw61细胞壁蛋白融合的ugt76g1酶(seqidno:3)的ugt76g1-gcw61核苷酸序列(seqidno:4)。在其它方面中,所述至少一个表达盒可包括两个拷贝的ugt76g1-gcw61核苷酸序列、三个拷贝的ugt76g1-gcw61核苷酸序列、四个拷贝的ugt76g1-gcw61核苷酸序列、五个拷贝的ugt76g1-gcw61核苷酸序列、六个拷贝的ugt76g1-gcw61核苷酸序列、七个拷贝的ugt76g1-gcw61核苷酸序列、八个拷贝的ugt76g1-gcw61核苷酸序列、九个拷贝的ugt76g1-gcw61核苷酸序列或十个拷贝的ugt76g1-gcw61核苷酸序列。
一方面,所述至少一个表达盒可包括一个拷贝的编码hv1-gcw61酶(seqidno:1)的hv1-gcw61核苷酸序列(seqidno:2)。在其它方面中,所述至少一个表达盒可包括两个拷贝的hv1-gcw61核苷酸序列、三个拷贝的hv1-gcw61核苷酸序列、四个拷贝的hv1-gcw61核苷酸序列、五个拷贝的hv1-gcw61核苷酸序列、六个拷贝的hv1-gcw61核苷酸序列、七个拷贝的hv1-gcw61核苷酸序列、八个拷贝的hv1-gcw61核苷酸序列、九个拷贝的hv1-gcw61核苷酸序列或十个拷贝的hv1-gcw61核苷酸序列。
一方面,所述至少一个表达盒可包括一个拷贝的编码mbsus1酶(seqidno:11)的mbsus1核苷酸序列(seqidno:12)。在其它方面中,所述至少一个表达盒可包括两个拷贝的mbsus1核苷酸序列、三个拷贝的mbsus1核苷酸序列、四个拷贝的mbsus1核苷酸序列、五个拷贝的mbsus1核苷酸序列、六个拷贝的mbsus1核苷酸序列、七个拷贝的mbsus1核苷酸序列、八个拷贝的mbsus1核苷酸序列、九个拷贝的mbsus1核苷酸序列或十个拷贝的mbsus1核苷酸序列。
一方面,表达盒可包括一个或多个拷贝的编码单个ugt序列的核苷酸序列,所述ugt序列编码选自如上文所述的ugt76g1、hv1或eugt11的ugt酶。另一方面,表达盒可包括一个或多个拷贝的选自ugt76g1、hv1、eugt11及其任何组合的两种或更多种ugt酶。在这样的其他方面,转化宿主细胞可根据需要表达两个或更多个类型的ugt酶。
一方面,表达载体可含多个表达盒,其中每个表达盒含单个拷贝的编码单个ugt序列或单个mbsus1序列的核苷酸序列,ugt序列编码选自如上文所述的ugt76g1、hv1或eugt11的ugt酶,mbsus1序列编码mbsus1酶。转化之后,可将多个表达盒整合到转化宿主细胞的基因组中。
生成甜菊醇糖苷的方法
在各个方面中,上文所述的全细胞催化剂可用于根据如本文所公开的方法由底物生成所需的甜菊醇糖苷化合物。图11是说明在一个方面中使用全细胞催化剂生成甜菊醇糖苷化合物的方法的方框图。参考图11,所述方法包括在步骤1102诱导编码至少一种ugt酶(ugt76g1、hv1和/或eugt11)和sus酶(mbsus1)的核苷酸序列在转化宿主细胞内表达。ugt和sus序列的表达产生全细胞催化剂,其特征在于多种ugt酶展示在细胞表面上和一定量的sus酶在全细胞催化剂内。
再参考图11,所述方法还包括在步骤1104用包括甜菊醇糖苷底物、udp或udp-葡萄糖和一定量蔗糖的培养基培育全细胞催化剂。如本文先前所述,ugt酶催化底物糖基化以生成所需的甜菊醇糖苷化合物。随时间推移,培养基内所含的底物转化为所需的甜菊醇糖苷化合物,其也含于培养基内。
图10说明了根据所述方法可以被培养基内的全细胞催化剂催化的示例性反应。在反应1中,培养基内的底物(reba)可经糖基化以生成rebd,正如使用udp-葡萄糖作为供体化合物通过ugt酶催化那样。在反应2中,反应1的udp副产物可与培养基内的蔗糖组合以再生额外udp-葡萄糖,用于根据反应1额外生成rebd。
再参考图11,在培养基中足以生成所需甜菊醇糖苷化合物的停留时间后,在步骤1106可以将至少一部分所需甜菊醇糖苷化合物从含全细胞催化剂的细胞培养物中分离。从水溶液中分离蛋白质的任何已知方法均可使用,且无限制。在一个非限制性实例中,可以使用高效液相色谱法(hplc)从细胞培养物中分离所需甜菊醇糖苷化合物。
在步骤1102中诱导的转化宿主细胞可以是使用上文所述任何材料和方法产生的任何转化宿主细胞。诱导转化宿主细胞的方法可根据至少几个因素中的任一个或多个而改变,包括但不限于:宿主细胞的类型、并入宿主细胞的所述至少一个表达盒的中包括的一个启动子或多个启动子,及任何其它相关因素。通过非限制性实例的方式,如果转化的宿主细胞是转化的巴斯德毕赤酵母菌株,其中引入的包括aox1启动子的表达盒可根据已知方法用一定量的足以诱导经由aox1启动子表达的甲醇来诱导。转化宿主细胞可诱导任何合适的时间段,且无限制,以确保生成足够量的所述至少一种ugt酶和所述sus酶。在各个方面中,转化宿主细胞可经诱导以表达ugt和sus酶至少1小时、至少2小时、至少4小时、至少6小时、至少8小时、至少10小时、至少12小时、至少16小时、至少20小时、至少24小时、至少30小时、至少36小时、至少42小时、至少48小时、至少60小时或至少72小时。
一方面,由步骤1102产生的全细胞催化剂可转移到含有待糖基化的底物的培养基中以根据上文所述的反应生成所需的甜菊醇糖苷化合物。一方面,全细胞催化剂可引入含培养基的体外反应体系中。任何已知的体外反应体系和培养基均可用于本文所述的方法中,且无限制。一方面,体外反应体系和培养基可根据至少几个因素中的任一个或多个进行选择,包括但不限于:宿主细胞类型、所需甜菊醇糖苷化合物的生产规模、待通过全细胞催化剂催化的特定糖基化反应及任何其它相关因素。通过非限制性实例的方式,如果全细胞催化剂是如上文所述的转化的毕赤酵母细胞,则可以使转化的毕赤酵母细胞悬浮在振荡式孵育器或含底物的培养基内的发酵罐中。
任何已知的培养基均可用于用底物培育全细胞催化剂,且无限制。在各个方面中,培养基的组成可根据至少几个因素中的任一个或多个变化,包括但不限于:宿主细胞类型、待通过全细胞催化剂催化的特定糖基化反应及任何其它相关因素。一方面,培养基可包括底物、蔗糖、udp-葡萄糖和udp。另一方面,培养基可包括额外化合物以维持全细胞催化剂的活力或以其它方式促进所需甜菊醇糖苷化合物的生成。合适的额外化合物的非限制性实例包括磷酸钾缓冲液、调节ph的化合物如酸或碱、mgcl2和任何其它合适的额外化合物。
可用含底物的培养基孵育全细胞催化剂任何合适的时间段,且无限制,以确保生成足够量的所需甜菊醇糖苷化合物。在各个方面中,全细胞催化剂可在培养基内孵育至少1小时、至少2小时、至少4小时、至少6小时、至少8小时、至少10小时、至少12小时、至少16小时、至少20小时、至少24小时、至少30小时、至少36小时、至少42小时、至少48小时、至少60小时或至少72小时。
所述方法的步骤1104中培养基内所包括的底物可根据ugt酶的类型和待生成的特定甜菊醇糖苷化合物来选择。一方面,底物可选自以下的任一种或多种:甜茶苷、莱鲍迪苷ka、甜菊苷、莱鲍迪苷a、莱鲍迪苷d和莱鲍迪苷e。参考图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h,这些底物可以根据示意性说明的任一个或多个反应糖基化以生成所需甜菊醇糖苷化合物,包括但不限于:莱鲍迪苷ka、莱鲍迪苷a、莱鲍迪苷d、莱鲍迪苷e和莱鲍迪苷m。可用于所述方法中的底物和全细胞催化剂的特定组合在下文中有更详细的描述。
由甜菊苷生成莱鲍迪苷a的方法。所公开的方法可用于由甜菊苷底物合成莱鲍迪苷a。所述方法包括用甜菊苷底物培养表达一定量的udp-糖基转移酶(ugt76g1)和一定量的蔗糖合成酶(mbsus1)的诱导的细胞足够的时间以生成莱鲍迪苷a,其中根据图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应1,葡萄糖与甜菊苷共价偶联以生成莱鲍迪苷a。一方面,全细胞催化剂可以是巴斯德毕赤酵母的诱导g/k4s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp或udp-葡萄糖及其任何组合。
由莱鲍迪苷d生成莱鲍迪苷m的方法。所公开的方法可用于由莱鲍迪苷d底物合成莱鲍迪苷m。所述方法包括用莱鲍迪苷d底物培养表达一定量的udp-糖基转移酶(ugt76g1)和一定量的蔗糖合成酶(mbsus1)的诱导的细胞足够的时间以生成莱鲍迪苷m底物,其中根据图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应3,葡萄糖与莱鲍迪苷d共价偶联以生成莱鲍迪苷m。一方面,全细胞催化剂可以是巴斯德毕赤酵母的诱导g/k4s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp或udp-葡萄糖及其任何组合。
由莱鲍迪苷e生成莱鲍迪苷d的方法。所公开的方法可用于由莱鲍迪苷e底物合成莱鲍迪苷d。所述方法包括用莱鲍迪苷e底物培养表达一定量的udp-糖基转移酶(ugt76g1)和一定量的蔗糖合成酶(mbsus1)的诱导的细胞足够的时间以生成莱鲍迪苷d,其中根据图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应2,葡萄糖与莱鲍迪苷e共价偶联以生成莱鲍迪苷d。一方面,全细胞催化剂可以是巴斯德毕赤酵母的g/k4s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp或udp-葡萄糖及其任何组合。
由莱鲍迪苷a生成莱鲍迪苷d的方法。所公开的方法可用于由莱鲍迪苷a底物合成莱鲍迪苷d。所述方法包括用莱鲍迪苷a底物培养表达一定量的udp-糖基转移酶(hv1)和一定量的蔗糖合成酶(mbsus1)的诱导的细胞足够的时间以生成莱鲍迪苷d,其中根据图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应4,葡萄糖与莱鲍迪苷a共价偶联以生成莱鲍迪苷d。一方面,全细胞催化剂可以是巴斯德毕赤酵母的g/h5s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp或udp-葡萄糖及其任何组合。
由甜菊苷生成莱鲍迪苷e的方法。所公开的方法可用于由甜菊苷底物合成莱鲍迪苷e。所述方法包括用甜菊苷底物培养表达一定量的udp-糖基转移酶(hv1)和一定量的蔗糖合成酶(mbsus1)的诱导的细胞足够的时间以生成莱鲍迪苷e,其中根据图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应5,葡萄糖与甜菊苷共价偶联以生成莱鲍迪苷e。一方面,全细胞催化剂可以是巴斯德毕赤酵母的g/h5s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp或udp-葡萄糖及其任何组合。
由甜菊苷和/或莱鲍迪苷a生成莱鲍迪苷m的方法。所公开的方法可用于由甜菊苷和/或莱鲍迪苷a底物合成莱鲍迪苷m。所述方法包括用甜菊苷和/或莱鲍迪苷a底物培养诱导的细胞足够的时间以生成莱鲍迪苷m,诱导的细胞包括表达一定量的udp-糖基转移酶(ugt76g1)和一定量的蔗糖合成酶(mbsus1)的第一诱导的细胞;和表达一定量的udp-糖基转移酶(hv1)和一定量的蔗糖合成酶(mbsus1)的第二诱导的细胞,其中根据图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应5、2和3和/或1、4和3,葡萄糖与甜菊苷和/或莱鲍迪苷a共价偶联以生成莱鲍迪苷m。一方面,第一诱导的细胞可为巴斯德毕赤酵母的g/k4s2菌株,第二诱导的细胞可为巴斯德毕赤酵母的g/h5s2菌株,并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp或udp-葡萄糖及其任何组合。
由莱鲍迪苷ka生成莱鲍迪苷e的方法。所公开的方法可用于由莱鲍迪苷ka底物合成莱鲍迪苷e。所述方法包括用莱鲍迪苷ka底物培养表达一定量的udp-糖基转移酶(hv1)和一定量的蔗糖合成酶(mbsus1)的诱导的细胞足够的时间以生成莱鲍迪苷e,其中根据图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应6,葡萄糖与莱鲍迪苷ka共价偶联以生成莱鲍迪苷e。一方面,全细胞催化剂可以是巴斯德毕赤酵母的g/h5s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp或udp-葡萄糖及其任何组合。
由甜茶苷生成莱鲍迪苷ka和莱鲍迪苷e的方法。所公开的方法可用于由甜茶苷底物合成莱鲍迪苷ka和莱鲍迪苷e。所述方法包括用甜茶苷底物培养表达一定量的udp-糖基转移酶(hv1)和一定量的蔗糖合成酶(mbsus1)的诱导的细胞足够的时间以生成莱鲍迪苷ka,其中根据图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应7,葡萄糖与甜茶苷共价偶联以生成莱鲍迪苷ka。此外,根据图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应6,葡萄糖与莱鲍迪苷ka共价价偶联以生成莱鲍迪苷e。一方面,全细胞催化剂可以是巴斯德毕赤酵母的g/h5s2菌株并且培养基还可包括:磷酸钾缓冲液、mgcl2、蔗糖、udp或udp-葡萄糖及其任何组合。
一方面,为生成所需甜菊醇糖苷化合物而进行的反应可按分批法进行,本文定义为向体外反应器引入离散量的全细胞催化剂并且从含有所需甜菊醇糖苷化合物的体外反应器中去除离散量的培养基。在这个方面,可以生成至少一个离散批次的所需甜菊醇糖苷化合物,或者可以通过多次更换培养基来重复所述分批法以生成多个离散批次的所需甜菊醇糖苷化合物。另一方面,为生成所需甜菊醇糖苷化合物而进行的反应可按连续进料法进行,本文定义为将一个菌落的全细胞催化剂保持在体外反应器中并且不断引入一定量含底物的培养基,同时不断地去除一定量含所需甜菊醇糖苷化合物的培养基。在两个方面中,分批或连续进料生物生产方法的任何已知方法均可使用,且无限制。
在各个方面中,对于牵涉多个糖基化反应的那些生成方法,例如由甜菊苷和/或莱鲍迪苷a底物生成莱鲍迪苷m,所有反应均可在如上文所述的相同体外反应器内进行。另一方面,多个糖基化反应中的每一个均可在至少两个相连的体外反应器之一中进行,其中含中间产物的细胞培养基可以从第一反应器转移到第二反应器。
通过非限制性实例的方式,根据图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应1、4和3,所公开的方法可用于在三个单独的体外反应器中由甜菊苷和/或莱鲍迪苷a合成莱鲍迪苷m。第一反应器可含表达一定量udp-糖基转移酶(ugt76g1)和一定量蔗糖合成酶(mbsus1)的全细胞催化剂。第二反应器可含表达一定量udp-糖基转移酶(hv1)和一定量蔗糖合成酶(mbsus1)的全细胞催化剂。第三反应器可含表达一定量udp-糖基转移酶(ugt76g1)和一定量蔗糖合成酶(mbsus1)的全细胞催化剂。用包括甜菊苷和/或莱鲍迪苷a底物的培养基培养足够时间以生成莱鲍迪苷m,其中根据图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应1、4和3,葡萄糖与甜菊苷和/或莱鲍迪苷a共价偶联以生成莱鲍迪苷m。第一反应器可进行图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应1以由甜菊苷底物生成莱鲍迪苷a。第二反应器可进行图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应4以由转移自第一反应器的莱鲍迪苷a生成莱鲍迪苷d。第三反应器可进行图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应3以由转移自第二反应器的莱鲍迪苷d生成莱鲍迪苷m。
通过非限制性实例的方式,根据图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应5、2和3,所公开的方法可用于在三个单独的体外反应器中由甜菊苷和/或莱鲍迪苷a合成莱鲍迪苷m。第一反应器可含表达一定量udp-糖基转移酶(hv1)和一定量蔗糖合成酶(mbsus1)的全细胞催化剂。第二反应器可含表达一定量udp-糖基转移酶(ugt76g1)和一定量蔗糖合成酶(mbsus1)的全细胞催化剂。第三反应器可含表达一定量udp-糖基转移酶(ugt76g1)和一定量蔗糖合成酶(mbsus1)的全细胞催化剂,用包括甜菊苷和/或莱鲍迪苷a底物的培养基培养足够时间以生成莱鲍迪苷m,其中根据图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应5、2和3,葡萄糖与甜菊苷共价偶联以生成莱鲍迪苷m。第一反应器可进行图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应5以由甜菊苷底物生成莱鲍迪苷e。第二反应器可进行图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应2以由转移自第一反应器的莱鲍迪苷e生成莱鲍迪苷d。第三反应器可进行图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应3以由转移自第二反应器的莱鲍迪苷d生成莱鲍迪苷m。
在这个非限制性实例中,每个反应器均可提供经单独调整以增强每个反应器的产物生成的反应条件。可单独调整的反应条件的非限制性实例包括:ph、温度、搅拌、培养基的组成、全细胞催化剂的密度及任何其它相关的反应条件。另外,每种全细胞催化剂的构型可单独调节以增强每个反应器的产物生成。调节每种全细胞催化剂构型的方法的非限制性实例包括:调节诱导期,改变转化宿主细胞的所述至少一个表达盒内的ugt和/或sus序列的拷贝数,改变宿主细胞的类型,及任何其它合适的方法。
在考虑了以下非限制性实施例后本公开将得到更全面的理解。
实施例
以下实施例证明了本公开的各个方面。
实施例1:巴斯德毕赤酵母转化产生工程化全细胞催化剂菌株。
为证明巴斯德毕赤酵母细胞转化产生几个适合用作全细胞催化剂的工程化毕赤酵母菌株,进行了以下实验。
合成候选ugt基因的全长dna片段用于巴斯德毕赤酵母细胞的转化。具体地,cdna经密码子优化用于巴斯德毕赤酵母表达(genscript,piscataway,nj)以生成两个udp-糖基转移酶序列:hv1(seqidno:8)和ugt76g1(seqidno:10)。在编码α因子信号肽的核苷酸序列和编码hv1(seqidno:8)或ugt76g1(seqidno:10)的核苷酸序列之间的框内插入flag标签。另外,在hv1或ugt76g1序列c端的框内插入编码毕赤酵母细胞壁蛋白的基因(gcw61,seqidno:6)以产生两个融合基因:α因子信号肽-flag-hv1-gcw61和α因子信号肽-flag-ugt76g1-gcw61。将合成的融合基因克隆到phka载体,即经修饰的毕赤酵母表达载体(ppiczαa,invitrogen)中。另外,将密码子优化的蔗糖合成酶cdna(mbsus1,seqidno:12)克隆到毕赤酵母表达载体ppiczαa(invitrogen)中。载体中,每个表达盒含有aox1启动子、基因和aox1转录终止子。
为生成多个拷贝的表达盒,用bamhi和bglii消化以上构建体。收集表达盒并插入经bamhi消化的质粒中。消化分析后,鉴定具有5个拷贝的hv1表达盒的质粒、具有4个拷贝的ugt76g1表达盒的质粒和具有2个拷贝的mbsus表达盒的质粒。
使用已知方法将线性化表达质粒转化到巴斯德毕赤酵母(gs115)细胞内并且将表达盒整合到毕赤酵母属基因组中。筛选后,鉴定阳性菌株,如表2所汇总的。
表2.巴斯德毕赤酵母菌株的汇总
实施例2:使用工程化巴斯德毕赤酵母菌株g/k4s2糖基化甜菊苷以生成莱鲍迪苷a
为证明使用甜菊苷底物由诱导的g/k4s2细胞酶促生成莱鲍迪苷a,进行了以下实验。将单个菌落的巴斯德毕赤酵母菌株g/k4s2接种在带挡板的烧瓶中的bmgy培养基中并使其在振荡式孵育器(250–300rpm)中在28–30℃下生长,直至培养物达到2-6的od600(对数期生长)。通过离心收获g/k4s2细胞并且重新悬浮在bmmy培养基中达到1.0的od600以诱导表达。每24小时向bmmy培养基中添加100%甲醇达到1%甲醇的最终浓度以维持表达的诱导。诱导后72小时,通过离心收获g/k4s2细胞并进行如下文所述的糖基化活性分析。
使用甜菊苷作为底物测定诱导的g/k4s2细胞的糖基化活性。在200μl含50mm磷酸钾缓冲液(ph7.2)、3mmmgcl2、3mg/ml甜菊苷(含95%甜菊苷的甜叶菊提取物;bluecalifornia,ca)、1mmudp或udp-葡萄糖(udpg)和250mm蔗糖的体外反应体系中测试诱导的g/k4s2细胞(60od)。将诱导的g/k4s2细胞保持在28-30℃的反应体系内24小时,之后通过添加1-丁醇来终止反应。用200μl1-丁醇萃取来自反应体系的样品三次。汇合的样品萃取成分经干燥并溶于80%甲醇中用于高效液相色谱(hplc)分析。
使用uplcultimate3000系统(dionex,sunnyvale,ca)进行hplc分析,所述系统包括四元泵、温控柱室、自动取样器和紫外吸光度检测器。具有保护柱的synergihydro-rp柱(phenomenex)用于表征汇合样品中的甜菊醇糖苷。水中的乙腈用于hplc分析中的等度洗脱。用于hplc分析的检测波长为210nm。
图2c是汇总在诱导的g/k4s2细胞引入后24小时从体外反应体系取样,由诱导的g/k4s2细胞酶促生成的莱鲍迪苷a的hplc检测的图表。
该实验的结果证明诱导的g/k4s2细胞酶促糖基化甜菊苷以生成莱鲍迪苷a。
实施例3:使用工程化巴斯德毕赤酵母菌株g/k4s2糖基化莱鲍迪苷d以生成莱鲍迪苷m
为证明使用莱鲍迪苷d底物由诱导的g/k4s2细胞酶促生成莱鲍迪苷m,进行了以下实验。在巴斯德毕赤酵母菌株g/k4s2的培养物中诱导表达72小时并且使用类似于实施例2所述的方法收获诱导的细胞。使用类似于实施例2所述的方法测定诱导的g/k4s2细胞对莱鲍迪苷d底物的糖基化。向200μl含50mm磷酸钾缓冲液(ph7.2)、3mmmgcl2、0.5mg/ml莱鲍迪苷d(98%纯度,bluecalifornia,ca)、1mmudp或udpg和250mm蔗糖的体外反应体系中引入诱导的g/k4s2细胞(60od)。将诱导的g/k4s2细胞保持在28-30℃的反应体系内24小时,之后通过添加1-丁醇来终止反应。提取来自反应体系的样品并且使用类似于实施例2所述的方法进行高效液相色谱(hplc)分析。
图3c是汇总在诱导的g/k4s2细胞引入后24小时从体外反应体系取样,由诱导的g/k4s2细胞酶促生成的莱鲍迪苷m的hplc检测的图表。
该实验的结果证明诱导的g/k4s2细胞酶促糖基化莱鲍迪苷d以生成莱鲍迪苷m。
实施例4:使用工程化巴斯德毕赤酵母菌株g/k4s2糖基化莱鲍迪苷e以生成莱鲍迪苷d
为证明使用莱鲍迪苷e底物由诱导的g/k4s2细胞酶促生成莱鲍迪苷d,进行了以下实验。在巴斯德毕赤酵母菌株g/k4s2的培养物中诱导表达72小时并且使用类似于实施例2所述的方法收获诱导的细胞。使用类似于实施例2所述的方法测定诱导的g/k4s2细胞对莱鲍迪苷e底物的糖基化。在200μl含50mm磷酸钾缓冲液(ph7.2)、3mmmgcl2、3mg/ml莱鲍迪苷e(95%bluecalifornia,ca)、1mmudp或udpg和250mm蔗糖的体外反应体系中测试诱导的g/k4s2细胞(60od)。将诱导的g/k4s2细胞保持在28-30℃的反应体系内24小时,之后通过添加1-丁醇来终止反应。提取来自反应体系的样品并且使用类似于实施例2所述的方法进行高效液相色谱(hplc)分析。
图4c是汇总在诱导的g/k4s2细胞引入后24小时从体外反应体系取样,由诱导的g/k4s2细胞酶促生成的莱鲍迪苷d的hplc检测的图表。
该实验的结果证明g/k4s2细胞酶促糖基化莱鲍迪苷e以生成莱鲍迪苷d。
实施例5:使用工程化巴斯德毕赤酵母菌株g/h5s2糖基化莱鲍迪苷a以生成莱鲍迪苷d
为证明使用莱鲍迪苷a底物由诱导的g/h5s2细胞酶促生成莱鲍迪苷d,进行了以下实验。在巴斯德毕赤酵母菌株g/h5s2中诱导表达72小时并且使用类似于实施例2所述的方法收获诱导的细胞。使用类似于实施例2所述的方法,使用莱鲍迪苷a作为底物测定诱导的g/h5s2细胞的糖基化活性。在200μl含50mm磷酸钾缓冲液(ph7.2)、3mmmgcl2、3mg/ml莱鲍迪苷a(99%bluecalifornia,ca)、1mmudp或udpg和250mm蔗糖的体外反应体系中测试诱导的g/h5s2细胞(60od)。将诱导的g/h5s2细胞保持在28-30℃的反应体系内24小时,之后通过添加1-丁醇来终止反应。提取来自反应体系的样品并且使用类似于实施例2所述的方法进行高效液相色谱(hplc)分析。为了比较,对两种另外的诱导巴斯德毕赤酵母菌株进行类似的测定和hplc分析:其中仅引入了hv1序列的h5菌株,及其中引入了空载体的phka(对照)菌株。
图5c、图5d和图5e是汇总在诱导的细胞引入后24小时从体外反应体系取样,分别由诱导的g/h5s2细胞、诱导的h5细胞和诱导的phka细胞酶促生成的莱鲍迪苷d的hplc检测的图表。如图5c和5d所示,诱导的g/h5s2细胞(图5c)和诱导的h5细胞(图5d)两者酶促生成莱鲍迪苷d,但诱导的g/h5s2细胞表现出更高的诱导的g/h5s2活性,表明在诱导的g/h5s2细胞中产生了ugt-sus偶联体系(参见图10)。由于这种ugt-sus偶联体系,如图10所示,诱导的g/h5s2细胞重复利用消耗的udp,而向诱导的h5细胞中添加额外udpg以使得能够生成额外莱鲍迪苷d。诱导的phka细胞未酶促生成莱鲍迪苷d,证明在不存在hv1序列时未生成莱鲍迪苷d。
该实验的结果证明g/h5s2细胞酶促糖基化莱鲍迪苷a以生成莱鲍迪苷d。
实施例6:使用工程化巴斯德毕赤酵母菌株g/h5s2糖基化甜菊苷以生成莱鲍迪苷e
为证明使用甜菊苷底物由g/h5s2细胞酶促生成莱鲍迪苷e,进行了以下实验。在巴斯德毕赤酵母菌株g/h5s2的培养物中诱导表达72小时并且使用类似于实施例2所述的方法收获诱导的细胞。使用类似于实施例2所述的方法,使用甜菊苷作为底物测定诱导的g/h5s2细胞的糖基化活性。在200μl含50mm磷酸钾缓冲液(ph7.2)、3mmmgcl2、3mg/ml甜菊苷(95%,bluecalifornia,ca)、1mmudp或udpg和250mm蔗糖的体外反应体系中测试诱导的g/h5s2细胞(60od)。将诱导的g/h5s2细胞保持在28-30℃的反应体系内24小时,之后通过添加1-丁醇来终止反应。提取来自反应体系的样品并且使用类似于实施例2所述的方法进行高效液相色谱(hplc)分析。为了比较,对其中引入了空载体的phka(对照)菌株进行类似的测定和hplc分析。
图6c和图6d是汇总在诱导的细胞引入后24小时从体外反应体系取样,分别由诱导的g/h5s2细胞和诱导的phka细胞酶促生成的莱鲍迪苷e的hplc检测的图表。如图6c所示,诱导的g/h5s2细胞酶促生成莱鲍迪苷e。如图6d所示,诱导的phka细胞未能生成莱鲍迪苷e。
该实验的结果证明g/h5s2细胞酶促糖基化甜菊苷以生成莱鲍迪苷e。
实施例7:使用工程化巴斯德毕赤酵母菌株g/h5s2和g/k4s2糖基化莱鲍迪苷a以生成莱鲍迪苷m。
为证明使用莱鲍迪苷a底物由诱导的g/k4s2细胞和诱导的g/h5s2细胞的组合酶促生成莱鲍迪苷m和莱鲍迪苷d,进行了以下实验。在巴斯德毕赤酵母菌株g/k4s2和g/h5s2的培养物中诱导表达72小时并且使用类似于实施例2所述的方法收获诱导的细胞。使用类似于实施例2所述的方法,使用莱鲍迪苷a作为底物测定诱导的g/k4s2和g/h5s2细胞的糖基化活性。在200μl含50mm磷酸钾缓冲液(ph7.2)、3mmmgcl2、1mg/ml莱鲍迪苷a(99%纯度,bluecalifornia,ca)、1mmudp或udpg和250mm蔗糖的体外反应体系中测试诱导的g/k4s2和g/h5s2细胞(60od)。将诱导的g/k4s2和g/h5s2细胞保持在28-30℃的反应体系内24小时和48小时,之后通过添加1-丁醇来终止反应。提取来自反应体系的样品并且使用类似于实施例2所述的方法进行高效液相色谱(hplc)分析。
图7d和图7e是汇总在诱导的g/k4s2和g/h5s2细胞引入后24小时(图7d)和48小时(图7e)从体外反应体系取样,由诱导的g/k4s2细胞和g/h5s2细胞酶促生成的莱鲍迪苷d和莱鲍迪苷m的hplc检测的图表。
该实验的结果证明诱导的g/k4s2和g/h5s2细胞的组合酶促糖基化莱鲍迪苷a以生成莱鲍迪苷m。rebd和rebm由诱导的g/k4s2和g/h5s2细胞生成,根据反应表明在反应中的hv1和ugt76g1糖基化活性。g/h5s2细胞最初将莱鲍迪苷a底物转化为莱鲍迪苷d,如图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应4所示,并且g/k4s2细胞催化莱鲍迪苷d转化为莱鲍迪苷m,如图9a、图9b、图9c、图9d、图9e、图9f、图9g和图9h的反应3所示。
实施例8:使用工程化巴斯德毕赤酵母菌株g/h5s2和g/k4s2糖基化莱鲍迪苷a以生成莱鲍迪苷m-g/h5s2:g/k4s2比率的影响
为证明使用莱鲍迪苷a底物在不同的g/k4s2和g/h5s2比率下,由诱导的g/k4s2细胞和诱导的g/h5s2细胞的组合酶促生成莱鲍迪苷m和莱鲍迪苷d的敏感性,进行了以下实验。对于g/h5s2:g/k4s2细胞的几个比率:2:1、1:1、1:2和1:3,使用类似于实施例7的方法评估使用莱鲍迪苷a作为底物,诱导的g/k4s2和g/h5s2细胞的组合的糖基化活性。另外,对于每个g/h5s2:g/k4s2细胞比率而言,在16小时、24小时和48小时的反应时间后从反应体系提取样品。所有样品使用类似于实施例2所述的方法进行高效液相色谱(hplc)分析。
图8a和图8b是汇总在诱导的g/k4s2和g/h5s2细胞引入后不同时间从体外反应体系取样,在不同g/h5s2:g/k4s2细胞比率下由诱导的g/k4s2细胞和g/h5s2细胞分别酶促生成的莱鲍迪苷d和莱鲍迪苷m的hplc检测的图表。如图8a所示,g/h5s2:g/k4s2比率越高可引起越多的rebd积聚。如图8b所示,g/h5s2:g/k4s2比率越低可引起越多的rebd转化为rebm。
该实验的结果证明g/k4s2和g/h5s2细胞的组合酶促糖基化莱鲍迪苷a以生成莱鲍迪苷m对g/h5s2:g/k4s2细胞比率敏感。
实施例9:使用工程化巴斯德毕赤酵母菌株g/h5s2和g/k4s2糖基化甜菊苷以生成莱鲍迪苷m。
为证明使用甜菊苷底物由诱导的g/k4s2细胞和诱导的g/h5s2细胞的组合酶促生成莱鲍迪苷m,进行了以下实验。在巴斯德毕赤酵母菌株g/k4s2和g/h5s2的培养物中诱导表达72小时并且使用类似于实施例2所述的方法收获诱导的细胞。使用类似于实施例2所述的方法,使用甜菊苷作为底物测定诱导的g/k4s2和g/h5s2细胞的糖基化活性。在200μl含50mm磷酸钾缓冲液(ph7.2)、3mmmgcl2、1mg/ml甜菊苷(95%纯度,bluecalifornia,ca)、1mmudp或udpg和250mm蔗糖的体外反应体系中测试诱导的g/k4s2和g/h5s2细胞(60od)。将诱导的g/k4s2和g/h5s2细胞保持在28-30℃的反应体系内10小时和24小时,之后通过添加1-丁醇来终止反应。提取来自反应体系的样品并且使用类似于实施例2所述的方法进行高效液相色谱(hplc)分析。
图12c和图12d是汇总在诱导的g/k4s2和g/h5s2细胞引入后10小时(图12c)和24小时(图12d)从体外反应体系取样,由诱导的g/k4s2细胞和g/h5s2细胞酶促生成的莱鲍迪苷d和莱鲍迪苷m的hplc检测的图表。
该实验的结果证明诱导的g/k4s2和g/h5s2细胞的组合酶促糖基化甜菊苷以生成莱鲍迪苷m。
实施例10:使用工程化巴斯德毕赤酵母菌株g/h5s2糖基化莱鲍迪苷ka以生成莱鲍迪苷e。
为证明使用莱鲍迪苷ka底物由g/h5s2细胞酶促生成莱鲍迪苷e,进行了以下实验。在巴斯德毕赤酵母菌株g/h5s2的培养物中诱导表达72小时并且使用类似于实施例2所述的方法收获诱导的细胞。使用类似于实施例2所述的方法,使用莱鲍迪苷ka作为底物测定诱导的g/h5s2细胞的糖基化活性。在200μl含50mm磷酸钾缓冲液(ph7.2)、3mmmgcl2、1mg/ml莱鲍迪苷ka(98%,bluecalifornia,ca)、1mmudp或udpg和250mm蔗糖的体外反应体系中测试诱导的g/h5s2细胞(60od)。将诱导的g/h5s2细胞保持在28-30℃的反应体系内24小时,之后通过添加1-丁醇来终止反应。提取来自反应体系的样品并且使用类似于实施例2所述的方法进行高效液相色谱(hplc)分析。
图13b是汇总在诱导的细胞引入后24小时从体外反应体系取样,由诱导的g/h5s2酶促生成的莱鲍迪苷e的hplc检测的图表。如图13b所示,诱导的g/h5s2细胞酶促生成莱鲍迪苷e。
该实验的结果证明g/h5s2细胞酶促糖基化莱鲍迪苷ka以生成莱鲍迪苷e。
实施例10:使用工程化巴斯德毕赤酵母菌株g/h5s2糖基化甜茶苷以生成莱鲍迪苷ka和莱鲍迪苷e。
为证明使用甜茶苷底物由g/h5s2细胞酶促生成莱鲍迪苷ka,进行了以下实验。在巴斯德毕赤酵母菌株g/h5s2的培养物中诱导表达72小时并且使用类似于实施例2所述的方法收获诱导的细胞。使用类似于实施例2所述的方法,使用甜茶苷作为底物测定诱导的g/h5s2细胞的糖基化活性。在200μl含50mm磷酸钾缓冲液(ph7.2)、3mmmgcl2、1mg/ml甜茶苷(98%,bluecalifornia,ca)、1mmudp或udpg和250mm蔗糖的体外反应体系中测试诱导的g/h5s2细胞(60od)。将诱导的g/h5s2细胞保持在28-30℃的反应体系内24小时,之后通过添加1-丁醇来终止反应。提取来自反应体系的样品并且使用类似于实施例2所述的方法进行高效液相色谱(hplc)分析。
图14b和图14c是汇总在诱导的细胞引入后14和24小时从体外反应体系取样,由诱导的g/h5s2酶促生成的莱鲍迪苷ka的hplc检测的图表。如图14b和图14c所示,诱导的g/h5s2细胞酶促生成莱鲍迪苷ka和莱鲍迪苷e。生成的莱鲍迪苷ka可转化为莱鲍迪苷e。
该实验的结果证明g/h5s2细胞酶促糖基化甜茶苷以生成莱鲍迪苷ka和莱鲍迪苷e。
鉴于以上所述,将会看到实现了本公开的几个优点并获得了其它有利结果。因为在不背离本公开范围的前提下可以在以上方法和体系上进行各种变化,所以意图是应将以上描述中所含的及附图中所示的所有问题均解释为说明性而非限制性意义。
在介绍本公开或其型式、实施方案或方面的要素时,冠词“一种”、“一个”、“该”和“所述”旨在意指存在一个或多个要素。术语“包含”、“包括”和“具有”旨在为包括性且意指可能存在除所列要素之外的额外要素。
序列表
<110>conageninc.
<120>使用工程化全细胞催化剂生产无热量甜味剂
<130>32559-24
<160>12
<170>patentin3.5版
<210>1
<211>508
<212>prt
<213>人工序列
<220>
<223>合成的
<400>1
metaspglyasnserserserserproleuhisvalvalilecyspro
151015
trpleualaleuglyhisleuleuprocysleuaspilealagluarg
202530
leualaserargglyhisargvalserphevalserthrproargasn
354045
ilealaargleuproproleuargproalavalalaproleuvalasp
505560
phevalalaleuproleuprohisvalaspglyleuprogluglyala
65707580
gluserthrasnaspvalprotyrasplysphegluleuhisarglys
859095
alapheaspglyleualaalapropheserglupheleuargalaala
100105110
cysalagluglyalaglyserargproasptrpleuilevalaspthr
115120125
phehishistrpalaalaalaalaalavalgluasnlysvalprocys
130135140
valmetleuleuleuglyalaalathrvalilealaglyphealaarg
145150155160
glyvalsergluhisalaalaalaalavalglylysgluargproala
165170175
alaglualaproserphegluthrgluargarglysleumetthrthr
180185190
glnasnalaserglymetthrvalalagluargtyrpheleuthrleu
195200205
metargseraspleuvalalaileargsercysalaglutrpglupro
210215220
gluservalalaalaleuthrthrleualaglylysprovalvalpro
225230235240
leuglyleuleuproproserprogluglyglyargglyvalserlys
245250255
gluaspalaalavalargtrpleuaspalaglnproalalysserval
260265270
valtyrvalalaleuglysergluvalproleuargalagluglnval
275280285
hisgluleualaleuglyleugluleuserglyalaargpheleutrp
290295300
alaleuarglysprothraspalaproaspalaalavalleupropro
305310315320
glypheglugluargthrargglyargglyleuvalvalthrglytrp
325330335
valproglnileglyvalleualahisglyalavalalaalapheleu
340345350
thrhiscysglytrpasnserthrilegluglyleuleupheglyhis
355360365
proleuilemetleuproileserseraspglnglyproasnalaarg
370375380
leumetgluglyarglysvalglymetglnvalproargaspgluser
385390395400
aspglyserpheargarggluaspvalalaalathrvalargalaval
405410415
alavalglugluaspglyargargvalphethralaasnalalyslys
420425430
metglngluilevalalaaspglyalacyshisgluargcysileasp
435440445
glypheileglnglnleuargsertyrlysalathrargasnasnleu
450455460
serasngluserasnglythrasnhisserasnhisthrserserval
465470475480
prothrglyalaalavalargalaserglymetglyalaglyleuleu
485490495
glyalaglyvalvalalaglyvalalaleuleuile
500505
<210>2
<211>1527
<212>dna
<213>人工序列
<220>
<223>合成的
<400>2
atggatggtaactcctcttcttcccctcttcacgtcgtcatttgcccttggcttgctctt60
ggacacttgttgccttgcttggacattgcagagagattggcttccagaggtcatagagtt120
tcatttgtcagtaccccaagaaacattgctagacttccacctttgagaccagctgttgcc180
cctttggttgatttcgtcgctcttccattgcctcatgttgacggtttgccagagggagct240
gaatcaacaaatgatgttccttacgacaagtttgaacttcacagaaaggcttttgatggt300
ttggctgccccattttctgagttccttagagcagcttgtgctgaaggtgccggttctaga360
ccagattggttgattgttgatacttttcatcactgggccgcagctgccgcagttgagaac420
aaggttccttgcgtcatgttgcttttgggtgctgccacagttatcgcaggtttcgctaga480
ggagtctctgaacacgcagctgccgcagttggaaaggagagaccagctgccgaagctcct540
tcatttgaaactgagagaagaaaattgatgactacacaaaatgcaagtggtatgactgtt600
gctgaaagatacttccttacattgatgagatcagatttggttgccatcagaagttgtgca660
gaatgggagccagaatctgttgcagctcttaccactttggctggtaaaccagttgtccct720
ttgggacttttgccaccttctcctgagggtggaagaggtgtttccaaggaagatgccgca780
gtcagatggttggacgcccaaccagcaaaatcagttgtctatgttgctcttggaagtgag840
gtccctttgagagccgagcaggttcatgaacttgctttgggtcttgaattgtctggagct900
agatttctttgggccttgagaaaaccaactgatgctcctgacgctgccgttttgccacct960
ggtttcgaagagagaaccagaggtagaggattggttgtcactggatgggttccacaaatt1020
ggtgtcttggctcatggagccgttgcagctttcttgacacactgcggttggaactccacc1080
atcgagggtcttttgttcggacacccacttattatgttgcctatctcttccgatcaaggt1140
ccaaatgctagattgatggaaggtagaaaagttggaatgcaggtccctagagatgagtct1200
gacggttcttttagaagagaagatgttgccgcaacagtcagagcagttgctgtcgaagag1260
gacggtagaagagtcttcaccgccaacgcaaagaaaatgcaggaaattgttgctgacgga1320
gcctgtcacgaaagatgtattgatggttttatccagcagttgagaagttacaaagcaacg1380
cgtaacaacctatcaaacgagagtaatggtactaatcactccaaccatacttcttccgtg1440
ccaactggagctgccgttcgtgcctctggtatgggagctggcttgttgggagctggtgtt1500
gtagccggtgttgctctattgatttaa1527
<210>3
<211>507
<212>prt
<213>人工序列
<220>
<223>合成的
<400>3
metgluasnlysthrgluthrthrvalargargargargargileile
151015
leupheprovalpropheglnglyhisileasnproileleuglnleu
202530
alaasnvalleutyrserlysglypheserilethrilephehisthr
354045
asnpheasnlysprolysthrserasntyrprohisphethrphearg
505560
pheileleuaspasnaspproglnaspgluargileserasnleupro
65707580
thrhisglyproleualaglymetargileproileileasngluhis
859095
glyalaaspgluleuargarggluleugluleuleumetleualaser
100105110
glugluaspglugluvalsercysleuilethraspalaleutrptyr
115120125
phealaglnservalalaaspserleuasnleuargargleuvalleu
130135140
metthrserserleupheasnphehisalahisvalserleuprogln
145150155160
pheaspgluleuglytyrleuaspproaspasplysthrargleuglu
165170175
gluglnalaserglypheprometleulysvallysaspilelysser
180185190
alatyrserasntrpglnileleulysgluileleuglylysmetile
195200205
lysglnthrlysalaserserglyvaliletrpasnserphelysglu
210215220
leugluglusergluleugluthrvalilearggluileproalapro
225230235240
serpheleuileproleuprolyshisleuthralaserserserser
245250255
leuleuasphisaspargthrvalpheglntrpleuaspglnglnpro
260265270
proserservalleutyrvalserpheglyserthrsergluvalasp
275280285
glulysasppheleugluilealaargglyleuvalaspserlysgln
290295300
serpheleutrpvalvalargproglyphevallysglyserthrtrp
305310315320
valgluproleuproaspglypheleuglygluargglyargileval
325330335
lystrpvalproglnglngluvalleualahisglyalaileglyala
340345350
phetrpthrhisserglytrpasnserthrleugluservalcysglu
355360365
glyvalprometilepheserasppheglyleuaspglnproleuasn
370375380
alaargtyrmetseraspvalleulysvalglyvaltyrleugluasn
385390395400
glytrpgluargglygluilealaasnalaileargargvalmetval
405410415
aspglugluglyglutyrileargglnasnalaargvalleulysgln
420425430
lysalaaspvalserleumetlysglyglysersertyrgluserleu
435440445
gluserleuvalsertyrileserserleuthrargasnasnleuser
450455460
asngluserasnglythrasnhisserasnhisthrserservalpro
465470475480
thrglyalaalavalargalaserglymetglyalaglyleuleugly
485490495
alaglyvalvalalaglyvalalaleuleuile
500505
<210>4
<211>1524
<212>dna
<213>人工序列
<220>
<223>合成的
<400>4
atggagaataagaccgagactaccgtcagaagaagaagaagaatcattttgttcccagtt60
cctttccaaggtcacatcaaccctatcttgcaacttgctaacgttttgtacagtaaggga120
ttctctatcactatcttccatacaaacttcaataagccaaagacctctaattaccctcac180
ttcacttttagattcattttggataacgacccacaggatgaaagaatctccaatttgcca240
actcatggacctcttgctggtatgagaatcccaattatcaacgaacacggtgccgacgag300
ttgagaagagaacttgagttgcttatgttggcctcagaagaggatgaagaggttagttgt360
ttgatcacagacgcactttggtactttgcacaatccgttgctgattcattgaaccttaga420
agattggtccttatgacctcttccttgttcaattttcatgctcacgtttcattgccacaa480
ttcgatgaattgggataccttgaccctgatgacaaaactagattggaagagcaggcttcc540
ggtttcccaatgttgaaggttaaggatatcaagtctgcctattccaactggcaaattttg600
aaggaaatccttggaaagatgattaaacagaccaaggcttcaagtggtgttatctggaac660
tcttttaaggaattggaggagtctgaacttgagactgtcattagagagatcccagcacct720
tctttcttgattccacttcctaaacatttgactgcttcttcctcaagtttgcttgatcac780
gacagaacagtttttcaatggttggatcaacagccaccttcttccgttttgtacgtctct840
ttcggttctacttctgaagttgatgagaaagactttttggaaattgctagaggtcttgtc900
gattcaaagcagagtttcttgtgggttgtcagaccaggatttgttaaaggttctacatgg960
gtcgaaccattgcctgacggtttccttggagagagaggtagaattgttaagtgggtccct1020
caacaggaagttttggcacatggagctatcggtgccttttggacacacagtggttggaac1080
tctaccttggaatccgtttgcgagggagtcccaatgattttctccgattttggtttggac1140
caacctcttaacgctagatatatgtctgatgttttgaaggttggagtctatcttgaaaac1200
ggatgggaaagaggagagattgctaatgccatcagaagagttatggtcgatgaagaggga1260
gagtatattagacaaaatgccagagttttgaagcagaaagcagacgtctctttgatgaaa1320
ggtggatcaagttacgaatctttggagtcccttgtttcatatatctcttccttgacgcgt1380
aacaacctatcaaacgagagtaatggtactaatcactccaaccatacttcttccgtgcca1440
actggagctgccgttcgtgcctctggtatgggagctggcttgttgggagctggtgttgta1500
gccggtgttgctctattgatttaa1524
<210>5
<211>47
<212>prt
<213>人工序列
<220>
<223>合成的
<400>5
asnasnleuserasngluserasnglythrasnhisserasnhisthr
151015
serservalprothrglyalaalavalargalaserglymetglyala
202530
glyleuleuglyalaglyvalvalalaglyvalalaleuleuile
354045
<210>6
<211>150
<212>dna
<213>人工序列
<220>
<223>合成的
<400>6
acgcgtaacaacctatcaaacgagagtaatggtactaatcactccaaccatacttcttcc60
gtgccaactggagctgccgttcgtgcctctggtatgggagctggcttgttgggagctggt120
gttgtagccggtgttgctctattgatttaa150
<210>7
<211>459
<212>prt
<213>人工序列
<220>
<223>合成的
<400>7
metaspglyasnserserserserproleuhisvalvalilecyspro
151015
trpleualaleuglyhisleuleuprocysleuaspilealagluarg
202530
leualaserargglyhisargvalserphevalserthrproargasn
354045
ilealaargleuproproleuargproalavalalaproleuvalasp
505560
phevalalaleuproleuprohisvalaspglyleuprogluglyala
65707580
gluserthrasnaspvalprotyrasplysphegluleuhisarglys
859095
alapheaspglyleualaalapropheserglupheleuargalaala
100105110
cysalagluglyalaglyserargproasptrpleuilevalaspthr
115120125
phehishistrpalaalaalaalaalavalgluasnlysvalprocys
130135140
valmetleuleuleuglyalaalathrvalilealaglyphealaarg
145150155160
glyvalsergluhisalaalaalaalavalglylysgluargproala
165170175
alaglualaproserphegluthrgluargarglysleumetthrthr
180185190
glnasnalaserglymetthrvalalagluargtyrpheleuthrleu
195200205
metargseraspleuvalalaileargsercysalaglutrpglupro
210215220
gluservalalaalaleuthrthrleualaglylysprovalvalpro
225230235240
leuglyleuleuproproserprogluglyglyargglyvalserlys
245250255
gluaspalaalavalargtrpleuaspalaglnproalalysserval
260265270
valtyrvalalaleuglysergluvalproleuargalagluglnval
275280285
hisgluleualaleuglyleugluleuserglyalaargpheleutrp
290295300
alaleuarglysprothraspalaproaspalaalavalleupropro
305310315320
glypheglugluargthrargglyargglyleuvalvalthrglytrp
325330335
valproglnileglyvalleualahisglyalavalalaalapheleu
340345350
thrhiscysglytrpasnserthrilegluglyleuleupheglyhis
355360365
proleuilemetleuproileserseraspglnglyproasnalaarg
370375380
leumetgluglyarglysvalglymetglnvalproargaspgluser
385390395400
aspglyserpheargarggluaspvalalaalathrvalargalaval
405410415
alavalglugluaspglyargargvalphethralaasnalalyslys
420425430
metglngluilevalalaaspglyalacyshisgluargcysileasp
435440445
glypheileglnglnleuargsertyrlysala
450455
<210>8
<211>1377
<212>dna
<213>人工序列
<220>
<223>合成的
<400>8
atggatggtaactcctcttcttcccctcttcacgtcgtcatttgcccttggcttgctctt60
ggacacttgttgccttgcttggacattgcagagagattggcttccagaggtcatagagtt120
tcatttgtcagtaccccaagaaacattgctagacttccacctttgagaccagctgttgcc180
cctttggttgatttcgtcgctcttccattgcctcatgttgacggtttgccagagggagct240
gaatcaacaaatgatgttccttacgacaagtttgaacttcacagaaaggcttttgatggt300
ttggctgccccattttctgagttccttagagcagcttgtgctgaaggtgccggttctaga360
ccagattggttgattgttgatacttttcatcactgggccgcagctgccgcagttgagaac420
aaggttccttgcgtcatgttgcttttgggtgctgccacagttatcgcaggtttcgctaga480
ggagtctctgaacacgcagctgccgcagttggaaaggagagaccagctgccgaagctcct540
tcatttgaaactgagagaagaaaattgatgactacacaaaatgcaagtggtatgactgtt600
gctgaaagatacttccttacattgatgagatcagatttggttgccatcagaagttgtgca660
gaatgggagccagaatctgttgcagctcttaccactttggctggtaaaccagttgtccct720
ttgggacttttgccaccttctcctgagggtggaagaggtgtttccaaggaagatgccgca780
gtcagatggttggacgcccaaccagcaaaatcagttgtctatgttgctcttggaagtgag840
gtccctttgagagccgagcaggttcatgaacttgctttgggtcttgaattgtctggagct900
agatttctttgggccttgagaaaaccaactgatgctcctgacgctgccgttttgccacct960
ggtttcgaagagagaaccagaggtagaggattggttgtcactggatgggttccacaaatt1020
ggtgtcttggctcatggagccgttgcagctttcttgacacactgcggttggaactccacc1080
atcgagggtcttttgttcggacacccacttattatgttgcctatctcttccgatcaaggt1140
ccaaatgctagattgatggaaggtagaaaagttggaatgcaggtccctagagatgagtct1200
gacggttcttttagaagagaagatgttgccgcaacagtcagagcagttgctgtcgaagag1260
gacggtagaagagtcttcaccgccaacgcaaagaaaatgcaggaaattgttgctgacgga1320
gcctgtcacgaaagatgtattgatggttttatccagcagttgagaagttacaaagca1377
<210>9
<211>458
<212>prt
<213>人工序列
<220>
<223>合成的
<400>9
metgluasnlysthrgluthrthrvalargargargargargileile
151015
leupheprovalpropheglnglyhisileasnproileleuglnleu
202530
alaasnvalleutyrserlysglypheserilethrilephehisthr
354045
asnpheasnlysprolysthrserasntyrprohisphethrphearg
505560
pheileleuaspasnaspproglnaspgluargileserasnleupro
65707580
thrhisglyproleualaglymetargileproileileasngluhis
859095
glyalaaspgluleuargarggluleugluleuleumetleualaser
100105110
glugluaspglugluvalsercysleuilethraspalaleutrptyr
115120125
phealaglnservalalaaspserleuasnleuargargleuvalleu
130135140
metthrserserleupheasnphehisalahisvalserleuprogln
145150155160
pheaspgluleuglytyrleuaspproaspasplysthrargleuglu
165170175
gluglnalaserglypheprometleulysvallysaspilelysser
180185190
alatyrserasntrpglnileleulysgluileleuglylysmetile
195200205
lysglnthrlysalaserserglyvaliletrpasnserphelysglu
210215220
leugluglusergluleugluthrvalilearggluileproalapro
225230235240
serpheleuileproleuprolyshisleuthralaserserserser
245250255
leuleuasphisaspargthrvalpheglntrpleuaspglnglnpro
260265270
proserservalleutyrvalserpheglyserthrsergluvalasp
275280285
glulysasppheleugluilealaargglyleuvalaspserlysgln
290295300
serpheleutrpvalvalargproglyphevallysglyserthrtrp
305310315320
valgluproleuproaspglypheleuglygluargglyargileval
325330335
lystrpvalproglnglngluvalleualahisglyalaileglyala
340345350
phetrpthrhisserglytrpasnserthrleugluservalcysglu
355360365
glyvalprometilepheserasppheglyleuaspglnproleuasn
370375380
alaargtyrmetseraspvalleulysvalglyvaltyrleugluasn
385390395400
glytrpgluargglygluilealaasnalaileargargvalmetval
405410415
aspglugluglyglutyrileargglnasnalaargvalleulysgln
420425430
lysalaaspvalserleumetlysglyglysersertyrgluserleu
435440445
gluserleuvalsertyrileserserleu
450455
<210>10
<211>1374
<212>dna
<213>人工序列
<220>
<223>合成的
<400>10
atggagaataagaccgagactaccgtcagaagaagaagaagaatcattttgttcccagtt60
cctttccaaggtcacatcaaccctatcttgcaacttgctaacgttttgtacagtaaggga120
ttctctatcactatcttccatacaaacttcaataagccaaagacctctaattaccctcac180
ttcacttttagattcattttggataacgacccacaggatgaaagaatctccaatttgcca240
actcatggacctcttgctggtatgagaatcccaattatcaacgaacacggtgccgacgag300
ttgagaagagaacttgagttgcttatgttggcctcagaagaggatgaagaggttagttgt360
ttgatcacagacgcactttggtactttgcacaatccgttgctgattcattgaaccttaga420
agattggtccttatgacctcttccttgttcaattttcatgctcacgtttcattgccacaa480
ttcgatgaattgggataccttgaccctgatgacaaaactagattggaagagcaggcttcc540
ggtttcccaatgttgaaggttaaggatatcaagtctgcctattccaactggcaaattttg600
aaggaaatccttggaaagatgattaaacagaccaaggcttcaagtggtgttatctggaac660
tcttttaaggaattggaggagtctgaacttgagactgtcattagagagatcccagcacct720
tctttcttgattccacttcctaaacatttgactgcttcttcctcaagtttgcttgatcac780
gacagaacagtttttcaatggttggatcaacagccaccttcttccgttttgtacgtctct840
ttcggttctacttctgaagttgatgagaaagactttttggaaattgctagaggtcttgtc900
gattcaaagcagagtttcttgtgggttgtcagaccaggatttgttaaaggttctacatgg960
gtcgaaccattgcctgacggtttccttggagagagaggtagaattgttaagtgggtccct1020
caacaggaagttttggcacatggagctatcggtgccttttggacacacagtggttggaac1080
tctaccttggaatccgtttgcgagggagtcccaatgattttctccgattttggtttggac1140
caacctcttaacgctagatatatgtctgatgttttgaaggttggagtctatcttgaaaac1200
ggatgggaaagaggagagattgctaatgccatcagaagagttatggtcgatgaagaggga1260
gagtatattagacaaaatgccagagttttgaagcagaaagcagacgtctctttgatgaaa1320
ggtggatcaagttacgaatctttggagtcccttgtttcatatatctcttccttg1374
<210>11
<211>805
<212>prt
<213>人工序列
<220>
<223>合成的
<400>11
metalathraspargleuthrargvalhisgluleuarggluargleu
151015
aspgluthrleuseralaasnargasngluileleualaleuleuser
202530
argilegluglylysglylysglyileleuglnhishisglnvalile
354045
alaglupheglugluileproglugluserargglnlysleuthrasp
505560
glyalapheglygluvalleuargserthrglnglualailevalleu
65707580
proprotrpvalalaleualavalargproargproglyvaltrpglu
859095
tyrleuargvalasnvalhisalaleuvalvalgluvalleuglnpro
100105110
alaglutyrleuargphelysglugluleuvalaspglyserserasn
115120125
glyasnphevalleugluleuaspphegluprophethralaserphe
130135140
proargprothrleuasnlysserileglyasnglyvalglnpheleu
145150155160
asnarghisleuseralalysleuphehisasplysgluserleuhis
165170175
proleuleuglupheleuargleuhisservallysglylysthrleu
180185190
metleuasnaspargileglnasnproaspalaleuglnhisvalleu
195200205
arglysalagluglutyrleuglythrvalproprogluthrprotyr
210215220
seralaphegluhislyspheglngluileglyleugluargglytrp
225230235240
glyaspasnalagluargvalleugluserileglnleuleuleuasp
245250255
leuleuglualaproaspprocysthrleugluthrpheleuglyarg
260265270
ileprometvalpheasnvalvalileleuserprohisglytyrphe
275280285
alaglnaspasnvalleuglytyrproaspthrglyglyglnvalval
290295300
tyrileleuaspglnvalargalaleugluasnglumetleuhisarg
305310315320
ilelysglnglnglyleuaspilevalproargileleuileilethr
325330335
argleuleuproaspalavalglythrthrcysglyglnargleuglu
340345350
lysvalpheglythrgluhisserhisileleuargvalprophearg
355360365
thrgluasnglyilevalarglystrpileserargphegluvaltrp
370375380
protyrleugluthrtyrthrgluaspvalalahisgluleualalys
385390395400
gluleuglnglylysproaspleuilevalglyasntyrseraspgly
405410415
asnilevalalaserleuleualahislysleuglyvalthrglncys
420425430
thrilealahisalaleuglulysthrlystyrprogluseraspile
435440445
tyrtrplyslysleuglugluargtyrhisphesercysglnphethr
450455460
alaaspleuphealametasnhisthrasppheileilethrserthr
465470475480
pheglngluilealaglyserlysaspthrvalglyglntyrgluser
485490495
histhralaphethrleuproglyleutyrargvalvalhisglyile
500505510
aspvalpheaspprolyspheasnilevalserproglyalaaspgln
515520525
thriletyrpheprohisthrgluthrserargargleuthrserphe
530535540
histhrgluileglugluleuleutyrserservalgluasngluglu
545550555560
hisilecysvalleulysaspargserlysproileilephethrmet
565570575
alaargleuaspargvallysasnilethrglyleuvalglutrptyr
580585590
glylysasnalalysleuarggluleuvalasnleuvalvalvalala
595600605
glyaspargarglysgluserlysaspleugluglulysalaglumet
610615620
lyslysmettyrserleuilegluthrtyrlysleuasnglyglnphe
625630635640
argtrpileserserglnmetasnargvalargasnglygluleutyr
645650655
argvalilealaaspthrlysglyalaphevalglnproalavaltyr
660665670
glualapheglyleuthrvalvalglualametthrcysglyleupro
675680685
thrphealathrcysasnglyglyproalagluileilevalhisgly
690695700
lysserglyphehisileaspprotyrhisglyaspargalaalaasp
705710715720
leuleuvalgluphepheglulysvallysvalaspproserhistrp
725730735
asplysileserglnalaglyleuglnargilegluglulystyrthr
740745750
trpglniletyrserglnargleuleuthrleuthrglyvaltyrgly
755760765
phetrplyshisvalserasnleuaspargarggluserargargtyr
770775780
leuglumetphetyralaleulystyrarglysleualagluserval
785790795800
proleualavalglu
805
<210>12
<211>2418
<212>dna
<213>人工序列
<220>
<223>合成的
<400>12
atggctactgacagattgactagagttcacgaattgagagaaagattggacgaaaccttg60
tctgctaatagaaacgagattttggccttgttgtctagaattgaaggaaagggtaagggt120
attttgcaacaccatcaggttattgcagagttcgaggaaatcccagaagagtctagacag180
aagttgactgatggagctttcggagaagtcttgagatccactcaagaggctattgtcttg240
ccaccatgggttgccttggcagtcagaccaagacctggtgtctgggaatacttgagagtt300
aatgtccatgctttggttgttgaggttttgcagcctgccgagtatttgagatttaaggaa360
gagttggttgatggatcttctaacggtaatttcgtcttggagttggatttcgagcctttt420
actgcctcttttcctagaccaacattgaataagtctatcggtaatggtgtccagtttttg480
aacagacatttgtctgccaaattgtttcatgataaggaatctttgcatccattgttggag540
ttcttgagattgcattctgttaagggaaaaactttgatgttgaacgatagaatccaaaac600
ccagacgcattgcagcatgttttgagaaaggctgaagagtacttgggaacagttccacca660
gaaacaccttactctgcattcgagcataagttccaggaaatcggattggagagaggttgg720
ggtgataacgctgagagagttttggaatctattcagttgttgttggacttgttggaggcc780
ccagacccatgtactttggagactttcttgggtagaatccctatggttttcaacgtcgtt840
atcttgtctccacatggttactttgctcaggataacgttttgggttaccctgacactgga900
ggtcaagtcgtttacattttggatcaagttagagccttggagaacgaaatgttgcacaga960
attaaacaacagggtttggatattgttccaagaatcttgattattactagattgttgcct1020
gacgccgttggaactacttgtggtcagagattggaaaaagtcttcggtacagaacactct1080
catattttgagagtcccatttagaactgaaaacggtattgttagaaagtggatctctaga1140
ttcgaggtttggccatacttggaaacttatacagaggatgttgctcatgaattggctaag1200
gagttgcagggaaagccagatttgatcgttggtaactactctgacggaaatatcgtcgct1260
tctttgttggcccacaaattgggtgttactcaatgtactattgctcacgcattggaaaag1320
acaaagtacccagaatctgatatttactggaaaaagttggaagagagataccacttctct1380
tgtcagtttacagctgatttgtttgctatgaaccatactgatttcattattacttctact1440
tttcaggaaatcgcaggttctaaggatactgttggtcaatacgaatctcacacagcattc1500
actttgccaggtttgtatagagttgttcacggaatcgatgtttttgatccaaagtttaac1560
attgtttctccaggagctgatcaaactatctatttcccacataccgagacctctagaaga1620
ttgacttctttccacacagagattgaggaattgttgtattcttctgttgaaaacgaggaa1680
cacatttgtgttttgaaagacagatccaagcctatcattttcactatggctagattggat1740
agagtcaagaacatcactggtttggtcgaatggtacggtaagaatgctaagttgagagaa1800
ttggttaacttggtcgttgttgccggtgatagaagaaaggaatctaaagacttggaggaa1860
aaggctgaaatgaagaagatgtactctttgattgaaacttacaaattgaacggtcaattc1920
agatggatctcttctcagatgaacagagtcagaaacggtgaattgtacagagttattgct1980
gatactaagggtgcatttgttcaaccagcagtctacgaagctttcggtttgactgttgtt2040
gaagctatgacttgtggtttgcctacatttgcaacttgtaatggtggaccagctgagatc2100
atcgttcatggaaagtctggttttcatattgatccttaccatggagatagagctgcagac2160
ttgttggttgagttcttcgagaaggttaaggttgacccatctcattgggataagatttct2220
caagctggattgcaaagaattgaagaaaaatacacttggcaaatttactctcaaagattg2280
ttgacattgactggagtttatggtttctggaagcatgtttctaatttggacagaagagaa2340
tctagaagatacttggaaatgttttacgctttgaaatatagaaaattggccgagtctgtt2400
ccattggctgttgagtaa2418
- 还没有人留言评论。精彩留言会获得点赞!