异戊烯化测定法的制作方法
本发明涉及在测定rab护送蛋白1(rep1)活性中使用的测定法。更具体地,本发明涉及rab6a在测定法中作为异戊烯化底物的用途,具体地其中已经使用基因治疗载体将rep1递送到细胞。
发明背景
无脉络膜(choroideremia)是一种罕见的疾病,其导致眼的脉络膜、视网膜色素上皮和光感受器的变性。患病的男性通常在青少年龄时期表现出夜盲症,在20和30岁期间逐渐丧失周边视力,在40岁完全失明。女性携带者有轻微的症状,最明显的是夜盲症,但偶尔可能有更严重的表型。
无脉络膜是由chm基因中的突变引起的,该chm基因编码rab护送蛋白(escortprotein)1(rep1)。rab护送蛋白2(rep2)(其与rep1是75%同源的)补偿身体的大多数细胞中的任何rep1缺陷。然而,rep2无法补偿眼中的rep1缺陷。这导致不足以维持靶rabgtp酶的正常异戊烯化的rab护送蛋白活性并引起细胞功能障碍并最终引起细胞死亡。
可以通过将rep1转基因的功能性拷贝提供给受影响的眼细胞成功地治疗无脉络膜(maclaren,r.e.etal.(2014)lancet383:1129-37)。具体地,已经显示腺伴随病毒(aav)基因治疗载体可以用于将编码功能性rep1的核苷酸序列递送至眼以治疗该疾病。
由于无脉络膜的基因治疗正在成为临床现实,因此需要可靠且灵敏的测定法来测定外源递送的rep1的活性,特别地以测试新的基因治疗载体和作为临床载体储液的质量控制筛选。
用于测定rep1的现有方法使用rab27a作为异戊烯化底物(tolmachova,t.etal.(2012)j.genemed.14:158-168;tolmachova,t.etal.(2013)j.mol.med.91:825-837;vasireddy,v.etal.(2013)plosone8:e61396;和black,a.etal.(2014)j.genemed.16:122-130)。这可能是从rab27a在无脉络膜发病机制中的众多影响(implication)得出。例如,已经显示rab27a在无脉膜细胞中以未异戊烯化存在,而其他rab是适当异戊烯化的(seabra,m.c.etal.(1995)j.biol.chem.270:24420-24427)。此外,rab27a在视网膜色素上皮和脉络膜毛细血管(无脉络膜中最早变性的两个部位)中以高水平表达。
然而,依赖于rab27a的异戊烯化的测定法产生非常弱的信号。结果,这些测定法的灵敏性是低的,并且它们可能不适合于临床基因治疗载体的可靠筛选。因此,非常需要更可靠且灵敏的测定法,其可以用于测定rep1活性和测试基因治疗载体。
发明概述
令人惊讶地,本发明人发现了尽管普遍认为rab27a提供了最合适的异戊烯化测定底物,但是在异戊烯化反应中使用rab6a作为底物提供了一种更灵敏的用于测定rab护送活性蛋白1(rep1)的方法。rab6a不仅在检测异戊烯化的测定法中提供增加的灵敏性,它还提供有益的信噪比、更好的动态信号范围和更好的一致性。
此外,本发明人已经证明了,可以利用基于rab6a的测定法的增加的灵敏性来准确且可靠测定编码rep1的载体的活性,特别是aav基因治疗载体,例如那些适用于临床的载体。
因此,一方面,本发明提供了测定rab护送蛋白1(rep1)活性的方法,包括以下步骤:
(a)提供包含rep1的样品;
(b)使步骤(a)的样品与rab6a、rab牻牛儿基牻牛儿基转移酶(rabgeranylgeranyltransferase)(rabggt酶)和脂质供体底物接触;并且
(c)检测脂质化rab6a产物。
例如,本发明的方法可以用于测试适合于将rep1递送至靶细胞的基因治疗载体或用于载体储液(例如药物储液)的质量控制分析。
基因治疗载体的验证对于在临床中基因治疗的安全且有效实施是强制性的。对于分析和质量控制步骤,与对照实验或参考水平的比较可以提供基因治疗载体或rep1相对于已知或公认标准的活性的量度(例如,比已知或公认的标准更好或更差)。这可以用于验证基因治疗载体储液是否符合特定目标和法规。
在一个实施方案中,比较rep1或编码rep1的aav载体的样品,其定义为主要参考标准品。可以例如在测试样品和主要参考标准样品上平行进行本发明的方法。可以例如相对于主要参考标准品定义测试样品的效力和/或行为。
换言之,本发明的方法可以例如用于质量控制分析和验证基因治疗载体为有效的(例如用于治疗无脉络膜),优选地包含编码rep1的核苷酸序列的aav载体颗粒,优选地,其中通过本发明的方法测定的载体的输出活性或功效高于阈值活性或在规定的靶标范围内(例如通过与对照实验或参照水平比较)指示载体适合于基因治疗目的。
因此,另一方面,本发明的方法用于rab护送蛋白1(rep1)编码基因治疗载体(优选aav载体)的质量控制分析。
另一方面,本发明提供了用于rab护送蛋白1(rep1)编码基因治疗载体(优选aav载体)的质量控制分析的方法,包括以下步骤:
(a)用载体转导细胞,在适合于表达rep1的条件下培养细胞并裂解细胞以提供含有rep1的样品;
(b)使步骤(a)的样品与rab6a、rab牻牛儿基牻牛儿基转移酶(rabggt酶)和脂质供体底物接触;并且
(c)检测脂质化rab6a产物。
因此,该方法可以包括进行包括步骤(a)至(c)的多个实验,其中涉及包含rep1的样品的参数是变化的,而其他参数(例如涉及rab6a、rabggt酶和脂质的参数)保持恒定。此类参数可以包括例如相关蛋白质(例如rep1)的氨基酸序列、用于在细胞中表达rep1的载体中包含的编码rep1的核苷酸序列、用于将编码rep1的核苷酸序列递送到细胞的载体类型(例如病毒载体的类型,例如腺伴随病毒(aav)载体的类型)、rep1的浓度和/或用于将编码rep1的核苷酸序列递送到细胞的载体的感染复数(moi)。在一个优选的实施方案中,该方法包括进行多个实验,包括在用于将编码rep1的核苷酸序列递送到细胞的载体的不同moi下的步骤(a)至(c)(例如,以产生剂量-响应曲线)。
在一个实施方案中,脂质化rab6a产物的检测包括量化脂质化rab6a产物的量。在优选的实施方案中,脂质化rab6a产物的检测包括相对于对照或参照水平量化脂质化rab6a产物的量。例如,可以相对于定义为主要参考标准品的rep1或编码rep1的aav载体的样品进行量化。可以例如在测试样品和主要参考标准样品上平行进行本发明的方法。可以例如相对于主要参考标准品定义测试样品的效力和/或行为。
在一个实施方案中,该方法包括比较脂质化rab6a产物(例如,异戊烯化的,例如牻牛儿基牻牛儿基化或生物素牻牛儿基化,rab6a)的量与自对照实验,例如使用rep1的已知或标准样品的实验确定的量的进一步步骤。
在另一个实施方案中,该方法还包括比较脂质化rab6a产物(例如,异戊烯化的,例如牻牛儿基牻牛儿基化或生物素牻牛儿基化,rab6a)的量与参考水平的进一步步骤。
在一个实施方案中,包含rep1的样品来自经遗传工程化改造以表达rep1的细胞。优选地,包含rep1的样品是经遗传工程化改造以表达rep1的细胞的裂解物。优选地,用包含编码rep1的核苷酸序列的载体转染或转导细胞,以提供经遗传工程化改造以表达rep1的细胞。优选地,载体是病毒载体。
在一个实施方案中,使用包含编码rep1的核苷酸序列的病毒载体表达rep1。
在一个实施方案中,病毒载体是腺伴随病毒(aav)载体。优选地,病毒载体是病毒载体颗粒的形式。
aav载体可以是任何血清型的(例如包含任何aav血清型基因组和/或壳体蛋白)。优选地,载体能够感染或转导眼细胞。
在一个实施方案中,aav载体包含aav血清型1、2、3、4、5、6、7、8、9、10或11基因组。在另一个实施方案中,aav载体包含aav血清型2、4、5或8基因组。优选地,aav载体包含aav血清型2基因组。
在一个实施方案中,aav载体颗粒包含aav血清型1、2、3、4、5、6、7、8、9、10或11壳体蛋白。在另一个实施方案中,aav载体颗粒包含aav血清型2、4、5或8壳体蛋白。例如,aav血清型8壳体蛋白可以是aav8/y733f突变体壳体蛋白。优选地,aav载体颗粒包含aav血清型2壳体蛋白。
在一个实施方案中,aav载体颗粒包含aav2基因组和aav2壳体蛋白(aav2/2);aav2基因组和aav5壳体蛋白(aav2/5);或aav2基因组和aav8壳体蛋白(aav2/8)。优选地,aav载体颗粒包含aav2基因组和aav2壳体蛋白(aav2/2)。
aav载体颗粒可以是一种或多种天然存在的aav的嵌合、改组或壳体修饰的衍生物。特别地,aav载体颗粒可以包含来自相同载体(即假型化载体)内的aav的不同血清型、进化枝、克隆或隔离群(isolates)的壳体蛋白序列。因此,在一个实施方案中,aav载体是假型化aav载体颗粒的形式。
在一个实施方案中,rep1是人rep1。
在一个实施方案中,rep1包含与seqidno:5具有至少70%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的氨基酸序列,优选其中氨基酸序列基本上保留以seqidno:5所示的蛋白质的天然功能。
在一个实施方案中,编码rep1的核苷酸序列包含与seqidno:6或7具有至少70%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的核苷酸序列,优选其中由核苷酸序列编码的蛋白质基本上保留以seqidno:5所示的蛋白质的天然功能。
在一个实施方案中,编码rep1的核苷酸序列包含核苷酸序列,所述核苷酸序列编码与seqidno:5具有至少70%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的氨基酸序列,优选其中氨基酸序列基本上保留以seqidno:5所示的蛋白质的天然功能。
在一个实施方案中,rab6a和/或rabggt酶是基本上纯的。
在一个实施方案中,rab6a包含与seqidno:1具有至少70%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的氨基酸序列,优选其中氨基酸序列基本上保留以seqidno:1所示的蛋白质的天然功能。
在一个实施方案中,rabggt酶包含与seqidno:3或8,优选seqidno:8具有至少70%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的氨基酸序列,优选其中氨基酸序列基本上保留以seqidno:8所示的蛋白质的天然功能;和/或与seqidno:4或9,优选seqidno:9具有至少70%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的氨基酸序列,优选其中氨基酸序列基本上保留以seqidno:9所示的蛋白质的天然功能。
在另一个实施方案中,rab6a:rabggt酶摩尔比率为约1:0.25-3、1:0.3-2.9、1:0.35-2.8、1:0.4-2.7、1:0.45-2.6或1:0.5-2.5,优选约1:0.5-2.5。
在一个实施方案中,rab6a:rabggt酶摩尔比率为约1:2-3、1:2.1-2.9、1:2.2-2.8、1:2.3-2.7或1:2.4-2.6,优选约1:2.4-2.6。在一个实施方案中,rab6a:rabggt酶摩尔比率为约1:2、1:2.1、1:2.2、1:2.3、1:2.4、1:2.5、1:2.6、1:2.7、1:2.8、1:2.9或1:3,优选约1:2.5。
在另一个实施方案中,rab6a:rabggt酶摩尔比率为约1:0.25-0.75、1:0.3-0.7、1:0.35-0.65、1:0.4-0.6或1:0.45-0.55,优选约1:0.4-0.6。在一个实施方案中,rab6a:rabggt酶摩尔比率为约1:0.25、1:0.3、1:0.35、1:0.4、1:0.45、1:0.5、1:0.55、1:0.6、1:0.65、1-0.7或1:0.75,优选约1:0.5。
在一个实施方案中,脂质供体底物是牻牛儿基牻牛儿基焦磷酸盐/酯(geranylgeranylpyrophosphate)(ggpp)或其类似物。优选地,用可检测标志物标记脂质供体底物。例如,脂质供体底物可以是同位素标记的(例如,脂质供体底物可以包含3h),或者可以包含荧光基团、表位或生物素部分。
在优选的实施方案中,脂质供体底物是生物素-牻牛儿基牻牛儿基焦磷酸盐/酯(bgpp)。
在一个实施方案中,使用酶联免疫吸附测定法(elisa)、western印迹分析或放射自显影术检测脂质化rab6a产物。在优选的实施方案中,使用elisa检测脂质化rab6a产物。elisa可以是例如夹心式elisa。
在一个优选的实施方案中,使用对生物素特异的检测试剂,例如链霉抗生物素蛋白检测生物素标记的脂质化rab6a产物。优选地,使用对生物素特异的检测试剂,例如链霉抗生物素蛋白(例如链霉抗生物素蛋白-辣根过氧化物酶缀合物),使用western印迹分析检测生物素标记的脂质化rab6a产物。更优选地,使用对生物素特异的检测试剂(例如链霉抗生物素蛋白),使用elisa检测生物素标记的脂质化rab6a产物。
在一个实施方案中,该方法用于测定用于治疗无脉膜的编码rep1的基因治疗载体的活性。
另一方面,本发明提供rab6a用于测定rab护送蛋白1(rep1)活性的用途。
测定rep1、rab6a、rabggt酶、脂质供体底物和rep1的活性的方法可以如本文所述。
另一方面,本发明提供了用于测定包含rab护送蛋白1(rep1)编码核苷酸序列的载体功效的方法,其中该方法包括以下步骤:
(a)提供包含rep1的样品,其中使用包含编码rep1的核苷酸序列的载体表达rep1;
(b)使步骤(a)的样品与rab6a、rab牻牛儿基牻牛儿基转移酶(rabggt酶)和脂质供体底物接触;并且
(c)检测脂质化rab6a产物。
另一方面,本发明提供rab6a用于测定包含rab护送蛋白1(rep1)编码核苷酸序列的载体功效的用途。
优选地,该方法和用途用于测定用于治疗无脉膜的载体的功效。
载体、rep1、rab6a、rabggt酶、脂质供体底物、脂质化rab6a产物、检测方法以及该方法和用途的其他特征可以如本文所述。
附图简述
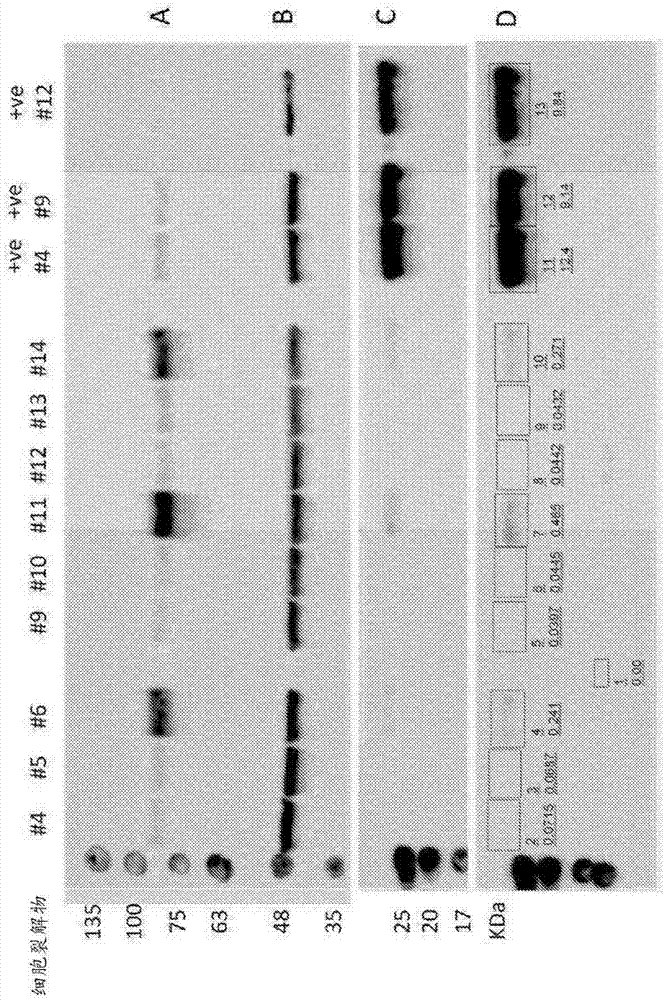
图1
使用rab27a作为底物进行体外异戊烯化反应,然后使用未转导的细胞和用aav-gfp或aav-rep1转导的细胞(moi=10000gp/细胞)对掺入的生物素化的脂质供体进行western印迹(wb)分析。
实验涉及3组独立制备的裂解物。使用总体积为12.5μl的10μg裂解物建立异戊烯化反应。对阳性对照掺加2μm鱼rep1。检测时间为2min。
(a)细胞裂解物(1:1000)中人rep1水平的wb分析。未转导细胞(#4,#9和#12)显示内源性rep1水平。用aav-gfp转导的细胞(#5,#10和#13)显示与未转导细胞相似的内源性rep1水平。与未转导的细胞和经aav-gfp转导的细胞相比,用aav-rep1转导的细胞(#6,#11和#14)显示rep1水平增加。阳性对照(+ve)显示内源性rep1水平。
(b)β-肌动蛋白作为上样对照(1:15000)的wb分析。在所有分析的样品中β-肌动蛋白的水平相似。
(c)在rab27a(1:10000)中掺入的生物素化的脂质供体的wb分析。未转导的细胞和经aav-gfp转导的细胞在rab27a中未显示可检测的生物素掺入。用aav-rep1转导的细胞显示一定水平的掺入rab27a中的生物素(#6,#11和#14)。由于鱼rep1活性,阳性对照显示所有中最强的条带。
(d)使用imagestudiolite软件半定量(c)中的wb分析。
图2
使用rab27a作为底物进行体外异戊烯化反应,然后使用未转导的细胞和用aav-gfp或aav-rep1转导的细胞(moi=10000gp/细胞)对掺入的生物素化的脂质供体进行western印迹(wb)分析。
实验涉及2组独立制备的裂解物。使用总体积为22μl的30μg裂解物建立异戊烯化反应。对阳性对照掺加1μm鱼rep1。检测时间为2分钟。
(a)细胞裂解物(1:1000)中人rep1水平的wb分析。未转导细胞(#9和#12)显示内源性rep1水平。用aav-gfp转导的细胞(#10和#13)显示与未转导细胞相似的内源性rep1水平。与未转导的细胞和经aav-gfp转导的细胞相比,用aav-rep1转导的细胞(#11和#14)显示rep1水平增加。阳性对照(+ve)显示内源性rep1水平。
(b)β-肌动蛋白作为上样对照(1:15000)的wb分析。在所有分析的样品中β-肌动蛋白的水平相似。
(c)在rab27a(1:10000)中掺入的生物素化的脂质供体的wb分析。未转导细胞和经aav-gfp转导的细胞在rab27a中未显示可检测的生物素掺入。用aav-rep1转导的细胞显示一定水平的掺入rab27a中的生物素(#11和#14)。由于鱼rep1活性,阳性对照显示所有中最强的条带。
(d)使用imagestudiolite软件半定量(c)中的wb分析。
图3
使用rab6a作为底物的体外异戊烯化反应,然后使用未转导的细胞和用aav-gfp或aav-rep1转导的细胞(moi=10000gp/细胞)对掺入的生物素化的脂质供体进行western印迹(wb)分析。
实验涉及2组独立制备的裂解物。使用总体积为20μl的20μg裂解物建立异戊烯化反应。对阳性对照掺加1μm鱼rep1。检测时间为2分钟。
(a)细胞裂解物(1:1000)中人rep1水平的wb分析。未转导细胞(#9和#12)显示内源性rep1水平。用aav-gfp转导的细胞(#10和#13)显示与未转导细胞相似的内源性rep1水平。与未转导的细胞和经aav-gfp转导的细胞相比,用aav-rep1转导的细胞(#11和#14)显示rep1水平增加。阳性对照(+ve)显示内源性rep1水平。
(b)β-肌动蛋白作为上样对照(1:15000)的wb分析。在所有分析的样品中β-肌动蛋白的水平相似。
(c)在rab6a(1:10000)中掺入的生物素化的脂质供体的wb分析。未转导的细胞和经aav-gfp转导的细胞在rab6a中显示非常低的生物素掺入。用aav-rep1转导的细胞显示掺入rab6a中的生物素(#11和#14)。由于鱼rep1活性,阳性对照显示最强的条带。
(d)使用imagestudiolite软件半定量(c)中的wb分析。
图4
使用rab6a作为底物的体外异戊烯化反应,然后使用未转导的细胞和用aav-rep1转导的细胞(moi=250、1000、5000、10000和20000gp/细胞)对掺入的生物素化的脂质供体进行western印迹(wb)分析。
使用总体积为15μl的20μg裂解物建立异戊烯化反应。对阳性对照掺加0.5μm鱼rep1。检测时间为2分钟。
(a)细胞裂解物(1:2500)中人rep1水平的wb分析。未转导的细胞显示内源性rep1水平。用aav-rep1转导的细胞显示rep1水平的增加,其与所用的moi直接相关。阳性对照(+ve)显示内源性rep1水平。
(b)β-肌动蛋白作为上样对照(1:50000)的wb分析。在所有分析的样品中β-肌动蛋白的水平相似。
(c)在rab6a(1:10000)中掺入的生物素化的脂质供体的wb分析。未转导的细胞在rab6a中显示非常低的生物素掺入。用aav-rep1转导的细胞显示越来越多的掺入rab6a中的生物素。由于鱼rep1活性,阳性对照显示所有中最强的条带。
(d)使用imagestudiolite软件半定量(c)中的wb分析。
图5
使用rab6a作为底物的体外异戊烯化反应,然后使用未转导的细胞和用aav-rep1转导的细胞(moi=10000gp/细胞)对掺入的生物素化的脂质供体进行western印迹(wb)分析。
使用总体积为15μl的20μg裂解物建立异戊烯化反应。对阳性对照掺加0.5μm鱼rep1。rep1/肌动蛋白的检测时间为2分钟,而生物素的检测时间为30秒。
(a)细胞裂解物(1:2500)中人rep1水平的wb分析。未转导的细胞显示内源性rep1水平。用aav-rep1转导的细胞显示rep1水平的增加。阳性对照(+ve)显示内源性rep1水平。
(b)β-肌动蛋白作为上样对照(1:50000)的wb分析。在所有分析的样品中β-肌动蛋白的水平相似。
(c)在rab6a(1:10000)中掺入的生物素化的脂质供体的wb分析。未转导的细胞在rab6a中显示非常低的生物素掺入。用aav-rep1转导的细胞显示增加的对rab6a的生物素掺入。由于鱼rep1活性,阳性对照显示所有中最强的条带。
(d)使用imagestudiolite软件半定量(c)中的wb分析。
图6
体外异戊烯化反应,然后在未转导的arpe-19细胞中掺入生物素化的脂质供体的western印迹(wb)分析,和用aav-rep1转导的arpe-19细胞。
使用总体积为45μl的15μg裂解物建立异戊烯化反应。对阳性对照掺加0.1μm的鱼rep1。rep1/肌动蛋白的检测时间:2分钟;生物素的检测时间:30秒。
使用以下细胞平行进行实验:
(a)未转导的细胞(#86和#87);和
(b)细胞+aav-rep1moi10,000(#90和#91)-r&d级载体。
(a)细胞裂解物(1:2,500)中人rep1水平的wb分析。未转导的细胞显示内源性rep1水平。用aav-rep1转导的细胞显示rep1水平的增加。阳性对照(+ve)显示内源性rep1水平。
(b)β-肌动蛋白作为上样对照(1:50,000)的wb分析。在所有分析的样品中β-肌动蛋白的水平相似。
(c)在rab6a(1:10,000)中掺入的生物素化的脂质供体的wb分析。未转导的细胞在rab6a中显示非常低的生物素掺入;用aav-rep1转导的细胞显示增加的对rab6a的生物素掺入。由于鱼rep1活性,阳性对照显示所有中最强的条带。
(d)使用imagestudiolite软件半定量(c)中的wb分析。
图7
体外异戊烯化反应,然后在未转导的ht1080细胞和用aav-rep1转导的ht1080细胞中掺入生物素化的脂质供体的western印迹(wb)分析。
使用总体积为20μl的20μg裂解物建立异戊烯化反应。对阳性对照掺加0.1μm的鱼rep1。rep1/肌动蛋白的检测时间:2分钟;生物素的检测时间:30秒。
使用以下细胞平行进行实验:
(a)未转导的细胞(#56和#57);
(b)细胞+aav-rep1moi10,000(#60和#61)-r&d级载体;和
(c)细胞+aav-rep1moi10,000(#64和#65)-临床级载体。
(a)细胞裂解物(1:2,500)中人rep1水平的wb分析。未转导的细胞显示内源性rep1水平。用aav-rep1转导的细胞显示rep1水平的增加。阳性对照(+ve)显示内源性rep1水平。
(b)β-肌动蛋白作为上样对照(1:50,000)的wb分析。在所有分析的样品中β-肌动蛋白的水平相似。
(c)在rab6a(1:10,000)中掺入的生物素化的脂质供体的wb分析。未转导的细胞显示rab6a中生物素掺入的基线水平;用aav-rep1转导的细胞显示增加的对rab6a的生物素掺入。
(d)使用imagestudiolite软件半定量(c)中的wb分析。
图8
体外异戊烯化反应,然后进行未转导的细胞和用aav-rep1转导的细胞(moi=250、1000、5000、10000和20000gp/细胞)中掺入的生物素化脂质供体的western印迹(wb)分析,比较rab6a和rab27a底物。
使用总体积为15μl的20μg裂解物和2种不同底物建立异戊烯化反应:rab27a(左手泳道和图;红色)和rab6a(右手泳道和图;蓝色)。对阳性对照(每种底物一种)掺加0.1μm鱼rep1。检测时间:2分钟。
(a)细胞裂解物(1:2,500)中人rep1水平的wb分析。未转导的细胞显示内源性rep1水平。用aav-rep1转导的细胞显示rep1水平的增加,其与所用的moi直接相关。阳性对照(+ve)显示内源性rep1水平。
(b)β-肌动蛋白作为上样对照(1:50,000)的wb分析。在所有分析的样品中β-肌动蛋白的水平相似。
(c)在rab27a和rab6a中掺入的生物素化脂质供体的wb分析。未转导的细胞显示非常低的生物素对rab蛋白的掺入,其随着moi的增加而增加。由于鱼rep1活性,阳性对照显示强烈的生物素掺入。
(d)使用imagestudiolite软件半定量(c)中的wb分析。使用prism软件绘制数据,如(e)中的四个图中所示。
(e)在使用的moi间的来自生物素化底物的条带密度值(上部两个图)和使用的moi间生物素化底物与rep1之间的比率(下部两个图)。
图9
体外异戊烯化反应,然后进行未转导的细胞中掺入的生物素化的脂质供体的western印迹(wb)分析,比较不同的异戊烯化反应条件。
使用总体积为15μl的20μg裂解物和2种不同的底物建立异戊烯化反应:rab27a(红色)和rab6a(蓝色)。对阳性对照(每种底物一种)掺加重组人rep1。检测时间:2分钟。
(a)在rab27a和rab6a中掺入的生物素化脂质供体的wb分析。生物素掺入水平与反应中总蛋白质的量成正比。由于鱼rep1活性,阳性对照显示强烈的生物素掺入。
(b)β-肌动蛋白作为上样对照(1:50,000)的wb分析。β-肌动蛋白的水平与反应中使用的总细胞裂解物的量相匹配,并且在样品之间是相似的。
(c)细胞裂解物(1:2,500)中人rep1水平的wb分析。未转导的细胞显示内源性rep1水平。阳性对照(+ve)显示更高的rep1密度。
(d)使用imagestudiolite软件半定量(a)中的wb分析。使用prism软件绘制数据。突出显示的值是针对检测到底物之间的较高差异的那些条件。
图10
作为经aav-rep1转导的细胞中异戊烯化底物的rab27a和rab6a之间的比较。
发明详述
现在将通过非限制性实例描述本发明的各种优选特征和实施方案。
除非另有指示,本发明的实践将采用化学、生物化学、分子生物学、微生物学和免疫学的常规技术,它们在本领域普通技术人员的能力范围内。此类技术在文献中有解释。参见,例如sambrook,j.,fritsch,e.f.andmaniatis,t.(1989)molecularcloning:alaboratorymanual,第2版,coldspringharborlaboratorypress;ausubel,f.m.etal.(1995及定期增补)currentprotocolsinmolecularbiology,第9、13和16章,johnwiley&sons;roe,b.,crabtree,j.andkahn,a.(1996)dnaisolationandsequencing:essentialtechniques,johnwiley&sons;polak,j.m.andmcgee,j.o’d.(1990)insituhybridization:principlesandpractice,oxforduniversitypress;gait,m.j.(1984)oligonucleotidesynthesis:apracticalapproach,irlpress;及lilley,d.m.anddahlberg,j.e.(1992)methodsinenzymology:dnastructuresparta:synthesisandphysicalanalysisofdna,academicpress。这些通用教科书中的每个通过引用并入本文。
异戊烯化
通过添加类异戊二烯部分使蛋白质脂质化是翻译后修饰,其影响高达2%的哺乳动物蛋白质组。此类脂质化能够使靶蛋白与细胞膜可逆结合,并且还可以调节蛋白质-蛋白质相互作用。
优选地,本文提及的脂质化是异戊烯化,使得脂质供体底物和脂质化rab6a产物分别是异戊烯基供体底物和异戊烯化的rab6a产物。
异戊烯化是一种特殊类型的翻译后修饰,其中牻牛儿基牻牛儿基或法呢基部分(或任一者的类似物)通过硫醚连接与蛋白质的一个或两个c端半胱氨酸残基连接。
优选地,异戊烯化是将牻牛儿基牻牛儿基部分或其类似物(例如生物素-牻牛儿基部分)添加至靶蛋白(例如rab6a)。
附着于蛋白质的牻牛儿基牻牛儿基部分(蛋白质用阴影圆圈示意性地描绘)是:
附着于蛋白质的法呢基部分(蛋白质用阴影圆圈示意性地描绘)是:
术语“类似物”在本文中就脂质(例如牻牛儿基牻牛儿基或法呢基)部分或脂质供体底物而言使用,指经过修饰以包含适合于特定目的,例如检测的官能团的化合物。类似物能够以基本上不受修饰阻碍(为了本发明的活性测定法的目的)的方式通过异戊烯化机器(即rep1和rabggt酶)被添加到底物蛋白质。
因此,上述部分的类似物包括为特定目的人工创建的那些(例如适于在测定法中检测的标记部分)。特别地,nguyen等人(nguyen,u.t.etal.(2009)nat.chem.biol.5:227-235)开发了以下生物素-牻牛儿基部分,其可以在体外蛋白质异戊烯化反应中检测到(生物素-牻牛儿基部分显示附着于蛋白质,其由阴影圆圈示意性地描绘):
rab6a
rab6a(ras相关蛋白rab-6a)是哺乳动物rabgtp酶家族的成员,其本身是gtp酶的ras样超家族中最大的。
rabgtp酶(也称为rab蛋白)是外周膜蛋白,并且参与膜运输的调节,包括囊泡形成、沿肌动蛋白和微管蛋白网络的囊泡运动和膜融合。rab6a的主要功能理解为调节从高尔基复合体到内质网的蛋白质转运。
rabgtp酶通常通过异戊二烯基团(特别是,牻牛儿基牻牛儿基)锚定到细胞膜,所述异戊二烯基团与两个c端半胱氨酸残基共价结合。
rabgtp酶表现出两种构象:无活性,gdp结合形式;和有活性,gtp结合形式。从gdp到gtp结合形式的转化由gdp/gtp交换因子(gef)催化,从而激活rabgtp酶。相反,rabgtp酶对gtp的水解可以通过gtp酶活化蛋白(gap)增强,从而导致rab失活。
在一个实施方案中,rab6a是人rab6a。
rab6a的示例性氨基酸序列是以ncbi登录号np_942599.1保藏的序列。
rab6a的示例性氨基酸序列是:
mstggdfgnplrkfklvflgeqsvgktslitrfmydsfdntyqatigidflsktmyledrtvrlqlwdtagqerfrslipsyirdstvavvvyditnvnsfqqttkwiddvrtergsdviimlvgnktdladkrqvsieegerkakelnvmfietsakagynvkqlfrrvaaalpgmestqdrsredmidiklekpqeqpvseggcsc
(seqidno:1)
编码rab6a的示例性核苷酸序列是在ncbi登录号nm_198896.1下描述的序列。
编码rab6a的示例性核苷酸序列是:
atgtccacgggcggagacttcgggaatccgctgaggaaattcaagctggtgttcctgggggagcaaagcgttggaaagacatctttgatcaccagattcatgtatgacagttttgacaacacctatcaggcaacaattggcattgactttttatcaaaaactatgtacttggaggatcgaacagtacgattgcaattatgggacacagcaggtcaagagcggttcaggagcttgattcctagctacattcgtgactccactgtggcagttgttgtttatgatatcacaaatgttaactcattccagcaaactacaaagtggattgatgatgtcagaacagaaagaggaagtgatgttatcatcatgctagtaggaaataaaacagatcttgctgacaagaggcaagtgtcaattgaggagggagagaggaaagccaaagagctgaatgttatgtttattgaaactagtgcaaaagctggatacaatgtaaagcagctctttcgacgtgtagcagcagctttgccgggaatggaaagcacacaggacagaagcagagaagatatgattgacataaaactggaaaagcctcaggagcaaccagtcagtgaaggaggctgttcctgctaa
(seqidno:2)
rab牻牛儿基牻牛儿基转移酶(rabggt酶)
rab牻牛儿基牻牛儿基转移酶(rabggt酶;也称为牻牛儿基牻牛儿基转移酶ii)是蛋白质异戊二烯基转移酶,其仅使rab家族的gtpa酶异戊烯化。
rabggt酶通常天然催化将两个牻牛儿基牻牛儿基基团转移到rabgtp酶的c端的半胱氨酸残基。每个牻牛儿基牻牛儿基基团经由与半胱氨酸残基的硫醚连接与rabgtp酶缀合。
已经显示了rabggt酶能够结合一大批衍生化的磷酸类异戊二烯,并且可以催化其添加到rabgtp酶底物(例如rab6a)。例如,nguyen等人(nguyen,u.t.etal.(2009)nat.chem.biol.5:227-235)证明了生物素-牻牛儿基部分对rabgtp酶的成功添加。
rabggt酶是一种由alpha和beta亚基构成的异二聚体酶。
在一个实施方案中,rabggt酶是人rabggt酶。在一个优选的实施方案中,rabggt酶是大鼠rabggt酶。
rabggt酶alpha亚基的示例性氨基酸序列是在ncbi登录号np_004572.3和np_113842.1下描述的序列。
rabggt酶alpha亚基的示例性氨基酸序列是:
mhgrlkvktseeqaeakrlereqklklyqsatqavfqkrqageldesvleltsqilganpdfatlwncrrevlqqletqkspeelaalvkaelgflesclrvnpksygtwhhrcwllgrlpepnwtrelelcarflevdernfhcwdyrrfvatqaavppaeelaftdslitrnfsnysswhyrscllpqlhpqpdsgpqgrlpedvllkelelvqnafftdpndqsawfyhrwllgradpqdalrclhvsrdeacltvsfsrpllvgsrmeilllmvddsplivewrtpdgrnrpshvwlcdlpaaslndqlpqhtfrviwtagdvqkecvllkgrqegwcrdsttdeqlfrcelsvekstvlqselesckelqelepenkwclltiillmraldpllyeketlqyfqtlkavdpmratylddlrskfllensvlkmeyaevrvlhlahkdltvlchleqlllvthldlshnrlrtlppalaalrclevlqasdnaiesldgvtnlprlqelllcnnrlqqpavlqplascprlvllnlqgnplcqavgileqlaellpsvssvlt
(seqidno:3)
和:
mhgrlkvktseeqaeakrlereqklklyqsatqavfqkrqageldesvleltsqilganpdfatlwncrrevlqhletekspeesaalvkaelgflesclrvnpksygtwhhrcwllsrlpepnwarelelcarfleadernfhcwdyrrfvaaqaavapaeelaftdslitrnfsnysswhyrscllpqlhpqpdsgpqgrlpenvllkelelvqnafftdpndqsawfyhrwllgraephdvlccvhvsreeaclsvcfsrpltvgsrmgtlllmvdeaplsvewrtpdgrnrpshvwlcdlpaaslndqlpqhtfrviwtgsdsqkecvllkdrpecwcrdsatdeqlfrcelsvekstvlqselesckelqelepenkwclltiillmraldpllyeketlqyfstlkavdpmraaylddlrskfllensvlkmeyadvrvlhlahkdltvlchleqlllvthldlshnrlralppalaalrclevlqasdnalenvdgvanlprlqelllcnnrlqqsaaiqplvscprlvllnlqgnslcqeegiqerlaemlpsvssilt
(seqidno:8)
rabggt酶beta亚基的示例性氨基酸序列是ncbi登录号np_004573.2.和np_619715.1下描述的序列。
rabggt酶beta亚基的示例性氨基酸序列是:
mgtpqkdviiksdapdtlllekhadyiasygskkddyeycmseylrmsgiywgltvmdlmgqlhrmnreeilafikscqhecggisasighdphllytlsavqiltlydsinvidvnkvveyvkglqkedgsfagdiwgeidtrfsfcavatlallgkldainvekaiefvlscmnfdggfgcrpgseshagqiycctgflaitsqlhqvnsdllgwwlcerqlpsgglngrpeklpdvcyswwvlaslkiigrlhwidreklrnfilacqdeetggfadrpgdmvdpfhtlfgiaglsllgeeqikpvnpvfcmpeevlqrvnvqpelvs
(seqidno:4)
和:
mgtqqkdvtiksdapdtlllekhadyiasygskkddyeycmseylrmsgvywgltvmdlmgqlhrmnkeeilvfikscqhecggvsasighdphllytlsavqiltlydsihvinvdkvvayvqslqkedgsfagdiwgeidtrfsfcavatlallgkldainvekaiefvlscmnfdggfgcrpgseshagqiycctgflaitsqlhqvnsdllgwwlcerqlpsgglngrpeklpdvcyswwvlaslkiigrlhwidreklrsfilacqdeetggfadrpgdmvdpfhtlfgiaglsllgeeqikpvspvfcmpeevlqrvnvqpelvs
(seqidno:9)
脂质供体底物
为了将脂质部分添加到rabgtp酶,rabggt酶可以以脂质(例如牻牛儿基牻牛儿基或生物素-牻牛儿基)供体底物形式使用脂质部分作为底物。这些通常是脂质部分的焦磷酸盐衍生物。
例如,牻牛儿基牻牛儿基焦磷酸盐/酯(ggpp)或生物素-牻牛儿基焦磷酸盐/酯(bgpp)可以被rabggt酶用作脂质供体底物以分别将牻牛儿基牻牛儿基或生物素-牻牛儿基部分转移到底物rabgtp酶。
牻牛儿基牻牛儿基焦磷酸盐具有结构:
生物素-牻牛儿基焦磷酸盐的示例性结构是:
rab护送蛋白1(rep1)
rab护送蛋白(rep)进行将未异戊烯化的rabgtp酶呈现到rabggt酶,并且将异戊烯化的rabgtp酶携带到其靶膜的功能。
rabgtp酶不包含可以由rabggt酶识别的异戊烯化位点处的共有序列。然而,底物识别经由rep实现,所述rep经由保守区结合rabgtp酶,然后给rabggt酶呈现其底物以进行异戊烯化。
一旦异戊烯化,脂质锚定使rabgtp酶变为不溶性的。因而,需要rep以结合并且溶解牻牛儿基牻牛儿基基团并且帮助将rabgtp酶递送到靶细胞膜。
rep1也可以称为rab蛋白牻牛儿基牻牛儿基转移酶组分a。此外,编码rep1的基因可以称为chm基因。
在一个实施方案中,rep1是人rep1。
rep1的示例性氨基酸序列是:
madtlpsefdvivigtglpesiiaaacsrsgrrvlhvdsrsyyggnwasfsfsgllswlkeyqensdivsdspvwqdqileneeaialsrkdktiqhvevfcyasqdlhedveeagalqknhalvtsansteaadsaflptedeslstmscemlteqtpssdpenalevngaevtgekenhcddktcvpstsaedmsenvpiaedtteqpkknritysqiikegrrfnidlvskllysrgllidlliksnvsryaefknitrilafregrveqvpcsradvfnskqltmvekrmlmkfltfcmeyekypdeykgyeeitfyeylktqkltpnlqyivmhsiamtsetasstidglkatknflhclgrygntpflfplygqgelpqcfcrmcavfggiyclrhsvqclvvdkesrkckaiidqfgqriisehflvedsyfpenmcsrvqyrqisravlitdrsvlktdsdqqisiltvpaeepgtfavrvielcsstmtcmkgtylvhltctssktaredlesvvqklfvpytemeieneqvekprilwalyfnmrdssdisrscyndlpsnvyvcsgpdcglgndnavkqaetlfqeicpnedfcppppnpediildgdslqpeasessaipeansetfkestnlgnleesse
(seqidno:5)
编码rep1的示例性核苷酸序列是:
atggcggatactctcccttcggagtttgatgtgatcgtaatagggacgggtttgcctgaatccatcattgcagctgcatgttcaagaagtggccggagagttctgcatgttgattcaagaagctactatggaggaaactgggccagttttagcttttcaggactattgtcctggctaaaggaataccaggaaaacagtgacattgtaagtgacagtccagtgtggcaagaccagatccttgaaaatgaagaagccattgctcttagcaggaaggacaaaactattcaacatgtggaagtattttgttatgccagtcaggatttgcatgaagatgtcgaagaagctggtgcactgcagaaaaatcatgctcttgtgacatctgcaaactccacagaagctgcagattctgccttcctgcctacggaggatgagtcattaagcactatgagctgtgaaatgctcacagaacaaactccaagcagcgatccagagaatgcgctagaagtaaatggtgctgaagtgacaggggaaaaagaaaaccattgtgatgataaaacttgtgtgccatcaacttcagcagaagacatgagtgaaaatgtgcctatagcagaagataccacagagcaaccaaagaaaaacagaattacttactcacaaattattaaagaaggcaggagatttaatattgatttagtatcaaagctgctgtattctcgaggattactaattgatcttctaatcaaatctaatgttagtcgatatgcagagtttaaaaatattaccaggattcttgcatttcgagaaggacgagtggaacaggttccgtgttccagagcagatgtctttaatagcaaacaacttactatggtagaaaagcgaatgctaatgaaatttcttacattttgtatggaatatgagaaatatcctgatgaatataaaggatatgaagagatcacattttatgaatatttaaagactcaaaaattaacccccaacctccaatatattgtcatgcattcaattgcaatgacatcagagacagccagcagcaccatagatggtctcaaagctaccaaaaactttcttcactgtcttgggcggtatggcaacactccatttttgtttcctttatatggccaaggagaactcccccagtgtttctgcaggatgtgtgctgtgtttggtggaatttattgtcttcgccattcagtacagtgccttgtagtggacaaagaatccagaaaatgtaaagcaattatagatcagtttggtcagagaataatctctgagcatttcctcgtggaggacagttactttcctgagaacatgtgctcacgtgtgcaatacaggcagatctccagggcagtgctgattacagatagatctgtcctaaaaacagattcagatcaacagatttccattttgacagtgccagcagaggaaccaggaacttttgctgttcgggtcattgagttatgttcttcaacgatgacatgcatgaaaggcacctatttggttcatttgacttgcacatcttctaaaacagcaagagaagatttagaatcagttgtgcagaaattgtttgttccatatactgaaatggagatagaaaatgaacaagtagaaaagccaagaattctgtgggctctttacttcaatatgagagattcgtcagacatcagcaggagctgttataatgatttaccatccaacgtttatgtctgctctggcccagattgtggtttaggaaatgataatgcagtcaaacaggctgaaacacttttccaggaaatctgccccaatgaagatttctgtccccctccaccaaatcctgaagacattatccttgatggagacagtttacagccagaggcttcagaatccagtgccataccagaggctaactcggagactttcaaggaaagcacaaaccttggaaacctagaggagtcctctgaataa
(seqidno:6)
编码rep1的进一步示例性核苷酸序列是:
gatatcgaattcctgcagcccggcggcaccatggcggatactctcccttcggagtttgatgtgatcgtaatagggacgggtttgcctgaatccatcattgcagctgcatgttcaagaagtggccggagagttctgcatgttgattcaagaagctactatggaggaaactgggccagttttagcttttcaggactattgtcctggctaaaggaataccaggaaaacagtgacattgtaagtgacagtccagtgtggcaagaccagatccttgaaaatgaagaagccattgctcttagcaggaaggacaaaactattcaacatgtggaagtattttgttatgccagtcaggatttgcatgaagatgtcgaagaagctggtgcactgcagaaaaatcatgctcttgtgacatctgcaaactccacagaagctgcagattctgccttcctgcctacggaggatgagtcattaagcactatgagctgtgaaatgctcacagaacaaactccaagcagcgatccagagaatgcgctagaagtaaatggtgctgaagtgacaggggaaaaagaaaaccattgtgatgataaaacttgtgtgccatcaacttcagcagaagacatgagtgaaaatgtgcctatagcagaagataccacagagcaaccaaagaaaaacagaattacttactcacaaattattaaagaaggcaggagatttaatattgatttagtatcaaagctgctgtattctcgaggattactaattgatcttctaatcaaatctaatgttagtcgatatgcagagtttaaaaatattaccaggattcttgcatttcgagaaggacgagtggaacaggttccgtgttccagagcagatgtctttaatagcaaacaacttactatggtagaaaagcgaatgctaatgaaatttcttacattttgtatggaatatgagaaatatcctgatgaatataaaggatatgaagagatcacattttatgaatatttaaagactcaaaaattaacccccaacctccaatatattgtcatgcattcaattgcaatgacatcagagacagccagcagcaccatagatggtctcaaagctaccaaaaactttcttcactgtcttgggcggtatggcaacactccatttttgtttcctttatatggccaaggagaactcccccagtgtttctgcaggatgtgtgctgtgtttggtggaatttattgtcttcgccattcagtacagtgccttgtagtggacaaagaatccagaaaatgtaaagcaattatagatcagtttggtcagagaataatctctgagcatttcctcgtggaggacagttactttcctgagaacatgtgctcacgtgtgcaatacaggcagatctccagggcagtgctgattacagatagatctgtcctaaaaacagattcagatcaacagatttccattttgacagtgccagcagaggaaccaggaacttttgctgttcgggtcattgagttatgttcttcaacgatgacatgcatgaaaggcacctatttggttcatttgacttgcacatcttctaaaacagcaagagaagatttagaatcagttgtgcagaaattgtttgttccatatactgaaatggagatagaaaatgaacaagtagaaaagccaagaattctgtgggctctttacttcaatatgagagattcgtcagacatcagcaggagctgttataatgatttaccatccaacgtttatgtctgctctggcccagattgtggtttaggaaatgataatgcagtcaaacaggctgaaacacttttccaggaaatctgccccaatgaagatttctgtccccctccaccaaatcctgaagacattatccttgatggagacagtttacagccagaggcttcagaatccagtgccataccagaggctaactcggagactttcaaggaaagcacaaaccttggaaacctagaggagtcctctgaataa
(seqidno:7)
rep1的示例性变体进一步描述于wo2012/114090(通过引用并入本文)。
活性测定
一方面,本发明提供了用于测定rab护送蛋白1(rep1)活性的方法,包括以下步骤:
(a)提供包含rep1的样品;
(b)使步骤(a)的样品与rab6a、rab牻牛儿基牻牛儿基转移酶(rabggt酶)和脂质供体底物接触;并且
(c)检测脂质化rab6a产物。
另一方面,本发明提供rab6a用于测定rab护送蛋白1(rep1)活性的用途。
测定法灵敏性是一个需要考虑的重要因素,因为它能够检测低水平的靶标,这在存在少量试剂时尤其相关(例如基因治疗试剂的情况可以就是如此)。然而,最大化测定信号的动态范围也是重要的,例如,其可以与提供低或高灵敏性的试剂无关。
本发明的方法和用途是用于测试rep1的活性,而不是测试参与rabgtp酶的异戊烯化的其他试剂,例如rabggt酶或脂质供体底物的活性,或rabgtp酶作为异戊烯化底物的活性。例如,本发明的方法可以用于测试适合于将rep1递送至靶细胞的基因治疗载体或用于载体储液(例如药物储液)的质量控制分析。
在一个实施方案中,包含rep1的样品来自经遗传工程化改造以表达rep1的细胞。优选地,用包含编码rep1的核苷酸序列的载体转染或转导细胞,以提供经遗传工程化改造以表达rep1的细胞。优选地,载体是病毒载体。
在一个实施方案中,使用包含编码rep1的核苷酸序列的病毒载体表达rep1。
经遗传工程化改造以表达rep1的细胞(其可以为此类细胞的群体)可以是适合于遗传工程和rep1表达的任何细胞,例如来自细胞系(例如hek293)的细胞。细胞可以是例如人或小鼠细胞。优选地,细胞是人细胞。例如,细胞可以是视网膜细胞,例如视网膜色素上皮细胞或光感受器细胞。在一个实施方案中,细胞是hek293细胞。在另一个实施方案中,细胞是arpe-19细胞。在另一个实施方案中,细胞是ht1080细胞。
优选地,rab6a和/或rabggt酶来自标准来源,使得它们在重复实验中提供最小的变化或没有变化。优选地,rab6a和/或rabggt酶是基本上纯的(即基本上不包含干扰本发明的方法或用途的蛋白质污染物)。
因此,该方法或用途可以包括进行多个实验(例如包括步骤(a)至(c)),其中涉及包含rep1的样品的参数是变化的,而其他参数(例如涉及rab6a、rabggt酶和脂质供体底物的参数保持恒定。此类参数可以包括例如相关蛋白质(例如rep1)的氨基酸序列、用于在细胞中表达rep1的载体中包含的编码rep1的核苷酸序列、用于将编码rep1的核苷酸序列递送到细胞的载体类型(例如病毒载体的类型,例如腺伴随病毒(aav)载体的类型)、rep1的浓度和/或用于将编码rep1的核苷酸序列递送到细胞的载体的感染复数(moi)。
术语“活性”在本文中用于指rep1促进rabgtp酶(例如rab6a)脂质化的能力。尽管rep1本身不催化脂质化,但是它是rabggt酶催化其底物rabgtp酶的脂质化需要的。因此,可以通过测定在某些条件下脂质化的rabgtp酶(即rab6a)的量来测量rep1的活性。
术语“功效”在本文中就包含编码rep1的核苷酸序列的载体的功效而言使用,指载体为由载体转染或转导的细胞提供rep1活性的能力。
如本文所用,术语“脂质化rab6a产物”是指已加入脂质部分的rab6a。优选地,脂质化rab6a产物是异戊烯化的rab6a,例如牻牛儿基牻牛儿基因rab6a或生物素-牻牛儿基化rab6a。
优选地,检测脂质化rab6a产物的步骤提供了脂质化rab6a产物的量的量化。
脂质化rab6a的检测可以通过任何合适的方法进行,例如酶联免疫吸附测定法(elisa)、western印迹、放射自显术术(例如利用同位素标记的,例如氚化的,脂质的供体底物),层析(例如hplc或fplc)和/或基于质谱术的方法(例如lc/ms)。
在一个实施方案中,使用western印迹检测脂质化rab6a产物。在优选的实施方案中,使用elisa检测脂质化rab6a产物。
举例来说,可以使用生物素-牻牛儿基焦磷酸盐/酯脂质供体底物,根据本发明的方法进行异戊烯化反应。可以对反应产物进行western印迹分析,其中可以通过与例如链霉抗生物素蛋白-辣根过氧化物酶缀合物直接温育来检测脂质化rab6a产物(即生物素-牻牛儿基化rab6a)。脂质化的rab6a(即生物素-牻牛儿基化rab6a)的量化可以通过所得western印迹的光密度分析来实现,其可以通过任何合适的手段(例如使用imagestudiolite软件(li-cor))进行。
作为另一个实例,可以使用生物素-牻牛儿基焦磷酸盐/酯脂质供体底物,根据本发明的方法进行异戊烯化反应。可以对反应产物进行elisa分析,其中rab6a可以直接固定在平板上或使用已附着到平板的抗体固定(即夹心elisa);然后可以通过与例如链霉抗生物素蛋白-辣根过氧化物酶缀合物一起温育来检测脂质化rab6a产物(即生物素-牻牛儿基化rab6a)。脂质化的rab6a(即生物素-牻牛儿基化rab6a)的量化可以通过任何合适的手段(例如使用分光光度计、荧光计或发光计进行检测)来实现。
根据需要(例如用于控制目的),可以将进一步的检测步骤掺入到本发明的方法中,例如检测反应中存在的rep1的量或检测β-肌动蛋白的量(例如作为上样对照)。
在一个实施方案中,该方法包括比较脂质化的rab6a产物(例如,异戊烯化的,例如牻牛儿基牻牛儿基化或生物素-牻牛儿基化rab6a)的量与从对照实验,例如使用rep1的已知或标准样品的实验确定的量的进一步的步骤。
在另一个实施方案中,该方法包括比较脂质化的rab6a产物(例如异戊烯化的,例如牻牛儿基牻牛儿基化或生物素-牻牛儿基化rab6a)的量与参考水平的进一步的步骤。
与此类对照实验或参考水平的比较可以提供rep1相对于已知或公认的标准品的活性的量度(例如,比已知或公认的标准品更好或更差)。
本发明的方法可以例如用于质量控制分析用于治疗无脉络膜的基因治疗载体,优选地包含编码rep1的核苷酸序列的aav载体颗粒,其中通过本发明的方法测定的载体的输出活性或功效高于阈值活性或在规定的靶标范围内(例如通过与对照实验或参照水平比较)指示载体适合于基因治疗目的。
异戊烯化反应的条件(例如在本发明方法的步骤(b)中发生)没有特别限制,条件是它们基本上不干扰rab6a的异戊烯化。
包含rep1的样品可以以任何合适的形式配制,例如可以在包含约50mmhepes、50mmnacl、2mmmgcl2、1mmdtt和蛋白酶抑制剂混合物(roche)的约ph7.5的异戊烯化缓冲液中制备样品。
包含rep1的样品可以例如包含约1-100、1-75、1-50、1-40、1-30、1-20或1-10μg总蛋白质。包含rep1的样品可以例如包含约10-100、10-75、10-50、10-40、10-30或10-20μg的总蛋白质。优选地,包含rep1的样品包含约10-30μg的总蛋白质,例如,约10、15、20、25或30μg的总蛋白质。
rab6a可以例如浓度为约1-25、1-20、1-15、1-10或1-5μm,优选约1-5μm。例如,rab6a可以浓度为约1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24或25μm,优选约4μm。
rabggt酶可以例如浓度为约1-25、1-20、1-15、1-10、1-5或1-2.5μm,优选约1-2.5μm。例如,rabggt酶可以浓度为约1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24或25μm,优选约2μm。
例如,脂质供体底物(例如生物素-牻牛儿基焦磷酸盐/酯(bgpp))浓度可以是约1-25、1-20、1-15、1-10或1-5μm,优选约1-5μm。例如,脂质供体底物(例如生物素-牻牛儿基焦磷酸盐/酯(bgpp))的浓度可以为约1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24或25μm,优选约4μm。
异戊烯化反应可以在任何合适的缓冲液中进行,例如,反应可以在包含约50mmhepes、50mmnacl、2mmmgcl2、1mmdtt和蛋白酶抑制剂混合物(roche)的约ph7.5的异戊烯化缓冲液中进行。
异戊烯化反应可以在任何合适的温度(例如约37℃)下进行任何合适的时间长度。例如,异戊烯化反应可以进行约1-10、1-7.5、1-5、1-2.5或1-2小时,优选约1-2小时。例如,异戊烯化反应可以进行约1、2、3、4、5、6、7、8、9或10小时,优选约2小时。
无脉络膜
无脉络膜是一种罕见的x连锁的进行性眼脉络膜、视网膜色素上皮和光感受器变性。患病男性的典型自然史是青少年时期夜盲症的发作,然后在20岁和30随期间逐渐丧失周边视力,导致40岁完全失明。女性携带者有轻微的症状,最明显的是夜盲症,但偶尔可能有更严重的表型。
无脉络膜是由chm基因突变引起的,该基因位于x染色体21q区域。rab护送蛋白2(rep2)(其与rep1是75%同源的)补偿身体的大多数细胞中的任何rep1缺陷。然而,出于尚不清楚的原因,rep2无法补偿眼中的rep1缺陷。这导致rab护送蛋白活性不足以维持靶rabgtp酶的正常异戊烯化并引起细胞功能障碍并最终导致细胞死亡,主要影响外视网膜和脉络膜。
可以通过将rep1转基因的功能性拷贝提供给受影响的眼细胞来成功地治疗无脉络膜(maclaren,r.e.etal.(2014)lancet383:1129-37)。
载体
载体是允许或促进实体从一个环境转移到另一个环境的工具。根据本发明,并且举例来说,重组核酸技术中使用的一些载体允许实体,例如核酸区段(例如异源dna区段,例如异源cdna区段)转移到靶细胞中。载体可以用于维持细胞内的异源核酸(例如dna或rna),促进包含核酸区段的载体的复制或促进由核酸区段编码的蛋白质的表达的目的。
载体可以是非病毒的或病毒的。用于重组核酸技术的载体的实例包括但不限于质粒、染色体、人工染色体和病毒。载体也可以是例如裸核酸(例如dna或rna)。在其最简单的形式中,载体本身可以是感兴趣的核苷酸。
用于本发明的载体可以是例如质粒或病毒载体,并且可以包含用于表达多核苷酸的启动子和任选的启动子调节物。
病毒载体
在优选的实施方案中,本发明的载体是病毒载体。优选地,病毒载体是病毒载体颗粒的形式。
病毒载体可以是例如腺伴随病毒(aav)、逆转录病毒、慢病毒或腺病毒载体。优选地,病毒载体是aav载体。
术语“基因治疗载体”在本文中用于指适合于用于基因治疗的载体,并且包括例如病毒(例如aav)载体和载体颗粒。
变体、衍生物、类似物、同源物和片段
除了本文提到的特定蛋白质和核苷酸外,本发明还涵盖其变体、衍生物、类似物、同源物和片段的用途。
在本发明的上下文中,任何给定序列的变体是下述序列,其中残基的特定序列(无论氨基酸或核酸残基)已经以使得所讨论的多肽或多核苷酸基本上保留其功能的方式修饰。可以通过天然存在的蛋白质中存在的至少一个残基的添加、缺失、取代、修饰、置换和/或变异来获得变体序列。
如本文所用,术语“衍生物”就本发明的蛋白质或多肽而言包含序列的一个(或多个)氨基酸残基的任何取代、变异、修饰、替换、缺失和/或添加,只要所得的蛋白质或多肽基本上保留其至少一种其内源功能。
如本文所用,术语“类似物”就多肽或多核苷酸而言包括任何模拟物,即拥有其模拟的多肽或多核苷酸的至少一种内源功能的化学化合物。
通常,可以进行氨基酸取代,例如1、2或3至10或20个取代,只要经修饰的序列基本上保留所需要的活性或能力。氨基酸取代可以包含使用非天然存在的类似物。
本发明中使用的蛋白质也可以具有氨基酸残基的缺失、插入或取代,其产生沉默变化并产生功能等同的蛋白质。可以基于残基的极性、电荷、溶解度、疏水性、亲水性和/或两亲性质的相似性来进行有意的氨基酸取代,只要保留内源功能。例如,带负电荷的氨基酸包括天冬氨酸和谷氨酸;带正电荷的氨基酸包括赖氨酸和精氨酸;并且具有相似亲水性值的不带电荷的极性首基的氨基酸包括天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸和酪氨酸。
可以进行保守取代,例如根据下表。第二列中相同块中并且优选在第三列中的相同行中的氨基酸可以彼此替换:
如本文所用,术语“同源物”是指与野生型氨基酸序列和野生型核苷酸序列具有一定同源性的实体。术语“同源性”可以等同于“同一性”。
同源序列可以包含与主题序列可以是至少50%,55%,60%,65%,70%,75%,80%,85%或90%相同,优选至少95%或97%或99%相同的氨基酸序列。通常,同源物将包含与主题氨基酸序列相同的活性位点等。尽管也可以根据相似性(即具有相似化学性质/功能的氨基酸残基)考虑同源性,但在本发明的上下文中,优选根据序列同一性表达同源性。
同源序列可以包含与主题序列可以至少50%,55%,60%,65%,70%,75%,80%,85%或90%相同,优选至少95%或97%或99%相同的核苷酸序列。虽然同源性也可以根据相似性考虑,但在本发明的上下文中,优选根据序列同一性表达同源性。
优选地,提及与本文详述的任一种seqidno具有百分比同一性的序列是指在提及的seqidno的整个长度里具有所述百分比同一性的序列。
可以通过眼,或者更通常地借助于容易获得的序列比较程序进行同源性比较。这些商品化的计算机程序可以计算两个或更多个序列之间的同源性或同一性百分比。
可以在连续序列里计算同源性百分比,即将一个序列与另一个序列比对,并且将一个序列中的每个氨基酸与另一个序列中的相应氨基酸直接比较,一次一个残基。这称为“无缺口(ungapped)”比对。通常,仅在相对少量的残基里进行此类无缺口比对。
虽然这是一种非常简单且一致的方法,但是它未能考虑例如在一对其他方面相同的序列中,核苷酸序列中的一个插入或缺失可以使以下密码子不对齐,因此当进行全局比对时潜在导致百分比同源性的大幅降低。因此,大多数序列比较方法设计为产生最佳比对,其考虑可能的插入和缺失而不对整体同源性评分过度罚分。这是通过在序列比对中插入“缺口”以试图最大化局部同源性来实现的。
然而,这些更复杂的方法对比对中出现的每个缺口分配“缺口罚分”,使得对于相同数目的相同氨基酸,具有尽可能少的缺口(反映两个比较序列之间较高的相关性)的序列比对将获得比具有许多缺口的序列比对更高的得分。通常使用“仿射缺口代价(affinegapcost)”,其对于缺口的存在收取相对高的代价并且对缺口中的每个后续残基收取较小的罚分。这是最常用的缺口评分系统。高缺口罚分当然会产生具有更少缺口的优化的比对。大多数比对程序允许修改缺口罚分。但是,当使用此类软件进行序列比较时优选使用默认值。例如,当使用gcgwisconsinbestfit包时,氨基酸序列的默认缺口罚分对于缺口为-12,并且对于每个延伸为-4。
因此,计算最大百分比同源性首先需要产生最佳比对,考虑缺口罚分。用于进行此类比对的合适的计算机程序是gcgwisconsinbestfit包(universityofwisconsin,u.s.a.;devereuxetal.(1984)nucleicacidsres.12:387)。可以进行序列比较的其他软件的实例包括但不限于blast包(参见ausubeletal.(1999)同上–第18章),fasta(atschuletal.(1990)j.mol.biol.403-410)和geneworks比较工具套件。blast和fasta两者可用于离线和在线搜索(参见ausubel等人(1999)同上,第7-58至7-60页)。然而,对于某些应用,优选使用gcgbestfit程序。另一种称为blast2序列的工具也可用于比较蛋白质和核苷酸序列(参见femsmicrobiol.lett.(1999)174:247-50;和femsmicrobiol.lett.(1999)177:187-8)。
虽然可以根据同一性测量最终的百分比同源性,但是比对过程本身通常不基于全有或全无对比较(all-or-nothingpaircomparison)。相反,通常使用缩放的相似性得分矩阵,其基于化学相似性或进化距离为每个成对比较分配得分。常用的此类矩阵的一个实例是blosum62矩阵-blast程序套件的默认矩阵。gcgwisconsin程序通常使用公共默认值或自定义符号比较表(如果提供的话)(关于更多详情,参见用户手册)。对于某些应用,优选使用gcg包的公共默认值,或者在其他软件的情况下,使用默认矩阵,例如blosum62。
一旦软件产生了最佳比对,就可能计算同源性百分比,优选序列同一性百分比。软件通常进行这点作为序列比较的一部分并生成数字结果。
本发明的全长多肽或多核苷酸的“片段”也是变体,并且该术语通常是指功能上或例如在测定法中感兴趣的多肽或多核苷酸的选定区域。因此,“片段”是指作为全长多肽或多核苷酸的一部分的氨基酸或核酸序列。
可以使用标准重组dna技术如定点诱变来制备此类变体。在要进行插入的情况下,可以制备编码插入以及对应于插入位点任一侧的天然存在序列的5'和3'侧翼区的合成dna。侧翼区会含有对应于天然存在序列中的位点的方便的限制性位点,以便可以用合适的酶切割序列,并将合成的dna连接到切口中。然后根据本发明表达dna以制备编码的蛋白质。这些方法仅阐述本领域已知的用于操纵dna序列的众多标准技术,并且也可以使用其他已知技术。
密码子优化
用于本发明的多核苷酸可以是经密码子优化的。密码子优化先前已在wo1999/41397和wo2001/79518中描述。不同的细胞在其特定密码子选择上不同。该密码子偏倚对应于细胞类型中特定trna的相对丰度的偏差。通过改变序列中的密码子以使它们适合于与相应trna的相对丰度相匹配,可以增加表达。出于同样的原因,可以通过有意选择已知相应的trna在特定细胞类型中罕见的密码子来降低表达。因此,可获得额外的翻译控制程度。
实施例
材料和方法(实施例1-2)
细胞转导和收获
以一系列感染复数(moi,基因组颗粒/细胞)用raav2/2-rep1处理培养的hek293细胞。将raav2/2-gfp平行用作对照载体,并监测荧光的转基因表达的开始。
平行进行对未转导的细胞和+aav-gfp转导的和+aav-rep1转导的细胞的实验。
使用以下方案在转导后第5天制备细胞裂解物:用pbs清洗细胞,并在冰上与ph7.5的异戊烯化缓冲液(50mmhepes,50mmnacl,2mmmgcl2,1mmdtt和蛋白酶抑制剂混合物(roche))一起温育5分钟;然后用细胞刮刀将细胞刮到1.5ml管中,并且在冰上温育15分钟;随后,通过连接至1ml注射器的26-g注射器针将细胞推动20次来破坏它们。
将裂解的细胞在4℃以1500xg离心5分钟。然后将上清液转移到丙酸纤维素管中并在4℃以100000xg离心1小时。将来自第二离心步骤的上清液用于体外异戊烯化反应(如下所述)。
总蛋白质量化
使用bradford方法根据制造商的说明书(quickstarttmbradford1xdyereagent,biorad,#500-0205)量化总细胞蛋白质浓度。从标准曲线外推样品值。
体外异戊烯化反应
使用异戊烯化缓冲液中的冷冻的细胞裂解物(10-30μg)、2μmrabggt酶、4μmrab蛋白(rab27a或rab6a)和作为脂质供体的5μm生物素-牻牛儿基焦磷酸盐(bgpp)建立异戊烯化反应。所有反应均补充有新鲜gdp(二磷酸鸟苷,20μm)和dtt(1mm)。
对于阳性对照样品,将鱼rep1(对于量,参见个别实验)加入到含有来自未转导的细胞的裂解物的异戊烯化反应。
将反应在37℃温育2小时,然后通过加入样品缓冲液(laemmli缓冲液,2x浓缩物,sigma#s3401)终止反应。该缓冲液含有4%sds、20%甘油、10%2-巯基乙醇、0.004%溴酚蓝和0.125mtrishcl,ph约6.8。
进行western印迹(wb)以检测人rep-1、β-肌动蛋白(作为上样对照)和生物素化的rab蛋白(rab27a或rab6a)。
为了检测人rep1,使用来自millipore的小鼠单克隆抗体(克隆2f1,#mabn52)。为了检测β-肌动蛋白,使用来自thermofisherscientific的小鼠单克隆抗体(克隆ac-15,#am4302)。这两次检测之后是二抗标记步骤(驴抗小鼠hrp,abcam,#ab98799)。
通过与链霉抗生物素蛋白-hrp(thermofisherscientific,#43-4323)直接温育来检测生物素化的脂质供体对合适的rab底物的掺入。
使用ecl底物和odysseyfc检测系统(li-cor)检测所有膜。使用imagestudiolite软件(li-cor)定量分析条带的强度。
实施例1:rab27a作为异戊烯化底物
为了测试使用rab27a作为底物的异戊烯化测定法的灵敏性,使用以下细胞平行进行实验:
(a)未转导的细胞;
(b)以10000的moi用aav-gfp转导的细胞;和
(c)以10000的moi用aav-rep1转导的细胞。
使用总体积为12.5μl的10μg裂解物建立异戊烯化反应。对阳性对照掺加2μm鱼rep1。
结果指示rab27a是用于评估用aav-rep1转导细胞后rep1功能的异戊烯化测定法的底物(图1)。然而,来自wb半量化的信号不是很强。
用增加量的总细胞蛋白重复该研究以增加wb条带强度。
使用总体积为22μl的30μg裂解物建立异戊烯化反应。对阳性对照掺加1μm鱼rep1。
结果证实rab27a作为用于异戊烯化测定法的底物起作用以评估用aav-rep1转导细胞后的rep1功能(图2)。此外,与使用10μg裂解物获得的数据相比,wb信号的强度已经增加。然而,信号仍然不是很强。理想地,观察到当用aav-rep1转导细胞时,异戊烯化的rab蛋白的较大增加。
实施例2:rab6a作为异戊烯化底物
为了使用rab6a作为底物测试异戊烯化测定法的灵敏性,使用以下细胞(实施例1中使用的相同细胞裂解物)平行进行实验:
(a)未转导的细胞;
(b)以10000的moi用aav-gfp转导的细胞;和
(c)以10000的moi用aav-rep1转导的细胞。
使用总体积为20μl的20μg裂解物建立异戊烯化反应。对阳性对照掺加1μm鱼rep1。
结果指示rab6a是用于评估用aav-rep1转导细胞后rep1功能的异戊烯化测定法的有效底物(图3)。
与图2中显示的数据相比,尽管使用较少的总蛋白,但是对于经aav-rep1转导的细胞,wb信号的强度增加了约10倍。此外,与图2中显示的数据相比,阳性对照的条带强度高约100倍,证实了基于rab6a的测定法的增加的灵敏性。
数据还证明增加的灵敏性也实现检测内源性水平的差异,从而使测定法更准确。
在通过使用rab6a作为异戊烯化底物提供的增加的灵敏性的测定法的成功演示后,我们使用aav-rep1研究的不同moi重复该测定法,无论rab6a异戊烯化是否与aav-rep1的量相关。
使用以下细胞平行进行实验:
(a)未转导的细胞;
(b)以250的moi用aav-rep1转导的细胞,;
(c)以1000的moi用aav-rep1转导的细胞;
(d)以5000的moi用aav-rep1转导的细胞;
(e)以10000的moi用aav-rep1转导的细胞;和
(f)以20000的moi用aav-rep1转导的细胞。
使用总体积为15μl的20μg裂解物建立异戊烯化反应。对阳性对照掺加0.5μm鱼rep1。
结果证实rab6a是用于评估用aav-rep1转导细胞后rep1功能的异戊烯化测定法的有效底物(图4),并且进一步证明生物素化脂质供体在rab6a中的掺入与用于细胞转导的aav-rep1的量相关。
在验证rab6a作为有效的测定法底物后,我们然后测试目前在我们的1期临床试验中使用的aav-rep1载体(maclaren,r.e.etal.(2014)lancet383:1129-37)。
使用以下细胞平行进行实验:
(a)未转导的细胞(#29、#30和#31);
(b)以10000的moi用aav-rep1转导的细胞(#32、#33和#34);和
(c)以10000的moi用gmp级aav-rep1转导的细胞(#35、#36和#37)。
使用总体积为15μl的20μg裂解物建立异戊烯化反应。对阳性对照掺加0.5μm鱼rep1。
结果与先前的实验一致,并证实生物素化的脂质供体在rab6a中的掺入与用于细胞转导的aav-rep1的量相关(图5)。
实施例3:rab6a作为使用arpe-19细胞的异戊烯化反应中的底物
细胞转导和收获
用raav2/2-rep1以10,000个基因组颗粒/细胞的moi处理培养的arpe-19细胞。在转导后第13天制备细胞裂解物:用pbs清洗细胞,并在冰上与ph7.5的异戊烯化缓冲液(50mmhepes,50mmnacl,2mmmgcl2,1mmdtt和蛋白酶抑制剂混合物(roche))一起温育。使用细胞刮刀将细胞刮入1.5ml管中,并在冰上温育15分钟。通过连接至1ml注射器的26-g注射器针将细胞推动20次来破坏它们。将细胞以1,500g在4℃旋转5分钟。然后将上清液转移到丙酸纤维素管中,并在4℃以100,000g离心1小时。将上清液用于体外异戊烯化反应。
总蛋白质量化
使用bradford方法根据制造商的说明书(quickstarttmbradford1xdyereagent,biorad,#500-0205)量化总细胞蛋白质。从标准曲线外推样品值。
体外异戊烯化反应
使用戊烯化缓冲液中的冷冻的细胞裂解物(15μg),2μmrabggt酶,4μmrab蛋白(rab6a)和5μm生物素-牻牛儿基焦磷酸盐作为脂质供体,建立异戊烯化反应。所有反应均补充有新鲜gdp(20μm)和dtt(1mm)。在阳性对照样品中,将鱼rep1(对于量,参见实验)加入到含有未转导的细胞裂解物的异戊烯化反应。
将反应在37℃温育2小时,然后通过加入sds-page样品缓冲液终止反应。
进行western印迹法(wb)以检测人rep-1、β-肌动蛋白(上样对照)和生物素化的rab蛋白(rab27a或rab6a)。为了检测人rep1,使用来自millipore的小鼠单克隆抗体(克隆2f1,#mabn52)。为了检测β-肌动蛋白,使用来自thermofisherscientific的小鼠单克隆抗体(克隆ac-15,#am4302)。这两次检测之后是二抗标记步骤(驴抗小鼠hrp,abcam,#ab98799)。通过与链霉抗生物素蛋白-hrp(thermofisherscientific,#43-4323)直接温育来检测生物素化的脂质供体对合适的rab底物的掺入。使用ecl底物和odysseyfc检测系统(li-cor)检测所有膜。使用imagestudilite软件(li-cor)定量分析条带的强度。
结果与讨论
为了测试在arpe-19细胞(人视网膜色素上皮细胞)中使用rab6a作为底物的异戊烯化测定法,使用以下细胞平行进行实验:
(a)未转导的细胞(#86和#87);和
(b)细胞+aav-rep1moi10,000(#90和#91)-r&d级载体。
使用总体积为45μl的15μg裂解物建立异戊烯化反应。对阳性对照掺加0.1μm的鱼rep1。
结果指示rab6a作为异戊烯化测定法的底物起作用,以评估用aav-rep1转导arpe-19细胞后的rep1功能(图6)。
实施例4:rab6a作为使用ht1080细胞的异戊烯化反应中的底物
细胞转导和收获
以10,000个基因组颗粒/细胞的moi用raav2/2-rep1处理培养的ht1080细胞。在转导后第5天制备细胞裂解物:用pbs清洗细胞,并在冰上与ph7.5的异戊烯化缓冲液(50mmhepes,50mmnacl,2mmmgcl2,1mmdtt和蛋白酶抑制剂混合物(roche))一起温育。使用细胞刮刀将细胞刮入1.5ml管中,并在冰上温育15分钟。通过连接至1ml注射器的26-g注射器针将细胞推动20次来破坏它们。将细胞以1,500g在4℃旋转5分钟。然后将上清液转移到丙酸纤维素管中,并在4℃以100,000g离心1小时。将上清液用于体外异戊烯化反应。
总蛋白质量化
使用bradford方法根据制造商的说明书(quickstarttmbradford1xdyereagent,biorad,#500-0205)量化总细胞蛋白质。从标准曲线外推样品值。
体外异戊烯化反应
使用异戊烯化缓冲液中的冷冻的细胞裂解物(20μg)、2μmrabggt酶、4μmrab蛋白(rab6a)和5μm作为脂质供体的生物素-牻牛儿基焦磷酸盐建立异戊烯化反应。所有反应均补充有新鲜gdp(20μm)和dtt(1mm)。在阳性对照样品中,将鱼rep1(对于量,参见实验)加入到含有未转导的细胞裂解物的异戊烯化反应。
将反应在37℃温育2小时,然后通过加入sds-page样品缓冲液终止反应。
进行western印迹法(wb)以检测人rep-1、β-肌动蛋白(上样对照)和生物素化的rab蛋白(rab27a或rab6a)。为了检测人rep1,使用来自millipore的小鼠单克隆抗体(克隆2f1,#mabn52)。为了检测β-肌动蛋白,使用来自thermofisherscientific的小鼠单克隆抗体(克隆ac-15,#am4302)。这两次检测之后是二抗标记步骤(驴抗小鼠hrp,abcam,#ab98799)。通过与链霉抗生物素蛋白-hrp(thermofisherscientific,#43-4323)直接温育来检测生物素化的脂质供体对合适的rab底物的掺入。使用ecl底物和odysseyfc检测系统(li-cor)检测所有膜。使用imagestudilite软件(li-cor)定量分析条带的强度。
结果与讨论
为了测试在ht1080细胞中使用rab6a作为底物的异戊烯化测定法,使用以下细胞平行进行实验:
(a)未转导的细胞(#56和#57);
(b)细胞+aav-rep1moi10,000(#60和#61)-r&d级载体;和
(c)细胞+aav-rep1moi10,000(#64和#65)-临床级载体。
使用总体积为20μl的20μg裂解物建立异戊烯化反应。对阳性对照掺加0.1μm的鱼rep1。
结果指示rab6a作为异戊烯化测定法的底物起作用,以评估用aav-rep1转导ht1080细胞后的rep1功能(图7)。
实施例5:rab27a和rab6a在异戊烯化反应中作为底物的比较
使用与图4所示实验中相同的细胞裂解物:
(a)未转导的细胞;
(a)细胞+aav-rep1moi250;
(a)细胞+aav-rep1moi1,000;
(a)细胞+aav-rep1moi5,000;
(a)细胞+aav-rep1moi10,000;和
(a)细胞+aav-rep1moi20,000。
使用总体积为15μl的20μg裂解物和2种不同底物:rab27a和rab6a建立异戊烯化反应。对阳性对照(每种底物一种)掺加0.1μm鱼rep1。在sds-page上平行运行样品并同时检测。
结果与讨论
rab27a和rab6a两者均作为异戊烯化测定法的底物起作用以评估用aav-rep1转导细胞后的rep1功能。
生物素化的脂质供体的掺入与用于每种所用底物的细胞转导的aav-rep1的量相关(图8)。
来自生物素化的rab6a的条带密度高于rab27a的条带密度,这指示rab6a是用于测定相对效力的平行线分析的更合适的底物。
实施例6:在使用不同条件的异戊烯化反应中作为底物的rab27a和rab6a性能的比较
制备未转导的293细胞裂解物以用于使用rab27a和rab6a的此实验。测试的条件显示在图9的表中。在sds-page上平行样品进行并同时检测。
结果与讨论
rab27a和rab6a两者均作为异戊烯化测定法的底物起作用,以评估内源rep1功能。
生物素化的脂质供体的掺入与所用每种底物的反应中总蛋白质的量相关。
比较条件,反应中rab底物的浓度看起来对信号的影响最大。
与rab27a相比,当使用rab6a时,生物素化的底物增加2.5倍。
实施例7:在用aav2-rep1转导的裂解物中的异戊烯化反应中rab27a和rab6a作为底物的比较
使用增加的aav2-rep1(r&d材料)的moi制备新的裂解物(一式三份):
(a)未转导的细胞;
(b)细胞+aav-rep1moi100;
(c)细胞+aav-rep1moi500;
(d)细胞+aav-rep1moi1,000;
(e)细胞+aav-rep1moi5,000;
(f)细胞+aav-rep1moi10,000;
(g)细胞+aav-rep1moi20,000;和
(h)细胞+aav-rep1moi50,000。
使用20μg总蛋白、2μmrab底物(rab27a或rab6a)和2μmrabggt酶,以10μl的总体积制备异戊烯化反应。对阳性对照(每种底物一种)掺加0.1μm鱼rep1。
在sds-page上平行运行来自每个重复的样品,并使用imagestudiolite软件检测生物素化的底物(1:10,000)、作为上样对照的β-肌动蛋白(1:50,000)和人rep1(1:2,500)。使用prism软件绘制来自生物素化底物的条带密度的半量化的数据(图10)。
结果
在所有分析的样品中β-肌动蛋白的水平是相似的。未转导细胞(和阳性对照样品)显示内源性rep1水平。用aav-rep1转导的细胞显示rep1水平的增加,其与所用的moi直接相关。由于鱼rep1活性,阳性对照显示更强的生物素掺入。
用底物和moi作为因子的所有三个重复的双因素anova分析发现这两者是高度显著的(p<0.0001)。在给定moi时底物效应的bonferroni多重比较检验在5,000(p=0.0023)以及所有以上(p<0.0001)的moi时显示显著成对差异。
讨论
rab27a和rab6a两者均作为异戊烯化测定法的底物起作用以评估用aav-rep1转导细胞后的rep1功能。
生物素化底物的条带密度的半量化仅显示rab6a的值显著高于对rab27a获得的值。
上述说明书中提及的所有出版物均通过引用并入本文。在不脱离本发明的范围和精神的情况下,本发明所述方法和用途的各种修改和变化对于本领域技术人员来说是显而易见的。尽管已经结合具体的优选实施方案描述了本发明,但是应当理解,要求保护的本发明不应过度限于此类具体的实施方案。实际上,对于生物化学和生物技术或相关领域的技术人员显而易见的、用于实施本发明的所述模式的各种修改旨在落入所附权利要求书的范围内。
序列表
<110>夜星有限公司
<120>异戊烯化测定法
<130>p108257pct
<150>gb1604146.9
<151>2016-03-10
<160>9
<170>patentinversion3.5
<210>1
<211>208
<212>prt
<213>智人
<400>1
metserthrglyglyasppheglyasnproleuarglysphelysleu
151015
valpheleuglygluglnservalglylysthrserleuilethrarg
202530
phemettyraspserpheaspasnthrtyrglnalathrileglyile
354045
asppheleuserlysthrmettyrleugluaspargthrvalargleu
505560
glnleutrpaspthralaglyglngluargpheargserleuilepro
65707580
sertyrileargaspserthrvalalavalvalvaltyraspilethr
859095
asnvalasnserpheglnglnthrthrlystrpileaspaspvalarg
100105110
thrgluargglyseraspvalileilemetleuvalglyasnlysthr
115120125
aspleualaasplysargglnvalserileglugluglygluarglys
130135140
alalysgluleuasnvalmetpheilegluthrseralalysalagly
145150155160
tyrasnvallysglnleupheargargvalalaalaalaleuprogly
165170175
metgluserthrglnaspargserarggluaspmetileaspilelys
180185190
leuglulysproglngluglnprovalsergluglyglycyssercys
195200205
<210>2
<211>627
<212>dna
<213>智人
<400>2
atgtccacgggcggagacttcgggaatccgctgaggaaattcaagctggtgttcctgggg60
gagcaaagcgttggaaagacatctttgatcaccagattcatgtatgacagttttgacaac120
acctatcaggcaacaattggcattgactttttatcaaaaactatgtacttggaggatcga180
acagtacgattgcaattatgggacacagcaggtcaagagcggttcaggagcttgattcct240
agctacattcgtgactccactgtggcagttgttgtttatgatatcacaaatgttaactca300
ttccagcaaactacaaagtggattgatgatgtcagaacagaaagaggaagtgatgttatc360
atcatgctagtaggaaataaaacagatcttgctgacaagaggcaagtgtcaattgaggag420
ggagagaggaaagccaaagagctgaatgttatgtttattgaaactagtgcaaaagctgga480
tacaatgtaaagcagctctttcgacgtgtagcagcagctttgccgggaatggaaagcaca540
caggacagaagcagagaagatatgattgacataaaactggaaaagcctcaggagcaacca600
gtcagtgaaggaggctgttcctgctaa627
<210>3
<211>567
<212>prt
<213>智人
<400>3
methisglyargleulysvallysthrserglugluglnalagluala
151015
lysargleugluarggluglnlysleulysleutyrglnseralathr
202530
glnalavalpheglnlysargglnalaglygluleuaspgluserval
354045
leugluleuthrserglnileleuglyalaasnproaspphealathr
505560
leutrpasncysargarggluvalleuglnglnleugluthrglnlys
65707580
serproglugluleualaalaleuvallysalagluleuglypheleu
859095
glusercysleuargvalasnprolyssertyrglythrtrphishis
100105110
argcystrpleuleuglyargleuprogluproasntrpthrargglu
115120125
leugluleucysalaargpheleugluvalaspgluargasnphehis
130135140
cystrpasptyrargargphevalalathrglnalaalavalpropro
145150155160
alaglugluleualaphethraspserleuilethrargasnpheser
165170175
asntyrsersertrphistyrargsercysleuleuproglnleuhis
180185190
proglnproaspserglyproglnglyargleuprogluaspvalleu
195200205
leulysgluleugluleuvalglnasnalaphephethraspproasn
210215220
aspglnseralatrpphetyrhisargtrpleuleuglyargalaasp
225230235240
proglnaspalaleuargcysleuhisvalserargaspglualacys
245250255
leuthrvalserpheserargproleuleuvalglyserargmetglu
260265270
ileleuleuleumetvalaspaspserproleuilevalglutrparg
275280285
thrproaspglyargasnargproserhisvaltrpleucysaspleu
290295300
proalaalaserleuasnaspglnleuproglnhisthrpheargval
305310315320
iletrpthralaglyaspvalglnlysglucysvalleuleulysgly
325330335
argglngluglytrpcysargaspserthrthraspgluglnleuphe
340345350
argcysgluleuservalglulysserthrvalleuglnsergluleu
355360365
glusercyslysgluleuglngluleugluprogluasnlystrpcys
370375380
leuleuthrileileleuleumetargalaleuaspproleuleutyr
385390395400
glulysgluthrleuglntyrpheglnthrleulysalavalasppro
405410415
metargalathrtyrleuaspaspleuargserlyspheleuleuglu
420425430
asnservalleulysmetglutyralagluvalargvalleuhisleu
435440445
alahislysaspleuthrvalleucyshisleugluglnleuleuleu
450455460
valthrhisleuaspleuserhisasnargleuargthrleupropro
465470475480
alaleualaalaleuargcysleugluvalleuglnalaseraspasn
485490495
alailegluserleuaspglyvalthrasnleuproargleuglnglu
500505510
leuleuleucysasnasnargleuglnglnproalavalleuglnpro
515520525
leualasercysproargleuvalleuleuasnleuglnglyasnpro
530535540
leucysglnalavalglyileleugluglnleualagluleuleupro
545550555560
servalserservalleuthr
565
<210>4
<211>331
<212>prt
<213>智人
<400>4
metglythrproglnlysaspvalileilelysseraspalaproasp
151015
thrleuleuleuglulyshisalaasptyrilealasertyrglyser
202530
lyslysaspasptyrglutyrcysmetserglutyrleuargmetser
354045
glyiletyrtrpglyleuthrvalmetaspleumetglyglnleuhis
505560
argmetasnargglugluileleualapheilelyssercysglnhis
65707580
glucysglyglyileseralaserileglyhisaspprohisleuleu
859095
tyrthrleuseralavalglnileleuthrleutyraspserileasn
100105110
valileaspvalasnlysvalvalglutyrvallysglyleuglnlys
115120125
gluaspglyserphealaglyaspiletrpglygluileaspthrarg
130135140
pheserphecysalavalalathrleualaleuleuglylysleuasp
145150155160
alaileasnvalglulysalailegluphevalleusercysmetasn
165170175
pheaspglyglypheglycysargproglysergluserhisalagly
180185190
glniletyrcyscysthrglypheleualailethrserglnleuhis
195200205
glnvalasnseraspleuleuglytrptrpleucysgluargglnleu
210215220
proserglyglyleuasnglyargproglulysleuproaspvalcys
225230235240
tyrsertrptrpvalleualaserleulysileileglyargleuhis
245250255
trpileaspargglulysleuargasnpheileleualacysglnasp
260265270
glugluthrglyglyphealaaspargproglyaspmetvalasppro
275280285
phehisthrleupheglyilealaglyleuserleuleuglygluglu
290295300
glnilelysprovalasnprovalphecysmetproglugluvalleu
305310315320
glnargvalasnvalglnprogluleuvalser
325330
<210>5
<211>653
<212>prt
<213>智人
<400>5
metalaaspthrleuproserglupheaspvalilevalileglythr
151015
glyleuprogluserileilealaalaalacysserargserglyarg
202530
argvalleuhisvalaspserargsertyrtyrglyglyasntrpala
354045
serpheserpheserglyleuleusertrpleulysglutyrglnglu
505560
asnseraspilevalseraspserprovaltrpglnaspglnileleu
65707580
gluasngluglualailealaleuserarglysasplysthrilegln
859095
hisvalgluvalphecystyralaserglnaspleuhisgluaspval
100105110
gluglualaglyalaleuglnlysasnhisalaleuvalthrserala
115120125
asnserthrglualaalaaspseralapheleuprothrgluaspglu
130135140
serleuserthrmetsercysglumetleuthrgluglnthrproser
145150155160
seraspprogluasnalaleugluvalasnglyalagluvalthrgly
165170175
glulysgluasnhiscysaspasplysthrcysvalproserthrser
180185190
alagluaspmetsergluasnvalproilealagluaspthrthrglu
195200205
glnprolyslysasnargilethrtyrserglnileilelysglugly
210215220
argargpheasnileaspleuvalserlysleuleutyrserarggly
225230235240
leuleuileaspleuleuilelysserasnvalserargtyralaglu
245250255
phelysasnilethrargileleualaphearggluglyargvalglu
260265270
glnvalprocysserargalaaspvalpheasnserlysglnleuthr
275280285
metvalglulysargmetleumetlyspheleuthrphecysmetglu
290295300
tyrglulystyrproaspglutyrlysglytyrglugluilethrphe
305310315320
tyrglutyrleulysthrglnlysleuthrproasnleuglntyrile
325330335
valmethisserilealametthrsergluthralaserserthrile
340345350
aspglyleulysalathrlysasnpheleuhiscysleuglyargtyr
355360365
glyasnthrpropheleupheproleutyrglyglnglygluleupro
370375380
glncysphecysargmetcysalavalpheglyglyiletyrcysleu
385390395400
arghisservalglncysleuvalvalasplysgluserarglyscys
405410415
lysalaileileaspglnpheglyglnargileilesergluhisphe
420425430
leuvalgluaspsertyrpheprogluasnmetcysserargvalgln
435440445
tyrargglnileserargalavalleuilethraspargservalleu
450455460
lysthraspseraspglnglnileserileleuthrvalproalaglu
465470475480
gluproglythrphealavalargvalilegluleucysserserthr
485490495
metthrcysmetlysglythrtyrleuvalhisleuthrcysthrser
500505510
serlysthralaarggluaspleugluservalvalglnlysleuphe
515520525
valprotyrthrglumetgluilegluasngluglnvalglulyspro
530535540
argileleutrpalaleutyrpheasnmetargaspserseraspile
545550555560
serargsercystyrasnaspleuproserasnvaltyrvalcysser
565570575
glyproaspcysglyleuglyasnaspasnalavallysglnalaglu
580585590
thrleupheglngluilecysproasngluaspphecyspropropro
595600605
proasnprogluaspileileleuaspglyaspserleuglnproglu
610615620
alasergluserseralaileproglualaasnsergluthrphelys
625630635640
gluserthrasnleuglyasnleuglugluserserglu
645650
<210>6
<211>1962
<212>dna
<213>智人
<400>6
atggcggatactctcccttcggagtttgatgtgatcgtaatagggacgggtttgcctgaa60
tccatcattgcagctgcatgttcaagaagtggccggagagttctgcatgttgattcaaga120
agctactatggaggaaactgggccagttttagcttttcaggactattgtcctggctaaag180
gaataccaggaaaacagtgacattgtaagtgacagtccagtgtggcaagaccagatcctt240
gaaaatgaagaagccattgctcttagcaggaaggacaaaactattcaacatgtggaagta300
ttttgttatgccagtcaggatttgcatgaagatgtcgaagaagctggtgcactgcagaaa360
aatcatgctcttgtgacatctgcaaactccacagaagctgcagattctgccttcctgcct420
acggaggatgagtcattaagcactatgagctgtgaaatgctcacagaacaaactccaagc480
agcgatccagagaatgcgctagaagtaaatggtgctgaagtgacaggggaaaaagaaaac540
cattgtgatgataaaacttgtgtgccatcaacttcagcagaagacatgagtgaaaatgtg600
cctatagcagaagataccacagagcaaccaaagaaaaacagaattacttactcacaaatt660
attaaagaaggcaggagatttaatattgatttagtatcaaagctgctgtattctcgagga720
ttactaattgatcttctaatcaaatctaatgttagtcgatatgcagagtttaaaaatatt780
accaggattcttgcatttcgagaaggacgagtggaacaggttccgtgttccagagcagat840
gtctttaatagcaaacaacttactatggtagaaaagcgaatgctaatgaaatttcttaca900
ttttgtatggaatatgagaaatatcctgatgaatataaaggatatgaagagatcacattt960
tatgaatatttaaagactcaaaaattaacccccaacctccaatatattgtcatgcattca1020
attgcaatgacatcagagacagccagcagcaccatagatggtctcaaagctaccaaaaac1080
tttcttcactgtcttgggcggtatggcaacactccatttttgtttcctttatatggccaa1140
ggagaactcccccagtgtttctgcaggatgtgtgctgtgtttggtggaatttattgtctt1200
cgccattcagtacagtgccttgtagtggacaaagaatccagaaaatgtaaagcaattata1260
gatcagtttggtcagagaataatctctgagcatttcctcgtggaggacagttactttcct1320
gagaacatgtgctcacgtgtgcaatacaggcagatctccagggcagtgctgattacagat1380
agatctgtcctaaaaacagattcagatcaacagatttccattttgacagtgccagcagag1440
gaaccaggaacttttgctgttcgggtcattgagttatgttcttcaacgatgacatgcatg1500
aaaggcacctatttggttcatttgacttgcacatcttctaaaacagcaagagaagattta1560
gaatcagttgtgcagaaattgtttgttccatatactgaaatggagatagaaaatgaacaa1620
gtagaaaagccaagaattctgtgggctctttacttcaatatgagagattcgtcagacatc1680
agcaggagctgttataatgatttaccatccaacgtttatgtctgctctggcccagattgt1740
ggtttaggaaatgataatgcagtcaaacaggctgaaacacttttccaggaaatctgcccc1800
aatgaagatttctgtccccctccaccaaatcctgaagacattatccttgatggagacagt1860
ttacagccagaggcttcagaatccagtgccataccagaggctaactcggagactttcaag1920
gaaagcacaaaccttggaaacctagaggagtcctctgaataa1962
<210>7
<211>1992
<212>dna
<213>智人
<400>7
gatatcgaattcctgcagcccggcggcaccatggcggatactctcccttcggagtttgat60
gtgatcgtaatagggacgggtttgcctgaatccatcattgcagctgcatgttcaagaagt120
ggccggagagttctgcatgttgattcaagaagctactatggaggaaactgggccagtttt180
agcttttcaggactattgtcctggctaaaggaataccaggaaaacagtgacattgtaagt240
gacagtccagtgtggcaagaccagatccttgaaaatgaagaagccattgctcttagcagg300
aaggacaaaactattcaacatgtggaagtattttgttatgccagtcaggatttgcatgaa360
gatgtcgaagaagctggtgcactgcagaaaaatcatgctcttgtgacatctgcaaactcc420
acagaagctgcagattctgccttcctgcctacggaggatgagtcattaagcactatgagc480
tgtgaaatgctcacagaacaaactccaagcagcgatccagagaatgcgctagaagtaaat540
ggtgctgaagtgacaggggaaaaagaaaaccattgtgatgataaaacttgtgtgccatca600
acttcagcagaagacatgagtgaaaatgtgcctatagcagaagataccacagagcaacca660
aagaaaaacagaattacttactcacaaattattaaagaaggcaggagatttaatattgat720
ttagtatcaaagctgctgtattctcgaggattactaattgatcttctaatcaaatctaat780
gttagtcgatatgcagagtttaaaaatattaccaggattcttgcatttcgagaaggacga840
gtggaacaggttccgtgttccagagcagatgtctttaatagcaaacaacttactatggta900
gaaaagcgaatgctaatgaaatttcttacattttgtatggaatatgagaaatatcctgat960
gaatataaaggatatgaagagatcacattttatgaatatttaaagactcaaaaattaacc1020
cccaacctccaatatattgtcatgcattcaattgcaatgacatcagagacagccagcagc1080
accatagatggtctcaaagctaccaaaaactttcttcactgtcttgggcggtatggcaac1140
actccatttttgtttcctttatatggccaaggagaactcccccagtgtttctgcaggatg1200
tgtgctgtgtttggtggaatttattgtcttcgccattcagtacagtgccttgtagtggac1260
aaagaatccagaaaatgtaaagcaattatagatcagtttggtcagagaataatctctgag1320
catttcctcgtggaggacagttactttcctgagaacatgtgctcacgtgtgcaatacagg1380
cagatctccagggcagtgctgattacagatagatctgtcctaaaaacagattcagatcaa1440
cagatttccattttgacagtgccagcagaggaaccaggaacttttgctgttcgggtcatt1500
gagttatgttcttcaacgatgacatgcatgaaaggcacctatttggttcatttgacttgc1560
acatcttctaaaacagcaagagaagatttagaatcagttgtgcagaaattgtttgttcca1620
tatactgaaatggagatagaaaatgaacaagtagaaaagccaagaattctgtgggctctt1680
tacttcaatatgagagattcgtcagacatcagcaggagctgttataatgatttaccatcc1740
aacgtttatgtctgctctggcccagattgtggtttaggaaatgataatgcagtcaaacag1800
gctgaaacacttttccaggaaatctgccccaatgaagatttctgtccccctccaccaaat1860
cctgaagacattatccttgatggagacagtttacagccagaggcttcagaatccagtgcc1920
ataccagaggctaactcggagactttcaaggaaagcacaaaccttggaaacctagaggag1980
tcctctgaataa1992
<210>8
<211>567
<212>prt
<213>褐家鼠
<400>8
methisglyargleulysvallysthrserglugluglnalagluala
151015
lysargleugluarggluglnlysleulysleutyrglnseralathr
202530
glnalavalpheglnlysargglnalaglygluleuaspgluserval
354045
leugluleuthrserglnileleuglyalaasnproaspphealathr
505560
leutrpasncysargarggluvalleuglnhisleugluthrglulys
65707580
serproglugluseralaalaleuvallysalagluleuglypheleu
859095
glusercysleuargvalasnprolyssertyrglythrtrphishis
100105110
argcystrpleuleuserargleuprogluproasntrpalaargglu
115120125
leugluleucysalaargpheleuglualaaspgluargasnphehis
130135140
cystrpasptyrargargphevalalaalaglnalaalavalalapro
145150155160
alaglugluleualaphethraspserleuilethrargasnpheser
165170175
asntyrsersertrphistyrargsercysleuleuproglnleuhis
180185190
proglnproaspserglyproglnglyargleuprogluasnvalleu
195200205
leulysgluleugluleuvalglnasnalaphephethraspproasn
210215220
aspglnseralatrpphetyrhisargtrpleuleuglyargalaglu
225230235240
prohisaspvalleucyscysvalhisvalserarggluglualacys
245250255
leuservalcyspheserargproleuthrvalglyserargmetgly
260265270
thrleuleuleumetvalaspglualaproleuservalglutrparg
275280285
thrproaspglyargasnargproserhisvaltrpleucysaspleu
290295300
proalaalaserleuasnaspglnleuproglnhisthrpheargval
305310315320
iletrpthrglyseraspserglnlysglucysvalleuleulysasp
325330335
argproglucystrpcysargaspseralathraspgluglnleuphe
340345350
argcysgluleuservalglulysserthrvalleuglnsergluleu
355360365
glusercyslysgluleuglngluleugluprogluasnlystrpcys
370375380
leuleuthrileileleuleumetargalaleuaspproleuleutyr
385390395400
glulysgluthrleuglntyrpheserthrleulysalavalasppro
405410415
metargalaalatyrleuaspaspleuargserlyspheleuleuglu
420425430
asnservalleulysmetglutyralaaspvalargvalleuhisleu
435440445
alahislysaspleuthrvalleucyshisleugluglnleuleuleu
450455460
valthrhisleuaspleuserhisasnargleuargalaleupropro
465470475480
alaleualaalaleuargcysleugluvalleuglnalaseraspasn
485490495
alaleugluasnvalaspglyvalalaasnleuproargleuglnglu
500505510
leuleuleucysasnasnargleuglnglnseralaalaileglnpro
515520525
leuvalsercysproargleuvalleuleuasnleuglnglyasnser
530535540
leucysglnglugluglyileglngluargleualaglumetleupro
545550555560
servalserserileleuthr
565
<210>9
<211>331
<212>prt
<213>褐家鼠
<400>9
metglythrglnglnlysaspvalthrilelysseraspalaproasp
151015
thrleuleuleuglulyshisalaasptyrilealasertyrglyser
202530
lyslysaspasptyrglutyrcysmetserglutyrleuargmetser
354045
glyvaltyrtrpglyleuthrvalmetaspleumetglyglnleuhis
505560
argmetasnlysglugluileleuvalpheilelyssercysglnhis
65707580
glucysglyglyvalseralaserileglyhisaspprohisleuleu
859095
tyrthrleuseralavalglnileleuthrleutyraspserilehis
100105110
valileasnvalasplysvalvalalatyrvalglnserleuglnlys
115120125
gluaspglyserphealaglyaspiletrpglygluileaspthrarg
130135140
pheserphecysalavalalathrleualaleuleuglylysleuasp
145150155160
alaileasnvalglulysalailegluphevalleusercysmetasn
165170175
pheaspglyglypheglycysargproglysergluserhisalagly
180185190
glniletyrcyscysthrglypheleualailethrserglnleuhis
195200205
glnvalasnseraspleuleuglytrptrpleucysgluargglnleu
210215220
proserglyglyleuasnglyargproglulysleuproaspvalcys
225230235240
tyrsertrptrpvalleualaserleulysileileglyargleuhis
245250255
trpileaspargglulysleuargserpheileleualacysglnasp
260265270
glugluthrglyglyphealaaspargproglyaspmetvalasppro
275280285
phehisthrleupheglyilealaglyleuserleuleuglygluglu
290295300
glnilelysprovalserprovalphecysmetproglugluvalleu
305310315320
glnargvalasnvalglnprogluleuvalser
325330
- 还没有人留言评论。精彩留言会获得点赞!