ShizukaolD在制备抗肿瘤靶点药物中的应用的制作方法
本发明属于医药和基因工程领域,具体而言,本发明涉及中药单体shizukaold在制备抗肿瘤靶点药物中的应用及其应用方法。
背景技术:
由于生活习惯的改变和环境污染的加剧,世界各国癌症发病率在不断上升。中国从上世纪70年代至今,癌症发病率已翻倍。全国2010年新发恶性肿瘤病例逾三百万例,死亡病例近两百万。对于国家而言,肿瘤的预防和治疗消耗了大量的人力、物力和财力。对于个人而言,肿瘤的危害在于:阻塞和压迫,如食管癌癌肿可以堵塞食管,造成病人吞咽困难;破坏所在器官的结构和功能;侵袭破坏邻近器官;坏死、出血、感染;疼痛;发热;恶病质,即指机体严重消瘦、无力、贫血和全身衰竭的状态,它是癌症病人死亡的重要原因。
开发全新来源的抗癌药物成为国内外药物学界的重要课题。全球约480亿美元的抗癌药物中,植物来源的抗癌药约占其中40%。与化学合成的抗癌药相比,植物抗癌药的毒副作用要小得多,植物抗癌药现已成为抗癌新药的重要来源。已经有一些中草药成分成功用于治疗肿瘤的实例。青蒿素的衍生物青蒿酯钠可诱导人肝癌细胞bel-7402凋亡。紫杉醇是从紫衫中提取的天然抗癌药物,其衍生物多西紫杉醇对肿瘤细胞的抑制过程中,存在氧化应急以及氧化—抗氧化失衡。苦参碱是从苦参中提取的生物碱,苦参碱和氧化苦参碱能显著抑制肝癌细胞增殖,促进其凋亡,并呈时间与剂量依赖性,其机制可能与下调stat3、stat5的基因表达,抑制细胞信号转导通路有关。随着对天然产物抗肿瘤活性的研究报道增加,中草药中具有抑制肿瘤细胞增殖的活性物质正逐步被解析。与传统的手术、放疗、化疗相比,中药具有多成分、多环节、多靶点效应的优势,可以通过作用于肿瘤的信号转导通路诱导肿瘤细胞的凋亡、减少肿瘤细胞的黏附、影响肿瘤血管生成。
shizukaold最早由日本学者川端(kawabata)从金粟兰科植物宽叶金粟兰(kawabataj,mizutanij.dimericsesquiterpenoidestersfromchloranthusserratus.[j].phytochemistry,1992,31(4):1293-6.)和银线草(kawabataj.sesquiterpenedimersfromchloranthusjaponicus[j].phytochemistry,1995,39,121-7.)中提取。金粟兰科植物主要分布在我国北方和日本、朝鲜等地(yimnh,hwangei,yunbs,etal.sesquiterpenefurancompoundcj-01,anovelchitinsynthase2inhibitorfromchloranthusjaponicussieb[j].biolpharmbull,2008,31(5):1041-4.),家喻户晓的代表性植物就是银线草。银线草全株供药用,能祛湿散寒、活血止痛、散瘀解毒,主治风寒咳嗽、风湿痛、闭经,外用治跌打损伤、瘀血肿痛、毒蛇咬伤等。
shizukaold是一种存在于金粟兰科植物的、典型的倍半萜二聚体;常温下为淡黄色粉末,极性溶液中溶解性差,可溶于dmso、氯仿等有机溶剂,避光保存;分子式:c33h38o9;分子量:578.65;cas号:142279-42-3。其活性报道较少。2009年10月13日申请的中国发明专利《乌药烷型二聚倍半萜类化合物,其制备方法及其在制药中的应用》(申请号为200910095051.6)中描述:shizukaold明显增加l6细胞葡萄糖的消耗量,提示能够促进胰岛素的作用,促进糖的摄取和转运。zhang等曾在2012年报道其抗炎症方面的作用(zhangm,imunekazu,wangjun-song,etal.terpenoidsfromchloranthusserratusandtheiranti-inflammatoryactivities[j].journalofnaturalproducts,2012,75(4):694-8.)。直到2014年1月,hu等在《plosone》杂志上发表了shizukaold通过诱导线粒体功能失调抑制肝细胞中脂内容物的报道(hur,yanh,haox,etal.shizukaoldisolatedfromchloranthusjaponicasinhibitsampk-dependentlipidcontentinhepaticcellsbyinducingmitochondrialdysfunction[j].plosone,2013,8(8):e73527.)。
但是,到目前为止,尚无关于shizukaold靶向抑制细胞信号通路方面的报道。
技术实现要素:
本发明要解决的技术问题是提供shizukaold的新用途。shizukaol是一种存在于金粟兰科植物的、典型的倍半萜二聚体,其功能活性报道集中在抗炎症方面。本发明提供了shizukaold在制备抗肿瘤靶点药物中的应用。
近年来,肿瘤信号通路成为新的研究热点。细胞通路的激活或抑制与肿瘤的发生、发展、消退等存在着紧密的联系。肝癌的发生与发展是癌细胞增殖旺盛与凋亡减少的结果,其中,信号通路在其中扮演着重要角色。众所周知的鸡尾酒法中的三氧化二砷,能够通过活化jnk信号转导通路激活caspase3,启动hepg2的凋亡。据统计,在三分之一的肝细胞癌患者中wnt通路呈激活状态。很多关于肿瘤的研究都证实,阻断wnt经典信号通路可以诱导相关病变细胞凋亡。例如,il-α2b可以抑制癌前病变的肝组织中的wnt/β-catenin信号通路,并能促进细胞发生凋亡。wnt信号通路抑制参与了姜黄素诱导的人端粒逆转酶引起的肝星状细胞凋亡。
wnt通路主要分为经典wnt通路和非经典wnt通路。前者的研究较多。经典wnt通路即wnt/β-catenin通路,通过β-catenin介导。正常体细胞中β-catenin大部分结合在细胞膜上,少部分与axin(轴蛋白)、apc(adenomatouspolyposiscoli,腺瘤性结肠息肉病蛋白)、ck(酪蛋白激酶)、gsk-3β(糖原合成酶-3β)结合形成降解复合物,并被其磷酸化,然后通过泛素蛋白酶体途径迅速降解,从而维持胞浆内游离β-catenin的低水平。当胞外wnt蛋白与fzds(卷曲蛋白)受体以及lrp5/6(低密度脂蛋白相关受体)复合物结合后,激活了细胞内的dsh(散乱蛋白)、抑制gsk-3β的活性,β-catenin从复合物中解离,其降解受到抑制并在细胞质中积累。β-catenin积累到一定程度进入细胞核,并与核内转录因子tcf/lef结合形成转录复合物,激活相关目的基因的表达,wnt通路被激活并引起细胞生物学活性的变化。
第二代测序(ngs,nextgenerationsequencing)表明,wnt通路是一个重要的致癌因素,wnt信号通路失调则可能导致肿瘤的发生、发展。shizukaold能够降低wnt通路信号,抑制β-catenin的表达,这可能是导致肿瘤细胞增殖受到抑制的重要因素。
一方面,本发明提供了一种抑制体外肿瘤细胞wnt信号通路的方法,在细胞培养液中加入shizukaold;其中,shizukaold的终浓度范围是0-25微摩尔/升且不包含0或者25微摩尔/升。较好的,所述药剂的有效浓度范围是0-12.5微摩尔/升,例如,不大于6.25微摩尔/升。
本发明提供了shizukaold在制备wnt通路抑制剂中的应用。
本发明中,所述的shizukaold常温下为淡黄色粉末,极性溶液中溶解性差,可溶于dmso、氯仿等有机溶剂,避光保存;分子式:c33h38o9;分子量:578.65;cas号:142279-42-3。
本发明中,所述的wnt通路通常是经典wnt通路。
本发明中,所述的wnt通路抑制剂是靶向β-catenin的抑制剂。
所述的细胞是肝源细胞或者肾源细胞。
所述的抑制wnt信号通路是靶向抑制β-catenin的表达水平。
本发明还提供了一种测试wnt通路活性的试剂盒,所述的试剂盒中含有中药单体shizukaold。
所述的试剂盒中含有制作测定wnt通路信号强度的标准曲线的试剂。
另一方面,本发明提供了一种抑制wnt通路的药物组合物,所述的抑制wnt通路的药物组合物中的活性成分是shizukaold。
所述的药物组合物是粉剂、软膏剂、气雾剂、片剂或者液体制剂。所述的药物组合物含有药剂学上常用的辅料。
所述的药物组合物含有抗肿瘤药物。
所述的抗肿瘤药物包括目前临床常规的化疗药物,如烷化剂、抗代谢药、抗癌抗生素、植物类、激素类和杂类等。其中,烷化剂有:尼莫司汀、卡莫司汀、洛莫司汀、环磷酰胺、异环磷酰胺、甘磷酰芥等;抗代谢药有:去氧氟鸟苷、多西氟鸟啶、5-氟尿嘧啶、巯嘌呤、硫鸟嘌呤、阿糖胞苷、氟鸟苷、替加氟、吉西他滨、卡莫氟、羟基脲、甲氨蝶呤、优福定、安西他滨等;抗肿瘤抗生素有:放线菌素d、多柔比星、柔红霉素、表柔比星、丝裂霉素、培洛霉素、平阳霉素、吡柔比星等抗肿瘤动植物成分药有:伊立替康、三尖杉酯碱、羟基喜树碱、长春瑞宾、紫杉醇、泰索帝、拓扑替康、长春新碱、长春地辛、长春酰胺、长春碱、替尼泊苷、依托泊苷等;抗肿瘤激素类有:阿他美坦、阿那曲唑、氨鲁米特、来曲唑、福美坦、甲他孕酮、他莫昔芬等;杂类药有:门冬酰胺酶、卡铂、顺铂、达卡巴嗪、奥沙利铂、乐沙定、可铂奥沙、米托蒽醌、丙卡巴肼等。
本发明提供了中药单体shizukaold在制备抗肿瘤靶点药物中的应用,尤其是shizukaold作为wnt通路抑制剂方面的应用。实验表明,shizukaold能够抑制top/fop荧光素酶活性。加入shizukaold处理的细胞样品,wnt通路关键基因及下游基因表达水平明显下降,激活wnt通路能够补偿shizukaold抑制细胞增殖的作用。尤其是加入shizukaold处理的细胞样品中,wnt通路核心效应物β-catenin的表达水平显著下降。这些表明,shizukaold能够抑制wnt通路。本发明wnt通路抑制剂提供了新的先导化合物,为抑制细胞的wnt通路提供新的方法和途径,为中药单体shizukaold提供新的应用,提升了其植物来源——银线草的药用价值。
附图说明
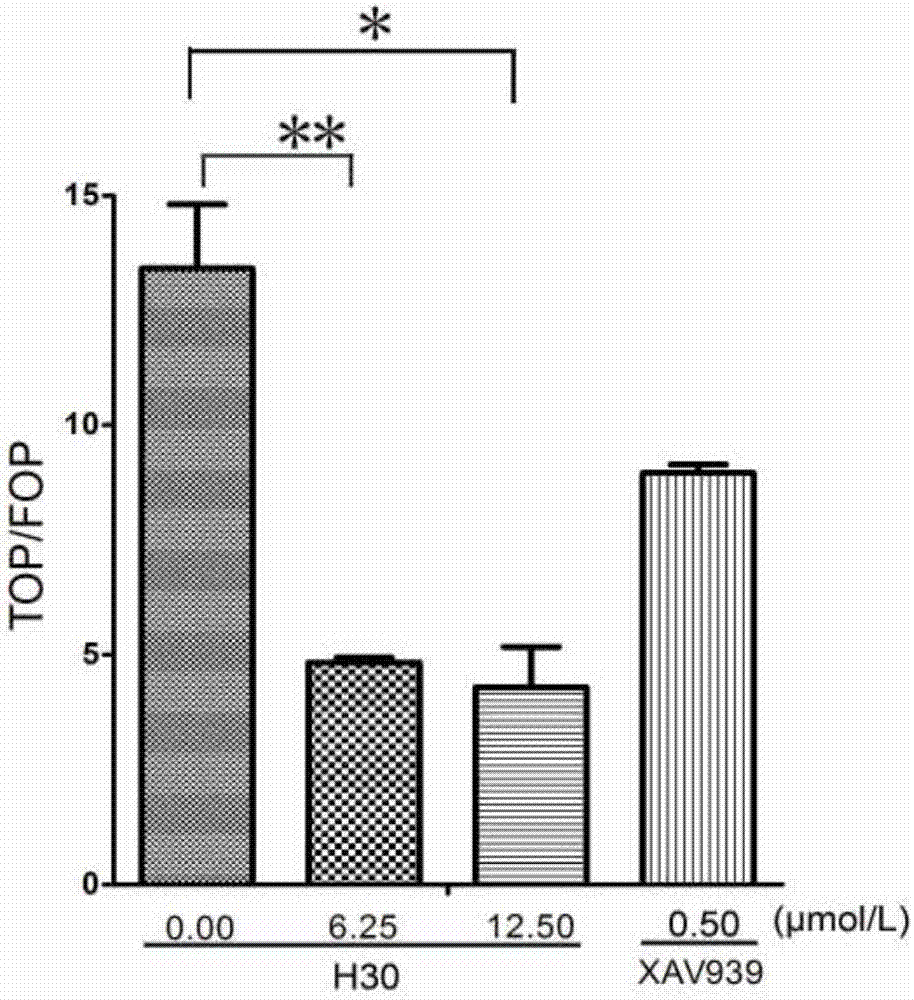
图1是shizukaold对top/fopflash荧光素酶活性的影响。纵坐标为top和fop的比值。横坐标为化合物的浓度。*表示0.01<p<0.05;**表示p<0.01。xav939是tankryase酶抑制剂,可以间接抑制wnt通路。xav-939通过抑制tankyrase1/2而选择性抑制wnt/β-catenin介导的转录,ic50为11nm/4nm。
图2是qpcr检测wnt通路相关蛋白表达水平。smmc-7721细胞贴壁后,加入shizukaold,分别在0、6和12小时收样,trizol法提取rna,去基因组rna,反转录后,通过实时pcr检测样品中基因表达量。以gapdh作为内参。**:p<0.01。
图3是wnt3a提高shizukaold作用下肝癌细胞的存活率。取smmc-7721细胞,贴壁后加入含有wnt3a的培养基,孵育过夜。加入6.25和12.50μmol/l的shizukaold,作用48小时后利用cck-8试剂盒检测细胞存活率。以0.1%的dmso为对照。
图4是外转β-catenin消减shizukaold对肝癌细胞增殖的抑制。取smmc-7721细胞,种24孔板(每孔约5×104个细胞)。贴壁后,转入野生型β-catenin、突变型β-catenin及pcdna3.0空载体质粒。隔天加入10μmol/l的shizukaold,以0.1%dmso作为药物对照,处理24小时,每个样做3个复孔。利用cck-8实验检测smmc-7721细胞的吸光值以表征存活率。**:p<0.01。
图5是shizukaold降低肝癌细胞中β-catenin的表达量。取肝癌细胞种于24孔板(3-6×104个/孔),待贴壁后加入6.25μmol/l、12.5μmol/l、25.0μmol/l的shizukaold,以0.1%dmso作为对照。化合物孵育48小时后,裂解细胞,配平总蛋白,sds-page电泳检测β-catenin的蛋白量。a和b:focus细胞,c和d:smmc-7721细胞。
具体实施方式
westernblotting(免疫印迹):
1)细胞处理方法与流式细胞术检测细胞周期相同。收细胞:胰酶消化细胞,含10%胎牛血清的dmem高糖培养基终止消化。室温离心1000r/min10分钟回收细胞。用0.5ml1×pbs清洗细胞两次,弃上清。
2)向沉淀中加入1ml裂解缓冲液,于4℃置于摇床,震荡30分钟。
3)将细胞裂解液于4℃12000r/min离心30分钟,收集上清。
4)10μl细胞裂解液上清与等量2×sds上样液混合后进行聚丙烯酰胺凝胶电泳。
5)按《分子克隆实验操作指南》方法,将蛋白从聚丙烯酰胺凝胶转至硝酸纤维素薄膜,转移时,恒流190ma,冰浴,2h。
6)剪下蛋白标准分子量条带,置于丽春红染色液中5~10分钟,置于水中洗去染料,显出蛋白标准分子量条带,并用铅笔进行标记。
7)将膜放置于20ml封闭缓冲液中,室温下置于摇床震荡1h。
8)一抗用封闭缓冲液稀释至合适浓度,室温下置于摇床震荡2h或4℃轻摇震荡过夜。
9)用洗涤缓冲液洗膜三次,每次10分钟,按适当比例将二抗稀释于封闭缓冲液,室温下置于摇床震荡1h。
10)洗涤缓冲液洗洗膜四次,每次10分钟。
11)1×tbs浸泡数秒,ecl法化学发光。
12)暗室显影、定影。
rna抽提:
准备工作:用depc(1ml/l)水溶液浸泡枪头、离心管等24h,然后高温灭菌2次,每次20分钟。
1)弃细胞培养基,pbs洗3次,每次10分钟。按照6孔板每个孔加入1ml的比例trizol,反复抽打均匀,转移到1.5ml离心管中,室温下放置5分钟。
2)加200μl氯仿,剧烈振荡15秒,颠倒几次,室温下放置15分钟。
3)4℃,12,000r/min离心15分钟。
4)小心转移400μl上清液至一新的离心管中,加400μl异丙醇,振荡混匀,室温下放置5~10分钟。
5)4℃,12,000r/min离心8分钟。
6)弃上清液,加1ml75%乙醇,漩涡混匀。
7)4℃,7,500r/min离心5分钟。
8)弃乙醇,空气风干3~5分钟,用depc处理过的水溶解,根据沉淀的量,一般加入30~50μl。
9)rna质量的检测,取1μlrna样品测定od值,od260/od280的比值大于1.8,说明制备的rna较纯,无蛋白质污染;取rna样品1μl,1%琼脂糖凝胶电泳80v电泳20分钟,用凝胶成像系统观察并拍照,总rna的5srrna,18srrna和28srrna条带,三条条带完整即可证明总rna抽提成功。
10)rna保存于-80℃冰箱。
rna去基因组:
使用rnase-free的dnasei(promega),按以下体系配置反应液,37℃消化30分钟,65℃灭活10分钟。
然后按以下步骤操作:
1)加入等体积的苯酚/氯仿,上下颠倒混匀,室温放置5分钟,14,000r/min离心15分钟,取上清。
2)加入等体积的氯仿,上下颠倒混匀,静置分层,14,000r/min离心15分钟,取上清。
3)加入等体积异丙醇,轻柔地充分混匀(颠倒6-8次),-20℃静置15分钟。
4)4℃下,14,000g离心15分钟,收集rna沉淀,去上清。
5)用75%乙醇洗涤两次(12,000g离心5分钟),超净台风干。
6)加入适量depc水(至少15μl)溶解沉淀。
rna反转录:
1)在离心管中配制下列混合液。
2)在pcr仪上进行变性、退火反应,65℃5分钟,4℃。
3)离心数秒钟,反应液聚集于离心管底中。
4)在上述离心管中配制下列反转录反应液。
5)在pcr仪上按下列条件进行反转录反应
42℃,15-30分钟;95℃,5分钟;4℃。
q-pcr实时定量pcr:
1)cdna用灭菌纯水稀释适当的浓度,一般为1:20稀释,如遇到基因表达低的样品,则适当降低稀释比例至1:10或1:5。
2)按照如下比例配制反应体系:
3)把384孔放在离心机上离心数秒,使反应液聚集在离心管底部。
4)上机扩增。
细胞免疫荧光定位:
1)在6孔板中每孔放入一片已经灭菌的盖玻片,按照1~2×105度均匀种植细胞。
2)待细胞贴壁后,加入刺激。
3)弃原培养基,pbs洗5分钟×3次,加入4%多聚甲醛固定30分钟。
4)pbs洗5分钟×3次,振荡洗涤制作好的细胞爬片,加入0.2%triton-x100打孔,室温静置15分钟。
5)pbs洗5分钟×3次,振荡洗涤,封闭液覆盖标本表面37℃孵育1h。
6)加入合适稀释浓度的一抗,4℃孵育过夜。
7)pbs洗5分钟×3次,振荡洗涤,加入fitc标记的二抗,37℃避光孵育1h。
8)pbs洗5分钟×3次,振荡洗涤,加入dapi,避光静置10分钟。
9)pbs洗5分钟×3次,避光、振荡洗涤。
10)在干净的载玻片上滴一滴封片剂,将爬片的细胞面扣在封片剂上。
11)镜检,拍照。
实施例1wnt通路的信号受到抑制
取293t细胞,种24孔板(每孔4~6×104个细胞)。贴壁后,用50mmol/l的licl处理12小时,然后转入topflash、fopflash及内参rl质粒。隔天加入shizukaold和dmso对照(即加入0.1%dmso作为对照)处理24小时,每个样做3个复孔。收样,检测荧光素酶活性。以xav939为阳性对照。xav939是tankryase酶抑制剂,可以间接抑制wnt通路(huangsm,mishinaym,lius,etal.tankyraseinhibitionstabilizesaxinandantagonizeswntsignalling[j].nature,2009,461(7264):614-20.)。xav-939通过抑制tankyrase1/2而选择性抑制wnt/β-catenin介导的转录,ic50为11nm/4nm,调节轴蛋白水平,但对cre,nf-κb和tgf-β无作用。
如图1所示,6.25和12.50μmol/l的shizukaold相比dmso对照,荧光素酶活性比值top/fop有显著下降。这表明shizukaold使wnt通路受到抑制。这与wnt通路潜在抑制剂xav939的表型一致,表明shizukaold具有抑制wnt通路的作用。
实施例2wnt通路的差异基因的验证
smmc-7721细胞贴壁后,加入shizukaold,分别在0、6和12小时收样,trizol法提取rna,去基因组rna,反转录后,通过实时pcr检测样品中基因表达量。以gapdh作为内参。利用real-timequantitypcr(q-pcr)检测wnt通路相关基因的扩增循环数差异(-δδct),由此判断该基因的表达量变化。
如图2所示,shizukaold加入12小时的样本与未加入shizukaold的样本相比,c-myc、cyclind、tcf-1、lef1、wnt3a和fgf18的表达量都呈下降。qpcr结果显示,shizukaold下调c-myc等基因的表达量。这可能是由于shizukaold能够抑制β-catenin的表达,从而减弱了其对下游靶基因c-myc的转录激活作用。c-myc是第一个被确定的wnt信号通路的靶基因。c-myc扩增、过表达的主要作用之一是人结节增生和肝癌发生进展。利用sirna等方法导致的c-myc下调使e2f1减少,阻滞人类和大鼠肝癌的体外生长。与缓慢生长型大鼠相比,化学诱导f344转基因鼠易产生不典型增生结节(dysplasticnodule)和肝细胞癌,易于发生肝癌,肿瘤生长快速,c-myc扩增、表达更迅速。研究发现,c-myc基因具有诱导细胞增殖和凋亡的双向功能,其作用取决于细胞接受的信号。当有生长因子如胰岛素生长因子1、表皮生长因子或致癌因素等存在时,c-myc促进细胞增殖;而当生长因子缺乏或有抑制细胞周期因子和抑癌因素等存在时,c-myc会促进细胞凋亡。
结论:shizukaold能够抑制wnt信号通路下游基因的表达水平。
实施例3加入wnt3a部分抵消shizukaold对肝癌细胞生长的抑制作用
加入wnt3a是经典的激活wnt通路的方法。l-wnt3a细胞的培养基中含有足量wnt3a,通常能够激活细胞的wnt通路。取smmc-7721细胞,贴壁后加入含有wnt3a的培养基,孵育过夜。加入6.25和12.50μmol/l的shizukaold,作用48小时后利用cck-8试剂盒检测细胞存活率。以0.1%的dmso为对照。
结果如图3所示,加入含wnt3a培养基能够显著提高shizukaold作用下smmc-7721细胞的存活率。无论加入6.25还是12.50μmol/l的shizukaold,一旦培养基中加入wnt3a,smmc-7721细胞的活力都能恢复到相同水平,这表明,培养基中加入wnt3a能够抵消这两组的化合物浓度差。
结论:激活wnt通路能够补偿shizukaold对细胞生长增殖的抑制作用。本发明的前期试验表明,shizukaold能够抑制肿瘤细胞的增殖。本发明的试验表明,shizukaold能够抑制wnt通路,降低β-catenin表达量,从而抑制肿瘤细胞的增殖。
实施例4外源β-catenin部分抵消shizukaold对细胞生长的抑制作用
取smmc-7721细胞,种24孔板(每孔约5×104个细胞)。贴壁后,转入野生型β-catenin、突变型β-catenin及pcdna3.0空载体质粒。隔天加入10μmol/l的shizukaold以0.1%dmso作为药物对照,处理24小时,每个样做3个复孔。利用cck-8实验检测smmc-7721细胞的吸光值以表征存活率。**:p<0.01。
如图4显示,在smmc-7721细胞中转入野生型β-catenin或者突变型β-catenin(试剂盒lipofectamineltx和plus,invitrogen,cat.noa12621)对肝癌细胞增殖的抑制。没有加入shizukaold的细胞的存活率在加入空载体、野生型β-catenin或者突变型β-catenin后,吸光值几乎没有变化。加入10μmol/l的shizukaold后,相对于空载体,野生型β-catenin组的细胞存活率增加,而突变型β-catenin组的吸光值有显著增加,表明细胞的增殖抑制现象被挽救。而且,加入shizukaold后细胞即使转入突变型β-catenin仍然无法达到不加药组的细胞活力。
同样条件下,在hek-293t细胞中检测外源β-catenin对shizukaold作用的影响。在hek-293t细胞中转入野生型β-catenin和突变型β-catenin,测试加入shizukaold对细胞增殖的影响。突变型β-catenin(pcdna3.0-stabilizedmutantβ-catenin)是s33a、s37a、t41a突变的β-catenin,该突变体相比于野生型β-catenin,不易被磷酸化降解,因而较稳定。结果与smmc-7721类似,没有加入shizukaold的细胞的存活率在加入空载体、野生型β-catenin或者突变型β-catenin后,吸光值几乎没有变化。加入10μmol/l的shizukaold后,相对于空载体,野生型β-catenin组和突变型β-catenin组的细胞存活率均有增加,而突变型β-catenin组的吸光值有显著增加,表明细胞的增殖抑制现象被挽救。
结论:激活wnt通路能够补偿shizukaold抑制细胞的作用;shizukaold能够抑制β-catenin的活性。
实施例5shizukaold导致β-catenin的蛋白量下降
取肝癌细胞种于24孔板(3-6×104个/孔),待贴壁后加入6.25μmol/l、12.5μmol/l、25.0μmol/l的shizukaold,以0.1%dmso作为对照。化合物孵育48小时后,裂解细胞,配平总蛋白,sds-page电泳检测β-catenin的蛋白量。β-catenin是wnt通路的核心,因此本研究检测了shizukaold对肝癌细胞中β-catenin表达的影响。
shizukaold处理48小时后,focus细胞中β-catenin蛋白量减少,当shizukaold的浓度达到25μmol/l时,β-catenin的蛋白量大幅下降(图5a);随着化合物作用时间的增加,β-catenin表达量进一步下降(图5b)。smmc-7721细胞中β-catenin蛋白量也逐步减少(图5c和d)。与focus细胞相比,smmc-7721细胞对shizukaold敏感度稍低,因此增加shizukaold后,β-catenin表达量下降但是不如focus细胞显著。
可见,shizukaold促进肝癌细胞中β-catenin表达量下降,这种效应具有时间和浓度依赖性,shizukaold能够抑制β-catenin的表达水平。
实施例6β-catenin在细胞核中的表达量下降
经典wnt通路激活后,细胞最直观的表现是β-catenin在细胞质中积累、聚集甚至入核,启动下游靶基因的转录。超过40%的肝癌患者中,β-catenin在核中聚集,这是wnt通路激活的标志之一。本研究利用免疫荧光定位方法检测加入shizukaold后β-catenin在细胞内蛋白量的变化。取肝癌细胞focus和smmc-7721种于6孔板(1-2×105/孔),板中预先植入盖玻片一片。待细胞贴壁后加入10μmol/l的shizukaold,以0.1%dmso作为对照。孵育到预定时间,pbs清洗,封闭。0.2%tritonx-100打孔,一抗鼠抗人β-catenin,4℃过夜。二抗羊抗鼠,室温1小时。dapi染色,最后清洗,confocal观察,拍照。红色染色β-catenin,蓝色染dapi。
结果显示,以10μmol/l的shizukaold处理24小时后,β-catenin的表达量下降,细胞核中β-catenin的表达量也明显下降,甚至几乎不可见。10μmol/l的shizukaold处理24小时后,smmc-7721细胞中β-catenin的表达量下降,细胞核中β-catenin的表达量也明显下降。
结论:shizukaold能够抑制β-catenin的表达量,阻止其入核从而激活下游基因的转录。
在本发明提及的所有文献都在本申请中引用作为参考,就如同每一篇文献被单独引用作为参考那样。此外应理解,在阅读了本发明的上述讲授内容之后,本领域技术人员可以对本发明作各种改动或修改,这些等价形式同样落于本申请所附权利要求书所限定的范围。
- 还没有人留言评论。精彩留言会获得点赞!