多表位抗原及其制备方法与在制备防治鹦鹉热衣原体感染的药物中的应用与流程
本发明涉及免疫学技术领域,尤其涉及多表位抗原及其制备方法与在制备防治鹦鹉热衣原体感染的药物中的应用。
背景技术:
鹦鹉热衣原体是一种人畜共患病原体,可导致禽类和鸟类的鹦鹉热、鸟疫,以及人类的非典型肺炎。鹦鹉热衣原体的特征包括临床疾病的爆发过程和非常广泛的宿主范围。人类可通过吸入感染鸟类的尿液、粪便或其他排泄物的污染气溶胶发生感染。在成功传播给人类后,主要表现为呼吸道感染、菌血症,而在感染早期主要引起社区获得性肺炎。如不及时治疗,则可能导致包括脑炎和心肌炎等更加严重的并发症。衣原体感染虽然可以通过抗生素治疗有效控制,但它很容易复发。因此,安全有效的疫苗被认为是预防鹦鹉热衣原体感染和减少衣原体疾病传播最有效的方法。
理想的候选抗原应该包含多个b细胞和t细胞表位,当抗原呈递细胞与mhci类、mhcii类分子结合呈现抗原时,可诱导产生免疫应答。以往的研究表明,鹦鹉热衣原体主要外膜蛋白(momp)是一种高度保守的蛋白,可诱导动物模型产生特异性的抗衣原体免疫应答。此外,鹦鹉热衣原体momp可能是存在较多t细胞和b细胞表位潜在候选抗原。质粒是衣原体中重要的毒力因子,包含8个开放阅读框(orfs),分别编码8个质粒蛋白。重组质粒蛋白pgp3或pgp4免疫可显著降低鼠衣原体载量,减轻感染小鼠肺部炎症反应。在我们之前的研究中,鹦鹉热衣原体的质粒编码cpsit_p8(pgp4)蛋白在balb/c小鼠体内免疫接种三次后,诱导了产生显著的免疫保护作用。cpsit_p6蛋白与cpsit_p8具有相似的功能,由于其序列较长,潜在表位较多,更适合于优势区域的筛选。
目前,鹦鹉热衣原体的疫苗都是以蛋白为抗原的,主要是以鹦鹉热衣原体momp和cpsit_p6蛋白。相较于常规的减毒活疫苗以及亚单位重组蛋白疫苗,多肽疫苗其优势在于:
1.特异性强、免疫靶向性好:针对复杂的非连续性抗原片段,构建相应的合成抗原多肽。其融合了不同的t细胞以及b细胞表位能够产生特异性更强的抗衣原体抗体,更具针对性,并能够直接靶向作用于体内的衣原体;
2.毒副作用小、排异反应少:由于合成过程我们只选取了抗原性较强的细胞表位片段,因此不含蛋白抗原中的高反应原性成分(脂多糖、毒素)。同时还能够去除对免疫个体本身分子具有致敏性和交叉反应活性的抗原片段,进而最大程度地减少了毒副反应以及排异反应的发生;
3.生产技术安全、高效:在多肽片段合成过程中,成熟的合成技术可以将不同抗优势抗原表位有效地融合在一个载体上,同时使片段纯度达到90%以上,远远高于蛋白抗原。而高纯度的融合多肽相较于纯度较低的重组蛋白而言,也能够大大降低非抗原物质对免疫过程影响;
4.结构稳定,不易变性:重组多肽的线性结构不同于蛋白的空间结构,其抗原结构更加稳定,在免疫过程中不会产生新的空间构象而影响免疫效果。
因此,与以蛋白为抗原的疫苗相比,多肽疫苗更适于作为候选的抗衣原体感染疫苗。但是,虽然多肽疫苗存在诸多优势,但优势表位的筛选始终是多肽疫苗开发中的难题之一,由于一个蛋白可存在多个表位,而多肽片段较短,其表位的筛选始终是本领域的研究热点。
技术实现要素:
有鉴于此,本发明要解决的技术问题在于提供多表位抗原及其制备方法与在制备防治鹦鹉热衣原体感染的药物中的应用。
本发明提供了如seqidno:1所示氨基酸序列的多表位抗原。
所述“抗原表位”指抗原上的一种位点,也称做“抗原决定簇”或“抗原决定族位点”,其实质是一种多肽。b细胞和t细胞对该位点可发生特异性应答反应。本发明选择两段来自cpsit_p6蛋白的表位和两段来自momp的表位,通过gggg(gly-gly-gly-gly)连接子进行连接,获得本发明所述的多表位抗原。该多表位抗原片段仅含有52个氨基酸,但其效果与大分子蛋白momp或cpsit_p6为抗原的相当。
本发明将上述抗原表位与载体蛋白偶联,形成融合多肽。所述载体蛋白可为klh(keyholelimpethemocyanin)或bsa。一些实施例中,选用klh蛋白作为载体蛋白。
本发明还提供了一种融合多肽,其包括如seqidno:1所示氨基酸序列和klh蛋白序列。
所述融合多肽中,klh可位于seqidno:1所示氨基酸序列的c端或n端。一些实施例中,klh位于seqidno:1所示氨基酸序列的c端。本发明还提供了编码所述抗原或融合多肽的dna分子。
本发明所述抗原或融合多肽可采用人工合成的方式获得,也可通过基因工程的手段合成。本发明对此不做限定。当其采用基因工程方式合成时,将编码的dna分子通过表达质粒载体转化入生物体内,诱导表达获得所述抗原或融合多肽。
本发明所述多表位抗原或融合多肽在制备治疗性抗体、疫苗和/或检测试剂盒中的应用。
以往的研究表明,igm和igg分别在衣原体感染的早期和晚期发挥保护作用。因此,血清中igg、iga和igm在末次免疫后14天从免疫小鼠中收集,并通过elisa检测这些抗体的水平。研究表明,以本发明提供的多表位抗原免疫组诱导igg、iga和igm的抗体水平明显高于fa-和pbc免疫组(p<0.01)。多表位抗原诱导产生的血清igg亚型(igg1、igg2a、igg2b、igg3抗体)水平也明显高于对照组。
本发明所述治疗性抗体或疫苗的作用包括:介导th1型细胞免疫应答、降低炎症因子水平、降低鹦鹉热衣原体载量、促进cd4+或cd8+细胞的免疫应答。
本发明还提供了一种预防鹦鹉热衣原体感染的疫苗,其包括本发明所述融合多肽。
所述的疫苗的制备采用本领域常规采用的各种方法,例如,将所述融合多肽与适宜的药用载体和/或媒介物配制本发明的疫苗。所述药物载体可采用无菌盐水,也可使用其它水性或非水性等渗无菌注射液以及水性和非水性无菌悬浮液。
另外,本发明的疫苗的配制还可含有其它药学上可接受的辅料,包括:佐剂、稳定剂、ph调节剂或防腐剂等。另外,在制剂中也可包含调节免疫应答的其它成分,例如:il-12、cpg寡脱氧核苷酸(cpg-odn)等。具体的佐剂采用弗氏佐剂。具体的,疫苗的免疫包括三次,初免采用的疫苗采用完全弗氏佐剂进行乳化,而第二次、第三次免疫采用与不完全弗氏佐剂乳化。
本发明还提供了一种预防鹦鹉热衣原体感染的方法,其为给予本发明所述的疫苗。所述治疗的方案中,疫苗可以单剂量或多剂量给予,且可以包括给予加强剂量以引发和/或维持免疫力。
本发明还提供了一种治疗鹦鹉热衣原体感染的药物,其包括由所述融合多肽制得的治疗性抗体。
本发明还提供了包含本发明所述融合多肽的各种组合物,特别是药物组合物或疫苗组合物。该组合物可有效用于治疗和/或防治鹦鹉热衣原体感染引起的相关的病症。所述组合物中,还包括其他治疗剂。所述其他治疗剂选自其他用于治疗或预防鹦鹉热衣原体感染的药物。本发明所述的组合物的剂型包括但不限于:注射剂、粒剂、片剂、丸剂、栓剂、胶囊、悬浮液、喷雾、栓剂、透皮药物(如贴片等)、油膏、洗剂等。
本发明还提供了一种治疗鹦鹉热衣原体感染的方法,其为给予本发明所述的药物。
本发明还提供了一种试剂盒,其包括所述融合多肽,和/或与所述融合多肽相互作用的抗体。
本发明中,所述试剂盒还包含检测与所述融合多肽和/或与融合多肽相互作用的抗体的检测试剂。所述检测试剂应用于本领域熟知的免疫检测方法:例如:酶联免疫分析法、免疫印迹法、免疫荧光或放射免疫分析法的试剂。一些实施例中,所述试剂盒中还包括用于固定所述融合多肽的固相支持物,所述“固相支持物”是指本文所述的多肽分子和/或抗原表位能够附着至其上的任何支持物,包括但不限于硝酸纤维素膜、聚偏二氟乙烯(pvdf)膜、ipdms芯片、微孔板、凝胶等。
本发明的试剂盒用于鉴定样品中是否存在融合多肽和/或融合多肽相互作用的抗体。具体的,可用于鉴定样品是否受到鹦鹉热衣原体的感染,或鉴定样品是否成功免疫获得抗性。
本发明还提供了一种检测所述融合多肽的抗体的方法,包括:以所述的试剂盒通过elisa法进行检测。
本发明提供了如seqidno:1所示氨基酸序列的多表位抗原、包括该抗原的融合多肽以及该融合多肽的用途。该抗原分子量小,制备简单,而免疫原性与大分子蛋白相当。实验表明,本发明提供的抗原能够诱导体液和th1细胞免疫反应产生,并产生具有显著意义的特异性抗体以及γ干扰素(ifn-γ)、白介素2(il-2)。疫苗接种能够显著降低感染小鼠肺组织的衣原体载量以及炎症侵润程度,并显著降低炎性因子ifn-γ和il-6水平。此外,从接种过疫苗的小鼠身上获得的cd4+t细胞,经过继转移后显著降低了感染小鼠肺组织的衣原体载量,表明其在细胞免疫应答中的具有重要意义。
附图说明
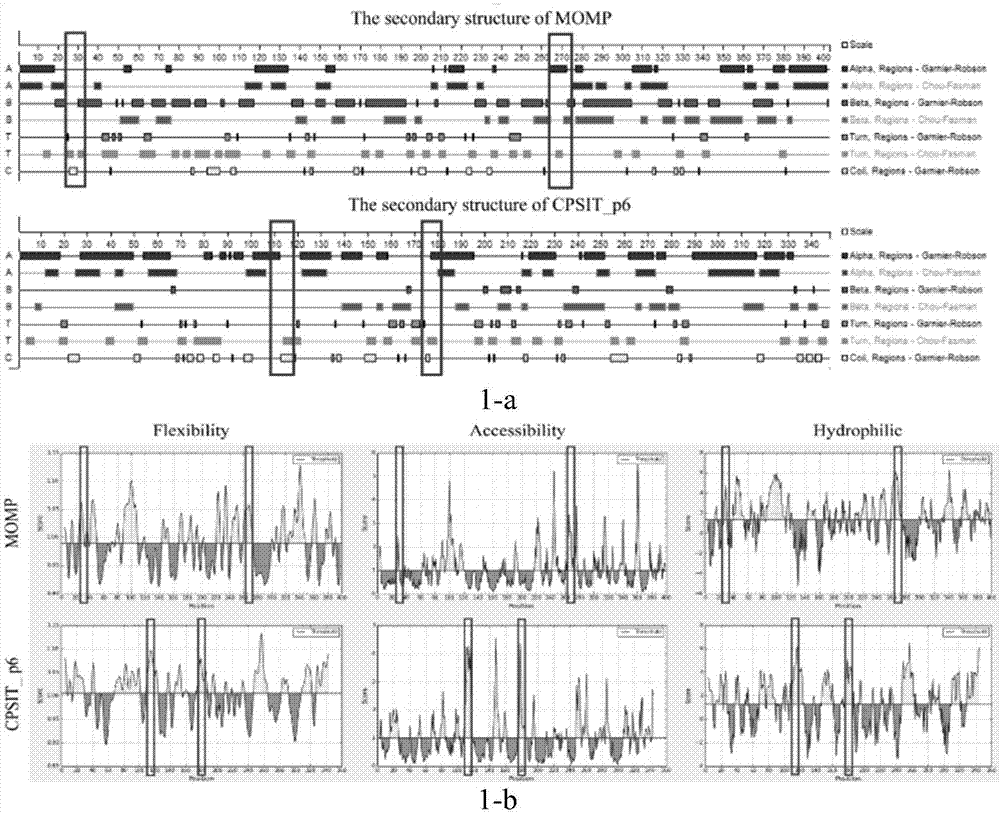
图1示鹦鹉热衣原体momp和cpsit_p6蛋白的生物信息学分析;其中,图1-a示利用protean软件预测momp和cpsit_p6蛋白的二级结构;红色区域:α螺旋;绿色区域:β折叠;蓝色区域:β转角;黄色区域:无规卷曲;图1-b示在iebd网站上预测了蛋白的亲水性、可及性和柔韧性;黄色峰值表示momp和cpsit_p6蛋白片段的亲水性、可及性和柔韧性较强;红色标记表示选定的片段;
图2示血清抗体和igg亚型的检测;balb/c小鼠免疫接种后14天,检测血清ig抗体、igg亚型抗体水平;图2-a示收集免疫小鼠血清,用elisa法测定1:200稀释后的特异性血清抗体(igg、igm、iga);图2-b示elisa法检测免疫小鼠igg亚型水平;图2-c~d示p-m、cpsit_p6和momp免疫小鼠中特异性血清抗体(igg、igm、iga)和igg亚型的抗体滴度;
图3示免疫小鼠脾脏细胞产生细胞因子;其中,图3-a~3-b示th1细胞因子,包括ifn-γ和2,测量使用ready-set-go!试剂盒;图3-c~3-d示elisa试剂盒检测含有il-4、il-10的th2细胞因子;每个柱状图表示6小鼠每组的平均数±标准差,三个独立检测的结果;*p<0.05;**p<0.01;
图4示免疫后免疫肺内衣原体滴度测定;感染后10天将免疫小鼠的肺匀浆接种到hela229细胞单层;然后用间接免疫荧光法鉴定衣原体包涵体数;每个柱状图表示6小鼠每组的平均数±标准差,三个独立检测的结果;**p<0.01;
图5示鹦鹉热衣原体感染后肺组织炎性侵润;其中,图5-a示每组小鼠肺组织的he染色图(放大200×400×);图5-b示感染小鼠的肺切片和免疫组织化学使用超灵敏tmsp(兔子)包含ihc试剂盒,兔抗为一抗,图中a示p-m组;b示cpsit_p6组;c示momp组;d示fa组;e示pbs组;棕色颗粒(黑色箭头)表明衣原体包裹在肺组织细胞的细胞核中;
图6示衣原体感染后肺组织炎性因子;提取衣原体感染后免疫小鼠肺匀浆,elisa检测ifn-γ、il-10、il-6、il-12、il-4的水平;每个柱状图表示每组6只小鼠肺组织细胞因子的平均数±标准差,三个独立检测的结果;*p<0.05;**p<0.01;
图7示脾细胞的流式检测结果;图7-a示cd4+细胞消耗后,采用流式细胞仪检测脾脏细胞;图7-b示cd8+细胞消耗后,采用流式细胞仪检测脾脏细胞;图7-c示采用流式细胞术检测未处理的脾脏细胞;图中为cd8+(r2)、cd4+(r3)和cd4-cd8-(r4)细胞的百分比;用pe抗cd4和fitc抗cd8单克隆抗体染色或用同型对照鸡抗体染色;
图8示脾脏细胞过继转移后肺内衣原体滴度的变化;免疫小鼠的脾脏细胞中cd4+或cd8+细胞被消耗后,转移至新的幼鼠中,免疫小鼠未处理脾脏细胞作为对照;在过继转移2天后,小鼠接感染衣原体,感染7天后通过间接免疫荧光检测肺组织中的衣原体包涵体滴度;每个柱状图表示每组6只小鼠肺组织衣原体滴度的平均数±标准差,三个独立检测的结果;**p<0.01。
具体实施方式
本发明提供了多表位抗原及其制备方法与在制备防治鹦鹉热衣原体感染的药物中的应用,本领域技术人员可以借鉴本文内容,适当改进工艺参数实现。特别需要指出的是,所有类似的替换和改动对本领域技术人员来说是显而易见的,它们都被视为包括在本发明。本发明的方法及应用已经通过较佳实施例进行了描述,相关人员明显能在不脱离本发明内容、精神和范围内对本文的方法和应用进行改动或适当变更与组合,来实现和应用本发明技术。
本发明采用的试材皆为普通市售品,皆可于市场购得。
下面结合实施例,进一步阐述本发明:
实施例
1、材料和方法
1.1多肽合成
对cpsit_p6蛋白和momp的t细胞表位进行鉴定、分析,选择momp24-32、momp262-272、cpsit_p6109-119和cpsit_p6173-181作为抗原表位,之间以gggg(gly-gly-gly-gly)连接子连接,c末端以keyholelimpethemocyanin(klh)偶联作为载体蛋白,将选定的抗原片段融合在一起。
形成的片段序列为kkyktsrnkneggggegltpkenkggggpvgnpaepsggggtteatdtksat-klh
重组抗原p-m通过lifeteinbiotechnology公司(中国,北京)合成,其纯度>90%。
1.2鹦鹉热衣原体培养
人宫颈癌上皮hela229细胞(atccccl-2.1)感染psittaci6bc(atccvr-125)后,于含10%胎牛血清的完全dmem培养基中培养。在35℃和5%的二氧化碳环境下培养36h后,通过梯度密度离心法收集衣原体eb。衣原体eb用spg缓冲液(0.25msucrose,10mmsodiumphosphate,5mmglutamicacid,ph7.5)收集并储存于-80℃。
1.3小鼠以及免疫
6-8w雌性balb/c小鼠购自湖南斯莱克景达动物有限公司。动物是在无菌条件下的温度26℃,并提供与无菌水和食物一天一次。5组小鼠(n=12)分别用重组抗原p-m、纯化后的cpsit_p6、momp、磷酸盐缓冲盐水(pbs)和freund佐剂(fa)腹腔注射免疫(i.p),间隔2周。免疫接种之前,将40μg重组抗原p-m,纯化的cpsit_p6和momp蛋白与0.1毫升的完全弗氏佐剂进行乳化后进行初免。而第二(14d)、第三次(28d)免疫则与不完全弗氏佐剂乳化。pbs组和fa组小鼠分别免疫接种0.1mlpbs和fa。末次免疫2w后(42天)每组处死6只小鼠取脾脏以及血清样本。血清样本收集后于-80℃保存。
1.4血清抗体检测
末次免疫接种两周后从实验组小鼠中收集血清,通过酶联免疫吸附法(elisa)测定抗p-m特异性抗体的水平。以抗momp特异性抗体和抗cpsit_p6特异性抗体为阳性对照。简而言之,96孔板与重组抗原p-m、纯化重组cpsit_p6或momp在碳酸氢钠缓冲液(ph9.5)中孵育过夜。第二天96孔板用含0.05%吐温20的pbs(pbst)清洗,然后用含3%牛血清白蛋白(250μl/)的pbst37℃封闭2h。pbst洗涤后,将100μl1:200稀释的血清或经过倍比稀释的血清添加到每个孔中37℃孵育1h。pbst洗涤后,分别孵育100μl辣根过氧化物酶(连接的羊抗鼠igg,iga,igm,igg1igg2a,igg2b,igg3抗体1h。再次洗板后,加入底物反应液室温孵育15min,随后加入50μl的终止液终止反应,最终抗体效价为高于阴性对照血清的均值+3个标准差的血清读数。
1.5脾脏细胞因子检测
根据之前的研究,每组小鼠在末次免疫接种两周后分别收集6只小鼠的脾脏细胞悬液。脾细胞(6×106/ml)接种到24孔细胞培养板中每孔800μl,随后用重组抗原、纯化的cpsit_p6蛋白质和momp(16μg/)进行刺激,并于37℃5%co2环境中培养48h。通过离心收获(6000rpm,10min)脾细胞上清,并通过ready-set-go!试剂盒检测th1(ifn-γ和il-2)以及th2(il-10andil-4)型细胞因子。
1.6滴鼻感染以及衣原体载量检测
末次免疫2w后,通过乙醚麻醉后滴鼻感染小鼠5×105ifu每只30μl(每组感染6只小鼠)。感染后1,4,7,10天分别观察小鼠体重变化。感染10天后处死小鼠,取肺组织,用spg缓冲液研磨成肺组织匀浆。将肺匀浆上清液连续稀释后接种于hela229细胞单层,并通过间接免疫荧光测定鹦鹉热衣原体滴度。随后通过he染色以及免疫组化对小鼠肺组织炎性侵润以及衣原体感染情况进行检测。
1.7磁珠分选以及过继转移
末次免疫2w后每组取3只小鼠的脾细胞进行过继转移实验。用红细胞裂解液裂解红细胞室温反应15min,并用pbs缓冲液清洗两遍后重悬。磁珠分选通过磁珠包被的cd4-antibody(l3t4)以及cd8a-antibody(ly-2)在磁珠细胞分选器中分别消耗掉脾细胞中的cd4+或者cd8+t细胞。并通过流式细胞术分别检测磁珠分选后脾细胞中cd4+或者cd8+t细胞的比例。分选后的cd4+或者cd8+t细胞的比例均要小于1%后方可进行过继转移实验。约1×106分选后的脾细胞通过尾缘静脉接种至新的balb/c小鼠体内,并将未经处理的细胞作为阳性对照。随后通过滴鼻感染小鼠,并于感染后第7天处死小鼠,取肺组织进行衣原体滴度检测。
1.8统计学分析
采用单因素方差分析(anova)和student-newman-keuls验计算这些组之间抗体、衣原体滴度和细胞因子水平的差异。统计学显著性标准设为p<0.05,所有计算均采用spss18.0软件进行。
2、结果
2.1重组抗原p-m的构建
分析的momp和cpsit_p6蛋白质的二级结构,筛选出4个片段,分别为:cpsit_p6109-119(kkyktsrnkne)、cpsit_p6173-181(egltpkenk)和momp24-32(pvgnpaeps)、momp262-272(tteatdtksat)为含有潜在b细胞表位的区域。我们将选定的抗原片段按照以下方式融合在一起kkyktsrnkneggggegltpkenkggggpvgnpaepsggggtteatdtksat-klh。并用质谱(ms)和高效液相色谱(hplc)验证重组抗原p-m的分子量以及纯度(数据未显示)。因此,基于momp和cpsit_p6的重组抗原p-m是一种包含t细胞和b细胞表位的新型融合抗原。
2.2重组抗原p-m的免疫原性
以往的研究表明,igm和igg分别在衣原体感染的早期和晚期发挥保护作用。因此,血清中igg、iga和igm在末次免疫后14天从免疫小鼠中收集,并通过elisa检测这些抗体的水平。如图2-a所示,重组抗原p-m免疫组诱导igg、iga和igm的抗体水平明显高于fa-和pbc免疫组(p<0.01)。重组抗原p-m诱导产生的血清igg亚型(igg1、igg2a、igg2b、igg3抗体)水平也明显高于对照组(p<0.01)(图2-b)。
为了比较p-m特异性抗体与阳性对照(cpsit_p6-和momp免疫组)的抗体水平,我们接下来通过血清的倍比稀释来确定特异性的抗体效价。从图2-c和2-d可以看出,重组抗原p-m诱导的特异性抗体滴度(包括igg、iga、igm和igg亚型)较抗momp免疫小鼠高,但低于cpsit_p6(尽管其并没有统计学差异)。
如图3-a~3-b所示,重组抗原p-m免疫后th1细胞因子较对照组(fa和pbs组)相比显著增加(p<0.01),但与cpsit_p6和momp组相比无显著差异。而p-m刺激的th2细胞因子(il-4和il-10)水平与其他组比较均无显著差异(图3-c和3-d)。因此,与cpsit_p6和momp蛋白相似,重组抗原p-m介导的主要为th1型细胞免疫应答。
2.3重组抗原p-m的免疫保护作用
免疫小鼠感染后1,4,7,10天分别观察体重变化。p-m组和阳性对照组体重减轻幅度小于阴性对照组,但无显著差异(未见数据)。在感染10天后处死小鼠,以测定肺中鹦鹉热衣原体的含量(图4)。我们观察到p-m免疫组与fa免疫组、pbs免疫组相比肺衣原体载量显著降低,但与cpsit_p6和momp组无显著差异。
接下来,我们通过比较小鼠肺部的炎症病理变化,进一步评估重组抗原p-m的保护作用。p-m组(图5a-a)、cpsit_p6(图5a-b)和momp(图5a-c)的感染小鼠肺组织炎症浸润要明显低于fa(图5a-d)和pbs(图5a-e)组,且实验组和阳性对照组的肺泡结构比阴性对照组更稳定。免疫组化检测肺组织感染情况显示,与fa-(图5b-d)和pbs-(图5b-e)免疫小鼠相比,p-m免疫组小鼠肺组织接近正常,肺组织炎症细胞较少(图5b-a)。此外,接种p-m的小鼠与阴性对照组相比,能够有效降低肺中的衣原体载量(棕色颗粒)。
诱导保护性免疫并有效抑制感染过程中的病理变化是潜在候选疫苗的必要条件。如图6所示,虽然p-m免疫小鼠与阴性对照组的il-10、il-12、il-4水平无显著差异。但是,重组抗原p-m却能够显著降低肺组织中ifn-γ(p<0.001)和il–6的水平(p<0.01)。
2.4免疫小鼠的脾细胞过继转移
为了确定重组抗原p-m特异性细胞免疫应答的保护作用,在末次免疫后14天,从p-m免疫组提取脾脏细胞进行过继转移。采用磁珠分选法对脾脏细胞进行cd4+,cd8+细胞的消耗,并采用流式细胞术进行收集验证。如图7所示,磁珠分选后脾脏细胞中cd4+(r3)和cd8+(r2)细胞均少于1%,而未处理脾脏细胞中分别为26%和21%。
消耗的脾脏细胞过继转移至新的幼鼠体内48小时后,小鼠感染鹦鹉热衣原体并于第7天实施安乐死。过继转移了消耗p-m特异性cd8+t的小鼠,其肺组织衣原体载量要显著低于pbs与fa组(p<0.01)(图8)。而继转移了消耗p-m特异性cd4+t的小鼠,其肺组织衣原体载量较对照组均无显著差异(图8)。因此,cd4+细胞在重组抗原p-m诱导的细胞免疫应答中可能发挥了重要作用。
3讨论
设计鹦鹉衣原体疫苗被认为是预防和控制这种呼吸道传播感染的最佳途径。早期衣原体的疫苗经历了活的、灭活的以及减毒的疫苗,其产生不良影响较大且存在较大的病理相关问题。虽然蛋白抗原已被证明是非常有效的,但由于免疫原的复杂性、有害的副作用以及需要有效的传递系统来维持免疫反应,使其不适合作为候选疫苗。近年来,多肽疫苗作为抑制传染病和癌症的潜在候选疫苗获得了较大成功。与灭活菌、亚单位免疫原和重组疫苗的特点相比,肽疫苗的优势在于生产工艺安全、免疫靶向性好、毒副作用少。此外,排除了高反应原性成分(脂多糖、毒素),有助于清除对免疫个体具有高致敏性的活性片段。因此,肽疫苗在衣原体疫苗开发项目中发挥着重要作用。
重组蛋白因其免疫原性而被广泛研究作为开发衣原体疫苗以控制感染或疾病相关病理的候选抗原。特别是近年来,融合抗原包括t细胞或b细胞表位丰富的肽库已被用作免疫原。然而,这些研究仅仅是从一种蛋白中筛选出抗原表位进行融合。因此,根据生物信息学分析,本研究从不同的蛋白免疫原中选取多个优势表位片段设计疫苗。由于重组抗原中含有多个t细胞和b细胞表位,我们可以评估肽疫苗对小鼠模型的免疫反应和保护作用。
此前已有研究表明,特异的igg可以更好地保护小鼠免受衣原体呼吸道感染。血清中产生的iga对消除原发性呼吸道感染无影响,但能有效增加体外的衣原体中和。在本研究中,我们发现重组多表位抗原与阴性对照相比,在小鼠血清中特异性igg、iga和igm水平显著高,这与之前的研究一致,即重组多表位抗原可以产生体液免疫反应,防止微生物附着。有趣的是,重组抗原p-m诱导的特异性抗体滴度比momp免疫组小鼠高,但低于cpsit_p6蛋白组。通过生物信息学分析,cpsit_p6中b细胞表位较多,因此p-m包含了其表位比momp能够产生更强的体液应答。此外,p-m中表位的较少似乎解释了其与cpsit_p6相比免疫原性更低。因此,在未来的研究中,确定最佳的融合策略能够诱发更强的免疫应答是一个至关重要的问题。
重组多表位抗原的免疫可诱导抗衣原体感染的体液免疫和细胞免疫应答。如图4、5、6所示,我们的研究结果表明接种疫苗可以显著降低感染小鼠肺部衣原体载量和病理改变,这与之前的研究一致。同样,我们也发现p-m-、momp-和cpsit_p6免疫的小鼠受到同样好的保护。推测影响血清抗原特异性抗体预防衣原体感染的因素有很多,包括血清中抗体的数量、抗体和中和能力。此外,除了体液免疫外,细胞免疫尤其是th2反应,对呼吸道衣原体感染也有一定的保护作用。然而,这些推测均需要进一步的研究来证实。尽管如此,基于momp和cpsit_p6的重组抗原p-m由于具多肽疫苗的优势,其比蛋白免疫原更适合作为候选疫苗。
cd4+t细胞是免疫系统中必不可少的一种淋巴细胞,在消除细菌感染方面具有重要作用。本研究中,p-m特异性cd4+细胞的转移显著降低了感染后衣原体的载量。相反,有研究显示cd4+t细胞在感染早期对人类病原体的清除并不重要。部分原因可能是特异性cd4+t细胞在不同病原菌中的作用不同。此外,一些研究表明,特定的cd8+细胞在减少衣原体负担或防止再感染的保护作用十分有限。同样,我们发现,从免疫小鼠转移到幼稚小鼠的特异性cd8+细胞并没有显著降低衣原体载量,尽管特定的cd8+细胞可能在某种程度上起到间接的保护作用。因此,cd8+细胞在消除衣原体或抵御感染方面的作用尚不明确。
综上所述,基于重组多表位抗原的多肽疫苗可诱导产生较强的体液和细胞免疫应答,同时能够有效降低感染后小鼠肺组织的衣原体载量,减轻肺组织的炎性侵润。此外,值得注意的是,cd4+细胞对重组抗原产生的抗鹦鹉衣原体感染免疫保护作用至关重要。因此,考虑到多肽疫苗的优势,我们的研究结果为抗衣原体感染重组多肽疫苗的研制提供了重要依据。
以上仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
序列表
<110>南华大学
<120>多表位抗原及其制备方法与在制备防治鹦鹉热衣原体感染的药物中的应用
<130>mp1827754
<160>1
<170>siposequencelisting1.0
<210>1
<211>52
<212>prt
<213>人工序列(artificialsequence)
<400>1
lyslystyrlysthrserargasnlysasngluglyglyglyglyglu
151015
glyleuthrprolysgluasnlysglyglyglyglyprovalglyasn
202530
proalagluproserglyglyglyglythrthrglualathraspthr
354045
lysseralathr
50
- 还没有人留言评论。精彩留言会获得点赞!