一种用于骨干载体的RNAi表达框及相应载体和应用
本发明属于生物,具体涉及提高引导编辑系统的编辑效率并且提供纯合品种的表达框、载体及其在基因组编辑中的应用。
背景技术:
1、2019年david liu课题组利用cas9-h840a蛋白融合逆转录酶mmlv,通过在单链引导rna的3’端引入引物结合位点pbs(prime binding sites)和逆转录模板rt(reversetemplate)实现突变的精准获得,其中pbs识别非靶标链并结合,rt上携带预设突变进行逆转录,经dna断裂后修复将突变精准的整合到基因组上。融合了pbs和rt的单链引导rna命名为pegrna。引导编辑系统具有更精准和可操作性强等优势,但是引导编辑的效率显著低于敲除和碱基编辑系统。随后,科学家们致力于引导编辑系统效率的优化工作:从稳定和增强pegrna表达角度出发,目前已知的策略主要有:1.pegrna的3’端融合motif基团稳定pegrna的结构和减少3’端pbs和rt序列被降解;2.使用双pegrna系统提高引导编辑效率,同时兼顾了pbs的退火温度等。3.通过使用复合型启动子增强pegrna的表达。4.在rt模板序列上置入无义的碱基突变,减少与野生型序列互补配对的干扰,从而稳定pegrna的结构。而优化另一重要结构域的策略也有许多报道,通过将ncas9蛋白点突变为ncas9-r221k,n394k稳定cas9-mmlv的复合体结构;同样的可以采取使用多肽来稳定结构和表达。
2、但是现有策略要么引导编辑效率偏低,难以应用,要么无法提供纯合的植物品种。
技术实现思路
1、针对上述问题,本技术的发明人在通过大量实验研究之后,研发出了一种用于骨干载体的rnai表达框,其可以抑制水稻本身mlh1基因的表达量,并且通过与另一表达框的配合能够实现自切除。动植物的机体为了抵御突变的引入,会启动错配修复途径进行修复。而mlh1基因是错配修复途径的关键因子,本技术在突变引入的关键时刻将该基因干扰,使得错配修复无法工作,有利于突变的引入,进而提升突变效率。本发明提供的表达框及相应载体既能够通过抑制水稻本身mlh1基因的表达量提高植物引导编辑系统的精确编辑效率,同时当实现这一目的后为了不影响mlh1抑制后对表型的影响,将mlh1干扰的表达框通过干旱诱导的cre-loxp系统进行切除,进而避免抑制mlh1之后带来植株生长问题,进而有利于纯合突变体的获得。
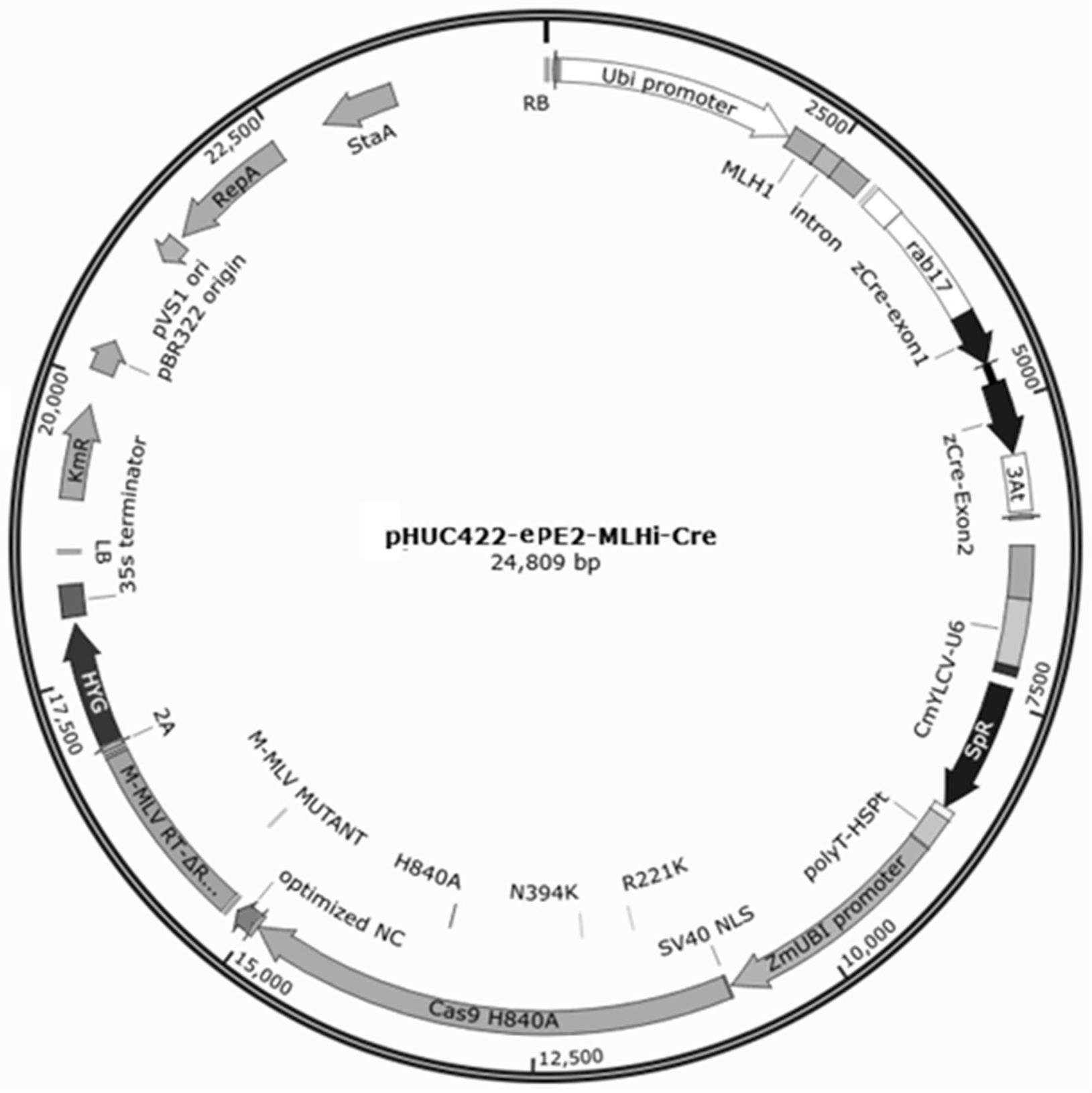
2、具体而言,本发明提供一种用于骨干载体的rnai表达框,所述表达框包含启动子、mlh基因互补序列、nos终止子序列,其中,所述启动子序列如序列表中seq id no.1的第59-2048位所示,所述mlh基因互补序列如序列表中seq id no.1序列表1的2058-2788位所示,所述nos终止子序列如序列表中seq idno.1的2902-3154位所示。
3、一种用于切除上述表达框的重组酶表达框,所述表达框包含干旱诱导的rab17启动子序列、cre重组酶和终止子序列,所述rab17干旱诱导启动子如序列表中seq id no.1的第3160-4137位所示,cre重组酶如序列表中seq id no.1的第4140-5365位所示,3at终止子如序列表中seq id no.1的第5380-5849位所示。
4、进一步地,所述重组酶表达框的特异识别位点包含方向一致的重组酶特异识别位点loxp,二者分别位于序列表1的1-34位和5879-5912位。
5、另一方面,本发明提供一种植物骨干载体,所述植物骨干载体包括权利要求1所述的抑制mlh基因表达的表达框和权利要求2所述的重组酶表达框。
6、进一步地,所述植物骨干载体包含识别并引入突变的pegrna表达框和用于cas9蛋白表达和反转录酶表达的pe蛋白复合体。
7、关于骨干载体需要说明的是,其长度为17562,由于系统原因,超过1万bp的序列无法上传,申请人在序列表中将其拆分成了9000bp的序列表2和8562bp的序列表3,但是为了表述方便,下述的骨干载体序列均指代的是将序列表中的序列表2和序列表3顺序连接所形成的17562bp的骨干载体。
8、进一步地,所述pegrna表达框包含35s-cmyl-u6复合型启动子,spr抗性基因,hdv-polyt核酶+终止子,所述pegrna表达框位于序列表骨干载体序列的第6263-8901位,所述壮观霉素抗性基因spr用于替换为靶向目的dna片段的sgrna、sgrna骨架序列、逆转录模板rt和引物结合位点pbs、8bp linker序列。
9、进一步地,所述融合蛋白表达框包括zmubi启动子、经改造后的cas9切刻酶或其变体编码序列、逆转录酶m-mlv rt编码序列和35s终止子,所述表达框位于序列表骨干载体序列的第9008-17279位
10、另一方面,本发明提供一种利用权所述的骨干载体构建靶向载体的方法,所述方法包括:
11、按照目的基因的编码序列和突变类型,选择sgrna序列,得到相应的逆转录模板rt和引物结合位点pbs序列,用bsai内切酶切开权利要求3所述的骨干载体,利用含bsai的golden gate反应,将sgrna序列、sgrna骨架序列、rt和pbs序列、8bp linker序列替换壮观霉素抗性基因,形成用于作物目的基因的引导编辑重组载体。
12、另一方面,本发明提供一种所述表达框、所述载体的应用,所述应用包括将所述表达框或所述重组载体转入植物细胞,使细胞同时含有针对靶标基因的pegrna和融合蛋白;并对生物体的基因组进行编辑,获得生物突变体,所述基因组序列的编辑为基因组序列的碱基替换、缺失和插入。。
13、rab17启动子序列受干旱特异诱导,当干旱处理时,诱导下游基因高效表达,诱导表达的基因为cre重组酶,通过识别同向的loxp识别位点并切割loxp之间的序列,同样包含3at终止子序列。
14、优选地,一种用于切除基因抑制表达框切除的重组酶特异识别位点包含方向一致的重组酶特异识别位点loxp,分别位于序列表1的1-34位和5879-5912位。可以供cre重组酶特异识别并高效切割。
15、此外该骨干载体包含识别并引入突变的pegrna表达框和用于cas9蛋白表达和反转录酶表达的pe蛋白复合体。
16、另一方面,本发明提供一种高效的植物引导编辑系统enppe。所述植物引导编辑系统含有rnai表达框、重组酶表达框、pegrna以及pe蛋白复合体,所述pegrna为由靶向目的dna片段的sgrna、逆转录模板(rt)和引物结合位点(pbs)连接得到的rna分子。此外,在pegrna的3’增加evopreq1,避免pegrna的3’被rna酶降解,其次使用35s-cmylcv-u6复合启动子驱动pegrna的表达,以增强pegrna的表达。
17、所述植物引导编辑系统还含有cas9切口酶(h840a)或变体与逆转录酶mmlv融合所形成的融合蛋白。通过使用cas9切口酶变体(h840a/r221k/n394k)以及不同类型的核定位信号nls,产生优化的融合蛋白结构,提高其作用活性。
18、在本发明中,spr基因两端分别存在反向排列的bsai内切酶识别位点(剪切位点如骨干载体序列第7335位和8548位所示),用于插入目的基因sgrna、sgrna骨架、相应的逆转录模板(rt)和引物结合位点(pbs)片段。
19、所述骨干质粒载体还包括t-dna的左、右边界序列,其中所述左边界的核苷酸序列如seq id no.1第17537至17562位所示,所述右边界的核苷酸序列如seq id no.1第1至26位所示;所述pegrna表达框和所述融合蛋白表达框位于所述左边界和所述右边界之间。
20、所述骨干质粒载体还可含有抗性标记基因。
21、本发明还提供了一种构建用于引导编辑的重组载体的方法,该方法包括:
22、按照目的基因的编码序列和突变类型,选择sgrna序列,得到相应的逆转录模板(rt)和引物结合位点(pbs)序列。用bsai内切酶切开本发明提供的骨干质粒载体,利用含bsai的golden gate反应,将sgrna序列、sgrna骨架序列、rt和pbs序列、8bp linker序列替换壮观霉素抗性基因,形成用于作物目的基因的引导编辑重组载体。
23、另一方面,本发明提供一种宿主菌,所述宿主菌包含上述的重组载体。
24、本发明的骨干质粒载体中,pegrna表达框和融合蛋白表达框及osmlhrnai表达框及cre重组酶表达框位于同一双元载体中,pegrna表达框依次由35s-cmylcv-u6复合启动子、trna基因序列、壮观霉素抗性基因spr、rna核酶hdv序列、eq序列和polyt-hspt复合终止子组成;cas9核酸酶表达框由zmubi启动子,融合蛋白编码序列和35s终止子依次构成。rnai干扰表达框主要由ubi启动子,osmlh1基因cds区200bp左右互补配对片段,中间由启动子串联,nos终止子序列组成。cre重组酶表达框由rab17干旱诱导启动子,cre重组酶和3at终止子组成。此外将重组酶识别切割位点loxp分别放置在ubi启动子前端和3at启动子后端。
25、本发明还提供了一种引导编辑系统在作物基因打靶中的应用,将所述重组载体应用于生物突变体获得,具体而言,将上面获得的重组载体转入植物细胞,比如,通过农杆菌介导法,使细胞同时含有针对靶标基因的pegrna和融合蛋白;并对生物体的基因组进行编辑,获得生物突变体。
26、上述的应用或方法中,基因组序列的编辑包括基因组序列的碱基替换(如单碱基替换和多碱基替换)、碱基插入(如单碱基插入和多碱基插入)和碱基删除(如单碱基删除和多碱基删除)。在本发明的具体实施例中,所述基因组序列的编辑为基因组序列的单个碱基替换、插入或缺失。
27、本发明通过特定时间内干扰错配修复途径中mlh1基因的表达进一步地提高水稻引导编辑系统的效率,并在达到这一目的后对干扰系统进行切除,减少由于mlh1表达的抑制可能造成的植物其他性状变化,具体而言,通过重组酶表达框控制,当pe的引入突变完成后,重组酶表达框是由干旱诱导启动子驱动的,将水稻愈伤干旱处理,诱导启动子驱动cre重组酶表达,cre酶切整个mlh1rnai表达框,停止干扰体内mlh1基因表达。利用本发明的引导编辑系统不仅可以有效地提高植物中引导编辑系统的效率,还能提高纯合突变的效率,具有很好的应用前景。
- 还没有人留言评论。精彩留言会获得点赞!