胺脱氢酶突变体及其在手性胺醇化合物合成中的应用
本发明涉及生物
技术领域:
中,胺脱氢酶突变体及其在手性胺醇化合物合成中的应用。
背景技术:
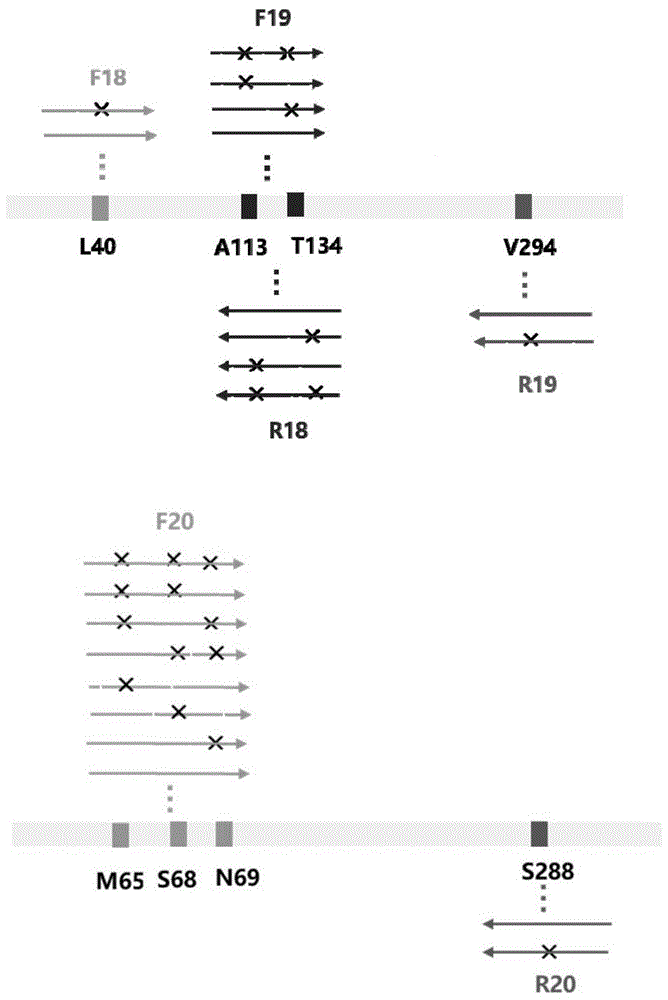
:手性胺醇是许多生物活性分子的结构单元,是重要的医药和精细化工中间体,其中(s)-2-氨基-1-丁醇主要用于合成盐酸乙胺丁醇(ethambutol),长期以来被用作抗结核杆菌一线药使用;(r)-3-氨基-1-丁醇是合成抗艾滋病整合酶抑制剂度鲁特韦(dolutegravir)的重要原料。(s)-2-氨基-1-丁醇主要有化学法和生物酶法合成,其中目前化学合成方法有:以丁醛、偶氮二甲酸二卞酯和d-脯氨酸为原料,在nabh4、h2和镍等催化作用下合成(s)-2-氨基-1-丁醇;用氢化铝锂还原2-氨基丁酸获得相应的手性胺醇产物;采用h2和镍在高压条件下还原l-2-氨基丁酸获得(s)-2-氨基-1-丁醇;采用nahb4在h2so4/thf条件下还原相应的α-氨基酸获得;利用联苯甲酰酒石酸拆分法制备,以及利用l-酒石酸拆分法制备(s)-2-氨基-1-丁醇等。生物酶法有:在氨基基团保护的情况下,由脂肪酶选择性水解羟基形成的酯键而获得手性单体;固定化青霉素g酰化酶选择性水解n-苯乙酰衍生2-氨基丁醇外消旋混合物中的(s)-构型获得(s)-2-氨基-1-丁醇;近期,weber等报道了利用酵母构建一条从苏氨酸通过四步酶法合成(s)-2-氨基-1-丁醇的新途径,但是(s)-2-氨基-1-丁醇的产量只有1.1mg/l。(r)-3-氨基-1-丁醇主要是化学法合成,收率为60%~70%。1998年,tatsuya等报道了一种合成(r)-3-氨基-1-丁醇的方法,但该方法中原料的价格昂贵,反应条件苛刻;2005年,achmatowicz等规避了昂贵的化学原料,以廉价易得的d-丙氨酸为起始原料合成(r)-3-氨基-1-丁醇,但没有报道具体产率和ee值。2011年,breuer等报道了利用扁桃酸来拆分消旋的3-氨基丁醇得到(r)-3-氨基-1-丁醇的方法。此外,2018年,khatik等通过一种具有高选择性的转氨酶,以4-羟基-2-丁酮为原料,一步合成(r)-3-氨基-1-丁醇。以上氨基丁醇的合成方法,化学法需要高温、高压和金属催化剂,反应条件苛刻,污染大,安全系数低。生物法反应条件温和,但转化率低,因此急需一种绿色高效合成手性胺醇化合物的方法。技术实现要素:本发明所要解决的技术问题是如何制备手性胺醇化合物,尤其是(s)-2-氨基-1-丁醇与(r)-3-氨基-1-丁醇。为解决上述技术问题,本发明首先提供了制备手性胺醇化合物的方法,所述方法包括:以羟基酮为底物,利用胺脱氢酶或其突变蛋白质进行催化反应,得到手性胺醇化合物;所述胺脱氢酶来源于嗜冷芽孢八叠球菌(sporosarcinapsychrophila),其名称为spamdh,其氨基酸序列为序列表中seqidno.2;所述spamdh突变蛋白质为对spamdh的第40、61、68、69、111、113、114、116、134、150、187、239、288、291、292、295、297和/或294位进行突变得到的蛋白质。上述方法中,所述手性胺醇化合物可为2-氨基-1-丁醇、3-氨基-1-丁醇、丙氨醇、苯甘胺醇或2-氨基-2-(4-氟苯基)-1-乙醇;所述羟基酮可为1-羟基-2-丁酮、4-羟基-2-丁酮、丙酮醇、2-羟基苯乙酮、1-(4-氟苯基)-2-羟基-1-乙酮。具体的,所述手性胺醇化合物可为(s)-2-氨基-1-丁醇、(r)-3-氨基-1-丁醇、(s)-丙氨醇、(s)-苯甘胺醇或(s)-2-氨基-2-(4-氟苯基)-1-乙醇。上述方法中,所述spamdh突变蛋白质可为对spamdh进行如下十八种的任一种、任两种、任三种、任四种、任五种、任六种、任七种、任八种、任九种、任十种、任十一种、任十二种、任十三种、任十四种、任十五种、任十六种、任十七种或全部改造得到的蛋白质:x1、将spamdh第40位的亮氨酸残基突变为缬氨酸残基、半胱氨酸残基或苯丙氨酸残基;x2、将spamdh第61位的亮氨酸残基突变为丙氨酸残基、半胱氨酸残基或缬氨酸残基;x3、将spamdh第68位的丝氨酸残基突变为半胱氨酸残基;x4、将spamdh第69位的天冬酰胺残基突变为丝氨酸残基或半胱氨酸残基;x5、将spamdh第111位的异亮氨酸残基突变为苯丙氨酸残基或色氨酸残基;x6、将spamdh第113位的丙氨酸残基突变为半胱氨酸残基;x7、将spamdh第114位的谷氨酸残基突变为缬氨酸残基或谷氨酰胺残基;x8、将spamdh第116位的缬氨酸残基突变为异亮氨酸残基、甲硫氨酸残基、丝氨酸残基或酪氨酸残基;x9、将spamdh第134位的苏氨酸残基突变为丙氨酸残基、苯丙氨酸残基、甘氨酸残基或半胱氨酸残基;x10、将spamdh第150位的苏氨酸残基突变为半胱氨酸残基;x11、将spamdh第187位的丙氨酸残基突变为赖氨酸残基、天冬酰胺残基、脯氨酸残基或苏氨酸残基;x12、将spamdh第239位的亮氨酸残基突变为苯丙氨酸残基或甲硫氨酸残基;x13、将spamdh第288位的丝氨酸残基突变为甲硫氨酸残基或半胱氨酸残基;x14、将spamdh第291位的缬氨酸残基突变为半胱氨酸残基或亮氨酸残基;x15、将spamdh第292位的异亮氨酸残基突变为甲硫氨酸残基或苏氨酸残基;x16、将spamdh第294位的缬氨酸残基突变为半胱氨酸残基或苯丙氨酸残基;x17、将spamdh第295位的丙氨酸残基突变为天冬酰胺残基、苏氨酸残基或色氨酸残基;x18、将spamdh第297位的谷氨酸残基突变为天冬酰胺残基。在本发明的一个实施例中,所述spamdh突变蛋白质为spamdh-l40v、spamdh-l61a、spamdh-l61v、spamdh-l61c、spamdh-s68c、spamdh-n69s、spamdh-i111f、spamdh-i111w、spamdh-a113c、spamdh-e114v、spamdh-e114q、spamdh-v116i、spamdh-v116m、spamdh-v116s、spamdh-v116y、spamdh-t134a、spamdh-t134f、spamdh-t134g、spamdh-t150c、spamdh-a187k、spamdh-a187n、spamdh-a187p、spamdh-a187t、spamdh-l239f、spamdh-l239m、spamdh-v291c、spamdh-v291l、spamdh-i292m、spamdh-i292t、spamdh-a295n、spamdh-a295t、spamdh-a295w、spamdh-s288m、spamdh-e297n、spamdh-n69c/v291c、spamdh-s68c/v291c、spamdh-l40c/i111f、spamdh-i111f/v294c、spamdh-e114v/v291c、spamdh-l40c/i111f/t134f、spamdh-s68c/n69c/v291c、spamdh-i111f/a113c/t134c、spamdh-l40c/i111f/t134f/v294c、spamdh-s68c/n69c/e114v/v291c、spamdh-s68c/n69c/i111f/v291c、spamdh-i111f/e114v/v294c、spamdh-l40f/s68c/n69c/t134c/v291c/v294c、spamdh-s68c/n69c/t134c/v291c、spamdh-l40f/s68c/n69c/a113c/t134c/v291c/v294f、spamdh-s68c/i111f/v294c或spamdh-i111f/s288c/v294c。所述spamdh-l40v为将spamdh的第40位突变为缬氨酸残基得到的蛋白质。所述spamdh-l61a为将spamdh的第61位突变为丙氨酸残基得到的蛋白质。所述spamdh-l61c为将spamdh的第61位突变为半胱氨酸残基得到的蛋白质。所述spamdh-l61v为将spamdh的第61位突变为缬氨酸残基得到的蛋白质。所述spamdh-s68c为将spamdh的第68位突变为半胱氨酸残基得到的蛋白质。所述spamdh-n69s为将spamdh的第69位突变为丝氨酸残基得到的蛋白质。所述spamdh-i111f为将spamdh的第111位突变为苯丙氨酸残基得到的蛋白质。所述spamdh-i111w为将spamdh的第111位突变为色氨酸残基得到的蛋白质。所述spamdh-a113c为将spamdh的第113位突变为半胱氨酸残基得到的蛋白质。所述spamdh-e114v为将spamdh的第114位突变为缬氨酸残基得到的蛋白质。所述spamdh-e114q为将spamdh的第114位突变为谷氨酰胺残基得到的蛋白质。所述spamdh-v116i为将spamdh的第116位突变为异亮氨酸残基得到的蛋白质。所述spamdh-v116m为将spamdh的第116位突变为甲硫氨酸残基得到的蛋白质。所述spamdh-v116s为将spamdh的第116位突变为丝氨酸残基得到的蛋白质。所述spamdh-v116y为将spamdh的第116位突变为酪氨酸残基得到的蛋白质。所述spamdh-t134a为将spamdh的第134位突变为丙氨酸残基得到的蛋白质。所述spamdh-t134f为将spamdh的第134位突变为苯丙氨酸残基得到的蛋白质。所述spamdh-t134g为将spamdh的第134位突变为甘氨酸残基得到的蛋白质。所述spamdh-t150c为将spamdh的第150位突变为半胱氨酸残基得到的蛋白质。所述spamdh-a187k为将spamdh的第187位突变为赖氨酸残基得到的蛋白质。所述spamdh-a187n为将spamdh的第187位突变为天冬酰胺残基得到的蛋白质。所述spamdh-a187p为将spamdh的第187位突变为脯氨酸残基得到的蛋白质。所述spamdh-a187t为将spamdh的第187位突变为苏氨酸残基得到的蛋白质。所述spamdh-l239f为将spamdh的第239位突变为苯丙氨酸残基得到的蛋白质。所述spamdh-l239m为将spamdh的第239位突变为甲硫氨酸残基得到的蛋白质。所述spamdh-v291c为将spamdh的第291位突变为半胱氨酸残基得到的蛋白质。所述spamdh-v291l为将spamdh的第291位突变为亮氨酸残基得到的蛋白质。所述spamdh-i292m为将spamdh的第292位突变为甲硫氨酸残基得到的蛋白质。所述spamdh-i292t为将spamdh的第292位突变为苏氨酸残基得到的蛋白质。所述spamdh-a295n为将spamdh的第295位突变为天冬酰胺氨酸残基得到的蛋白质。所述spamdh-a295t为将spamdh的第295位突变为苏氨酸残基得到的蛋白质。所述spamdh-a295w为将spamdh的第295位突变为色氨酸残基得到的蛋白质。所述spamdh-s288m为将spamdh的第288位突变为甲硫氨酸残基得到的蛋白质。所述spamdh-e297n为将spamdh的第297位突变为天冬酰胺残基得到的蛋白质。所述spamdh-n69c/v291c为将spamdh的第69、291位均突变为半胱氨酸残基得到的蛋白质。所述spamdh-s68c/v291c为将spamdh的第68、291位均突变为半胱氨酸残基得到的蛋白质。所述spamdh-l40c/i111f为将spamdh的第40、111位分别突变为半胱氨酸和苯丙氨酸残基得到的蛋白质。所述spamdh-i111f/v294c为将spamdh的第111、294位分别突变为苯丙氨酸和半胱氨酸残基得到的蛋白质。所述spamdh-e114v/v291c为将spamdh的第114、291位分别突变为缬氨酸和半胱氨酸残基得到的蛋白质。所述spamdh-l40c/i111f/t134f为将spamdh的第40、111和134位分别突变为半胱氨酸、苯丙氨酸和苯丙氨酸残基得到的蛋白质。所述spamdh-s68c/n69c/v291c为将spamdh的第68、69和291位均突变为半胱氨酸残基得到的蛋白质。所述spamdh-i111f/a113c/t134c为将spamdh的第111、113、134位分别突变为苯丙氨酸、半胱氨酸和半胱氨酸残基得到的蛋白质。所述spamdh-l40c/i111f/t134f/v294c为将spamdh的第40、111、134和294位分别突变为半胱氨酸、苯丙氨酸、苯丙氨酸和半胱氨酸残基得到的蛋白质。所述spamdh-s68c/n69c/e114v/v291c为将spamdh的第68、69、114、291位分别突变为半胱氨酸、半胱氨酸、缬氨酸和半胱氨酸残基得到的蛋白质。所述spamdh-s68c/n69c/i111f/v291c为将spamdh的第68、69、111、291位分别突变为半胱氨酸、半胱氨酸、苯丙氨酸和半胱氨酸残基得到的蛋白质。所述spamdh-i111f/e114v/v294c为将spamdh的第111、114、294位分别突变为苯丙氨酸、缬氨酸和半胱氨酸残基得到的蛋白质。所述spamdh-l40f/s68c/n69c/t134c/v291c/v294c为将spamdh的第40、68、69、134、291和294位分别突变为苯丙氨酸、半胱氨酸、半胱氨酸、半胱氨酸、半胱氨酸、半胱氨酸残基得到的蛋白质。所述spamdh-s68c/n69c/t134c/v291c为将spamdh的第68、69、134、291位均突变为半胱氨酸残基得到的蛋白质。所述spamdh-l40f/s68c/n69c/a113c/t134c/v291c/v294f为将spamdh的第40、68、69、113、134、291、294位分别突变为苯丙氨酸、半胱氨酸、半胱氨酸、半胱氨酸、半胱氨酸、半胱氨酸和苯丙氨酸残基得到的蛋白质。所述spamdh-s68c/i111f/v294c为将spamdh的第68、111、294位分别突变为半胱氨酸、苯丙氨酸和半胱氨酸残基得到的蛋白质。所述spamdh-i111f/s288c/v294c为将spamdh的第111、288、294位分别突变为苯丙氨酸、半胱氨酸和半胱氨酸残基得到的蛋白质。本发明还提供了另一种制备手性胺醇化合物的方法,所述方法包括:以羟基酮为底物,利用表达spamdh或所述spamdh突变蛋白质的重组细胞或所述重组细胞的裂解物进行催化反应,得到手性胺醇化合物。上述方法中,所述重组细胞可通过向生物细胞中导入能表达spamdh或所述spamdh突变蛋白质的重组载体获得。所述生物细胞可为微生物。所述微生物可为大肠杆菌,也可以是其他菌。在本发明的一个实施例中,所述微生物为大肠杆菌bl21(de3)。所述重组载体可为将pet24a(+)载体中ndei和xhoi识别序列间的小dna片段替换为spamdh基因或所述spamdh突变蛋白质的编码基因得到的重组质粒。所述spamdh基因可为序列表中seqidno.1所示的核酸分子。所述spamdh突变蛋白质的编码基因可为spamdh-l40v基因、spamdh-l61a基因、spamdh-l61c基因、spamdh-l61v基因、spamdh-s68c基因、spamdh-n69s基因、spamdh-i111f基因、spamdh-i111w基因、spamdh-a113c基因、spamdh-e114v基因、spamdh-e114q基因、spamdh-v116i基因、spamdh-v116m基因、spamdh-v116s基因、spamdh-v116y基因、spamdh-t134a基因、spamdh-t134f基因、spamdh-t134g基因、spamdh-t150c基因、spamdh-a187k基因、spamdh-a187n基因、spamdh-a187p基因、spamdh-a187t基因、spamdh-l239f基因、spamdh-l239m基因、spamdh-v291c基因、spamdh-v291l基因、spamdh-i292m基因、spamdh-i292t基因、spamdh-a295n基因、spamdh-a295t基因、spamdh-a295w基因、spamdh-s288m基因、spamdh-e297n基因、spamdh-n69c/v291c基因、spamdh-s68c/v291c基因、spamdh-l40c/i111f基因、spamdh-i111f/v294c基因、spamdh-e114v/v291c基因、spamdh-l40c/i111f/t134f基因、spamdh-s68c/n69c/v291c基因、spamdh-i111f/a113c/t134c基因、spamdh-l40c/i111f/t134f/v294c基因、spamdh-s68c/n69c/e114v/v291c基因、spamdh-s68c/n69c/i111f/v291c基因、spamdh-i111f/e114v/v294c基因、spamdh-l40f/s68c/n69c/t134c/v291c/v294c基因、spamdh-s68c/n69c/t134c/v291c基因、spamdh-l40f/s68c/n69c/a113c/t134c/v291c/v294f基因、spamdh-s68c/i111f/v294c基因或spamdh-i111f/s288c/v294c基因。所述spamdh-l40v基因为将spamdh基因的第118位由c突变为了g得到的双链dna分子。所述spamdh-l61a基因为将spamdh基因的第181位由c突变为了g、第182位由t突变为了c得到的双链dna分子。所述spamdh-l61c基因为将spamdh基因的第181位由c突变为了t、第182位由t突变为了g、第183位由g突变为了t得到的双链dna分子。所述spamdh-l61v基因为将spamdh基因的第181位由c突变为了g得到的双链dna分子。所述spamdh-s68c基因为将spamdh基因的第202位由a突变为了t得到的双链dna分子。所述spamdh-n69s基因为将spamdh基因的第206位由a突变为了g得到的双链dna分子。所述spamdh-i111f基因为将spamdh基因的第331位由a突变为了t得到的双链dna分子。所述spamdh-i111w基因为将spamdh基因的第331位由a突变为了t、第332位由t突变为了g、第333位由t突变为了g得到的双链dna分子。所述spamdh-a113c基因为将spamdh基因的第337位由g突变为了t、第338位由c突变为了g、第339位由a突变为了t得到的双链dna分子。所述spamdh-e114v基因为将spamdh基因的第341位由a突变为了t、第342位由a突变为了g得到的双链dna分子。所述spamdh-e114q基因为将spamdh基因的第340位由g突变为了c、第342位由a突变为了g得到的双链dna分子。所述spamdh-v116i基因为将spamdh基因的第346位由g突变为了a得到的双链dna分子。所述spamdh-v116m基因为将spamdh基因的第346位由g突变为了a、第348位由t突变为了g得到的双链dna分子。所述spamdh-v116s基因为将spamdh基因的第346位由g突变为了a、第347位由t突变为了g得到的双链dna分子。所述spamdh-v116y基因为将spamdh基因的第346位由g突变为了t、第347位由t突变为了a得到的双链dna分子。所述spamdh-t134a基因为将spamdh基因的第400位由a突变为了g、第402位由c突变为了g得到的双链dna分子。所述spamdh-t134f基因为将spamdh基因的第400位由a突变为了t、第401位由c突变为了t、第402位由c突变为了t得到的双链dna分子。所述spamdh-t134g基因为将spamdh基因的第400位由a突变为了g、第401位由c突变为了g、第402位由c突变为了t得到的双链dna分子。所述spamdh-t150c基因为将spamdh基因的第448位由a突变为了t、第449位由c突变为了g、第450位由c突变为了t得到的双链dna分子。所述spamdh-a187k基因为将spamdh基因的第559位由g突变为了a、第560位由c突变为了a、第561位由c突变为了a得到的双链dna分子。所述spamdh-a187n基因为将spamdh基因的第559位由g突变为了a、第560位由c突变为了a、第561位由c突变为了t得到的双链dna分子。所述spamdh-a187p基因为将spamdh基因的第559位由g突变为了c、第561位由c突变为了g得到的双链dna分子。所述spamdh-a187t基因为将spamdh基因的第559位由g突变为了a、第561位由c突变为了g得到的双链dna分子。所述spamdh-l239f基因为将spamdh基因的第715位由c突变为了t、第717位由g突变为了t得到的双链dna分子。所述spamdh-l239m基因为将spamdh基因的第715位由c突变为了a得到的双链dna分子。所述spamdh-v291c基因为将spamdh基因的第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t得到的双链dna分子。所述spamdh-v291l基因为将spamdh基因的第871位由g突变为了c得到的双链dna分子。所述spamdh-i292m基因为将spamdh基因的第876位由t突变为了g得到的双链dna分子。所述spamdh-i292t基因为将spamdh基因的第875位由t突变为了c、第876位由t突变为了g得到的双链dna分子。所述spamdh-a295n基因为将spamdh基因的第883位由g突变为了a、第884位由c突变为了a、第885位由c突变为了t得到的双链dna分子。所述spamdh-a295t基因为将spamdh基因的第883位由g突变为了a、第885位由c突变为了g得到的双链dna分子。所述spamdh-a295w基因为将spamdh基因的第883位由g突变为了t、第884位由c突变为了g、第885位由c突变为了g得到的双链dna分子。所述spamdh-s288m基因为将spamdh基因的第863位由g突变为了t、第864位由t突变为了g得到的双链dna分子。所述spamdh-e297n基因为将spamdh基因的第889位由g突变为了a、第891位由a突变为了t得到的双链dna分子。所述spamdh-n69c/v291c基因为将spamdh基因的第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t得到的双链dna分子。所述spamdh-s68c/v291c基因为将spamdh基因的第202位由a突变为了t、第204位由t突变为了c、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t得到的双链dna分子。所述spamdh-l40c/i111f基因为将spamdh基因的第118位由c突变为了t、第119位由t突变为了g、第120位由g突变为了c、第331位由a突变为了t得到的双链dna分子。所述spamdh-i111f/v294c基因为将spamdh基因的第331位由a突变为了t、第880位由g突变为了t、第881位由t突变为了g、第882位由t突变为了c得到的双链dna分子。所述spamdh-e114v/v291c基因为将spamdh基因的第341位由a突变为了t、第342位由a突变为了g、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了c得到的双链dna分子。所述spamdh-l40c/i111f/t134f基因为将spamdh基因的第118位由c突变为了t、第119位由t突变为了g、第120位由g突变为了c、第331位由a突变为了t、第400位由a突变为了t、第401位由c突变为了t、第402位由c突变为了t得到的双链dna分子。所述spamdh-s68c/n69c/v291c基因为将spamdh基因的第202位由a突变为了t、第204位由t突变为了c、第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t得到的双链dna分子。所述spamdh-i111f/a113c/t134c基因为将spamdh基因的第331位由a突变为了t、第337位由g突变为了t、第338位由c突变为了g、第339位由a突变为了c、第400位由a突变为了t、第401位由c突变为了g得到的双链dna分子。所述spamdh-l40c/i111f/t134f/v294c基因为将spamdh基因的第118位由c突变为了t、第119位由t突变为了g、第120位由g突变为了c、第331位由a突变为了t、第400位由a突变为了t、第401位由c突变为了t、第402位由c突变为了t、第880位由g突变为了t、第881位由t突变为了g、第882位由t突变为了c得到的双链dna分子。所述spamdh-s68c/n69c/e114v/v291c基因为将spamdh基因的第202位由a突变为了t、第204位由t突变为了c、第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第341位由a突变为了t、第342位由a突变为了g、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t得到的双链dna分子。所述spamdh-s68c/n69c/i111f/v291c基因为将spamdh基因的第202位由a突变为了t、第204位由t突变为了c、第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第331位由a突变为了t、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t得到的双链dna分子。所述spamdh-i111f/e114v/v294c基因为将spamdh基因的第331位由a突变为了t、第341位由a突变为了t、第342位由a突变为了g、第880位由g突变为了t、第881位由t突变为了g、第882位由t突变为了c得到的双链dna分子。所述spamdh-l40f/s68c/n69c/t134c/v291c/v294c基因为将spamdh基因的第118位由c突变为了t、第120位由g突变为了t、第202位由a突变为了t、第204位由t突变为了c、第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第400位由a突变为了t、第401位由c突变为了g,第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t、第880位由g突变为了t、第881位由t突变为了g、第882位由t突变为了c得到的双链dna分子。所述spamdh-s68c/n69c/t134c/v291c基因为将spamdh基因的第202位由a突变为了t、第204位由t突变为了c、第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第400位由a突变为了t、第401位由c突变为了g、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t得到的双链dna分子。所述spamdh-l40f/s68c/n69c/a113c/t134c/v291c/v294f基因为将spamdh基因的第118位由c突变为了t、第120位由g突变为了t、第202位由a突变为了t、第204位由t突变为了c、第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第337位由g突变为了t、第338位由c突变为了g、第339位由a突变为了c、第400位由a突变为了t、第401位由c突变为了g、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t、第880位由g突变为了t得到的双链dna分子。所述spamdh-s68c/i111f/v294c基因为将spamdh基因的第202位由a突变为了t、第204位由t突变为了c、第331位由a突变为了t、第880位由g突变为了t、第881位由t突变为了g、第882位由t突变为了c得到的双链dna分子。所述spamdh-i111f/s288c/v294c基因为将spamdh基因的第331位由a突变为了t、第862位由a突变为了t、第864位由t突变为了c、第880位由g突变为了t、第881位由t突变为了g、第882位由t突变为了c得到的双链dna分子。上文中,以spamdh或其突变体作为生物酶催化底物羟基酮生成手性胺醇化合物的反应中,反应体系中除了含有所述底物和所述spamdh或其突变体外,还可含有spamdh或其突变体的辅酶nad+和/或nh4+。所述底物在所述反应体系中浓度可为1-100mmol/l(如40mmol/l)。所述重组细胞在所述反应体系中的浓度可为50-500g/l(如100g/l)。所述裂解物在所述反应体系中的浓度可为10-50g/l(如20g/l)。所述胺脱氢酶纯酶在所述反应体系中的浓度可为0.1-2g/l(如0.5g/l)。所述nad+在所述反应体系中的浓度可为0.1-2.0mmol/l(如1.0mmol/l)。所述nh4+在所述反应体系中的浓度可为100mmol/l~4mol/l。所述催化反应的反应体系可为向100mmol/l~4mol/l(如1mol/l)的氯化铵/氨水缓冲液(氯化铵和氨水等摩尔比混合得到)中添加如下物质得到:所述羟基酮、所述重组细胞或所述重组细胞的裂解物、nad+(以氧化型辅酶i水溶液形式存在),葡萄糖脱氢酶,葡萄糖,溶菌酶,dnasei(脱氧核糖核酸酶)。各物质的添加量可为:所述羟基酮1-100mmol/l(如20mmol/l)、所述重组细胞50-500g/l(如100g/l)或所述重组细胞的裂解物10-50g/l(如20g/l)、nad+(以氧化型辅酶i水溶液形式存在)0.1-2.0mmol/l(如1.0mmol/l),葡萄糖脱氢酶粉2g/l,葡萄糖100mmol/l,溶菌酶1g/l,dnasei(脱氧核糖核酸酶)6u/ml。所述氯化铵/氨水缓冲液的ph可为7-11。所述葡萄糖脱氢酶粉按照包括如下步骤的方法制备得到:将葡萄糖脱氢酶基因(编码基因的核苷酸序列为seqidno.3)替换pet24a(+)载体ndei和xhoi识别序列间的dna片段,得到重组载体,记为pet24a-gdh;将pet24a-gdh导入大肠杆菌bl21(de3),得到重组菌,记为bl21(de3)/pet24a-gdh。培养bl21(de3)/pet24a-gdh,将所得菌体破碎后离心,将上清液置于冷冻干燥机中获得所述葡萄糖脱氢酶粉。所述反应的温度可为20-35℃,具体可为30℃;所述反应的时间以反应完全为准,一般可为0.5-24小时,具体可为12小时。所述重组细胞的裂解物可由裂解所述重组细胞得到。所述spamdh突变蛋白质,也属于本发明的保护范围。本发明还提供了与spamdh或所述spamdh突变蛋白质相关的生物材料,所述生物材料为下述b1)至b4)中的任一种:b1)编码spamdh或所述spamdh突变蛋白质的核酸分子;b2)含有b1)所述核酸分子的表达盒;b3)含有b1)所述核酸分子的重组载体、或含有b2)所述表达盒的重组载体;b4)含有b1)所述核酸分子的重组微生物、或含有b2)所述表达盒的重组微生物、或含有b3)所述重组载体的重组微生物。其中,所述核酸分子可以是dna,如cdna、基因组dna或重组dna;所述核酸分子也可以是rna,如mrna或hnrna等。b1)所述核酸分子可为所述spamdh基因、所述spamdh-l40v基因、所述spamdh-l61a基因、所述spamdh-l61c基因、所述spamdh-l61v基因、所述spamdh-s68c基因、所述spamdh-n69s基因、所述spamdh-i111f基因、所述spamdh-i111w基因、所述spamdh-a113c基因、所述spamdh-e114v基因、所述spamdh-e114q基因、所述spamdh-v116i基因、所述spamdh-v116m基因、所述spamdh-v116s基因、所述spamdh-v116y基因、所述spamdh-t134a基因、所述spamdh-t134f基因、所述spamdh-t134g基因、所述spamdh-t150c基因、所述spamdh-a187k基因、所述spamdh-a187n基因、所述spamdh-a187p基因、所述spamdh-a187t基因、所述spamdh-l239f基因、所述spamdh-l239m基因、所述spamdh-v291c基因、所述spamdh-v291l基因、所述spamdh-i292m基因、所述spamdh-i292t基因、所述spamdh-a295n基因、所述spamdh-a295t基因、所述spamdh-a295w基因、所述spamdh-s288m基因、所述spamdh-e297n基因、所述spamdh-n69c/v291c基因、所述spamdh-s68c/v291c基因、所述spamdh-l40c/i111f基因、所述spamdh-i111f/v294c基因、所述spamdh-e114v/v291c基因、所述spamdh-l40c/i111f/t134f基因、所述spamdh-s68c/n69c/v291c基因、所述spamdh-i111f/a113c/t134c基因、所述spamdh-l40c/i111f/t134f/v294c基因、所述spamdh-s68c/n69c/e114v/v291c基因、所述spamdh-s68c/n69c/i111f/v291c基因、所述spamdh-i111f/e114v/v294c基因、所述spamdh-l40f/s68c/n69c/t134c/v291c/v294c基因、所述spamdh-s68c/n69c/t134c/v291c基因、所述spamdh-l40f/s68c/n69c/a113c/t134c/v291c/v294f基因、所述spamdh-s68c/i111f/v294c基因或所述spamdh-i111f/s288c/v294c基因。所述的核酸分子的表达盒是指能够在宿主细胞中表达spamdh或所述spamdh突变蛋白质的dna,该dna不但可包括启动spamdh或所述spamdh突变蛋白质编码基因转录的启动子,还可包括spamdh或所述spamdh突变蛋白质转录的终止子。更进一步的,所述表达盒还可包括增强子序列。所述的核酸分子的重组载体可为携带有spamdh或所述spamdh突变蛋白质编码基因的细菌质粒(如在细菌中表达的基于t7启动子的表达载体,具体如pet-24a等)、噬菌体、酵母质粒(如yep系列载体等)或逆转录病毒包装质粒。所述重组载体为将所述核酸分子插入表达载体中得到的载体。所述重组载体可为pet24a-spamdh-l40v、pet24a-spamdh-l61a、pet24a-spamdh-l61c、pet24a-spamdh-l61v、pet24a-spamdh-s68c、pet24a-spamdh-n69s、pet24a-spamdh-i111f、pet24a-spamdh-i111w、pet24a-spamdh-a113c、pet24a-spamdh-e114v、pet24a-spamdh-e114q、pet24a-spamdh-v116i、pet24a-spamdh-v116m、pet24a-spamdh-v116s、pet24a-spamdh-v116y、pet24a-spamdh-t134a、pet24a-spamdh-t134f、pet24a-spamdh-t134g、pet24a-spamdh-t150c、pet24a-spamdh-a187k、pet24a-spamdh-a187n、pet24a-spamdh-a187p、pet24a-spamdh-a187t、pet24a-spamdh-l239f、pet24a-spamdh-l239m、pet24a-spamdh-v291c、pet24a-spamdh-v291l、pet24a-spamdh-i292m、pet24a-spamdh-i292t、pet24a-spamdh-a295n、pet24a-spamdh-a295t、pet24a-spamdh-a295w、pet24a-spamdh-s288m、pet24a-spamdh-e297n、pet24a-spamdh-n69c/v291c、pet24a-spamdh-s68c/v291c、pet24a-spamdh-l40c/i111f、pet24a-spamdh-i111f/v294c、pet24a-spamdh-e114v/v291c、pet24a-spamdh-l40c/i111f/t134f、pet24a-spamdh-s68c/n69c/v291c、pet24a-spamdh-i111f/a113c/t134c、pet24a-spamdh-l40c/i111f/t134f/v294c、pet24a-spamdh-s68c/n69c/e114v/v291c、pet24a-spamdh-s68c/n69c/i111f/v291c、pet24a-spamdh-i111f/e114v/v294c、pet24a-spamdh-l40f/s68c/n69c/t134c/v291c/v294c、pet24a-spamdh-s68c/n69c/t134c/v291c、pet24a-spamdh-l40f/s68c/n69c/a113c/t134c/v291c/v294f、pet24a-spamdh-s68c/i111f/v294c或pet24a-spamdh-i111f/s288c/v294c。所述的含有编码spamdh或所述spamdh突变蛋白质的核酸分子的重组微生物可为携带有spamdh或所述spamdh突变蛋白质编码基因的酵母、细菌、藻或真菌,如大肠杆菌等。所述重组微生物可为bl21(de3)/pet24a-spamdh-l40v、bl21(de3)/pet24a-spamdh-l61a、bl21(de3)/pet24a-spamdh-l61c、bl21(de3)/pet24a-spamdh-l61v、bl21(de3)/pet24a-spamdh-s68c、bl21(de3)/pet24a-spamdh-n69s、bl21(de3)/pet24a-spamdh-i111f、bl21(de3)/pet24a-spamdh-i111w、bl21(de3)/pet24a-spamdh-a113c、bl21(de3)/pet24a-spamdh-e114v、bl21(de3)/pet24a-spamdh-e114q、bl21(de3)/pet24a-spamdh-v116i、bl21(de3)/pet24a-spamdh-v116m、bl21(de3)/pet24a-spamdh-v116s、bl21(de3)/pet24a-spamdh-v116y、bl21(de3)/pet24a-spamdh-t134a、bl21(de3)/pet24a-spamdh-t134f、bl21(de3)/pet24a-spamdh-t134g、bl21(de3)/pet24a-spamdh-t150c、bl21(de3)/pet24a-spamdh-a187k、bl21(de3)/pet24a-spamdh-a187n、bl21(de3)/pet24a-spamdh-a187p、bl21(de3)/pet24a-spamdh-a187t、bl21(de3)/pet24a-spamdh-l239f、bl21(de3)/pet24a-spamdh-l239m、bl21(de3)/pet24a-spamdh-v291c、bl21(de3)/pet24a-spamdh-v291l、bl21(de3)/pet24a-spamdh-i292m、bl21(de3)/pet24a-spamdh-i292t、bl21(de3)/pet24a-spamdh-a295n、bl21(de3)/pet24a-spamdh-a295t、bl21(de3)/pet24a-spamdh-a295w、bl21(de3)/pet24a-spamdh-s288m、bl21(de3)/pet24a-spamdh-e297n、bl21(de3)/pet24a-spamdh-n69c/v291c、bl21(de3)/pet24a-spamdh-s68c/v291c、bl21(de3)/pet24a-spamdh-l40c/i111f、bl21(de3)/pet24a-spamdh-i111f/v294c、bl21(de3)/pet24a-spamdh-e114v/v291c、bl21(de3)/pet24a-spamdh-l40c/i111f/t134f、bl21(de3)/pet24a-spamdh-s68c/n69c/v291c、bl21(de3)/pet24a-spamdh-i111f/a113c/t134c、bl21(de3)/pet24a-spamdh-l40c/i111f/t134f/v294c、bl21(de3)/pet24a-spamdh-s68c/n69c/e114v/v291c、bl21(de3)/pet24a-spamdh-s68c/n69c/i111f/v291c、bl21(de3)/pet24a-spamdh-i111f/e114v/v294c、bl21(de3)/pet24a-spamdh-l40f/s68c/n69c/t134c/v291c/v294c、bl21(de3)/pet24a-spamdh-s68c/n69c/t134c/v291c、bl21(de3)/pet24a-spamdh-l40f/s68c/n69c/a113c/t134c/v291c/v294f、bl21(de3)/pet24a-spamdh-s68c/i111f/v294c或bl21(de3)/pet24a-spamdh-i111f/s288c/v294c。本发明还提供了下述任一产品:m1、由1-羟基-2-丁酮与spamdh组成的成套产品;m2、由1-羟基-2-丁酮与所述spamdh突变蛋白质组成的成套产品;m3、由1-羟基-2-丁酮与权利要求7中所述生物材料组成的成套产品;m4、由4-羟基-2-丁酮与spamdh组成的成套产品;m5、由4-羟基-2-丁酮与所述spamdh突变蛋白质组成的成套产品;m6、由4-羟基-2-丁酮与权利要求7中所述生物材料组成的成套产品;m7、由丙酮醇与spamdh组成的成套产品;m8、由丙酮醇与所述spamdh突变蛋白质组成的成套产品;m9、由丙酮醇与权利要求7中所述生物材料组成的成套产品;m10、由2-羟基苯乙酮与spamdh组成的成套产品;m11、由2-羟基苯乙酮与所述spamdh突变蛋白质组成的成套产品;m12、由2-羟基苯乙酮与权利要求7中所述生物材料组成的成套产品;m13、由1-(4-氟苯基)-2-羟基-1-乙酮与spamdh组成的成套产品;m14、由1-(4-氟苯基)-2-羟基-1-乙酮与所述spamdh突变蛋白质组成的成套产品;m15、由1-(4-氟苯基)-2-羟基-1-乙酮与权利要求7中所述生物材料组成的成套产品。m1-m3所述成套产品均可用于制备(s)-2-氨基-1-丁醇。m4-m6所述成套产品均可用于制备(r)-3-氨基-1-丁醇。m7-m9所述成套产品均可用于制备(s)-丙氨醇。m10-m12所述成套产品均可用于制备(s)-苯甘胺醇。m13-m15所述成套产品均可用于制备(s)-2-氨基-2-(4-氟苯基)-1-乙醇。本发明还提供了spamdh、所述spamdh突变蛋白质、所述生物材料或所述产品的下述任一应用:z1、在制备手性胺醇化合物中的应用;z2、在制备生产手性胺醇化合物产品中的应用;z3、在催化羟基酮生成手性胺醇化合物中的应用;z4、在制备催化羟基酮生成手性胺醇化合物产品中的应用。本发明中,所述手性胺醇化合物可为2-氨基-1-丁醇、3-氨基-1-丁醇、丙氨醇、苯甘胺醇或2-氨基-2-(4-氟苯基)-1-乙醇;所述羟基酮可为1-羟基-2-丁酮、4-羟基-2-丁酮、丙酮醇、2-羟基苯乙酮、1-(4-氟苯基)-2-羟基-1-乙酮。具体的,所述手性胺醇化合物可为(s)-2-氨基-1-丁醇、(r)-3-氨基-1-丁醇、(s)-丙氨醇、(s)-苯甘胺醇或(s)-2-氨基-2-(4-氟苯基)-1-乙醇。本发明应用定向进化技术和方法对来源于嗜冷芽孢八叠球菌(sporosarcinapsychrophila)的胺脱氢酶spamdh进行酶改造,获得了其突变体,所得突变体可以催化羟基酮底物合成以1-羟基-2-丁酮、4-羟基-2-丁酮为代表的手性胺醇化合物,并且具有高产率、高ee值的特点。本发明操作方便,具有产物光学纯度高等优点,在生物催化制备手性胺醇化合物具有较好的工业应用前景。附图说明图1为胺脱氢酶l40/a113/t134/v294突变体文库a和m65/s68/n69/s288突变体文库a构建引物设计示意图。f18与r18、f19与r19、f20与r20分别为表1中的spamdh-f18与spamdh-r18、spamdh-f19与spamdh-r19、spamdh-f20与spamdh-r20。图2为胺脱氢酶spamdh或其突变体催化还原1-羟基-2-丁酮生成(s)-2-氨基-1-丁醇和4-羟基-2-丁酮生成(r)-3-氨基-1-丁醇示意图。图3为胺脱氢酶spamdh或其突变体催化还原1-羟基-2-丁酮生成(s)-2-氨基-1-丁醇和4-羟基-2-丁酮生成(r)-3-氨基-1-丁醇反应的hplc检测结果图谱。a:混旋-2-氨基-1-丁醇标准品液相色谱结果;b:样品(s)-2-氨基-1-丁醇液相色谱结果;c:混旋3-氨基-1-丁醇标准品液相色谱结果;d:样品(r)-3-氨基-1-丁醇液相色谱结果。图4为胺脱氢酶spamdh或其突变体催化羟基酮底物检测结果,百分号后的“c”表示转化率。图5为胺脱氢酶spamdh及其突变体催化1-羟基-2-丁酮的检测结果。具体实施方式下面结合具体实施方式对本发明进行进一步的详细描述,给出的实施例仅为了阐明本发明,而不是为了限制本发明的范围。以下提供的实施例可作为本
技术领域:
普通技术人员进行进一步改进的指南,并不以任何方式构成对本发明的限制。下述实施例中的实验方法,如无特殊说明,均为常规方法,按照本领域内的文献所描述的技术或条件或者按照产品说明书进行。下述实施例中所用的材料、试剂、仪器等,如无特殊说明,均可从商业途径得到。以下实施例中的定量试验,均设置三次重复实验,结果取平均值。下述实施例中,如无特殊说明,序列表中各核苷酸序列的第1位均为相应dna/rna的5′末端核苷酸,末位均为相应dna/rna的3′末端核苷酸。2×high-fidelitymastermix:擎科生物科技有限公司,货号:tp001;1-羟基-2-丁酮:上海毕得医药科技有限公司,货号:bd59017;4-羟基-2-丁酮:上海阿拉丁生化科技股份有限公司,货号:h106330;丙酮醇:上海阿拉丁生化科技股份有限公司,货号:h111149;2-羟基苯乙酮:上海艾览化工科技有限公司,货号:a42334;1-(4-氟苯基)-2-羟基-1-乙酮:北京百灵威科技有限公司,货号:042861。实施例1、胺脱氢酶spamdh基因或其突变体的工程菌的制备一、胺脱氢酶spamdh基因工程菌株的制备将来源于嗜冷芽孢八叠球菌(sporosarcinapsychrophila)的胺脱氢酶(spamdh)的编码基因以大肠杆菌为宿主细胞进行密码子优化,得到序列表中seqidno.1所示的spamdh基因。spamdh的氨基酸序列如序列表中seqidno.2所示。全基因合成序列表中seqidno.1所示的spamdh基因,将pet24a(+)载体(novagen)中ndei和xhoi识别序列间的小dna片段替换为seqidno.1所示的spamdh基因,得到重组载体,将该重组载体记为pet24a-spamdh,pet24a-spamdh能表达序列表中seqidno.2所示的spamdh,spamdh基因的表达由t7启动子驱动。将重组载体pet24a-spamdh导入大肠杆菌bl21(de3)中,得到重组菌,记为bl21(de3)/pet24a-spamdh,即为胺脱氢酶spamdh基因工程菌株。将pet24a(+)导入大肠杆菌bl21(de3)中,得到重组菌bl21(de3)/pet24a,作为对照菌株。二、胺脱氢酶spamdh基因突变体工程菌株的制备分别构建各个胺脱氢酶spamdh突变体的胺脱氢酶spamdh基因突变体工程菌株。胺脱氢酶spamdh的突变体是通过设计简并引物构建突变体文库筛选获得,或者通过设计突变引物获得。设计的简并引物和突变引物序列如表1和表2所示。表1突变体文库构建所用引物序列注:表1中,有多条单链dna的引物,引物spamdh-f1、spamdh-f2、spamdh-f3、spamdh-f4、spamdh-f5、spamdh-f6、spamdh-f7、spamdh-f8、spamdh-f9、spamdh-f10、spamdh-r11、spamdh-r12、spamdh-r13、spamdh-r14、spamdh-r15、spamdh-r16、spamdh-r17、spamdh-r18各单链dna按照表中从上至下的顺序按照摩尔比:12:6:1:1混合,引物spamdh-f18、spamdh-r18、spamdh-f19、spamdh-r19、spamdh-f21、spamdh-r21、spamdh-f22、spamdh-r22各单链dna按照表中从上至下的顺序按照摩尔比:2:1混合,引物spamdh-f20、spamdh-f23各单链dna按照表中从上至下的顺序按照摩尔比:4:2:2:1混合;r表示a或g,y表示c或t,m表示a或c,k表示g或t,s表示g或c,w表示a或t,h表示a或t或c,b表示g或t或c,v表示g或a或c,d表示g或a或t,n表示a或t或g或c。表2突变体及所用引物序列其中,l40/a113/t134/v294突变体文库a和m65/s68/n69/s288突变体文库a构建的引物设计如图1所示。具体构建方法如下:1、两轮mega-pcr反应构建l40突变体文库、l61突变体文库、s68突变体文库、n69突变体文库、i111突变体文库、a113突变体文库、e114突变体文库、v116突变体文库、t134突变体文库、t150突变体文库、a187突变体文库、l239突变体文库、s288突变体文库、v291突变体文库、i292突变体文库、a295突变体文库和e297突变体文库以步骤一得到的重组载体pet24a-spamdh为模板,构建m65/s68/n69/s288突变体文库a以下文的重组载体pet24a-spamdh-v291c为模板,构建m65/s68/n69/s288突变体文库b以下文的重组载体pet24a-spamdh-i111f/v294c为模板,构建spamdh-e114v/v291c突变体以下文的重组载体pet24a-spamdh-v291c为模板,构建spamdh-i111f/e114v/v294c突变体以下文的重组载体pet24a-spamdh-i111f/v294c为模板,构建spamdh-s68c/n69c/e114v/v291c和spamdh-s68c/n69c/i111f/v291c突变体均以下文的重组载体pet24a-spamdh-s68c/n69c/v291c为模板,分别采用各突变体对应的引物对进行两轮mega-pcr反应。同一突变体库或突变体构建时,第一轮pcr与第二轮pcr的模板相同。pcr反应体系及程序如下:第一轮pcr体系为50μl:模板(150ng/μl)1μl;2×high-fidelitymastermix25μl;ddh2o22μl;前引物(10μmol/l)1μl;后引物(10μmol/l)1μl。所用引物具体序列如表1及表2中所示,同一引物对中的前引物和后引物分别标有“f”和“r”。第一轮pcr程序:预变性98℃2分钟;变性98℃10秒,退火55℃15秒,延伸72℃30秒或50秒,循坏30次;终延伸72℃5分钟。第二轮pcr体系为50μl:模板(150ng/μl)1μl;2×high-fidelitymastermix25μl;ddh2o22μl;第一轮pcr产物2μl。第二轮pcr程序:预变性98℃2分钟;变性98℃10秒,退火60℃15秒,延伸72℃2分钟,循坏30次;终延伸72℃5分钟。2、overlappcr反应构建l40/a113/t134/v294突变体文库a以下文的重组载体pet24a-spamdh-i111f为模板(即第一轮pcr和第三轮pcr的模板均为pet24a-spamdh-i111f),构建l40/a113/t134/v294突变体文库b以下文的重组载体pet24a-spamdh-s68c/n69c/v291c为模板(即第一轮pcr和第三轮pcr的模板均为pet24a-spamdh-s68c/n69c/v291c)。以构建l40/a113/t134/v294突变体文库a为例,突变体文库构建引物设计示意图如图1所示,进行overlappcr反应。pcr反应体系及程序如下:第一轮pcr反应扩增获得f18r18、f19r19产物。pcr体系为50μl:模板(150ng/μl)1μl;2×high-fidelitymastermix25μl;ddh2o22μl;前引物(10μmol/l)1μl;后引物(10μmol/l)1μl。所用引物具体序列如表1中所示,同一引物对中的前引物和后引物分别标有“f”和“r”。第一轮pcr程序:预变性98℃2分钟;变性98℃10秒,退火55℃15秒,延伸72℃30秒,循坏次数30次;终延伸72℃5分钟。回收第一轮所得pcr产物,将由spamdh-f18与spamdh-r18得到的产物记为f18r18,将由spamdh-f19与spamdh-r19得到的产物记为f19r19。第二轮pcr反应扩增获得f18r19产物。pcr体系为50μl:模板(第一轮获得的f18r18:f19r19摩尔混合的混合液)1μl;2×high-fidelitymastermix25μl;ddh2o22μl;前引物spamdh-f18(10μmol/l)1μl;后引物spamdh-r19(10μmol/l)1μl。第二轮pcr程序:预变性98℃2分钟;变性98℃10秒;退火55℃15秒;延伸72℃1分钟,循坏30次;终延伸72℃5分钟。将由spamdh-f18与spamdh-r19得到的产物记为f18r19。第三轮pcr反应扩增获得重组质粒。pcr体系为50μl:模板(150ng/μl)1μl;2×high-fidelitymastermix25μl;ddh2o22μl;第二轮pcrf18r19产物2μl。第三轮pcr程序:预变性98℃2分钟;变性98℃10秒,退火60℃15秒,延伸72℃2分钟,循坏30次;终延伸72℃5分钟。l40/a113/t134/v294突变体文库b的构建按照上述方法进行,将spamdh-f18、spamdh-r18、spamdh-f19、spamdh-r19分别替换为spamdh-f21、spamdh-r21、spamdh-f22、spamdh-r22,得到pcr产物。3、胺脱氢酶spamdh基因突变体工程菌株的获得最后一轮pcr结束后,向各反应体系中加入dpni酶2μl,37℃消化2小时,并取1μl电转入e.colibl21(de3)感受态细胞中,放入37℃培养箱倒置培养12-16小时,待转化子长出,挑取转化子提取质粒进行测序,获得目的核苷酸发生不定向突变或定向突变的转化子,即为胺脱氢酶spamdh基因突变体工程菌株,该菌株所含有的目的核苷酸发生不定向突变或定向突变的重组质粒,即为表达胺脱氢酶spamdh基因突变体的表达载体。实施例2、胺脱氢酶spamdh或其突变体的表达及全细胞、粗酶粉与纯酶液的制备对实施例1中制备的重组菌bl21(de3)/pet24a-spamdh、bl21(de3)/pet24a以及各个胺脱氢酶spamdh基因突变体工程菌株进行诱导表达,获得各菌株的全细胞、粗酶粉与纯酶液,每个菌株的操作步骤均如下:挑取菌株至含有50μg/ml卡那霉素的5mllb液体培养基中,37℃、220rpm震荡过夜12h,再按照体积百分含量1%接种量接种到含有50μg/ml卡那霉素的tb液体培养基中,37℃培养至od600为0.7时,加入终浓度为0.1mmol/l的iptg,20℃、220rpm诱导表达12h后,4℃、4,000rpm离心10min,收集沉淀菌体(即全细胞),并将收集的菌体用磷酸盐缓冲液(50mmol/l,ph7.4)重悬得到菌体悬液,然后在冰浴条件下超声破碎菌体细胞,得到超声破碎后的样品(即为粗酶液)。将粗酶液于8,000rpm,4℃离心30min收集上清,将所得上清液置于-80℃冷冻后再用真空干燥仪冷冻干燥获得粗酶粉。将粗酶液于12,000rpm,4℃离心60min收集上清,将所得上清液用0.45μm水系滤膜过滤后用atka蛋白纯化仪进行纯化,所用层析柱为histraphp5ml预装柱(ge),货号17524802。上样及平衡缓冲液a(50mmol/l磷酸盐缓冲液,0.5mol/lnacl,20mmol/l咪唑,ph8.0);洗脱缓冲液b(50mmol/l磷酸盐缓冲液,0.5mol/lnacl,500mmol/l咪唑,ph8.0),洗脱流速2ml/min;收集洗脱液(即为纯酶液)。实施例3、胺脱氢酶spamdh或其突变体催化1-羟基-2-丁酮生成(s)-2-氨基-1-丁醇用实施例2制备的胺脱氢酶spamdh或其突变体的粗酶粉或全细胞催化1-羟基-2-丁酮生成(s)-2-氨基-1-丁醇,示意图如图2所示,并利用bl21(de3)/pet24a作为对照。spamdh或其突变体的催化反应的反应体系通过向1mol/l的氯化铵/氨水缓冲液(氯化铵和氨水等摩尔比混合得到,ph8.5)中添加如下量的物质得到:底物1-羟基-2-丁酮20mmol/l或40mmol/l,胺脱氢酶spamdh或其突变体的粗酶粉20g/l或全细胞100g/l,nad+(以氧化型辅酶i水溶液形式存在)1mmol/l,gdh粗酶粉2g/l,葡萄糖100mmol/l,溶菌酶1g/l(北京索莱宝科技有限公司,cas:12650-88-3,比活力为20000u/mg),dnasei(脱氧核糖核酸酶,北京鼎国昌盛生物技术有限责任公司,cas:9003-98-9,比活力为2000u/mg)6u/ml。上述gdh粗酶粉为葡萄糖脱氢酶酶粉,能够催化底物葡萄糖和nad+生成葡萄糖酸和nadh,加该酶是为了达到辅酶nadh再生的目的,该酶粉按照如下方法制备:将葡萄糖脱氢酶基因(编码基因的核苷酸序列为seqidno.3)替换pet24a(+)载体ndei和xhoi识别序列间的dna片段,得到重组载体,记为pet24a-gdh;将pet24a-gdh导入大肠杆菌bl21(de3),得到重组菌,记为bl21(de3)/pet24a-gdh。培养bl21(de3)/pet24a-gdh,将所得菌体破碎后离心,将上清液置于冷冻干燥机中获得gdh粗酶粉。检测gdh粗酶粉,比酶活为2.04u/mg,酶活测定方法如下:通过监测nadh在340nm处吸光度的减少来计算酶活,酶活定义:在ph为8.5、温度为30℃,底物为1mmol/lnad+,10mmol/l的葡萄糖的条件下,每分钟内消耗1μmol/lnadh所需要的酶量。酶活计算公式:酶活(u)=ew×v×103/(6220×0.3),式中,ew表示每分钟内340nm处吸光值的变化,v表示反应液体积,单位ml,6220为辅酶的摩尔消光系数,单位l/(mol×cm),0.3为光程距离,单位cm)。将上述spamdh或其突变体的催化反应的反应体系于30℃反应12小时。反应结束后,计算转化率并进行立体选择性分析,具体方法如下:将上述反应得到的反应液煮沸5分钟,12,000rpm离心10分钟,去除沉淀,保留上清液,用衍生剂衍生(衍生剂配制方法:取0.343g邻苯二甲醛+5ml无水乙醇+0.147gn-乙酰-l半胱氨酸,用0.4mol/l硼酸缓冲液(ph=9.5)定容到25ml,避光备用;衍生方法:300μl0.4mol/l硼酸缓冲液(ph=9.5)+150μl超纯水+200μl衍生剂+100μl反应液,混匀并静置2分钟,12,000rpm离心10分钟后,取上清,用滤膜过滤后上样),然后进行hplc检测反应。hplc检测条件:agilentsb-aqc18柱(4.6*250mm,5μm),检测波长334nm,柱温:35℃,流速:1ml/min,上样量:10μl。梯度洗脱程序如表3所示。表3hplc的梯度洗脱程序注:表3中的%表示体积百分含量。hplc检测结果如表4所示。转化率=a1/a2×100%;a1:液相色谱分析获得的(s)-2-氨基-1-丁醇峰面积值;a2:液相色谱分析获得的标准品(s)-2-氨基-1-丁醇的峰面积值。bl21(de3)/pet24a的反应产物中并无(s)-2-氨基-1-丁醇的生成,胺脱氢酶spamdh或其突变体催化还原1-羟基-2-丁酮生成(s)-2-氨基-1-丁醇反应的hplc检测结果图谱如图3所示,a:混旋-2-氨基-1-丁醇标准品液相色谱结果;b:样品(s)-2-氨基-1-丁醇液相色谱结果。结果显示,胺脱氢酶spamdh或其突变体可以不对称还原催化1-羟基-2-丁酮生成(s)-2-氨基-1-丁醇,转化率为47-99%,立体选择性大于99%(s)。表4为胺脱氢酶spamdh及其突变体催化1-羟基-2-丁酮检测结果注:a反应体系中底物1-羟基-2-丁酮20mmol/l、胺脱氢酶spamdh或其突变体的粗酶粉20g/l;b反应体系中底物1-羟基-2-丁酮40mmol/l、胺脱氢酶spamdh或其突变体的全细胞100g/l;-表示没有测定;ee(立体选择性)=(as-ar)/(as+ar)×100%;as:液相色谱分析获得的(s)-2-氨基-1-丁醇的峰面积值;ar:液相色谱分析获得的(r)-2-氨基-1-丁醇的峰面积值。表4第1列中,数字为胺脱氢酶spamdh突变位点,数字前后的字母分别表示突变前后的氨基酸。相应菌株的dna测序结果显示,将表达spamdh-l40v的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-l40v和pet24a-spamdh-l40v,该菌株和载体中spamdh基因的第118位由c突变为了g,将突变后的spamdh基因记为spamdh-l40v基因;将表达spamdh-l61a的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-l61a和pet24a-spamdh-l61a,该菌株和载体中spamdh基因的第181位由c突变为了g、第182位由t突变为了c,将突变后的spamdh基因记为spamdh-l61a基因;将表达spamdh-l61c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-l61c和pet24a-spamdh-l61c,该菌株和载体中spamdh基因的第181位由c突变为了t、第182位由t突变为了g、第183位由g突变为了t,将突变后的spamdh基因记为spamdh-l61c基因;将表达spamdh-l61v的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-l61v和pet24a-spamdh-l61v,该菌株和载体中spamdh基因的第181位由c突变为了g,将突变后的spamdh基因记为spamdh-l61v基因;将表达spamdh-s68c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-s68c和pet24a-spamdh-s68c,该菌株和载体中spamdh基因的第202位由a突变为了t,将突变后的spamdh基因记为spamdh-s68c基因;将表达spamdh-n69s的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-n69s和pet24a-spamdh-n69s,该菌株和载体中spamdh基因的第206位由a突变为了g,将突变后的spamdh基因记为spamdh-n69s基因;将表达spamdh-i111f的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-i111f和pet24a-spamdh-i111f,该菌株和载体中spamdh基因的第331位由a突变为了t,将突变后的spamdh基因记为spamdh-i111f基因;将表达spamdh-i111w的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-i111w和pet24a-spamdh-i111w,该菌株和载体中spamdh基因的第331位由a突变为了t、第332位由t突变为了g、第333位由t突变为了g,将突变后的spamdh基因记为spamdh-i111w基因;将表达spamdh-a113c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-a113c和pet24a-spamdh-a113c,该菌株和载体中spamdh基因的第337位由g突变为了t、第338位由c突变为了g、第339位由a突变为了t,将突变后的spamdh基因记为spamdh-a113c基因;将表达spamdh-e114v的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-e114v和pet24a-spamdh-e114v,该菌株和载体中spamdh基因的第341位由a突变为了t、第342位由a突变为了g,将突变后的spamdh基因记为spamdh-e114v基因;将表达spamdh-e114q的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-e114q和pet24a-spamdh-e114q,该菌株和载体中spamdh基因的第340位由g突变为了c、第342位由a突变为了g,将突变后的spamdh基因记为spamdh-e114q基因;将表达spamdh-v116i的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-v116i和pet24a-spamdh-v116i,该菌株和载体中spamdh基因的第346位由g突变为了a,将突变后的spamdh基因记为spamdh-v116i基因;将表达spamdh-v116m的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-v116m和pet24a-spamdh-v116m,该菌株和载体中spamdh基因的第346位由g突变为了a、第348位由t突变为了g,将突变后的spamdh基因记为spamdh-v116m基因;将表达spamdh-v116s的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-v116s和pet24a-spamdh-v116s,该菌株和载体中spamdh基因的第346位由g突变为了a、第347位由t突变为了g,将突变后的spamdh基因记为spamdh-v116s基因;将表达spamdh-v116y的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-v116y和pet24a-spamdh-v116y,该菌株和载体中spamdh基因的第346位由g突变为了t、第347位由t突变为了a,将突变后的spamdh基因记为spamdh-v116y基因;将表达spamdh-t134a的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-t134a和pet24a-spamdh-t134a,该菌株和载体中spamdh基因的第400位由a突变为了g、第402位由c突变为了g,将突变后的spamdh基因记为spamdh-t134a基因;将表达spamdh-t134f的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-t134f和pet24a-spamdh-t134f,该菌株和载体中spamdh基因的第400位由a突变为了t、第401位由c突变为了t、第402位由c突变为了t,将突变后的spamdh基因记为spamdh-t134f基因;将表达spamdh-t134g的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-t134g和pet24a-spamdh-t134g,该菌株和载体中spamdh基因的第400位由a突变为了g、第401位由c突变为了g、第402位由c突变为了t,将突变后的spamdh基因记为spamdh-t134g基因;将表达spamdh-t150c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-t150c和pet24a-spamdh-t150c,该菌株和载体中spamdh基因的第448位由a突变为了t、第449位由c突变为了g、第450位由c突变为了t,将突变后的spamdh基因记为spamdh-t150c基因;将表达spamdh-a187k的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-a187k和pet24a-spamdh-a187k,该菌株和载体中spamdh基因的第559位由g突变为了a、第560位由c突变为了a、第561位由c突变为了a,将突变后的spamdh基因记为spamdh-a187k基因;将表达spamdh-a187n的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-a187n和pet24a-spamdh-a187n,该菌株和载体中spamdh基因的第559位由g突变为了a、第560位由c突变为了a、第561位由c突变为了t,将突变后的spamdh基因记为spamdh-a187n基因;将表达spamdh-a187p的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-a187p和pet24a-spamdh-a187p,该菌株和载体中spamdh基因的第559位由g突变为了c、第561位由c突变为了g,将突变后的spamdh基因记为spamdh-a187p基因;将表达spamdh-a187t的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-a187t和pet24a-spamdh-a187t,该菌株和载体中spamdh基因的第559位由g突变为了a、第561位由c突变为了g,将突变后的spamdh基因记为spamdh-a187t基因;将表达spamdh-l239f的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-l239f和pet24a-spamdh-l239f,该菌株和载体中spamdh基因的第715位由c突变为了t、第717位由g突变为了t,将突变后的spamdh基因记为spamdh-l239f基因;将表达spamdh-l239m的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-l239m和pet24a-spamdh-l239m,该菌株和载体中spamdh基因的第715位由c突变为了a,将突变后的spamdh基因记为spamdh-l239m基因;将表达spamdh-v291c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-v291c和pet24a-spamdh-v291c,该菌株和载体中spamdh基因的第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t,将突变后的spamdh基因记为spamdh-v291c基因;将表达spamdh-v291l的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-v291l和pet24a-spamdh-v291l,该菌株和载体中spamdh基因的第871位由g突变为了c,将突变后的spamdh基因记为spamdh-v291l基因;将表达spamdh-i292m的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-i292m和pet24a-spamdh-i292m,该菌株和载体中spamdh基因的第876位由t突变为了g,将突变后的spamdh基因记为spamdh-i292m基因;将表达spamdh-i292t的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-i292t和pet24a-spamdh-i292t,该菌株和载体中spamdh基因的第875位由t突变为了c、第876位由t突变为了g,将突变后的spamdh基因记为spamdh-i292t基因;将表达spamdh-a295n的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-a295n和pet24a-spamdh-a295n,该菌株和载体中spamdh基因的第883位由g突变为了a、第884位由c突变为了a、第885位由c突变为了t,将突变后的spamdh基因记为spamdh-a295n基因;将表达spamdh-a295t的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-a295t和pet24a-spamdh-a295t,该菌株和载体中spamdh基因的第883位由g突变为了a、第885位由c突变为了g,将突变后的spamdh基因记为spamdh-a295t基因;将表达spamdh-a295w的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-a295w和pet24a-spamdh-a295w,该菌株和载体中spamdh基因的第883位由g突变为了t、第884位由c突变为了g、第885位由c突变为了g,将突变后的spamdh基因记为spamdh-a295w基因;将表达spamdh-s288m的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-s288m和pet24a-spamdh-s288m,该菌株和载体中spamdh基因的第863位由g突变为了t、第864位由t突变为了g,将突变后的spamdh基因记为spamdh-s288m基因;将表达spamdh-e297n的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-e297n和pet24a-spamdh-e297n,该菌株和载体中spamdh基因的第889位由g突变为了a、第891位由a突变为了t,将突变后的spamdh基因记为spamdh-e297n基因;将表达spamdh-n69c/v291c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-n69c/v291c和pet24a-spamdh-n69c/v291c,该菌株和载体中spamdh基因的第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t,将突变后的spamdh基因记为spamdh-n69c/v291c基因;将表达spamdh-s68c/v291c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-s68c/v291c和pet24a-spamdh-s68c/v291c,该菌株和载体中spamdh基因的第202位由a突变为了t、第204位由t突变为了c、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t,将突变后的spamdh基因记为spamdh-s68c/v291c基因;将表达spamdh-l40c/i111f的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-l40c/i111f和pet24a-spamdh-l40c/i111f,该菌株和载体中spamdh基因的第118位由c突变为了t、第119位由t突变为了g、第120位由g突变为了c、第331位由a突变为了t,将突变后的spamdh基因记为spamdh-l40c/i111f基因;将表达spamdh-i111f/v294c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-i111f/v294c和pet24a-spamdh-i111f/v294c,该菌株和载体中spamdh基因的第331位由a突变为了t、第880位由g突变为了t、第881位由t突变为了g、第882位由t突变为了c,将突变后的spamdh基因记为spamdh-i111f/v294c基因;将表达spamdh-e114v/v291c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-e114v/v291c和pet24a-spamdh-e114v/v291c,该菌株和载体中spamdh基因的第341位由a突变为了t、第342位由a突变为了g、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了c,将突变后的spamdh基因记为spamdh-e114v/v291c基因;将表达spamdh-l40c/i111f/t134f的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-l40c/i111f/t134f和pet24a-spamdh-l40c/i111f/t134f,该菌株和载体中spamdh基因的第118位由c突变为了t、第119位由t突变为了g、第120位由g突变为了c、第331位由a突变为了t、第400位由a突变为了t、第401位由c突变为了t、第402位由c突变为了t,将突变后的spamdh基因记为spamdh-l40c/i111f/t134f基因;将表达spamdh-s68c/n69c/v291c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-s68c/n69c/v291c和pet24a-spamdh-s68c/n69c/v291c,该菌株和载体中spamdh基因的第202位由a突变为了t、第204位由t突变为了c、第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t,将突变后的spamdh基因记为spamdh-s68c/n69c/v291c基因;将表达spamdh-i111f/a113c/t134c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-i111f/a113c/t134c和pet24a-spamdh-i111f/a113c/t134c,该菌株和载体中spamdh基因的第331位由a突变为了t、第337位由g突变为了t、第338位由c突变为了g、第339位由a突变为了c、第400位由a突变为了t、第401位由c突变为了g,将突变后的spamdh基因记为spamdh-i111f/a113c/t134c基因;将表达spamdh-l40c/i111f/t134f/v294c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-l40c/i111f/t134f/v294c和pet24a-spamdh-l40c/i111f/t134f/v294c,该菌株和载体中spamdh基因的第118位由c突变为了t、第119位由t突变为了g、第120位由g突变为了c、第331位由a突变为了t、第400位由a突变为了t、第401位由c突变为了t、第402位由c突变为了t、第880位由g突变为了t、第881位由t突变为了g、第882位由t突变为了c,将突变后的spamdh基因记为spamdh-l40c/i111f/t134f/v294c基因;将表达spamdh-s68c/n69c/e114v/v291c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-s68c/n69c/e114v/v291c和pet24a-spamdh-s68c/n69c/e114v/v291c,该菌株和载体中spamdh基因的第202位由a突变为了t、第204位由t突变为了c、第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第341位由a突变为了t、第342位由a突变为了g、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t,将突变后的spamdh基因记为spamdh-s68c/n69c/e114v/v291c基因;将表达spamdh-s68c/n69c/i111f/v291c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-s68c/n69c/i111f/v291c和pet24a-spamdh-s68c/n69c/i111f/v291c,该菌株和载体中spamdh基因的第202位由a突变为了t、第204位由t突变为了c、第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第331位由a突变为了t、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t,将突变后的spamdh基因记为spamdh-s68c/n69c/i111f/v291c基因;将表达spamdh-i111f/e114v/v294c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-i111f/e114v/v294c和pet24a-spamdh-i111f/e114v/v294c,该菌株和载体中spamdh基因的第331位由a突变为了t、第341位由a突变为了t、第342位由a突变为了g、第880位由g突变为了t、第881位由t突变为了g、第882位由t突变为了c,将突变后的spamdh基因记为spamdh-i111f/e114v/v294c基因;将表达spamdh-l40f/s68c/n69c/t134c/v291c/v294c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-l40f/s68c/n69c/t134c/v291c/v294c和pet24a-spamdh-l40f/s68c/n69c/t134c/v291c/v294c,该菌株和载体中spamdh基因的第118位由c突变为了t、第120位由g突变为了t、第202位由a突变为了t、第204位由t突变为了c、第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第400位由a突变为了t、第401位由c突变为了g,第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t、第880位由g突变为了t、第881位由t突变为了g、第882位由t突变为了c,将突变后的spamdh基因记为spamdh-l40f/s68c/n69c/t134c/v291c/v294c基因;将表达spamdh-s68c/n69c/t134c/v291c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-s68c/n69c/t134c/v291c和pet24a-spamdh-s68c/n69c/t134c/v291c,该菌株和载体中spamdh基因的第202位由a突变为了t、第204位由t突变为了c、第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第400位由a突变为了t、第401位由c突变为了g、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t,将突变后的spamdh基因记为spamdh-s68c/n69c/t134c/v291c基因;将表达spamdh-l40f/s68c/n69c/a113c/t134c/v291c/v294f的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-l40f/s68c/n69c/a113c/t134c/v291c/v294f和pet24a-spamdh-l40f/s68c/n69c/a113c/t134c/v291c/v294f,该菌株和载体中spamdh基因的第118位由c突变为了t、第120位由g突变为了t、第202位由a突变为了t、第204位由t突变为了c、第205位由a突变为了t、第206位由a突变为了g、第207位由t突变为了c、第337位由g突变为了t、第338位由c突变为了g、第339位由a突变为了c、第400位由a突变为了t、第401位由c突变为了g、第871位由g突变为了t、第872位由t突变为了g、第873位由g突变为了t、第880位由g突变为了t,将突变后的spamdh基因记为spamdh-l40f/s68c/n69c/a113c/t134c/v291c/v294f基因;将表达spamdh-s68c/i111f/v294c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-s68c/i111f/v294c和pet24a-spamdh-s68c/i111f/v294c,该菌株和载体中spamdh基因的第202位由a突变为了t、第204位由t突变为了c、第331位由a突变为了t、第880位由g突变为了t、第881位由t突变为了g、第882位由t突变为了c,将突变后的spamdh基因记为spamdh-s68c/i111f/v294c基因;将表达spamdh-i111f/s288c/v294c的菌株和重组载体分别记为bl21(de3)/pet24a-spamdh-i111f/s288c/v294c和pet24a-spamdh-i111f/s288c/v294c,该菌株和载体中spamdh基因的第331位由a突变为了t、第862位由a突变为了t、第864位由t突变为了c、第880位由g突变为了t、第881位由t突变为了g、第882位由t突变为了c,将突变后的spamdh基因记为spamdh-i111f/s288c/v294c基因。实施例4、胺脱氢酶spamdh或其突变体催化4-羟基-2-丁酮生成(r)-3-氨基-1-丁醇用实施例2制备的胺脱氢酶spamdh或其突变体的全细胞催化4-羟基-2-丁酮生成(r)-3-氨基-1-丁醇,并利用bl21(de3)/pet24a作为对照。spamdh或其突变体的催化反应的反应体系通过向1mol/l的氯化铵/氨水缓冲液(ph8.5)中添加如下量的物质得到:底物4-羟基-2-丁酮20mmol/l、胺脱氢酶spamdh或其突变体的全细胞100g/l、nad+(以氧化型辅酶i水溶液形式存在)1mmol/l、实施例3的gdh粗酶粉2g/l,葡萄糖100mmol/l,溶菌酶1g/l,dnasei(脱氧核糖核酸酶)为6u/ml。将上述spamdh或其突变体的催化反应的反应体系于30℃反应12小时。反应结束后将所得反应液煮沸5分钟,12,000rpm离心10分钟,去除沉淀,保留上清,用邻苯二甲醛衍生(同实施例3)后进行hplc检测反应,hplc的梯度洗脱程序同表3。hplc检测结果如表5所示。转化率=a1/a2×100%;a1:液相色谱分析获得的(r)-3-氨基-1-丁醇峰面积值;a2:液相色谱分析获得的标准品(r)-3-氨基-1-丁醇的峰面积值。bl21(de3)/pet24a的反应产物中并无(r)-3-氨基-1-丁醇的生成,胺脱氢酶spamdh或其突变体催化还原4-羟基-2-丁酮生成(r)-3-氨基-1-丁醇反应的hplc检测结果图谱如图3所示,c:混旋3-氨基-1-丁醇标准品液相色谱结果;d:样品(r)-3-氨基-1-丁醇液相色谱结果。结果显示,胺脱氢酶spamdh或其突变体可以不对称还原催化4-羟基-2-丁酮生成(r)-3-氨基-1-丁醇,转化率为14-29%,立体选择性大于99%(r)。表5为胺脱氢酶spamdh及其突变体催化4-羟基-2-丁酮检测结果胺脱氢酶spamdh及其突变体转化率(%)ee(%)spamdh23>99(r)spamdh-i111f14>99(r)spamdh-v291c17>99(r)spamdh-i111f/v294c18>99(r)spamdh-e114v/v291c20>99(r)spamdh-i111f/e114v/v294c29>99(r)实施例5、胺脱氢酶spamdh或其突变体催化1-羟基-2-丁酮生成(s)-2-氨基-1-丁醇根据辅酶nadh在340nm的吸收值来检测酶活,摩尔吸光系数ε340=6.22x103(m-1cm-1)。酶活单位定义为:一分钟内转化产生或消耗1μmol/lnadh所需要的酶量。酶活计算公式:酶活(u)=ew×v×103/(6220×0.3),式中,ew表示每分钟内340nm处吸光值的变化,v表示反应液体积,单位ml,6220为辅酶的摩尔消光系数,单位l/(mol×cm),0.3为光程距离,单位cm。所用反应体系通过向1mol/l的氯化铵/氨水缓冲液(氯化铵和氨水等摩尔比混合得到,ph8.5)中添加如下量的物质得到:0.2mmol/lnadh,1-30mmol/l1-羟基-2-丁酮,适量实施例2的纯酶液。在30℃下反应,测nadh在5min内的吸光值变化,测定不同底物浓度下的反应初速率,然后用origin进行米氏方程非线性拟合。检测结果如表6所示。结果显示,胺脱氢酶spamdh或其突变体催化1-羟基-2-丁酮的km值为9.45-21.5mm,kcat值为0.83-6.07s-1,kcat/km值为0.088-0.346s-1mm-1。表6为胺脱氢酶spamdh及其突变体的米氏常数胺脱氢酶spamdh及其突变体km(mm)kcat(s-1)kcat/km(s-1mm-1)spamdh9.45±1.440.83±0.050.088spamdh-i111f18.52±1.421.74±0.070.094spamdh-v291c10.01±0.061.22±0.030.122spamdh-i111f/v294c21.5±3.373.14±0.270.146spamdh-e114v/v291c10.74±1.041.60±0.110.149spamdh-i111f/e114v/v294c17.53±1.116.07±0.190.346实施例6、胺脱氢酶spamdh或其突变体催化羟基酮底物用实施例2制备的胺脱氢酶spamdh或其突变体(spamdh-e114v/v291c、spamdh-i111f/e114v/v294c)催化羟基酮底物,分别为1-羟基-2-丁酮(1a)、4-羟基-2-丁酮(1b)、丙酮醇(1c)、2-羟基苯乙酮(1d)、1-(4-氟苯基)-2-羟基-1-乙酮(1e),对应生成的产物分别为(s)-2-氨基-1-丁醇(2a)、(r)-3-氨基-1-丁醇(2b)、(s)-丙氨醇(2c)、(s)-苯甘胺醇(2d)、(s)-2-氨基-2-(4-氟苯基)-1-乙醇(2e)。上述催化反应的反应体系通过向1mol/l的氯化铵/氨水缓冲液(ph8.5)中添加如下量的物质得到:底物20mmol/l、胺脱氢酶spamdh或其突变体(spamdh-e114v/v291c、spamdh-i111f/e114v/v294c)的全细胞100g/l、nad+(以氧化型辅酶i水溶液形式存在)1mmol/l、实施例3的gdh粗酶粉2g/l,葡萄糖100mmol/l,溶菌酶1g/l,dnasei为6u/ml。上述反应的条件为30℃反应12小时。检测催化底物1-羟基-2-丁酮(1a)、4-羟基-2-丁酮(1b)和丙酮醇(1c)生成的产物用邻苯二甲醛衍生后进行hplc检测反应,检测催化底物2-羟基苯乙酮(1d)和1-(4-氟苯基)-2-羟基-1-乙酮(1e)生成的产物用marfey’s试剂衍生后(衍生方法:取50μl上述反应液+50μl1mol/l氯化铵/氨水缓冲液(ph8.5)+500μl乙腈,然后过滤。取20μl上述过滤后的混合液+20μlmarfey’s试剂(14mmol/l,溶于乙腈)+36μlnahco3(1mol/l)+100μldmso混匀,40℃,1,000rpm反应2h后,加入40μlhcl(1mol/l)终止)进行hplc检测反应,hplc检测条件如表7所示。表7为hplc检测方法注:表7中,ahplc检测条件:agilentsb-aqc18柱(4.6*250mm,5μm),检测波长334nm,柱温:35℃,流速:1ml/min,上样量:10μl;流动相a:甲醇,流动相b:0.05mol/l乙酸钠;bhplc检测条件:zorbaxsb-c18柱(4.6*150mm,5μm),检测波长340nm,柱温:25℃,流速:0.45ml/min,上样量:10μl;流动相a:含0.1%(v/v)三氟乙酸的甲醇,流动相b:含0.1%(v/v)三氟乙酸的双蒸水;a标准品为混旋2-氨基-1-丁醇,玛雅试剂,货号:maya-cr-3330;b标准品为混旋3-氨基-1-丁醇,上海韶远试剂,货号:sy030187;c标准品为混旋丙氨醇,上海毕得医药科技有限公司,货号:bd70946;d标准品为混旋苯甘胺醇上海韶远试剂,货号:sy002083;e标准品为混旋2-氨基-2-(4-氟苯基)-1-乙醇,上海毕得医药科技有限公司,货号:bd177526。检测结果如表8和图4所示,可以看出,胺脱氢酶spamdh突变体(spamdh-i111f/e114v/v294c)对底物1-羟基-2-丁酮有很好的催化活性及对映体选择性之外,对其它羟基酮底物同样具有良好的催化活性。wt:spamdh;m1:突变体spamdh-e114v/v291c;m2:突变体spamdh-i111f/e114v/v294c。表8为胺脱氢酶spamdh及其突变体催化羟基酮底物检测结果注:表8中的转化率与ee,从上至下各行分别为spamdh、spamdh-e114v/v291c或spamdh-i111f/e114v/v294c催化羟基酮底物1-羟基-2-丁酮(1a)、4-羟基-2-丁酮(1b)、丙酮醇(1c)、2-羟基苯乙酮(1d)、1-(4-氟苯基)-2-羟基-1-乙酮(1e)的转化率和立体选择性。实施例7、胺脱氢酶spamdh或其突变体催化1-羟基-2-丁酮生成(s)-2-氨基-1-丁醇放大反应用实施例2制备的胺脱氢酶spamdh或其突变体(spamdh-e114v/v291c、spamdh-i111f/e114v/v294c)催化1-羟基-2-丁酮生成(s)-2-氨基-1-丁醇放大反应,将底物1-羟基-2-丁酮的浓度提高到200mmol/l。反应体系通过向1mol/l的氯化铵/氨水缓冲液(ph8.5)中添加如下量的物质得到:底物1-羟基-2-丁酮200mmol/l,全细胞100g/l,nad+(氧化型辅酶i水溶液的形式存在)1mmol/l,实施例3的gdh粗酶粉2g/l,葡萄糖100mmol/l,溶菌酶1g/l,dnasei(脱氧核糖核酸酶)为6u/ml。上述不对称还原反应的条件为30℃反应0.5h、1h、2h、4h、8h、12h、18h、24h。反应结束后将反应液煮沸5分钟,12,000rpm离心10分钟,去除沉淀,保留上清,用邻苯二甲醛衍生后进行hplc检测反应。检测结果图5所示,可以看出,胺脱氢酶spamdh突变体(spamdh-i111f/e114v/v294c)催化底物1-羟基-2-丁酮(200mmol/l)转化率高达91%,立体选择性大于99%(s)。以上对本发明进行了详述。对于本领域技术人员来说,在不脱离本发明的宗旨和范围,以及无需进行不必要的实验情况下,可在等同参数、浓度和条件下,在较宽范围内实施本发明。虽然本发明给出了特殊的实施例,应该理解为,可以对本发明作进一步的改进。总之,按本发明的原理,本申请欲包括任何变更、用途或对本发明的改进,包括脱离了本申请中已公开范围,而用本领域已知的常规技术进行的改变。按以下附带的权利要求的范围,可以进行一些基本特征的应用。<110>中国科学院天津工业生物技术研究所<120>胺脱氢酶突变体及其在手性胺醇化合物合成中的应用<160>3<170>patentinversion3.5<210>1<211>1095<212>dna<213>人工序列(artificialsequence)<400>1atggaaatcttcaagtacatggaacatcaggattatgaacagctggttatttgtcaggat60aaagccagtggtctgaaagcaattattgccattcatgataccaccctgggcccggccctg120ggtggtacacgtatgtggacctatgccagtgaagaagaagcaattgaagatgcactgcgc180ctggcacgtggtatgacctatagtaatgcagcagcaggcctgaatctgggtggcggcaaa240accgttattattggtaatccgaaaaccgataaaaacgatgaaatgtttcgtgcatttggt300cgctatattgaaggtctgaatggtcgttatattaccgcagaagatgttggcaccaccgaa360gcagatatggatctgattaatctggaaaccgattatgttaccggcaccagcgcaggtgca420ggtagtagtggcaatccgagcccggtgaccgcatacggtatctattatggcatgaaagcc480gccgccaaagaagcctttggcgatgatagcctggcaggtaaaaccgttgccgttcagggc540gttggtaatgttgcctatgccctgtgcgaatatctgcatgaagaaggcgcaaaactgatt600attaccgatattaatgaggaggccgtgcagcgcgccgtggatgcttttggcgccaccgca660gttggtattaatgaaatctatagtcaggaagccgatatttttgccccgtgcgcactgggt720gcaattattaatgatgaaaccattccgcagctgaaagccaaagttattgccggtagcgca780ctgaatcagctgaaagaaacccgccacggtgacctgattcatgaaatgggcattgtgtat840gcaccggattatgttattaatagtggcggtgtgattaatgttgccgatgaactggatggc900tataatcgtgaacgcgcactgaaacgtgtggaaggcatctatgatgttattggcaaaatt960tttgcgatcagcaaacgtgataatattccgacctatgttgcagccgatcgcatggccgaa1020gaacgtattgcacgcgtggcaaatacccgcagtacctttctgcagaatgaaaaaagtgtg1080ctgagccgtcgttaa1095<210>2<211>364<212>prt<213>嗜冷芽孢八叠球菌(sporosarcinapsychrophila)<400>2metgluilephelystyrmetgluhisglnasptyrgluglnleuval151015ilecysglnasplysalaserglyleulysalaileilealailehis202530aspthrthrleuglyproalaleuglyglythrargmettrpthrtyr354045alaserglugluglualailegluaspalaleuargleualaarggly505560metthrtyrserasnalaalaalaglyleuasnleuglyglyglylys65707580thrvalileileglyasnprolysthrasplysasnaspglumetphe859095argalapheglyargtyrilegluglyleuasnglyargtyrilethr100105110alagluaspvalglythrthrglualaaspmetaspleuileasnleu115120125gluthrasptyrvalthrglythrseralaglyalaglysersergly130135140asnproserprovalthralatyrglyiletyrtyrglymetlysala145150155160alaalalysglualapheglyaspaspserleualaglylysthrval165170175alavalglnglyvalglyasnvalalatyralaleucysglutyrleu180185190hisglugluglyalalysleuileilethraspileasnglugluala195200205valglnargalavalaspalapheglyalathralavalglyileasn210215220gluiletyrserglnglualaaspilephealaprocysalaleugly225230235240alaileileasnaspgluthrileproglnleulysalalysvalile245250255alaglyseralaleuasnglnleulysgluthrarghisglyaspleu260265270ilehisglumetglyilevaltyralaproasptyrvalileasnser275280285glyglyvalileasnvalalaaspgluleuaspglytyrasnargglu290295300argalaleulysargvalgluglyiletyraspvalileglylysile305310315320phealaileserlysargaspasnileprothrtyrvalalaalaasp325330335argmetalaglugluargilealaargvalalaasnthrargserthr340345350pheleuglnasnglulysservalleuserargarg355360<210>3<211>786<212>dna<213>人工序列(artificialsequence)<400>3atgtatccggatttaaaaggaaaagtcgtcgctattacaggagctgcttcagggctcgga60aaggcgatggccattcgcttcggcaaggagcaggcaaaagtggttatcaactattatagt120aataaacaagatccgaacgaggtaaaagaagaggtcatcaaggcgggcggtgaagctgtt180gtcgtccaaggagatgtcacgaaagaggaagatgtaaaaaatatcgtgcaaacggcaatt240aaggagttcggcacactcgatattatgattaataatgccggtcttgaaaatcctgtgcca300tctcacgaaatgccgctcaaggattgggataaagtcatcggcacgaacttaacgggtgcc360tttttaggaagccgtgaagcgattaaatatttcgtagaaaacgatatcaagggaaatgtc420attaacatgtccagtgtgcacgaagtgattccttggccattatttgtccactatgcggca480agtaaaggcgggatgaagctgatgacagaaacattagcgttggaatacgcgccgaagggc540attcgcgtcaataatattgggccaggtgcgatcaacacgacgatcaataaggagaaattt600gctgaccctgaacagagagctgatgtagaaagcatgattccaatgggatatatcggcgaa660ccggaggagatcgccgcagtagcagcctggcttgcttcgaaggaagccagctacgtcaca720ggcatcacgttattcgcggacggcggtatgacacaatatccttcattccaggcaggccgc780ggttaa786当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1