使用全乙酰基半乳糖调节重组蛋白的蛋白质半乳糖基化概况的方法与流程
本发明涉及用于在细胞培养过程中使用全乙酰基半乳糖(peracetylgalactose)调节由哺乳动物宿主细胞表达的重组蛋白的半乳糖基化概况的方法和组合物。
背景技术:
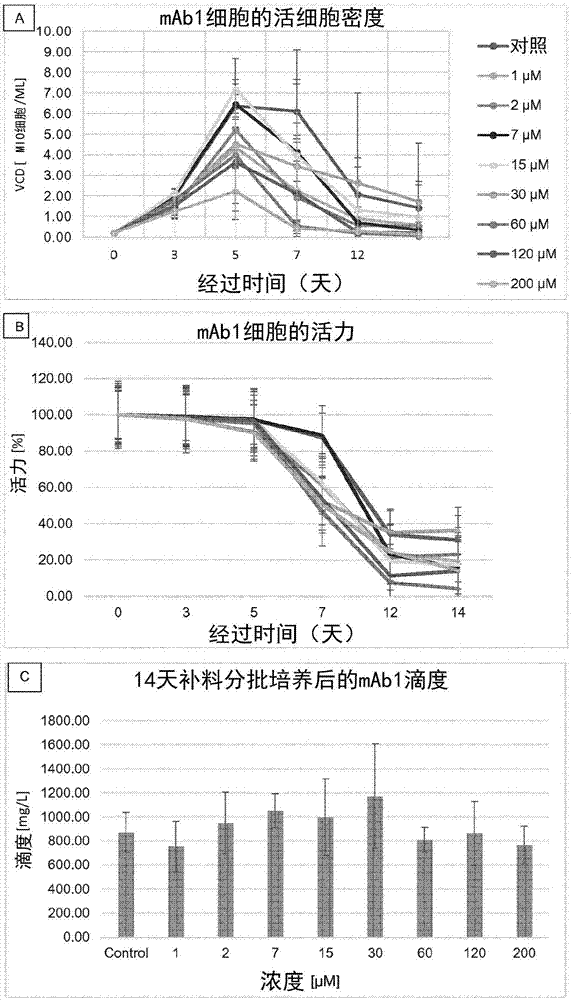
蛋白质如治疗性蛋白质或抗体的糖基化状况是重要的特征,由于其对例如折叠、稳定性和抗体依赖性细胞毒性(adcc,负责抗体治疗效果的机制之一)的影响而导致半衰期和亲和力变化,进而影响蛋白质的生物学活性(eon-duval等,2012)。糖基化高度依赖于用来生产感兴趣蛋白质的细胞系以及细胞培养过程(ph、温度、细胞培养基组成、原料批次间区别、培养基过滤材料、空气等)。adcc活性受与fc区的寡糖连接的岩藻糖和/或甘露糖的量影响,随着岩藻糖减少和/或甘露糖增加可观察到活性增强。事实上,例如,与岩藻糖基化的igg相比,非岩藻糖基化形式表现出显着增强的adcc,这是由于fcγriiia结合能力的增强并且没有任何补体依赖性细胞毒性(cdc)或抗原结合能力的可检测到的变化(yamane-ohnuki和satoh,2009)。类似地,显示高水平甘露糖-5聚糖的抗体也呈现更高的adcc(yu等,2012)。因此,在adcc反应是抗体活性的主要治疗机制的情况下,提供方法来制备糖基化状况以岩藻糖基化降低和/或甘露糖基化升高为特征的重组治疗性蛋白质是有益的。非岩藻糖基化和/或高甘露糖基化抗体的优点还包括在低剂量下达到治疗效果。然而,目前市场上的许多治疗性抗体都是高度岩藻糖基化的,因为它们由哺乳动物细胞系产生,这些细胞系具有负责产物的fcn-聚糖的核心岩藻糖基化的内在酶活性。半乳糖基化:对抗体的活性也有影响。尽管它似乎不影响抗体与抗原的结合,但据报道,半乳糖基化的变化可能导致一些重组igg的cdc活性发生显着变化(hodoniczky等,2005)。实际上,已证明相对于未修饰的(可变半乳糖基化的)对照mab,rituxan的去半乳糖基化使cdc减少约一半。由于糖基化(例如半乳糖基化)可影响治疗效用和安全性,因此蛋白质糖基化的调整对于市售治疗性蛋白质或抗体具有特殊意义。此外,在生物仿制药化合物框架内,重组蛋白糖基化概况的控制是至关重要的,因为所述重组蛋白的糖基化概况必须与参照产物的糖基化概况相当。因此,仍然需要培养条件和生产方法,其允许控制重组蛋白的糖基化概况,例如半乳糖基化概况。本发明通过提供用于调节重组蛋白糖基化,例如重组蛋白半乳糖基化的方法和组合物满足了这种需要。技术实现要素:一方面,本发明提供了一种产生具有经调节的半乳糖基化概况的重组蛋白的方法,所述方法包括在补充有全乙酰基半乳糖的细胞培养基中培养表达所述蛋白的宿主细胞。或者,这里公开了一种产生具有经调节的半乳糖基化概况的重组蛋白的方法,所述方法包括在补充有至少一种包含全乙酰基半乳糖的补料的细胞培养基中培养表达所述蛋白的宿主细胞。在另一方面,本发明提供了一种组合物,其包含含有全乙酰基半乳糖的细胞培养基。另一方面,本发明提供了药物组合物,其包含通过本发明的方法产生的具有经调节的半乳糖基化概况的重组蛋白和药学上可接受的运载体。另一方面,本发明提供了组合物,其包含通过本发明的方法产生的具有经调节的半乳糖基化概况的重组蛋白。在另一方面,本发明提供了全乙酰基半乳糖用于调节重组蛋白的半乳糖基化概况的用途。全乙酰基半乳糖例如是α-2-f全乙酰基半乳糖或β-2-f全乙酰基半乳糖。附图说明图1显示了在微孔板(dwp)中以不同α-2f-pgal浓度培养的mab1细胞的活细胞密度与时间的关系(图1a;)和活力与时间的关系(图1b;)以及第14天的滴度(图1c;)。结果表示为平均值±标准偏差。图1a的图例也适用于图1b。提到的浓度是指刚接种后第0天的浓度。图2显示了在微孔板(dwp)中以不同β-2f-pgal浓度培养的mab1细胞的活细胞密度与时间的关系(图2a;)和活力与时间的关系(图2b;)以及第14天的滴度(图2c;)。结果表示为平均值±标准偏差。图2a的图例也适用于图2b。提到的浓度是指刚接种后第0天的浓度。图3显示了根据β-2f-pgal培养基浓度,与对照相比半乳糖基化概况的绝对变化(图3a);以及与根据β-2f-pgal培养基浓度,与对照相比糖基化概况的绝对变化(图3b)。图4显示了在中以不同α-2f-pgal浓度培养的mab1细胞的活细胞密度与时间的关系(图4a;)和活力与时间的关系(图4b;)以及滴度与经过时间的关系(图4c;)。结果表示为平均值±标准偏差。图4a的图例也适用于图4b。提到的浓度是指刚接种后第0天的浓度。图5显示了在中以不同β-2f-pgal浓度培养的mab1细胞的活细胞密度与时间的关系(图5a;)和活力与时间的关系(图5b;)以及滴度与经过时间的关系(图5c;)。结果表示为平均值±标准偏差。提到的浓度是指刚接种后第0天的浓度。图6显示了根据α-2f-pgal培养基浓度,与对照相比糖基化概况的绝对变化(图6a);以及与根据α-2f-pgal培养基浓度,与对照相比半乳糖基化概况的绝对变化(图6b)。图7显示在摇动管中补充有0-90μmα-2f-全乙酰基-半乳糖或60μmβ-2f-全乙酰基-半乳糖的细胞系a培养物的(a)活细胞密度。(b)活力。(c)培养第5天,第7天,第10天,第12天和第14天每种浓度的蛋白质滴度。每种条件一式两份进行。所有点都是相应条件的平均值,并且误差条报告最大值和最小值。图8显示(a)随着细胞系a培养物中培养基中α-和β-2f-β-半乳糖浓度变化,与对照相比总糖基化模式图(pattern)的绝对变化。(b)随着培养基中α-和β-2f-β-半乳糖浓度变化,与对照相比半乳糖基化的绝对变化。每种条件一式两份进行。所有柱都是通过cge-lif分析的相应条件的平均值,并且误差条报告最大值和最小值。图9显示在摇管中补充有0-90μmα-2f-全乙酰基-半乳糖的细胞系b培养物的(a)活细胞密度。(b)活力。(c)培养第5天,第7天,第10天,第12天和第14天每种浓度的蛋白质滴度。每种条件一式两份进行。所有点都是相应条件的平均值,并且误差条报告最大值和最小值。图10显示(a)随着细胞系b培养物的培养基中α-2f-p-半乳糖浓度变化,与对照相比总糖基化模式图的绝对变化。(b)随着细胞系b培养物的培养基中α-2f-p-半乳糖浓度变化,与对照相比半乳糖基化的绝对变化。每种条件一式两份进行并通过2ab-uplc分析。所有柱都是相应条件的平均值,并且误差条报告最大值和最小值。图11显示摇管中随着β-2f-p-半乳糖的补料时间变化的细胞系a培养物的(a)活细胞密度。(b)活力。(c)培养第5天,第7天,第10天,第12天和第14天每种条件的蛋白质滴度。实验一式两份进行。所有点都是相应条件的平均值,并且误差条报告最大值和最小值。图12显示(a)随着细胞系a培养物中β-2f-p-半乳糖补料时间变化,与对照相比总糖基化模式图的绝对变化。(b)随着细胞系a培养物中β-2f-p-半乳糖补料时间变化,与对照相比半乳糖基化的绝对变化。实验一式两份进行,并通过cge-lif分析上清液。所有柱都是相应条件的平均值,并且误差条报告最大值和最小值。发明详述本文中述及的所有出版物、专利申请、专利和其他参考文献都通过引用全文纳入本文。提供本文讨论的出版物和申请仅针对其在本申请提交日之前的公开内容。本文中所有内容均不应解释为承认本发明不能凭借在先发明而先于这些出版物。此外,材料、方法和实施例都仅是说明性,并不意在构成限制。除非另外定义,否则本文使用的所有技术和科学术语具有与本文主题所属领域的技术人员通常理解的相同的含义。如本文所使用的,提供以下定义以便于理解本发明。在例如措辞“a和/或b”中使用的术语“和/或”旨在包括“a和b”、“a或b”、“a”和“b”。术语“细胞培养”或“培养”是指细胞在体外(即在生物体外或组织外)的生长和繁殖。哺乳动物细胞的合适培养条件是本领域已知的,例如在《用于制药和细胞疗法的细胞培养技术》(cellculturetechnologyforpharmaceuticalandcell-basedtherapies)(2005)中教导的。哺乳动物细胞可以悬浮培养或与固体基质附连。术语“细胞培养基”、“培养基”及其任何复数形式是指可培养任何类型细胞的任何培养基。“基础培养基”是指含有对细胞代谢有用的所有必需成分的细胞培养基。这包括例如氨基酸、脂质、碳源、维生素和矿物盐。dmem(达氏修正依氏培养基),rpmi(洛斯维公园纪念研究所培养基)或培养基f12[翰氏(ham's)f12培养基]是市售基础培养基的示例。或者,所述基础培养基可以是完全内部开发的专有培养基,在此也称为“化学组成明确的培养基”(chemicallydefinedmedium或chemicallydefinedculturemedium),其中所有成分都可以用化学式来描述并且以已知浓度存在。培养基可以不含蛋白质和/或不含血清,并且可以补充有任何其他化合物(例如氨基酸、盐、糖、维生素、激素、生长因子),这取决于培养细胞的需要。术语“补料培养基”或“补料”(及其复数)是指在培养过程中用作补充物以补充所消耗的营养素的培养基。补料培养基可以是市售的补料培养基或专有补料培养基(或者在本文中称为化学组成明确的补料培养基)。术语“生物反应器”或“培养系统”是指可以培养细胞的任何系统,优选以分批或分批补料方式培养。该术语包括但不限于烧瓶、静态烧瓶、旋转瓶、管、摇管、摇瓶、摇摆袋(wavebag)、生物反应器、纤维生物反应器、流化床生物反应器以及带或不带微载体的搅拌罐生物反应器。或者,术语“培养系统”还包括微量滴定板、毛细管或多孔板。可以使用任意尺寸的生物反应器,例如0.1毫升(0.1ml,非常小规模)至20000升(20000l或20kl,大规模),例如0.1ml、0.5ml、1ml、5ml、0.01l、0.1l、1l、2l、5l、10l、50l、100l、500l、1000l(或1kl)、2000l(或2k)、5000l(或5kl)、10000l(或10kl)、15000l(或15kl)或20000l(20kl)。术语“分批补料培养”是指一种生长细胞的方法,其中包括团注(bolus)或连续补充补料培养基以补充消耗的营养素。根据培养基配方、细胞系和其他细胞生长条件,这种细胞培养技术能获得大致超过10x106至30x106个细胞/ml的高细胞密度。可采用各种补料策略和培养基配方来创建和维持双相培养条件。或者可以使用灌注培养。灌注培养中,细胞培养物接受新鲜灌注补料培养基,同时除去废培养基。灌注可以是连续的,分步的,间歇的,或以上若干或全部的任意组合。灌注速率可以为每天小于一工作体积至多个工作体积。优选地,细胞保留在培养物中,且被去除的废培养基基本不含细胞或所含细胞显著少于培养物。灌注可以通过许多细胞截留技术来完成,包括离心、沉淀或过滤(参见例如voisard等,2003)。使用本发明的方法和/或细胞培养技术时,具有经调节的半乳糖基化概况的蛋白质一般直接分泌到培养基中。一旦所述蛋白质分泌到培养基中,就可以使用市售蛋白质浓缩滤器首先浓缩来自这种表达系统的上清液。本文中,“细胞密度”是指给定体积的培养基中细胞的数量。“活细胞密度”是指通过标准活力测定法测定的给定体积培养基中的活细胞数。如果细胞密度与对照培养条件下的细胞密度相比,变化在约-10%至+10%的范围内,则认为细胞密度保持不变。术语“活力”或“细胞活力”是指培养物中活细胞总数与细胞总数之间的比率。只要活力与培养开始时的活力相比不低于60%,则活力通常是可接受的(然而,可接受的阈值可以根据具体情况确定)。活力通常用于确定收获时间。例如,在分批补料培养中,一旦活力达到60%或培养14天后,就可以进行收获。术语“滴度”是指溶液中的物质(此处为感兴趣的蛋白质)的量或浓度。其指示溶液可被稀释且仍含有可检测量的感兴趣分子的稀释倍数。其计算是常规的,例如通过连续稀释(1:2、1:4、1:8、1:16等)含有感兴趣蛋白质的样品,然后使用适当的检测方法(比色法、色谱法等)测定各稀释液是否存在可检测水平的感兴趣蛋白质。还可以采用诸如fortébio或手段来测量滴度,如实施例部分中所示。术语“比生产力”是指每个细胞每天产出的产物(本文为感兴趣蛋白质)的量。如果与对照培养条件的滴度或比生产力相比,滴度或比生产力的变化在约-10%至+10%的范围内,则认为滴度或比生产力保持不变。术语“经调节的糖基化概况”或“经调节的糖基化水平”包括与通过在不补充有全乙酰基半乳糖(如α-或β-2-f-全乙酰基半乳糖)的细胞培养基中培养表达重组蛋白质的重组细胞所产生的相同蛋白质的糖基化概况/水平相比,被调节的该重组蛋白质(例如治疗性蛋白质或抗体)的糖基化概况/水平。经调节的糖基化概况/水平例如是对所述蛋白质中半乳糖基化概况/水平的调节。在一个实施方式中,经调节的糖基化概况/水平,例如经调节的半乳糖基化概况/水平,可以包括蛋白质的半乳糖基化水平的总体降低。本文中,术语“蛋白质”包括肽和多肽,并且指包含两个或更多个氨基酸残基的化合物。本发明所述的蛋白质包括但不限于细胞因子、生长因子、激素、融合蛋白、抗体或其片段。治疗性蛋白质指用于或可以用于治疗的蛋白质。术语“重组蛋白”是指通过重组技术产生的蛋白质。重组技术为本领域技术人员所熟知和掌握(参见例如sambrook等,1989和更新版)。术语“抗体”及其复数形式包括但不限于多克隆抗体、亲和纯化的多克隆抗体、单克隆抗体和抗原结合片段,如f(ab')2、fab蛋白酶水解片段以及单链可变区片段(scfv)。还包括遗传工程改造的完整抗体或片段,例如嵌合抗体、scfv和fab片段,以及合成的抗原结合肽和多肽。术语“人源化”免疫球蛋白指包含人框架区和来自非人(通常是小鼠或大鼠)免疫球蛋白的一个或多个cdr的免疫球蛋白。提供cdr的非人免疫球蛋白称为“供体”且提供框架的人免疫球蛋白称为“受体”(通过将非人cdr移植到人框架和恒定区上,或通过将整个非人可变结构域整合在人恒定区上(嵌合化)来进行人源化)。恒定区无需存在,但如果存在,其必须与人免疫球蛋白恒定区基本相同,即至少约85-90%、优选至少约95%或更高的相同程度。因此,人源化免疫球蛋白的所有部分与天然人免疫球蛋白序列的相应部分基本相同,可能为根据需要调整效应子功能而改变的若干重链恒定区残基和cdr除外。通过将抗体人源化可提高生物半衰期,并降低了给予人后不良免疫反应的可能性。术语“全人”免疫球蛋白指同时包含人框架区和人cdr的免疫球蛋白。恒定区无需存在,但如果存在,其必须与人免疫球蛋白恒定区基本相同,即至少约85-90%、优选至少约95%或更高的相同程度。因此,全人免疫球蛋白的所有部分与天然人免疫球蛋白序列的相应部分基本相同,可能为根据需要调整效应子功能或药代动力学特性而改变的若干重链恒定区残基除外。在一些情况下,可在cdr、框架区或恒定区内引入氨基酸突变以改进抗体的结合亲和力和/或降低抗体的免疫原性和/或改进抗体的生物化学/生物物理学特性。术语“重组抗体”是指通过重组技术产生的抗体。由于抗体生成中涉及重组dna技术,因此无需局限于天然抗体中发现的氨基酸的序列;可以重新设计抗体以获得所需的特性。可能的变化形式有很多,包括仅改变(例如可变结构域或恒定区的)一个或数个氨基酸到完全重新设计(例如可变结构域或恒定区)。通常,进行恒定区的变化是为改进、降低或改变一些特性,这些特性诸如补体固定(例如补体依赖的细胞毒性,cdc)、与fc受体的相互作用,以及其他效应子功能(例如抗体依赖的细胞毒性,adcc)、药代动力学特性(例如与新生儿fc受体(fcrn)结合)。进行可变结构域的变化是为改进抗原结合特性。除抗体外,免疫球蛋白还可以多种其他形式存在,包括例如单链或fv、fab和(fab')2以及双抗体、线性抗体、多价或多特异性杂交抗体。术语“抗体部分”指完整或全长链或抗体的片段,通常是结合区或可变区。所述部分或片段应保留完整链/抗体的至少一种活性,即其是“功能性部分”或“功能性片段”。如果其保留至少一种活性,则优选保留靶标结合特性。抗体部分(或抗体片段)的示例包括但不限于“单链fv”、“单链抗体”、“fv”或“scfv”。这些术语指在单个多肽链内包含来自重链和轻链的可变结构域但缺少恒定区的抗体片段。通常,单链抗体还包含vh和vl结构域之间的多肽接头,其能够令单链抗体形成抗原结合所需的结构。在具体的实施方式中,单链抗体还可以是双特异性和/或人源化的。“fab片段”包含一条重链的可变结构域和ch1结构域和一条轻链。fab分子的重链无法与另一重链分子形成二硫键。含有一条轻链和一条重链且含有更多恒定区(ch1和ch2结构域之间部分)从而在两条重链之间形成链间二硫键的“fab’片段”称作f(ab')2分子。“f(ab')2”含有两条轻链和两条重链,其含有恒定区中ch1和ch2结构域之间的部分,使得两条重链之间形成链间二硫键。在定义了若干重要术语后,现在可以着重讨论本发明的具体实施方式了。可根据本发明生产的已知抗体的示例包括但不限于阿达木单抗、阿仑单抗、贝利单抗、贝伐单抗、卡那单抗(canakinumab)、聚乙二醇化塞妥珠单抗(certolizumabpegol)、西妥昔单抗、地诺单抗、依库珠单抗、戈利木单抗、英夫利昔单抗、那他珠单抗、奥法木单抗、奥马珠单抗、帕妥珠单抗、雷珠单抗、利妥昔单抗、司妥昔单抗、托珠单抗、曲妥珠单抗、优特克单抗(ustekinumab)或维多珠单抗(vedolizomab)。大多数天然存在的蛋白质包含碳水化合物或糖部分,它们与主肽链上选定数目的氨基酸通过特异性连接从而连接至肽。因此,许多天然存在的肽被称为“糖肽”或“糖蛋白”或被称为“糖基化”的蛋白质或肽。在糖蛋白上发现的主要的糖有:岩藻糖、半乳糖、葡萄糖、甘露糖、n-乙酰半乳糖胺(“galnac”)、n-乙酰葡糖胺(“glcnac”)、木糖和唾液酸。连接到肽链上的寡糖结构被称为“聚糖”分子。聚糖的性质影响它们所连接的蛋白质的三维结构和稳定性。天然糖肽中发现的聚糖结构分为两大类:“n-连接聚糖”或“n-连接寡糖”(真核细胞中的主要形式)和“o-连接聚糖”或“o-连接寡糖”。在真核细胞中表达的肽通常包含n-聚糖。n-连接的糖蛋白的糖基团的加工发生在内质网(er)的腔中,并在高尔基体中继续。这些n-连接的糖基化发生在肽一级结构中的天冬酰胺残基上,在含有氨基酸序列天冬酰胺-x-丝氨酸/苏氨酸(x是除脯氨酸和天冬氨酸以外的任何氨基酸残基)的位点上。表1列出了cho细胞分泌的抗体或其片段上可见的主要聚糖:表1——主要聚糖结构(图例:灰色方块:glcnac;中灰色圆:甘露糖,浅灰色圆:半乳糖;灰色三角形:岩藻糖;灰色菱形:唾液酸)“糖型”是指蛋白质例如抗体或其片段的一种同种型,其区别仅在于连接的聚糖的数量和/或类型。通常,包含糖蛋白的组合物包含所述糖蛋白的多种不同糖型。用于测定聚糖一级结构的技术在本领域中是公知的,详细描述可见例如roth等(2012)或song等(2014)。分离细胞产生的蛋白质并确定其n-聚糖的结构是常规的。n-聚糖在含糖分支(也称为“天线(antennae)”)的数目以及所述分支的性质上各不相同,这些分支除了man3glcnac2核心结构之外还可包括例如n-乙酰葡糖胺、半乳糖、n-乙酰半乳糖胺、n-乙酰神经氨酸、岩藻糖和/或唾液酸。关于标准糖生物学命名法的综述,参见《糖生物学基础》(essentialsofglycobioloby),1999。半乳糖基化蛋白质包含至少一个半乳糖残基,并且包括如g1、g1f和/或g2f之类聚糖(参见上表1)。蛋白质上的n-聚糖结构包含至少三个甘露糖残基。这些结构可以进一步被甘露糖基化。甘露糖基化的聚糖如man5、man6或man7被称为高甘露糖聚糖(见上表1)。术语“对象”旨在包括(但不限于)哺乳动物,例如人、狗、牛、马、绵羊、山羊、猫、小鼠、兔或大鼠。更优选地,对象是人。本发明提供了用于调节重组蛋白如治疗性蛋白质或抗体的糖基化概况,例如调节半乳糖基化概况的方法和组合物。本发明基于蛋白质制造的细胞培养条件的优化,例如抗体或抗原结合片段的产生,导致产生具有经调节的糖基化概况,优选具有降低的半乳糖基化的重组蛋白。观察到在补充有半乳糖类似物,更具体地补充有全乙酰基半乳糖(例如氟化全乙酰基半乳糖)的细胞培养条件下,重组蛋白质的半乳糖基化糖型含量降低。因此,在细胞培养生产过程中,当需要调节重组蛋白的糖基化概况,例如正在生产的重组蛋白中的半乳糖基化水平时,细胞培养物可以补充有全乙酰基半乳糖,例如α-或β-2-f-全乙酰基半乳糖或可以用含有全乙酰基半乳糖,例如α-或β-2-f-全乙酰基半乳糖的补料培养基补料。或者,细胞培养基可以已经包含所述全乙酰基半乳糖。还观察到在补充有全乙酰基半乳糖的细胞培养条件下,细胞生长、活力和滴度没有受到影响(即,与没有全乙酰基半乳糖的细胞生长相比,变化维持在+/-10%的范围内;注意到这类没有全乙酰基半乳糖的条件下的细胞生长对应于根据本发明的对照)。2-f-全乙酰半乳糖(具有一个氟原子的全乙酰半乳糖;α/β-d-吡喃半乳糖,2-脱氧-2-氟-,1,3,4,6-四乙酸酯;或者在本文中为α-或β-2f-pgal;)一方面,本发明提供了一种产生具有经调节的糖基化概况(例如经调节的半乳糖基化概况)的重组蛋白的方法,所述方法包括在包含或补充有全乙酰基半乳糖的细胞培养基中培养表达所述蛋白的宿主细胞。优选的全乙酰基半乳糖化合物是α-2-f-全乙酰基半乳糖或β-2-f-全乙酰基半乳糖。或者,本发明描述了一种产生具有经调节的糖基化概况(例如经调节的半乳糖基化概况)的重组蛋白的方法,所述方法包括在补充有包含全乙酰基半乳糖的至少一种补料的细胞培养基中培养表达所述蛋白的宿主细胞。优选的全乙酰基半乳糖化合物是α-2-f-全乙酰基半乳糖或β-2-f-全乙酰基半乳糖。在一个实施方式中,本文提供了全乙酰基半乳糖在细胞培养基或补料培养基中用于调节哺乳动物细胞中产生的重组蛋白的糖基化概况(例如调节半乳糖基化概况)的用途。优选的全乙酰基半乳糖化合物是α-2-f-全乙酰基半乳糖或β-2-f-全乙酰基半乳糖。在另一方面,本发明提供了一种组合物,其包含含有全乙酰基半乳糖的细胞培养基或补料培养基。优选的全乙酰基半乳糖化合物是α-2-f-全乙酰基半乳糖或β-2-f-全乙酰基半乳糖。在另一方面,本发明提供了全乙酰基半乳糖,例如α-2-f-全乙酰基半乳糖或β-2-f-全乙酰基半乳糖用于调节哺乳动物细胞中产生的重组蛋白的糖基化概况(例如调节半乳糖基化概况)的用途。优选地,就本发明上下文整体而言,蛋白质的经调节的糖基化概况包括调节所述蛋白质中的半乳糖基化概况,即,调节半乳糖基化水平。具体地,与对照(即在没有全乙酰基半乳糖的情况下生长的细胞)相比,半乳糖基化水平的调节是重组蛋白中总半乳糖基化水平的降低。更具体地,半乳糖基化水平的降低至少归因于g1f和g2f形式的降低。优选地,与对照相比,总半乳糖基化水平降低约5%至约75%,例如约5%、7.5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%和75%(相对变化)。如果半乳糖残基完全消失或接近于零(即总半乳糖基化水平降低超过75%且最多至100%,则蛋白质将被去半乳糖基化(agalactosylated)。根据定义部分,糖基化水平(例如半乳糖基化水平)的调节或半乳糖基化水平的降低相对于通过在未补充全乙酰基半乳糖的细胞培养基中培养表达所述重组蛋白的重组细胞所产生的相同蛋白质的糖基化水平或半乳糖基化水平来表示。就本发明上下文整体而言,待生产的重组蛋白可以是治疗性蛋白质、抗体或其抗原结合片段,例如人抗体或其抗原结合部分、人源化抗体或其抗原结合部分、嵌合抗体或其抗原结合部分。优选地,它是抗体或其抗原结合片段。本发明的方法可用于产生具有降低的半乳糖残基量或水平的蛋白质,例如抗体。实际上可能需要调节抗体的半乳糖基水平以达到或维持例如某一cdc和/或adcc水平。在本发明的整体上下文中,优选将全乙酰基半乳糖化合物,例如α-2-f-全乙酰基半乳糖或β-2-f-全乙酰基半乳糖以0.1至200μm或约0.1至200μm,优选1至120μm或约1至120μm,甚至优选20至100μm或约20至100μm的接种前(即在接种之前)的浓度,例如以以下浓度或约以下浓度:20、25、30、35、40、45、50、60、65、70、80、90或100μm浓度添加到细胞培养基中。例如,通过调整全乙酰基半乳糖化合物的浓度,可以调节半乳糖基化概况。或者,如果在接种后将全乙酰基半乳糖化合物加入细胞培养基(作为补充物或作为补料),则必须考虑与接种本身相关的稀释因子。通过了解接种前的培养基体积和补充时的细胞培养物的体积(或接种前添加到培养基中的接种体积),容易获取稀释因子。所述稀释因子通常为10至15%。然而,在扩增阶段细胞生长较少的情况下,它可以高达20%或甚至更高。例如,如果在接种前目标等于60μm且稀释因子为10%或20%,则必须分别以54μm或48μm的最终浓度(一旦其在接种培养物中)加入全乙酰基半乳糖化合物。通常,并且考虑到稀释因子在10%和20%之间,如果在接种后将全乙酰基半乳糖化合物加入细胞培养基(作为补充物或作为补料),其在培养基中的最终浓度将是0.08和180μm或约0.08和180μm,甚至优选是0.4至108μm或约0.4至108μm。该最终浓度必须被理解为给定补充物/补料的最终浓度。实际上,并且作为实例,技术人员将理解,如果在培养期间以2次补料加入全乙酰基半乳糖化合物,则总终浓度将对应于叠加到第一次补充物/补料的剩余最终浓度的第二次补充物/补料的最终浓度。就本发明而言,细胞培养基是适合动物细胞如哺乳动物细胞在体外细胞培养中生长的培养基。细胞培养基配方在本领域中是众所周知的。根据培养细胞的需要,细胞培养基可补充额外的组分,如糖、维生素、激素和生长因子。优选地,细胞培养基不含动物性组分;它们可以不含血清和/或不含蛋白质。在本发明的某些实施方式中,向细胞培养基补充全乙酰基半乳糖,例如在培养开始时,并且/或者以分批补料或连续的方式补充。可以基于测得的中间糖基化概况(例如中间半乳糖基化概况/水平)添加全乙酰基半乳糖补充物。培养期间的所述添加可以通过仅由全乙酰基半乳糖化合物组成的补料或通过除包含除其他组分外还包含全乙酰基半乳糖化合物的补充物的补料来完成。在本发明的一个实施方式中,宿主细胞优选哺乳动物宿主细胞(本文也称为哺乳动物细胞),包括但不限于hela、cos、3t3、骨髓瘤细胞系(例如ns0、sp2/0)和中国仓鼠卵巢(cho)细胞。在优选的实施方式中,宿主细胞是中国仓鼠卵巢(cho)细胞,例如cho-s细胞和cho-k1细胞。就本发明上下文整体而言,重组细胞(优选哺乳动物细胞)在培养系统例如生物反应器中生长。生物反应器接种有在包含或补充有全乙酰基半乳糖的培养基中的活细胞。培养基优选无血清和/或无蛋白质。一旦接种到生产性生物反应器中,重组细胞经历指数生长阶段。可以采用分批补料方法团注补以任选性补充有所述全乙酰基半乳糖的补料培养基或由全乙酰基半乳糖组成的补料来维持生长期。补料培养基优选无血清和/或无蛋白质。这些补充性团注补料通常开始于细胞接种到生物反应器后不久,在预计或确定细胞培养物需要补料的时候进行。例如,补充性补料可以从培养开始后的第3天、第4天或第5天或约第3天、第4天或第5天,或者更早或更晚一天或两天开始。在生长和生产阶段,培养物可接受两次,三次或更多的团注补料。这些团注补料中的任何一次可任选地补充有全乙酰基半乳糖。用全乙酰基半乳糖进行的补料或补充可以以分批补料和/或连续的方式在培养开始时进行。或者,可以仅在培养开始后进行全乙酰基半乳糖的补充:在这种情况下,在培养开始时(例如在接种时),不会在培养基中加入全乙酰基半乳糖。当全乙酰基半乳糖(如α-2-f-全乙酰基半乳糖或β-2-f-全乙酰基半乳糖)作为补料加入时,可单独补充(作为单组分补料)或与通常的补充补料一起补充(作为另一类补料的一部分)。所述全乙酰基半乳糖的补料可以从培养开始后的第3天、第4天或第5天或者约第3天、第4天或第5天,或者更早或更晚一天或两天开始。在生长和生产阶段,培养物可接受两次,三次或更多的团注补料。例如,但不应被视为限制性实例,1)可在第3天加入第一次全乙酰基半乳糖补料,然后在第5天、第7天和第10天加入额外全乙酰基半乳糖补料,或2)可在第5天第一次加入全乙酰基半乳糖补料,然后在第7天和第10天加入额外全乙酰基半乳糖补料。本发明的方法、组合物和用途可用于改善多步培养过程中重组蛋白的生产。在多阶段过程中,细胞培养包括两个或更多不同阶段。例如,细胞首先在一个或多个生长阶段中培养(该生长阶段处于改善细胞增殖和活力的条件下),然后转入生产阶段(该生产阶段处于改善蛋白质生产的条件下)。在多步培养过程中,某些条件在各个步骤(或各个阶段)之间可有所不同:培养基组成、ph变化、温度变化等。生长阶段可以在高于生产阶段的温度下进行。例如,生长阶段可以在约35℃至约38℃的第一温度下进行,然后为进入生产阶段而将温度变为约29℃至约37℃的第二温度。收获前,细胞培养物可以在生产阶段维持数天或者甚至数周。用于本发明的细胞系(也称为“重组细胞”)经遗传工程改造而表达具有商业或科研价值的蛋白质。用于表达感兴趣多肽的细胞和/或细胞系的基因工程方法和载体是本领域技术人员熟知的;例如,ausubel等(1988年和更新版)或sambrook等(1989年和更新版)阐述了各种技术。本发明的方法可用于培养表达感兴趣重组蛋白的细胞。通常令重组蛋白分泌到可从中回收重组蛋白的培养基中。然后可以使用可从供应商获得的产品和已知方法将回收的蛋白质纯化或者部分纯化。然后可以将纯化的蛋白质配制成药物组合物。用于药物组合物的合适制剂包括《雷明顿药物科学》(remington'spharmaceuticalsciences,1995)中描述的那些。另一方面,本发明提供了组合物,其包含通过本发明的方法产生的具有经调节的糖基化概况(例如经调节的半乳糖基化概况)的重组蛋白。包含具有经调节的糖基化概况的重组蛋白,例如半乳糖基化水平或量降低的抗体或其抗原结合片段可用于治疗对包含于组合物中的治疗性蛋白质(例如抗体或其抗原结合片段)适合的对象的任何病症。另一方面,本发明提供了药物组合物,其包含通过本发明的方法产生的具有经调节的糖基化概况(例如,经调节的半乳糖基化概况)的重组蛋白和药学上可接受的运载体。重组蛋白优选为治疗性蛋白,可以是抗体或其抗原结合片段,如人抗体或其抗原结合部分、人源化抗体或其抗原结合部分、嵌合抗体或其抗原结合部分。优选地,它是抗体或其抗原结合片段,并且与在不存在全乙酰基半乳糖的补充物中产生的相同抗体或其抗原结合片段相比具有降低的半乳糖基化水平或量。在某些实施方式中,包含具有经调节的糖基化概况的重组蛋白的本发明药物组合物可以与药学上可接受的运载体一起配制成药物(治疗性)组合物,并且可以通过本领域已知的多种方法施用(参见例如《雷明顿药物科学》(remington'spharmaceuticalsciences,1995)。这样的药物组合物可以包含下述的任何一种:盐、缓冲剂、表面活性剂、增溶剂、多元醇、氨基酸、防腐剂、相容性运载体、任选的其他治疗剂,以及它们的组合。包含具有经调节的糖基化概况的重组蛋白的本发明药物组合物以本领域已知并且可用于治疗用途的形式存在,例如液体制剂或冻干制剂。本领域技术人员能够理解,在特定说明的情形以外对本文所述的发明进行变化和修改是容易的。应该理解的是,本发明包括所有这些符合本发明的精神或必要特征的变化和修改。本发明还包括说明书中以单独或组合方式提到或指示的所有步骤、特征、组合物和化合物,以及所述步骤或特征中任意两种或更多种的任意组合和全部组合。因此,本公开在所有方面均为说明性而非限制性,本发明的范围如所附权利要求书所述,并且包含所有等同含义和范围内的所有改变。通过参考以下实施例将更全面地理解前述公开。然而,这些实施例是实施本发明的方法的示例,无意于限制本发明的范围。实施例材料和方法i.细胞、细胞扩增和细胞生长1)细胞用在cho细胞系中产生的抗体进行第一次测定,该cho细胞系为:表达igg1mab1的cho-s细胞,本文为“细胞mab1”或“mab1细胞”。“mab1”是抗可溶性蛋白的完全人单克隆抗体。其等电点(pi)约为8.20-8.30。用在另一cho细胞系中产生的另一抗体进行另外测定,该另一cho细胞系为:表达igg1mab2的cho-k1细胞,本文为“细胞mab2”或“mab2细胞”。“mab2”是抗细胞膜上受体的人源化单克隆抗体。其等电点(pi)约为9.30。2)细胞扩增在管中在适合细胞扩增的培养基中进行细胞扩增。在扩增至少一周后开始分批补料试验。3)接种按0.3x106个细胞/毫升(ml)接种表达mab1的细胞,而按0.2x106个细胞/毫升(ml)接种表达mab2的细胞。4)分批补料所有试验均以分批补料培养进行。对于实施例1-3:使用无血清的化学组成明确的培养基。它原样使用,或者补充有不同浓度(接种前0-200μm)的α-或β-2-f-全乙酰基半乳糖(本文为α-或β-2f-pgal;biosynth和ax分子公司(biosynth&axmolecules))。图1、2、3和4中所示的浓度是刚好在接种前第0天培养基中α-或β-2f-pgal的浓度。定期向培养基中补入化学组成明确的补料培养基和葡萄糖,以维持所述葡萄糖水平在>0至约8g/l的范围内(补料在第3天,第5天,第7天,第10天和第12天进行)。对于实施例4:除了补充α-或β-2-f-全乙酰基半乳糖外,在类似条件下进行另外的实验。在这种情况下,使用无血清的化学组成明确的培养基,如下表2中提到的,用α-或β-2-f-全乙酰基半乳糖补充。培养在深孔板(dwp)中进行,工作体积为450μl。36.5℃、5%co2、90%湿度下孵育并以320rpm振荡。各分批补料培养持续14天。众所周知,dwp中的实验是人工的,尽管它们允许首次筛选培养条件。为了证实dwp中获得的结果在更高规模下是可重复的,在(本文中也称为摇管或st)中也进行了类似的实验,其工作体积为30ml细胞培养物(与深孔板相同的培养条件)。实验第3天第5天第7天第10天β-2f-pgal-d3√√√√β-2f-pgal-d5-√√√β-2f-pgal-d7--√√表2——实施例4的实验条件ii.分析方法:对于dwp实验,使用guava流式细胞仪测量活细胞密度和活力。使用fortébio测量抗体滴度。对于实验,使用测量活细胞密度和活力。使用测量抗体滴度。通过毛细管凝胶电泳和激光诱导荧光(cge-lif;用于dwp和摇管实验)或通过超高效液相色谱法-2-氨基-苯甲酰胺标记技术(2ab-uplc;用于摇管实验)确立糖基化概况。聚糖组别如下表3所示。表3——鉴定的聚糖主要基团(图例:灰色方块:glcnac;中灰色圆:甘露糖,浅灰色圆:半乳糖;灰色三角形:岩藻糖;)实施例1——在dwp中2-f-全乙酰基半乳糖对mab1抗体的影响培养细胞并如材料和方法部分中所公开的那样分析结果。应注意,根据本发明的对照(或对照条件)对应于在没有全乙酰基半乳糖的情况下生长的细胞。活细胞密度和活力:随经过时间变化的活细胞密度和活力,以及补料分批培养结束时的抗体滴度显示在图1(对于α-2f-pgal)和图2(对于β-2f-pgal)中。在高达30μm的浓度下,α-和β-2f-pgal在培养结束前对细胞生长没有影响(图1a和1b;图2a和2b)。在约60μm和更高的浓度下,这两种化合物对细胞生长具有负面影响。平行地,在高达60μm的浓度下,无法在第10天(数据未显示)或第14天(图1c和2c)区分滴度趋势。如图1c和2c所示,在2f-pgal培养基浓度高达60μm时,蛋白质滴度在第10天和第14天保持高于对照的蛋白质滴度。在60μm时,生长细胞的减少对滴度没有实际影响。然而,在较高浓度下,细胞数量的减少反映在滴度上,与对照相比,滴度降低约1.2倍。对于该抗体,对生长和滴度的影响被认为是可接受的,直到约60μm。糖基化概况:通过cge-lif分析获得的糖基化概况显示在图3中(对于β-2f-pgal,图3a和3b)。获得的数据强调α-和β-2f-pgal都能够调节抗体的糖基化,特别是能够降低半乳糖基化的聚糖,而不影响甘露糖基化的,非岩藻糖基化的和岩藻糖基化的糖型。具体地,对照蛋白质具有10%的半乳糖基化n-寡糖,而通过在含有30、60、120和200μmα-2f-pgal的培养基中进行细胞培养获得的蛋白质分别含有8.3%、6.8%、6.0%和4.8%的半乳糖基化n-寡糖(数据未显示)。对于30、60、120和200μmα-2f-pgal浓度,与对照相比,半乳糖基化n-寡糖的绝对变化分别为1.3%、2.8%、3.5%和4.8%(这些变化分别对应于与对照相比相对减少17%、32%、40%和52%)。对于g1f,与对照相比,半乳糖基化n-寡糖的绝对变化分别为1.1%、2.5%、3.1%和4.4%;对于g2f,30、60、120和200μmα-2f-pgal的绝对变化分别为0.2%、0.3%、0.35%和0.4%。对于β-2f-pgal获得了类似的结果(参见图3a和3b)。对照蛋白质含有10.6%的半乳糖基化n-寡糖,而含有30、60、120和200β-2f-pgal的培养基中产生的蛋白质分别含有8.3%、6.4%、5.1%和3.9%的半乳糖基化n-寡糖(这些变化分别对应于与对照相比减少22%、40%、52%和64%)。对于30、60、120和200μmβ-2f-pgal浓度,与对照相比,半乳糖基化n-寡糖的绝对变化分别为2.2%、4.2%、5.4%和6.7%。对于g1f,与对照相比,半乳糖基化n-寡糖的绝对变化分别为1.1%、2.5%、3.1%和4.4%;对于g2f,30、60、120和200μmβ-2f-pgal的绝对变化分别为0.20%、0.30%、0.33%和0.4%。对于这两种化合物,与对照相比的绝对变化包括对甘露糖基化、非岩藻糖基化和岩藻糖基化寡糖的可能的分析噪音(小于1%)内。因此,这些质量属性不受α-或β-2f-pgal中的任何一个的影响。dwp中mab1的结论:低于30μm的α-和β-2f-pgal培养基浓度不影响细胞密度、细胞活力和蛋白质滴度。然而,低于30μm时,α-或β-2f-pgal对半乳糖基化都没有显著的抑制作用。因此,低于30μm时,α-或β-2f-pgal都不是良好的半乳糖基化抑制剂。当浓度高于30μm时,在半乳糖基化上观察到显著的抑制效果。然而,细胞密度、细胞活力和蛋白质滴度可能受到负面影响,尤其是在浓度高于60μm时。浓度范围在30至60μm之间时,α-和β-2f-pgal对蛋白质滴度均没有影响,但它们影响半乳糖基化,相对降低范围为约20%至40%。因此,考虑到对细胞密度、细胞活力、蛋白质滴度和半乳糖基化抑制的影响,30至60μm之间的浓度范围似乎是最佳浓度范围。实施例1显示α-或β-2f-pgal中的任一种特异性地抑制半乳糖基化,而对甘露糖基化和岩藻糖基化没有任何影响。实施例2-在中2-f-全乙酰基半乳糖对mab1抗体的影响培养细胞并如材料和方法部分中所公开的那样分析结果。实施例2.1——初步评价活细胞密度和活力:随经过时间变化的活细胞密度和活力,以及补料分批培养结束时的抗体滴度显示在图4(对于α-2f-pgal)和图5(对于β-2f-pgal)中。在测试的任何一种浓度,即30、60和90μm,α-2f-pgal在培养结束前对细胞生长或蛋白质滴度没有负面影响:与对照相比,这些值是稳定的。对于β-2f-pgal观察到类似的结果,尽管仅测试了一种浓度(即60μm)。糖基化概况:糖基化概况显示在图6中(对于α-2f-pga,图6a和6b)。获得的数据确认了dwp中获得的结果:α-和β-2f-pgal都能够调节抗体的糖基化,特别是能够降低半乳糖基化的聚糖,而不影响甘露糖基化的、非岩藻糖基化的和岩藻糖基化的糖型。根据图6a和6b,可以看到在α-2f-pgal存在下产生的蛋白质的糖基化概况的绝对变化:这导致半乳糖基化的显著降低。对于半乳糖基化的n-寡糖,对于30、60和90μmα-2f-pgal培养基浓度,与对照相比的绝对变化分别为2.0%、4.61%、4.5%(这些变化分别对应于与对照相比相对减少约16%、37%和36%)。关于g1f,与对照相比,30、60和90μmα-2f-pgal培养基浓度的绝对变化分别为1.9%、4.3%、4.3%。关于g2f,与对照相比,30、60和90μmα-2f-pgal培养基浓度的绝对变化为0.11%、0.27%、0.33%。用β-2f-pgal获得了类似的结果(数据未显示):在60μm的浓度下,半乳糖基化的绝对变化为约4%。对于这两种化合物,与对照相比的绝对变化包括对甘露糖基化、非岩藻糖基化和岩藻糖基化寡糖的可能的分析噪音(小于1%)内,也证实了在dwp中获得的结果。实施例2.2确认评价活细胞密度和活力:图7a显示培养基中30、60和90μmα-2f-p-半乳糖下细胞系1培养物的活细胞密度。对照培养物在第7天达到最高密度,攀升至19.5×106活细胞/ml。α-半乳糖类似物补充在培养的第一部分中没有表现出生长抑制作用。尽管如此,对于浓度为30和90μm的α-半乳糖类似物浓度,从第10天到培养结束时细胞密度略低。在第14天,以11.5×106活细胞/ml的密度收获对照培养物。补充的培养物在8.4至9.9×106活细胞/ml的范围内。使用60μmβ-2f-全乙酰基-半乳糖的实验在第7天达到较低峰值,为12.8×106活细胞/ml。然而,之前的数据点与对照相当。对于所有培养条件,活力在第7天是相当的(图7b)。因此,图7a中观察到的差异可能是由于基于成像的细胞荧光分析仪产生的分析假象。在培养第5天、第7天、第10天、第12天和第14天,含有β-2f-p-半乳糖的培养物上清液中的蛋白质浓度与对照相当(在第14天产生2225mg/l),这一事实进一步证实了这一假设。同样地,-α-半乳糖类似物的产物滴度(图7c)在所有时间点都是相当的,在30、60和90μm下,在对照中分别在培养结束时达到2115、2050和2130、2225mg/l。在0至90μm之间的α-2f-p-半乳糖和60μm的β-2f-p-半乳糖的整个浓度范围内未观察到对细胞培养性能的不利影响。糖基化概况:根据图8b,在摇管中证实了在96-dwp中观察到的半乳糖类似物的半乳糖基化抑制作用。半乳糖基化的总水平在30、60和90μm的α-2f-全乙酰基-半乳糖下分别下降:-2.0%、-4.6%和-4.5%,并且用60μm的β-2f-全乙酰基-半乳糖时的下降:-5.0。单半乳糖基化形式分别在30、60和90μm的α-2f-全乙酰基-半乳糖下改变-1.9%、-4.3%、-4.3%,在β-类似物存在下改变-4.7%。完全半乳糖基化的糖型略微降低:-0.1%至-0.3%。总的来说,两种端基异构体的性能相当。2f-p-半乳糖的使用是值得的,因为它具有高特异性,限制了对其他聚糖种类的影响。中mab1的结论:与96dwp平台相比,无论α-和β-2-f-全乙酰基-半乳糖培养基浓度(测试范围:30至90μm之间)如何,使用旋转管平台可获得更稳定的细胞密度、细胞活力和蛋白质滴度结果。半乳糖基化抑制是显着的,对于分别为30和60μm的α-2f-pgal培养基浓度,绝对变化约为2%和4.5%。因此,考虑到对细胞密度、细胞活力、蛋白质滴度和半乳糖基化抑制的影响,30至60μm的浓度范围似乎是最佳浓度范围,因为90μm的α-2f-pgal培养基浓度的半乳糖基化的绝对变化与60μm的相同。添加β-2f-pgal也观察到相同的情况。实施例2确认α-和β-2f-pgal特异性地抑制半乳糖基化,而对甘露糖基化和岩藻糖基化没有任何影响。实施例3——在中2-f-全乙酰基半乳糖对mab2抗体的影响培养细胞并如材料和方法部分中所公开的那样分析结果。活细胞密度和活力:如图9a所强调的,30和60μm的α-2f-p-半乳糖培养物的总活细胞密度与对照相当,其在第7天达到11.2×106活细胞/ml的最大细胞密度。90μm的水平导致细胞生长略有减少,峰值为10.0×106活细胞/ml。整个补充物浓度范围的活力与对照条件(图9b)相当(第7天约97%的细胞活力)。中间浓度(30和60μm)使生产力保持总体不变(图9c)。例如,在第7天,对照的滴度为约730mg/l,对于30和60μm的α-2f-p-半乳糖,滴度为720mg/l。类似地,在第14天,对照的滴度为约3300mg/l,对于30和60μm的α-2f-p-半乳糖,滴度为约3400-3420mg/l。最高的抑制剂浓度需要略微降低滴度,尽管认为不显着(例如:第7天约670mg/l或第14天3200mg/l)。糖基化概况:图10a显示随培养基中α-2f-全乙酰基-半乳糖浓度变化的绝对聚糖变化。与细胞系1一样,半乳糖类似物减少了半乳糖基化。在30、60和90μm下,减少量分别为:-0.95%、-2.05%和-1.4%(绝对变化)。它的存在对整个岩藻糖基化物质的影响也不可忽略(+0.5%)。放大单个末端半乳糖物质的图10b显示分别在30、60和90μm时,对于fa2g1的-0.85%、-1.85和-1.2%减少,和对于fa2g2丰度的-0.09%、-0.19%和-0.19%的小变化。应该记住,使用2ab-uplc而不是cge-lif来定量细胞系2培养物的聚糖模式图。摇管中mab1和mab2的结论:实施例3证实α-和β-2f-pgal特异性地抑制半乳糖基化,对甘露糖基化和岩藻糖基化没有任何影响,无论抗体(这里,用mab1和mab2显示)和细胞系(这里,用细胞mab1和细胞mab2显示)是何种。实施例4——2-f-全乙酰基半乳糖对mab1抗体的影响-在培养开始后以补料补充活细胞密度和活力:与在接种前补充培养基不同,决定在第3、5或7天开始加入半乳糖类似物(分别为β-2f-pgal-d3、β-2f-pgal-d5和β-2f-pgal-d7;见表2)。细胞系1用于这些实验。图11a显示了β-2f-p-半乳糖如何随着添加时间变化而影响活细胞密度。在第7天达到峰值细胞密度。未观察到对照和补充β-2f-pgal-d3的培养物之间的显着差异。它们分别在20.8和21.2×106活细胞/ml下稳定。第5天的补料降低了最大细胞密度,达到18.6×106活细胞/ml。总的来说,可以假设β-2f-p-半乳糖的存在诱导细胞生长的有限变化直至第7天。在培养的后半部分,细胞密度和活力(图11b)与补料时间相关。第3天的补充最强烈地影响了培养进程。在收获时,细胞密度达到9.8×106活细胞/ml(对照:11.8×106活细胞/ml)。活力下降明显快于对照,降至60%以下。分别在第5天和第7天添加补料导致第14天的78%和90%(对照:93%)的活力。根据图11c,没有明确的滴度趋势出现。早期补充在第7天和第10天引起轻微的滴度增加,但随后在培养的后期阶段降低了生产力。对照产生2870mg/l的滴度,第3天补充(β-2f-pgal-d3)产生2560mg/l的滴度。在第5天添加β-2f-p-半乳糖(β-2f-pgal-d5)有利于抗体表达,产生3215mg/l的滴度。在第7天添加(β-2f-pgal-d7),导致显着降低的蛋白质滴度:2100mg/l。与培养基补充相比,通过补料引入β-2f-p-半乳糖限制了对细胞性能的不利影响。即使在第7天开始补料时,上清液中的最终添加剂浓度也显着高于培养基补充物,其中如前所述导致重要的生长和生产力降低。糖基化概况:使用β-2f-p-半乳糖的补料优化导致对半乳糖基化糖型的重要抑制,如图12a所示。对照中11.5%的分泌抗体是半乳糖基化的。从第3天开始添加补料(β-2f-pgal-d3)导致最强的抑制:总体半乳糖基化降低:-8.5%。在第5天(β-2f-pgal-d5)和第7天(β-2f-pgal-d7)开始补料的条件分别导致-7.0%和-4.6%的降低。像在培养基补充实验中一样(在培养开始时),β-2f-p-半乳糖特异性地靶向半乳糖基化。对其他聚糖种类的影响仍然很小。当在第3、5和7天开始补料时,单半乳糖基化物质下降:-8.0%、-6.7%和-4.4%,而二-半乳糖基化实体分别下降:-0.41、-0.33和-0.19(图12b)。半乳糖基化抑制的幅度与补充补料的开始日期相关,因此与上清液中的--2f-p-半乳糖的水平相关。实施例4的结论:补料优化证明是进一步增强补充效果的优秀策略。此外,补料补充允许增加添加剂的总量,与培养基补充相比,对细胞培养性能产生相当小的不利影响。总结本发明实施例证明α-或β-2f-pgal特异性地抑制半乳糖基化,而对甘露糖基化和岩藻糖基化没有影响。本领域技术人员将从上述实施例的结果中理解,可以使用α-和β-2f-pgal中的任何一种来调节任何抗体和任何蛋白质的半乳糖基化概况,无论何种细胞系用于生产,并且特别是降低了整体半乳糖基化水平。补料优化证明是进一步增强补充效果的优秀策略。细胞培养基中待加入的α-和β-2f-pgal的确切浓度,以及补充的时间(在培养开始时,例如在实施例1至3中所示,或作为后面时间点的补料,如实施例4中所示)必须根据具体情况确定,这取决于技术人员希望每分子获得分子的半乳糖基化概况。基于本发明的教导能够完成所述确定而无需任何创造性技术。本领域技术人员还将理解,即使不打算达到特定的糖基化概况,也可以在任何产生蛋白质如抗体的方法中使用α-或β-2f-pgal。参考文献1)eon-duval等,2012.重组治疗性蛋白质量属性:通过设计开发方法评估对安全性和有效性的影响作为质量的一部分(qualityattributesofrecombinanttherapeuticproteins:anassessmentofimpactonsafetyandefficacyaspartofaqualitybydesigndevelopmentapproach).biotechnol.prog.28(3):608-6222)n.yamane-ohnukietm.satoh,2009.产生具有受控岩藻糖基化的治疗性抗体(productionoftherapeuticantibodieswithcontrolledfucosylation);mabs,1(3):230‐2363)yu等,2012.具有n-连接的甘露糖-5聚糖的抗体的表征和药代动力学特性(characterizationandpharmacokineticpropertiesofantibodieswithn-linkedmannose-5glycans);mabs,4(4):475-487.4)hodoniczkyj,zhengyz,jamesdc.通过体外fcn-聚糖重塑控制重组单克隆抗体效应功能(controlofrecombinantmonoclonalantibodyeffectorfunctionsbyfcn-glycanremodelinginvitro).biotechnolprog.2005;21:1644–52.5)zivroth等,2012.蛋白质糖基化的鉴定和定量(identificationandquantificationofproteinglycosylation);internationaljournalofcarbohydratechemistry,articleid640923.6)tingsong等,2014.用于表征制造的重组单克隆抗体药物中糖基化的深入方法(in-depthmethodforthecharacterizationofglycosylationinmanufacturedrecombinantmonoclonalantibodydrugs);anal.chem.,86(12):5661–56667)voisard等,2003,biotechnol.bioeng.82:751-7658)ausubel等,1988和更新版本,《新编分子生物学实验方法》(currentprotocolsinmolecularbiology),纽约约翰韦利森出版公司.9)sambrook等,1989和更新版本,《分子克隆:实验室手册》(molecularcloning:alaboratorymanual),冷泉港实验室出版社.10)《雷明顿药物科学》(remington'spharmaceuticalsciences),1995,第18版,宾夕法尼亚州伊斯顿的马克出版公司当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1