合理的多倍体腺相关病毒载体及其制造和使用方法与流程
相关申请的交叉引用
本申请根据35u.s.c.§119(e)要求2018年5月7日提交的美国临时申请号62/668,056;和2018年5月31日提交的美国临时申请号62/678,675的权益,所述美国临时申请各自的内容通过引用以其整体并入本文。
政府支持声明
本发明根据由美国国立卫生研究院(nationalinstitutesofhealth)授予的拨款号dk084033、ai117408、ai072176、ca016086、ca151652、hl125749和hl112761在政府支持下进行。政府在本发明中拥有一定权利。
关于电子提交的序列表的声明
提供根据37c.f.r.§1.821提交、标题为5470-786wo2_st25.txt、大小为111,597字节、在2018年7月31日生成且经由efs-web提交的ascii文本格式的序列表,以代替纸质副本。该序列表在此通过引用并入本说明书作为其公开内容。
本发明涉及用于产生具有期望特性的合理的多倍体病毒粒子的方法、病毒粒子、此类病毒粒子的基本上均质的群体、用于产生基本上均质的群体的方法及其用途。
发明背景
腺相关病毒(aav)载体已用于超过100个临床试验中,其具有有希望的结果,尤其是用于治疗失明和血友病b。aav是非致病性的,具有广泛的组织向性,并且可以感染分裂或不分裂的细胞。更重要的是,aav载体转导已在临床前和临床试验中诱导长期治疗性转基因表达。目前,存在12种分离用于基因递送的aav血清型。其中,已显示aav8对于小鼠肝脏靶向是最佳的。已经使用aav2和aav8实施具有fix缺乏的临床前动物中的广泛研究和在具有血友病b的患者中的i/ii期临床试验。来自这些试验的结果是非常有希望的;然而,即使使用相同的载体剂量/kg,接受aav/fix的患者的fix表达与在动物模型中实现的表达也不成比例。当在fix敲除小鼠中使用1x1011的编码fix的aav8颗粒用于全身施用时,在血液中检测到正常水平的160%的fix。然而,当施用2x1011的aav8/fix颗粒时,在灵长类动物中仅实现40%的fix,并且在人中发现少于1%的fix。在这些物种中的aav载体转导后的不一致的fix表达可能是由于不同物种中肝细胞向性的改变。来自aavfix临床试验的另一个有趣发现是衣壳特异性细胞毒性t淋巴细胞(ctl)应答,其消除aav转导的肝细胞,导致治疗失败。在aav递送后的动物模型中尚未看到该现象,其指出临床前研究和临床研究之间的另一种变化。当使用高得多剂量的aav/fix载体时,在使用aav2或aav8的两个临床试验中均检测到fix表达;然而,血液fix水平分别在注射后第4周或第9周下降。进一步的研究表明,aav载体感染引发衣壳特异性ctl应答,其似乎消除aav转导的肝细胞。因此,来自这些临床试验的结果突出了探索在不增加载体衣壳负荷的情况下增强aav转导的有效方法的必要性。降低aav衣壳抗原效应的任何载体改进也将影响令人生畏的载体生产问题,并且是可行的基因治疗药物开发的令人欢迎的补充。
由于其安全性和简易性,腺相关病毒(aav)(一种需要辅助病毒用于有效复制的非致病性的依赖性细小病毒)被用作用于基因疗法的病毒载体。aav具有广泛的宿主和细胞类型向性,其能够转导分裂和非分裂的细胞。迄今为止,已鉴定12种aav血清型和多于100种变体。不同的血清型衣壳在组织或培养细胞中具有不同的感染性,其取决于细胞表面上的主要受体和共受体或细胞内运输途径本身。已经确定aav的一些血清型的主要受体,诸如aav2和aav3的硫酸肝素蛋白聚糖(hspg)和aav5的n-连接的唾液酸,而尚未鉴定到aav7和aav8的主要受体。有趣的是,培养细胞中aav载体的转导效率可能不总是转化为动物中的效率。例如,在小鼠肝脏中,aav8比其他血清型诱导高得多的转基因表达,但在培养细胞系中却没有。
在上述12种血清型中,几种aav血清型和变体已用于临床试验中。作为首先表征的衣壳,aav2已最广泛用于基因递送中,诸如莱伯氏先天性黑蒙症的rpe65和血友病b的因子ix(fix)。尽管aav载体的应用已证明是安全的并且在这些临床试验中已实现治疗效果,但aav载体的主要挑战之一是其低感染性,其需要相对大量的病毒基因组。aav8载体是已在具有血友病b的患者中在临床试验中使用的另一种载体。来自aav8/fix肝脏靶向递送的结果已表明,小鼠、非人灵长类动物和人之间的转基因表达存在独特的物种特异性差异。尽管1010vg的具有fix基因的aav8可以在fix敲除小鼠中达到超生理水平(>100%)的fix表达,但只有高剂量(2x1012vg/kg体重)才可以诱导人中可检测到的fix表达。基于上述这些结果,增强aav转导的有效策略的开发仍然是必要的。
大部分人已自然暴露于aav。作为结果,人群中的大部分已在血液和其他体液中发展出针对特定血清型aav的中和抗体(nabs)。在未来的临床试验中,nab的存在为更广泛的aav应用提出另一个主要挑战。已经探索许多方法来增强aav转导或逃逸nab活性,尤其是基于合理设计和定向进化的aav衣壳的基因修饰。尽管几种aav突变体已在体外或动物模型中表明高转导,连同逃逸nab的能力,但衣壳组成的修饰提供了改变亲本aav的细胞向性的能力。
本发明解决了本领域对具有组合的期望特征的aav载体的需求。
发明概述
我们先前的研究已显示,来自不同aav血清型(aav1至aav5)的衣壳与组装aav病毒粒子(术语病毒粒子、衣壳、病毒颗粒和颗粒在本申请中可互换使用)相容,并且大多数分离的aav单克隆抗体识别位于不同aav亚基上的几个位点。另外,来自嵌合aav衣壳的研究表明,可以通过引入主要受体的结构域或来自其他血清型的组织特异性结构域实现更高的转导。aav9聚糖受体引入aav2衣壳增强aav2的转导。将100aa结构域从aav6取代至aav2衣壳中增加肌肉向性。我们发现由来自两种或更多种aav血清型的衣壳构成的多倍体aav载体可能采用来自个别血清型的优势用于更高的转导,但在某些实施方案中不能消除来自亲本的向性。此外,这些多倍体病毒可能具有逃逸被nabs中和的能力,因为大多数nab识别构象表位,并且多倍体病毒粒子可以改变其表面结构。
一种用于生成具有混合或镶嵌衣壳壳的raav的方法是添加编码来自aav血清型的混合物的衣壳蛋白(vp1、vp2和vp3)的aav辅助质粒。这种方法有时被称为交叉装饰(cross-dressing)。在某些实施方案中,其可以改变某些病毒粒子的抗原性模式。然而,产生各种各样的病毒粒子。此外,产生的病毒粒子是具有血清型的混合物的镶嵌体。因此,产生的病毒粒子的群体保留一些将引发抗原应答的颗粒。因此,获得预定病毒粒子的基本上均质的群体是期望的。
我们现在已经发现允许合理设计和产生此类嵌合或改组的病毒粒子的方法。所得病毒粒子有时被称为多倍体、单倍体或三倍体,以指衣壳蛋白vp1、vp2和vp3来自至少两种不同血清型的事实。所述衣壳可以来自任何aav血清型,包括为基因疗法而分离的12种aav血清型、其他物种、突变血清型、此类基因的改组血清型,例如,aav2、vp1.5和aav4vp2、aav4vp3或任何其他期望的aav血清型。该方法允许产生仅期望的病毒粒子的感染性病毒,其产生病毒粒子的基本上均质的群体。
aav病毒粒子具有t=1二十面体对称性,并且由三种结构病毒蛋白vp1、vp2和vp3构成。比率为1:1:8至10(分别为vp1:vp2:vp3)的三种病毒蛋白的60个拷贝形成病毒粒子(rayaprolu,v.,等人,j.virol.87(24):13150-13160(2013)。
在一个实施方案中,aav病毒粒子是分离的病毒粒子,其具有来自不同于其他vp的血清型的病毒结构蛋白vp1、vp2和vp3中的至少一种,且每种vp仅来自一种血清型。例如,vp1仅来自aav2,vp2仅来自aav4,且vp3仅来自aav8。
在一个替代实施方案中,可以构建病毒粒子,其中来自由aav衣壳蛋白vp1、vp2和vp3组成的组的至少一种病毒蛋白不同于形成能够使aav基因组衣壳化的病毒粒子所需的其他病毒蛋白中的至少一种。对于存在的每种病毒蛋白(vp1、vp2和/或vp3),该蛋白是相同的类型(例如,所有aav2vp1)。在一种情况下,所述病毒蛋白中的至少一种是嵌合病毒蛋白,且其他两种病毒蛋白中的至少一种不是嵌合的。在一个实施方案中,vp1和vp2是嵌合的,且仅vp3是非嵌合的。例如,仅由来自嵌合aav2/8的vp1/vp2(aav2的n-末端和aav8的c-末端)构成的病毒颗粒与仅来自aav2的vp3配对;或仅嵌合vp1/vp228m-2p3(来自aav8的n-末端和来自aav2的c-末端,没有vp3起始密码子的突变)与仅来自aav2的vp3配对。在另一个实施方案中,仅vp3是嵌合的且vp1和vp2是非嵌合的。在另一个实施方案中,所述病毒蛋白中的至少一种来自完全不同的血清型。例如,仅嵌合vp1/vp228m-2p3与仅来自aav3的vp3配对。在另一个实例中,不存在嵌合体。
在一个实施方案中,可以用仅两种病毒结构蛋白vp1和vp3形成使aav基因组(在2个aavitr之间包括异源基因)衣壳化的aav病毒粒子。在一个实施方案中,该病毒粒子是构象上正确的,即具有t=1二十面体对称性。在一个实施方案中,所述病毒粒子是感染性的。
所述群体是至少101个病毒粒子,至少102个病毒粒子,至少103个病毒粒子,至少104个病毒粒子,至少105个病毒粒子,…至少1010个病毒粒子,至少1011个病毒粒子,至少1012个病毒粒子,至少1015个病毒粒子,至少1017个病毒粒子。在一个实施方案中,所述群体是至少100个病毒颗粒。在一个实施方案中,所述群体是109至1012个病毒粒子。
在一个实施方案中,所述群体是至少1x104病毒基因组(vg)/ml,是至少1x105病毒基因组(vg)/ml,是至少1x106病毒基因组(vg)/ml,至少1x107病毒基因组(vg)/ml,至少1x108病毒基因组(vg)/ml,至少1x109病毒基因组(vg)/ml,至少1x1010vg/每ml,至少1x1011vg/每ml,至少1x1012vg/每ml。在一个实施方案中,所述群体范围为约1x105vg/ml至约1x1013vg/ml。
基本上均质的群体是至少90%、至少91%、至少93%、至少95%、至少97%、至少99%、至少99.5%或至少99.9%的仅期望的病毒粒子。在一个实施方案中,所述群体是完全均质的。
aav2和aav8已经用于临床应用。在一个实施方案中,我们首先表征来自aav2和aav8的单倍体aav病毒在体外和体内的转导效率以及nab逃逸能力,即免疫应答,诸如抗原应答。在该研究中,我们发现单倍体载体的病毒产率未被损害,并且肝素结合概况与aav2衣壳亚基蛋白的并入相关。单倍体载体aav2/8在小鼠肌肉和肝脏中引发更高的转导。当应用于具有fix缺乏的小鼠模型时,与aav8组相比,在单倍体载体治疗的小鼠中观察到更高的fix表达和改善的出血表型校正。重要的是,单倍体病毒aav2/8具有对a20的低结合亲和力,并且能够逃逸来自抗aav2血清的中和。下一多倍体病毒aav2/8/9由三种血清型(aav2、8和9)的衣壳制成。证明单倍体aav2/8/9针对用亲本血清型免疫的血清的中和抗体逃逸能力显著提高。
因此,在一个实施方案中,本发明提供了腺相关病毒(aav)衣壳,其中所述衣壳包含衣壳蛋白vp1,其中所述衣壳蛋白vp1来自一种或多于一种第一aav血清型,和衣壳蛋白vp3,其中所述衣壳蛋白vp3来自一种或多于一种第二aav血清型,且其中所述第一aav血清型中的至少一种不同于所述第二aav血清型中的至少一种,其呈任何组合。优选地,这种群体是基本上均质的。在一些实施方案中,本发明的衣壳包含衣壳蛋白vp2,其中所述衣壳蛋白vp2来自一种或多于一种第三aav血清型,其中所述一种或多于一种第三aav血清型中的至少一种不同于所述第一aav血清型和/或所述第二aav血清型,其呈任何组合。
在一些实施方案中,aav病毒粒子可以通过多于3种典型的病毒结构蛋白vp1、vp2和vp3形成(参见例如,wang,q.等人,“syngeneicaavpseudo-particlespotentiategenetransductionofaavvectors,”moleculartherapy:methodsandclinicaldevelopment,vol.4,149-158(2017))。此类病毒衣壳也落入本发明内。例如,分离的aav病毒粒子,其具有足以形成将aav基因组衣壳化的aav病毒粒子的病毒衣壳结构蛋白,其中所述病毒衣壳结构蛋白中的至少一种不同于其他病毒衣壳结构蛋白,且其中每种病毒衣壳结构蛋白只有相同的类型。在进一步实施方案中,分离的aav病毒粒子具有至少两种来自由aav衣壳蛋白vp1、vp2、vp1.5和vp3组成的组的病毒结构蛋白,其中所述两种病毒蛋白足以形成将aav基因组衣壳化的aav病毒粒子,且其中存在的病毒结构蛋白中的至少一种来自与另一种病毒结构蛋白不同的血清型,且其中vp1仅来自一种血清型,vp2仅来自一种血清型,vp1.5仅来自一种血清型,且vp3仅来自一种血清型。例如,vp1.5可以来自aav血清型2,且vp3可以来自aav血清型8。
在一些实施方案中,本发明的衣壳包含衣壳蛋白vp1.5,其中所述衣壳蛋白vp1.5来自一种或多于一种第四aav血清型,其中所述一种或多于一种第四aav血清型中的至少一种不同于所述第一aav血清型和/或所述第二aav血清型,其呈任何组合。在一些实施方案中,本文所述的aav衣壳蛋白可包含衣壳蛋白vp2。
因此,在某些实施方案中,病毒结构蛋白中的至少一种可以是嵌合病毒结构蛋白,即,可以含有来自多于一种蛋白的区段。在一个实施方案中,嵌合病毒结构蛋白全部来自相同血清型。在另一个实施方案中,嵌合病毒结构蛋白由来自多于一种血清型的组分构成,但这些血清型不同于至少一种其他血清型。在一个实施方案中,病毒结构蛋白不是嵌合的。在一个实施方案中,嵌合aav结构蛋白不包含来自犬细小病毒的结构氨基酸。在一个实施方案中,嵌合aav结构蛋白不包含来自b19细小病毒的结构氨基酸。在一个实施方案中,嵌合aav结构蛋白不包含来自犬细小病毒或b19细小病毒的结构氨基酸。在一个实施方案中,嵌合aav结构蛋白仅包含来自aav的结构氨基酸。
在一些实施方案中,仅产生含有至少一种不同于其他病毒蛋白的病毒蛋白的病毒粒子。例如,vp1和vp2来自相同血清型,且vp3仅来自替代血清型。在其他实施方案中,vp1仅来自一种血清型,且vp2和vp3仅来自另一种血清型。在另一个实施方案中,仅产生其中vp1来自一种血清型、vp2来自第二血清型且vp3来自又另一种血清型的颗粒。
这可以通过例如位点特异性缺失和/或添加、改变剪接供体位点、剪接受体位点、起始密码子及其组合来进行。
使用aav血清型2作为示例性病毒,m11是vp1起始密码子,m138是vp2起始密码子,且m203是vp3起始密码子。尽管通常通过取代m11和m138的起始密码子的缺失将使vp1和vp2的表达不起作用,但vp3起始密码子的类似缺失是不够的。这是因为病毒衣壳orf含有许多具有不同强度的atg密码子作为起始密码子。因此,在设计不表达vp3的构建体中,必须小心以确保不产生替代的vp3种类。对于vp3,m138的消除是必需的,或如果需要vp2,但不是vp3,则除了m203以外,m211和235的缺失通常是最好的方法(warrington,k.h.jr.,等人,j.ofvirol.78(12):6595-6609(june2004))。这可以通过突变诸如取代或本领域已知的其他方式来进行。可以容易地确定其他血清型中相应的起始密码子,以及诸如vp3中的额外atg序列是否可以充当替代起始密码子。
这允许用于产生多倍体病毒粒子–诸如单倍体或三倍体病毒颗粒的基本上均质的群体的方法。
本发明还提供了aav衣壳,其中所述衣壳包含衣壳蛋白vp1,其中所述衣壳蛋白vp1来自一种或多于一种第一aav血清型,和衣壳蛋白vp2,其中所述衣壳蛋白vp2来自一种或多于一种第二aav血清型,且其中所述第一aav血清型中的至少一种不同于所述第二aav血清型中的至少一种,其呈任何组合。
在一些实施方案中,所述衣壳包含衣壳蛋白vp3,其中所述衣壳蛋白vp3来自一种或多于一种第三aav血清型,其中所述一种或多于一种第三aav血清型中的至少一种不同于所述第一aav血清型和/或所述第二aav血清型,其呈任何组合。在一些实施方案中,本文所述的aav衣壳可包含衣壳蛋白vp1.5。
本发明进一步提供了腺相关病毒(aav)衣壳,其中所述衣壳包含衣壳蛋白vp1,其中所述衣壳蛋白vp1来自一种或多于一种第一aav血清型,和衣壳蛋白vp1.5,其中所述衣壳蛋白vp1.5来自一种或多于一种第二aav血清型,且其中所述第一aav血清型中的至少一种不同于所述第二aav血清型中的至少一种,其呈任何组合。
在额外实施方案中,本发明提供了病毒载体,其包含:(a)本发明的aav衣壳;和(b)包含至少一个末端重复序列的核酸,其中所述核酸由aav衣壳包裹。所述病毒载体可以是aav颗粒,且本发明的衣壳蛋白、衣壳、病毒载体和/或aav颗粒可以存在于进一步包含药学上可接受的载体的组合物中。
本文进一步提供了制备包含任一前述权利要求的aav衣壳的aav颗粒的方法,其包括:(a)用一种或多种质粒转染宿主细胞,所述质粒组合提供组装aav颗粒所需的所有功能和基因;(b)将一种或多种核酸构建体引入包装细胞系或生产细胞系,以组合提供组装aav颗粒所需的所有功能和基因;(c)将一种或多种重组杆状病毒载体引入宿主细胞,所述杆状病毒载体组合提供组装aav颗粒所需的所有功能和基因;和/或(d)将一种或多种重组疱疹病毒载体引入宿主细胞,所述重组疱疹病毒载体组合提供组装aav颗粒所需的所有功能和基因。
在进一步实施方案中,本发明进一步提供了将核酸施用于细胞的方法,所述方法包括使所述细胞与本发明的病毒载体和/或本发明的组合物接触。
本文还提供了将核酸递送至受试者的方法,所述方法包括向受试者施用本发明的病毒载体和/或本发明的组合物。
另外,本文提供了本发明的衣壳蛋白、衣壳、病毒载体、aav颗粒和/或组合物,其用作在疾病或疾病的有益治疗中的药物。
本发明的这些及其他方面在下文阐述的本发明的说明书中更详细地描述。
附图简述
图1:单倍体病毒的体外转导概况。将单倍体或亲本病毒以104vg/细胞添加至huh7或c2c12细胞。转导后48h,裂解细胞用于荧光素酶测定。数据代表三次分开感染的平均值,其中标准偏差由误差条指示。
图2:小鼠肌肉中的单倍体病毒的转导。经由直接肌肉注射将1x1010vg的单倍体病毒、亲本病毒或与aav2和aav8混合的病毒注射入c57bl/6小鼠。每组包括4只小鼠。(小图a)一周后,通过ivis成像系统对荧光素酶基因表达进行成像。(小图b)测量并计算光子信号。数据代表每组中4只注射的小鼠的荧光素酶基因表达值的平均值,其中标准偏差由误差条指示。面朝上:左腿-aav8或单倍体或混合病毒,右腿-aav2。
图3:小鼠肝脏中的单倍体病毒的转导。经由静脉内注射施用3x1010vg的单倍体病毒。注射后第1周,通过ivis成像系统对荧光素酶表达进行成像(小图a),并测量并计算光子信号(小图b)。注射后第2周,对小鼠安乐死,并收获其肝脏用于dna提取,通过qpcr测量肝脏中的aav基因组拷贝((小图c),并计算每个aav基因组拷贝数的相对荧光素酶表达(小图d)。数据代表4只小鼠的平均值和标准偏差。
图4:经由单倍体病毒递送的fix的治疗水平。fix敲除小鼠经由尾静脉注射1x1010vg的每种载体。在注射后1、2和4周,收集血液样品。(小图a)通过酶联免疫吸附测定法测试hfix蛋白水平。(小图b)通过hfix特异性的一级凝血测定法测试hfix功能。注射后第6周,通过测量盐水溶液中血红蛋白含量的a575处的吸光度确定失血量(小图c)。数据代表来自5只小鼠(敲除小鼠和正常小鼠,未经aav处理,作为对照)或8只小鼠(aav8fix或aav2/81:3/fix处理组)的平均值和标准偏差。
图5:来自aav2和aav8的单倍体aav82的转导。小图a.aav衣壳亚基的组成。小图b.单倍体病毒的western印迹。小图c.代表性成像和肝脏转导的定量。小图d.代表性成像和肌肉转导的定量。
图6:用三倍体病毒aav2/8/9的肝脏转导。经由眼眶后静脉注射3x1010vg的单倍体病毒。注射后第1周,通过ivis成像系统对荧光素酶基因表达进行成像(小图a),并测量并计算光子信号(小图b)。数据代表5只小鼠的平均值和标准偏差。
图7:aav针对热的稳定性。
图8:通过突变衣壳蛋白vp1的起始密码子的单倍体设计。
图9:通过突变剪接受体位点a2的单倍体设计。
图10:通过突变剪接受体位点a1的单倍体设计。
图11:通过突变vp2/vp3的衣壳蛋白的起始密码子和剪接受体位点a2的单倍体设计。
图12:通过突变衣壳蛋白vp1的起始密码子和剪接受体位点a1的单倍体设计。
图13:使用两种质粒的单倍体载体产生。
图14:使用三种质粒的单倍体载体产生。
图15:使用四种质粒的单倍体载体产生。
图16:显示使用dna改组以获得具有期望特征的病毒粒子的示意图。
图17:包括aav2衣壳蛋白的dna序列(seqidno:139)的质粒,其中vp1和vp2的起始密码子已被突变。
图18:包括aav2衣壳蛋白的dna序列(seqidno:140)的质粒,其中vp1的起始密码子已被突变。
图19:包括aav2衣壳蛋白的dna序列(seqidno:141)的质粒,其中vp2和vp3的起始密码子已被突变。
图20:包括aav2衣壳蛋白的dna序列(seqidno:142)的质粒,其中vp2的起始密码子已被突变。
图21:被取代以生成新型多倍体aav衣壳的单个或多个亚基。
图22a-c:单倍体载体h-aav82的肝脏转导。(22a)aav衣壳亚基的组成。从两种质粒(一种编码vp1和vp2,另一种编码vp3)的共转染产生单倍体aav病毒。(22b)将3x1010aav载体的颗粒经由眼眶后静脉注射至c57bl小鼠中。一周后进行成像。(22c)肝脏转导的定量。数据代表5只小鼠的平均值和标准偏差。
图23a-b:单倍体载体h-aav82的肌肉转导。将1x109aav/luc的颗粒注射至小鼠后腿肌肉中。注射后第3周,进行成像3分钟。面朝上:左腿-单倍体aav,右腿-aav2。(23a)代表性成像。(23b)肌肉注射后来自4只小鼠的数据。通过来自单倍体aav至aav2的转导来计算转导的倍数增加。
图24a-c:单倍体载体h-aav92的肝脏转导。(24a)aav衣壳亚基的组成。从两种质粒(一种编码aav9vp1和vp2,另一种编码aav2vp3)的共转染产生单倍体aav病毒。(24b)将3x1010aav载体的颗粒经由眼眶后静脉注射至c57bl小鼠中。一周后进行成像。(24c)肝脏转导的定量。数据代表5只小鼠的平均值和标准偏差。
图25a-c:单倍体载体h-aav82g9的肝脏转导。(25a)aav衣壳亚基的组成。从两种质粒(一种编码aav8vp1和vp2,另一种编码aav2g9vp3)的共转染产生单倍体aav病毒。(25b)将3x1010aav载体的颗粒经由眼眶后静脉注射至c57bl小鼠中。aav施用后第1周,实施成像。(25c)肝脏转导的定量。数据代表5只小鼠的平均值和标准偏差。
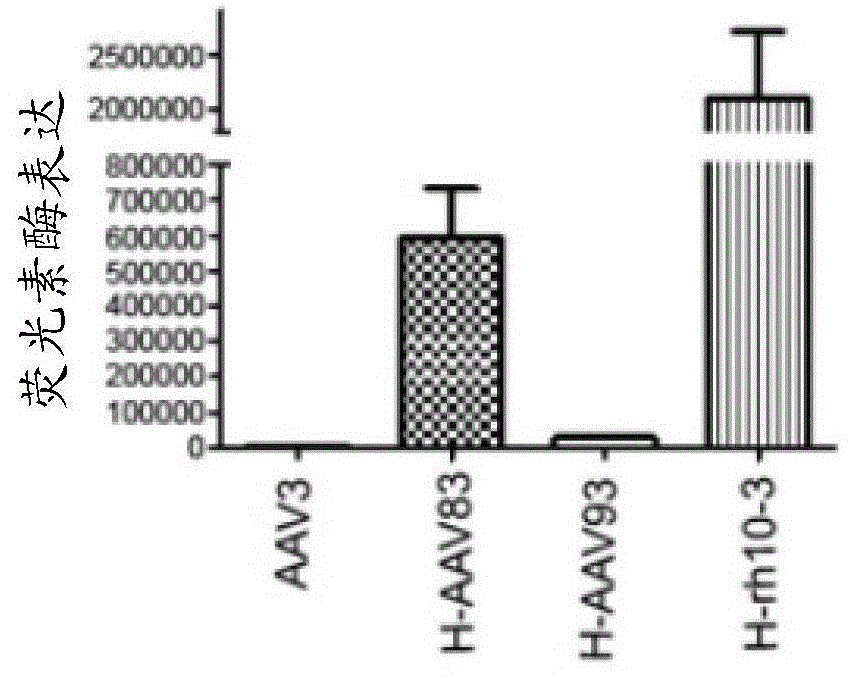
图26a-d:单倍体aav83、aav93和aavrh10-3的肝脏转导。(26a)aav衣壳亚基的组成。(26b)代表性成像。(26c)肝脏转导的定量。(26d)与小鼠核纤层蛋白(表达水平的内部对照)相比,指定器官中病毒基因组的定量。
图27a-d:来自aav2和aav8的单倍体aav82的转导。(27a)aav衣壳亚基的组成。(27b)单倍体病毒的western印迹。(27c)代表性成像和肝脏转导的定量。(27d)代表性成像和肌肉转导的定量。
图28:单倍体结合和运输的能力的分析。
图29:aav针对热的稳定性。
图30:在不同ph下的n-末端暴露的检测。
发明详述
本发明现在参考附图进行描述,其中显示了本发明的代表性实施方案。然而,本发明可以以不同形式体现,并且不应解释为限制于本文阐述的实施方案。相反,提供这些实施方案,以便本公开内容将是彻底和完整的,并且将本发明的范围完全传达给本领域技术人员。
除非另有定义,否则本文使用的所有技术和科学术语均具有与本发明所属领域普通技术人员通常理解相同的含义。在本文中本发明的说明书中使用的术语仅用于描述具体实施方案的目的,并且不预期限制本发明。本文提及的所有出版物、专利申请、专利、登录号及其他参考文献通过引用以其整体并入本文。
在本发明描述和所附权利要求书中在aav衣壳病毒结构蛋白中的所有氨基酸位置的命名都是关于vp1衣壳亚基编号(天然aav2vp1衣壳蛋白:genbank登录号aac03780或yp680426)。本领域技术人员将理解,如果插入aavcap基因中,本文描述的修饰可以导致构成衣壳亚基的结构病毒蛋白vp1、vp2和/或vp3中的修饰。或者,可以独立地表达衣壳亚基以在仅一种或两种衣壳亚基(vp1、vp2、vp3、vp1+vp2、vp1+vp3或vp2+vp3)中实现修饰。
定义
下述术语用于本文说明书和所附的权利要求中:
单数形式“一个/种(a)”、“一个/种(an)”和“该/所述”预期还包括复数形式,除非上下文另有明确说明。
此外,如本文所用,当提及可测量值诸如多核苷酸或多肽序列的长度、剂量、时间、温度等的量时,术语“约”意欲涵盖指定量±20%、±10%、±5%、±1%、±0.5%或甚至±0.1%的变化。
还如本文所用,“和/或”是指且涵盖相关列出项中的一种或多种的任何和所有可能组合,以及当以替代(“或”)解释时的组合缺乏。
如本文所用,连接词“基本上由...组成”意味着权利要求的范围被解释为涵盖权利要求中所述的指定材料或步骤,和“不实质上影响要求保护的发明的基本和新颖特征的那些”。参见,inreherz,537f.2d549,551-52,190uspq461,463(ccpa1976)(原文中强调);还参见mpep§2111.03。因此,术语“基本上由...组成”当在本发明的权利要求中使用时,并不旨在被解释为等同于“包含”。除非上下文另有说明,特别预期本文描述的本发明的各种特征可以以任何组合使用。
此外,本发明还考虑了在本发明的一些实施方案中,可以排除或省略本文阐述的任何特征或特征的组合。
为了进一步举例说明,如果例如说明书指示特定氨基酸可以选自a、g、i、l和/或v,则该措辞还指示氨基酸可以选自这些氨基酸的任何子集,例如a、g、i或l;a、g、i或v;a或g;仅l;等,如同每种此类子组合在本文中明确阐述一样。此外,此类措辞还指示指定氨基酸中的一个或多个可以被放弃(例如,通过否定性附带条件)。例如,在具体实施方案中,氨基酸不是a、g或i;不是a;不是g或v;等,如同每种此类可能的放弃在本文中明确阐述一样。
如本文所用,术语“降低(reduce)”、“降低(reduces)”、“降低(reduction)”和相似术语意指至少约25%、35%、50%、75%、80%、85%、90%、95%、97%或更多的减少。
如本文所用,术语“增强(enhance)”、“增强(enhances)”、“增强(enhancement)”和相似术语指示至少约25%、50%、75%、100%、150%、200%、300%、400%、500%或更多的增加。
如本文所用的术语“细小病毒(parvovirus)”涵盖细小病毒科(parvoviridae),包括自主性复制细小病毒和依赖病毒。自主性细小病毒包括细小病毒属(parvovirus)、红病毒属(erythrovirus)、浓核病毒属(densovirus)、艾特拉病毒属(iteravirus)和康特拉病毒属(contravirus)的成员。示例性自主性细小病毒包括但不限于小鼠的微小病毒、牛细小病毒、犬细小病毒、鸡细小病毒、猫泛白细胞减少症病毒、猫细小病毒、鹅细小病毒、h1细小病毒、番鸭细小病毒、b19病毒和现在已知或后来发现的任何其他自主性细小病毒。其他自主性细小病毒是本领域技术人员已知的。参见例如bernardn.fields等人,virology,第2卷,第69章(第4版,lippincott-ravenpublishers)。
如本文所用,术语“腺相关病毒”(aav)包括但不限于aav1型、aav2型、aav3型(包括3a和3b型)、aav4型、aav5型、aav6型、aav7型、aav8型、aav9型、aav10型、aav11型、禽aav、牛aav、犬aav、马aav、绵羊aav及任何其他目前已知或以后发现的aav。参见例如fields等人virology,第2卷,第69章(第4版,lippincott-ravenpublishers)。许多相对新的aav血清型和进化枝已得到鉴定(参见例如gao等人,(2004)j.virology78:6381-6388;moris等人,(2004)virology33-:375-383;和表3)。
aav和自主性细小病毒的多种血清型的基因组序列,以及天然末端重复(trs)、rep蛋白和衣壳亚基的序列是本领域已知的。此类序列可以在文献或公共数据库诸如genbank中找到。参见例如,genbank登录号nc_002077、nc_001401、nc_001729、nc_001863、nc_001829、nc_001862、nc_000883、nc_001701、nc_001510、nc_006152、nc_006261、af063497、u89790、af043303、af028705、af028704、j02275、j01901、j02275、x01457、af288061、ah009962、ay028226、ay028223、nc_001358、nc_001540、af513851、af513852、ay530579;其公开内容通过引用并入本文用于教导细小病毒和aav核酸和氨基酸序列。还参见例如srivistava等人,(1983)j.virology45:555;chiarini等人,(1998)j.virology71:6823;chiarini等人,(1999)j.virology73:1309;bantel-schaal等人,(1999)j.virology73:939;xiao等人,(1999)j.virology73:3994;muramatsu等人,(1996)virology221:208;shade等人,(1986)j.virol.58:921;gao等人,(2002)proc.nat.acad.sci.usa99:11854;moris等人,(2004)virology33-:375-383;国际专利公开wo00/28061,wo99/61601,wo98/11244;和美国专利号6,156,303;其公开内容通过引用并入本文用于教导细小病毒和aav核酸和氨基酸序列。还参见表1。
自主性细小病毒和aav的衣壳结构更详细地描述于bernardn.fields等人,virology,第2卷,第69&70章(第4版,lippincott-ravenpublishers)。还参见aav2(xie等人,(2002)proc.nat.acad.sci.99:10405-10)、aav4(padron等人,(2005)j.virol.79:5047-58)、aav5(walters等人,(2004)j.virol.78:3361-71)和cpv(xie等人,(1996)j.mol.biol.6:497-520和tsao等人,(1991)science251:1456-64)的晶体结构的描述。
如本文所用的术语“向性(tropism)”是指病毒优先进入某些细胞或组织内,任选随后为由病毒基因组携带的序列在细胞中的表达(例如转录和任选翻译),例如对于重组病毒,异源目标核酸的表达。
如本文所用,“全身性向性”和“全身性转导”(和等同术语)表明,本发明的病毒衣壳或病毒载体表现出对全身的组织(例如,脑、肺、骨骼肌、心脏、肝脏、肾脏和/或胰腺)的向性和/或转导全身的组织(例如,脑、肺、骨骼肌、心脏、肝脏、肾脏和/或胰腺)。在本发明的实施方案中,观察到中枢神经系统(例如,脑、神经细胞等)的全身性转导。在其他实施方案中,实现心肌组织的全身性转导。
如本文所用,“选择性向性”或“特异性向性”意指病毒载体向某些靶细胞和/或某些组织的递送和/或某些靶细胞和/或某些组织的特异性转导。
除非另外指出,否则“有效转导”或“有效向性”或类似术语,可通过参考合适的对照而确定(例如,分别为对照的至少约50%、60%、70%、80%、85%、90%、95%、100%、125%、150%、175%、200%、250%、300%、350%、400%、500%或更多的转导或向性)。在具体实施方案中,病毒载体有效地转导神经细胞和心肌细胞或对神经细胞和心肌细胞具有有效的向性。合适的对照将取决于各种因素,包括期望的向性和/或转导概况。
类似地,通过参考合适对照,可以确定病毒对于靶组织是否“未有效转导”或“不具有有效向性”或类似术语。在具体实施方案中,所述病毒载体未有效转导肝、肾、性腺和/或生殖细胞(即已对其不具有有效向性)。在具体实施方案中,组织(例如肝)的转导(例如不期望的转导)是期望靶组织(例如,骨骼肌、膈肌、心肌和/或中枢神经系统的细胞)的转导水平的20%或更少、10%或更少、5%或更少、1%或更少、0.1%或更少。
在本发明的一些实施方案中,包含本发明的衣壳的aav颗粒可以表明某些组织/细胞的有效转导和对于其转导是不期望的某些组织/细胞的非常低水平的转导(例如,减少的转导)的多种表型。
如本文所用,术语“多肽”涵盖肽和蛋白两者,除非另有说明。
“多核苷酸”是核苷酸碱基的序列,并且可以是rna、dna或dna-rna杂合序列(包括天然存在和非天然存在的核苷酸两者),但在代表性实施方案中,是单链或双链dna序列。
如本文所用,“分离的”多核苷酸(例如“分离的dna”或“分离的rna”)意指至少部分与天然存在的生物或病毒的至少一些其他组分,例如通常发现与多核苷酸结合的细胞或病毒结构组分或者其他多肽或核酸分开的多核苷酸。在代表性实施方案中,与起始材料相比,“分离的”核苷酸富集至少约10倍、100倍、1000倍、10,000倍或更多倍。
同样地,“分离的”多肽意指至少部分与天然存在的生物或病毒的至少一些其他组分,例如通常发现与多肽结合的细胞或病毒结构组分或者其他多肽或核酸分开的多肽。在代表性实施方案中,与起始材料相比,“分离的”多肽富集至少约10倍、100倍、1000倍、10,000倍或更多倍。
“分离的细胞”是指与通常以其天然状态与其缔合的其他组分分离的细胞。例如,分离的细胞可以是在培养基中的细胞和/或在本发明的药学上可接受的载体中的细胞。因此,分离的细胞可以被递送至和/或引入受试者。在一些实施方案中,分离的细胞可以是从受试者移出并如本文所述离体操作且然后返回受试者的细胞。
病毒粒子的群体可以通过本文所述的任何方法生成。在一个实施方案中,所述群体是至少101个病毒粒子。在一个实施方案中,所述群体是至少102个病毒粒子、至少103个病毒粒子、至少104个病毒粒子、至少105个病毒粒子、至少106个病毒粒子、至少107个病毒粒子、至少108个病毒粒子、至少109个病毒粒子、至少1010个病毒粒子、至少1011个病毒粒子、至少1012个病毒粒子、至少1013个病毒粒子、至少1014个病毒粒子、至少1015个病毒粒子、至少1016个病毒粒子或至少1017个病毒粒子。病毒粒子的群体可以是异质的或可以是均质的(例如,基本上均质的或完全均质的)。
如本文所用的术语“基本上均质的群体”是指大部分相同的病毒粒子的群体,其中污染的病毒粒子(不相同的那些)很少至没有。基本上均质的群体是至少90%的相同病毒粒子(例如,期望的病毒粒子),并且可以是至少91%、至少92%、至少93%、至少94%、至少95%、至少96%、至少97%、至少98%、至少99%、至少99.5%、至少99.9%的相同病毒粒子。
完全均质的病毒粒子的群体仅含有相同的病毒粒子。
如本文所用,“分离”或“纯化”(或语法同义词)病毒载体或病毒颗粒或病毒颗粒群体意指病毒载体或病毒颗粒或病毒颗粒群体至少部分与起始材料中的至少一些其他组分分开。在代表性实施方案中,与起始材料相比,“分离的”或“纯化的”病毒载体或病毒颗粒或病毒颗粒群体富集至少约10倍、100倍、1000倍、10,000倍或更多倍。
“治疗多肽”是可以减轻、降低、预防、延迟和/或稳定症状的多肽,所述症状产生于蛋白在细胞或受试者中的不存在或缺陷,和/或是以其他方式对受试者赋予益处,例如抗癌效应或移植物存活性的改善或免疫应答的诱导的多肽。
术语“治疗(treat)”、“治疗(treating)”或“……的治疗”(及其语法变体)意指受试者状况的严重程度降低,至少部分改善或稳定,和/或实现至少一种临床症状中的一定减轻、缓和、减少或稳定,和/或在疾病或病症的进展中存在延迟。
术语“预防(prevent)”、“预防(preventing)”和“预防(prevention)”(及其语法变体)是指相对于在不存在本发明的方法的情况下出现的那种,受试者中的疾病、病症和/或临床症状的发作预防和/或延迟,和/或疾病、病症和/或临床症状的发作严重程度的降低。预防可以是完全的,例如疾病、病症和/或临床症状的完全不存在。预防还可以是部分的,使得受试者中的疾病、病症和/或临床症状的出现和/或发作的严重程度显著小于在不存在本发明的情况下出现的那种。
如本文所用,“治疗有效”量是足以对受试者提供一定改善或益处的量。或者说,“治疗有效”量是提供受试者中的至少一种临床症状中的一定减轻、缓和、减少或稳定的量。本领域技术人员应当理解疗效无需是完全或治愈的,只要对受试者提供一定益处。
如本文所用,“预防有效”量是相对于在不存在本发明的方法的情况下出现的那种,足以预防和/或延迟受试者中的疾病、病症和/或临床症状的发作,和/或降低和/或延迟受试者中的疾病、病症和/或临床症状的发作严重程度的量。本领域技术人员应当理解预防水平无需是完全的,只要对受试者提供一定预防性益处。
术语“异源核苷酸序列”和“异源核酸”在本文中可互换使用,并且是指没有在病毒中天然存在的核酸序列。通常,异源核酸分子或异源核苷酸序列包含编码目标多肽或非翻译rna(例如用于递送至细胞和/或受试者)的开放读码框。
如本文所用,术语“病毒载体”、“载体”或“基因递送载体”是指病毒(例如aav)颗粒,其充当核酸递送媒介物,并且包含包装在病毒粒子内的载体基因组(例如病毒dna[vdna])。或者,在一些背景下,术语“载体”可以用于指单独的载体基因组/vdna。
“raav载体基因组”或“raav基因组”是包含一种或多种异源核酸序列的aav基因组(即vdna)。raav载体一般仅需要顺式(incis)的末端重复(tr(s)),以生成病毒。所有其他病毒序列是可有可无的,并且可以以反式(intrans)供应(muzyczka,(1992)curr.topicsmicrobiol.immunol.158:97)。通常,raav载体基因组将仅保留一种或多种tr序列,以便使可以由载体有效包装的转基因大小最大化。结构和非结构蛋白编码序列可以以反式提供(例如由载体,诸如质粒,或通过将序列稳定整合到包装细胞内)。在本发明的实施方案中,raav载体基因组包含至少一个tr序列(例如aavtr序列),任选两个tr(例如两个aavtr),其通常将在载体基因组的5'和3'端处,并且侧接异源核酸,但无需与之邻接。tr可以彼此相同或不同。
术语“末端重复”或“tr”包括任何病毒末端重复或合成序列,其形成发夹结构且充当反向末端重复(即,介导所需功能诸如复制、病毒包装、整合和/或原病毒拯救等等)。tr可以是aavtr或非aavtr。例如,非aavtr序列诸如其他细小病毒(例如犬细小病毒(cpv)、小鼠细小病毒(mvm)、人细小病毒b-19)的那些,或任何其他合适的病毒序列(例如充当sv40复制起点的sv40发夹)可以用作tr,其可以进一步通过截短、取代、缺失、插入和/或添加进行修饰。进一步地,tr可以部分或完全是合成的,诸如如授予samulski等人的美国专利号5,478,745中所述的“双重d序列”。
“aav末端重复”或“aavtr”可以来自任何aav,包括但不限于血清型1、2、3、4、5、6、7、8、9、10、11或12或任何其他目前已知或以后发现的aav(参见例如表1)。aav末端重复无需具有天然末端重复序列(例如天然aavtr序列可以通过插入、缺失、截短和/或错义突变进行改变),只要末端重复介导所需功能,例如复制、病毒包装、整合和/或原病毒拯救等等。
aav蛋白vp1、vp2和vp3是一起相互作用以形成二十面体对称的aav衣壳的衣壳蛋白。vp1.5是美国公开号2014/0037585中描述的aav衣壳蛋白。
本发明的病毒载体可以进一步是“靶向的”病毒载体(例如具有指定向性)和/或“杂合”细小病毒(即,其中病毒tr和病毒衣壳来自不同细小病毒),如国际专利公开wo00/28004和chao等人(2000)moleculartherapy2:619中所述。
本发明的病毒载体可以进一步是双链体化的细小病毒颗粒,如国际专利公开wo01/92551(其公开内容通过引用以其整体并入本文)中所述。因此,在一些实施方案中,双链(双链体)基因组可以包装到本发明的病毒衣壳内。
进一步地,病毒衣壳或基因组元件可以含有其他修饰,包括插入、缺失和/或取代。
如本文所用的“嵌合的”病毒结构蛋白意指已通过衣壳蛋白的氨基酸序列中的一个或多个(例如,2、3、4、5、6、7、8、9、10个等)氨基酸残基相对于野生型的取代,以及氨基酸序列中的一个或多个(例如,2、3、4、5、6、7、8、9、10个等)氨基酸残基相对于野生型的插入和/或缺失而修饰的aav病毒结构蛋白(衣壳)。在一些实施方案中,来自一种aav血清型的完全或部分结构域、功能区域、表位等可以以任何组合替换不同aav血清型的相应的野生型结构域、功能区域、表位等,以产生本发明的嵌合衣壳蛋白。在其他实施方案中,所述取代全部来自相同的血清型。在其他实施方案中,所述取代全部来自aav或是合成的。嵌合衣壳蛋白的产生可以根据本领域中众所周知的方案实施,并且大量的嵌合衣壳蛋白在文献以及本文中描述,其可以包括在本发明的衣壳中。
在一个替代实施方案中,可以构建病毒粒子,其中来自由aav衣壳蛋白vp1、vp2和vp3组成的组的至少一种病毒蛋白不同于形成能够使aav基因组衣壳化的病毒粒子所需的其他病毒蛋白中的至少一种。对于存在的每种病毒蛋白(vp1、vp2和/或vp3),该蛋白是相同的类型(例如,所有aav2vp1)。在一种情况下,所述病毒蛋白中的至少一种是嵌合病毒蛋白,且其他两种病毒蛋白中的至少一种不是嵌合的。在一个实施方案中,vp1和vp2是嵌合的,且仅vp3是非嵌合的。例如,仅由来自嵌合aav2/8的vp1/vp2(aav2的n-末端和aav8的c-末端)构成的病毒颗粒与仅来自aav2的vp3配对;或仅嵌合vp1/vp228m-2p3(来自aav8的n-末端和来自aav2的c-末端,没有vp3起始密码子的突变)与仅来自aav2的vp3配对。在另一个实施方案中,仅vp3是嵌合的且vp1和vp2是非嵌合的。在另一个实施方案中,所述病毒蛋白中的至少一种来自完全不同的血清型。例如,仅嵌合vp1/vp228m-2p3与仅来自aav3的vp3配对。在另一个实例中,不存在嵌合体。
如本文所用,术语“氨基酸”涵盖任何天然存在的氨基酸、其修饰形式和合成氨基酸。
天然存在的左旋(l-)氨基酸显示于表2中。
或者,氨基酸可以是修饰的氨基酸残基(非限制性实例显示于表4中)和/或可以是通过翻译后修饰(例如乙酰化、酰胺化、甲酰化、羟基化、甲基化、磷酸化或硫酸化)修饰的氨基酸。
进一步地,非天然存在的氨基酸可以是如wang等人,annurevbiophysbiomolstruct.35:225-49(2006)所述的“非天然”氨基酸。这些非天然氨基酸可以有利地用于将目标分子化学连接至aav衣壳蛋白。
如本文所用,术语“同源重组”意指一种类型的遗传重组,其中核苷酸序列在两个相似或相同的dna分子之间交换。同源重组也产生dna序列的新组合。这些dna的新组合代表遗传变异。同源重组还用于水平基因转移中,以在病毒的不同毒株和物种之间交换遗传物质。
如本文所用,术语“基因编辑”、“基因组编辑”或“基因组工程改造”意指一种类型的基因工程改造,其中使用工程改造的核酸酶或“分子剪刀”在活生物体的基因组中插入、缺失或替换dna。这些核酸酶在基因组中的期望位置处产生位点特异性双链断裂(dsb)。
如本文所用,术语“基因递送”意指将外源dna转移至宿主细胞用于基因疗法的应用的过程。
如本文所用,术语“crispr”代表成簇的规则间隔的短回文重复,其是形成crispr-cas9基因组编辑技术的基础的细菌防御系统的标志。
如本文所用,术语“锌指”意指小的蛋白结构基序,其特征在于一个或多个锌离子的配位以稳定折叠。
在一些实施方案中,本发明的aav颗粒可以是合成的病毒载体,其被设计为展现适合于不同的体内和体外应用的一系列期望表型。因此,在一个实施方案中,本发明提供了包含腺相关病毒(aav)的aav颗粒。
本发明提供了一系列合成的病毒载体,其展现适合于不同的体内和体外应用的一系列期望表型。具体而言,本发明基于以下意料之外的发现:在个别衣壳中组合来自不同aav血清型的衣壳蛋白允许开发在每个个别衣壳中具有多种期望表型的改进的aav衣壳。此类嵌合或改组的病毒粒子有时被称为多倍体、单倍体或三倍体,以指衣壳蛋白vp1、vp2和vp3来自至少两种不同血清型的事实。本文描述了产生此类病毒粒子的新方法。通过防止不期望的开放阅读框的翻译,这些方法导致生成的病毒粒子的均质群体的产生。
生成重组病毒粒子的(例如,基本上或完全)均质的群体的能力显著减少或消除不期望的/污染的病毒粒子的特性(例如,转导特异性或抗原性)的残留。
aav病毒粒子具有t=1二十面体对称性,并且由三种结构病毒蛋白vp1、vp2和vp3构成。比率为1:1:8至10(分别为vp1:vp2:vp3)的三种病毒蛋白的60个拷贝形成病毒粒子(rayaprolu,v.,等人,j.virol.87(24):13150-13160(2013)。
在一个实施方案中,aav病毒粒子是分离的病毒粒子,其具有来自不同于其他vp的血清型的病毒结构蛋白vp1、vp2和vp3中的至少一种,且每种vp仅来自一种血清型。例如,vp1仅来自aav2,vp2仅来自aav4,且vp3仅来自aav8。
在一个实施方案中,可以用仅两种病毒结构蛋白vp1和vp3形成将在2个aavitr之间包括异源基因的aav基因组衣壳化的aav病毒粒子。在一个实施方案中,该病毒粒子是构象上正确的,即具有t=1二十面体对称性。在一个实施方案中,所述病毒粒子是感染性的。
感染性病毒粒子包括vp1/vp3vp1/vp2/vp3。通常,vp2/vp3和仅vp3的病毒粒子不是感染性的。
用于生成这些病毒粒子群体的病毒结构蛋白可以来自为基因疗法而分离的12种aav血清型中的任一种、其他物种、突变血清型、此类基因的改组血清型,例如,aav2、vp1.5和aav4vp2、aav4vp3或任何其他期望的aav血清型。
例如,通过共转染来自血清型2、8和9的aav辅助质粒产生的本文所述的三倍体aav2/8/9载体具有比aav2高得多的小鼠肝脏转导,类似于aav8。重要的是,三倍体aav2/8/9载体具有提高的逃逸来自用亲本血清型免疫的血清的中和抗体的能力。尽管aav3在全身施用后转导小鼠全身的效率较低,但本文所述的单倍体载体h-aav83或h-aav93或h-rh10-3(其中vp3来自aav3,且vp1/vp2来自aav8、9或rh10)诱导全身转导以及与aav3相比在肝脏和其他组织中的高得多的转导。
因此,在一个实施方案中,本发明提供了具有病毒衣壳的腺相关病毒(aav),其中所述衣壳包含蛋白vp1,其中所述vp1来自一种或多于一种第一aav血清型,和衣壳蛋白vp3,其中所述衣壳蛋白vp3来自一种或多于一种第二aav血清型,且其中所述第一aav血清型中的至少一种不同于所述第二aav血清型中的至少一种,其呈任何组合。当至少一种病毒结构蛋白来自多于一种血清型时,我们是指有时被称为“交叉装饰(crossdressing)”的现象,其导致镶嵌衣壳。另一方面,当病毒衣壳蛋白各自来自相同血清型时,即使病毒蛋白中的至少一种来自不同血清型,也不产生镶嵌衣壳。例如,vp1来自aav2,vp2来自aav6,且vp3来自aav8。
在一些实施方案中,本发明的衣壳包含衣壳蛋白vp2,其中所述衣壳蛋白vp2来自一种或多于一种第三aav血清型,其中所述一种或多于一种第三aav血清型中的至少一种不同于所述第一aav血清型和/或所述第二aav血清型,其呈任何组合。在一些实施方案中,本文所述的aav衣壳可包含衣壳蛋白vp1.5。vp1.5描述于美国专利公开号2014/0037585中,且本文提供了vp1.5的氨基酸序列。
在一些实施方案中,仅产生含有至少一种不同于其他病毒蛋白的病毒蛋白的病毒粒子。例如,vp1和vp2来自相同血清型,且vp3仅来自替代血清型。在其他实施方案中,vp1仅来自一种血清型,且vp2和vp3仅来自另一种血清型。在另一个实施方案中,仅产生其中vp1来自一种血清型、vp2来自第二血清型且vp3来自又另一种血清型的颗粒。
这可以通过例如位点特异性缺失和/或添加、改变剪接供体位点、剪接受体位点、起始密码子及其组合来进行。
这允许用于产生多倍体病毒粒子–诸如单倍体或三倍体病毒颗粒的基本上均质的群体的方法。
在一些实施方案中,aav病毒粒子可以通过多于典型的3种病毒结构蛋白vp1、vp2和vp3形成(参见例如,wang,q.等人,“syngeneicaavpseudo-particlespotentiategenetransductionofaavvectors,”moleculartherapy:methodsandclinicaldevelopment,vol.4,149-158(2017))。此类病毒衣壳也落入本发明内。例如,分离的aav病毒粒子,其具有足以形成将aav基因组衣壳化的aav病毒粒子的病毒衣壳结构蛋白,其中所述病毒衣壳结构蛋白中的至少一种不同于其他病毒衣壳结构蛋白,且其中每种病毒衣壳结构蛋白只有相同的类型。在进一步实施方案中,分离的aav病毒粒子具有至少两种来自由aav衣壳蛋白vp1、vp2、vp1.5和vp3组成的组的病毒结构蛋白,其中所述两种病毒蛋白足以形成将aav基因组衣壳化的aav病毒粒子,且其中存在的病毒结构蛋白中的至少一种来自与另一种病毒结构蛋白不同的血清型,且其中vp1仅来自一种血清型,vp2仅来自一种血清型,vp1.5仅来自一种血清型,且vp3仅来自一种血清型。例如,vp1.5可以来自aav血清型2,且vp3可以来自aav血清型8。
在一些实施方案中,本发明的衣壳包含衣壳蛋白vp1.5,其中所述衣壳蛋白vp1.5来自一种或多于一种第四aav血清型,其中所述一种或多于一种第四aav血清型中的至少一种不同于所述第一aav血清型和/或所述第二aav血清型,其呈任何组合。在一些实施方案中,本文所述的aav病毒结构蛋白可包含病毒结构蛋白vp2。
本发明还提供了aav衣壳,其中所述衣壳包含衣壳蛋白vp1,其中所述衣壳蛋白vp1来自一种或多于一种第一aav血清型,和衣壳蛋白vp2,其中所述衣壳蛋白vp2来自一种或多于一种第二aav血清型,且其中所述第一aav血清型中的至少一种不同于所述第二aav血清型中的至少一种,其呈任何组合。在一些实施方案中,病毒粒子中不存在嵌合病毒结构蛋白。
在一些实施方案中,本发明的aav颗粒可包含含有衣壳蛋白vp3的衣壳,其中所述衣壳蛋白vp3来自一种或多于一种第三aav血清型,其中所述一种或多于一种第三aav血清型中的至少一种不同于所述第一aav血清型和/或所述第二aav血清型,其呈任何组合。在一些实施方案中,本文所述的aav衣壳可包含衣壳蛋白vp1.5。
本发明进一步提供了包含腺相关病毒(aav)衣壳的aav颗粒,其中所述衣壳包含衣壳蛋白vp1,其中所述衣壳蛋白vp1来自一种或多于一种第一aav血清型,和衣壳蛋白vp1.5,其中所述衣壳蛋白vp1.5来自一种或多于一种第二aav血清型,且其中所述第一aav血清型中的至少一种不同于所述第二aav血清型中的至少一种,其呈任何组合。
在一些实施方案中,所述衣壳包含衣壳蛋白vp3,其中所述衣壳蛋白vp3来自一种或多于一种第三aav血清型,其中所述一种或多于一种第三aav血清型中的至少一种不同于所述第一aav血清型和/或所述第二aav血清型,其呈任何组合。在一些实施方案中,本文所述的aav衣壳可包含衣壳蛋白vp1.5。
本发明进一步提供了腺相关病毒(aav)衣壳,其中所述衣壳包含衣壳蛋白vp1,其中所述衣壳蛋白vp1来自一种或多于一种第一aav血清型,和衣壳蛋白vp1.5,其中所述衣壳蛋白vp1.5来自一种或多于一种第二aav血清型,且其中所述第一aav血清型中的至少一种不同于所述第二aav血清型中的至少一种,其呈任何组合。
在一些实施方案中,本发明的aav衣壳包含衣壳蛋白vp3,其中所述衣壳蛋白vp3来自一种或多于一种第三aav血清型,其中所述一种或多于一种第三aav血清型中的至少一种不同于所述第一aav血清型和/或所述第二aav血清型,其呈任何组合。在一些实施方案中,本文所述的aav衣壳蛋白可包含衣壳蛋白vp2。
在本发明的衣壳的一些实施方案中,所述一种或多于一种第一aav血清型、所述一种或多于一种第二aav血清型、所述一种或多于一种第三aav血清型和所述一种或多于一种第四aav血清型以任何组合选自表1中列出的aav血清型。
在本发明的一些实施方案中,本文所述的aav衣壳缺乏衣壳蛋白vp2。
在本发明的衣壳的一些实施方案中,包含嵌合衣壳vp1蛋白、嵌合衣壳vp2蛋白、嵌合衣壳vp3蛋白和/或嵌合衣壳vp1.5蛋白。
在一些实施方案中,本发明的aav衣壳可以是aavaav2/8/9、h-aav82、h-aav92、h-aav82g9、aav2/83:1、aav2/81:1、aav2/81:3或aav8/9,其全部描述于本文提供的实施例部分中。
可以以与本文描述的其他衣壳蛋白和/或与现在已知或以后开发的其他衣壳蛋白的任何组合包括于本发明的衣壳中的aav衣壳蛋白的非限制性实例包括lk3、lk01-19、aav-dj、olig001、raav2-retro、aav-lic、aav0kera1、aav-kera2、aav-kera3、aav7m8、aav1,9、aavr3.45、aav克隆32、aav克隆83、aav-u87r7-c5、aavshh13、aavshh19、aavl1-12、aavhae-1、aavhae-2、aav变体shh10、aav2.5t、aavls1-4、aavlsm、aav1289、aavhsc1-17、aav2rec1-4、aav8bp2、aav-b1、aav-php.b、aav9.45、aav9.61、aav9.47、aavm41、aav2展示肽、aav2-gmn、aav9-展示肽、aav8和aav9展示肽、aavpo2.1、aavpo4、aavpo5、aavpo6、aavrh、aavhu、aav-go.1、aav-mo.1、baav、aaav、aav8k137r、aavanc80l65、aav2g9、aav2265插入-aav2/265d、aav2.5、aav3sastg、aav2i8、aav8g9、aav2酪氨酸突变体aav2y-f、aav8y-f、aav9y-f、aav6y-f、aav6.2及其任何组合。
作为一个非限制性实例,本发明的aav衣壳蛋白和病毒衣壳可以是嵌合的,因为它们可以包含来自另一种病毒(任选地另一种细小病毒或aav)的衣壳亚基的全部或一部分,例如如国际专利公开wo00/28004中所述。
以下出版物描述可以与野生型衣壳蛋白和/或现在已知或以后鉴定的其他嵌合或变体衣壳蛋白以任何组合并入本发明的aav衣壳中的嵌合或变体衣壳蛋白。
llisowski,apdane,kchu,yzhang,sccunninghamm,emwilson,等人,selectionandevaluationofclinicallyrelevantaavvariantsinaxenograftlivermodel.nature,506(2014),pp.382–386(lk03和其他lk01-19)。
grimmd,leejs,wangl,desait,akacheb,stormta,kayma.invitroandinvivogenetherapyvectorevolutionviamultispeciesinterbreedingandretargetingofadeno-associatedviruses.j.virol.2008jun:82(12):5887-911.(aav-dj)。
powellsk,khann,parkercl,samulskirj,matsushimag,graysj,mccowntj.characterizationofanoveladeno-associatedviralvectorwithpreferentialoligodendrocytetropism.genether.2016nov:23(11):807-814.(olig001)。
tervodg,hwangby,viswanathans,gajt,lavzinm,ritolakd,lindos,michaels,kuleshovae,ojalad,huangcc,gerfencr,schillerj,dudmanjt,hantmanaw,loogerll,schafferdv,karpovaay.adesigneraavvariantpermitsefficientretrogradeaccesstoprojectionneurons.neuron.2016oct19:92(2):372-382.(raav2-retro)。
marsicd,govindasamyl,currlins,markusicdm,tsengys,herzogrw,agbandje-mckennam,zolotukhins.vectordesigntourdeforce:integratingcombinatorialandrationalapproachestoderivenoveladeno-associatedvirusvariants.molther.2014nov:22(11):1900-9.(aav-lic)。
sallachj,dipasqualeg,larcherf,niehoffn,rübsamm,hubera,chiorinij,almarzad,emingsa,ulush,nishimuras,hackerut,hallekm,niessencm,büningh.tropism-modifiedaavvectorsovercomebarrierstosuccessfulcutaneoustherapy.molther.2014may:22(5):929-39.(aav-kera1、aav-kera2和aav-kera3)。
dalkarad,byrnelc,klimczakrr,viselm,yinl,meriganwh,flanneryjg,schafferdv.invivo-directedevolutionofanewadeno-associatedvirusfortherapeuticouterretinalgenedeliveryfromthevitreous.scitranslmed.2013jun12:5(189):189ra76.(aav7m8)。
asurip,bartelma,vazint,jangjh,wongtb,schafferdv.directedevolutionofadeno-associatedvirusforenhancedgenedeliveryandgenetargetinginhumanpluripotentstemcells.molther.2012feb:20(2):329-38.(aav1.9)。
jangjh,koerberjt,kimjs,asurip,vazint,bartelm,keunga,kwoni,parkki,schafferdv.anevolvedadeno-associatedviralvariantenhancesgenedeliveryandgenetargetinginneuralstemcells.molther.2011apr:19(4):667-75.doi:10.1038/mt.2010.287.(aavr3.45)。
graysj,blakebl,criswellhe,nicolsonsc,samulskirj,mccowntj,liw.directedevolutionofanoveladeno-associatedvirus(aav)vectorthatcrossestheseizure-compromisedblood-brainbarrier(bbb).molther.2010mar:18(3):570-8.(aav克隆32和83)。
maguireca,giannid,meijerdh,shaketla,wakimotoh,rabkinsd,gaog,sena-estevesm.directedevolutionofadeno-associatedvirusforgliomacelltransduction.j.neurooncol.2010feb:96(3):337-47.(aav-u87r7-c5)。
koerberjt,klimczakr,jangjh,dalkarad,flanneryjg,schafferdv.molecularevolutionofadeno-associatedvirusforenhancedglialgenedelivery.molther.2009dec:17(12):2088-95.(aavshh13、aavshh19、aavl1-12)。
liw,zhangl,johnsonjs,zhijianw,griegerjc,ping-jiex,drouinlm,agbandje-mckennam,picklesrj,samulskirj.generationofnovelaavvariantsbydirectedevolutionforimprovedcftrdeliverytohumanciliatedairwayepithelium.molther.2009dec:17(12):2067-77.(aavhae-1、aavhae-2)。
klimczakrr,koerberjt,dalkarad,flanneryjg,schafferdv.anoveladeno-associatedviralvariantforefficientandselectiveintravitrealtransductionofratmüllercells.plosone.2009oct14:4(10):e7467.(aav变体shh10)。
excoffonkj,koerberjt,dickeydd,murtham,keshavjees,kasparbk,zabnerj,schafferdv.directedevolutionofadeno-associatedvirustoaninfectiousrespiratoryvirus.procnatlacadsciusa.2009mar10:106(10):3865-70.(aav2.5t)。
sellnerl,stiefelhagenm,kleinschmidtja,laufss,wenzf,fruehaufs,zellerwj,veldwijkmr.generationofefficienthumanbloodprogenitor-targetedrecombinantadeno-associatedviralvectors(aav)byapplyinganaavrandompeptidelibraryonprimaryhumanhematopoieticprogenitorcells.exphematol.2008aug:36(8):957-64.(aavls1-4,aavlsm)。
liw,asokana,wuz,vandyket,diprimion,johnsonjs,govindaswamyl,agbandje-mckennam,leichtles,redmonddejr,mccowntj,petermannkb,sharplessne,samulskirj.engineeringandselectionofshuffledaavgenomes:anewstrategyforproducingtargetedbiologicalnanoparticles.molther.2008jul:16(7):1252-60.(aav1289)。
charbelissap,desilvasr,lipinskidm,singhms,mouravleva,youq.assessmentoftropismandeffectivenessofnewprimate-derivedhybridrecombinantaavserotypesinthemouseandprimateretina.plosone.2013:8:e60361.(aavhsc1-17)。
huangw,mcmurphyt,liux,wangc,caol.geneticmanipulationofbrownfatviaoraladministrationofanengineeredrecombinantadeno-associatedviralserotypevector.mol.ther.2016jun:24(6):1062-9.(aav2rec1-4)。
cronint,vandenberghelh,hantzp,等人,efficienttransductionandoptogeneticstimulationofretinalbipolarcellsbyasyntheticadeno-associatedviruscapsidandpromoter.embomol.med.2014:6:1175–1190.(aav8bp2)。
choudhurysr,fitzpatrickz,harrisaf,maitlandsa,ferreirajs,zhangy,mas,sharmarb,gray-edwardshl,johnsonja,johnsonak,alonsolc,punzoc,wagnerkr,maguireca,kotinrm,martindr,sena-estevesm.invivoselectionyieldsaav-b1capsidforcentralnervoussystemandmusclegenetherapy.molther.2016aug:24(7):1247-57.(aav-b1)。
devermanbe,pravdopl,simpsonbp,kumarsr,chanky,banerjeea,wuwl,yangb,hubern,pascasp,gradinaruv.cre-dependentselectionyieldsaavvariantsforwidespreadgenetransfertotheadultbrain.natbiotechnol.2016feb:34(2):204-9.doi:10.1038/nbt.3440.(aav-php.b)。
pulicherlan,shens,yadavs,debbinkk,govindasamyl,agbandje-mckennam,asokana.engineeringliver-detargetedaav9vectorsforcardiacandmusculoskeletalgenetransfer.molther.2011jun:19(6):1070-8.(aav9衍生的突变体-aav9.45、aav9.61和aav9.47)。
yangl,jiangj,drouinlm,agbandje-mckennam,chenc,qiaoc,pud,hux,wangdz,lij,xiaox.amyocardiumtropicadeno-associatedvirus(aav)evolvedbydnashufflingandinvivoselection.procnatlacadsciusa.2009mar10:106(10):3946-51.(aavm41)。
körbelinj,siebert,michelfelders,lundingl,spiese,hungera,alawim,raptik,indenbirkend,mülleroj,pasqualinir,arapw,kleinschmidtja,trepelm.pulmonarytargetingofadeno-associatedviralvectorsbynext-generationsequencing-guidedscreeningofrandomcapsiddisplayedpeptidelibraries.molther.2016jun:24(6):1050-61.(aav2展示肽)。
geogheganjc,keisernw,okulista,martinsi,wilsonms,davidsonbl.chondroitinsulfateistheprimaryreceptorforapeptide-modifiedaavthattargetsbrainvascularendotheliuminvivo.molthernucleicacids.2014oct14:3:e202.(aav2-gmn)。
varadik,michelfelders,korfft,heckerm,trepelm,katusha,kleinschmidtja,mülleroj.novelrandompeptidelibrariesdisplayedonaavserotype9forselectionofendothelialcell-directedgenetransfervectors.genether.2012aug:19(8):800-9.(aav9-展示肽)。
michelfelders,varadik,rauppc,hungera,körbelinj,pahrmannc,schrepfers,mülleroj,kleinschmidtja,trepelm.peptideligandsincorporatedintothethreefoldspikecapsiddomaintore-directgenetransductionofaav8andaav9invivo.plosone.2011:6(8):e23101.(aav8和aav9展示肽)。
yucy,yuanz,caoz,wangb,qiaoc,lij,xiaox.amuscle-targetingpeptidedisplayedonaav2improvesmuscletropismonsystemicdelivery.genether.2009aug:16(8):953-62。
michelfelders,leemk,delima-hahne,wilmest,kaulf,müllero,kleinschmidtja,trepelm.vectorsselectedfromadeno-associatedviraldisplaypeptidelibrariesforleukemiacell-targetedcytotoxicgenetherapy.exphematol.2007dec:35(12):1766-76。
mülleroj,kaulf,weitzmanmd,pasqualinir,arapw,kleinschmidtja,trepelm.randompeptidelibrariesdisplayedonadeno-associatedvirustoselectfortargetedgenetherapyvectors.natbiotechnol.2003sep:21(9):1040-6。
grifmanm,trepelm,speecep,gilbertlb,arapw,pasqualinir,weitzmanmd.incorporationoftumor-targetingpeptidesintorecombinantadeno-associatedviruscapsids.molther.2001jun:3(6):964-75。
annegirod,martinried,christianewobus,haraldlahm,kristinleike,jürgenkleinschmidt,gilbertdeléageandmichaelhallek.geneticcapsidmodificationsallowefficientre-targetingofadeno-associatedvirustype2.naturemedicine,1052-1056(1999)。
belloa,chanda,avilesj,souleg,auricchioa,kobingergp.noveladeno-associatedvirusesderivedfrompigtissuestransducemostmajororgansinmice.scirep.2014oct22:4:6644.(aavpo2.1、-po4、-po5和-po6)。
gaog,vandenberghelh,alviramr,luy,calcedor,zhoux,wilsonjm.cladesofadeno-associatedvirusesarewidelydisseminatedinhumantissues.j.virol.2004jun:78(12):6381-8.(aavrh和aavhu)。
arbetmanae,lochriem,zhous,wellmanj,scallanc,doroudchimm,等人,novelcaprineadeno-associatedvirus(aav)capsid(aav-go.1)iscloselyrelatedtotheprimateaav-5andhasuniquetropismandneutralizationproperties.j.virol.2005:79:15238–15245.(aav-go.1)。
lochriema,tatsunogp,arbetmanae,jonesk,paterc,smithph,等人,adeno-associatedvirus(aav)capsidgenesisolatedfromratandmouselivergenomicdnadefinetwonewaavspeciesdistantlyrelatedtoaav-5.virology.2006:353:68–82.(aav-mo.1)。
schmidtm,katanoh,bossisi,chiorinija.cloningandcharacterizationofabovineadeno-associatedvirus.j.virol.2004:78:6509–6516.(baav)。
bossisi,chiorinija.cloningofanavianadeno-associatedvirus(aaav)andgenerationofrecombinantaaavparticles.j.virol.2003:77:6799–6810.(aaav)。
chencl,jensenrl,schneppbc,connellmj,shellr,sferratj,bartlettjs,clarkkr,johnsonpr.molecularcharacterizationofadeno-associatedvirusesinfectingchildren.j.virol.2005dec:79(23):14781-92.(aav变体)。
send,gadkarira,sudhag,gabrieln,kumarys,selotr,samuelr,rajalingams,ramyav,nairsc,srinivasann,srivastavaa,jayandharangr.targetedmodificationsinadeno-associatedvirusserotype8capsidimprovesitshepaticgenetransferefficiencyinvivo.humgenethermethods.2013apr:24(2):104-16.(aav8k137r)。
lib,maw,lingc,vanvlietk,huangly,agbandje-mckennam,srivastavaa,aslanidigv.site-directedmutagenesisofsurface-exposedlysineresiduesleadstoimprovedtransductionbyaav2,butnotaav8,vectorsinmurinehepatocytesinvivo.humgenethermethods.2015dec:26(6):211-20。
gabrieln,hareendrans,send,gadkarira,sudhag,selotr,hussainm,dhaksnamoorthyr,samuelr,srinivasann,等人,bioengineeringofaav2capsidatspecificserine,threonine,orlysineresiduesimprovesitstransductionefficiencyinvitroandinvivo.humgenethermethods.2013apr:24(2):80-93。
zinne,pacourets,khaychukv,turunenht,carvalhols,andres-mateose,shahs,shelker,maurerac,ploviee,xiaor,vandenberghelh.insilicoreconstructionoftheviralevolutionarylineageyieldsapotentgenetherapyvector.cellrep.2015aug11:12(6):1056-68.(aavanc80l65)。
shens,horowitzed,troupesan,brownsm,pulicherlan,samulskirj,agbandje-mckennam,asokana.engraftmentofagalactosereceptorfootprintontoadeno-associatedviralcapsidsimprovestransductionefficiency.jbiolchem.2013oct4:288(40):28814-23.(aav2g9)。
lic,diprimion,bowlesde,hirschml,monahanpe,asokana,rabinowitzj,agbandje-mckennam,samulskirj.singleaminoacidmodificationofadeno-associatedviruscapsidchangestransductionandhumoralimmuneprofiles.j.virol.2012aug:86(15):7752-9.(aav2265插入-aav2/265d)。
bowlesde,mcpheesw,lic,graysj,samulskijj,campas,lij,wangb,monahanpe,rabinowitzje,等人,phase1genetherapyforduchennemusculardystrophyusingatranslationaloptimizedaavvector.mol.ther.2012feb:20(2):443-55.(aav2.5)。
messinael,nienaberj,daneshmandm,villamizarn,samulskij,milanoc,bowlesde.adeno-associatedviralvectorsbasedonserotype3busecomponentsofthefibroblastgrowthfactorreceptorsignalingcomplexforefficienttransduction.hum.genether.2012oct:23(10):1031-42.(aav3sastg)。
asokana,conwayjc,phillipsjl,lic,heggej,sinnottr,yadavs,diprimion,namhj,agbandje-mckennam,mcphees,wolffj,samulskirj.reengineeringareceptorfootprintofadeno-associatedvirusenablesselectiveandsystemicgenetransfertomuscle.natbiotechnol.2010jan:28(1):79-82.(aav2i8)。
vancem,llangat,bennettw,woodardk,murlidharang,chungfatn,asokana,gilgerb,kurtzbergj,samulskirj,hirschml.aavgenetherapyformps1-associatedcornealblindness.scirep.2016feb22:6:22131.(aav8g9)。
zhongl,lib,mahcs,govindasamyl,agbandje-mckennam,cooperm,herzogrw,zolotukhini,warringtonkhjr,weigel-vanakenka,hobbsja,zolotukhins,muzyczkan,srivastavaa.nextgenerationofadeno-associatedvirus2vectors:pointmutationsintyrosinesleadtohigh-efficiencytransductionatlowerdoses.procnatlacadsciusa.2008jun3:105(22):7827-32.(aav2酪氨酸突变体aav2y-f)。
petrs-silvah,dinculescua,liq,minsh,chiodov,pangjj,zhongl,zolotukhins,srivastavaa,lewinas,hauswirthww.high-efficiencytransductionofthemouseretinabytyrosine-mutantaavserotypevectors.mol.ther.2009mar:17(3):463-71.(aav8y-f和aav9y-f)。
qiaoc,zhangw,yuanz,shinjh,lij,jayandharangr,zhongl,srivastavaa,xiaox,duand.adeno-associatedvirusserotype6capsidtyrosine-to-phenylalaninemutationsimprovegenetransfertoskeletalmuscle.humgenether.2010oct:21(10):1343-8(aav6y-f)。
carlonm,toelenj,vanderperrena,vandenberghelh,reumersv,sbragial,gijsbersr,baekelandtv,himmelreichu,wilsonjm,deprestj,debyserz.efficientgenetransferintothemouselungbyfetalintratrachealinjectionofraav2/6.2.mol.ther.2010dec:18(12):2130-8.(aav6.2)。
pct公开号wo2013158879a1.(赖氨酸突变体)。
在uspto授予的专利和公开申请的文件包装中列出的以下生物序列文件描述可以与现在已知或以后鉴定的野生型衣壳蛋白和/或其他嵌合或变体衣壳蛋白以任何组合并入本发明的aav衣壳中的嵌合或变体衣壳蛋白(出于说明性目的,美国专利申请号11/486,254对应于美国专利申请号11/486,254):11486254.raw、11932017.raw、12172121.raw、12302206.raw、12308959.raw、12679144.raw、13036343.raw、13121532.raw、13172915.raw、13583920.raw、13668120.raw、13673351.raw、13679684.raw、14006954.raw、14149953.raw、14192101.raw、14194538.raw、14225821.raw、14468108.raw、14516544.raw、14603469.raw、14680836.raw、14695644.raw、14878703.raw、14956934.raw、15191357.raw、15284164.raw、15368570.raw、15371188.raw、15493744.raw、15503120.raw、15660906.raw和15675677.raw。
应当理解,可以采用vp1和vp3的任何组合,且当存在时,来自aav血清型的任何组合的vp1.5和vp2,以产生本发明的aav衣壳。例如,来自aav血清型的任何组合的vp1蛋白可以与来自aav血清型的任何组合的vp3蛋白组合,且相应的vp1蛋白可以存在于任何比率的不同血清型中,且相应的vp3蛋白可以存在于任何比率的不同血清型中,且vp1和vp3蛋白可以存在于任何比率的不同血清型中。应进一步理解,当存在时,来自aav血清型的任何组合的vp1.5和/或vp2蛋白可以与来自aav血清型的任何组合的vp1和vp3蛋白组合,且相应的vp1.5蛋白可以存在于任何比率的不同血清型中,且相应的vp2蛋白可以存在于任何比率的不同血清型中,且相应的vp1蛋白可以存在于任何比率的不同血清型中,且相应的vp3蛋白可以存在于任何比率的不同血清型中,且vp1.5和/或vp2蛋白可以与vp1和vp3蛋白组合存在于任何比率的不同血清型中。
例如,相应的病毒蛋白和/或相应的aav血清型可以以任何比率组合,其可以是以下比率:a:b、a:b:c、a:b:c:d、a:b:c:d:e、a:b:c:d:e:f、a:b:c:d:e:f:g、a:b:c:d:e:f:g:h、a:b:c:d:e:f:g:h:i或a:b:c:d:e:f:g:h:i:j,其中a可以是1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、20、25、30、35、40、50、60、70、80、90、100等;b可以是1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、20、25、30、35、40、50、60、70、80、90、100等;c可以是1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、20、25、30、35、40、50、60、70、80、90、100等;d可以是1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、20、25、30、35、40、50、60、70、80、90、100等;e可以是1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、20、25、30、35、40、50、60、70、80、90、100等;f可以是1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、20、25、30、35、40、50、60、70、80、90、100等;g可以是1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、20、25、30、35、40、50、60、70、80、90、100等;h可以是1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、20、25、30、35、40、50、60、70、80、90、100等;i可以是1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、20、25、30、35、40、50、60、70、80、90、100等;且j可以是1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、20、25、30、35、40、50、60、70、80、90、100等。
还应理解,vp1、vp1.5、vp2和/或vp3衣壳蛋白中的任一种可以作为嵌合衣壳蛋白存在于本发明的衣壳中,其呈任何组合,且呈相对于相同蛋白类型和/或相对于不同的衣壳蛋白的任何比率。
在进一步实施方案中,本发明提供了病毒载体,其包含以下,基本上由以下组成和/或由以下组成:(a)本发明的aav衣壳;和(b)包含至少一个末端重复序列的核酸分子,其中所述核酸分子由aav衣壳衣壳化。在一些实施方案中,所述病毒载体可以是aav颗粒。
在一些实施方案中,本发明的病毒载体可具有对骨骼肌、心肌和/或膈肌的全身性或选择性向性。在一些实施方案中,本发明的病毒载体可具有降低的对肝的向性。
本发明进一步提供了组合物,其可以是药物制剂,其包含本发明的衣壳蛋白、衣壳、病毒载体、aav颗粒组合物和/或药物制剂以及药学上可接受的载体。
在一些非限制性实例中,本发明提供了包含三倍轴环4的氨基酸序列中的修饰的aav衣壳蛋白(vp1、vp1.5、vp2和/或vp3)(opie等人,j.viral.77:6995-7006(2003))和包含修饰的aav衣壳蛋白的病毒衣壳和病毒载体。本发明人已经发现,该环中的修饰可以对包含修饰的aav衣壳蛋白的病毒载体赋予一种或多种所需特性,包括但不限于:(i)肝脏的转导减少,(ii)跨内皮细胞的运动增强,(iii)全身转导;(iv)肌肉组织(例如骨骼肌、心肌和/或膈肌)的转导增强,和/或(v)脑组织(例如,神经元)的转导减少。因此,本发明解决了与常规aav载体相关的一些限制。例如,基于aav8和raav9载体的载体对于全身性核酸递送是有吸引力的,因为它们容易穿过内皮细胞屏障;然而,raav8或raav9的全身施用导致大部分载体被递送至肝脏,由此减少其他重要靶组织(诸如骨骼肌)的转导。
在一个实施方案中,修饰的aav衣壳可以由vp1、vp2和/或vp3构成,其通过dna改组以通过定向进化来开发细胞类型特异性载体来产生。用aav进行dna改组通常描述于li,w.等人,mol.ther.16(7):1252–12260(2008),其通过引用并入本文。在一个实施方案中,dna改组可以用于使用来自两种或更多种不同的aav血清型、aav嵌合体或其他aav的衣壳基因的dna序列来产生vp1、vp2和/或vp3。在一个实施方案中,单倍体aav可以由通过dna改组产生的vp1、通过dna改组产生的vp2和/或通过dna改组产生的vp3构成。
在一个实施方案中,来自单倍体aav的vp1可以通过使用限制酶和/或dna酶随机断裂aav2、aav8和aav9的衣壳基因组以生成由aav2/8/9的部分构成的vp1衣壳蛋白文库来产生。在该实施方案中,通过dna改组产生的aav2/8/9vp1衣壳蛋白可以与来自不同血清型(在一个实施方案中,来自aav3)的vp2和/或vp3蛋白组合。这将导致单倍体aav,其中衣壳由vp1、vp2和/或vp3构成,所述vp1包括通过dna改组随机连接在一起的来自aav2、aav8和aav9的氨基酸,且vp2和/或vp3仅包含来自天然aav3vp2和/或vp3的氨基酸。在一个实施方案中,用于产生vp1、vp2和/或vp3的供体可以是任何aav,包括aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10、aav11、aav嵌合体或其他aav,或选自表1或表3的那些。在某些实施方案中,改组的vp1表达仅vp1,或仅vp1/vp2,或仅vp3。
在另一个实施方案中,编码vp1、vp2和/或vp3的核酸可以通过dna改组产生。在一个实施方案中,通过dna改组产生的第一核酸将编码vp1。在该相同实施方案中,通过dna改组产生的第二核酸将编码vp2和vp3。在另一个实施方案中,通过dna改组产生的第一核酸将编码vp1。在该相同实施方案中,通过dna改组产生的第二核酸将编码vp2,且第三核酸将编码vp3。在一个进一步实施方案中,通过dna改组产生的第一核酸将编码vp1和vp2,且通过dna改组产生的第二核酸将编码vp3。在一个额外实施方案中,通过dna改组产生的第一核酸将编码vp1和vp3,且通过dna改组产生的第二核酸将编码vp2。
在本发明的实施方案中,心肌和/或骨骼肌的转导(基于个别骨骼肌、多种骨骼肌或整个范围的骨骼肌确定)是肝脏中的转导水平的至少约五倍、十倍、50倍、100倍、1000倍或更高倍。
在具体实施方案中,本发明的修饰的aav衣壳蛋白在三倍轴环4的氨基酸序列(例如,天然aav2vp1衣壳蛋白的氨基酸位置575至600[包括端值]或来自另一种aav的衣壳蛋白的相应区域)中包含一个或多个修饰。如本文所用,氨基酸序列中的“修饰”包括取代、插入和/或缺失,其各自可以涉及1、2、3、4、5、6、7、8、9、10或更多个氨基酸。在具体实施方案中,所述修饰是取代。例如,在具体实施方案中,来自一种aav的三倍轴环4的1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25或更多个氨基酸可被取代至天然aav2衣壳蛋白的氨基酸位置575-600或来自另一种aav的衣壳蛋白的相应位置。然而,本发明的修饰的病毒衣壳不限于aav衣壳,其中来自一种aav衣壳的氨基酸被取代至另一种aav衣壳中,并且取代和/或插入的氨基酸可以来自任何来源,并且可以进一步是天然存在的或部分或完全合成的。
如本文所述,来自许多aav的衣壳蛋白的核酸和氨基酸序列是本领域已知的。因此,对于任何其他aav,可以容易地确定“对应”于天然aav2衣壳蛋白的氨基酸位置575至600(包括端值)或氨基酸位置585至590(包括端值)的氨基酸(例如,通过使用序列比对)。
在一些实施方案中,本发明考虑,本发明的修饰的衣壳蛋白可以通过修饰现在已知或以后发现的任何aav的衣壳蛋白来生产。进一步地,待修饰的aav衣壳蛋白可以是天然存在的aav衣壳蛋白(例如aav2、aav3a或3b、aav4、aav5、aav8、aav9、aav10、aav11或aav12衣壳蛋白或表3中所示的任何aav),但不限于此。本领域技术人员将理解,对aav衣壳蛋白的各种操作是本领域已知的,并且本发明不限于天然存在的aav衣壳蛋白的修饰。例如,待修饰的衣壳蛋白与天然存在的aav相比可能已经具有改变(例如,衍生自天然存在的aav衣壳蛋白,例如aav2、aav3a、aav3b、aav4、aav5、aav6、aav7、aav8、aav9、aav10、aav11和/或aav12或现在已知或后来发现的任何其他aav)。此类aav衣壳蛋白也在本发明的范围内。
例如,在一些实施方案中,待修饰的aav衣壳蛋白可包含直接在天然aav2衣壳蛋白序列的氨基酸264之后的氨基酸插入(参见,例如,pct公开wo2006/066066)和/或可以是如pct公开wo2009/108274中所述的具有改变的hi环的aav和/或可以是经修饰以含有多聚-his序列以便于纯化的aav。作为另一个说明性实例,aav衣壳蛋白可具有作为插入或取代并入其中的肽靶向序列。进一步,aav衣壳蛋白可包含已被取代和/或插入至衣壳蛋白中的来自另一种aav的大结构域。
因此,在具体实施方案中,待修饰的aav衣壳蛋白可以衍生自天然存在的aav,但进一步包含被插入和/或取代至衣壳蛋白中和/或已经通过缺失一个或多个氨基酸而被改变的一个或多个外源序列(例如,对于天然病毒是外源的)。
因此,当在本文中提及特定aav衣壳蛋白(例如,aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10、aav11或aav12衣壳蛋白或来自表1中所示的任何aav的衣壳蛋白,等)时,其意图涵盖天然衣壳蛋白以及除了本发明的修饰以外具有改变的衣壳蛋白。此类改变包括取代、插入和/或缺失。在具体实施方案中,与天然aav衣壳蛋白序列相比,所述衣壳蛋白包含(除了本发明的插入以外)其中插入的1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20个、少于20个、少于30个、少于40个、少于50个、少于60个或少于70个氨基酸。在本发明的实施方案中,与天然aav衣壳蛋白序列相比,所述衣壳蛋白包含(除了根据本发明的氨基酸取代以外)1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20个、少于20个、少于30个、少于40个、少于50个、少于60个或少于70个氨基酸取代。在本发明的实施方案中,与天然aav衣壳蛋白序列相比,所述衣壳蛋白包含(除了本发明的氨基酸缺失以外)1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19或20个、多于20个、多于30个、多于40个、多于50个、多于60个或多于70个氨基酸的缺失。
使用aav血清型2作为示例性病毒,m11是vp1起始密码子,m138是vp2起始密码子,且m203是vp3起始密码子。尽管通常通过取代m11和m138的起始密码子的缺失将使vp1和vp2的表达不起作用,但vp3起始密码子的类似缺失是不够的。这是因为病毒衣壳orf含有许多具有不同强度的atg密码子作为起始密码子。因此,在设计不表达vp3的构建体中,必须小心以确保不产生替代的vp3种类。对于vp3,m138的消除是必需的,或如果需要vp2,但不是vp3,则除了m203以外,m211和235的缺失通常是最好的方法(warrington,k.h.jr.,等人,j.ofvirol.78(12):6595-6609(june2004))。这可以通过突变诸如取代或本领域已知的其他方式来进行。可以容易地确定其他血清型中相应的起始密码子,以及诸如vp3中的额外atg序列是否可以充当替代起始密码子。
因此,例如,术语“aav2衣壳蛋白”包括具有天然aav2衣壳蛋白序列(参见genbank登录号aac03780)的aav衣壳蛋白以及在天然aav2衣壳蛋白序列中包含取代、插入和/或缺失的那些(如前述段落中所述)。
在具体实施方案中,所述aav衣壳蛋白具有天然aav衣壳蛋白序列或与天然aav衣壳蛋白序列具有至少约70%、75%、80%、85%、90%、95%、97%、98%或99%相似性或同一性的氨基酸序列。例如,在具体实施方案中,“aav2”衣壳蛋白涵盖天然aav2衣壳蛋白序列以及与天然aav2衣壳蛋白序列具有至少约75%、80%、85%、90%、95%、97%、98%或99%相似性或同一性的序列。
确定两个或更多个氨基酸序列之间的序列相似性或同一性的方法是本领域已知的。序列相似性或同一性可以使用本领域已知的标准技术来确定,所述标准技术包括但不限于smith&waterman,adv.appl.math.2,482(1981)的局部序列同一性算法,needleman&wunsch,j.mol.biol.48,443(1970)的序列同一性比对算法,pearson&lipman,proc.natl.acad.sci.usa85,2444(1988)的相似性搜索方法,这些算法的计算机化实施(wisconsingenetics软件包,geneticscomputergroup,575sciencedrive,madison,wi中的gap、bestfit、fasta和tfasta),由devereux等人,nucl.acidres.12,387-395(1984)描述的bestfit序列程序,或通过检查。
另一种合适的算法是blast算法,其描述于altschul等人,j.mol.biol.215,403-410,(1990)和karlin等人,proc.natl.acad.sci.usa90,5873-5787(1993)。一种特别有用的blast程序是从altschul等人,methodsinenzymology,266,460-480(1996);http://blast.wustl/edu/blast/readme.html获得的wu-blast-2程序。wu-blast-2使用几个搜索参数,其可以任选地被设置为默认值。所述参数是动态值,并且由程序本身根据特定序列的组成和特定数据库(针对所述特定数据库搜索目标序列)的组成来确定;然而,可以调整所述值以增加灵敏度。
进一步地,额外有用的算法是如altschul等人,(1997)nucleicacidsres.25,3389-3402所报道的空位blast(gappedblast)。
在本发明的一些实施方案中,可以在天然aav2衣壳蛋白的氨基酸位置585至590(包括端值)(使用vp1编号)或其他aav的相应位置(天然aav2vp1衣壳蛋白:genbank登录号aac03780或yp680426),即对应于天然aav2衣壳蛋白的氨基酸位置585至590(vp1编号)的氨基酸处进行修饰。“对应于”天然aav2衣壳蛋白的位置585至590的其他aav血清型或修饰的aav衣壳中的氨基酸位置对于本领域技术人员而言是显而易见的,并且可以使用序列比对技术(参见,例如,wo2006/066066的图7)和/或晶体结构分析(padron等人,(2005)j.virol.79:5047-58)容易地确定。
为了举例说明,可以将修饰引入已经含有插入和/或缺失、使得所有下游序列的位置均被移位的aav衣壳蛋白中。在该情况下,对应于aav2衣壳蛋白中的氨基酸位置585至590的氨基酸位置对于本领域技术人员仍将是容易鉴定的。为了举例说明,衣壳蛋白可以是aav2衣壳蛋白,其含有在氨基酸位置264之后的插入(参见,例如,wo2006/066066)。在位置585至590发现的氨基酸(例如,天然aav2衣壳蛋白中的rgnrqa(seqidno:1))将在位置586至591处,但是对于本领域技术人员来说仍然是可以鉴定的。
本发明还提供了包含本发明的修饰的aav衣壳蛋白、基本上由其组成或由其组成的病毒衣壳。在具体实施方案中,所述病毒衣壳是细小病毒衣壳,其可以进一步是自主性细小病毒衣壳或依赖病毒衣壳。任选地,所述病毒衣壳是aav衣壳。在具体实施方案中,所述aav衣壳是aav1、aav2、aav3a、aav3b、aav4、aav5、aav6、aav7、aav8、aav9、aav10、aav11、aav12或表1中所示或另外已知或后来发现的任何其他aav或通过一个或多个插入、取代和/或缺失而衍生自前述任一种。
在本发明的实施方案中,分离的aav病毒粒子或基本上均质的aav病毒粒子不是表达一种血清型的vp1、vp2和vp3的一种核酸辅助质粒与表达另一种血清型的vp1、vp2和vp3的另一种核酸辅助质粒的混合物的表达产物,这种表达称为“交叉装饰(cross-dressing)”。
在本发明的实施方案中,分离的aav病毒粒子不包含镶嵌衣壳,且aav病毒粒子的基本上均质的群体不包含镶嵌衣壳的基本上均质的群体。
在2018年3月15日提交的pct/us18/22725中的任何公开内容落入如本申请的权利要求中的任何一项或多项中所定义的发明内或在将来可在本申请或由其衍生的任何专利中提交的修改的权利要求所定义的任何发明内的程度上,以及在该或那些权利要求提交的任何一个或多个相关国家的法律提供了pct/us18/22725的公开内容是针对该或那些权利要求针对或该或那些国家的现有技术的部分的程度上,我们在此在防止本申请或由其衍生的任何专利无效的程度上保留将所述公开内容从本申请或由其衍生的任何专利的权利要求免于请求保护的权利。
例如,且非限制性地,我们保留将以下主题中的任何一项或多项从现在或将来修改的本申请的任何权利要求或由其衍生的任何专利免于请求保护的权利:
a.pct/us18/22725的实施例9中公开的任何主题;或
b.载体病毒粒子,称为多倍体载体病毒粒子,其通过转染两种aav辅助质粒或三种质粒以产生由来自不同血清型的不同衣壳亚基构成的个别多倍体载体病毒粒子而产生或可产生;或
c.载体病毒粒子,称为多倍体载体病毒粒子,其通过转染作为aav2和aav8或aav9的两种aav辅助质粒以产生由来自不同血清型的不同衣壳亚基构成的个别多倍体载体病毒粒子而产生或可产生;或
d.载体病毒粒子,称为多倍体载体病毒粒子,其通过转染作为aav2、aav8和aav9的三种aav辅助质粒以产生由来自不同血清型的不同衣壳亚基构成的个别多倍体载体病毒粒子而产生或可产生;或
e.载体病毒粒子,称为单倍体载体,其具有来自一种aav载体衣壳或aav血清型的vp1/vp2和来自替代aav载体衣壳或aav血清型的vp3,例如来自仅一种aav血清型(的衣壳)的vp1/vp2和来自仅一种替代aav血清型的vp3;或
f.选自以下的任何一种或多种aav载体病毒粒子:
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav8的vp1衣壳亚基和来自aav2的vp2/vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8或单倍体aav8/2或单倍体aav82或h-aav82),且具有来自aav8的vp1/vp2衣壳亚基和来自aav2的vp3衣壳亚基;或
载体,其中vp1/vp2衍生自不同血清型;或
载体(称为单倍体aav92或h-aav92),其具有来自aav9的vp1/vp2衣壳亚基和来自aav2的vp3衣壳亚基;或
载体(称为单倍体aav2g9或h-aav2g9),其具有来自aav8的vp1/vp2衣壳亚基和来自aav2g9的vp3衣壳亚基,其中将aav9聚糖受体结合位点移植至aav2中;或
载体(称为单倍体aav83或h-aav83),其具有来自aav8的vp1/vp2衣壳亚基和来自aav3的vp3衣壳亚基;或
载体(称为单倍体aav93或h-aav93),其具有来自aav9的vp1/vp2衣壳亚基和来自aav3的vp3衣壳亚基;或
载体(称为单倍体aavrh10-3或h-aavrh10-3),其具有来自aavrh10的vp1/vp2衣壳亚基和来自aav3的vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav2的vp1衣壳亚基和来自aav8的vp2/vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav2的vp1/vp2衣壳亚基和来自aav8的vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav8的vp1衣壳亚基和来自aav2的vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav2的vp1衣壳亚基和来自aav8的vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav2的vp1/vp2/vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav8的vp1/vp2/vp3衣壳亚基;或
称为28m-2vp3或单倍体2m-2vp3或单倍体载体28m-2vp3的载体,其中嵌合vp1/vp2衣壳亚基具有来自aav2的n-末端和来自aav8的c-末端,且vp3衣壳亚基来自aav2;或
称为嵌合aav8/2或嵌合aav82的载体,其中嵌合vp1/vp2衣壳亚基具有来自aav8的n-末端和来自aav2的c-末端且没有vp3起始密码子的突变,且vp3衣壳亚基来自aav2;或
载体,其中嵌合vp1/vp2衣壳亚基具有来自aav2的n-末端和来自aav8的c-末端;或
g.f的载体中任一种的群体,例如基本上均质的群体,例如1010个颗粒的群体,例如1010个颗粒的基本上均质的群体;或
h.产生a和/或b和/或c和/或d和/或e和/或f和/或g的载体或载体群体中任一种的方法;或
i.其任何组合。
在没有限制的情况下,我们声明,上述免于请求保护的权利的保留至少适用于本申请所附的权利要求1-30和[00437]中所述的段落1-83。修饰的病毒衣壳可用作“衣壳媒介物”,如例如在美国专利号5,863,541中所述。可被修饰的病毒衣壳包装并转移到细胞中的分子包括异源dna、rna、多肽、有机小分子、金属或其组合。
异源分子被定义为并未在aav感染中天然发现的那些,例如,未被野生型aav基因组编码的那些。进一步地,治疗有用的分子可以与病毒衣壳外面缔合,用于将分子转移至宿主靶细胞中。此类缔合的分子可以包括dna、rna、小有机分子、金属、碳水化合物、脂质和/或多肽。在本发明的一个实施方案中,将所述治疗有用的分子与衣壳蛋白共价连接(即缀合或化学偶联)。将分子共价连接的方法是本领域技术人员已知的。
本发明的修饰的病毒衣壳在产生针对新的衣壳结构的抗体中也找到用途。作为进一步的替代方案,可将外源氨基酸序列插入修饰的病毒衣壳,用于将抗原呈递给细胞,例如用于施用于受试者以产生对外源氨基酸序列的免疫应答。
在其他实施方案中,可以在施用递送编码目标多肽或功能性rna的核酸的病毒载体之前和/或同时(例如,在彼此的数分钟或数小时内)施用病毒衣壳以阻断某些细胞位点。例如,可以递送本发明的衣壳以阻断肝细胞上的细胞受体,并且可以随后或同时施用递送载体,这可以减少肝细胞的转导,并且增强其他靶标(例如骨骼、心脏和/或膈肌)的转导。
根据代表性实施方案,可以在根据本发明的修饰的病毒载体之前和/或同时将修饰的病毒衣壳施用于受试者。进一步地,本发明提供了包含本发明的修饰的病毒衣壳的组合物和药物制剂;任选地,所述组合物还包含本发明的修饰的病毒载体。
本发明也提供编码本发明的修饰的病毒衣壳和衣壳蛋白的核酸分子(任选地,分离的核酸分子)。进一步提供的是包含所述核酸分子的载体和包含本发明的核酸分子和/或载体的细胞(在体内或在培养中)。合适的载体包括但不限于病毒载体(例如腺病毒、aav、疱疹病毒、甲病毒、牛痘、痘病毒、杆状病毒等)、质粒、噬菌体、yac、bac等。此类核酸分子、载体和细胞可用作例如用于产生如本文所述的修饰的病毒衣壳或病毒载体的试剂(例如,辅助包装构建体或包装细胞)。
可以使用本领域已知的任何方法,例如通过从杆状病毒表达(brown等人,(1994)virology198:477-488)来产生根据本发明的病毒衣壳。
在一些实施方案中,对本发明的aav衣壳蛋白的修饰是“选择性”修饰。该方法与用aav血清型之间的整个亚基或大结构域交换的先前工作形成对比(参见例如国际专利公开wo00/28004和hauck等人,(2003)j.virology77:2768-2774)。在具体实施方案中,“选择性”修饰导致少于约20、18、15、12、10、9、8、7、6、5、4、3或2个连续氨基酸的插入和/或取代和/或缺失。
本发明的修饰的衣壳蛋白和衣壳可以进一步包含现在已知或后来鉴定的任何其他修饰。
所述病毒衣壳可以是靶向的病毒衣壳,其包含指导病毒衣壳与存在于期望靶组织上的细胞-表面分子相互作用的靶向序列(例如,取代或插入病毒衣壳中)(参见例如,国际专利公开号wo00/28004和hauck等人,(2003)j.virology77:2768-2774);shi等人,humangenetherapy17:353-361(2006)[描述在aav衣壳亚基的位置520和/或584插入整联蛋白受体结合基序rgd];和美国专利号7,314,912[描述在aav2衣壳亚基的氨基酸位置447、534、573和587之后插入含有rgd基序的p1肽])。aav衣壳亚基内耐受插入的其他位置是本领域已知的(例如,grifman等人,moleculartherapy3:964-975(2001)描述的位置449和588)。
例如,本发明的一些病毒衣壳对于大多数目标靶组织(例如肝、骨骼肌、心脏、膈肌、肾、脑、胃、肠、皮肤、内皮细胞和/或肺)具有相对低效的向性。可以将靶向序列有利地并入这些低转导载体中,由此为病毒衣壳赋予期望的向性和任选对特定组织的选择性向性。包含靶向序列的aav衣壳蛋白、衣壳和载体例如描述于国际专利公开wo00/28004中。作为另一种可能性,如wang等人,annurevbiophysbiomolstruct.35:225-49(2006))描述的一个或多个天然存在的氨基酸可以作为将低转导载体重新指导至期望的靶组织的方式在正交位点处并入aav衣壳亚基中。这些非天然氨基酸可以有利地用于将目标分子化学连接至aav衣壳蛋白,包括但不限于:聚糖(甘露糖-树突细胞靶向);用于靶向递送至特定癌细胞类型的rgd、铃蟾肽或神经肽;靶向至特定细胞表面受体诸如生长因子受体、整联蛋白等的选自噬菌体展示的rna适体或肽。化学修饰氨基酸的方法是本领域已知的(参见例如,gregt.hermanson,bioconjugatetechniques,第1版,academicpress,1996)。
在代表性实施方案中,所述靶向序列可以是指导感染至特定细胞类型的病毒衣壳序列(例如,自主性细小病毒衣壳序列、aav衣壳序列或任何其他病毒衣壳序列)。
作为另一个非限制性实例,可以将肝素结合结构域(例如,呼吸道合胞病毒肝素结合结构域)插入或取代至通常不结合hs受体的衣壳亚基(例如aav4、aav5)以便为所得突变体赋予肝素结合。
b19使用红细胞糖苷酯作为其受体来感染原代红细胞祖细胞(brown等人,(1993)science262:114)。b19的结构已被确定为8å分辨率(agbandje-mckenna等人,(1994)virology203:106)。结合红细胞糖苷酯的b19衣壳的区域已经被定位在氨基酸399-406之间(chapman等人,(1993)virology194:419),环出区域已经被定位在β-桶状结构e和f之间(chipman等人,(1996)proc.nat.acad.sci.usa93:7502)。因此,b19衣壳的红细胞糖苷酯受体结合结构域可被取代至aav衣壳蛋白中,以将病毒衣壳或包含其的病毒载体靶向至红细胞样细胞。
在代表性实施方案中,所述外源靶向序列可以是编码改变包含修饰的aav衣壳蛋白的病毒衣壳或病毒载体的向性的肽的任何氨基酸序列。在具体实施方案中,所述靶向肽或蛋白可以是天然存在的,或者可替代地是完全或部分合成的。示例性靶向序列包括结合细胞表面受体和糖蛋白的配体和其他肽,诸如rgd肽序列、缓激肽、激素、肽生长因子(例如,表皮生长因子、神经生长因子、成纤维细胞生长因子、血小板衍生生长因子、胰岛素样生长因子i和ii等)、细胞因子、黑素细胞激素(例如α、β或γ)、神经肽和内啡肽等,以及其保留将细胞靶向至其同源受体的能力的片段。其他说明性的肽和蛋白包括p物质、角质形成细胞生长因子、神经肽y、胃泌素释放肽、白介素2、鸡蛋清溶菌酶、促红细胞生成素、促性腺激素释放激素、皮质抑素、β-内啡肽、亮内啡肽、强腓肽b、α-新-内啡肽、血管紧张素、肺炎球菌素、血管活性肠肽、神经降压素、胃动素及其片段,如上所述。作为又一个进一步替代实施方案,来自毒素(例如破伤风毒素或蛇毒素,诸如α-银环蛇毒素等)的结合结构域可以作为靶向序列被取代至衣壳蛋白中。在又另一个代表性实施方案中,aav衣壳蛋白可以通过将如cleves(currentbiology7:r318(1997))所述的“非经典”输入/输出信号肽(例如,成纤维细胞生长因子-1和-2、白介素1、hiv-1tat蛋白、疱疹病毒vp22蛋白等)取代至aav衣壳蛋白中来修饰。还涵盖指导特定细胞摄取的肽基序,例如fvflp肽基序触发肝细胞摄取。
噬菌体展示技术以及本领域已知的其他技术可用于鉴定识别任何目标细胞类型的肽。
所述靶向序列可以编码靶向至细胞表面结合位点、包括受体(例如,蛋白、碳水化合物、糖蛋白或蛋白聚糖)的任何肽。细胞表面结合位点的实例包括但不限于硫酸乙酰肝素、硫酸软骨素和其他糖胺聚糖、粘蛋白上发现的唾液酸部分、糖蛋白和神经节苷脂、mhci糖蛋白、膜糖蛋白上发现的碳水化合物组分,包括甘露糖、n-乙酰基-半乳糖胺、n-乙酰基-葡糖胺、岩藻糖、半乳糖等。
在具体实施方案中,将硫酸乙酰肝素(hs)或肝素结合结构域取代至病毒衣壳中(例如,在否则不结合hs或肝素的aav中)。本领域已知hs/肝素结合由富含精氨酸和/或赖氨酸的“碱性贴剂”介导。在示例性实施方案中,序列遵循基序bxxb,其中“b”是碱性残基并且x是中性和/或疏水性的。作为一个非限制性实例,bxxb是rgnr。在具体实施方案中,用bxxb取代天然aav2衣壳蛋白中的氨基酸位置262至265或另一种aav的衣壳蛋白中的对应位置。
合适的靶向序列的其他非限制性实例包括由müller等人,naturebiotechnology21:1040-1046(2003)鉴定的靶向冠状动脉内皮细胞的肽(共有序列nsvrdlg/s(seqidno:2)、prsvtvp(seqidno:3)、nsvssxs/a(seqidno:4));由grifman等人,moleculartherapy3:964-975(2001)所述的肿瘤靶向肽(例如,ngr、ngraha(seqidno:5));由work等人,moleculartherapy13:683-693(2006)所述的肺或脑靶向序列(qpehsst(seqidno:6)、vntanst(seqidno:7)、hgpmqks(seqidno:8)、phkppla(seqidno:9)、iknnemw(seqidno:10)、rnldtpm(seqidno:11)、vdshrqs(seqidno:12)、ydsktkt(seqidno:13)、sqlphqk(seqidno:14)、stmqqnt(seqidno:15)、terymtq(seqidno:16)、qpehsst(seqidno:6)、daslsts(seqidno:17)、dlpnkkt(seqidno:18)、dltaarl(seqidno:19)、ephqfny(seqidno:20)、epqsnht(seqidno:21)、msswpsq(seqidno:22)、npkhnat(seqidno:23)、pdgmrtt(seqidno:24)、pnnnktt(seqidno:25)、qstthds(seqidno:26)、tgskqkq(seqidno:27)、slkhqal(seqidno:28)和spidgeq(seqidno:29));由hajitou等人,tcm16:80-88(2006)描述的血管靶向序列(wifpwiql(seqidno:30)、cdcrgdcfc(seqidno:31)、cngrc(seqidno:32)、cpreces(seqidno:33)、gsl、ctthwgftlc(seqidno:34)、cgrraggsc(seqidno:35)、ckggrakdc(seqidno:36)和cvpelghec(seqidno:37));由koivunen等人,j.nucl.med.40:883-888(1999)所述的靶向肽(crretawak(seqidno:38)、kgd、vswfshryspfavs(seqidno:39)、gyrdgyagpilyn(seqidno:40)、xxxy*xxx[其中y*是磷酸化-tyr](seqidno:41)、y*e/mnw(seqidno:42)、rplpplp(seqidno:43)、applppr(seqidno:44)、dvfypypyasgs(seqidno:45)、mywypy(seqidno:46)、ditwdqlwdlmk(seqidno:47)、cwddg/lwlc(seqidno:48)、ewceylggylrcya(seqidno:49)、yxcxxgpxtwxcxp(seqidno:50)、iegptlrqwlaara(seqidno:51)、lwxxy/w/f/h(seqidno:52)、xfxxylw(seqidno:53)、ssiishfrwglcd(seqidno:54)、msrpacppndkye(seqidno:55)、clrsgrgc(seqidno:56)、chwmfspwc(seqidno:57)、wxxf(seqidno:58)、cssrldac(seqidno:59)、clpvasc(seqidno:60)、cgfecvrqcperc(seqidno:61)、cvalcreacgegc(seqidno:62)、swcepgwcr(seqidno:63)、ysgkwgw(seqidno:64)、glsggrs(seqidno:65)、lmlprad(seqidno:66)、cscfrdvcc(seqidno:67)、crdvvsvic(seqidno:68)、cngrc(seqidno:32)和gsl);和由newton&deutscher,phagepeptidedisplayinhandbookofexperimentalpharmacology,第145-163页,springer-verlag,berlin(2008)所述的肿瘤靶向肽(marsgl(seqidno:69)、marake(seqidno:70)、msrtms(seqidno:71)、kccysl(seqidno:72)、wrr、wkr、wvr、wvk、wik、wtr、wvl、wll、wrt、wrg、wvs、wva、mywgdshwlqywye(seqidno:73)、mqlplat(seqidno:74)、ewls(seqidno:75)、snew(seqidno:76)、tnyl(seqidno:77)、wifpwiql(seqidno:30)、wdlawmfrlpvg(seqidno:78)、ctvalpggyvrvc(seqidno:79)、cvpelghec(seqidno:37)、cgrraggsc(seqidno:35)、cvayciehhcwtc(seqidno:80)、cvfahnydylvc(seqidno:81)和cvftsnyafc(seqidno:82)、vhspnkk(seqidno:83)、cdcrgdcfc(seqidno:31)、crgdgwc(seqidno:84)、xrgcdx(seqidno:85)、p:xxs/t(seqidno:86)、ctthwgftlc(seqidno:34)、sgkgprqital(seqidno:87)、a9a/q)(n/a)(l/y)(tn/m/r)(r/k)(seqidno:88)、vymspf(seqidno:89)、mqlplat(seqidno:74)、atwlppr(seqidno:90)、htmyyhhyqhhl(seqidno:91)、sevgcragplqwlcekyfg(seqidno:92)、cgllpvgrpdrnvwrwlc(seqidno:93)、ckgqcdrfkglpwec(seqidno:94)、sgrsa(seqidno:95)、wgfp(seqidno:96)、lwxxar[ar=y、w、f、h)(seqidno:97)、xf:xxylw(seqidno:98)、aepmphslnfsqylwyt(seqidno:99)、way(w/f)sp(seqidno:100)、iellqar(seqidno:101)、ditwdqlwdlmk(seqidno:102)、aytkcsrqwrtcmtth(seqidno:103)、pqnskipgptfldph(seqidno:104)、smepalpdwwwkmfk(seqidno:105)、antpcgpythdcpvkr(seqidno:106)、tachqhvrmvrp(seqidno:107)、vpwmepayqrfl(seqidno:108)、dpratpgs(seqidno:109)、frpnraqdyntn(seqidno:110)、ctknsylmc(seqidno:111)、c(r/q)l/rt(g/n)xxg(an)gc(seqidno:112)、cpiedrpmc(seqidno:113)、hewsylapypwf(seqidno:114)、mcpkhplgc(seqidno:115)、rmwpsstvnlsagrr(seqidno:116)、saktavsqrvwlpshrggep(seqidno:117)、ksrehvnnsacpskritaal(seqidno:118)、egfr(seqidno:119)、rvs、ags、aglgvr(seqidno:120)、ggr、ggl、gsv、gvs、gtrqghtmrlgvsdg(seqidno:121)、iaglatpgwshwlal(seqidno:122)、smsiarl(seqidno:123)、htfepgv(seqidno:124)、ntslkrisnkrirrk(seqidno:125)、lrikrkrrkrkktrk(seqidno:126)、ggg、gfs、lws、egg、llv、lsp、lbs、agg、grr、ggh和gtv)。
作为又一个进一步替代方案,所述靶向序列可以是可以用于与另一种靶向进入细胞的分子化学偶联的肽(例如,可以包含可以通过它们的r基团化学偶联的精氨酸和/或赖氨酸残基)。
作为另一个选项,本发明的aav衣壳蛋白或病毒衣壳可以包含如wo2006/066066中所述的突变。例如,所述衣壳蛋白可以包含天然aav2衣壳蛋白的氨基酸位置263、705、708和/或716处的选择性氨基酸取代或来自另一种aav的衣壳蛋白中的相应改变。另外或可替代地,在代表性实施方案中,衣壳蛋白、病毒衣壳或载体包含直接在aav2衣壳蛋白的氨基酸位置264之后的选择性氨基酸插入或来自其他aav的衣壳蛋白的相应改变。“直接在氨基酸位置x之后”意指插入紧紧在指定氨基酸位置之后(例如,“氨基酸位置264之后”指示在位置265处的点插入或更大的插入,例如从位置265至268等)。本发明的前述实施方案可用于将异源核酸递送至细胞或受试者,如本文所述。例如,修饰的载体可用于治疗如本文所述的溶酶体贮积症诸如粘多糖贮积症(例如斯莱综合征[β-葡糖醛酸酶]、胡尔勒综合征[α-l-艾杜糖醛酸酶]、沙伊综合征[α-l-艾杜糖醛酸酶]、胡-射二氏(hurler-scheie)综合征[α-l-艾杜糖醛酸酶]、亨特氏综合征[艾杜糖醛酸硫酸酯酶]、圣菲利波综合征a[乙酰肝素磺酰胺酶]、b[n-乙酰氨基葡糖苷酶]、c[乙酰-coa:α-氨基葡糖苷乙酰基转移酶]、d[n-乙酰氨基葡糖-6-硫酸酯酶]、莫尔基奥氏综合征a[半乳糖-6-硫酸盐硫酸酯酶]、b[β-半乳糖苷酶]、马-兰二氏(maroteaux-lamy)综合征[n-乙酰氨基半乳糖-4-硫酸酯酶]等)、法布里病(α-半乳糖苷酶)、戈谢病(葡糖脑苷脂酶)、或糖原贮积症(例如庞贝氏症;溶酶体酸性α-葡糖苷酶)。
本领域技术人员将理解,对于一些aav衣壳蛋白,相应的修饰将是插入和/或取代,这取决于相应的氨基酸位置是否部分或完全存在于病毒中,或者可替代地完全不存在。同样,当修饰除了aav2以外的aav时,特定氨基酸位置可能不同于aav2中的位置(参见例如,表3)。如本文别处所讨论,使用众所周知的技术,相应的氨基酸位置对于本领域技术人员将是显而易见的。
在代表性实施方案中,衣壳蛋白中的插入和/或取代和/或缺失导致如下氨基酸的插入、取代和/或重新定位:(i)维持该区域中的亲水性环结构的氨基酸;(ii)改变环结构的构型的氨基酸;(iii)带电荷的氨基酸;和/或(iv)可以被磷酸化或硫酸化或者以其他方式通过在aav2衣壳蛋白中的264位后的翻译后修饰(例如糖基化)或另一种aav的衣壳蛋白中的相应变化而获得电荷的氨基酸。用于插入/取代的合适氨基酸包括天冬氨酸、谷氨酸、缬氨酸、亮氨酸、赖氨酸、精氨酸、苏氨酸、丝氨酸、酪氨酸、甘氨酸、丙氨酸、脯氨酸、天冬酰胺、苯丙氨酸、酪氨酸或谷氨酰胺。在具体实施方案中,将苏氨酸插入或取代入衣壳亚基中。许多其他aav中对应位置的非限制性实例显示于表3(位置2)中。在具体实施方案中,氨基酸插入或取代是苏氨酸、天冬氨酸、谷氨酸或苯丙氨酸(除了在该位置分别具有苏氨酸、谷氨酸或苯丙氨酸的aav以外)。
根据本发明的该方面,在一些实施方案中,aav衣壳蛋白在aav2、aav3a或aav3b衣壳蛋白中的氨基酸位置264之后或在已经修饰以分别包含非aav2、aav3a或aav3b序列和/或已通过缺失一个或多个氨基酸而修饰(即,衍生自aav2、aav3a或aav3b)的aav2、aav3a或aav3b衣壳蛋白中的相应位置中包含氨基酸插入。对应于aav2(或aav3a或aav3b)衣壳亚基中的位置264的氨基酸将容易在已衍生自aav2(或aav3a或aav3b)的起始病毒中可鉴定,其然后可以根据本发明进一步修饰。用于插入的合适氨基酸包括天冬氨酸、谷氨酸、缬氨酸、亮氨酸、赖氨酸、精氨酸、苏氨酸、丝氨酸、酪氨酸、甘氨酸、丙氨酸、脯氨酸、天冬酰胺、苯丙氨酸、酪氨酸或谷氨酰胺。
在其他实施方案中,aav衣壳蛋白包含在aav1衣壳蛋白中的氨基酸位置265处、在aav8衣壳蛋白中的氨基酸位置266处的氨基酸取代,或在aav9衣壳蛋白中的氨基酸位置265处或在已经修饰以分别包含非aav1、非aav8或非aav9序列和/或已通过缺失一个或多个氨基酸而修饰(即,衍生自aav1、aav8或aav9)的aav1、aav8或aav9衣壳蛋白中的相应位置中的氨基酸取代。对应于aav1和aav9衣壳亚基中的位置265或aav8衣壳亚基中的位置266的氨基酸将容易在已衍生自aav1、aav8或aav9的起始病毒中可鉴定,其然后可以根据本发明进一步修饰。用于插入的合适氨基酸包括天冬氨酸、谷氨酸、缬氨酸、亮氨酸、赖氨酸、精氨酸、苏氨酸、丝氨酸、酪氨酸、甘氨酸、丙氨酸、脯氨酸、天冬酰胺、苯丙氨酸、酪氨酸或谷氨酰胺。
在本发明的代表性实施方案中,衣壳蛋白在aav2衣壳蛋白的氨基酸位置264(即,插入)或另一种衣壳蛋白的相应位置之后包含苏氨酸、天冬氨酸、谷氨酸或苯丙氨酸。
在其他代表性实施方案中,本发明的修饰的衣壳蛋白或病毒衣壳进一步包含一个或多个如wo2007/089632中所述的突变(例如,aav2衣壳蛋白的氨基酸位置531或来自另一种aav的衣壳蛋白的相应位置处的e7k突变)。
在进一步实施方案中,修饰的衣壳蛋白或衣壳可以包含如wo2009/108274中所述的突变。
作为另一种可能性,aav衣壳蛋白可以包含如zhong等人(virology381:194-202(2008);proc.nat.acad.sci.105:7827-32(2008))所述的突变。例如,aav衣壳蛋白可以在氨基酸位置730处包含yf突变。
上述修饰可以彼此组合和/或与现在已知或后来发现的任何其它修饰组合并入本发明的衣壳蛋白或衣壳中。
本发明还涵盖了包含本发明的修饰的衣壳蛋白和衣壳的病毒载体。在具体实施方案中,病毒载体是细小病毒载体(例如包含细小病毒衣壳和/或载体基因组),例如aav载体(例如包含aav衣壳和/或载体基因组)。在代表性实施方案中,病毒载体含有包含本发明的修饰的衣壳蛋白亚基的修饰的aav衣壳和载体基因组。
例如,在代表性实施方案中,病毒载体包含:(a)包含本发明的修饰的衣壳蛋白的修饰的病毒衣壳(例如修饰的aav衣壳);(b)包含末端重复序列(例如aavtr)的核酸,其中包含末端重复序列的所述核酸由修饰的病毒衣壳衣壳化。核酸可以任选包含两个末端重复(例如两个aavtr)。
在代表性实施方案中,病毒载体是包含编码目的多肽或功能rna的异源核酸的重组病毒载体。重组病毒载体在下文更详细地描述。
在一些实施方案中,本发明的病毒载体(i)与通过不具有本发明的修饰的衣壳蛋白的病毒载体的转导水平相比,具有降低的肝转导;(ii)与通过不具有本发明的修饰的衣壳蛋白的病毒载体观察到的水平相比,表现出病毒载体在动物受试者中增强的全身转导;(iii)与不具有本发明的修饰的衣壳蛋白的病毒载体的运动水平相比,证明经内皮细胞的运动增强,和/或(iv)表现出肌肉组织(例如骨骼肌、心肌和/或膈肌)的转导的选择性增强,和/或(v)与不具有本发明的修饰的衣壳蛋白的病毒载体转导水平相比,脑组织(例如神经元)的转导减少。在一些实施方案中,病毒载体具有朝向肌肉的全身转导,例如其转导遍及全身的多重骨骼肌群,并且任选转导心肌和/或膈肌。
进一步,在本发明的一些实施方案中,修饰的病毒载体表明靶组织的有效转导。
本领域技术人员应当理解本发明的修饰的衣壳蛋白、病毒衣壳、病毒载体或aav颗粒排除如其在其天然状态下存在或发现的那些衣壳蛋白、衣壳、病毒载体或aav颗粒。
生产病毒载体的方法。
本发明进一步提供了生产本发明的创造性病毒载体作为aav颗粒的方法。因此,本发明提供了制备包含本发明的aav衣壳的aav颗粒的方法,其包括:(a)用一种或多种质粒转染宿主细胞,所述质粒组合提供组装aav颗粒所需的所有功能和基因;(b)将一种或多种核酸构建体引入包装细胞系或生产细胞系,以组合提供组装aav颗粒所需的所有功能和基因;(c)将一种或多种重组杆状病毒载体引入宿主细胞,所述杆状病毒载体组合提供组装aav颗粒所需的所有功能和基因;和/或(d)将一种或多种重组疱疹病毒载体引入宿主细胞,所述重组疱疹病毒载体组合提供组装aav颗粒所需的所有功能和基因。用于形成aav病毒粒子的条件是用于在细胞(例如哺乳动物或昆虫细胞)中产生aav载体的标准条件,其作为非限制性实例包括在ad辅助质粒或其他辅助病毒诸如hsv存在的情况下转染细胞。
制备本发明的病毒载体的各种方法的非限制性实例描述于clement和grieger("manufacturingofrecombinantadeno-associatedviralvectorsforclinicaltrials"mol.ther.methodsclindev.3:16002(2016))和grieger等人("productionofrecombinantadeno-associatedvirusvectorsusingsuspensionhek293cellsandcontinuousharvestofvectorfromtheculturemediaforgmpfixandflt1clinicalvector"molther24(2):287-297(2016)),全部内容其通过引用并入本文。
在一个代表性实施方案中,本发明提供了生产病毒载体的方法,该方法包括对细胞提供:(a)包含至少一个tr序列(例如aavtr序列)的核酸模板,和(b)对于核酸模板复制和包入aav衣壳内足够的aav序列(例如编码本发明的aav衣壳的aavrep序列和aavcap序列)。任选地,核酸模板进一步包含至少一种异源核酸序列。在具体实施方案中,核酸模板包含两个aavitr序列,其位于异源核酸序列(如果存在的话)的5’和3’,尽管它们无需与之直接邻接。
核酸模板以及aavrep和cap序列在这样的条件下提供,使得在细胞中产生包含在aav衣壳内包装的核酸模板的病毒载体。该方法可以进一步包括从细胞中收集病毒载体的步骤。病毒载体可以从培养基中和/或通过使细胞裂解进行收集。
在一个实施方案中,改变核酸模板,使得cap序列不能从仅来自一种血清型(第一核酸序列)的核酸序列表达所有三种病毒结构蛋白vp1、vp2和vp3。这种改变可以例如通过消除病毒结构蛋白中的至少一种的起始密码子来实现。该模板还将含有来自不同血清型的至少一种额外的核酸序列(第二核酸序列),其编码且能够表达不能由第一核酸序列表达的病毒结构蛋白,其中所述第二核酸序列不能表达能够通过第一核酸序列表达的病毒结构蛋白。在一个实施方案中,所述第一核酸序列能够表达病毒结构蛋白中的两种,而所述第二核酸序列能够仅表达剩余的病毒序列。例如,所述第一核酸序列能够表达来自一种血清型的vp1和vp2,但不是vp3,且所述第二核酸序列能够表达来自于替代血清型的vp3,但不是vp1或vp2。模板不能表达三种病毒结构蛋白中的任何其他蛋白。在一个实施方案中,所述第一核酸序列仅能够表达三种病毒结构蛋白之一,所述第二核酸序列能够仅表达其他两种病毒结构蛋白,而不是第一种。
在另一个实施方案中,存在来自第三血清型的第三核酸序列。在该实施方案中,三种核酸序列各自仅能够表达三种衣壳病毒结构蛋白vp1、vp2和vp3之一,且各自都不表达由另一序列表达的病毒结构蛋白,使得总的来说,产生包含vp1、vp2和vp3的衣壳,其中衣壳中的每种病毒结构蛋白都来自相同血清型,且在该实施方案中,vp1、vp2和vp3都来自不同血清型。
防止表达的改变可以通过本领域已知的任何方式进行。例如,消除起始密码子、剪接受体、剪接供体及其组合。可以使用缺失和添加以及改变阅读框的位点特异性改变。也可以合成产生核酸序列。这些辅助模板通常不含itr。
细胞可以是对于aav病毒复制许可的细胞。可以采用本领域已知的任何合适的细胞。在具体实施方案中,细胞是哺乳动物细胞。作为另一个选项,细胞可以是反式补充包装细胞系(trans-complementingpackagingcellline),其提供从复制缺陷辅助病毒中缺失的功能,例如293细胞或其他ela反式补充细胞。
aav复制和衣壳序列可以通过本领域已知的任何方法提供。目前方案通常在单一质粒上表达aavrep/cap基因。aav复制和包装序列无需一起提供,尽管这样做可以是方便的。aavrep和/或cap序列可以通过任何病毒或非病毒载体提供。例如,rep/cap序列可以通过杂合腺病毒或疱疹病毒载体提供(例如插入缺失的腺病毒载体的ela或e3区内)。ebv载体还可以用于表达aavcap和rep基因。该方法的一个优点是ebv载体是附加型,在相继细胞分裂自始至终仍维持高拷贝数(即作为染色体外元件稳定整合到细胞内,指定为“基于ebv的核附加体”,参见margolski(1992)curr.top.microbiol.immun.158:67)。
作为进一步的替代方案,可以将rep/cap序列稳定并入细胞内。通常,aavrep/cap序列不由tr侧接,以预防这些序列的援救和/或包装。
核酸模板可以使用本领域已知的任何方法提供给细胞。例如,模板可以由非病毒(例如质粒)或病毒载体供应。在具体实施方案中,核酸模板由疱疹病毒或腺病毒载体供应(例如插入缺失腺病毒的ela或e3区内)。作为另一个举例说明,palombo等人(1998)j.virology72:5025,描述了携带由aavtr侧接的报道基因的杆状病毒载体。ebv载体还可以用于如上所述就rep/cap基因而言递送模板。
在另一个代表性实施方案中,核酸模板通过复制raav病毒提供。在另外其他实施方案中,包含核酸模板的aav原病毒稳定整合到细胞的染色体内。
为了增强病毒滴度,促进生产性aav感染的辅助病毒功能(例如腺病毒或疱疹病毒)可以提供给细胞。aav复制所需的辅助病毒序列是本领域已知的。通常,这些序列由辅助腺病毒或疱疹病毒载体提供。或者,腺病毒或疱疹病毒序列可以通过另一种非病毒或病毒载体例如作为非感染性腺病毒小质粒提供,所述非感染性腺病毒小质粒携带促进有效aav生产的所有辅助基因,如由ferrari等人(1997)naturemed.3:1295;以及美国专利号6,040,183和6,093,570描述的。
进一步地,辅助病毒功能可以通过包装细胞提供,所述包装细胞具有在染色体中包埋或作为稳定的染色体外元件维持的辅助序列。一般地,辅助病毒序列不能包装到aav病毒粒子内,例如不由tr侧接。
本领域技术人员应当理解,在单一辅助构建体上提供aav复制和衣壳序列以及辅助病毒序列(例如腺病毒序列)可以是有利的。该辅助构建体可以是非病毒或病毒构建体。作为一个非限制性举例说明,辅助构建体可以是包含aavrep/cap基因的杂合腺病毒或杂合疱疹病毒。
在一个具体实施方案中,aavrep/cap序列和腺病毒辅助序列由单一腺病毒辅助载体供应。该载体还可以进一步包含核酸模板。aavrep/cap序列和/或raav模板可以插入腺病毒的缺失区(例如e1a或e3区)内。
在进一步的实施方案中,aavrep/cap序列和腺病毒辅助序列由单一腺病毒辅助载体供应。根据该实施方案,raav模板可以作为质粒模板提供。
在另一个说明性实施方案中,aavrep/cap序列和腺病毒辅助序列由单一腺病毒辅助载体供应,并且raav模板作为原病毒整合到细胞内。或者,raav模板由ebv载体提供,所述ebv载体作为染色体外元件(例如作为基于ebv的核附加体)维持在细胞内。
在进一步的示例性实施方案中,aavrep/cap序列和腺病毒辅助序列由单一腺病毒辅助载体提供。raav模板可以作为分开的复制病毒载体提供。例如,raav模板可以由raav颗粒或第二重组腺病毒颗粒提供。
根据前述方法,杂合腺病毒载体通常包含对于腺病毒复制和包装足够的腺病毒5’和3’顺式序列(即,腺病毒末端重复和pac序列)。aavrep/cap序列和raav模板(如果存在的话)包埋在腺病毒骨架中,并且由5'和3'顺式序列侧接,使得这些序列可以包装到腺病毒衣壳内。如上所述,腺病毒辅助序列和aavrep/cap序列一般不由tr侧接,使得这些序列不包装到aav病毒粒子内。
zhang等人((2001)genether.18:704-12)描述了包含腺病毒以及aavrep和cap基因两者的嵌合辅助者。
疱疹病毒还可以用作aav包装方法中的辅助病毒。
编码aavrep蛋白的杂合疱疹病毒可以有利地促进可扩大的aav载体生产方案。表达aav-2rep和cap基因的杂合单纯疱疹病毒i型(hsv-1)载体已得到描述(conway等人(1999)genetherapy6:986和wo00/17377。
作为进一步的替代方案,本发明的病毒载体可以使用杆状病毒载体在昆虫细胞中产生,以递送rep/cap基因和raav模板,如例如urabe等人(2002)humangenetherapy13:1935-43中所述。
不含污染性辅助病毒的aav载体储备物可以通过本领域已知的任何方法获得。例如,aav和辅助病毒可以基于大小容易地区分。aav还可以基于对于肝素底物的亲和力与辅助病毒分离开(zolotukhin等人(1999)genetherapy6:973)。可以使用缺失的复制缺陷辅助病毒,使得任何污染性辅助病毒不是有复制能力的。作为进一步的替代方案,可以采用缺乏晚期基因表达的腺病毒辅助者,因为仅需要腺病毒早期基因表达以介导aav病毒的包装。对于晚期基因表达缺陷的腺病毒突变体是本领域已知的(例如ts100k和ts149腺病毒突变体)。
重组病毒载体
本发明提供了将核酸分子施用于细胞的方法,所述方法包括使所述细胞与本发明的病毒载体、aav颗粒和/或组合物或药物制剂接触。
本发明进一步提供了将核酸递送至受试者的方法,所述方法包括向所述受试者施用本发明的病毒载体、aav颗粒和/或组合物或药物制剂接触。
在具体实施方案中,所述受试者是人,且在一些实施方案中,所述受试者具有可以通过基因疗法方案治疗的病症或处于可以通过基因疗法方案治疗的病症的风险中。此类病症的非限制性实例包括肌营养不良症(包括杜兴氏或贝克氏肌营养不良症),血友病a,血友病b,多发性硬化症,糖尿病,高雪氏病,法布里病,庞贝病,癌症,关节炎,肌肉萎缩,心脏病(包括充血性心力衰竭或周围动脉疾病),内膜增生,神经系统病症,包括:癫痫病,亨廷顿氏病,帕金森氏病或阿尔茨海默氏病,自身免疫性疾病,囊性纤维化,地中海贫血,霍勒氏综合征,斯莱综合征,谢伊综合征,hurler-scheie综合征,亨特氏综合征,沙费利波综合征a、b、c、d,莫基奥综合征,maroteaux-lamy综合征,克腊伯氏病,苯酮尿症,巴滕氏病,脊髓性共济失调,ldl受体缺乏症,高氨血症,贫血,关节炎,视网膜退行性病症,包括黄斑变性,腺苷脱氨酶缺乏,代谢病症,和癌症,包括形成肿瘤的癌症。
在本发明的方法的一些实施方案中,可以将本发明的病毒载体、aav颗粒和/或组合物或药物制剂施用于骨骼肌、心肌和/或膈肌。
在本文所述的方法中,可以将本发明的病毒载体、aav颗粒和/或组合物或药物制剂经由全身途径(例如,静脉内、动脉内、腹膜内等)施用/递送至本发明的受试者。在一些实施方案中,可以将所述病毒载体和/或组合物经由脑室内、脑池内、实质内、颅内和/或鞘内途径施用于受试者。在具体实施方案中,静脉内施用本发明的病毒载体和/或药物制剂。
本发明的病毒载体可用于在体外、离体和在体内将核酸分子递送至细胞。特别地,病毒载体可以有利地用于将核酸分子递送或转移至动物细胞,包括哺乳动物细胞。
任何目标异源核酸序列均可在本发明的病毒载体中递送。目标核酸分子包括编码多肽的核酸分子,包括治疗(例如用于医学或兽医学用途)和/或免疫原性(例如用于疫苗)多肽。
治疗多肽包括但不限于囊性纤维化跨膜调节蛋白(cftr)、抗肌萎缩蛋白(包括小和微小抗肌萎缩蛋白,参见例如vincent等人(1993)naturegenetics5:130;美国专利公开号2003/017131;国际专利公开号wo/2008/088895,wang等人proc.natl.acad.sci.usa97:13714-13719(2000);和gregorevic等人mol.ther.16:657-64(2008))、肌肉生长抑制素前肽、卵泡抑素、激活素ii型可溶性受体、igf-1、抗炎多肽例如iκb显性突变体、sarcospan、肌营养相关蛋白(utrophin)(tinsley等人(1996)nature384:349)、小肌营养相关蛋白、凝血因子、(例如因子viii、因子ix、因子x等)、促红细胞生成素,血管抑素、内皮抑素、过氧化氢酶、酪氨酸羟化酶、超氧化物歧化酶、瘦素、ldl受体、脂蛋白脂肪酶、鸟氨酸氨甲酰基转移酶、β-珠蛋白、α-珠蛋白、血影蛋白、α1-抗胰蛋白酶、腺苷脱氨酶、次黄嘌呤鸟嘌呤磷酸核糖基转移酶、β-葡糖脑苷脂酶、鞘磷脂酶、溶酶体氨基己糖苷酶a、支链酮酸脱氢酶、rp65蛋白、细胞因子(例如α-干扰素、β-干扰素、干扰素-γ、白介素-2、白介素-4、粒细胞-巨噬细胞集落刺激因子、淋巴毒素等)、肽生长因子、神经营养因子和激素(例如生长激素、胰岛素、胰岛素样生长因子1和2、血小板来源的生长因子、表皮生长因子、成纤维细胞生长因子、神经生长因子、神经营养因子-3和-4、脑来源的神经营养因子、骨形态发生蛋白[包括rankl和vegf]、神经胶质来源的生长因子、转化生长因子-α和-β等)、溶酶体酸性α-葡糖苷酶、α-半乳糖苷酶a、受体(例如肿瘤坏死生长因子α可溶性受体)、s100a1、小白蛋白、腺苷酸环化酶6型、调节钙处理的分子(例如pp1的serca2a抑制剂1及其片段[例如wo2006/029319和wo2007/100465])、影响g蛋白偶联受体激酶2型敲低的分子例如截短的组成型活性barkct、抗炎因子例如irap、抗肌肉生长抑制素蛋白、天冬氨酸酰酶、单克隆抗体(包括单链单克隆抗体;示例性mab是herceptin®mab)、神经肽及其片段(例如甘丙肽、神经肽y(参见美国专利号7,071,172)、血管生成抑制剂例如vasohibins及其他vegf抑制剂(例如vasohibin2[参见wojp2006/073052])。其他说明性异源核酸序列编码自杀基因产物(例如胸苷激酶、胞嘧啶脱氨酶、白喉毒素和肿瘤坏死因子),赋予对在癌症疗法中使用的药物抗性的蛋白、肿瘤抑制基因产物(例如p53、rb、wt-1)、trail、fas-配体、以及在有需要的受试者中具有疗效的任何其他多肽。aav受试者还可以用于递送单克隆抗体和抗体片段,例如针对肌肉生长抑制素的抗体或抗体片段(参见例如fang等人naturebiotechnology23:584-590(2005))。
编码多肽的异源核酸序列包括编码报道多肽(例如酶)的那些。报道多肽是本领域已知的,并且包括但不限于绿色荧光蛋白(gfp)、荧光素酶、β-半乳糖苷酶、碱性磷酸酶、萤光素酶和氯霉素乙酰转移酶基因。
任选地,异源核酸分子编码分泌多肽(例如作为以其天然状态的分泌多肽,或例如通过与如本领域已知的分泌信号序列可操作地结合已工程改造为被分泌的多肽)。
或者,在本发明的具体实施方案中,异源核酸分子可以编码反义核酸分子,核酶(例如如美国专利号5,877,022中所述),实现剪接体介导的反式剪接的rna(参见puttaraju等人(1999)naturebiotech.17:246;美国专利号6,013,487;美国专利号6,083,702),干扰rna(rnai)包括介导基因沉默的sirna、shrna或mirna(参见sharp等人(2000)science287:2431),以及其他非翻译rna,例如“引导”rna(gorman等人(1998)proc.nat.acad.sci.usa95:4929;给予yuan等人的美国专利号5,869,248)等。示例性非翻译rna包括针对多药抗性(mdr)基因产物的rnai(例如以治疗和/或预防肿瘤和/或用于施用于心脏以预防通过化学疗法的损害)、针对肌肉生长抑制素的rnai(例如用于杜兴肌营养不良)、针对vegf的rnai(例如以治疗和/或预防肿瘤)、针对受磷蛋白的rnai(例如以治疗心血管疾病,参见例如andino等人j.genemed.10:132-142(2008)和li等人actapharmacolsin.26:51-55(2005));受磷蛋白抑制或显性失活分子例如受磷蛋白s16e(例如以治疗心血管疾病,参见例如hoshijima等人nat.med.8:864-871(2002)),针对腺苷激酶的rnai(例如用于癫痫症),以及针对致病性生物和病毒(例如乙型和/或丙型肝炎病毒、人免疫缺陷病毒、cmv、单纯疱疹病毒、人乳头状瘤病毒等)的rnai。
进一步地,可以递送指导可变剪接的核酸序列。为了举例说明,与抗肌萎缩蛋白外显子51的5'和/或3'剪接位点互补的反义序列(或其他抑制序列)可以与u1或u7小核(sn)rna启动子结合递送,以诱导该外显子的跳跃。例如,包含位于反义/抑制序列5'的u1或u7snrna启动子的dna序列可以在本发明的修饰的衣壳内包装且递送。
病毒载体还可以包含异源核酸分子,其与宿主细胞染色体上的基因座共享同源性且与之重组。该方法可以例如用于校正宿主细胞中的遗传缺陷。
本发明还提供了表达免疫原性多肽、肽和/或表位的病毒载体,例如用于疫苗接种。核酸分子可以编码本领域已知的任何目标免疫原,包括但不限于来自人免疫缺陷病毒(hiv)、猴免疫缺陷病毒(siv)、流感病毒的免疫原,hiv或sivgag蛋白,肿瘤抗原,癌抗原,细菌抗原,病毒抗原等。
细小病毒作为疫苗载体的用途是本领域已知的(参见例如miyamura等人,(1994)proc.nat.acad.sciusa91:8507;授予young等人的美国专利号5,916,563、授予mazzara等人的美国专利号5,905,040、授予samulski等人的美国专利号5,882,652和美国专利号5,863,541)。抗原可以存在于细小病毒衣壳内。或者,免疫原或抗原可以由引入重组载体基因组内的异源核酸分子表达。如本文描述的和/或如本领域已知的任何目标免疫原或抗原可以由本发明的病毒载体提供。
免疫原性多肽可以是适合于引发免疫应答和/或保护受试者不受感染和/或疾病的任何多肽、肽和/或表位,所述感染和/或疾病包括但不限于微生物、细菌、原生动物、寄生虫、真菌和/或病毒感染和疾病。例如,免疫原性多肽可以是正粘病毒免疫原(例如流感病毒免疫原,例如流感病毒血凝素(ha)表面蛋白或流感病毒核蛋白、或马流感病毒免疫原)、或慢病毒免疫原(例如马传染性贫血病毒免疫原、猴免疫缺陷病毒(siv)免疫原、或人免疫缺陷病毒(hiv)免疫原,例如hiv或siv包膜gp160蛋白,hiv或siv基质/衣壳蛋白,和hiv或sivgag、pol和env基因产物)。免疫原性多肽还可以是沙粒病毒免疫原(例如拉沙热病毒免疫原,例如拉沙热病毒核衣壳蛋白和拉沙热包膜糖蛋白)、痘病毒免疫原(例如牛痘病毒免疫原,例如牛痘l1或l8基因产物)、黄病毒免疫原(例如黄热病毒免疫原或日本脑炎病毒免疫原)、丝状病毒免疫原(例如埃博拉病毒免疫原或马尔堡病毒免疫原,例如np和gp基因产物)、布尼亚病毒免疫原(例如rvfv、cchf和/或sfs病毒免疫原)、或冠状病毒免疫原(例如传染性人冠状病毒免疫原例如人冠状病毒包膜糖蛋白、或猪传播性胃肠炎病毒免疫原、或禽传染性支气管炎病毒免疫原)。免疫原性多肽可以进一步是脊髓灰质炎免疫原、疱疹免疫原(例如cmv、ebv、hsv免疫原)、腮腺炎免疫原、麻疹免疫原、风疹免疫原、白喉毒素或其他白喉免疫原、百日咳抗原、肝炎(例如甲型肝炎、乙型肝炎、丙型肝炎等)免疫原、和/或本领域目前已知或后来鉴定为免疫原的任何其他疫苗免疫原。
或者,免疫原性多肽可以是任何肿瘤或癌细胞抗原。任选地,肿瘤或癌抗原在癌细胞的表面上表达。示例性癌症和肿瘤细胞抗原在s.a.rosenberg(immunity10:281(1991))中描述。其他说明性癌症和肿瘤抗原包括但不限于brca1基因产物、brca2基因产物、gp100、酪氨酸酶、gage-1/2、bage、rage、lage、ny-eso-1、cdk-4、β-连环蛋白、mum-1、半胱天冬酶-8、kiaa0205、hpve、sart-1、prame、p15、黑素瘤肿瘤抗原(kawakami等人(1994)proc.natl.acad.sci.usa91:3515;kawakami等人(1994)j.exp.med.,180:347;kawakami等人(1994)cancerres.54:3124)、mart-1、gp100mage-1、mage-2、mage-3、cea、trp-1、trp-2、p-15、酪氨酸酶(brichard等人(1993)j.exp.med.178:489);her-2/neu基因产物(美国专利号4,968,603)、ca125、lk26、fb5(内皮唾液酸蛋白)、tag72、afp、ca19-9、nse、du-pan-2、ca50、span-1、ca72-4、hcg、stn(唾液酸tn抗原)、c-erbb-2蛋白、psa、l-canag、雌激素受体、乳脂球蛋白、p53肿瘤抑制蛋白(levine,(1993)ann.rev.biochem.62:623);粘蛋白抗原(国际专利公开号wo90/05142);端粒酶;核基质蛋白;前列腺酸性磷酸酶;乳头状瘤病毒抗原;和/或目前已知或以后发现与下述癌症相关的抗原:黑素瘤、腺癌、胸腺瘤、淋巴瘤(例如非霍奇金氏淋巴瘤、霍奇金氏淋巴瘤)、肉瘤、肺癌、肝癌、结肠癌、白血病、子宫癌、乳腺癌、前列腺癌、卵巢癌、宫颈癌、膀胱癌、肾癌、胰腺癌、脑癌和目前已知或后来鉴定的任何其他癌症或恶性状况(参见例如rosenberg,(1996)ann.rev.med.47:481-91)。
作为进一步的替代方案,异源核酸分子可以编码在体外、离体或在体内在细胞中期望地产生的任何多肽、肽和/或表位。例如,病毒载体可以引入培养的细胞内,并且由其分离所表达的基因产物。
本领域技术人员应当理解,目标异源核酸分子可以与适当的控制序列可操作地结合。例如,异源核酸分子可以与表达控制元件可操作地结合,所述表达控制元件例如转录/翻译控制信号、复制起点、多聚腺苷酸化信号、内部核糖体进入位点(ires)、启动子和/或增强子等。
进一步地,例如通过选择性阻断在特异性位点处的剪接活性的寡核苷酸、小分子和/或其他化合物(例如如wo2006/119137中所述)的存在或不存在,通过调节不同内含子的选择性剪接,可以在转录后水平下实现目标异源核酸分子的调节表达。
本领域技术人员应当理解,取决于所需水平和组织特异性表达,可以使用各种启动子/增强子元件。取决于所需表达模式,启动子/增强子可以是组成型或诱导型的。启动子/增强子可以是天然或外源的,并且可以是天然或合成序列。外源预期转录起始区在转录起始区引入其内的野生型宿主中未发现。
在具体实施方案中,启动子/增强子元件对于待处理的靶细胞或受试者可以是天然的。在代表性实施方案中,启动子/增强子元件对于异源核酸序列可以是天然的。
启动子/增强子元件一般这样进行选择,使得它在目标靶细胞中起作用。进一步地,在具体实施方案中,启动子/增强子元件是哺乳动物启动子/增强子元件。启动子/增强子元件可以是组成型或诱导型的。
诱导型表达控制元件通常在其中期望提供异源核酸序列的过表达调节的那些应用中是有利的。用于基因递送的诱导型启动子/增强子元件可以是组织特异性或优选的启动子/增强子元件,并且包括肌肉特异性或优选的(包括心肌、骨骼肌和/或平滑肌特异性或优选的)、神经组织特异性或优选的(包括脑特异性或优选的)、眼特异性或优选的(包括视网膜特异性和角膜特异性的)、肝特异性或优选的、骨髓特异性或优选的、胰腺特异性或优选的、脾特异性或优选的、和肺特异性或优选的启动子/增强子元件。其他诱导型启动子/增强子元件包括激素诱导型和金属诱导型元件。示例性诱导型启动子/增强子元件包括但不限于tet开/关元件、ru486诱导型启动子、蜕皮激素诱导型启动子、雷帕霉素诱导型启动子和金属硫蛋白启动子。
在其中异源核酸序列在靶细胞中转录且随后翻译的实施方案中,一般包括特异性起始信号用于所插入的蛋白编码序列的有效翻译。这些外源翻译控制序列可以包括atg起始密码子和邻近序列,可以具有各种起源,天然和合成的两者。
根据本发明的病毒载体提供了用于将异源核酸分子递送到广泛范围的细胞包括分裂和不分裂细胞内的手段。病毒载体可以用于将目标核酸分子在体外递送至细胞,例如在体外产生多肽或用于离体或体内基因疗法。病毒载体另外可用于将核酸递送至有需要的受试者的方法中,例如以表达免疫原性或治疗多肽或功能rna。以这种方式,多肽或功能rna可以在受试者中在体内产生。因为受试者具有多肽的缺乏,所以受试者可以需要该多肽。
进一步地,因为多肽或功能rna在受试者中的产生可以赋予一些有利效应,所以可以实践该方法。
病毒载体还可以用于在培养的细胞或受试者中产生目标多肽或功能rna(例如使用受试者作为生物反应器来产生多肽,或例如与筛选方法结合观察功能rna对受试者的作用)。
一般而言,本发明的病毒载体可以用于递送编码多肽或功能rna的异源核酸分子,以治疗和/或预防对其而言递送治疗多肽或功能rna是有利的任何病症或疾病状态。说明性疾病状态包括但不限于:囊性纤维化(囊性纤维化跨膜调节蛋白)和肺的其他疾病、血友病a(因子viii)、血友病b(因子ix)、地中海贫血(β-珠蛋白)、贫血(促红细胞生成素)及其他血液病症、阿尔茨海默氏病(gdf;脑啡肽酶)、多发性硬化(β-干扰素)、帕金森氏病(神经胶质细胞系来源的神经营养因子[gdnf])、亨廷顿氏病(rnai以去除重复)、肌萎缩侧索硬化、癫痫症(甘丙肽、神经营养因子)、及其他神经病症、癌症(内皮抑素、血管抑素、trail、fas-配体、细胞因子包括干扰素;rnai包括针对vegf或多药抗性基因产物的rnai、mir-26a[例如用于肝细胞癌])、糖尿病(胰岛素)、肌营养不良包括杜兴(抗肌萎缩蛋白、小抗肌萎缩蛋白、胰岛素样生长因子i、肌聚糖[例如α、β、γ]、针对肌肉生长抑制素的rnai、肌肉生长抑制素前肽、卵泡抑素、激活素ii型可溶性受体、抗炎多肽例如iκb显性突变体、sarcospan、肌营养相关蛋白、小肌营养相关蛋白、针对抗肌萎缩蛋白基因中的剪接点的反义或rnai以诱导外显子跳跃[参见例如wo/2003/095647]、针对u7snrna的反义以诱导外显子跳跃[参见例如wo/2006/021724]、或针对肌肉生长抑制素或肌肉生长抑制素前肽的抗体或抗体片段)和贝克尔、戈谢病(葡糖脑苷脂酶)、胡尔勒病(α-l-艾杜糖醛酸酶)、腺苷脱氨酶缺乏症(腺苷脱氨酶)、糖原贮积病(例如法布里病[-α半乳糖苷酶]和庞贝氏症[溶酶体酸性α葡糖苷酶])及其他代谢病症、先天性肺气肿(α1-抗胰蛋白酶)、莱-尼二氏(lesch-nyhan)综合征(次黄嘌呤鸟嘌呤磷酸核糖基转移酶)、尼曼-皮克病(鞘磷脂酶)、泰萨克斯病(tay-sachsdisease)(溶酶体氨基己糖苷酶a)、枫糖尿症(支链酮酸脱氢酶)、视网膜变性疾病(以及眼和视网膜的其他疾病;例如pdgf用于黄斑变性和/或vasohibin或其他vegf抑制剂或其他血管生成抑制剂,以治疗/预防视网膜病症,例如在i型糖尿病中)、实体器官例如脑(包括帕金森氏病[gdnf]、星形细胞瘤[内皮抑素、血管抑素和/或针对vegf的rnai]、成胶质细胞瘤[内皮抑素、血管抑素和/或针对vegf的rnai])、肝、肾、心脏的疾病包括充血性心力衰竭或外周动脉疾病(pad)(例如通过递送蛋白磷酸酶抑制剂i(i-1)及其片段(例如i1c)、serca2a、调节受磷蛋白基因的锌指蛋白、barkct、β2-肾上腺素受体、β2-肾上腺素受体激酶(bark)、磷脂酰肌醇-3激酶(pi3激酶)、s100a1s100al、小白蛋白、腺苷酸环化酶6型、影响g蛋白偶联受体激酶2型敲低的分子例如截短的组成型活性barkct;calsarcin、针对受磷蛋白的rnai;受磷蛋白抑制或显性失活分子例如受磷蛋白s16e等)、关节炎(胰岛素样生长因子)、关节病症(胰岛素样生长因子1和/或2)、内膜增生(例如通过递送enos、inos)、改善心脏移植的存活(超氧化物歧化酶)、aids(可溶性cd4)、肌萎缩(胰岛素样生长因子i)、肾虚(促红细胞生成素)、贫血(促红细胞生成素)、关节炎(抗炎因子如irap和tnfα可溶性受体)、肝炎(α-干扰素)、ldl受体缺乏(ldl受体)、高氨血症(鸟氨酸氨甲酰基转移酶)、克拉伯病(半乳糖脑苷脂酶)、巴滕病、脊髓小脑性共济失调包括sca1、sca2和sca3、苯丙酮尿症(苯丙氨酸羟化酶)、自身免疫疾病等。本发明可以进一步在器官移植后使用,以增加移植的成功和/或降低器官移植或辅助疗法的不良副作用(例如通过施用免疫抑制剂或抑制核酸以阻断细胞因子生产)。作为另一个实例,例如在骨折后或在癌症患者中的手术摘除后,骨形态发生蛋白(包括bnp2、7等,rankl和/或vegf)可以与骨异体移植物一起施用。
本发明还可以用于生产诱导的多能干细胞(ips)。例如,本发明的病毒载体可以用于将干细胞相关核酸递送到非多能细胞内,例如成年成纤维细胞、皮肤细胞、肝细胞、肾细胞、脂肪细胞、心肌细胞、神经细胞、上皮细胞、内皮细胞等。编码与干细胞相关的因子的核酸是本领域已知的。与干细胞和多能性相关的此类因子的非限制性实例包括oct-3/4、sox家族(例如sox1、sox2、sox3和/或sox15)、klf家族(例如klf1、klf2、klf4和/或klf5)、myc家族(例如c-myc、l-myc和/或n-myc)、nanog和/或lin28。
本发明还可以进行实践,以治疗和/或预防代谢病症例如糖尿病(例如胰岛素)、血友病(例如因子ix或因子viii)、溶酶体贮积症例如粘多糖贮积症(例如斯莱综合征[β-葡糖醛酸酶]、胡尔勒综合征[α-l-艾杜糖醛酸酶]、沙伊综合征[α-l-艾杜糖醛酸酶]、胡-射二氏(hurler-scheie)综合征[α-l-艾杜糖醛酸酶]、亨特氏综合征[艾杜糖醛酸硫酸酯酶]、圣菲利波综合征a[乙酰肝素磺酰胺酶]、b[n-乙酰氨基葡糖苷酶]、c[乙酰-coa:α-氨基葡糖苷乙酰基转移酶]、d[n-乙酰氨基葡糖-6-硫酸酯酶]、莫尔基奥氏综合征a[半乳糖-6-硫酸盐硫酸酯酶]、b[β-半乳糖苷酶]、马-兰二氏(maroteaux-lamy)综合征[n-乙酰氨基半乳糖-4-硫酸酯酶]等)、法布里病(α-半乳糖苷酶)、戈谢病(葡糖脑苷脂酶)、或糖原贮积症(例如庞贝氏症;溶酶体酸性α-葡糖苷酶)。
基因转移具有用于理解且提供疾病状态疗法的基本潜在用途。存在其中缺陷基因是已知的且已得到克隆的许多遗传性疾病。一般而言,上述疾病状态落入两个类别内:通常为酶的缺乏状态,其一般以隐性遗传方式遗传,和不平衡状态,其可以涉及调节或结构蛋白,并且通常以显性方式遗传。对于缺乏状态疾病,基因转移可以用于将正常基因带入受累组织内用于替代疗法,以及使用反义突变产生疾病的动物模型。对于不平衡疾病状态,基因转移可以用于产生在模型系统中的疾病状态,其随后可以用于对抗疾病状态的努力中。因此,根据本发明的病毒载体允许遗传疾病的治疗和/或预防。
根据本发明的病毒载体还可以用于在体外或在体内将功能rna提供给细胞。功能rna在细胞中的表达例如可以缩小特定靶蛋白通过细胞的表达。相应地,可以施用功能rna,以减少特定蛋白在有需要的受试者中的表达。功能rna还可以在体外施用于细胞,以调节基因表达和/或细胞生理学,例如以优化细胞或组织培养系统或筛选方法。
另外,根据本发明的病毒载体在诊断和筛选方法中有用,由此目标核酸在细胞培养系统中或可替代的转基因动物模型中瞬时或稳定表达。
本发明的病毒载体还可以用于各种非治疗目的,包括但不限于在评价基因靶向、清除、转录、翻译等的方案中使用,如对于本领域技术人员显而易见的。病毒载体还可以用于评估安全(传播、毒性、免疫原性等)的目的。此类数据例如由美国食品和药物管理局(unitedstatesfoodanddrugadministration)视为在临床功效评估前的监管审批过程的部分。
作为进一步方面,本发明的病毒载体可以用于在受试者中产生免疫应答。根据该实施方案,包含编码免疫原性多肽的异源核酸序列的病毒载体可以施用于受试者,或活性免疫应答通过受试者针对免疫原性多肽产生。免疫原性多肽如上文描述。在一些实施方案中,引发保护性免疫应答。
或者,病毒载体可以离体施用于细胞,并且将改变的细胞施用于受试者。将包含异源核酸的病毒载体引入细胞内,并且将细胞施用于受试者,编码免疫原的异源核酸可以在其中表达且在受试者中诱导针对免疫原的免疫应答。在具体实施方案中,细胞是抗原呈递细胞(例如树突细胞)。
“主动免疫应答”或“主动免疫”的特征在于“在遇到免疫原后宿主组织和细胞的参与。它涉及免疫活性细胞在淋巴网状组织中的分化和增殖,其导致抗体合成或细胞介导反应性的发展,或两者”。herbertb.herscowitz,immunophysiology:cellfunctionandcellularinteractionsinantibodyformation,inimmunology:basicprocesses117(josepha.bellanti编辑1985)。或者说,在通过感染或疫苗接种暴露于免疫原后,通过宿主产生主动免疫应答。主动免疫可以与被动免疫相比,所述被动免疫通过“预先形成的物质(抗体、转移因子、胸腺移植物和白介素-2)从主动免疫的宿主转移到非免疫宿主”获得。同上。
如本文所用,“保护性”免疫应答或“保护性”免疫指示免疫应答对受试者赋予一定益处,因为它预防或降低疾病的发生率。或者,保护性免疫应答或保护性免疫可以用于治疗和/或预防疾病,特别是癌症或肿瘤(例如通过预防癌症或肿瘤形成,通过引起癌症或肿瘤消退,和/或通过预防转移和/或通过预防转移结节的生长)。保护效应可以是完全或部分的,只要治疗的益处超过其任何缺点。
在具体实施方案中,包含异源核酸分子的病毒载体或细胞可以以免疫原性有效量施用,如下面描述的。
通过施用表达一种或多种癌细胞抗原(或免疫相似分子),或产生针对癌细胞的免疫应答的任何其他免疫原的病毒载体,还可以施用本发明的病毒载体用于癌症免疫疗法。为了举例说明,通过施用包含编码癌细胞抗原的异源核酸的病毒载体,可以在受试者中产生针对癌细胞抗原的免疫应答,例如以治疗患有癌症的患者和/或预防癌症在受试者中的发展。病毒载体可以在体内或通过使用离体方法施用于受试者,如本文描述的。或者,癌抗原可以作为病毒衣壳的部分表达,或以其他方式与病毒衣壳结合(例如如上所述)。
作为另一个替代方案,可以施用本领域已知的任何其他治疗核酸(例如rnai)或多肽(例如细胞因子),以治疗和/或预防癌症。
如本文所用,术语“癌症”涵盖肿瘤形成癌症。
同样地,术语“癌组织”涵盖肿瘤。“癌细胞抗原”涵盖肿瘤抗原。
术语“癌症”具有其在本领域中理解的含义,例如不受控制的组织生长,其具有传播到机体的遥远部位(即转移)的潜力。示例性癌症包括但不限于黑素瘤、腺癌、胸腺瘤、淋巴瘤(例如非霍奇金氏淋巴瘤、霍奇金氏淋巴瘤)、肉瘤、肺癌、肝癌、结肠癌、白血病、子宫癌、乳腺癌、前列腺癌、卵巢癌、宫颈癌、膀胱癌、肾癌、胰腺癌、脑癌和目前已知或后来鉴定的任何其他癌症或恶性状况。在代表性实施方案中,本发明提供了治疗和/或预防肿瘤形成癌症的方法。
术语“肿瘤”还在本领域中例如理解为在多细胞生物内的未分化细胞的异常团块。肿瘤可以是恶性的或良性的。在代表性实施方案中,本文公开的方法用于预防和治疗恶性肿瘤。
术语“治疗癌症”、“癌症的治疗”和等价术语预期癌症的严重程度得到降低或至少部分消除,和/或疾病的进展减慢和/或得到控制和/或疾病得到稳定。在具体实施方案中,这些术语指示癌症的转移得到预防或降低或至少部分消除,和/或转移结节的生长得到预防或降低或至少部分消除。
术语“癌症的预防”或“预防癌症”和等价术语预期该方法至少部分消除或降低和/或延迟癌症的发生率和/或癌症发作的严重程度。或者说,受试者中的癌症发作可以在可能性或概率中得到降低和/或延迟。
在具体实施方案中,细胞可以从患有癌症的受试者中取出,并且与根据本发明的表达癌细胞抗原的病毒载体接触。随后将修饰的细胞施用于受试者,由此引发针对癌细胞抗原的免疫应答。该方法可以有利地对免疫妥协的受试者使用,所述免疫妥协的受试者不能在体内产生足够的免疫应答(即,不能产生足够数量的增强抗体)。
本领域已知免疫应答可以通过免疫调节细胞因子(例如α-干扰素、β-干扰素、γ-干扰素、ω-干扰素、τ-干扰素、白介素-1α、白介素-1β、白介素-2、白介素-3、白介素-4、白介素5、白介素-6、白介素-7、白介素-8、白介素-9、白介素-10、白介素-11、白介素12、白介素-13、白介素-14、白介素-18、b细胞生长因子、cd40配体、肿瘤坏死因子-α、肿瘤坏死因子-β、单核细胞趋化蛋白-1、粒细胞-巨噬细胞集落刺激因子和淋巴毒素)得到增强。相应地,免疫调节细胞因子(优选地ctl诱导细胞因子)可以与病毒载体结合施用于受试者。
细胞因子可以通过本领域已知的任何方法进行施用。外源细胞因子可以施用于受试者,或可替代地,编码细胞因子的核酸可以使用合适载体递送至受试者,并且在体内产生细胞因子。
受试者、药物制剂和施用模式
根据本发明的病毒载体、aav颗粒和衣壳在兽医学和医学应用两者中有用。合适的受试者包括禽类和哺乳动物两者。如本文所用,术语“禽类”包括但不限于鸡、鸭、鹅、鹌鹑、火鸡、雉鸡、鹦鹉、长尾鹦鹉等。如本文所用,术语“哺乳动物”包括但不限于人、非人灵长类动物、牛、绵羊、山羊、马、猫、犬、兔类动物等。
人受试者包括新生儿、婴幼儿、青少年、成人和老年受试者。
在代表性实施方案中,受试者“需要”本发明的方法。
在具体实施方案中,本发明提供了药物组合物,其包含在药学上可接受的载体中的本发明的病毒载体和/或衣壳和/或aav颗粒,以及任选的其他医学试剂、药物试剂、稳定剂、缓冲剂、载体、佐剂、稀释剂等。对于注射,载体通常为液体。对于其他施用模式,载体可以是固体或液体。对于吸入施用,载体将是呼吸性的,并且任选可以采取固体或液体微粒形式。对于施用于受试者或对于其他药物用途,所述载体将是无菌和/或生理相容的。
“药学上可接受的”意指无毒或在其他方面不期望的材料,即材料可以施用于受试者,而不引起任何不期望的生物效应。
本发明的一个方面是在体外将核酸分子转移至细胞的方法。根据适合于特定靶细胞的标准转导方法,病毒载体可以以适当的感染复数引入细胞内。取决于靶细胞类型和数目,以及特定的病毒载体,待施用的病毒载体滴度可以改变,并且可以由本领域技术人员无需过度实验而决定。在代表性实施方案中,将至少约103感染单位、任选至少约105感染单位引入细胞。
病毒载体引入其内的细胞可以具有任何类型,包括但不限于神经细胞(包括外周和中枢神经系统的细胞,特别是脑细胞例如神经元和少突胶质细胞)、肺细胞、眼细胞(包括视网膜细胞、视网膜色素上皮和角膜细胞)、上皮细胞(例如肠道和呼吸道上皮细胞)、肌细胞(例如骨骼肌细胞、心肌细胞、平滑肌细胞和/或膈肌细胞)、树突细胞、胰腺细胞(包括胰岛细胞)、肝细胞、心肌细胞、骨细胞(例如骨髓干细胞)、造血干细胞、脾细胞、角化细胞、成纤维细胞、内皮细胞、前列腺细胞、生殖细胞等。在代表性实施方案中,细胞可以是任何祖细胞。作为进一步可能性,细胞可以是干细胞(例如神经干细胞、肝干细胞)。作为再进一步的替代实施方案,细胞可以是癌症或肿瘤细胞。此外,细胞可以来自任何起源的物种,如上所述。
病毒载体可以在体外引入细胞内,用于将修饰的细胞施用于受试者的目的。在具体实施方案中,细胞已从受试者中取出,并且将病毒载体引入其内,并且随后将细胞施用回受试者。从受试者中取出细胞用于离体操作,随后引入回受试者内的方法是本领域已知的(参见例如美国专利号5,399,346)。或者,重组病毒载体可以引入来自供体受试者的细胞、培养的细胞或来自其他合适来源的细胞内,并且将细胞施用于有需要的受试者(即,“接受者”受试者)。
用于离体核酸递送的合适细胞如上所述。待施用于受试者的细胞剂量根据受试者的年龄、状况和物种,细胞类型,由细胞表达的核酸,施用模式等而改变。通常,施用在药学上可接受的载体中的至少约102至约108细胞或至少约103至约106细胞/剂量。在具体实施方案中,用病毒载体转导的细胞以治疗有效或预防有效量与药学载体组合施用于受试者。
在一些实施方案中,将病毒载体引入细胞内,并且将细胞施用于受试者,以引发针对所递送多肽(例如作为转基因表达或在衣壳中)的免疫应答。通常,施用表达与药学上可接受的载体组合的免疫原性有效量的多肽的细胞数量。“免疫有效量”是所表达多肽的量,其足以在药物制剂施用于其的受试者中诱发针对多肽的主动免疫应答。在具体实施方案中,剂量足以产生保护性免疫应答(例如上文定义的)。
赋予的保护程度无需是完全或永久的,只要施用免疫原性多肽的益处超过其任何缺点。
本发明的进一步方面是将病毒载体和/或病毒衣壳施用于受试者的方法。根据本发明的病毒载体和/或衣壳施用于有需要的人受试者或动物可以通过本领域已知的任何手段。任选地,病毒载体和/或衣壳以在药学上可接受的载体中的治疗有效或预防有效剂量递送。
可以进一步施用本发明的病毒载体和/或衣壳,以引发免疫应答(例如作为疫苗)。通常,本发明的免疫原性组合物包含与药学上可接受的载体组合的免疫原性有效量的病毒载体和/或衣壳。任选地,剂量足以产生保护性免疫应答(例如上文定义的)。赋予的保护程度不必是完全或永久的,只要施用免疫原性多肽的益处超过其任何缺点。受试者和免疫原如上所述。
待施用于受试者的病毒载体和/或衣壳的剂量取决于施用模式、待治疗和/或预防的疾病或状况、各个受试者的状况、特定病毒载体或衣壳和待递送的核酸等,并且可以以常规方式测定。用于实现疗效的示例性剂量是至少约105、106、107、108、109、1010、1011、1012、103、1014、1015转导单位,任选约108至1013转导单位的滴度。
在具体实施方案中,多于一次施用(例如两次、三次、四次、五次、六次、七次、八次、九次、十次等或更多次施用)可以用于实现在各种间隔的时期上的所需基因表达水平,例如每小时一次、每天一次、每周一次、每月一次、每年一次等。给药可以是单剂量或累积的(连续给药),并且可以由本领域技术人员容易地确定。例如,疾病或病症的治疗可包括一次性施用有效剂量的本文公开的药物组合物病毒载体。或者,疾病或病症的治疗可包括经一定范围的时间段实施的有效剂量的病毒载体的多次施用,诸如例如每天一次、每天两次、每天三次、每几天一次或每周一次。施用的时机可以因个体而变化,这取决于诸如个体症状的严重程度等因素。例如,本文公开的病毒载体的有效剂量可以无限期地每六个月一次施用于个体,或者直到个体不再需要疗法为止。本领域普通技术人员将认识到,可以在整个治疗过程中监测个体的状况,并且可以相应地调节施用的本文公开的病毒载体的有效量。
在一个实施方案中,病毒载体的施用时段持续1天、2天、3天、4天、5天、6天、7天、8天、9天、10天、11天、12天、13天、14天、3周、4周、5周、6周、7周、8周、9周、10周、11周、12周、4个月、5个月、6个月、7个月、8个月、9个月、10个月、11个月、12个月或更长时间。在一个进一步实施方案中,期间停止施用的时段持续1天、2天、3天、4天、5天、6天、7天、8天、9天、10天、11天、12天、13天、14天、3周、4周、5周、6周、7周、8周、9周、10周、11周、12周、4个月、5个月、6个月、7个月、8个月、9个月、10个月、11个月、12个月或更长时间。
示例性施用模式包括口服、直肠、经粘膜、鼻内、吸入(例如经由气溶胶)、经颊(例如舌下)、阴道、鞘内、眼内、经皮、在子宫内(或在卵内)、肠胃外(例如静脉内、皮下、皮内、肌内[包括施用于骨骼肌、膈肌和/或心肌]、皮内、胸膜内、脑内和关节内)、局部(例如对皮肤和粘膜表面两者,包括气道表面和经皮施用)、淋巴管内等,以及直接的组织或器官注射(例如对肝、骨骼肌、心肌、膈肌或脑)。施用还可以针对肿瘤(例如在肿瘤或淋巴结中或者接近肿瘤或淋巴结)。在任何给定情况下的最合适途径取决于待治疗和/或预防状况的性质和严重程度,以及使用的特定载体的性质。
根据本发明对骨骼肌的施用包括但不限于对肢体(例如上臂、下臂、大腿和/或小腿)、背部、颈部、头部(例如舌)、胸部、腹部、骨盆/会阴、和/或手指中的骨骼肌的施用。合适的骨骼肌包括但不限于小指展肌(在手中)、小趾展肌(在足中)、拇展肌、外展小趾肌、拇短展肌、拇长展肌、短收肌、拇收肌、长收肌、大收肌、拇内收肌、肘肌、前斜角肌、膝关节肌、肱二头肌、股二头肌、肱肌、肱桡肌、颊肌、喙肱肌、皱眉肌、三角肌、降口角肌、降下唇肌、二腹肌、背侧骨间肌(在手中)、背侧骨间肌(在足中)、桡侧腕短伸肌、桡侧腕长伸肌、尺侧腕伸肌、小指伸肌、指伸肌、趾短伸肌、趾长伸肌、短伸肌、长伸肌、食指伸肌、拇短伸肌、拇长伸肌、桡侧腕屈肌、尺侧腕屈肌、小指短屈肌(在手中)、小趾短屈肌(在足中)、趾短屈肌、趾长屈肌、指深屈肌、指浅屈肌、短屈肌、长屈肌、拇短屈肌、拇长屈肌、额肌、腓肠肌、颏舌骨肌、臀大肌、臀中肌、臀小肌、股薄肌、颈髂肋肌、腰髂肋肌、胸髂肋肌、骼肌、下孖肌、下斜肌、下直肌、冈下肌、棘突间肌、横突间肌、翼外肌、外直肌、背阔肌、提口角肌、提上唇肌、提上唇鼻翼肌、提上睑肌、肩胛提肌、长回旋肌、头最长肌、颈最长肌、胸最长肌、头长肌、颈长肌、蚓状肌(在手中)、蚓状肌(在足中)、咬肌、翼内肌、内直肌、中斜角肌、多裂肌、下颌舌骨肌、头下斜肌、头上斜肌、闭孔外肌、闭孔内肌、枕肌、肩胛舌骨肌、小指对掌肌、拇对掌肌、眼轮匝肌、口轮匝肌、骨间掌侧肌、掌短肌、掌长肌、耻骨肌、胸大肌、胸小肌、腓骨短肌、腓骨长肌、第三腓骨肌、梨状肌、骨间足底肌、跖肌、颈阔肌、腘肌、后斜角肌、旋前方肌、旋前圆肌、腰大肌、股方肌、跖方肌、头前直肌、头外侧直肌、头后大直肌、头后小直肌、股直肌、大菱形肌、小菱形肌、笑肌、缝匠肌、小斜角肌、半膜肌、头半棘肌、颈半棘肌、胸半棘肌、半腱肌、前锯肌、短回旋肌、比目鱼肌、头棘肌、颈棘肌、胸棘肌、头夹肌、颈夹肌、胸锁乳突肌、胸骨舌骨肌、胸骨甲状肌、茎突舌骨肌、锁骨下肌、肩胛下肌、上孖肌、上斜肌、上直肌、旋后肌、冈上肌、颞肌、阔筋膜张肌、大圆肌、小圆肌、胸肌(thoracis)、甲状舌骨肌、胫骨前肌、胫骨后肌、斜方肌、肱三头肌、股中间肌、股外侧肌、股内侧肌、颧大肌和颧小肌,以及如本领域已知的任何其他合适的骨骼肌。
病毒载体和/或衣壳可以通过静脉内施用、动脉内施用、腹膜内施用、肢体灌注(任选地,腿和/或臂的隔离肢体灌注;参见例如arruda等人(2005)blood105:3458-3464)、和/或直接肌内注射而递送至骨骼肌。在具体实施方案中,病毒载体和/或衣壳通过肢体灌注,任选隔离肢体灌注(例如通过静脉内或动脉内施用)施用于受试者(例如患有肌营养不良例如dmd的受试者)的肢体(臂和/或腿)。在本发明的实施方案中,本发明的病毒载体和/或衣壳可以无需采用“水动力”技术而有利地施用。现有技术载体的组织递送(例如对肌肉)通常通过水动力技术(例如以大体积的静脉内/静脉内施用)得到增强,所述水动力技术增加脉管系统中的压力,并且促进载体跨越内皮细胞屏障的能力。在具体实施方案中,本发明的病毒载体和/或衣壳可以在不存在水动力技术的情况下施用,例如高体积输注和/或升高的血管内压(例如大于正常收缩压,例如在血管内压中相对于正常收缩压的小于或等于5%、10%、15%、20%、25%增加)。此类方法可以降低或避免与水动力技术相关的副作用,例如水肿、神经损害和/或筋膜室综合征。
对心肌的施用包括对左心房、右心房、左心室、右心室和/或隔膜的施用。病毒载体和/或衣壳可以通过静脉内施用、动脉内施用例如主动脉内施用、直接心脏注射(例如进入左心房、右心房、左心室、右心室内)、和/或冠状动脉灌注而递送至心肌。
对膈肌的施用可以通过任何合适的方法,包括静脉内施用、动脉内施用和/或腹膜内施用。
对靶组织的递送还可以通过递送包含病毒载体和/或衣壳的储库来实现。在代表性实施方案中,将包含病毒载体和/或衣壳的储库植入骨骼肌、心肌和/或膈肌组织内,或组织可以与包含病毒载体和/或衣壳的薄膜或其他基质接触。此类可植入基质或基底在美国专利号7,201,898中描述。
在具体实施方案中,根据本发明的病毒载体和/或病毒衣壳施用于骨骼肌、膈肌和/或心肌(例如以治疗和/或预防肌营养不良、心脏病[例如pad或充血性心力衰竭])。
在代表性实施方案中,本发明用于治疗和/或预防骨骼肌、心肌和/或膈肌的病症。
在代表性实施方案中,本发明提供了治疗和/或预防有需要的受试者中的肌营养不良的方法,该方法包括:给哺乳动物受试者施用治疗或预防有效量的本发明的病毒载体,其中所述病毒载体包含编码下述的异源核酸:抗肌萎缩蛋白、小抗肌萎缩蛋白、微小抗肌萎缩蛋白、肌肉生长抑制素前肽、卵泡抑素、激活素ii型可溶性受体、igf-1、抗炎多肽如iκb显性突变体、sarcospan、肌营养相关蛋白、微小肌营养不良蛋白、层粘连蛋白α2、α-肌聚糖、β-肌聚糖、γ-肌聚糖、δ-肌聚糖、igf-1、针对肌肉生长抑制素或肌肉生长抑制素前肽的抗体或抗体片段、和/或针对肌抑制素的rnai。在具体实施方案中,病毒载体可以施用于骨骼肌、膈肌和/或心肌,如本文其他地方描述的。
或者,本发明可以进行实践,以将核酸递送至骨骼肌、心肌或膈肌,其用作多肽(例如酶)或功能rna(例如rnai、微小rna、反义rna)的生产平台,所述多肽或功能rna通常在血液中循环或用于全身递送至其他组织,以治疗和/或预防病症(例如代谢病症,例如糖尿病[例如胰岛素]、血友病[例如因子ix或因子viii]、粘多糖症[例如斯莱综合征,胡尔勒综合征,沙伊综合征,胡-射二氏综合征,亨特氏综合征,圣菲利波综合征a、b、c、d,莫尔基奥氏综合征,马-兰二氏综合征等]或溶酶体贮积症,例如戈谢病[葡糖脑苷脂酶]或法布里病[α-半乳糖苷酶a]或糖原贮积症,例如庞贝氏症[溶酶体酸性α葡糖苷酶])。用于治疗和/或预防代谢病症的其他合适蛋白在本文中描述。肌肉用作表达目标核酸的平台的用途在美国专利公开us2002/0192189中描述。
因此,作为一个方面,本发明进一步涵盖治疗和/或预防有需要的受试者中的代谢病症的方法,该方法包括:给受试者的骨骼肌施用治疗或预防有效量的本发明的病毒载体,其中所述病毒载体包含编码多肽的异源核酸,其中所述代谢病症是多肽中的缺乏和/或缺陷的结果。说明性代谢病症和编码多肽的异源核酸在本文中描述。任选地,多肽是分泌的(例如作为以其天然状态的分泌多肽,或例如通过与如本领域已知的分泌信号序列可操作地结合已工程改造为被分泌的多肽)。不受本发明的任何具体理论束缚,根据该实施方案,对骨骼肌的施用可以导致多肽分泌到全身循环内,并且递送至靶组织。将病毒载体递送至骨骼肌的方法在本文中更详细地描述。
本发明还可以进行实践,以产生反义rna、rnai或其他功能rna(例如核酶)用于全身递送。
本发明还提供了治疗和/或预防有需要的受试者中的先天性心力衰竭或pad的方法,该方法包括给哺乳动物受试者施用治疗或预防有效量的本发明的病毒载体,其中所述病毒载体包含编码例如下述的异源核酸:肌浆内质网(sarcoplasmicendoreticulum)ca2+-atp酶(serca2a)、血管生成因子、磷酸酶抑制剂i(i-1)及其片段(例如i1c)、针对受磷蛋白的rnai;受磷蛋白抑制或显性失活分子例如受磷蛋白s16e、调节受磷蛋白基因的锌指蛋白、β2-肾上腺素受体、β2-肾上腺素受体激酶(bark)、pi3激酶、calsarcan、β-肾上腺素受体激酶抑制剂(βarkct)、蛋白磷酸酶1的抑制剂1及其片段(例如i1c)、s100a1、小白蛋白、腺苷酸环化酶6型、影响g蛋白偶联受体激酶2型敲低的分子例如截短的组成型活性barkct、pim-1、pgc-1α、sod-1、sod-2、ec-sod、激肽释放酶、hif、胸腺肽-β4、mir-1、mir-133、mir-206、mir-208和/或mir-26a。
注射剂可以以常规形式制备为液体溶液或悬浮液,适合于在注射前溶解或悬浮于液体中的固体形式,或制备为乳状液。或者,可以以局部方式而不是全身方式,例如在储库或持续释放制剂中,施用本发明的病毒载体和/或病毒衣壳。进一步地,病毒载体和/或病毒衣壳可以附着至可手术植入的基质进行递送(例如如美国专利公开号us2004/0013645描述的)。本文公开的病毒载体和/或病毒衣壳可以通过任何合适手段施用于受试者的肺,任选通过施用受试者吸入的呼吸性颗粒的气溶胶悬浮液,所述呼吸性颗粒包含病毒载体和/或病毒衣壳。呼吸性颗粒可以是液体或固体。包含病毒载体和/或病毒衣壳的液体颗粒的气溶胶可以通过任何合适手段产生,例如使用压力驱动的气溶胶雾化器或超声波雾化器,如本领域技术人员已知的。参见例如美国专利号4,501,729。同样地可以通过药学领域已知的技术,用任何固体微粒药剂气溶胶发生器,来产生包含病毒载体和/或衣壳的固体颗粒的气溶胶。
病毒载体和病毒衣壳可以施用于cns的组织(例如脑、眼),并且可以有利地导致比在不存在本发明的情况下观察到的更广泛的病毒载体或衣壳分布。
在具体实施方案中,本发明的递送载体可以施用以治疗cns的疾病,包括遗传病症、神经变性病症、精神障碍和肿瘤。说明性cns疾病包括但不限于阿尔茨海默氏病、帕金森氏病、亨廷顿氏病、卡纳万病、利氏病、雷弗素姆病、图雷特综合征、原发性侧索硬化、肌萎缩侧索硬化、进行性肌萎缩、皮克氏病、肌营养不良、多发性硬化、重症肌无力、宾斯万格病、由于脊髓或头部损伤的外伤、泰萨克斯病、莱-尼二氏病、癫痫症、脑梗塞、精神障碍包括情绪障碍(例如抑郁症、双相情感障碍、持续性情感障碍、继发性情感障碍)、精神分裂症、药物依赖(例如酗酒及其他物质依赖)、神经症(例如焦虑、强迫症、躯体形式障碍、分离性障碍、悲伤、产后抑郁症)、精神病(例如幻觉和妄想)、痴呆、偏执狂、注意力缺陷障碍、性心理障碍、睡眠障碍、疼痛障碍、进食障碍或体重病症(例如肥胖、恶病质、神经性厌食症、和诱吐症(bulemia))以及癌症和cns肿瘤(例如垂体瘤)。
cns病症包括涉及视网膜、后束和视神经的眼病症(例如色素性视网膜炎、糖尿病视网膜病变及其他视网膜变性疾病、葡萄膜炎、年龄相关性黄斑变性、青光眼)。
大多数(如果并非全部)眼疾病和病症与三类适应症中的一类或多类相关:(1)血管生成、(2)炎症和(3)变性。本发明的递送载体可以用于递送抗血管生成因子;抗炎因子;延缓细胞变性,促进细胞保留,或促进细胞生长和上述的组合的因子。
例如,糖尿病视网膜病变的特征在于血管生成。糖尿病视网膜病变可以通过眼内(例如在玻璃体中)或眼周(例如在筋膜下区中)递送一种或多种抗血管生成因子进行治疗。一种或多种神经营养因子还可以眼内(例如玻璃体内)或眼周共递送。
葡萄膜炎涉及炎症。一种或多种抗炎因子可以通过眼内(例如玻璃体或眼前房)施用本发明的递送载体进行施用。
相比之下,色素性视网膜炎的特征在于视网膜变性。在代表性实施方案中,色素性视网膜炎可以通过编码一种或多种神经营养因子的递送载体的眼内施用(例如玻璃体施用)进行治疗。
年龄相关性黄斑变性涉及血管生成和视网膜变性两者。该病症可以通过眼内(例如玻璃体)施用编码一种或多种神经营养因子的本发明的递送载体,和/或眼内或眼周(例如在筋膜下区中)施用编码一种或多种抗血管生成因子的本发明的递送载体进行治疗。
青光眼的特征在于增加的眼内压和视网膜神经节细胞的丧失。青光眼的治疗包括使用本发明的递送载体施用一种或多种神经保护剂,其保护细胞免于兴奋性毒素损害。此类试剂包括眼内任选玻璃体内递送的n-甲基-d-天冬氨酸(nmda)拮抗剂、细胞因子和神经营养因子。
在其他实施方案中,本发明可以用于治疗癫痫发作,例如以降低癫痫发作的发病、发病率或严重程度。癫痫发作的治疗性处理的功效可以通过行为(例如摇晃、眼睛或口轻拍(ticks))和/或电子照相手段(大多数癫痫发作具有特色脑电图异常)进行评价。因此,本发明还可以用于治疗癫痫症,其通过随着时间过去的多重癫痫发作进行标记。
在一个代表性实施方案中,生长抑素(或其活性片段)使用本发明的递送载体施用于脑,以治疗垂体瘤。根据该实施方案,编码生长抑素(或其活性片段)的递送载体通过显微输注施用到垂体内。同样地,此类治疗可以用于治疗肢端肥大症(来自垂体的异常生长激素分泌)。生长抑素的核酸(例如genbank登录号j00306)和氨基酸(例如genbank登录号p01166;含有经加工的活性肽生长抑素-28和生长抑素-14)序列是本领域已知的。
在具体实施方案中,载体可以包含如美国专利号7,071,172中所述的分泌信号。
在本发明的代表性实施方案中,将病毒载体和/或病毒衣壳施用于cns(例如脑或眼)。病毒载体和/或衣壳可以引入脊髓、脑干(延髓、脑桥)、中脑(下丘脑、丘脑、上丘脑、脑垂体、黑质、松果体)、小脑、端脑(纹状体,大脑包括枕叶、颞叶、顶叶和额叶,皮层,基底神经节,海马和杏仁核(portaamygdala))、边缘系统、新皮层、纹状体、大脑和下丘。病毒载体和/或衣壳还可以施用于不同眼区,例如视网膜、角膜和/或视神经。
病毒载体和/或衣壳可以递送到脑脊髓液内(例如通过腰椎穿刺术)用于递送载体的更分散施用。
在其中血脑屏障已被扰乱(例如脑瘤或脑梗塞)的情况下,病毒载体和/或衣壳可以进一步血管内施用于cns。
病毒载体和/或衣壳可以通过本领域已知的任何途径施用于所需cns区,包括但不限于鞘内、眼内、脑内、心室内、静脉内(例如在糖如甘露醇的存在下)、鼻内、耳内、眼内(例如玻璃体内、视网膜下、眼前房)和眼周(例如筋膜下区(sub-tenon'sregion))递送以及肌内递送与逆行递送至运动神经元。
在具体实施方案中,病毒载体和/或衣壳通过直接注射(例如立体定位注射)在液体制剂中施用于所需cns区或区室。在其他实施方案中,病毒载体和/或衣壳可以通过局部应用提供给所需区,或通过气溶胶制剂的鼻内施用提供。对眼的施用可以通过液体小滴的局部应用。作为进一步的替代方案,病毒载体和/或衣壳可以作为固体、缓慢释放制剂进行施用(参见例如美国专利号7,201,898)。
在再另外的实施方案中,病毒载体可以用于逆向转运,以治疗和/或预防涉及运动神经元的疾病和病症(例如肌萎缩侧索硬化(als);脊髓性肌萎缩(sma)等)。例如,病毒载体可以递送至肌肉组织,它可以从其中迁移到神经元内。
在该实施方案的其他方面,病毒载体将疾病或病症的严重程度降低,例如,至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%或至少95%。在该实施方案的还有其他方面,病毒载体将疾病或病症的严重程度降低,例如,约5%至约100%、约10%至约100%、约20%至约100%、约30%至约100%、约40%至约100%、约50%至约100%、约60%至约100%、约70%至约100%、约80%至约100%、约10%至约90%、约20%至约90%、约30%至约90%、约40%至约90%、约50%至约90%、约60%至约90%、约70%至约90%、约10%至约80%、约20%至约80%、约30%至约80%、约40%至约80%、约50%至约80%、或约60%至约80%、约10%至约70%、约20%至约70%、约30%至约70%、约40%至约70%或约50%至约70%。
本文公开的病毒载体可以包含足以溶解本文公开的病毒载体的量的溶剂、乳液或其他稀释剂。在该实施方案的其他方面,本文公开的病毒载体可以包含以下量的溶剂、乳液或稀释剂:例如,小于约90%(v/v)、小于约80%(v/v)、小于约70%(v/v)、小于约65%(v/v)、小于约60%(v/v)、小于约55%(v/v)、小于约50%(v/v)、小于约45%(v/v)、小于约40%(v/v)、小于约35%(v/v)、小于约30%(v/v)、小于约25%(v/v)、小于约20%(v/v)、小于约15%(v/v)、小于约10%(v/v)、小于约5%(v/v)或小于约1%(v/v)。在该实施方案的其他方面,本文公开的病毒载体可以包含以下范围内的量的溶剂、乳液或其他稀释剂:例如,约1%(v/v)至90%(v/v)、约1%(v/v)至70%(v/v)、约1%(v/v)至60%(v/v)、约1%(v/v)至50%(v/v)、约1%(v/v)至40%(v/v)、约1%(v/v)至30%(v/v)、约1%(v/v)至20%(v/v)、约1%(v/v)至10%(v/v)、约2%(v/v)至50%(v/v)、约2%(v/v)至40%(v/v)、约2%(v/v)至30%(v/v)、约2%(v/v)至20%(v/v)、约2%(v/v)至10%(v/v)、约4%(v/v)至50%(v/v)、约4%(v/v)至40%(v/v)、约4%(v/v)至30%(v/v)、约4%(v/v)至20%(v/v)、约4%(v/v)至10%(v/v)、约6%(v/v)至50%(v/v)、约6%(v/v)至40%(v/v)、约6%(v/v)至30%(v/v)、约6%(v/v)至20%(v/v)、约6%(v/v)至10%(v/v)、约8%(v/v)至50%(v/v)、约8%(v/v)至40%(v/v)、约8%(v/v)至30%(v/v)、约8%(v/v)至20%(v/v)、约8%(v/v)至15%(v/v)或约8%(v/v)至12%(v/v)。
本说明书的方面部分地公开了治疗患有疾病或病症的个体。如本文所用,术语“治疗”是指减少或消除个体中的疾病或病症的临床症状;或延迟或预防个体中的疾病或病症的临床症状的发作。例如,术语“治疗”可以意指使特征在于疾病或病症的病况的症状减少,例如,至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%或至少100%。与特定疾病或病症相关的实际症状是众所周知的,并且可以由本领域普通技术人员通过考虑因素来确定,所述因素包括但不限于疾病或病症的位置,疾病或病症的原因,疾病或病症的严重程度和/或受疾病或疾病影响的组织或器官。本领域技术人员将知道与特定类型的疾病或病症相关的适当症状或指标,并且将知道如何确定个体是否是如本文公开的治疗的候选者。
在该实施方案的方面,治疗有效量的本文公开的病毒载体使与疾病或病症相关的症状降低,例如,至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%或至少100%。在该实施方案的其他方面,治疗有效量的本文公开的病毒载体使与疾病或病症相关的症状降低,例如,至多10%、至多15%、至多20%、至多25%、至多30%、至多35%、至多40%、至多45%、至多50%、至多55%、至多60%、至多65%、至多70%、至多75%、至多80%、至多85%、至多90%、至多95%或至多100%。在该实施方案的还有其他方面,治疗有效量的本文公开的病毒载体使与疾病或病症相关的症状降低,例如,约10%至约100%、约10%至约90%、约10%至约80%、约10%至约70%、约10%至约60%、约10%至约50%、约10%至约40%、约20%至约100%、约20%至约90%、约20%至约80%、约20%至约20%、约20%至约60%、约20%至约50%、约20%至约40%、约30%至约100%、约30%至约90%、约30%至约80%、约30%至约70%、约30%至约60%或约30%至约50%。
在一个实施方案中,与未接受相同治疗的患者相比,本文公开的病毒载体能够使施用于患者的病毒载体中编码的蛋白的水平和/或量增加,例如,至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%或至少95%。在该实施方案的其他方面,与未接受相同治疗的患者相比,病毒载体能够使患有疾病或病症的个体中的疾病或病症的严重程度降低,例如,约10%至约100%、约20%至约100%、约30%至约100%、约40%至约100%、约50%至约100%、约60%至约100%、约70%至约100%、约80%至约100%、约10%至约90%、约20%至约90%、约30%至约90%、约40%至约90%、约50%至约90%、约60%至约90%、约70%至约90%、约10%至约80%、约20%至约80%、约30%至约80%、约40%至约80%、约50%至约80%、或约60%至约80%、约10%至约70%、约20%至约70%、约30%至约70%、约40%至约70%或约50%至约70%。
在该实施方案的方面,与未接受相同治疗的个体相比,治疗有效量的本文公开的病毒载体使个体中的病毒载体内编码的蛋白的量增加,例如,至少10%、至少15%、至少20%、至少25%、至少30%、至少35%、至少40%、至少45%、至少50%、至少55%、至少60%、至少65%、至少70%、至少75%、至少80%、至少85%、至少90%、至少95%或至少100%。在该实施方案的其他方面,治疗有效量的本文公开的病毒载体使个体中的疾病或病症的严重程度降低或使个体中的疾病或病症的严重程度维持,例如,至多10%、至多15%、至多20%、至多25%、至多30%、至多35%、至多40%、至多45%、至多50%、至多55%、至多60%、至多65%、至多70%、至多75%、至多80%、至多85%、至多90%、至多95%或至多100%。在该实施方案的还有其他方面,治疗有效量的本文公开的病毒载体使个体中的疾病或病症的严重程度降低或维持,例如,约10%至约100%、约10%至约90%、约10%至约80%、约10%至约70%、约10%至约60%、约10%至约50%、约10%至约40%、约20%至约100%、约20%至约90%、约20%至约80%、约20%至约20%、约20%至约60%、约20%至约50%、约20%至约40%、约30%至约100%、约30%至约90%、约30%至约80%、约30%至约70%、约30%至约60%或约30%至约50%。
将病毒载体施用于个体或患者。个体或患者通常是人类,但可以是动物,包括但不限于狗、猫、禽类、牛、马、绵羊、山羊、爬行动物和其他动物,无论是否驯养。
在一个实施方案中,本发明的病毒载体可用于产生靶向特定组织的aav,所述特定组织包括但不限于中枢神经系统、视网膜、心脏、肺、骨骼肌和肝脏。这些靶向的病毒载体可用于治疗组织特异性的疾病,或用于产生在特定正常组织中内源性产生的蛋白,诸如因子ix(fix)、因子viii、fviii和本领域中已知的其他蛋白。
中枢神经系统疾病
在一个实施方案中,中枢神经系统疾病可以使用aav治疗,其中所述aav包含可以是任何aav血清型的受体aav和选自aav1、aav2、aav3、aav4、aav5、aav7、aav8、aav9或aav10中的一种或多种的供体衣壳。在一个实施方案中,所述受体aav是aav2,且所述供体衣壳选自aav1、aav2、aav3、aav4、aav5、aav7、aav8、aav9或aav10中的一种或多种。在另一个实施方案中,所述受体aav是aav3,且所述供体衣壳选自aav1、aav2、aav3、aav4、aav5、aav7、aav8、aav9或aav10中的一种或多种。
视网膜疾病
在一个实施方案中,视网膜疾病可以使用aav治疗,其中所述aav包含可以是任何aav血清型的受体aav和选自aav1、aav2、aav3、aav4、aav5、aav7、aav8、aav9或aav10中的一种或多种的供体衣壳。在一个实施方案中,所述受体aav是aav2,且所述供体衣壳选自aav1、aav2、aav3、aav4、aav5、aav7、aav8、aav9或aav10中的一种或多种。在另一个实施方案中,所述受体aav是aav3,且所述供体衣壳选自aav1、aav2、aav3、aav4、aav5、aav7、aav8、aav9或aav10。
心脏疾病
在一个进一步实施方案中,心脏疾病可以使用aav治疗,其中所述aav包含可以是任何aav血清型的受体aav和选自aav1、aav3、aav4、aav6或aav9中的一种或多种的供体衣壳。在一个额外实施方案中,所述受体aav是aav2,且所述供体衣壳选自aav1、aav3、aav4、aav6或aav9中的一种或多种。在另一个实施方案中,所述受体aav是aav3,且所述供体衣壳选自aav1、aav3、aav4、aav6或aav9中的一种或多种。
肺部疾病
在一个实施方案中,肺部疾病可以使用aav治疗,其中aav血清型包含可以是任何aav血清型的受体aav和选自aav1、aav5、aav6、aav9或aav10中的一种或多种的供体衣壳。在另一个实施方案中,所述受体aav是aav2,且所述供体衣壳选自aav1、aav5、aav6、aav9或aav10中的一种或多种。在一个进一步实施方案中,所述受体aav是aav3,且所述供体衣壳选自aav1、aav5、aav6、aav9或aav10中的一种或多种。
骨骼肌疾病
在一个进一步实施方案中,骨骼肌疾病可以使用aav治疗,其中aav血清型包含可以是任何aav血清型的受体aav和选自aav1、aav2、aav6、aav7、aav8或aav9中的一种或多种的供体衣壳。在另一个实施方案中,所述受体aav是aav2,且所述供体衣壳选自aav1、aav2、aav6、aav7、aav8或aav9中的一种或多种。在一个实施方案中,所述受体aav是aav3,且所述供体衣壳选自aav1、aav2、aav6、aav7、aav8或aav9中的一种或多种。
肝脏疾病
在一个实施方案中,肝脏疾病可以使用aav治疗,其中aav血清型包含可以是任何aav血清型的受体aav和选自aav2、aav3、aav6、aav7、aav8或aav9中的一种或多种的供体衣壳。在一个额外实施方案中,所述受体aav是aav2,且所述供体衣壳选自aav2、aav3、aav6、aav7、aav8或aav9中的一种或多种。在一个进一步实施方案中,所述受体aav是aav3,且所述供体衣壳选自aav2、aav3、aav6、aav7、aav8或aav9中的一种或多种。
在一些实施方案中,本申请可以在任何以下段落中定义:
1.分离的aav病毒粒子,其具有至少两种来自由aav衣壳蛋白vp1、vp2和vp3组成的组的病毒结构蛋白,其中所述两种病毒蛋白足以形成将aav基因组衣壳化的aav病毒粒子,且其中存在的病毒结构蛋白中的至少一种来自与另一种病毒结构蛋白不同的血清型,且其中vp1仅来自一种血清型,vp2仅来自一种血清型,且vp3仅来自一种血清型。
2.段落1的分离的aav病毒粒子,其中所有三种病毒结构蛋白均存在。
3.段落2的分离的aav病毒粒子,其中所有三种病毒结构蛋白均来自不同的血清型。
4.段落2的分离的aav病毒粒子,其中三种结构蛋白中仅一种来自不同的血清型。
5.段落4的分离的aav病毒粒子,其中与其他两种病毒结构蛋白不同的一种病毒结构蛋白是vp1。
6.段落4的分离的aav病毒粒子,其中与其他两种病毒结构蛋白不同的一种病毒结构蛋白是vp2。
7.段落4的分离的aav病毒粒子,其中与其他两种病毒结构蛋白不同的一种病毒结构蛋白是vp3。
8.段落1-7的病毒粒子的基本上均质的群体,其中所述群体是至少101个病毒粒子。
9.段落8的病毒粒子的基本上均质的群体,其中所述群体是至少107个病毒粒子。
10.段落8的病毒粒子的基本上均质的群体,其中所述群体是至少107至1015个病毒粒子。
11.段落8的病毒粒子的基本上均质的群体,其中所述群体是至少109个病毒粒子。
12.段落8的病毒粒子的基本上均质的群体,其中所述群体是至少1010个病毒粒子。
13.段落8的病毒粒子的基本上均质的群体,其中所述群体是至少1011个病毒粒子。
14.段落10的病毒粒子的基本上均质的群体,其中病毒粒子的群体是至少95%均质的。
15.段落10的病毒粒子的基本上均质的群体,其中病毒粒子的群体是至少99%均质的。
16.产生腺相关病毒(aav)病毒粒子的方法,其包括在形成aav病毒粒子的条件下使细胞与第一核酸序列和第二核酸序列接触,其中所述aav病毒粒子由至少vp1和vp3病毒结构蛋白形成,其中第一核酸仅编码来自第一aav血清型的vp1,但不能表达vp3,且第二核酸序列编码来自不同于第一aav血清型的仅第二aav血清型的vp3,且进一步不能表达vp1,且其中,所述aav病毒粒子包含来自仅第一血清型的vp1和来自仅第二血清型的vp3,且如果表达vp2,则其仅来自一种血清型。
17.段落16的方法,其中所述第一核酸在vp2和vp3的起始密码子中具有突变,所述突变阻止从由第一核酸转录的rna翻译vp2和vp3,且进一步其中,所述第二核酸在vp1的起始密码子中具有突变,所述突变阻止从由第二核酸转录的rna翻译vp1。
18.段落16的方法,其中表达来自仅一种血清型的vp2。
19.段落18的方法,其中vp2来自与vp1不同的血清型和与vp3不同的血清型。
20.段落18的方法,其中vp2来自与vp1相同的血清型。
21.段落18的方法,其中vp2来自与vp3相同的血清型。
22.段落16的方法,其中第一aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10、aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
23.段落16的方法,其中第二aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
24.段落18的方法,其中aav病毒粒子由vp1、vp2和vp3衣壳蛋白形成,其中所述病毒结构蛋白在来自仅第一aav血清型的第一核酸和来自不同于第一aav血清型的仅第二aav血清型的第二核酸中编码,且进一步其中,所述第一核酸在a2剪接受体位点中具有突变,且进一步其中,所述第二核酸在a1剪接受体位点中具有突变,且其中,所述多倍体aav病毒粒子包含来自仅第一血清型的vp1和来自仅第二血清型的vp2和vp3。
25.段落24的方法,其中第一aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
26.段落24的方法,其中第二aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
27.段落18的方法,其中所述病毒结构蛋白在来自不同于第二和第三血清型的仅第一aav血清型的第一核酸序列、来自不同于第一和第三aav血清型的仅第二aav血清型的第二核酸序列和来自不同于第一和第二aav血清型的仅第三aav血清型的第三核酸序列中编码,且,进一步其中,所述第一核酸序列在vp2和vp3的起始密码子中具有突变,所述突变阻止从由第一核酸转录的rna翻译vp2和vp3,且进一步其中,所述第二核酸序列在vp1和vp3的起始密码子中具有突变,所述突变阻止从由第二核酸序列转录的rna翻译vp1和vp3,且进一步其中,所述第三核酸序列在vp1和vp2的起始密码子中具有突变,所述突变阻止从由第三核酸转录的rna翻译vp1和vp2,且其中,所述aav病毒粒子包含来自仅第一血清型的vp1、来自仅第二血清型的vp2和来自仅第三血清型的vp3。
28.段落27的方法,其中第一aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
29.段落27的方法,其中第二aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
30.段落27的方法,其中第三aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
31.段落18的方法,其中,所述第一核酸序列具有vp2和vp3的起始密码子中的阻止从由第一核酸序列转录的rna翻译vp2和vp3的突变和a2剪接受体位点中的突变,且进一步其中,所述第二核酸序列具有vp1的起始密码子中的阻止从由第二核酸序列转录的rna翻译vp1的突变和a1剪接受体位点的突变,且其中,所述aav多倍体衣壳包含来自仅第一血清型的vp1和来自仅第二血清型的vp2和vp3。
32.段落31的方法,其中第一aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
33.段落31的方法,其中第二aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
34.段落18的方法,其中所述病毒结构蛋白在第一核酸序列中编码,所述第一核酸序列通过两种或更多种不同aav血清型的dna改组来产生,且进一步其中,vp2和vp3的起始密码子被突变,使得vp2和vp3不能从由第一核酸序列转录的rna翻译,且进一步其中,衣壳蛋白在来自仅单一aav血清型的第二核酸中编码,其中,所述第二核酸在vp1的起始密码子中具有突变,所述突变阻止从由第二核酸转录的rna翻译vp1,且其中,所述多倍体aav衣壳包含来自通过dna改组产生的第一核酸序列的vp1和来自仅第二血清型的vp2和vp3。
35.段落18的方法,其中所述病毒结构蛋白在第一核酸序列中编码,所述第一核酸序列通过两种或更多种不同aav血清型的dna改组来产生,且进一步其中,vp2和vp3的起始密码子被突变,使得vp2和vp3不能从由第一核酸转录的rna翻译,且第一核酸的a2剪接受体位点被突变,且进一步其中,衣壳蛋白在来自仅单一aav血清型的第二核酸序列中编码,其中所述第二核酸具有vp1的起始密码子中的阻止从由第二核酸转录的rna翻译vp1的突变和a1剪接受体位点的突变,且其中,所述多倍体aav衣壳包含来自通过dna改组产生的第一核酸的vp1和来自仅第二血清型的vp2和vp3。
36.段落15的病毒粒子,其中所述aav血清型选自aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10、aav11,选自表1或表3的aav,和每种aav的任何嵌合体。
37.通过段落16的方法产生的病毒粒子的基本上均质的群体。
38.通过段落18的方法产生的病毒粒子的基本上均质的群体。
39.段落38的aav病毒粒子,其中所述异源基因编码用于治疗疾病的蛋白。
40.段落39的aav病毒粒子,其中所述疾病选自溶酶体贮积症诸如粘多糖贮积症(例如斯莱综合征[β-葡糖醛酸酶]、胡尔勒综合征[α-l-艾杜糖醛酸酶]、沙伊综合征[α-l-艾杜糖醛酸酶]、胡-射二氏(hurler-scheie)综合征[α-l-艾杜糖醛酸酶]、亨特氏综合征[艾杜糖醛酸硫酸酯酶]、圣菲利波综合征a[乙酰肝素磺酰胺酶]、b[n-乙酰氨基葡糖苷酶]、c[乙酰-coa:α-氨基葡糖苷乙酰基转移酶]、d[n-乙酰氨基葡糖-6-硫酸酯酶]、莫尔基奥氏综合征a[半乳糖-6-硫酸盐硫酸酯酶]、b[β-半乳糖苷酶]、马-兰二氏(maroteaux-lamy)综合征[n-乙酰氨基半乳糖-4-硫酸酯酶]等)、法布里病(α-半乳糖苷酶)、戈谢病(葡糖脑苷脂酶)、或糖原贮积症(例如庞贝氏症;溶酶体酸性α-葡糖苷酶)。
41.段落1-7的分离的aav病毒粒子,其中所述病毒结构蛋白中的至少一种是嵌合病毒结构蛋白。
42.段落41的分离的aav病毒粒子,其中所述嵌合病毒结构蛋白来自aav血清型,但不同于其他病毒结构蛋白。
43.段落1-7的分离的aav病毒粒子,其中所述病毒结构蛋白中无一是嵌合病毒结构蛋白。
44.段落41的分离的aav病毒粒子,其中在所述嵌合病毒结构蛋白和至少一种其他病毒结构蛋白之间的血清型没有重叠。
45.使用段落16-35的方法调节转导的方法。
46.段落45的方法,其中所述方法增强转导。
47.改变aav病毒粒子的向性的方法,其包括使用段落16-35的方法。
48.改变aav病毒粒子的免疫原性的方法,其包括使用段落16-35的方法。
49.增加组织中的载体基因组拷贝数的方法,其包括使用段落16-35的方法。
50.用于增加转基因表达的方法,其包括使用段落16-35的方法。
51.治疗疾病的方法,其包括向具有所述疾病的受试者施用有效量的段落1-7、36、43和44的病毒粒子,段落8-15、37-42和44的病毒粒子的基本上均质的群体或通过段落16-35的方法制成的病毒粒子,其中所述异源基因编码用于治疗适合通过基因疗法治疗的疾病的蛋白。
52.段落51的方法,其中所述疾病选自遗传疾病、癌症、免疫疾病、炎症、自身免疫性疾病和退行性疾病。
53.段落51和52的方法,其中进行多次施用。
54.段落53的方法,其中不同的多倍体病毒粒子用于逃逸响应于先前施用而形成的中和抗体。
55.相对于具有其全部病毒结构蛋白仅来自一种血清型的颗粒的aav载体增加转导、拷贝数和转基因表达中的至少一种的方法,其包括施用段落1-15和36-44的aav病毒粒子。
56.分离的aav病毒粒子,其具有足以形成将aav基因组衣壳化的aav病毒粒子的病毒衣壳结构蛋白,其中所述病毒衣壳结构蛋白中的至少一种不同于其他病毒衣壳结构蛋白,且其中所述病毒粒子仅含有相同类型的每种病毒衣壳蛋白。
57.段落56的分离的aav病毒粒子,其具有至少两种来自由aav衣壳蛋白vp1、vp2和vp3组成的组的病毒结构蛋白,其中所述两种病毒蛋白足以形成将aav基因组衣壳化的aav病毒粒子,且其中存在的其他病毒结构蛋白中的至少一种不同于另一种病毒结构蛋白,且其中病毒粒子仅含有相同类型的每种结构蛋白。
58.段落57的分离的aav病毒粒子,其中所有三种病毒结构蛋白均存在。
59.段落58的分离的aav病毒粒子,其进一步包含第四种aav结构蛋白。
60.段落56的分离的aav病毒粒子,其具有至少两种来自由aav衣壳蛋白vp1、vp2、vp1.5和vp3组成的组的病毒结构蛋白,其中所述两种病毒蛋白足以形成将aav基因组衣壳化的aav病毒粒子,且其中存在的病毒结构蛋白中的至少一种来自与另一种病毒结构蛋白不同的血清型,且其中vp1仅来自一种血清型,vp2仅来自一种血清型,vp1.5仅来自一种血清型,且vp3仅来自一种血清型。
61.段落57-60的分离的aav病毒粒子,其中所述病毒结构蛋白中的至少一种是不同于其他病毒结构蛋白中的至少一种的嵌合蛋白。
62.段落61的病毒粒子,其中仅vp3是嵌合的且vp1和vp2是非嵌合的。
63.段落61的病毒粒子,其中仅vp1和vp2是嵌合的,且仅vp3是非嵌合的。
64.段落63的病毒粒子,其中所述嵌合体由来自aav血清型2和8的亚基构成,且vp3来自aav血清型2。
65.段落56-64的分离的aav病毒粒子,其中所有病毒结构蛋白均来自不同的血清型。
66.段落56-64的分离的aav病毒粒子,其中所述结构蛋白中仅一种来自不同的血清型。
67.段落56-66的病毒粒子的基本上均质的群体,其中所述群体是至少107个病毒粒子。
68.段落67的病毒粒子的基本上均质的群体,其中所述群体是至少107至1015个病毒粒子。
69.段落67的病毒粒子的基本上均质的群体,其中所述群体是至少109个病毒粒子。
70.段落67的病毒粒子的基本上均质的群体,其中所述群体是至少1010个病毒粒子。
71.段落67的病毒粒子的基本上均质的群体,其中所述群体是至少1011个病毒粒子。
72.段落67-71的病毒粒子的基本上均质的群体,其中病毒粒子的群体是至少95%均质的。
73.段落72的病毒粒子的基本上均质的群体,其中病毒粒子的群体是至少99%均质的。
74.段落56-73的病毒粒子,其中所述aav血清型选自aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10、aav11,选自表1或表3的aav,和每种aav的任何嵌合体。
75.段落73的病毒粒子的基本上均质的群体。
76.段落56-74的aav病毒粒子,其中所述异源基因编码用于治疗疾病的蛋白。
77.段落76的aav病毒粒子,其中所述疾病选自溶酶体贮积症诸如粘多糖贮积症(例如斯莱综合征[β-葡糖醛酸酶]、胡尔勒综合征[α-l-艾杜糖醛酸酶]、沙伊综合征[α-l-艾杜糖醛酸酶]、胡-射二氏(hurler-scheie)综合征[α-l-艾杜糖醛酸酶]、亨特氏综合征[艾杜糖醛酸硫酸酯酶]、圣菲利波综合征a[乙酰肝素磺酰胺酶]、b[n-乙酰氨基葡糖苷酶]、c[乙酰-coa:α-氨基葡糖苷乙酰基转移酶]、d[n-乙酰氨基葡糖-6-硫酸酯酶]、莫尔基奥氏综合征a[半乳糖-6-硫酸盐硫酸酯酶]、b[β-半乳糖苷酶]、马-兰二氏(maroteaux-lamy)综合征[n-乙酰氨基半乳糖-4-硫酸酯酶]等)、法布里病(α-半乳糖苷酶)、戈谢病(葡糖脑苷脂酶)、或糖原贮积症(例如庞贝氏症;溶酶体酸性α-葡糖苷酶)。
78.段落56-60和66-77的分离的aav病毒粒子,其中所述病毒结构蛋白中无一是嵌合病毒结构蛋白。
79.段落57-78的分离的aav病毒粒子,其中在所述嵌合病毒结构蛋白和至少一种其他病毒结构蛋白之间的血清型没有重叠。
80.治疗疾病的方法,其包括向具有所述疾病的受试者施用有效量的段落56-66、74、76-79的病毒粒子或段落67-73和75的病毒粒子的基本上均质的群体,其中所述异源基因编码用于治疗适合通过基因疗法治疗的疾病的蛋白。
81.段落80的方法,其中所述疾病选自遗传疾病、癌症、免疫疾病、炎症、自身免疫性疾病和退行性疾病。
82.段落80和81的方法,其中进行多次施用。
83.段落82的方法,其中不同的多倍体病毒粒子用于逃逸响应于先前施用而形成的中和抗体。
84.段落1-7、36、39-44、56-66、74、76-79的分离的aav病毒粒子,段落8-15、37-38、67-73、75的基本上均质的群体和16-35、45-55和80-83的方法,其中申请人免于请求保护如下:在2018年3月15日提交的pct/us18/22725中的任何公开内容落入如本申请的权利要求中的任何一项或多项中所定义的发明内或在将来可在本申请或由其衍生的任何专利中提交的修改的权利要求所定义的任何发明内的程度上,以及在该或那些权利要求提交的任何一个或多个相关国家的法律提供了pct/us18/22725的公开内容是针对该或那些权利要求针对或该或那些国家的现有技术的部分的程度上,我们在此在防止本申请或由其衍生的任何专利无效的程度上保留将所述公开内容从本申请或由其衍生的任何专利的权利要求免于请求保护的权利。
例如,且非限制性地,我们保留将以下主题中的任何一项或多项从现在或将来修改的本申请的任何权利要求或由其衍生的任何专利免于请求保护的权利:
a.pct/us18/22725的实施例9中公开的任何主题;或
b.载体病毒粒子,称为多倍体载体病毒粒子,其通过转染两种aav辅助质粒或三种质粒以产生由来自不同血清型的不同衣壳亚基构成的个别多倍体载体病毒粒子而产生或可产生;或
c.载体病毒粒子,称为多倍体载体病毒粒子,其通过转染作为aav2和aav8或aav9的两种aav辅助质粒以产生由来自不同血清型的不同衣壳亚基构成的个别多倍体载体病毒粒子而产生或可产生;或
d.载体病毒粒子,称为多倍体载体病毒粒子,其通过转染作为aav2、aav8和aav9的三种aav辅助质粒以产生由来自不同血清型的不同衣壳亚基构成的个别多倍体载体病毒粒子而产生或可产生;或
e.载体病毒粒子,称为单倍体载体,其具有来自一种aav载体衣壳或aav血清型的vp1/vp2和来自替代aav载体衣壳或aav血清型的vp3,例如来自仅一种aav血清型(的衣壳)的vp1/vp2和来自仅一种替代aav血清型的vp3;或
f.选自以下的任何一种或多种aav载体病毒粒子:
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav8的vp1衣壳亚基和来自aav2的vp2/vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8或单倍体aav8/2或单倍体aav82或h-aav82),且具有来自aav8的vp1/vp2衣壳亚基和来自aav2的vp3衣壳亚基;或
载体,其中vp1/vp2衍生自不同血清型;或
载体(称为单倍体aav92或h-aav92),其具有来自aav9的vp1/vp2衣壳亚基和来自aav2的vp3衣壳亚基;或
载体(称为单倍体aav2g9或h-aav2g9),其具有来自aav8的vp1/vp2衣壳亚基和来自aav2g9的vp3衣壳亚基,其中将aav9聚糖受体结合位点移植至aav2中;或
载体(称为单倍体aav83或h-aav83),其具有来自aav8的vp1/vp2衣壳亚基和来自aav3的vp3衣壳亚基;或
载体(称为单倍体aav93或h-aav93),其具有来自aav9的vp1/vp2衣壳亚基和来自aav3的vp3衣壳亚基;或
载体(称为单倍体aavrh10-3或h-aavrh10-3),其具有来自aavrh10的vp1/vp2衣壳亚基和来自aav3的vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav2的vp1衣壳亚基和来自aav8的vp2/vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav2的vp1/vp2衣壳亚基和来自aav8的vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav8的vp1衣壳亚基和来自aav2的vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav2的vp1衣壳亚基和来自aav8的vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav2的vp1/vp2/vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav8的vp1/vp2/vp3衣壳亚基;或
称为28m-2vp3或单倍体2m-2vp3或单倍体载体28m-2vp3的载体,其中嵌合vp1/vp2衣壳亚基具有来自aav2的n-末端和来自aav8的c-末端,且vp3衣壳亚基来自aav2;或
称为嵌合aav8/2或嵌合aav82的载体,其中嵌合vp1/vp2衣壳亚基具有来自aav8的n-末端和来自aav2的c-末端且没有vp3起始密码子的突变,且vp3衣壳亚基来自aav2;或
载体,其中嵌合vp1/vp2衣壳亚基具有来自aav2的n-末端和来自aav8的c-末端;或
g.f的载体中任一种的群体,例如基本上均质的群体,例如1010个颗粒的群体,例如1010个颗粒的基本上均质的群体;或
h.产生a和/或b和/或c和/或d和/或e和/或f和/或g的载体或载体群体中任一种的方法;或
i.其任何组合。
在没有限制的情况下,我们声明,上述免于请求保护的权利的保留至少适用于本申请所附的权利要求1-30和[00437]中所述的段落1-83。修饰的病毒衣壳可以用作“衣壳媒介物”,如例如在美国专利号5,863,541中已经描述。可以被修饰的病毒衣壳包装并转移至细胞中的分子包括异源dna、rna、多肽、有机小分子、金属或其组合。
在一些实施方案中,本申请可以在任何以下段落中定义:
1.分离的aav病毒粒子,其具有三种来自由aav衣壳蛋白vp1、vp2和vp3组成的组的病毒结构蛋白,其中所述病毒蛋白足以形成将aav基因组衣壳化的aav病毒粒子,且其中存在的vp1和vp2病毒结构蛋白来自相同血清型,且vp3血清型来自替代血清型,且其中vp1和vp2仅来自单一血清型,且vp3仅来自单一血清型。
2.段落1的分离的aav病毒粒子,其中vp1和vp2来自aav血清型8或9,且vp3来自aav血清型3或2。
3.段落1的分离的aav病毒粒子,其中vp1和vp2来自aav血清型8,且vp3来自aav血清型2g9。
4.分离的aav病毒粒子,其具有三种来自由aav衣壳蛋白vp1、vp2和vp3组成的组的病毒结构蛋白,其中所述病毒蛋白足以形成将aav基因组衣壳化的aav病毒粒子,且其中存在的vp1和vp2病毒结构蛋白来自相同嵌合血清型,且vp3血清型不是嵌合血清型,且其中vp1和vp2仅来自单一嵌合血清型,且vp3仅来自单一血清型,其中vp1和vp2来自嵌合aav血清型28m,且vp3来自aav血清型2。
5.段落1的分离的aav病毒粒子,其中vp1和vp2来自aav血清型aavrh10,且vp3来自aav血清型2g9。
6.产生腺相关病毒(aav)病毒粒子的方法,其包括在形成aav病毒粒子的条件下使细胞与第一核酸序列和第二核酸序列接触,其中所述aav病毒粒子由vp1、vp2和vp3病毒结构蛋白形成,其中第一核酸仅编码来自第一aav血清型的vp1和vp2,但不能表达vp3,且第二核酸序列编码来自不同于第一aav血清型的替代aav血清型的vp3,且进一步不能表达vp1或vp2,且其中,所述aav病毒粒子包含仅来自第一血清型的vp1和vp2和仅来自第二血清型的vp3。
7.通过段落6的方法产生的aav病毒粒子。
8.段落2的方法,其中vp1和vp2来自aav血清型8或9,且vp3来自aav血清型3或2。
9.段落2的方法,其中vp1和vp2来自aav血清型8,且vp3来自aav血清型2g9。
10.产生腺相关病毒(aav)病毒粒子的方法,其包括在形成aav病毒粒子的条件下使细胞与第一核酸序列和第二核酸序列接触,其中所述aav病毒粒子由vp1、vp2和vp3病毒结构蛋白形成,其中第一核酸仅编码来自第一嵌合aav血清型的vp1和vp2,但不能表达vp3,且第二核酸序列编码来自替代aav血清型的vp3,且进一步不能表达vp1或vp2,其中vp1和vp2来自aav血清型28m,且vp3来自aav血清型2。
11.段落2的方法,其中vp1和vp2来自aav血清型aavrh10,且vp3来自aav血清型2g9。
12.单倍体载体,其具有来自一种aav载体衣壳的vp1/vp2和来自替代avp载体衣壳的vp3。
13.单倍体载体aav82(h-aav82),具有来自aav8的vp1/vp2和来自aav2的vp3。
14.单倍体载体aav92(h-aav92),具有来自aav9的vp1/vp2和来自aav2的vp3。
15.单倍体载体aav82g9(h-aav82g9),其中vp1/vp2来自aav8,且vp3来自aav2g9,其中aav2g9已将aav9聚糖受体结合位点移植至aav2中。
16.单倍体载体aav83(h-aav83),其中vp1/vp2来自aav8,且vp3来自aav3。
17.单倍体载体aav93(h-aav93),其中vp1/vp2来自aav9,且vp3来自aav3。
18.单倍体载体aavrh10-3(h-aavrh10-3),其中vp1/vp2来自aavrh10,且vp3来自aav3。
19.载体28m-2vp3(h-28m-2vp3),其中嵌合vp1/vp2衣壳亚基具有来自aav2的n-末端和来自aav8的c-末端,且vp3衣壳亚基来自aav2。
20.称为嵌合aav8/2或嵌合aav82的载体,其中嵌合vp1/vp2衣壳亚基具有来自aav8的n-末端和来自aav2的c-末端而没有vp3起始密码子的突变,且vp3衣壳亚基来自aav2。
在一些实施方案中,本申请可以在任何以下段落中定义:
1.产生多倍体腺相关病毒(aav)衣壳的方法,其包括在形成aav病毒粒子的条件下使细胞与第一核酸序列和第二核酸序列接触,其中aav衣壳由vp1、vp2和vp3衣壳蛋白形成,其中所述衣壳蛋白在来自仅第一aav血清型的第一核酸和来自不同于第一aav血清型的仅第二aav血清型的第二核酸中编码,且进一步其中,所述第一核酸在vp2和vp3的起始密码子中具有突变,所述突变阻止从由第一核酸转录的rna翻译vp2和vp3,且进一步其中,所述第二核酸在vp1的起始密码子中具有突变,所述突变阻止从由第二核酸转录的rna翻译vp1,且其中,所述多倍体aav衣壳包含来自仅第一血清型的vp1和来自仅第二血清型的vp2和vp3。
2.段落1的方法,其中第一aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10、aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
3.段落1的方法,其中第二aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
4.产生多倍体腺相关病毒(aav)衣壳的方法,其包括在形成aav病毒粒子的条件下使细胞与第一核酸序列和第二核酸序列接触,其中aav衣壳由vp1、vp2和vp3衣壳蛋白形成,其中所述衣壳蛋白在来自仅第一aav血清型的第一核酸和来自不同于第一aav血清型的仅第二aav血清型的第二核酸中编码,且进一步其中,所述第一核酸在a2剪接受体位点中具有突变,且进一步其中,所述第二核酸在a1剪接受体位点中具有突变,且其中,所述多倍体aav衣壳包含来自仅第一血清型的vp1和来自仅第二血清型的vp2和vp3。
5.段落4的方法,其中第一aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
6.段落4的方法,其中第二aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
7.产生多倍体腺相关病毒(aav)衣壳的方法,其包括在形成aav病毒粒子的条件下使细胞与第一核酸序列、第二核酸序列和第三核酸序列接触,其中aav衣壳由vp1、vp2和vp3衣壳蛋白形成,其中所述衣壳蛋白在来自不同于第二和第三血清型的仅第一aav血清型的第一核酸、来自不同于第一和第三aav血清型的仅第二aav血清型的第二核酸和来自不同于第一和第二aav血清型的仅第三aav血清型的第三核酸中编码,且,进一步其中,所述第一核酸在vp2和vp3的起始密码子中具有突变,所述突变阻止从由第一核酸转录的rna翻译vp2和vp3,且进一步其中,所述第二核酸在vp1和vp3的起始密码子中具有突变,所述突变阻止从由第二核酸转录的rna翻译vp1和vp3,且进一步其中,所述第三核酸在vp1和vp2的起始密码子中具有突变,所述突变阻止从由第三核酸转录的rna翻译vp1和vp2,且其中,所述多倍体aav衣壳包含来自仅第一血清型的vp1、来自仅第二血清型的vp2和来自仅第三血清型的vp3。
8.段落7的方法,其中第一aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
9.段落7的方法,其中第二aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
10.段落7的方法,其中第三aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
11.产生多倍体腺相关病毒(aav)衣壳的方法,其包括在形成aav病毒粒子的条件下使细胞与第一核酸序列和第二核酸序列接触,其中aav衣壳由vp1、vp2和vp3衣壳蛋白构建,其中所述衣壳蛋白在来自仅第一aav血清型的第一核酸和来自不同于第一aav血清型的仅第二aav血清型的第二核酸中编码,且进一步其中,所述第一核酸具有vp2和vp3的起始密码子中的阻止从由第一核酸转录的rna翻译vp2和vp3的突变和a2剪接受体位点中的突变,且进一步其中,所述第二核酸具有vp1的起始密码子中的阻止从由第二核酸转录的rna翻译vp1的突变和a1剪接受体位点的突变,且其中,所述aav多倍体衣壳包含来自仅第一血清型的vp1和来自仅第二血清型的vp2和vp3。
12.段落11的方法,其中第一aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
13.段落11的方法,其中第二aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
14.产生多倍体腺相关病毒(aav)衣壳的方法,其包括在形成aav病毒粒子的条件下使细胞与第一核酸和第二核酸接触,其中aav衣壳由vp1、vp2和vp3衣壳蛋白形成,其中所述衣壳蛋白在第一核酸中编码,所述第一核酸通过两种或更多种不同aav血清型的dna改组来产生,且进一步其中,vp2和vp3的起始密码子被突变,使得vp2和vp3不能从由第一核酸转录的rna翻译,且进一步其中,衣壳蛋白在来自仅单一aav血清型的第二核酸中编码,其中,所述第二核酸在vp1的起始密码子中具有突变,所述突变阻止从由第二核酸转录的rna翻译vp1,且其中,所述多倍体aav衣壳包含来自通过dna改组产生的第一核酸的vp1和来自仅第二血清型的vp2和vp3。
15.产生多倍体腺相关病毒(aav)衣壳的方法,其包括在形成aav病毒粒子的条件下使细胞与第一核酸和第二核酸接触,其中aav衣壳由vp1、vp2和vp3衣壳蛋白形成,其中所述衣壳蛋白在第一核酸中编码,所述第一核酸通过两种或更多种不同aav血清型的dna改组来产生,且进一步其中,vp2和vp3的起始密码子被突变,使得vp2和vp3不能从由第一核酸转录的rna翻译,且第一核酸的a2剪接受体位点被突变,且进一步其中,衣壳蛋白在来自仅单一aav血清型的第二核酸中编码,其中所述第二核酸具有vp1的起始密码子中的阻止从由第二核酸转录的rna翻译vp1的突变和a1剪接受体位点的突变,且其中,所述多倍体aav衣壳包含来自通过dna改组产生的第一核酸的vp1和来自仅第二血清型的vp2和vp3。
16.段落14和15的方法,其中aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
17.段落1-16中任一项的方法,其中所述aav衣壳具有基本上均质的衣壳蛋白。
18.段落17的方法,其中所述多倍体腺相关病毒(aav)基本上均质的衣壳蛋白是vp1。
19.段落17的方法,其中所述基本上均质的衣壳蛋白是vp2。
20.段落17的方法,其中所述基本上均质的衣壳蛋白是vp3。
21.段落17的方法,其中所述基本上均质的衣壳蛋白是vp1和vp2,vp1和vp3,vp2和vp3,或vp1和vp2和vp3。
22.段落1-21中任一项的方法,其中所述多倍体腺相关病毒(aav)在aav衣壳的基本上均质的群体中。
23.段落22的方法,其中所述多倍体腺相关病毒(aav)在包含仅一种血清型的衣壳蛋白vp1的aav病毒粒子的基本上均质的群体中。
24.段落22的方法,段落17的方法,其中所述多倍体腺相关病毒(aav)在包含仅一种血清型的衣壳蛋白vp2的aav病毒粒子的基本上均质的群体中。
25.段落22的方法,其中所述多倍体腺相关病毒(aav)在包含仅一种血清型的衣壳蛋白vp3的aav病毒粒子的基本上均质的群体中。
26.段落22的方法,其中所述多倍体腺相关病毒(aav)在包含仅一种血清型的衣壳蛋白vp1和vp2或仅一种血清型的vp1和vp3或仅一种血清型的vp2和vp3或仅一种血清型的vp1的aav病毒粒子基本上均质的群体中。
27.多倍体aav,其中所述多倍体aav使用段落1-26中任一项的方法制备。
28.段落1-27中任一项的多倍体aav,其中所述多倍体aav仅由vp1和vp3构建。
29.多倍体aav,其中所述多倍体aav使用段落1-28中任一项的方法制备,且进一步其中,所述多倍体aav包括异源基因。
30.段落29的多倍体aav,其中所述异源基因编码用于治疗疾病的蛋白。
31.段落30的多倍体aav,其中所述疾病选自溶酶体贮积症诸如粘多糖贮积症(例如斯莱综合征[β-葡糖醛酸酶]、胡尔勒综合征[α-l-艾杜糖醛酸酶]、沙伊综合征[α-l-艾杜糖醛酸酶]、胡-射二氏(hurler-scheie)综合征[α-l-艾杜糖醛酸酶]、亨特氏综合征[艾杜糖醛酸硫酸酯酶]、圣菲利波综合征a[乙酰肝素磺酰胺酶]、b[n-乙酰氨基葡糖苷酶]、c[乙酰-coa:α-氨基葡糖苷乙酰基转移酶]、d[n-乙酰氨基葡糖-6-硫酸酯酶]、莫尔基奥氏综合征a[半乳糖-6-硫酸盐硫酸酯酶]、b[β-半乳糖苷酶]、马-兰二氏(maroteaux-lamy)综合征[n-乙酰氨基半乳糖-4-硫酸酯酶]等)、法布里病(α-半乳糖苷酶)、戈谢病(葡糖脑苷脂酶)、或糖原贮积症(例如庞贝氏症;溶酶体酸性α-葡糖苷酶)。
在一些实施方案中,本申请可以在任何以下段落中定义:
1.分离的aav病毒粒子,其具有至少两种来自由aav衣壳蛋白vp1、vp2和vp3组成的组的病毒结构蛋白,其中所述两种病毒蛋白足以形成将aav基因组衣壳化的aav病毒粒子,且其中存在的其他病毒结构蛋白中的至少一种不同于另一种病毒结构蛋白,且其中病毒粒子仅含有相同类型的每种结构蛋白。
2.段落1的分离的aav病毒粒子,其中所有三种病毒结构蛋白均存在。
3.段落1和2的分离的aav病毒粒子,其中所述病毒结构蛋白中的至少一种是不同于其他病毒结构蛋白中的至少一种的嵌合蛋白。
4.段落3的病毒粒子,其中仅vp3是嵌合的且vp1和vp2是非嵌合的。
5.段落3的病毒粒子,其中仅vp1和vp2是嵌合的,且仅vp3是非嵌合的。
6.段落5的病毒粒子,其中所述嵌合体由来自aav血清型2和8的亚基构成,且vp3来自aav血清型2。
7.段落1-6的分离的aav病毒粒子,其中所有三种病毒结构蛋白均来自不同的血清型。
8.段落1-6的分离的aav病毒粒子,其中三种结构蛋白中仅一种来自不同的血清型。
9.段落8的病毒粒子的基本上均质的群体,其中所述群体是至少107个病毒粒子。
10.段落8的病毒粒子的基本上均质的群体,其中所述群体是至少107至1015个病毒粒子。
11.段落8的病毒粒子的基本上均质的群体,其中所述群体是至少109个病毒粒子。
12.段落8的病毒粒子的基本上均质的群体,其中所述群体是至少1010个病毒粒子。
13.段落8的病毒粒子的基本上均质的群体,其中所述群体是至少1011个病毒粒子。
14.段落9-13的病毒粒子的基本上均质的群体,其中病毒粒子的群体是至少95%均质的。
15.段落14的病毒粒子的基本上均质的群体,其中病毒粒子的群体是至少99%均质的。
16.段落1-15的病毒粒子,其中aav血清型是aav1、aav2、aav3、aav4、aav5、aav6、aav7、aav8、aav9、aav10或aav11,或选自表1或表3的aav,或每种aav的任何嵌合体。
17.段落16的病毒粒子的基本上均质的群体。
18.段落1-17的aav病毒粒子,其中所述异源基因编码用于治疗疾病的蛋白。
19.段落18的aav病毒粒子,其中所述疾病选自溶酶体贮积症诸如粘多糖贮积症(例如斯莱综合征[β-葡糖醛酸酶]、胡尔勒综合征[α-l-艾杜糖醛酸酶]、沙伊综合征[α-l-艾杜糖醛酸酶]、胡-射二氏(hurler-scheie)综合征[α-l-艾杜糖醛酸酶]、亨特氏综合征[艾杜糖醛酸硫酸酯酶]、圣菲利波综合征a[乙酰肝素磺酰胺酶]、b[n-乙酰氨基葡糖苷酶]、c[乙酰-coa:α-氨基葡糖苷乙酰基转移酶]、d[n-乙酰氨基葡糖-6-硫酸酯酶]、莫尔基奥氏综合征a[半乳糖-6-硫酸盐硫酸酯酶]、b[β-半乳糖苷酶]、马-兰二氏(maroteaux-lamy)综合征[n-乙酰氨基半乳糖-4-硫酸酯酶]等)、法布里病(α-半乳糖苷酶)、戈谢病(葡糖脑苷脂酶)、或糖原贮积症(例如庞贝氏症;溶酶体酸性α-葡糖苷酶)。
20.段落1-2和8-19的分离的aav病毒粒子,其中所述病毒结构蛋白中无一是嵌合病毒结构蛋白。
21.段落1-19的分离的aav病毒粒子,其中在所述嵌合病毒结构蛋白和至少一种其他病毒结构蛋白之间的血清型没有重叠。
22.治疗疾病的方法,其包括向具有所述疾病的受试者施用有效量的段落1-9、16、18-21的病毒粒子或段落10-15和17的病毒粒子的基本上均质的群体,其中所述异源基因编码用于治疗适合通过基因疗法治疗的疾病的蛋白。
23.段落22的方法,其中所述疾病选自遗传疾病、癌症、免疫疾病、炎症、自身免疫性疾病和退行性疾病。
24.段落22和23的方法,其中进行多次施用。
25.段落24的方法,其中不同的多倍体病毒粒子用于逃逸响应于先前施用而形成的中和抗体。
26.段落1-25的分离的aav病毒粒子,其中申请人免于请求保护如下:在2018年3月15日提交的pct/us18/22725中的任何公开内容落入如本申请的权利要求中的任何一项或多项中所定义的发明内或在将来可在本申请或由其衍生的任何专利中提交的修改的权利要求所定义的任何发明内的程度上,以及在该或那些权利要求提交的任何一个或多个相关国家的法律提供了pct/us18/22725的公开内容是针对该或那些权利要求针对或该或那些国家的现有技术的部分的程度上,我们在此在防止本申请或由其衍生的任何专利无效的程度上保留将所述公开内容从本申请或由其衍生的任何专利的权利要求免于请求保护的权利。
例如,且非限制性地,我们保留将以下主题中的任何一项或多项从现在或将来修改的本申请的任何权利要求或由其衍生的任何专利免于请求保护的权利:
a.pct/us18/22725的实施例9中公开的任何主题;或
b.载体病毒粒子,称为多倍体载体病毒粒子,其通过转染两种aav辅助质粒或三种质粒以产生由来自不同血清型的不同衣壳亚基构成的个别多倍体载体病毒粒子而产生或可产生;或
c.载体病毒粒子,称为多倍体载体病毒粒子,其通过转染作为aav2和aav8或aav9的两种aav辅助质粒以产生由来自不同血清型的不同衣壳亚基构成的个别多倍体载体病毒粒子而产生或可产生;或
d.载体病毒粒子,称为多倍体载体病毒粒子,其通过转染作为aav2、aav8和aav9的三种aav辅助质粒以产生由来自不同血清型的不同衣壳亚基构成的个别多倍体载体病毒粒子而产生或可产生;或
e.载体病毒粒子,称为单倍体载体,其具有来自一种aav载体衣壳或aav血清型的vp1/vp2和来自替代aav载体衣壳或aav血清型的vp3,例如来自仅一种aav血清型(的衣壳)的vp1/vp2和来自仅一种替代aav血清型的vp3;或
f.选自以下的任何一种或多种aav载体病毒粒子:
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav8的vp1衣壳亚基和来自aav2的vp2/vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8或单倍体aav8/2或单倍体aav82或h-aav82),且具有来自aav8的vp1/vp2衣壳亚基和来自aav2的vp3衣壳亚基;或
载体,其中vp1/vp2衍生自不同血清型;或
载体(称为单倍体aav92或h-aav92),其具有来自aav9的vp1/vp2衣壳亚基和来自aav2的vp3衣壳亚基;或
载体(称为单倍体aav2g9或h-aav2g9),其具有来自aav8的vp1/vp2衣壳亚基和来自aav2g9的vp3衣壳亚基,其中将aav9聚糖受体结合位点移植至aav2中;或
载体(称为单倍体aav83或h-aav83),其具有来自aav8的vp1/vp2衣壳亚基和来自aav3的vp3衣壳亚基;或
载体(称为单倍体aav93或h-aav93),其具有来自aav9的vp1/vp2衣壳亚基和来自aav3的vp3衣壳亚基;或
载体(称为单倍体aavrh10-3或h-aavrh10-3),其具有来自aavrh10的vp1/vp2衣壳亚基和来自aav3的vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav2的vp1衣壳亚基和来自aav8的vp2/vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav2的vp1/vp2衣壳亚基和来自aav8的vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav8的vp1衣壳亚基和来自aav2的vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav2的vp1衣壳亚基和来自aav8的vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav2的vp1/vp2/vp3衣壳亚基;或
载体,其通过转染aav2辅助质粒和aav8辅助质粒生成(称为单倍体aav2/8),且具有来自aav8的vp1/vp2/vp3衣壳亚基;或
称为28m-2vp3或单倍体2m-2vp3或单倍体载体28m-2vp3的载体,其中嵌合vp1/vp2衣壳亚基具有来自aav2的n-末端和来自aav8的c-末端,且vp3衣壳亚基来自aav2;或
称为嵌合aav8/2或嵌合aav82的载体,其中嵌合vp1/vp2衣壳亚基具有来自aav8的n-末端和来自aav2的c-末端且没有vp3起始密码子的突变,且vp3衣壳亚基来自aav2;或
载体,其中嵌合vp1/vp2衣壳亚基具有来自aav2的n-末端和来自aav8的c-末端;或
g.f的载体中任一种的群体,例如基本上均质的群体,例如1010个颗粒的群体,例如1010个颗粒的基本上均质的群体;或
h.产生a和/或b和/或c和/或d和/或e和/或f和/或g的载体或载体群体中任一种的方法;或
i.其任何组合。
在没有限制的情况下,我们声明,上述免于请求保护的权利的保留至少适用于本申请所附的权利要求1-30和[00437]中所述的段落1-83。修饰的病毒衣壳可以用作“衣壳媒介物”,如例如在美国专利号5,863,541中已经描述。可以被修饰的病毒衣壳包装并转移至细胞中的分子包括异源dna、rna、多肽、有机小分子、金属或其组合。
实施例
实施例1:多倍体腺相关病毒载体用于转导增强和中和抗体逃逸的应用
腺相关病毒(aav)载体已成功用于具有血友病和失明的患者的临床试验中。增强aav转导和逃逸中和抗体活性的有效策略的探索仍然势在必行。先前的研究已显示,来自aav血清型的衣壳的相容性和位于一个病毒粒子的不同衣壳亚基上的aavnab的识别位点。在该研究中,我们以不同比率(3:1、1:1和1:3)共转染aav2和aav8辅助质粒,以组装单倍体衣壳,并研究其转导和nab逃逸活性。单倍体病毒产率与亲本病毒相似,并且硫酸肝素结合能力与aav2衣壳输入量正相关。为了确定是否通过混合衣壳蛋白来改变这些单倍体载体的向性,通过转导人huh7和小鼠c2c12细胞系来分析单倍体病毒的转导效力(图1)。尽管在huh7细胞中单倍体载体转导低于aav2,但在c2c12细胞中,单倍体载体aav2/83:1诱导了与aav2转导相比3倍更高的转导。
在肌肉注射后,所有单倍体病毒诱导的转导都高于亲本aav载体(aav2的2至9倍),其中这些中最高的是单倍体载体aav2/81:3。在全身施用后,用单倍体aav2/81:3观察到与单独aav8相比在肝脏中4倍更高的转导。将单倍体aav2/89及其亲本载体直接注射至c57b16小鼠中的后腿的肌肉中。作为对照,还研究比率为3:1、1:1和1:3的aav2和aav8病毒的混合物。为了方便比较,一只腿用aav2注射,且对侧腿用单倍体载体注射。与aav2相比,对于亲本aav8衣壳实现类似的肌肉转导(图2)。与c2c12细胞中的结果相反,从所有单倍体病毒观察到增强的肌肉转导(图2)。单倍体载体aav2/91:1和aav2/81:3分别实现aav2的4倍和2倍更高的转导。值得注意的是,单倍体载体aav2/83:1的肌肉转导比aav2高超过6倍。然而,所有对照(注射物是物理混合亲本载体的结果)具有与aav2载体相似的转导效率。
进一步,我们将治疗因子ix盒包装至单倍体aav2/81:3衣壳中,并经由尾静脉将其注射入fix敲除小鼠中。与aav8的相比,用单倍体aav2/81:3病毒载体实现更高的fix表达和改进的表型校正。此外,单倍体病毒aav2/81:3能够逃逸aav2中和,并且具有与aav2的非常低的nab交叉反应性。
为了提高多倍体病毒的nab逃逸能力,我们通过以1:1:1的比率共转染aav2、aav8和aav9辅助质粒来产生三倍体载体aav2/8/9载体。在全身施用后,用三倍体载体aav2/8/9观察到与aav8相比在肝脏中2倍更高的转导。中和抗体分析表明,aav2/8/9载体能够从用亲本血清型免疫的小鼠血清逃逸中和抗体活性。这些结果表明,多倍体病毒可能从亲本血清型获得优势,用于增强转导和逃逸nab识别。应当在具有中和抗体阳性的患者中的未来临床试验中探索该策略。
具有不同cap基因的辅助质粒的数目不受限制,并且可以基于特定治疗方案的特定要求进行混合和匹配。
细胞系.将hek293细胞、huh7细胞和c2c12细胞在具有10%胎牛血清和10%青霉素-链霉素的dulbecco氏改良eagle氏培养基中在37℃、5%co2中维持。
重组aav病毒产生.通过三质粒转染系统产生重组aav。用9µgaav转基因质粒ptr/cba-luc、12µgaav辅助质粒和15μgad辅助质粒xx680转染hek293细胞的15-cm皿。为了生成三倍体aav2/8病毒粒子,将用于转染的aav2或aav8的量的每种辅助质粒以1:1、1:3和3:1的三种不同比率共转染。为了制备单倍体aav2/8/9载体,每种血清型的辅助质粒的比率为1:1:1。转染后60小时,收集并裂解hek293细胞。将上清液进行cscl梯度超离心。通过定量pcr测定病毒滴度。
western和免疫印迹.根据病毒滴度,在每个泳道中上样相同量的病毒粒子,随后在nupage4-10%聚丙烯酰胺bis-tris凝胶(invitrogen,carlsbad,ca)上电泳,且然后经由iblot®2dry印迹系统(invitrogen,carlsbad,ca)转移至pvdf膜。将膜与针对aav衣壳蛋白特异性的b1抗体一起孵育。
如先前所述进行天然免疫印迹测定。简言之,通过使用真空点印器将纯化的衣壳转移至hybond-ecl膜(amersham,piscataway,nj)。将膜在10%乳pbs中封闭1小时,且然后与单克隆抗体a20或adk8孵育。将膜与过氧化物酶偶联的山羊抗小鼠抗体孵育1小时。蛋白通过amershamimager600(gehealthcarebiosciences,pittsburg,pa)显现。
体外转导测定.将huh7和c2c12细胞通过重组病毒在平底、24孔板中以1x104vg/细胞进行转导。48小时后,收获细胞并通过荧光素酶测定系统(promega,madison,wi)评估。
肝素抑制测定.测定可溶性肝素抑制重组病毒与huh7或c2c12细胞的结合的能力。简言之,将aav2、aav8、单倍体病毒aav2/81:1、aav2/81:3和aav2/83:1在可溶性hs存在或不存在的情况下在dmem中在37℃下孵育1h。预孵育后,将重组病毒和可溶性hs的混合物添加至huh7或c2c12细胞中。在转导后48小时,收获细胞并通过荧光素酶测定法评估。
在体内来自单倍体aav衣壳的抗原呈递类似于aav8的抗原呈递.为了研究衣壳抗原呈递的效力,我们通过以1:3的比率转染pxr2-ova和pxr8-ova来产生单倍体aav2/8ova1:3载体。在c57bl/6小鼠中经由眼眶后注射施用1x1011vg的aav2/8-ova和aav8-ova载体。三天后,将cfse标记的ot-1小鼠脾细胞转移至c57bl/6小鼠中。在转移ot-1脾细胞后第10天,通过流式细胞术测量t细胞增殖。当与未施用aav载体的对照组小鼠相比时,ot-1t细胞增殖在接受aav2/8-ova1:3或aav8-ova的小鼠中显著增加(图5)。然而,aav2/8-ova1:3和aav8-ova组之间的ot-1细胞增殖没有差异。
动物研究.在该研究中进行的动物实验用c57bl/6小鼠和fix-/-小鼠进行。根据由unch机构动物护理和使用委员会(iacuc)批准的nih指南维持小鼠。经由眼眶后注射向六只三周龄雌性c57bl/6小鼠注射3x1010vg重组病毒。在i.p.注射d-荧光素底物(nanolightpinetop,az)后,在注射后一周,使用xenogenivislumina(caliperlifesciences,waltham,ma)对荧光素酶表达进行成像。使用livingimage(perkinelmer,waltham,ma)分析生物发光图像。对于肌肉传导,将1x1010个aav/luc颗粒注射至6周龄c57bl/6雌性的腓肠肌中。在指定的时间点对小鼠成像。
接下来,评估单倍体病毒在小鼠肝脏中的转导效率。还注射aav2和aav8病毒的混合物作为对照。经由眼眶后静脉向c57bl/6小鼠注射剂量为3x1010vg的重组病毒,并且在aav注射后第3天实施成像。在小鼠肝脏中,单倍体病毒aav2/81:3诱导最高的转导效率,甚至高于其他单倍体组合、亲本病毒和亲本aav8的混合物(图3a和3b)。单倍体载体aav2/81:3的转导效率是aav8的转导效率的约4倍(图3b)。来自其他单倍体病毒的肝脏转导低于来自亲本aav8的肝脏转导,但高于aav2的肝脏转导(图3a和3b)。注射后第7天,处死小鼠,收获肝脏,并分离基因组dna。通过qpcr测定肝脏中的荧光素酶基因拷贝数。与肝转导效率的结果不同,无论病毒组成如何,在肝脏中都发现相似的aav载体基因组拷贝数(图3c)。当将转基因表达针对基因拷贝数归一化时,单倍体载体aav2/81:3诱导比任何其他单倍体载体组合或亲本血清型最高的相对转基因表达(图3d)。
fix敲除的雄性小鼠(fixko小鼠)经由尾静脉注射接受1x1010vg。在注射后的各个时间点,从眼眶后丛收集血液。在第6周,进行小鼠出血分析。
肝脏中荧光素酶表达的定量.用于成像研究的动物在重组病毒注射后第4周处死,并收集肝脏。将肝脏切碎并在被动裂解缓冲液中均质化。在将肝脏裂解物离心后,检测上清液中的荧光素酶活性。使用bradford测定(biorad,hercules,ca)测量组织裂解物中的总蛋白浓度。
检测肝脏中的aav基因组拷贝数.通过蛋白酶k处理切碎的肝脏。通过purelinkgenomicdnaminikit(invitrogen,carlsbad,ca)分离总基因组dna。通过qpcr测定法检测荧光素酶基因。小鼠核纤层蛋白基因用作内部对照。
人fix表达、功能和尾巴出血时间测定.如先前所述进行人fix表达、一级hfix活性测定和尾巴出血时间测定。中和测定.将huh7细胞以每孔105个细胞的密度接种于48孔板中。将小鼠抗体的两倍稀释液与aav-luc(1x108vg)在37℃下孵育1小时。将混合物添加至细胞中,并在37℃下孵育48小时。细胞用被动裂解缓冲液(promega,madison,wi)裂解,并测量荧光素酶活性。将nab滴度定义为荧光素酶活性为无血清对照的50%的最高稀释度。
统计分析.数据呈现为平均值±sd。studentt检验用于实施所有统计分析。p值<0.05被认为是统计学显著性差异。
测试aav2/81:3以确定其是否会增加动物疾病模型中的治疗性转基因表达。使用人fix(hfix或人因子ix)作为治疗基因,并以1x1010vg/小鼠的剂量经由尾静脉将单倍体载体aav2/81:3/hfix注射至fix敲除(ko)小鼠中。单倍体载体编码人优化的fix转基因,并由肝脏特异性启动子ttr驱动。注射后第1、2和4周,elisa和一级因子活性分别分析循环中的hfix表达和活性。在第6周,使用尾夹测定评估体内hfix功能的失血量。与在野生型c57bl/6小鼠中用单倍体aav载体的高肝脏转导的观察一致,注射后2周后,单倍体载体aav2/81:3肝靶向产生的hfix比aav8载体高得多(图4a)。aav2/81:3的更高hfix蛋白表达与用高fix活性所预测的相关(图4b)。注射aav2/81:3/hfix的小鼠的失血量与野生型c57bl/6小鼠的失血量相似,并且远小于ko小鼠的失血量(图4c)。尽管统计上注射aav8和aav2/81:3/hfix的小鼠之间的失血量没有显著差异,但aav8小鼠的失血量比aav2/81:3小鼠的失血量多一点(图4c)。
单倍体病毒aav2/8逃逸nab的能力.为了研究单倍体病毒是否能够逃逸响应于亲本载体而生成的nab,通过免疫印迹测定法使用单克隆抗体进行nab结合测定。将含病毒基因组的颗粒的三倍稀释液吸附至硝酸纤维素膜,并用分别识别完整aav2或aav8的naba20或adk8进行探测。针对a20和adk9的单倍体病毒的中和概况与来自天然免疫印迹的数据相似。(表5)。单倍体aav2/81:3几乎完全逃逸aav2血清和a20中和,其表明该单倍体病毒具有用于具有抗aav2nab的个体的潜能(表5)。
体外单倍体病毒的表征.我们先前的研究已表明aav1、2、3和5衣壳间的衣壳相容性。通过用aav转基因和腺病毒辅助pxx6-80以不同比率转染来自两种血清型的aav辅助质粒来产生单倍体病毒。与亲本载体相比,在某些细胞系中观察到来自单倍体病毒的增强的转导。aav2针对其生物学良好表征并表征为基因递送媒介物,并且aav8由于在小鼠肝脏中的高转导而引起广泛的关注。在几项临床试验中在具有血友病的患者中已使用两种血清型。为了研究aav2型和8型衣壳形成单倍体病毒的可能性及其转导概况,我们以3:1、1:1和1:3的比率转染aav2和aav8的辅助质粒,以制备单倍体载体。所有单倍体病毒使用铯梯度纯化,并通过q-pcr滴定。单倍体病毒和亲本aav2或aav8之间的病毒产率没有显著差异。为了确定是否表达单倍体病毒的衣壳蛋白,使用识别aav2和aav8的衣壳蛋白的单克隆抗体b1对来自纯化的单倍体病毒的等效病毒基因组进行western印迹分析。在所有单倍体病毒中,都观察来自aav2和aav8的vp2衣壳的混合物,单倍体病毒中来自aav2或aav8的vp2衣壳的强度与两种辅助质粒的比率相关。这些结果表明,来自aav2和aav8的衣壳是相容的,并且能够整合至aav病毒粒子中。
为了确定是否通过混合衣壳蛋白来改变单倍体病毒的向性,通过转导人huh7和小鼠c2c12细胞系来分析单倍体病毒的转导效力。在两种细胞系中,aav8的转导效率均远低于aav2。在两种细胞系中,来自所有单倍体载体的转导均高于来自aav8的转导,并且效率与添加aav2衣壳正相关。尽管在huh7细胞中单倍体载体转导低于aav2,但在c2c12细胞中,单倍体载体aav2/83:1诱导的转导是aav2的3倍。
该体外转导数据支持病毒制备物由单倍体载体构成,但不是由个别血清型载体的混合物构成,并表明单倍体载体可以增强aav转导。硫酸肝素蛋白聚糖已被鉴定为aav2的主要受体。接下来,我们研究肝素结合能力的抑制是否改变单倍体病毒的转导。aav载体与可溶性肝素的预孵育在huh7和c2c12细胞两者中将aav2转导阻断接近100%,并且在huh7和c2c12细胞中将aav8转导分别阻断37%和56%。可溶性肝素对单倍体载体转导的抑制取决于两种细胞系中aav2衣壳的输入量。用更多的aav2衣壳输入量观察到更高的转导抑制。该结果表明,单倍体病毒可使用来自亲本载体的两种主要受体用于有效转导[图1]。
单倍体病毒的增加的肌肉转导.如上所述,在肌肉细胞系c2c12中,单倍体病毒aav2/83:1的转导效率高于aav2和aav8的转导效率。接下来,我们研究体外高传导是否被转化至小鼠肌肉组织。将aav2/8单倍体和亲本载体直接注射至c57bl/6小鼠中的后腿的肌肉中。作为对照,还研究比率为3:1、1:1和1:3的aav2和aav8病毒的混合物。为了方便比较,一只腿用aav2注射,且另一只腿用测试载体注射。对于每种病毒施用1x1010vg的总载体。与aav2相比,对于aav8实现类似的肌肉转导。与c2c12细胞中的结果相反,从所有单倍体病毒观察到增强的肌肉转导[图2]。
单倍体载体aav2/81:1和aav2/81:3分别实现aav2的4倍和2倍的转导。值得注意的是,单倍体载体aav2/83:1的肌肉转导是aav2的超过6倍。然而,所有混合病毒都具有与aav2类似的转导效率。这些结果表明单倍体病毒能够增加肌肉转导,并进一步支持由两种衣壳质粒的共转染产生的病毒是单倍体。
单倍体病毒的增强的肝脏转导.在几项临床试验中在具有血友病b的患者中,aav2和aav8已用于肝脏靶向。我们还评估单倍体病毒在小鼠肝脏中的转导效率。还注射与aav2和aav8混合的病毒作为对照。经由眼眶后静脉向c57bl/6小鼠施用剂量为3x1010vg的aav/luc载体;在aav注射后第3天实施成像。在小鼠肝脏中,单倍体病毒aav2/81:3诱导比其他单倍体、混合物病毒和甚至亲本aav8最高的转导效率[图3a和3b]。单倍体载体aav2/81:3的转导效率是aav8的转导效率的约4倍[图3b]。来自其他单倍体病毒的肝脏转导低于来自亲本载体aav8的肝脏转导,但高于aav2[图3a和3b]。注射后第7天,处死小鼠,收获肝脏,并分离基因组dna。通过qpcr测定肝脏中的荧光素酶基因拷贝数。与肝转导效率的结果不同,无论单倍体病毒还是aav血清型2和8,在肝脏中发现相似的aav载体基因组拷贝数[图3c]。当将转基因表达针对基因拷贝数归一化时,与肝脏中的转基因表达一致,单倍体载体aav2/81:3诱导比任何其他单倍体载体和血清型最高的相对转基因表达[图3d]。肝脏中单倍体病毒的转导概况与肌肉转导中的转导概况不同,在肌肉转导中,所有单倍体病毒都诱导比来自亲本血清型的转基因表达更高的转基因表达,其中来自aav2/83:1的最佳。
在血友病b小鼠模型中用单倍体载体的增强的治疗性fix表达和改进的出血表型校正.基于以上结果,单倍体载体aav2/81:3诱导比aav8高得多的肝脏转导。接下来,我们进一步测试单倍体载体aav2/81:3是否可以增加动物疾病模型中的治疗性转基因表达。我们使用人fix(hfix)作为治疗基因,并将编码人优化的fix转基因并由肝脏特异性启动子ttr驱动的单倍体载体aav2/81:3/hfix以1x1010vg/小鼠的剂量经由尾静脉注射至fix敲除(ko)小鼠中。注射后第1、2和4周,分别通过elisa和一级因子活性分析循环中的hfix表达和活性。在第6周,使用尾夹测定评估体内hfix功能的失血量。与在野生型c57bl/6小鼠中用单倍体aav载体的高肝脏转导的观察一致,注射后2周后,单倍体载体aav2/81:3肝靶向产生的hfix比aav8载体多得多[图4a]。aav2/81:3的更高hfix蛋白表达与高fix活性密切相关[图4b]。注射aav2/81:3/hfix的小鼠的失血量与野生型c57bl/6小鼠的失血量相似,并且小于ko小鼠的失血量[图4c]。然而,aav8治疗的小鼠具有比野生型小鼠更多的失血量[图4c}。这些数据现实,单倍体载体aav2/81:3增加来自肝脏的治疗性转基因表达并改进疾病表型校正。
单倍体病毒aav2/8逃逸中和抗体的能力.每种个别单倍体病毒病毒粒子由来自不同aav血清型衣壳的60个亚基构成。将来自一种血清型的一些衣壳亚基插入来自不同血清型的其他衣壳亚基可改变病毒粒子表面结构。众所周知,大多数aav单克隆抗体识别一个单个病毒粒子的不同亚基上的残基。为了研究单倍体病毒是否能够逃逸由亲本载体生成的nab,我们首先通过免疫印迹测定法使用单克隆抗体进行nab结合测定。将含病毒基因组的颗粒的三倍稀释液吸附至硝酸纤维素膜,并用分别识别完整aav2或aav8的naba20或adk8进行探测。单克隆抗体adk8或a20识别所有的单倍体病毒和具有aav2和aav8的混合物的病毒。单倍体病毒与a20的反应性通过将更多的aav2衣壳并入单倍体病毒病毒粒子中而增加。然而,无论衣壳比率如何,单倍体病毒间的抗aav8nabadk8的识别没有明显变化。值得注意的是,单倍体aav2/81:3与a20的结合远弱于亲本aav2和具有比率为1:3的aav2和8的混合物的病毒,其表明a20结合位点在单倍体aav2/81:3病毒粒子表面上被耗尽。
接下来,我们分析单倍体病毒针对来自aav免疫的小鼠的血清的免疫学概况。nab滴度用于评估血清抑制载体转导的能力。在注射后第4周从用亲本病毒处理的小鼠中收集血清。如表5中所示,针对a20或adk8的单倍体病毒的中和概况与来自天然免疫印迹的数据相似。aav8和aav2之间没有nab交叉反应性。有趣的是注意到,aav8免疫的小鼠血清针对aav8病毒和所有单倍体病毒具有相似的中和活性,而无论aav8衣壳并入的量如何,但对于与aav2和aav8混合的病毒却没有。aav8血清对混合病毒没有抑制作用,可以通过在测试的细胞系中来自aav2至aav8的优异转导来解释。然而,单倍体病毒部分逃逸来自aav2血清的中和作用。孵育病毒和抗aav2血清后,单倍体aav2/81:1的转导比亲本aav2降低16倍。单倍体病毒逃逸aav2血清nab的能力比与aav2和aav8混合的病毒高得多。令人惊讶的是,单倍体aav2/81:3几乎完全逃逸aav2血清和a20中和,表明该单倍体病毒具有用于具有抗aav2nab的个体的潜能(表5)。
用由三种血清型制成的三倍体载体提高中和抗体逃逸能力.我们上面描述的数据表明,单倍体aav2/8病毒不能逃逸aav8中和抗体活性,但具有逃逸aav2中和抗体的能力,这取决于来自aav8的整合衣壳的量。为了研究由更多血清型衣壳制成的多倍体病毒是否提高nab逃逸能力,我们以1:1:1的比率制备三倍体病毒aav2/8/9。在将三倍体载体aav2/8/9注射入小鼠后,与aav2相比,三倍体病毒aav2/8/9在肝脏中诱导的转导是aav8的2倍。在aav8和单倍体载体aav2/9和aav8/9(其中三倍体载体以1:1的比率由两种aav辅助质粒制成)间没有观察到肝脏转导的差异。注意到aav9全身性施用诱导比aav8更高的肝脏转导。当进行中和抗体测定时,当与aav2、8和9相比时,单倍体aav2/8/9载体分别使其nab逃逸能力提高约20倍、32倍和8倍(表6)。
在该研究中,从2种血清型或3种血清型的衣壳组装了多倍体aav病毒粒子。单倍体病毒与aav2主要受体肝素的结合能力取决于aav2衣壳输入量。在小鼠肌肉和肝脏中,所有单倍体病毒都实现比亲本aav2载体更高的转导效力,而单倍体病毒aav2/81:3具有比亲本aav8载体显著增强的肝脏转导。与aav8相比,单倍体病毒aav2/81:3的全身性施用以递送人fix诱导fix-/-小鼠中更高的fix表达和改善的血友病表型校正。重要的是,单倍体病毒aav2/81:3能够逃逸抗aav2血清的中和作用。将aav9衣壳整合至单倍体aav2/8病毒粒子中进一步改善中和抗体逃逸能力。
aav2的主要受体是hspg,而aav8的主要受体仍不清楚。为了研究单倍体病毒是否可以使用来自aav2和aav8两者的受体,我们进行肝素抑制测定以测试单倍体病毒结合肝素受体基序的能力。在huh7和c2c12细胞系中的肝素抑制结果支持单倍体病毒使用aav2衣壳的肝素受体基序用于有效转导。在一定程度上,在肝素存在的情况下,aav8也显示降低的转导效率,但转导效率仍高于aav2的转导效率。
在临床试验中有效转导的最具挑战性的方面之一是针对aav载体的中和抗体的广泛存在。nab介导的aav载体的清除已变为重复施用aav基因转移的限制因素。几项研究已经探索通过识别位点的中和抗体的合理突变或定向进化方法,对aav衣壳进行遗传修饰用于nab逃逸。衣壳突变可改变aav向性和转导效率。另外,aav病毒粒子上nab结合位点的鉴定远远落后于临床试验中的载体应用,并且不可能从多血清找出所有nab结合位点。先前的研究已表明,几种aav单克隆抗体的识别位点在一个病毒粒子的不同亚基上旋转。当将aav8衣壳引入aav2病毒粒子后,单倍体病毒的a20结合能力和来自aav2免疫血清的中和活性急剧降低。将aav2衣壳整合至aav8病毒粒子中没有降低结合完整aav8单克隆抗体adk8的能力,并且没有逃逸抗aav8血清的中和活性(表5)。这表明来自多血清的所有nab识别位点可能位于aav8病毒粒子的同一亚基上。同样,该结果表明,整合至aav2病毒粒子中的aav8衣壳可在病毒细胞内运输中起主要作用。
当由三种血清型aav2、8和9的衣壳制备三倍体病毒时,不同于三倍体载体aav2/8,单倍体aav2/8/9病毒具有逃逸来自aav2、8或9免疫的小鼠的中和抗体活性血清的能力,这提示aav8和aav9共有相似的转导途径。
来自该研究的一些证据支持来自两种或三种aav辅助质粒的转染的多倍体病毒粒子组装。(1)使用western印迹分析从单倍体病毒展示两个大小不同的vp2条带。这些vp2匹配来自不同血清型的大小。(2)c2c12中的转导概况与huh7细胞相比不同。具体而言,单倍体aav2/83:1载体在huh7细胞中的转导低于aav2,但在c2c12细胞中更高。(3)与亲本载体aav2和aav8以及具有aav2和aav8的混合物的病毒相比,所有单倍体aav2/8病毒均表明更高的肌肉转导。(4)当与aav8相比时,三倍体病毒aav2/81:3具有增强的肝脏向性。(5)单倍体病毒与a20和adk8的结合模式不同于具有aav2和aav8的混合物的病毒。(6)aav2血清中和活性的概况在单倍体病毒和混合物病毒之间是不同的。(7)三倍体aav2/8/9病毒逃逸来自用任何亲本血清型免疫的小鼠的血清的中和抗体活性。
这些多倍体病毒增强体内和体外的转导效率,并且甚至逃逸来自亲本载体免疫的血清的中和。多倍体病毒用于递送治疗性转基因fix的应用能够在具有fix缺陷的小鼠中增加fix表达并改善血友病表型校正。这些结果表明,单倍体aav载体具有增强转导和逃逸nab的能力。
实施例2:通过应用合理的多倍体方法,通过将aav病毒粒子与来自一种aav载体的vp1/vp2和来自替代aav载体的vp3组装来增强来自单倍体aav载体的aav转导
在以上研究中,我们已表明已使用多倍体载体实现增加的aav转导,所述多倍体载体通过转染两种aav辅助质粒(aav2和aav8或aav9)或三种质粒(aav2、aav8和aav9)来产生。这些个别多倍体载体病毒粒子可以由来自不同血清型的不同衣壳亚基构成。例如,通过转染aav2辅助质粒和aav8辅助质粒而生成的单倍体aav2/8,可以具有在一个病毒粒子中具有不同组合的衣壳亚基,用于有效转导:来自aav8的vp1和来自aav2的vp2/vp3,或来自aav8的vp1/vp2和来自aav2的vp3,或来自aav2的vp1和来自aav8的vp2/vp3,或来自aav2的vp1/vp2和来自aav8的vp3,或来自aav8的vp1和来自aav2的vp3,或来自aav2的vp1和来自aav8的vp3,或来自aav2的vp1/vp2/vp3,或来自aav8的vp1/vp2/vp3。在以下研究中,我们发现可以从具有来自一种aav载体衣壳的vp1/vp2和来自替代aav载体衣壳的vp3的单倍体载体获得增强的转导。
通过不同aav血清型生成vp1、vp2和vp3提供两种不同的策略用于产生这些不同蛋白。有趣的是,vp蛋白从具有vp1、vp2和vp3的重叠序列的单个cap核苷酸序列翻译。
cap基因编码3种蛋白-vp1、vp2和vp3。如上图中所示,vp1含有vp2和vp3蛋白,且vp2含有vp3蛋白。因此,cap基因具有3个区段,vp1的起点–vp2的起点–vp3的起点–所有3种vp蛋白的终点。
在作为来自两种不同aav血清型(称为a和b)的cap基因的来源的情况下,存在三种cap蛋白的6种可能的组合。在一种情况下,被鉴定为血清型a(其可以是任何血清型(或嵌合或其他非天然存在的aav))的vp1仅来自第一血清型a,而被鉴定为血清型b的vp2/vp3仅来自血清型b,并且是与vp1的血清型(或嵌合或其他非天然存在的aav)不同的血清型。在一种情况下,vp1和vp2两者都仅来自第一血清型a,且vp3仅来自血清型b。产生第一血清型的vp1和第二血清型的vp2/vp3;或来自第一血清型的vp1/vp2和来自第二血清型的vp3的方法公开于本文阐述的实施例中。在一种情况下,vp1和vp3仅来自第一血清型,且vp2仅来自第二血清型。
在作为来自三种不同aav血清型(称为a、b和c)的cap基因的来源的情况下,存在三种cap蛋白的6种可能的组合。在该情况下,被鉴定为血清型a(其可以是任何血清型(或嵌合或其他非天然存在的aav))的vp1来自不同于vp2和vp3的血清型的第一血清型;被鉴定为血清型b(其为不同于vp1和vp3的血清型(或嵌合或其他非天然存在的aav)的血清型)的vp2来自第二血清型;且,被鉴定为血清型c(其为不同于vp1的血清型(或嵌合或其他非天然存在的aav)和vp2的血清型的血清型)的vp3的血清型来自第三血清型。产生第一血清型的vp1、第二血清型的vp2和第三血清型的vp3的方法公开于本文阐述的实施例中。
在一个实施方案中,当vp1被鉴定为第一血清型a且vp2和vp3被鉴定为第二血清型b时,应理解在一个实施方案中,这将意味着vp1仅来自血清型a且vp2和vp3仅来自血清型b。在另一个实施方案中,当vp1被鉴定为第一血清型a、vp2被鉴定为第二血清型b且vp3被鉴定为第三血清型c时,应理解在一个实施方案中,这将意味着vp1仅来自血清型a;vp2仅来自血清型b;且vp3仅来自血清型c。如以下实施例中更详细描述,在一个实施方案中,为了使用两种不同的血清型产生单倍体载体,可以包括仅表达来自血清型a的vp1的来自血清型a(或嵌合或其他非天然存在的aav)的vp1的核苷酸序列,和仅来自第二血清型的vp2和/或vp3或可替代地来自第二血清型的vp2和仅来自第三血清型的vp3的第二核苷酸序列(参见例如,图13-15)。在一个实施方案中,vp1/vp2仅来自第一血清型,且vp3仅来自第二血清型。
在3种不同的cap基因的情况下,可以用来自三种aav血清型的特定vp蛋白的核苷酸序列的完整拷贝来生成辅助质粒。个别cap基因将生成与该特定aav血清型(称为a、b和c)相关的vp蛋白。
在一个实施方案中,当vp1被鉴定为第一血清型a且vp2被鉴定为第二血清型b且vp3被鉴定为第三血清型c时,应理解在一个实施方案中,这将意味着vp1仅来自血清型a;vp2仅来自血清型b且vp3仅来自血清型c。如以下实施例中更详细描述,为了产生这种单倍体载体,将包括仅表达来自血清型a的vp1而不表达来自血清型a的vp2或vp3的来自血清型a的vp1的核苷酸序列;表达核苷酸b的vp2而不表达血清型b的vp3的第二核苷酸序列;和表达血清型c的vp3的第三核苷酸序列。
在某些实施方案中,单倍体病毒粒子仅包含vp1和vp3衣壳蛋白。在某些实施方案中,单倍体病毒粒子包含vp1、vp2和vp3衣壳蛋白。
应注意,在vp1与vp3形成单倍体病毒粒子的各种组合;或vp1/vp2/vp3形成单倍体病毒粒子的各种血清型组合的这些实施方案的每一个中,表达衣壳蛋白的核苷酸序列可以从一种或多种载体、例如质粒表达。在一个实施方案中,对表达vp1、vp2或vp3的核酸序列进行密码子优化,使得核苷酸序列之间的重组显著减少,尤其是当从一种载体(例如质粒等)表达时。
具有来自aav8的vp1/vp2和来自aav2的vp3的c-末端的合理单倍体载体增强aav转导.已经表明,以aav2衣壳与aav8衣壳的任何比率的单倍体载体aav2/8比aav2或具有相同比率的aav2载体和aav8载体的混合物的病毒诱导更高的肝脏转导。为了阐明个别单倍体aav2/8载体中的哪些aav亚基促成比aav2更高的转导,我们制成不同的构建体,其表达仅aav8vp1/vp2、仅aav2vp3、具有来自aav2的n-末端和来自aav8的c-末端的嵌合vp1/vp2(28m-2vp3)或来自aav8的n-末端和来自aav2的c-末端且没有vp3起始密码子的突变的嵌合aav8/2。这些质粒用于产生具有不同组合的单倍体aav载体。在经由眼眶后静脉在小鼠中注射lx1010个这些单倍体载体的颗粒后,评估肝转导效率。嵌合aav82载体(aav82)诱导比aav2略高的肝脏转导。然而,单倍体aav82(h-aav82)具有比aav2高得多的肝脏转导。观察到单倍体载体28m-2vp3的肝脏转导进一步增加。我们还将这些单倍体载体施用至小鼠的肌肉中。为了便于比较,当小鼠面朝上时,向右腿注射aav2载体,并向左腿注射单倍体载体。aav注射后第3周,拍摄图像。与在肝脏中的观察一致,所有单倍体载体和嵌合载体均具有较高的肌肉转导,其中来自单倍体载体28m-2vp3的最佳。该结果表明,具有来自aav2的n-末端和来自aav8的c-末端的嵌合vp1/vp2归因于单倍体aav82载体的高肝脏转导。
来自具有来自其他血清型的vp1/vp2和来自aav2的vp3的单倍体载体的增强的aav肝脏转导.我们已经显示,具有来自aav8的vp1/vp2和来自aav2的vp3的单倍体载体aav82如上所述增加肝脏转导。接下来,我们想检查其中vp1/vp2衍生自不同血清型的其他单倍体病毒粒子是否也增加转导。在临床前研究中,已显示aav9有效地转导不同组织。我们已制成单倍体aav92载体(h-aav92),其中vp1/vp2来自aav9,且vp3来自aav2。在全身施用后,在第1周进行成像。用h-aav92实现的肝脏转导率是aav2的约4倍。该数据表明来自其他血清型的vp1/vp2也能够增加aav2转导。
来自具有来自aav2突变体或其他血清型的vp3的单倍体载体的增强的aav肝脏转导.aav9使用聚糖作为有效转导的主要受体。在我们先前的研究中,我们已将aav9聚糖受体结合位点植入aav2中以制成aav2g9,并且发现aav2g9具有比aav2更高的肝脏向性。在本文中,我们制成单倍体载体(h-aav82g9),其中vp1/vp2来自aav8且vp3来自aav2g9。在全身性注射入小鼠后,与aav2g9相比,在h-aav82g9应用后的第1周和第2周都观察到超过10倍的肝脏转导。为了研究其中vp3来自其他血清型且vp1/vp2来自不同血清型或变体的单倍体载体,我们克隆其他构建体:仅aav3vp3、仅aavrh10vp1/vp2,并制成具有各种组合的不同单倍体载体(h-aav83、h-aav93和h-aavrh10-3)。在全身注射入小鼠后,在第1周实施成像。与从其他单倍体载体获得的结果一致,用单倍体载体(h-aav83、h-aav93和h-aavrh10-3)实现比aav3更高的肝脏转导。有趣的是注意到,基于成像特征,这些单倍体载体还诱导全身性转导,这不同于来自具有来自aav2的vp3的单倍体载体5的结果,其仅有效地转导肝脏。总之,具有来自一种血清型的vp1/vp2和来自替代血清型的vp3的单倍体载体能够增强转导并可能改变向性。
具有来自一种aav血清型的vp1/vp3和来自另一种aav血清型的vp2的单倍体载体增强aav转导并逃逸抗体中和.为了研究其中vp2来自一种血清型且vp1/vp3来自不同血清型的单倍体载体,将生成几种构建体。将生成仅表达aav2vp2的构建体。这将通过并入aav2vp1起始密码子的突变和/或aav2vp1剪接受体位点的突变(例如图10中所示)与vp3起始密码子的组合来完成。还将生成仅表达aav8vp1/3的构建体。这将通过并入aav8vp2起始密码子的突变来完成。类似地,将生成仅表达aav2vp1/3的构建体,和仅表达aav8vp2的构建体。
将使用适当的质粒和辅助病毒从这些构建体制成编码荧光素酶转基因且具有aav2vp1和aav8vp1/3或具有aav8vp1和aav2vp1/3的单倍体载体的基本上均质的群体。lx1010个这些单倍体载体的颗粒将经由眼眶后静脉注射至小鼠中,并在1周后通过成像评估肝脏转导效率。预期用单倍体载体的均质群体将实现比aav2更高的肝脏转导,并且与aav2或aav8的活性相比,用单倍体载体将看到低得多的nab交叉反应性。进一步,均质的单倍体载体群体也可以诱导全身转导(例如,如基于成像概况所鉴定),这不同于使用aav2或aav8的结果。
在这些实施例中,我们表明由来自相容血清型的vp1/vp2和vp3s制成的单倍体病毒也增加转导。将2x1010vg的aav载体全身注射入小鼠后,当与仅aav2和仅aav3衣壳相比时,发现由来自血清型7、8、9和rh10的vp1/vp2和来自aav2或aav3的vp3构成的单倍体aav载体跨多种组织类型(包括肝脏、心脏和脑)展现转导的2至7倍增加。这些组织另外在这些组织中具有更高的载体基因组拷贝数,表明非同源vp1/vp2的并入可影响aav受体结合和细胞内运输。另外,用与aav2或aav8vp3组合的aav2或aav8vp1/vp2产生嵌合和单倍体衣壳。当将这些单倍体aav载体注射入小鼠后,由aav8vp1/2和aav2vp3构成的单倍体aav载体的转导是仅由aav2vps构成的病毒的5倍。值得注意的是,与由与aav2vp3配对的aav8vp1/vp2构成的衣壳相比,由与来自aav2的vp3配对的来自嵌合aav2/8的vp1/vp2(aav2的n-末端和aav8的c-末端)构成的单倍体载体的转基因表达增加50倍。鉴于来自aav8vp3的衣壳的比例相同,差异位于aav2和aav8之间的vp1/2n-末端区域中,这可能表明aav2的vp1/2n-末端与其同源vp3之间的“通信”。总之,本文呈现的工作提供了对可增加跨多种组织类型的转导的当前aav生产策略的洞见。
单倍体载体也将被注射至小鼠的肌肉中。为了便于比较,当小鼠面朝上时,向右腿注射aav2载体,并向左腿注射单倍体载体。aav注射后第3周,拍摄图像。还预期通过单倍体载体的增强的肌肉中的转导。
单倍体病毒的均质群体逃逸中和抗体的能力.为了研究单倍体病毒是否能够逃逸由亲本载体生成的nab,通过免疫印迹测定法使用单克隆抗体进行nab结合测定。将含病毒基因组的颗粒的三倍稀释液吸附至硝酸纤维素膜,并用分别识别完整aav2或aav8的naba20或adk8进行探测。预期单倍体病毒的均质群体将大大减少至通过单克隆抗体adk8或a20不可检测的识别。
接下来,生成使用来自aav免疫的小鼠的血清的单倍体病毒的均质群体的免疫学概况。nab滴度用于评估血清抑制载体转导的能力。在注射后第4周从用亲本病毒处理的小鼠中收集血清。比较针对a20或adk8的单倍体病毒的中和概况,并预期其与从天然免疫印迹获得的数据相似。预期在aav8和aav2之间看不得nab交叉反应性。预期单倍体病毒的均质群体至少部分地且可能完全逃逸来自av2血清或aav8血清的中和。
具有来自一种aav血清型的vp2/vp3和来自另一种aav血清型的vp1的单倍体载体增强aav转导并逃逸抗体中和.为了研究其中vp1来自一种血清型且vp2/vp3来自不同血清型的单倍体载体,将生成几种构建体。将生成仅表达aav2vp1的构建体。这将通过并入aav2vp2起始密码子的突变、aav2vp3起始密码子的突变(例如,如图7和图21中所示)或vp2和vp3剪接受体位点的突变(例如,如图9中所示)或两者的突变(例如,如图11中所示)来完成。将生成仅表达aav8vp2/3的构建体。这将通过并入aav8vp1起始密码子的突变(例如,参见图21)和/或剪接受体位点(例如,参见图12)来完成。类似地,将生成仅表达aav2vp2/3的构建体,并将生成仅表达aav8vp1的构建体。
将使用适当的质粒和辅助病毒从这些构建体制成编码荧光素酶转基因且具有aav2vp1和aav8vp2/3或具有aav8vp1和aav2vp2/3的单倍体载体的基本上均质的群体。lx1010个这些单倍体载体的颗粒将经由眼眶后静脉注射至小鼠中,并在1周后通过成像评估肝脏转导效率。预期用单倍体载体的均质群体将实现比aav2更高的肝脏转导,并且与aav2或aav8的活性相比,用单倍体载体将看到低得多的nab交叉反应性。进一步,均质的单倍体载体群体也可以诱导全身转导(例如,如基于成像概况所鉴定),这不同于使用aav2或aav8的结果。
单倍体载体也将被注射至小鼠的肌肉中。为了便于比较,当小鼠面朝上时,向右腿注射aav2载体,并向左腿注射单倍体载体。aav注射后第3周,拍摄图像。还预期通过单倍体载体的增强的肌肉中的转导。
单倍体病毒的均质群体逃逸中和抗体的能力.为了研究单倍体病毒是否能够逃逸由亲本载体生成的nab,通过免疫印迹测定法使用单克隆抗体进行nab结合测定。将含病毒基因组的颗粒的三倍稀释液吸附至硝酸纤维素膜,并用分别识别完整aav2或aav8的naba20或adk8进行探测。预期单倍体病毒的均质群体将大大减少至通过单克隆抗体adk8或a20不可检测的识别。
接下来,生成使用来自aav免疫的小鼠的血清的单倍体病毒的均质群体的免疫学概况。nab滴度用于评估血清抑制载体转导的能力。在注射后第4周从用亲本病毒处理的小鼠中收集血清。比较针对a20或adk8的单倍体病毒的中和概况,并预期其与从天然免疫印迹获得的数据相似。预期在aav8和aav2之间看不得nab交叉反应性。预期单倍体病毒的均质群体至少部分地且可能完全逃逸来自av2血清或aav8血清的中和。
具有来自一种aav血清型的vp1、来自另一种aav血清型的vp2和来自第三aav血清型的vp3的三倍体载体增强aav转导并逃逸抗体中和。
为了研究其中vp1、vp2和vp各自来自不同aav血清型的三倍体载体,将生成几种构建体。将生成仅表达aav2vp1的构建体。这将通过并入aav2vp2起始密码子的突变和vp3起始密码子的突变(例如,如图7中所示)或并入vp2/3的剪接受体位点的突变(例如,如图9中所示)来完成。将生成仅表达aav9vp2的构建体。这将通过并入aav9vp1起始密码子中的突变和/或并入aav9vp1剪接受体位点中的突变和vp3起始密码子的突变来完成。或者,这将通过合成省略vp1的上游编码序列的aav9cap编码序列的片段和vp3起始密码子的突变来完成。将生成仅表达aav8vp3的构建体。这将通过并入aav8vp1起始密码子和/或剪接受体位点中的突变和并入aav8vp2起始密码子中的突变来完成。或者,这将通过合成省略vp1和vp2的上游编码序列的aav8cap编码序列的片段来完成。
将使用适当的质粒和辅助病毒从这些构建体制成编码荧光素酶转基因且具有aav2vp1、aav9vp2和aav8vp3的三倍体载体的基本上均质的群体。lx1010个这些三倍体载体的颗粒将经由眼眶后静脉注射至小鼠中,并在1周后通过成像评估肝脏转导效率。预期用三倍体载体的均质群体将实现比aav2、aav9或aav8更高的肝脏转导,并且与用aav2、aav8或aav8的活性相比,用三倍体载体将看到低得多的nab交叉反应性。进一步,均质的三倍体载体群体也可以诱导全身转导(例如,如基于成像概况所鉴定)。
三倍体载体也将被注射至小鼠的肌肉中。为了便于比较,当小鼠面朝上时,向右腿注射aav2载体、aav9载体或aav8载体,并向左腿注射三倍体载体。aav注射后第3周,拍摄图像。预期通过三倍体载体的增强的肌肉中的转导。
三倍体病毒的均质群体逃逸中和抗体的能力.每种个别单倍体病毒病毒粒子由来自相应不同aav血清型衣壳的60个亚基构成。预期组合衍生自三种不同血清型的血清型衣壳蛋白改变病毒粒子表面结构。众所周知,大多数aav单克隆抗体识别一个单个病毒粒子的不同亚基上的残基。为了研究三倍体病毒是否能够逃逸由亲本载体生成的nab,通过免疫印迹测定法使用单克隆抗体进行nab结合测定。将含病毒基因组的颗粒的三倍稀释液吸附至硝酸纤维素膜,并用分别识别完整aav2或aav8的naba20或adk8进行探测。预期三倍体病毒的均质群体将大大减少至通过单克隆抗体adk8或a20不可检测的识别。
接下来,生成使用来自aav免疫的小鼠的血清的三倍体病毒的均质群体的免疫学概况。nab滴度用于评估血清抑制载体转导的能力。在注射后第4周从用亲本病毒处理的小鼠中收集血清。比较针对a20或adk8的三倍体病毒的中和概况,并预期其与从天然免疫印迹获得的数据相似。预期在aav8和aav2之间看不得nab交叉反应性。预期三倍体病毒的均质群体至少部分地且可能完全逃逸来自aav2血清、aav9血清或aav8血清的中和。
实施例3:多倍体腺相关病毒载体增强转导并逃逸中和抗体
腺相关病毒(aav)载体已成功用于具有血友病和失明的患者的临床试验中。尽管aav载体的应用在这些临床试验中已证明是安全的并显示治疗效果,但主要挑战之一是其低感染性,其需要相对大量的病毒基因组。另外,群体中的大部分在血液和其他体液中具有针对aav的中和抗体(nabs)。在未来的临床试验中,nab的存在为更广泛的aav应用提出另一个主要挑战。迫切需要增强aav转导和逃逸中和抗体活性的有效策略。先前的研究已显示,来自aav血清型的衣壳的相容性和位于一个病毒粒子的不同衣壳亚基上的aavnab的识别位点。在该研究中,我们提议研究由共转染不同aav辅助质粒产生的多倍体aav病毒是否具有增强aav转导和逃逸nab的能力。我们以不同比率(3:1、1:1和1:3)共转染aav2和aav8辅助质粒,以组装单倍体衣壳。单倍体病毒产率与亲本病毒相似,表明这两种aav衣壳是相容的。在huh7和c2cl2细胞系中,aav8的转导效率远低于来自aav2的那些;然而,来自所有单倍体载体的转导高于来自aav8的转导。单倍体载体的转导效率和硫酸肝素结合能力与整合的aav2衣壳的量正相关。这些结果表明单倍体病毒载体保留其亲本病毒特性,并利用亲本载体用于增强的转导。在肌肉注射后,所有单倍体病毒诱导的转导都高于亲本aav载体(aav2的2至9倍),其中这些中最高的是单倍体载体aav2/83:1。
在全身施用后,用单倍体载体aav2/81:3观察到在肝脏中的转导是单独的aavs的4倍。重要的是,我们将治疗因子ix盒包装至单倍体载体aav2/81:3衣壳中,并经由尾静脉将其注射入fix敲除小鼠中。与aavs的相比,用单倍体载体aav2/81:3病毒载体实现更高的fix表达和改进的表型校正。引人注目的是,单倍体病毒aav2/81:3能够逃逸aav2中和,并且具有与aav2的非常低的nab交叉反应性。但aavs中和抗体可以抑制单倍体载体aav2/8转导,其效率与aav8相同。接下来,我们通过以1:1:1的比率共转染aav2、aav8和aav9辅助质粒来产生三倍体载体aav2/8/9载体。在全身施用后,用三倍体载体aav2/8/9观察到在肝脏中的转导是aav8的2倍(图6)。中和抗体分析表明,aav2/8/9载体能够从用亲本血清型免疫的小鼠血清逃逸中和抗体活性,这不同于aav2/8三倍体载体。结果表明,多倍体病毒可能从亲本血清型获得优势,用于增强转导,且具有逃逸nab识别的能力。应当在具有中和抗体阳性的患者中的未来临床试验中探索该策略。
实施例4:aav衣壳亚基的取代增强转导并逃逸中和抗体
使用腺相关病毒(aav)载体在具有血液疾病和盲症的患者中在临床试验中已实现治疗效果。然而,两个问题限制扩展aav载体应用:aav衣壳特异性细胞毒性t细胞(ctl)和中和抗体(nabs)。用低剂量的aav载体增强aav转导将可能降低衣壳抗原载量,并希望消除衣壳ctl介导的aav转导的靶细胞的清除而不损害转基因表达。目前,由于其不同的组织向性和转导效率,已经探索12种血清型和超过100种变体或突变体用于基因递送。已经表明在aav血清型间存在衣壳的相容性,并且来自一种血清型的特定氨基酸整合至另一种aav衣壳中增强aav转导。通过利用来自不同血清型的有效aav转导的不同机制,在体内和体外使用其中aav衣壳亚基衍生自不同血清型的镶嵌病毒实现增强的aav转导。最近关于aav载体与单克隆中和抗体的相互作用的结构研究表明,nab结合一种病毒粒子表面的几种不同亚基上的残基,这表明aav病毒粒子的亚基组装的改变可能消除aavnab结合位点,且然后逃逸nab活性。我们已经表明,镶嵌aav载体能够逃逸nab活性。这些结果表明,aav衣壳亚基的取代具有增强aav转导的潜能和中和抗体逃逸的能力。
腺相关病毒(aav)载体已成功应用于具有血液疾病和盲症的患者的临床试验中。两个问题限制广泛的aav载体应用:aav衣壳特异性细胞毒性t细胞(ctl)应答介导的aav转导的靶细胞的消除和中和抗体(nabs)介导的aav转导的阻断。已经表明衣壳抗原呈递是剂量依赖性的,这表明用低剂量的aav载体增强aav转导将潜在地降低衣壳抗原载荷,并希望消除衣壳ctl介导的aav转导的靶细胞的清除而不损害转基因表达。为此目的已探索几种方法,包括:转基因盒的优化,aav衣壳的修饰以及用药剂对aav运输的干扰。aav衣壳的修饰可改变aav向性;特别是在人组织中,aav转导效率不清楚。尽管正在进行几项临床试验,但基于来自动物模型的观察凭经验选择aav载体。用于增强aav转导的药理学试剂通常具有不想要的副作用。必须开发理想策略来增强aav转导,但不因衣壳的修饰改变其向性,并且不因药物治疗而具有副作用。目前,已经探索12种血清型和超过100种变体或突变体用于基因递送。有效的aav转导涉及以下步骤,包括:经由受体和共受体在靶细胞表面上结合,内吞入内体,从内体逃逸,核进入,aav病毒粒子脱衣壳,随后进行转基因表达。为了合理地设计新型aav载体用于增强转导,我们开发了嵌合病毒:aav2.5(其中aav2突变体具有来自aav1的5aa取代)和aav2g9(其中将来自aav9的半乳糖受体移植至aav2衣壳中)。两种嵌合突变体分别在小鼠肌肉和肝脏中诱导比aav2高得多的转导。这些观察表明,这些嵌合病毒可以使用来自两种aav血清型的特性用于增强转导(例如,aav2g9使用两种主要受体-肝素和半乳糖用于有效的细胞表面结合)。基于用于病毒组装的来自不同aav血清型的衣壳亚基间的相容性和我们的初步结果(这表明来自其他血清型(1或9)的特定氨基酸整合至aav血清型2中增强肌肉和肝脏中的aav2转导),我们推理,来自其他血清型的一些衣壳亚基的取代能够通过利用来自不同血清型的有效aav转导的不同机制来增强aav转导。另外,针对天然存在的aav的预先存在的抗体已影响血友病b和其他aav基因转移研究的成功。在一般人群中,约50%携带中和抗体。已经考虑几种方法来设计逃逸nab的aav载体,包括化学修饰、aav载体的不同血清型、原位衣壳的合理设计和组合诱变以及nab滴度的生物学耗尽(空衣壳利用、b细胞耗尽和血浆置换)。这些方法具有低效率或副作用或aav向性的改变。最近关于aav载体与单克隆中和抗体的相互作用的结构研究表明,nab结合一种病毒粒子表面的几种不同亚基上的残基,这表明aav病毒粒子的亚基组装的改变可能消除aavnab结合位点,且然后逃逸nab活性。我们具有的结果强烈地支持这样的观点:新型镶嵌aav载体具有增强各种组织中的转导的潜能并能够逃逸中和抗体活性。
疾病的治疗
在用于治疗疾病(例如中枢神经系统、心脏、肺、骨骼肌和肝脏;包括例如帕金森氏症、阿尔茨海默氏病、囊性纤维化、als、杜兴氏肌营养不良症、肢带肌营养不良症、重症肌无力和血友病a或b)的以下实施例5-6的每一个中;使用实施例2的合理多倍体方法可替代地生成使用指定的aav血清型和镶嵌性生成的其中描述的衣壳病毒粒子,以生成单倍体衣壳,其中vp1仅来自第一血清型,且vp2和/或vp3仅来自第二种血清型;或,例如vp1、vp2和vp3分别来自不同的血清型。产生此类病毒粒子的替代方法例如也描述于实施例7-15中。
实施例5:用来自两种或更多种不同aav血清型的vp1/vp2/vp3治疗中枢神经系统(cns)疾病
在第一个实验中,使用两种辅助质粒。第一辅助质粒具有来自aav2的rep和cap基因,且第二辅助质粒具有来自aav2的rep基因和来自aav4的cap基因。第三质粒编码谷氨酸脱羧酶65(gad65)和/或谷氨酸脱羧酶67(gad67)的核苷酸序列,所述核苷酸序列插入两个itr之间。多倍体病毒粒子可用于将治疗性gad65和/或gad67的核酸序列衣壳化。在以下实例中,衣壳可以使用例如实施例2的合理的多倍体方法制备,以产生例如这样的单倍体衣壳,其中vp1仅来自一种血清型,vp3仅来自替代血清型,且vp2可以存在或可以不存在。当存在vp2时,其仅来自可与vp1或vp3相同、或者可来自第三血清型的一种血清型,或者衣壳可以通过导致镶嵌单倍体衣壳的上述交叉装饰方法制备。从三种质粒生成的单倍体aav含有用于治疗帕金森氏病的gad65和/或gad67蛋白的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与帕金森氏病相关的中枢神经系统组织的特异性。实际上,通过该方法产生用于治疗帕金森氏病的单倍体病毒对相关组织的特异性高于仅由aav2或aav4构成的病毒载体。
在进一步实验中,再次使用两种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav3的rep和cap基因,且第二辅助质粒具有来自aav3的rep基因和来自aav5的cap基因。第三质粒编码用于治疗巴滕氏病的cln2的核苷酸序列,其包含在第三质粒中,并已插入两个itr之间。从三种质粒生成的单倍体aav含有用于治疗巴滕氏病的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与帕金森氏病相关的中枢神经系统组织的特异性。实际上,通过该方法产生用于治疗巴滕氏病的单倍体病毒对相关中枢神经系统组织的特异性高于仅由aav3或aav5构成的病毒载体。
在另一个实验中,使用三种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav3的rep和cap基因,且第二辅助质粒具有来自aav3的rep基因和来自aav4的cap基因。第三辅助质粒具有来自aav3的rep基因和来自aav5的cap基因。第四质粒编码用于治疗阿尔茨海默氏病的神经生长因子(ngf)的核苷酸序列,其包含在第三质粒中,并已插入两个itr之间。从四种质粒生成的三倍体aav含有用于治疗阿尔茨海默氏病的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3、aav4和aav5)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与阿尔茨海默氏病相关的中枢神经系统组织的特异性。实际上,通过该方法产生用于治疗阿尔茨海默氏病的三倍体病毒对相关中枢神经系统组织的特异性高于仅由aav3、aav4或aav5构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav2的rep和来自aav2的vp1,来自aav4的vp2和来自aav5的vp3。第二质粒编码插入两个itr之间以治疗卡纳万氏病的aac的核苷酸序列。从两种质粒生成的三倍体aav含有用于治疗卡纳万氏病的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav2、aav4和aav5)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与卡纳万氏病相关的中枢神经系统组织的特异性。实际上,通过该方法产生用于治疗卡纳万氏病的三倍体病毒对相关中枢神经系统组织的特异性高于仅由aav2、aav4或aav5构成的病毒载体。
用来自两种或更多种不同aav血清型的vp1/vp2/vp3治疗心脏疾病.在一个实验中,使用两种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav2的rep和cap基因,且第二辅助质粒具有来自aav2的rep基因和来自aav6的cap基因。第三质粒编码用于治疗心脏病的蛋白的核苷酸序列,其包含在第三质粒中,并已插入两个itr之间。从三种质粒生成的单倍体aav含有用于治疗心脏病的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与心脏病相关的心脏组织的特异性。实际上,通过该方法产生用于治疗心脏病的单倍体病毒对相关心脏组织的特异性高于仅由aav2或aav6构成的病毒载体。
在进一步实验中,使用两种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav3的rep和cap基因,且第二辅助质粒具有来自aav3的rep基因和来自aav9的cap基因。第三质粒编码用于治疗心脏病的蛋白的核苷酸序列,其包含在第三质粒中,并已插入两个itr之间。从三种质粒生成的单倍体aav含有编码用于治疗心脏病的蛋白的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与心脏病相关的心脏组织的特异性。实际上,通过该方法产生用于治疗心脏病的单倍体病毒对相关心脏组织的特异性高于仅由aav3或aav9构成的病毒载体。
在一个实验中,使用三种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav3的rep和cap基因,且第二辅助质粒具有来自aav3的rep基因和来自aav6的cap基因。第三辅助质粒具有来自aav3的rep基因和来自aav9的cap基因。第四质粒含有编码用于治疗心脏病的蛋白的核苷酸序列,其包含在第三质粒中,并已插入两个itr之间。从四种质粒生成的三倍体aav含有用于治疗心脏病的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3、aav6和aav9)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与心脏病相关的心脏组织的特异性。实际上,通过该方法产生用于治疗心脏病的三倍体病毒对相关心脏组织的特异性高于仅由aav3、aav6或aav9构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav2的rep和来自aav2的vp1,来自aav3的vp2和来自aav9的vp3。第二质粒含有插入两个itr之间的编码用于治疗心脏病的蛋白的核苷酸序列。从两种质粒生成的三倍体aav编码用于治疗心脏病的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav2、aav3和aav9)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与心脏病相关的心脏组织的特异性。实际上,通过该方法产生用于治疗心脏病的三倍体病毒对相关心脏组织的特异性高于仅由aav2、aav3或aav9构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav3的rep和来自aav3的vp1,来自aav6的vp2和来自aav6的vp3。第二质粒含有插入两个itr之间的编码用于治疗心脏病的蛋白的核苷酸序列。从两种质粒生成的单倍体aav编码用于治疗心脏病的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3和aav6)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与心脏病相关的心脏组织的特异性。实际上,通过该方法产生用于治疗心脏病的单倍体病毒对相关心脏组织的特异性高于仅由aav2或aav6构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav3的rep和来自aav3的vp1,来自aav6的vp2和来自aav9的vp3。第二质粒含有插入两个itr之间的编码用于治疗心脏病的蛋白的核苷酸序列。从两种质粒生成的三倍体aav编码用于治疗心脏病的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3、aav6和aav9)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与心脏病相关的心脏组织的特异性。实际上,通过该方法产生用于治疗心脏病的三倍体病毒对相关心脏组织的特异性高于仅由aav3、aav6或aav9构成的病毒载体。
用来自两种或更多种不同aav血清型的vp1/vp2/vp3治疗肺病.在一个实验中,再次使用两种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav2的rep和cap基因,且第二辅助质粒具有来自aav9的cap基因。第三质粒编码用于治疗囊性纤维化的cftr的核苷酸序列,其插入两个itr之间。从三种质粒生成的单倍体aav含有用于治疗囊性纤维化的cftr的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与囊性纤维化相关的肺组织的特异性。实际上,通过该方法产生用于治疗囊性纤维化的单倍体病毒对相关组织的特异性高于仅由aav2或aav9构成的病毒载体。
在一个实验中,再次使用两种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav3的rep和cap基因,且第二辅助质粒具有来自aav3的rep和来自aav10的cap基因。第三质粒编码用于治疗囊性纤维化的cftr的核苷酸序列,其插入两个itr之间。从三种质粒生成的单倍体aav含有用于治疗囊性纤维化的cftr的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与囊性纤维化相关的肺组织的特异性。实际上,通过该方法产生用于治疗囊性纤维化的单倍体病毒对相关组织的特异性高于仅由aav3或aav10构成的病毒载体。
在一个实验中,使用三种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav3的rep和cap基因,且第二辅助质粒具有来自aav3的rep基因和来自aav9的cap基因。第三辅助质粒具有来自aav3的rep基因和来自aav10的cap基因。第四质粒编码用于治疗囊性纤维化的cftr的核苷酸序列,其包含在第三质粒中,并已插入两个itr之间。从四种质粒生成的三倍体aav含有用于治疗囊性纤维化的cftr的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3、aav9和aav10)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与囊性纤维化相关的肺组织的特异性。实际上,通过该方法产生用于治疗囊性纤维化的三倍体病毒对相关组织的特异性高于仅由aav3、aav9或aav10构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav2的rep和来自aav2的vp1,来自aav9的vp2和来自aav9的vp3。第二质粒编码插入两个itr之间以治疗囊性纤维化的cftr的核苷酸序列。从两种质粒生成的单倍体aav含有用于治疗囊性纤维化的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav2和aav9)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与囊性纤维化相关的中枢神经系统组织的特异性。实际上,通过该方法产生用于治疗囊性纤维化的单倍体病毒对相关组织的特异性高于仅由aav2或aav9构成的病毒载体。
在进一步实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav3的rep和来自aav2的vp1,来自aav10的vp2和来自aav10的vp3。第二质粒编码插入两个itr之间以治疗囊性纤维化的cftr的核苷酸序列。从两种质粒生成的单倍体aav含有用于治疗囊性纤维化的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3和aav10)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与囊性纤维化相关的中枢神经系统组织的特异性。实际上,通过该方法产生用于治疗囊性纤维化的单倍体病毒对相关组织的特异性高于仅由aav3或aav10构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav2的rep和来自aav2的vp1,来自aav9的vp2和来自aav10的vp3。第二质粒编码插入两个itr之间以治疗囊性纤维化的cftr的核苷酸序列。从两种质粒生成的三倍体aav含有用于治疗囊性纤维化的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav2、aav9和aav9)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与卡纳万氏病相关的中枢神经系统组织的特异性。实际上,通过该方法产生用于治疗囊性纤维化的三倍体病毒对相关组织的特异性高于仅由aav2、aav9或aav10构成的病毒载体。
用来自两种或更多种不同aav血清型的vp1/vp2/vp3治疗骨骼肌疾病.对于以下实验,骨骼肌疾病可以是,但不限于,杜兴氏肌营养不良症、肢带肌营养不良症、脑麻痹、重症肌无力和肌萎缩性侧索硬化症(als)。
在一个实验中,再次使用两种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav2的rep和cap基因,且第二辅助质粒具有来自aav2的rep和来自aav8的cap基因。第三质粒编码用于治疗骨骼肌疾病的蛋白的核苷酸序列,其插入两个itr之间。从三种质粒生成的单倍体aav含有用于治疗骨骼肌疾病的蛋白的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与骨骼肌疾病相关的骨骼肌的特异性。实际上,通过该方法产生用于治疗骨骼肌疾病的单倍体病毒对相关骨骼肌组织的特异性高于仅由aav2或aav8构成的病毒载体。
在一个实验中,再次使用两种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav3的rep和cap基因,且第二辅助质粒具有来自aav3的rep和来自aav9的cap基因。第三质粒编码用于治疗骨骼肌疾病的蛋白的核苷酸序列,其插入两个itr之间。从三种质粒生成的单倍体aav含有用于治疗骨骼肌疾病的蛋白的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与骨骼肌疾病相关的骨骼肌的特异性。实际上,通过该方法产生用于治疗骨骼肌疾病的单倍体病毒对相关骨骼肌组织的特异性高于仅由aav3或aav9构成的病毒载体。
在一个实验中,使用三种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav3的rep和cap基因,且第二辅助质粒具有来自aav3的rep基因和来自aav8的cap基因。第三辅助质粒具有来自aav3的rep基因和来自aav9的cap基因。第四质粒编码用于治疗骨骼肌疾病的蛋白的核苷酸序列,其插入两个itr之间。从四种质粒生成的三倍体aav含有用于治疗骨骼肌疾病的蛋白的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3、aav8和aav9)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与骨骼肌疾病相关的骨骼肌的特异性。实际上,通过该方法产生用于治疗骨骼肌疾病的三倍体病毒对相关组织的特异性高于仅由aav3、aav8或aav9构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav3的rep和来自aav3的vp1,来自aav9的vp2和来自aav9的vp3。第二质粒编码用于治疗骨骼肌疾病的蛋白的核苷酸序列,其插入两个itr之间。从两种质粒生成的单倍体aav含有用于治疗骨骼肌疾病的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3和aav9)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与骨骼肌疾病相关的骨骼肌组织的特异性。实际上,通过该方法产生用于治疗骨骼肌疾病的单倍体病毒对相关骨骼肌组织的特异性高于仅由aav3或aav9构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav3的rep和来自aav3的vp1,来自aav8的vp2和来自aav8的vp3。第二质粒编码用于治疗骨骼肌疾病的蛋白的核苷酸序列,其插入两个itr之间。从两种质粒生成的单倍体aav含有用于治疗骨骼肌疾病的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3和aav8)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与骨骼肌疾病相关的骨骼肌组织的特异性。实际上,通过该方法产生用于治疗骨骼肌疾病的单倍体病毒对相关骨骼肌组织的特异性高于仅由aav3或aav8构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav3的rep和来自aav3的vp1,来自aav8的vp2和来自aav9的vp3。第二质粒编码用于治疗骨骼肌疾病的蛋白的核苷酸序列,其插入两个itr之间。从两种质粒生成的三倍体aav含有用于治疗骨骼肌疾病的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3、aav8和aav9)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与骨骼肌疾病相关的骨骼肌组织的特异性。实际上,通过该方法产生用于治疗骨骼肌疾病的三倍体病毒对相关骨骼肌组织的特异性高于仅由aav3、aav8或aav9构成的病毒载体。
用来自两种或更多种不同aav血清型的vp1/vp2/vp3治疗肝脏疾病.在一个实验中,再次使用两种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav2的rep和cap基因,且第二辅助质粒具有来自aav2的rep和来自aav6的cap基因。第三质粒编码用于治疗血友病b的因子ix(fix)的核苷酸序列,其插入两个itr之间。从三种质粒生成的单倍体aav含有用于治疗骨骼肌疾病的蛋白的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与血友病b相关的fix的特异性。实际上,通过该方法产生用于治疗患有血友病b的患者中的肝脏组织的单倍体病毒对相关组织的特异性高于仅由aav2或aav6构成的病毒载体。
在一个实验中,再次使用两种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav2的rep和cap基因,且第二辅助质粒具有来自aav3的rep和来自aav7的cap基因。第三质粒编码用于治疗血友病b的因子ix(fix)的核苷酸序列,其插入两个itr之间。从三种质粒生成的单倍体aav含有用于治疗骨骼肌疾病的蛋白的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与血友病b相关的fix的特异性。实际上,通过该方法产生用于治疗患有血友病b的患者中的肝脏组织的单倍体病毒对相关组织的特异性高于仅由aav3或aav7构成的病毒载体。
在一个实验中,使用三种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav3的rep和cap基因,且第二辅助质粒具有来自aav3的rep基因和来自aav6的cap基因。第三辅助质粒具有来自aav3的rep基因和来自aav7的cap基因。第四质粒编码用于治疗血友病b的因子ix(fix)的核苷酸序列,其插入两个itr之间。从四种质粒生成的三倍体aav含有用于治疗血友病b的蛋白的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3、aav6和aav7)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与血友病b相关的肝脏组织的特异性。实际上,通过该方法产生用于治疗患有血友病b的患者中的肝脏组织的三倍体病毒对相关组织的特异性高于仅由aav3、aav6或aav7构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav2的rep和来自aav2的vp1,来自aav6的vp2和来自aav6的vp3。第二质粒编码用于治疗血友病b的fix的核苷酸序列,其插入两个itr之间。从两种质粒生成的单倍体aav含有用于治疗血友病b的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav2和aav6)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与血友病b相关的肝脏组织的特异性。实际上,通过该方法产生用于治疗患有血友病b的患者中的肝脏组织的单倍体病毒对相关组织的特异性高于仅由aav2或aav6构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav2的rep和来自aav3的vp1,来自aav7的vp2和来自aav7的vp3。第二质粒编码用于治疗血友病b的fix的核苷酸序列,其插入两个itr之间。从两种质粒生成的单倍体aav含有用于治疗血友病b的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3和aav7)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与血友病b相关的肝脏组织的特异性。实际上,通过该方法产生用于治疗患有血友病b的患者中的肝脏组织的单倍体病毒对相关组织的特异性高于仅由aav3或aav7构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav2的rep和来自aav3的vp1,来自aav6的vp2和来自aav7的vp3。第二质粒编码用于治疗血友病b的fix的核苷酸序列,其插入两个itr之间。从两种质粒生成的三倍体aav含有用于治疗血友病b的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3、aav6和aav7)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与血友病b相关的肝脏组织的特异性。实际上,通过该方法产生用于治疗患有血友病b的患者中的肝脏组织的三倍体病毒对相关组织的特异性高于仅由aav3、aav6或aav7构成的病毒载体。
在一个实验中,再次使用两种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav2的rep和cap基因,且第二辅助质粒具有来自aav2的rep和来自aav6的cap基因。第三质粒编码用于治疗血友病a的因子viii(fviii)的核苷酸序列,其插入两个itr之间。从三种质粒生成的单倍体aav含有用于治疗骨骼肌疾病的蛋白的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与血友病a相关的fviii的特异性。实际上,通过该方法产生用于治疗患有血友病a的患者中的肝脏组织的单倍体病毒对相关组织的特异性高于仅由aav2或aav6构成的病毒载体。
在一个实验中,再次使用两种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav2的rep和cap基因,且第二辅助质粒具有来自aav3的rep和来自aav7的cap基因。第三质粒编码用于治疗血友病a的fviii的核苷酸序列,其插入两个itr之间。从三种质粒生成的单倍体aav含有用于治疗骨骼肌疾病的蛋白的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与血友病a相关的fviii的特异性。实际上,通过该方法产生用于治疗患有血友病a的患者中的肝脏组织的单倍体病毒对相关组织的特异性高于仅由aav3或aav7构成的病毒载体。
在一个实验中,使用三种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。第一辅助质粒具有来自aav3的rep和cap基因,且第二辅助质粒具有来自aav3的rep基因和来自aav6的cap基因。第三辅助质粒具有来自aav3的rep基因和来自aav7的cap基因。第四质粒编码用于治疗血友病a的fviii的核苷酸序列,其插入两个itr之间。从四种质粒生成的三倍体aav含有用于治疗血友病a的fviii蛋白的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3、aav6和aav7)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与血友病b相关的肝脏组织的特异性。实际上,通过该方法产生用于治疗患有血友病a的患者中的肝脏组织的三倍体病毒对相关组织的特异性高于仅由aav3、aav6或aav7构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav2的rep和来自aav2的vp1,来自aav6的vp2和来自aav6的vp3。第二质粒编码用于治疗血友病b的fviii的核苷酸序列,其插入两个itr之间。从两种质粒生成的单倍体aav含有用于治疗血友病a的fviii的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav2和aav6)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与血友病a相关的肝脏组织的特异性。实际上,通过该方法产生用于治疗患有血友病a的患者中的肝脏组织的单倍体病毒对相关组织的特异性高于仅由aav2或aav6构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav2的rep和来自aav3的vp1,来自aav7的vp2和来自aav7的vp3。第二质粒编码用于治疗血友病a的fviii的核苷酸序列,其插入两个itr之间。从两种质粒生成的单倍体aav含有用于治疗血友病a的fviii的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3和aav7)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与血友病b相关的肝脏组织的特异性。实际上,通过该方法产生用于治疗患有血友病a的患者中的肝脏组织的单倍体病毒对相关组织的特异性高于仅由aav3或aav7构成的病毒载体。
在另一个实验中,使用一种辅助质粒,其以不同aav血清型作为rep和cap基因的来源。辅助质粒具有来自aav2的rep和来自aav3的vp1,来自aav6的vp2和来自aav7的vp3。第二质粒编码用于治疗血友病a的fviii的核苷酸序列,其插入两个itr之间。从两种质粒生成的三倍体aav含有用于治疗血友病b的fviii的核苷酸序列,这部分通过经由根据本发明的方法使用多种aav血清型(例如,aav3、aav6和aav7)来作为编码vp1、vp2和vp3的蛋白的来源来增加对于与血友病a相关的肝脏组织的特异性。实际上,通过该方法产生用于治疗患有血友病a的患者中的肝脏组织的三倍体病毒对相关组织的特异性高于仅由aav3、aav6或aav7构成的病毒载体。
实施例6:本发明的aav治疗疾病的用途
帕金森氏病的治疗.患有帕金森氏病的45岁的男性患者用aav治疗,所述aav由细胞系、诸如具有atccno.pta13274的分离的hek293细胞系生成(参见,例如,美国专利号9,441,206),所述aav含有具有来自aav2的rep和cap基因的第一辅助质粒和具有来自aav2的rep基因和来自aav4的cap基因的第二辅助质粒以及编码谷氨酸脱羧酶65(gad65)和/或谷氨酸脱羧酶67(gad67)的核苷酸序列的第三质粒,所述核苷酸序列插入两个itr之间。由三种质粒生成的单倍体aav含有用于治疗帕金森氏病的gad65和/或gad67蛋白的核苷酸序列。将aav施用于患者,其在施用后不久显示震颤的频率降低和患者的平衡的改善。随着时间,患者还看到患者在施用aav前患有的幻觉和幻想的数量和严重程度的降低。
巴滕病的治疗.患有巴滕病的8岁的男性患者用aav治疗,所述aav由细胞系、诸如具有atccno.pta13274的分离的hek293细胞系生成(参见,例如,美国专利号9,441,206),所述aav含有具有来自aav3的rep和cap基因的第一辅助质粒和具有来自aav3的rep基因和来自aav5的cap基因的第二辅助质粒。第三质粒编码用于治疗巴滕氏病的cln2的核苷酸序列,其中cln2基因已插入两个itr之间。由三种质粒生成的单倍体aav含有用于治疗巴滕氏病的核苷酸序列。将aav施用于患者,其在施用后不久显示精神敏锐度的增加。此外,患者还看到患者在施用aav前患有的癫痫的减少以及体征和运动技能的改善。
阿尔茨海默氏病的治疗.患有阿尔茨海默氏病的73岁的女性患者用aav治疗,所述aav由细胞系、诸如具有atccno.pta13274的分离的hek293细胞系生成
(参见,例如,美国专利号9,441,206),所述aav含有具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep基因和来自aav4的cap基因的第二辅助质粒;和具有来自aav3的rep基因和来自aav5的cap基因的第三辅助质粒。第四质粒编码用于治疗阿尔茨海默氏病的神经生长因子(ngf)的核苷酸序列,其中ngf已插入两个itr之间。将三倍体aav施用于患者,其在施用后不久显示精神敏锐度和短期记忆的增加。与施用aav前相比,患者还能够更好地与他人沟通,并开始更加独立地运作。
心脏病的治疗.患有心脏病的63岁的男性患者用aav治疗,所述aav由细胞系、诸如具有atccno.pta13274的分离的hek293细胞系生成(参见,例如,美国专利号9,441,206),所述aav含有以下中任一者:
(1)具有来自aav2的rep和cap基因的第一辅助质粒;具有来自aav2的rep基因和来自aav6的cap基因的第二辅助质粒;且第三质粒编码用于治疗心脏病的蛋白的核苷酸序列,其包含在第三质粒中,并已插入两个itr之间;
(2)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep基因和来自aav9的cap基因的第二辅助质粒;且第三质粒编码用于治疗心脏病的蛋白的核苷酸序列,其包含在第三质粒中,并已插入两个itr之间;
(3)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep基因和来自aav6的cap基因的第二辅助质粒;具有来自aav3的rep基因和来自aav9的cap基因的第三辅助质粒;且第四质粒含有编码用于治疗心脏病的蛋白的核苷酸序列,其包含在第三质粒中,并已插入两个itr之间;
(4)具有来自aav2的rep和来自aav2的vp1、来自aav3的vp2和来自aav9的vp3的辅助质粒;和含有插入两个itr之间的编码用于治疗心脏病的蛋白的核苷酸序列的第二质粒;
(5)具有来自aav3的rep和来自aav3的vp1、来自aav6的vp2和来自aav6的vp3的辅助质粒;且第二质粒含有插入两个itr之间的编码用于治疗心脏病的蛋白的核苷酸序列;或
(6)具有来自aav3的rep和来自aav3的vp1、来自aav6的vp2和来自aav9的vp3的辅助质粒;且第二质粒含有插入两个itr之间的编码用于治疗心脏病的蛋白的核苷酸序列,其中
将多倍体aav施用于患者,其在施用后不久显示与心脏病相关的症状的减少,并显示患者的心脏健康的相称改善。
囊性纤维化的治疗.患有囊性纤维化的19岁女性用aav治疗,所述aav由细胞系、诸如具有atccno.pta13274的分离的hek293细胞系生成(参见,例如,美国专利号9,441,206),所述aav含有以下中任一者:
(1)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep和来自aav10的cap基因的第二辅助质粒;和编码插入两个itr之间的cftr的核苷酸序列的第三质粒;
(2)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep基因和来自aav9的cap基因的第二辅助质粒;具有来自aav3的rep基因和来自aav10的cap基因的第三辅助质粒;和编码已插入两个itr之间的cftr的核苷酸序列的第四质粒;
(3)具有来自aav2的rep和来自aav2的vp1、来自aav9的vp2和来自aav9的vp3的辅助质粒;和编码插入两个itr之间的cftr的核苷酸序列的第二质粒;
(4)具有来自aav3的rep和来自aav2的vp1、来自aav10的vp2和来自aav10的vp3的辅助质粒;和编码插入两个itr之间的cftr的核苷酸序列的第二质粒;或,
(7)具有来自aav2的rep和来自aav2的vp1、来自aav9的vp2和来自aav10的vp3的辅助质粒;且第二质粒编码插入两个itr之间的cftr的核苷酸序列,其中
将aav施用于患者,其在施用后不久显示对患者肺部的损伤增加的减慢;肺功能丧失增加的降低和肝受损的速度的降低,和肝硬化严重程度增加的减缓。同一患者还看到该患者已开始患有的囊性纤维化相关的糖尿病的严重程度的降低。
骨骼肌疾病-肌萎缩性侧索硬化症(als)的治疗.患有肌萎缩性侧索硬化症(als)的33岁的男性用aav治疗,所述aav由细胞系、诸如具有atccno.pta13274的分离的hek293细胞系生成(参见,例如,美国专利号9,441,206),所述aav含有以下中任一者:
(1)具有来自aav2的rep和cap基因的第一辅助质粒;具有来自aav2的rep和来自aav8的cap基因的第二辅助质粒;和编码插入两个itr之间的超氧化物歧化酶1(sod1)的核苷酸序列的第三质粒;
(2)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep和来自aav9的cap基因的第二辅助质粒;和编码插入两个itr之间的sod1的核苷酸序列的第三质粒;
(3)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep基因和来自aav8的cap基因的第二辅助质粒;具有来自aav3的rep基因和来自aav9的cap基因的第三辅助质粒;和编码插入两个itr之间的sod1的核苷酸序列的第四质粒;
(4)具有来自aav3的rep和来自aav3的vp1、来自aav9的vp2和来自aav9的vp3的辅助质粒;和编码插入两个itr之间的sod1的核苷酸序列的第二质粒;
(5)具有来自aav3的rep和来自aav3的vp1、来自aav8的vp2和来自aav8的vp3的辅助质粒;和编码插入两个itr之间的sod1的核苷酸序列的第二质粒;或,
(6)具有来自aav3的rep和来自aav3的vp1、来自aav8的vp2和来自aav的vp3的辅助质粒;和编码插入两个itr之间的sod1的核苷酸序列的第二质粒,其中
将aav施用于患者,其在施用后不久显示与als相关的症状的减轻,包括对大脑和脊髓中的运动神经元的损害进展的减缓或停止以及患者的脑和肌肉之间的通讯的维持。
杜兴氏肌营养不良症的治疗.患有杜兴氏肌营养不良症(dmd)的5岁的男性用aav治疗,所述aav由细胞系、诸如具有atccno.pta13274的分离的hek293细胞系生成,所述aav含有以下中任一者:
(1)具有来自aav2的rep和cap基因的第一辅助质粒;具有来自aav2的rep和来自aav8的cap基因的第二辅助质粒;和编码插入两个itr之间的肌营养不良蛋白的核苷酸序列的第三质粒;
(2)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep和来自aav9的cap基因的第二辅助质粒;和编码插入两个itr之间的肌营养不良蛋白的核苷酸序列的第三质粒;
(3)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep基因和来自aav8的cap基因的第二辅助质粒;具有来自aav3的rep基因和来自aav9的cap基因的第三辅助质粒;和编码插入两个itr之间的肌营养不良蛋白的核苷酸序列的第四质粒;
(4)具有来自aav3的rep和来自aav3的vp1、来自aav9的vp2和来自aav9的vp3的辅助质粒;和编码插入两个itr之间的肌营养不良蛋白的核苷酸序列的第二质粒;
(5)具有来自aav3的rep和来自aav3的vp1、来自aav8的vp2和来自aav8的vp3的辅助质粒;和编码插入两个itr之间的肌营养不良蛋白的核苷酸序列的第二质粒;或,
(6)具有来自aav3的rep和来自aav3的vp1、来自aav8的vp2和来自aav的vp3的辅助质粒;和编码插入两个itr之间的肌营养不良蛋白的核苷酸序列的第二质粒,其中
将aav施用于患者,其在施用后不久显示患者的骨骼肌的损害和消耗增加的减慢,以及由于杜兴氏肌营养不良症导致的心脏和肺部遭受的损害的减慢或停止。
重症肌无力的治疗.患有重症肌无力(mg)的33岁的女性用aav治疗,所述aav由细胞系、诸如具有atccno.pta13274的分离的hek293细胞系生成,所述aav含有以下中任一者:
(1)具有来自aav2的rep和cap基因的第一辅助质粒;具有来自aav2的rep和来自aav8的cap基因的第二辅助质粒;和编码插入两个itr之间的使得患者将不再患有mg的基因的核苷酸序列的第三质粒;
(2)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep和来自aav9的cap基因的第二辅助质粒;和编码插入两个itr之间的使得患者将不再患有mg的基因的第三质粒;
(3)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep基因和来自aav8的cap基因的第二辅助质粒;具有来自aav3的rep基因和来自aav9的cap基因的第三辅助质粒;和编码插入两个itr之间的使得患者将不再患有mg的基因的第四质粒;
(4)具有来自aav3的rep和来自aav3的vp1、来自aav9的vp2和来自aav9的vp3的辅助质粒;和编码插入两个itr之间的使得患者将不再患有mg的基因的第二质粒;
(5)具有来自aav3的rep和来自aav3的vp1、来自aav8的vp2和来自aav8的vp3的辅助质粒;且第二质粒编码插入两个itr之间的使得患者将不再患有mg的基因;或,
(6)具有来自aav3的rep和来自aav3的vp1、来自aav8的vp2和来自aav的vp3的辅助质粒;且第二质粒编码插入两个itr之间的使得患者将不再患有mg的基因,其中
将aav施用于患者,其在施用后不久显示患者身体的肌肉和神经之间的通讯的增加崩溃的减慢,导致肌肉控制丧失的严重程度的减慢或停止。在施用aav后,患者的活动力稳定并且不再恶化,并且在施用aav后,患者的呼吸也没有恶化。
肢带肌营养不良症的治疗.患有肢带肌营养不良症(lgmd)的13岁的男性用aav治疗,所述aav由细胞系、诸如具有atccno.pta13274的分离的hek293细胞系生成,所述aav含有以下中任一者:
(1)具有来自aav2的rep和cap基因的第一辅助质粒;具有来自aav2的rep和来自aav8的cap基因的第二辅助质粒;和编码插入两个itr之间的具有与lgmd相关的突变的十五种基因之一(包括但不限于肌节蛋白、视松蛋白、需钙蛋白酶-3、α-肌聚糖和β-肌聚糖)的核苷酸序列的第三质粒;
(2)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep和来自aav9的cap基因的第二辅助质粒;和编码插入两个itr之间的具有与lgmd相关的突变的十五种基因之一(包括但不限于肌节蛋白、视松蛋白、需钙蛋白酶-3、α-肌聚糖和β-肌聚糖)的核苷酸序列的第三质粒;
(3)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep基因和来自aav8的cap基因的第二辅助质粒;具有来自aav3的rep基因和来自aav9的cap基因的第三辅助质粒;和编码插入两个itr之间的具有与lgmd相关的突变的十五种基因之一(包括但不限于肌节蛋白、视松蛋白、需钙蛋白酶-3、α-肌聚糖和β-肌聚糖)的核苷酸序列的第四质粒;
(4)具有来自aav3的rep和来自aav3的vp1、来自aav9的vp2和来自aav9的vp3的辅助质粒;和编码插入两个itr之间的具有与lgmd相关的突变的十五种基因之一(包括但不限于肌节蛋白、视松蛋白、需钙蛋白酶-3、α-肌聚糖和β-肌聚糖)的核苷酸序列的第二质粒;
(5)具有来自aav3的rep和来自aav3的vp1、来自aav8的vp2和来自aav8的vp3的辅助质粒;且第二质粒编码插入两个itr之间的具有与lgmd相关的突变的十五种基因之一(包括但不限于肌节蛋白、视松蛋白、需钙蛋白酶-3、α-肌聚糖和β-肌聚糖)的核苷酸序列;或
(6)具有来自aav3的rep和来自aav3的vp1、来自aav8的vp2和来自aav的vp3的辅助质粒;且第二质粒编码插入两个itr之间的具有与lgmd相关的突变的十五种基因之一(包括但不限于肌节蛋白、视松蛋白、需钙蛋白酶-3、α-肌聚糖和β-肌聚糖)的核苷酸序列,其中
将aav中的一种或多种(其各自编码与lgmd相关的15种不同基因之一)施用于患者,其在施用后不久显示额外的肌肉消瘦和萎缩的减慢或停止。
肝脏疾病-血友病b的治疗.患有由于因子ix(fix)的缺乏导致的血友病b的9岁的男性用aav治疗,所述aav由细胞系、诸如具有atccno.pta13274的分离的hek293细胞系生成,所述aav含有以下中任一者:
(1)具有来自aav2的rep和cap基因的第一辅助质粒;具有来自aav2的rep和来自aav6的cap基因的第二辅助质粒;和编码插入两个itr之间的用于治疗血友病b的fix的核苷酸序列的第三质粒;
(2)具有来自aav2的rep和cap基因的第一辅助质粒;具有来自aav3的rep和来自aav7的cap基因的第二辅助质粒;和编码插入两个itr之间的用于治疗血友病b的fix的核苷酸序列的第三质粒;
(3)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep基因和来自aav6的cap基因的第二辅助质粒;具有来自aav3的rep基因和来自aav7的cap基因的第三辅助质粒;和编码插入两个itr之间的fix的核苷酸序列的第四质粒;
(4)具有来自aav2的rep和来自aav2的vp1、来自aav6的vp2和来自aav6的vp3的辅助质粒;和编码插入两个itr之间的fix的核苷酸序列的第二质粒;
(5)具有来自aav2的rep和来自aav3的vp1、来自aav7的vp2和来自aav7的vp3的辅助质粒;和编码插入两个itr之间的fix的核苷酸序列的第二质粒;或
(6)具有来自aav2的rep和来自aav3的vp1、来自aav6的vp2和来自aav7’的vp3的辅助质粒;和编码插入两个itr之间的fix的核苷酸序列的第二质粒,其中
将aav施用于患者,其在施用后不久显示血友病b的严重程度的降低,包括出血发作的降低。
血友病a的治疗.患有由于因子viii(fviii)的缺乏导致的血友病a的8岁的男性用aav治疗,所述aav由细胞系、诸如具有atccno.pta13274的分离的hek293细胞系生成,所述aav含有以下中任一者:
(1)具有来自aav2的rep和cap基因的第一辅助质粒;具有来自aav2的rep和来自aav6的cap基因的第二辅助质粒;和编码插入两个itr之间的fviii的核苷酸序列的第三质粒;
(2)具有来自aav2的rep和cap基因的第一辅助质粒;具有来自aav3的rep和来自aav7的cap基因的第二辅助质粒;和编码插入两个itr之间的fviii的核苷酸序列的第三质粒;
(3)具有来自aav3的rep和cap基因的第一辅助质粒;具有来自aav3的rep基因和来自aav6的cap基因的第二辅助质粒;具有来自aav3的rep基因和来自aav7的cap基因的第三辅助质粒;和编码插入两个itr之间的fviii的核苷酸序列的第四质粒;
(4)具有来自aav2的rep和来自aav2的vp1、来自aav6的vp2和来自aav6的vp3的辅助质粒;和编码插入两个itr之间的fviii的核苷酸序列的第二质粒;
(5)具有来自aav2的rep和来自aav3的vp1、来自aav7的vp2和来自aav7的vp3的辅助质粒;和编码插入两个itr之间的fviii的核苷酸序列的第二质粒;或
(6)具有来自aav2的rep和来自aav3的vp1、来自aav6的vp2和来自aav7’的vp3的辅助质粒;和编码插入两个itr之间的fviii的核苷酸序列的第二质粒,其中
将aav施用于患者,其在施用后不久显示血友病a的严重程度的降低,包括出血发作的降低。
实施例7.由两种不同血清型产生单倍体衣壳和起始密码子的突变。
在本实施例中,多倍体aav病毒粒子由两种不同血清型的衣壳组装。将来自仅第一aav血清型的vp1、vp2和vp3的核苷酸序列连接至辅助质粒中,且将来自仅第二aav血清型的vp1、vp2和vp3的核苷酸序列连接至相同或不同的辅助质粒中,使得辅助质粒包括来自两种不同血清型的vp1、vp2和vp3衣壳蛋白的核酸序列。在将编码vp1、vp2和vp3衣壳蛋白的第一和第二血清型核苷酸序列连接至辅助质粒中之前或之后,改变衣壳核苷酸序列以提供来自仅第一血清型的vp1和来自仅第二血清型的vp2和vp3。在本实施例中,已通过突变vp2和vp3衣壳蛋白的起始密码子来改变第一血清型的vp1核苷酸序列,如图7中所示。在本实施例中,突变vp2的acg起始位点和vp3的三个atg起始位点,使得这些密码子不能起始从来自第一血清型的vp2和vp3衣壳蛋白的核苷酸序列转录的rna的翻译。类似地,如图8中所示,在编码第二血清型的衣壳蛋白的核苷酸序列中突变vp1的atg起始位点,使得该密码子无法起始编码vp1的rna的翻译,但可以对于vp2和vp3两者起始翻译。因此,在本实施例中,产生多倍体aav病毒粒子,其包括来自仅第一血清型的vp1,但不是vp2或vp3,和来自仅第二血清型的vp2和vp3,但不是vp1。
在应用这种通过起始密码子的突变产生多倍体aav病毒粒子的技术中,突变aav2的vp2和vp3的起始密码子(如图19中突出显示所示),使得仅vp1从由图19中所示的质粒转录的rna翻译。在该技术的进一步应用中,突变aav2vp1的起始密码子(如图18中突出显示所示),使得vp2和vp3而非vp1从由图19中所示的质粒转录的rna翻译。因此,起始密码子的突变提供敲除vp1、vp2和vp3中的一种或多种的表达的方法。
实施例8.由两种不同血清型产生单倍体衣壳和起始密码子的突变。
在本实施例中,多倍体aav病毒粒子由两种不同血清型的衣壳组装。将来自仅第一aav血清型的vp1、vp2和vp3的核苷酸序列连接至辅助质粒中,且将来自仅第二aav血清型的vp1、vp2和vp3连接至相同或不同的辅助质粒中,使得辅助质粒包括来自两种不同血清型的vp1、vp2和vp3衣壳蛋白。在将编码vp1、vp2和vp3衣壳蛋白的第一和第二血清型核苷酸序列连接至辅助质粒中之前或之后,改变衣壳核苷酸序列以提供来自仅第一血清型的vp1和vp3和来自仅第二血清型的vp2。在本实施例中,突变vp2的acg起始位点,使得该密码子不能起始从来自第一血清型的vp2衣壳蛋白的核苷酸序列转录的rna的翻译。类似地,在编码第二血清型的衣壳蛋白的核苷酸序列中突变vp1和vp3的atg起始位点,使得这些密码子无法起始编码vp1和vp3的rna的翻译,但可以对于vp2起始翻译。因此,在本实施例中,产生多倍体aav病毒粒子,其包括来自仅第一血清型的vp1和vp3,但不是vp2,和来自仅第二血清型的vp2,但不是vp1和vp3。
在应用这种通过起始密码子的突变产生多倍体aav病毒粒子的技术中,突变aav2的vp2的起始密码子(如图20中突出显示所示),使得vp1和vp3从由图20中所示的质粒转录的rna翻译。因此,起始密码子的突变提供敲除vp1、vp2和vp3中的一种或多种的表达的方法。
实施例9.由两种不同血清型产生单倍体衣壳和剪接受体位点的突变。
在本实施例中,多倍体aav病毒粒子由两种不同血清型的衣壳组装。将来自仅第一aav血清型的vp1、vp2和vp3的核苷酸序列连接至辅助质粒中,且将来自仅第二aav血清型的vp1、vp2和vp3连接至相同或不同的辅助质粒中,使得辅助质粒包括来自两种不同血清型的vp1、vp2和vp3衣壳蛋白。在将编码vp1、vp2和vp3衣壳蛋白的第一和第二血清型核苷酸序列连接至辅助质粒中之前或之后,改变衣壳核苷酸序列以提供来自仅第一血清型的vp1和来自仅第二血清型的vp2和vp3。在本实施例中,已通过突变a2剪接受体位点改变第一血清型的核苷酸序列,如图9中所示。在本实施例中,通过突变a2剪接受体位点,不产生来自第一血清型的vp2和vp3衣壳蛋白。类似地,如图10中所示,通过突变a1剪接受体位点,不产生来自第二血清型的vp1衣壳蛋白,而产生vp2和vp3衣壳蛋白。因此,在本实施例中,产生多倍体aav病毒粒子,其包括来自仅第一血清型的vp1,但不是vp2或vp3,和来自仅第二血清型的vp2和vp3,但不是vp1。
实施例10.由两种不同血清型产生单倍体衣壳和起始密码子和剪接受体位点的突变。
在本实施例中,多倍体aav病毒粒子由两种不同血清型的衣壳组装。将来自仅第一aav血清型的vp1、vp2和vp3的核苷酸序列连接至辅助质粒中,且将来自仅第二aav血清型的vp1、vp2和vp3连接至相同或不同的质粒中,使得辅助质粒包括来自两种不同血清型的vp1、vp2和vp3衣壳蛋白。在将编码vp1、vp2和vp3衣壳蛋白的第一和第二血清型核苷酸序列连接至辅助质粒中之前或之后,改变衣壳核苷酸序列以提供来自仅第一血清型的vp1和来自仅第二血清型的vp2和vp3。在本实施例中,已通过突变vp2和vp3衣壳蛋白的起始密码子和突变a2剪接受体位点来改变第一血清型的核苷酸序列,如图11中所示。在本实施例中,突变vp2的acg起始位点和vp3的三个atg起始位点连同a2剪接受体位点。作为结果,仅产生第一血清型的vp1衣壳蛋白。均不产生来自第一血清型的vp2和vp3衣壳蛋白。类似地,如图12中所示,突变vp1的atg起始位点连同a1剪接受体位点。作为结果,仅产生第二血清型的vp2和vp3衣壳蛋白。不产生第二血清型的vp1衣壳蛋白。因此,在本实施例中,产生多倍体aav病毒粒子,其包括来自仅第一血清型的vp1,但不是vp2或vp3,和来自仅第二血清型的vp2和vp3,但不是vp1。
实施例11.使用两种质粒由两种不同血清型产生单倍体衣壳。
在本实施例中,使用两种质粒产生包含来自aav5的vp1和来自aav9的vp2/vp3的单倍体aav病毒粒子。如图13中所示,产生辅助质粒,其包括质粒骨架连同adearlygenes和rep(例如,来自aav2)。该辅助质粒已向其中连接编码来自仅aav5的衣壳蛋白的核苷酸序列和编码仅aav9的衣壳蛋白的分开核苷酸序列。关于编码aav5的衣壳蛋白的核苷酸序列,该核苷酸序列已突变vp2/vp3的起始密码子以防止翻译和/或已突变a2剪接受体位点以防止剪接。关于编码aav9的衣壳蛋白的核苷酸序列,该核苷酸序列已突变vp1的起始密码子以防止翻译和/或已突变vp1剪接受体位点以防止剪接。将辅助质粒连同编码具有两个itr的转基因的质粒转染至具有atccno.pta13274的hek293细胞系中(参见例如,美国专利号9,441,206)。从上清液纯化病毒并表征。如图13中所示,病毒衣壳包括aa9的vp2/vp3(以浅灰色显示)和aav5的vp1(以深灰色显示),如图13的底部所示的病毒粒子中所见。
实施例12.使用三种质粒由两种不同血清型产生单倍体衣壳。
在本实施例中,使用三种质粒产生包含来自aav5的vp1和来自aav9的vp2/vp3的单倍体aav病毒粒子。如图14中所示,产生第一辅助质粒,其包括adearlygenes。产生第二辅助质粒,其包括质粒骨架连同rep(例如,aav2)。该第二辅助质粒已向其中连接编码来自仅aav5的衣壳蛋白的核苷酸序列和编码仅aav9的衣壳蛋白的分开核苷酸序列。关于编码aav5的衣壳蛋白的核苷酸序列,该核苷酸序列已突变vp2/vp3的起始密码子以防止翻译和/或已突变a2剪接受体位点以防止剪接。关于编码aav9的衣壳蛋白的核苷酸序列,该核苷酸序列已突变vp1的起始密码子以防止翻译和/或已突变vp1剪接受体位点以防止剪接。将辅助质粒连同编码具有两个itr的转基因的质粒转染至具有atccno.pta13274的hek293细胞系中(参见例如,美国专利号9,441,206)。从上清液纯化病毒并表征。如图14中所示,病毒衣壳包括aav9的vp2/vp3(以浅灰色显示)和aav5的vp1(以深灰色显示),如图13的底部所示的病毒粒子中所见。
实施例13.使用四种质粒由两种不同血清型产生单倍体衣壳。
在本实施例中,使用四种质粒产生包含来自aav5的vp1和来自aav9的vp2/vp3的单倍体aav病毒粒子。如图15中所示,产生第一辅助质粒,其包括adearlygenes。产生第二辅助质粒,其包括质粒骨架连同rep(例如,aav2)。该第二辅助质粒已向其中连接编码来自仅aav5的衣壳蛋白的核苷酸序列。产生第三辅助质粒,其包括质粒骨架连同rep。该第三辅助质粒已向其中连接编码来自仅aav9的衣壳蛋白的核苷酸序列。第四质粒包括转基因和两个itr。关于编码aav5的衣壳蛋白的核苷酸序列,该核苷酸序列已突变vp2/vp3的起始密码子以防止翻译和/或已突变a2剪接受体位点以防止剪接。关于编码aav9的衣壳蛋白的核苷酸序列,该核苷酸序列已突变vp1的起始密码子以防止翻译和/或已突变vp1剪接受体位点以防止剪接。将辅助质粒连同编码具有两个itr的转基因的质粒转染至具有atccno.pta13274的hek293细胞系中(参见例如,美国专利号9,441,206)。从上清液纯化病毒并表征。如图14中所示,病毒衣壳包括aa9的vp2/vp3(以浅灰色显示)和aav5的vp1(以深灰色显示),如图13的底部所示的病毒粒子中所见。
实施例14.由三种不同血清型产生单倍体衣壳和起始密码子的突变。
在本实施例中,多倍体aav病毒粒子由三种不同血清型的衣壳组装。构建辅助质粒,使得来自仅第一aav血清型的vp1、vp2和vp3、来自仅第二aav血清型的vp1、vp2和vp3和来自仅第三aav血清型的vp1、vp2和vp3的核苷酸序列连接至辅助质粒中,使得辅助质粒包括来自三种不同血清型的vp1、vp2和vp3衣壳蛋白的核酸序列。在将编码来自三种不同血清型中每一种的vp1、vp2和vp3衣壳蛋白的核苷酸序列连接至辅助质粒中之前或之后,改变衣壳核苷酸序列以提供来自仅第一血清型的vp1、来自仅第二血清型的vp2和来自仅第三血清型的vp3。在本实施例中,已通过突变vp2和vp3衣壳蛋白的起始密码子来改变第一血清型的vp1核苷酸序列。在本实施例中,突变vp2的acg起始密码子和vp3的三个atg起始密码子,使得这些密码子不能起始从来自第一血清型的vp2和vp3衣壳蛋白的核苷酸序列转录的rna的翻译。类似地,已通过突变vp1和vp3衣壳蛋白的起始密码子来改变第二血清型的vp1和vp3核苷酸序列。在本实施例中,突变vp1的atg起始位点和vp3的三个atg起始密码子,使得这些密码子不能起始从vp1和vp3衣壳蛋白的核苷酸序列转录的rna的翻译。进一步,已通过突变vp1和vp2衣壳蛋白的起始密码子来改变第三血清型的vp1和vp2核苷酸序列。在本实施例中,突变vp1的atg起始密码子和vp2的acg起始密码子,使得这些密码子不能起始从vp1和vp2衣壳蛋白的核苷酸序列转录的rna的翻译。因此,在本实施例中,产生多倍体aav病毒粒子,其包括来自仅第一血清型的vp1,但不是vp2,也不是vp3;来自仅第二血清型的vp2,但不是vp1,也不是vp2;和来自仅第三血清型的vp3,但不是vp1,也不是vp2。
实施例15.使用dna改组由两种不同血清型产生单倍体衣壳。
在该实验中,多倍体aav病毒粒子由来自仅一种aav血清型的aav衣壳蛋白和由三种不同aav血清型的dna改组产生的核酸产生。在本实施例中,对aav1、aav2和aav8的核苷酸衣壳蛋白序列进行用一种或多种限制酶和/或dna酶处理,并且将dna切割成长度为50-100bp的dna片段。然后对dna片段的混合物在没有引物的情况下进行聚合酶链式反应(pcr)。pcr重复多次或直到通过pcr产生的dna分子达到编码衣壳基因的核酸的大小。此时,进行另一轮pcr,其中添加引物,其包括用于限制酶识别位点的序列,以允许将新产生的dna连接至辅助质粒中。在连接至辅助质粒中之前,将aav1/2/8核苷酸序列进行测序,并将可以起始从由该序列转录的rna翻译vp2和vp3衣壳蛋白的核苷酸序列内的任何起始密码子突变以防止翻译。以这种方式,aav1/2/8只能产生vp1,并且将aav1/2/8核苷酸序列连接至辅助质粒中。在该实验中,还将编码aav9的衣壳蛋白(vp1、vp2和vp3)的核苷酸序列连接至相同或不同的辅助质粒中。为了产生具有来自通过dna改组产生的aav1/2/8核苷酸序列的vp1和来自仅aav9的vp2和vp3的多倍体aav病毒粒子,将aav9的vp1的atg起始密码子突变,使得不能翻译编码vp1的rna。因此,在本实施例中,产生多倍体aav病毒粒子,其包括来自通过dna改组aav1/2/8的衣壳蛋白核苷酸序列产生的核苷酸序列的vp1,但不是vp2或vp3,和来自仅aav9的vp2和vp3,但不是vp1。
dna改组的实例阐述于图16中,其开始于来自八种aav血清型的编码vp1、vp2和vp3的核酸,并且首先通过dna酶i片段化、随后组装和组装来自八种aav的核酸的各种片段来处理核酸。生成的dna改组的核酸编码aav衣壳蛋白,其然后表达以产生衣壳的文库。然后,在动物上测试这些衣壳,以筛选显示特定组织向性和/或免疫原性降低的那些衣壳,并选择显示希望的那些用于进一步开发(图16)。
实施例16.单倍体载体h-aav829的肝脏转导。
用三种aav进行实验。在图22a中,显示aav衣壳亚基的组成。显示杂合aav,其将来自aav8的仅vp1氨基酸与编码来自aav2的vp2和vp3的那些组合(aav82)。通过将两种质粒(一种编码vp1和vp2,另一种编码vp3)共转染至hek293细胞中,产生两种单倍体aav病毒。将三种aav,aav82、28m-2vp3和h-aav82,连同aav2亲本对照经由眼眶后静脉以3x1010个颗粒的剂量注射于c57bl6小鼠中(图22b)。一周后进行成像(图22b)。基于代表5只小鼠的平均值和标准偏差的数据定量肝脏转导(图22c)。
实施例17.单倍体载体h-aav82的肌肉转导。
接下来将来自实施例23的三种aav(aav82、h-aav82和28m-vp3)以1x109个aav/luc颗粒的剂量注射至小鼠后腿肌肉中。注射后第3周,进行成像3分钟的时段,如图23a中所见。面朝上进行成像:左腿-aav82、h-aav82或28m-vp3,和右腿-aav2亲本aav。图23b提供了肌肉注射后来自4只小鼠的数据,其具有通过从aav82、h-aav82或28m-vp3至亲本aav2的通过转导计算的转导倍数增加。
实施例18.单倍体载体h-aav92的肝脏转导。
在该实验中,产生单倍体aav92,其中vp1和vp2来自仅aav9,且vp3来自仅aav3(图24a)。通过将两种质粒(一种编码aav9vp1和vp2,另一种编码aav2vp3)共转染至hek293细胞中,产生h-aav92。将h-aav92和亲本aav2经由眼眶后静脉以3x1010个颗粒的剂量注射至c57bl6小鼠中(图24b)。一周后进行成像(图24b)。基于代表5只小鼠的平均值和标准偏差的数据定量肝脏转导(图24c)。
实施例19.单倍体载体h-aav82g9的肝脏转导。
在该实验中,产生单倍体aav82g9,其中vp1和vp2来自仅aav8,且vp3来自仅aav2g9(图25a)。通过将两种质粒(一种编码aav8vp1和vp2,另一种编码aav2g9vp3)共转染至hek293细胞中,产生h-aav82g9。将h-aav82g9和aav2g9经由眼眶后静脉以3x1010个颗粒的剂量注射至c57bl6小鼠中(图25b)。一周后进行成像(图25b)。基于代表5只小鼠的平均值和标准偏差的数据定量肝脏转导(图25c)。
最后,应当理解,尽管通过参考具体实施方案突出本说明书的各方面,但本领域技术人员将容易理解,这些公开的实施方案仅举例说明本文公开的主题的原理。因此,应当理解,公开的主题绝不限于本文所述的具体方法、方案和/或试剂等。因此,在不脱离本说明书的精神的情况下,可以根据本文的教导作出公开的主题的各种修改或改变或替代配置。最后,本文使用的术语仅用于描述具体实施方案的目的,且不旨在限制本发明的范围,其仅通过权利要求限定。因此,本发明不限于精确地如所显示和描述的那样。
本文描述本发明的某些实施方案,包括本发明人已知用于用于实施本发明的最佳模式。当然,在阅读前面描述后,对这些描述的实施方案的变型对于本领域普通技术人员将变得显而易见。本发明人期望技术人员适当地采用此类变型,且本发明人意欲以不同于本文具体描述的方式来实施本发明。相应地,如适用法律所允许,本发明包括其随附权利要求中所述的主题的所有修改和等效方案。此外,本发明涵盖其所有可能的变型中的上述实施方案的任何组合,除非本文另有指明或另外与上下文明显矛盾。
本发明的替代实施方案、要素或步骤的分组不应被解释为限制。每个组成员可以单独或与本文中公开的其他组成员进行任何组合来提及和请求保护。预期出于方便和/或可专利性的原因,组中的一个或多个成员可以包括在组中或从其中删除。当发生任何这种包括或删除时,说明书被认为含有如修改的组,因此满足所附权利要求中使用的所有马库什基团的书面描述。
除非另有说明,否则表示本说明书和权利要求中使用的特征、项目、数量、参数、特性、术语等的所有数字在所有情况下均应理解为通过术语“约”修饰。如本文所用,术语“约”意味着如此限定的特征、项目、数量、参数、特性或项目涵盖在所述特征、项目、数量、参数、特性或项目的值之上和之下正负百分之十的范围。因此,除非相反指示,否则说明书和所附权利要求中阐述的数字参数是可以变化的近似值。至少,并且不试图将等同原则的应用限制于权利要求的范围,每个数字指示至少应根据所报告的有效数字的数量并通过应用普通的舍入技术来解释。尽管阐述本发明的广泛范围的数值范围和值是近似值,但在具体实施方案中阐述的数值范围和值被尽可能精确地报告。然而,任何数值范围或值都固有地含有某些误差,所述误差必然由它们各自的测试测量中发现的标准偏差导致。本文中值的数值范围的叙述仅意欲用作分别指代落在该范围内的每个分开数值的简写方法。除非本文另外指出,否则数值范围的每个个别值都被并入本说明书中,如同其在本文中被个别引用一样。
本文描述的所有方法都可以以任何合适的顺序进行,除非本文另有指明或另外与上下文明显矛盾。本文提供的任何和所有实施例或示例性语言(例如,“诸如”)的使用仅仅意欲更好地说明本发明,并且不构成对本发明的范围的限制,除非另有要求。本说明书中的任何语言都不应理解为指示对实施本发明必不可少的任何未请求保护的要素。
本文公开的具体实施方案可以使用由语言组成或基本上由语言组成而在权利要求中进一步限制。当在权利要求中使用时,无论是提交还是根据修改增加,过渡术语“由…组成”不包括权利要求中未指定的任何要素、步骤或成分。过渡术语“基本上由…组成”将权利要求的范围限制为指定的材料或步骤以及不实质性影响基本和新型特征的那些。如此请求保护的本发明的实施方案在本文中被固有地或清楚地描述和实施。
本说明书中引用和标识的所有专利、专利出版物和其他出版物均通过引用以其整体个别地和明确地并入本文,用于描述和公开例如在此类出版物中描述的可能与本发明结合使用的组合物和方法的目的。仅仅提供这些出版物用于其在本申请的申请日之前的公开。在这方面,任何内容都不应被解释为承认本发明人无权凭借在先发明或任何其他原因而先于这种公开。关于这些文件的日期或内容的代表的所有陈述都基于申请人可得的信息,并且不构成关于这些文件的日期或内容的正确性的承认。
实施例20.嵌合衣壳蛋白和aav单倍体病毒载体转导。
如上所解释,用衣壳orf的起始密码子中的突变体制备一系列aav辅助质粒的构建体,其中仅表达一种或两种病毒vp蛋白。还制备嵌合aav辅助构建体,其中vp1/2蛋白由两种不同的血清型(aav2和aav8)驱动。这些构建体用于产生一堆单倍体病毒载体,并评估它们在小鼠中的转导效力。当与仅aav2和仅aav3载体相比时,发现从具有来自血清型7、8、9和rh10的vp1/vp2和来自aav2或aav3的vp3的单倍体载体获得增强的转导。进一步显示,由具有来自aav2的n-末端和来自aav8的c-末端的嵌合vp1/vp2衣壳以及来自aav2的vp3制成的aav载体诱导高得多的转导。本文提供的数据显示增强aav转导用于进一步应用aav载体的简单且有效的方法。
具有来自其他血清型的vp1/vp2和来自aav2的vp3的单倍体载体增强aav肝脏转导。
通过以1:1的比率共转染质粒表达的aav8vp1/2和aav2vp3产生单倍体病毒。结果显示,具有来自aav8的vp1/vp2和来自aav2的vp3的单倍体载体aav82增加肝脏转导(图22b和22c)。
使用aav9的vp1/vp2和aav2的vp3产生单倍体aav92载体(h-aav92)(图24a)。在全身施用后,在第1周进行成像。用h-aav92实现的肝脏转导率是aav2的约4倍(图24b和24c)。该数据表明来自其他血清型的vp1/vp2能够增加aav2转导。
来自具有来自aav2突变体的vp3的单倍体载体的增强的aav肝脏转导。
aav9载体使用聚糖作为其有效转导的主要受体。在先前的研究中,将aav9聚糖受体结合位点移植至aav2衣壳中以制成aav2g9载体,并且发现aav2g9具有比aav2更高的肝脏向性。本文描述了单倍体载体(h-aav82g9),其中vp1/vp2来自aav8且vp3来自aav2g9(图25a)。在全身性注射入小鼠后,与aav2g9相比,在h-aav82g9应用后的第1周和第2周都观察到超过10倍的肝脏转导(图25b和25c)。该数据表明来自其他血清型的vp1/vp2整合至aav2突变体vp3中能够增加肝脏转导。
来自具有来自aav3的vp3的单倍体载体的增强的aav肝脏转导。
制成单倍体载体(其中vp3来自其他血清型且vp1/vp2来自不同血清型或变体,其中起始密码子被突变)和vp蛋白构建体以表达仅aav3vp3或仅aavrh10vp1/vp2。产生不同的单倍体h-aav83(来自aav8的vp1/vp2和来自aav3的vp3)、h-aav93(来自aav9的vp1/vp2和来自aav3的vp3)和h-aavrh10-3(来自aavrh10的vp1/vp2和来自aav3的vp3)载体(图26a),并经由全身施用注射至小鼠中。在第1周实施成像。如图26b和26c中所示,用单倍体载体(h-aav83、h-aav93和h-aavrh10-3)实现比aav3更高的肝脏转导。这与从其他单倍体载体获得的结果一致。此外,这些单倍体载体还增强来自其他组织的转导,如图26b和26d中所示。有趣的是,基于成像特征,这些单倍体载体还诱导全身性转导,这不同于来自具有来自aav2的vp3的单倍体载体的结果,其仅有效地转导肝脏(图22和24)。总之,具有来自一种血清型的vp1/vp2和来自替代血清型的vp3的单倍体载体能够增强转导并可能改变其向性。
具有来自aav8的vp1/vp2和来自aav2的vp3的c-末端的单倍体载体增强aav转导。
生成一系列构建体,其表达仅aav8vp1/vp2、仅aav2vp3、具有来自aav2的n-末端和来自aav8的c-末端的嵌合vp1/vp2(28m-2vp3)或具有来自aav8的n-末端和来自aav2的c-末端且没有vp3起始密码子的突变的嵌合aav8/2(图27a)。这些质粒用于以1:1的质粒比率产生具有不同组合的单倍体aav载体(图27b)。在经由眼眶后静脉在小鼠中注射1x1010个这些单倍体载体的颗粒后,评估肝转导效率(图27c)。嵌合aav82载体(aav82)诱导比aav2略高的肝脏转导。然而,单倍体aav82(h-aav82)具有比aav2高得多的肝脏转导。观察到单倍体载体28m-2vp3的肝脏转导进一步增加。将这些单倍体载体施用至小鼠的肌肉中。为了便于比较,当小鼠面朝上时,向右腿注射aav2载体,并向左腿注射单倍体载体。aav注射后第3周,拍摄图像。与在肝脏中的观察一致,所有单倍体载体和嵌合载体均具有较高的肌肉转导,其中来自单倍体载体28m-2vp3的最佳(图27d)。该结果表明,具有来自aav2的n-末端和来自aav8的c-末端的嵌合vp1/vp2归因于单倍体aav82载体的高肝脏转导。
从嵌合单倍体载体向核的病毒粒子运输增加。
aav转导涉及许多步骤。结合后,经由内吞作用将aav病毒粒子吸收至内体中。从内体逃逸后,aav病毒粒子到达核用于转基因表达。确定哪些步骤导致来自单倍体载体的高转导。首先,进行aav载体结合测定,并且发现与其他载体相比,更少的28m-2vp3病毒粒子与huh7细胞结合(图28)。接下来,在核中检测到aav基因组拷贝数,并且在不同的aav载体之间未发现差异。有趣的是注意到,当将aav基因组拷贝数与结合的病毒粒子进行比较时,在核中观察到更多的aav病毒粒子(图28)。这些结果表明,aav载体28m-2vp3对于运输更有效。
单倍体aav载体的高转导不是由病毒粒子稳定性导致。
通过加热病毒病毒粒子进行以下实验。将病毒在不同温度下加热半小时,且然后使用识别完整或不完整病毒粒子的一抗a20adk8或b1进行western印迹。如图29中所示,当将病毒在70℃下加热时,所有病毒病毒粒子都崩解。aav单倍体载体之间的针对热的稳定性没有差异,而无论不同温度如何,除了aav82载体。该数据表明增强的转导可能与单倍体病毒粒子稳定性无关。
酸性条件对单倍体载体的vp1n-末端暴露的影响。
已经表明,在aav载体内吞后,在酸性内体中,vp1/vp2n-末端暴露于病毒粒子表面上。vp1/vp2末端含有aav载体的磷脂酶a2和nls结构域,其帮助aav病毒从内涵体逃逸并到达核。将aav单倍体载体与pbs在不同ph值下孵育30分钟,然后应用于western印迹分析,以使用抗体a1检测vp1的n-末端。结果显示,当用不同的ph处理病毒时,没有暴露任何vp1n-末端(图30)。
本中呈现的数据显示,可以从具有来自一种aav载体衣壳的vp1/vp2和来自替代aav载体衣壳的vp3的单倍体载体获得增强的转导。
质粒和定点诱变.用于表达vp12和vp3的所有质粒均通过定点诱变制成。使用quikchangeiixl定点诱变试剂盒(agilent)根据制造商的手册进行诱变。通过重叠pcr生成含有aav2衣壳的n-末端(1201aa)和aav8衣壳的c-末端的片段。然后,将该片段克隆至pxr的swai和noti位点中。通过dna测序验证所有突变和构建体。
病毒产生.通过三质粒转染系统产生重组aav。用9µgaav转基因质粒ptr/cba-luc、12µg含有aavrep和cap基因的aav辅助质粒和15μgad辅助质粒pxx6-80转染hek293细胞的15-cm皿。转染后60小时,收集并裂解hek293细胞。将上清液进行cscl梯度超离心。通过定量pcr测定病毒滴度。
体外转导测定.将huh7和c2c12细胞通过重组病毒在平底、24孔板中以1x104vg/细胞进行转导。48小时后,收获细胞并通过荧光素酶测定系统(promega,madison,wi)评估。
动物研究.在该研究中进行的动物实验用c57bl/6小鼠和fix-/-小鼠进行。根据由unch机构动物护理和使用委员会(iacuc)批准的nih指南维持小鼠。经由眼眶后注射向六只三周龄雌性c57bl/6小鼠注射1x1010vg重组病毒。在i.p.注射d-荧光素底物(nanolightpinetop,az)后,在注射后1周,使用xenogenivislumina(caliperlifesciences,waltham,ma)对荧光素酶表达进行成像。使用livingimage(perkinelmer,waltham,ma)分析生物发光图像。对于肌肉传导,将5×109个aav/luc颗粒注射至6周龄c57bl/6雌性的腓肠肌中。在指定的时间点对小鼠成像。
检测肝脏中的aav基因组拷贝数.用蛋白酶k处理切碎的肝脏,且通过purelinkgenomicdnaminikit(invitrogen,carlsbad,ca)分离总基因组dna。通过qpcr测定法检测荧光素酶基因。小鼠核纤层蛋白基因用作内部对照。
统计分析.数据呈现为平均值±sd。studentt检验用于实施所有统计分析。<0.05的p值被认为是统计学显著性差异的。
参考文献
- 还没有人留言评论。精彩留言会获得点赞!