一种腈水解酶及其应用的制作方法
本发明涉及生物技术领域,具体而言,涉及一种腈水解酶及其应用。
背景技术:
(r)-(-)-邻氯扁桃酸是一种具有广泛用途的医药中间体和精细化工品,主要用于抗血小板凝集药物氯吡格雷的合成。氯吡格雷作为全球重磅级药物,2010年全球销售额近100亿美元,2011和2012年的销售额也都在90亿美元以上。随着2012年5月17号氯吡格雷专利到期,全球掀起了一股仿制热潮。而作为其重要中间体的(r)-(-)-邻氯扁桃酸随之也水涨船高,国内外需求不断增长,因此研究如何获得高浓度、高纯度的(r)-(-)-邻氯扁桃酸将具有重要的市场意义。
国内外通过腈水解酶酶法制备(r)-(-)-邻氯扁桃酸已有报道,但数量不多。张陈胜通过传统的基因组探矿技术从stappiaaggregata中筛选到对邻氯扁桃腈具有对映体选择性的腈水解酶lan,产物(r)-(-)-邻氯扁桃酸的ee值为96.5%,但活力不高。basf公司构建了来源于a.faecalisatcc8750的腈水解酶y296位的不同突变体,其中y296a突变体对外消旋的邻氯扁桃腈的相对比活力为y296野生型的近10倍。y296c突变体对所检测的每一个取代的扁桃腈都具有相当高的活力,而对于未发生取代的扁桃腈活力则没有改变。
有鉴于此,特提出本发明。
技术实现要素:
本发明的目的在于提供一种新型的重组腈水解酶,以解决上述问题。
为了实现本发明的上述目的,特采用以下技术方案:
一种腈水解酶,其氨基酸序列如seqidno:1所示。
如上所述的腈水解酶在催化合成r-邻氯扁桃酸中的应用。
如上所述的腈水解酶在催化合成r-邻氯扁桃酸的中间产物中的应用。
本发明以源于bradyrhizobiumjaponicumusda110的腈水解酶(bll6402,np_773042)氨基酸序列为模板进行改造,获得的新型重组腈水解酶的氨基酸序列如seqidno.1所示。本发明可推动腈水解酶在手性医药中间体合成中的应用,极大降低r-邻氯扁桃酸的生产成本,进而降低氯吡格雷的价格,具有很好的示范作用,将会产生良好的社会效益与经济效益。而通过生物催化法的温和反应条件将显著降低能耗及三废排放量。项目的顺利实施不仅可以实现r-邻氯扁桃酸的绿色生产,还可以为建立以腈水解酶为基础的生物催化法制备手性药物中间体等手性化合物的技术平台提供技术支撑,能够提高我国的生物、化工、医药等领域的手性药物、手性食品添加剂和手性农药及其中间体等的研发水平,有利于我国的绿色生产、工业生物技术的发展,有着广阔的应用前景,对国民经济的可持续发展具有重要意义。
本发明还请求保护编码权利要求1所述的腈水解酶的基因。
优选的,所述基因的核苷酸序列如seqidno:2所示。
需要注意的是,本领域技术人员可根据本发明提供的腈水解酶的氨基酸,根据密码子的简并性原则等自行调整其核苷酸序列以及功能等价体,这些经过调整的序列也在本发明请求保护的范围内。
此外,本发明还请求保护在严格条件下能够与seqidno:2所示核苷酸序列杂交的核苷酸序列;以及与上述序列互补的核苷酸序列。
功能等价体序列还包括与seqidno:2所示序列有至少90%、95%、96%、97%、98%、或99%序列同一性,且具有腈水解酶活性的基因序列,可以从任何植物中分离获得。其中,序列同一性的百分比可以通过公知的生物信息学算法来获得,包括myers和miller算法(bioinformatics,4(1):11-17,1988)、needleman-wunsch全局比对法(j.mol.biol.,48(3):443-53,1970)、smith-waterman局部比对法(j.mol.biol.,147:195-197,1981)、pearson和lipman相似性搜索法(pnas,85(8):2444-2448,1988)、karlin和altschul的算法(altschul等,j.mol.biol.,215(3):403-410,1990;pnas,90:5873-5877,1993)。这对于本领域技术人员来说是熟悉的。
一种表达载体,其包含如上所述基因的序列。
优选的,所述表达载体为pqe30。
一种工程菌,其被如上所述的表达载体所转化。
优选的,所述工程菌为大肠杆菌。
更优选的,所述工程菌为m15大肠杆菌菌株。
与现有技术相比,本发明提供的腈水解酶可在12h反应时间里,共将600mm的底物完全水解,由于底物流加和ph调控的稀释作用,累计产生了112g/l的产物邻氯扁桃酸。整个反应过程中反应转化率保持在90%以上。反应10h后停止流加,继续反应2h后底物全部转化完。反应速率非常快,反应转化率也很高。
附图说明
为了更清楚地说明本发明具体实施方式或现有技术中的技术方案,下面将对具体实施方式或现有技术描述中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图是本发明的一些实施方式,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他的附图。
图1为发酵过程中od600的增长情况;
图2为发酵过程中dcw的增长情况;
图3为发酵过程中酶产量的增长情况;
图4为连续流加反应制备r-邻氯扁桃酸进程结果图。
具体实施方式
下面将结合实施例对本发明的实施方案进行详细描述,但是本领域技术人员将会理解,下列实施例仅用于说明本发明,而不应视为限制本发明的范围。实施例中未注明具体条件者,按照常规条件或制造商建议的条件进行。所用试剂或仪器未注明生产厂商者,均为可以通过市售购买获得的常规产品。
1.1新型腈水解酶的基因挖掘
1.1.1序列筛选
以源于bradyrhizobiumjaponicumusda110的腈水解酶(bll6402,np_773042)氨基酸序列为模板,通过blastp(http://www.ncbi.nlm.nih.gov/blast/blast.cgi)程序搜索潜在的扁桃腈腈水解酶。我们设立了三个标准对blastp的结果进行筛选。首先氨基酸序列一致性高于90%或者低于30%的都被剔除掉,将与bll6402具有高度相同性和不同性的酶过滤掉。具有相同序列一致性的酶只保留一到两个。第二个标准是序列的来源。来源于同一种属的序列只保留一个。因为他们往往具有非常高的序列一致性和相同的酶学特性。来源于同一菌株的所有不同序列全部保留。第三个标准是来源菌株的可获得性。
除去上面三个标准,所有经过实验验证具有腈水解酶活性的序列都被挑出来。这些酶的底物特异性大部分都经过了实验验证,为后面的系统进化分析提供分类依据。expasy蛋白质组学服务器上的scanprosite程序用来检测筛选出的序列是否具有glu-lys-cys催化三联体。
1.1.2数据分析
筛选出来的氨基酸序列用clustalw先进行聚类分析。聚类后的数据用mega4.0软件包中的邻接法进行系统进化分析。bootstrap值设定为1000。
1.2腈水解酶的定向进化
1.2.1突变子文库构建
突变引物含有酶切位点(bamhi/hindiii)及保护碱基。以质粒bcj2315/pqe30作为模板,采用天恩泽的易错pcr试剂盒进行扩增。带有突变碱基的pcr产物经bamhi/hindiii双酶切后连于经同样酶切过的线性载体pqe30,转化大肠杆菌m15。
1.2.2突变子筛选
从平板上挑取单克隆(8000个转化子),接种于24孔培养板中,30℃培养36h。离心收集菌体。培养基采用自诱导培养基。将上述离心后的菌体用1ml生理盐水清洗一遍,离心。菌体用1ml100mm磷酸钠缓冲液(ph8.0)悬浮,30℃温育10min后加入20μl1m的邻氯扁桃腈起始反应。反应12h后加入100μl2mhcl终止反应。取200μl上清液13,000g离心10min后用测氨法检测酶活,有活性的反应液通过hplc检测ee值。
1.3结果与分析
通过优化易错反应pcr条件,突变频率控制在3-7碱基/kb,转化效率为1000克隆/10μl连接液,有效克隆率达到70%以上。
通过14轮共8000个突变子的筛选,我们得到了4个有益突变株,ee值均在95%以上。通过测序发现,四个突变体最少含有一个突变子。其中突变体18和1332,其49位的苏氨酸都发生了突变。在突变体18中,49位的苏氨酸突变成了丝氨酸。在突变体1332中,49位的苏氨酸突变成异亮氨酸。在突变体338和1332中,113位的异亮氨酸都突变成了天冬酰胺。虽然ee值都提高了,但四个突变体在水解邻氯扁桃腈时都表现出了活力的下降。
表1随机突变产生的腈水解酶bcj2315突变体及单点突变体酶活及对映体选择性比较
为了验证四个突变体中的有益突变子,通过单点突变,我们得到了四个突变体中的所有突变子并对酶进行了纯化。所有突变子的水解活力及ee值如表6.4所示。在突变体18中,只有一个突变位点,该突变位点突变后相对酶活为87%,ee值提高到95.9%。突变体338中,a91s为无义突变,i113n提高了产物的ee值,但活力都有所下降。在突变体101中,m98k和r345c为无义突变。y199c和t310p表现出了ee值的提高。其中t310p的相对活力为33%,y199c的活力则提高到了201%。在突变体1332中,y85f为无义突变。t49i和i113n均表现出了ee值的提高和活力的下降。
腈水解酶bcj2315的催化三联体为e48-k130-c164,在我们所得到的有益突变体中,离催化三联体最近的位点是第49位的t。而另外一个对对映体选择性有影响的位点是t310,该位点在氨基酸序列上远离腈水解酶的催化活性中心。因此,靠近和远离腈水解酶催化中心的氨基酸突变都能影响到酶的对映体选择性。研究者往往倾向通过对催化中心附近的位点进行点突变来达到改善酶某个性质的目的,因为对靠近催化中心的位点进行突变比对远离催化中心的位点进行突变具有更高的效率。我们更倾向于通过随机突变来达到提高bcj2315选择性的目的。因为靠近催化中心的位点毕竟是有限的,而随机突变作用于整个基因序列,能够产生更多的远离催化活性中心的有益突变。
通过定点突变,共有4个位点在提高bcj2315对映体选择性方面做出了有益的贡献。其中i113n表现出了最高的ee值,t49i的ee值提高的最低,为95.7%。在所有突变子当中,y199c表现出了最高的相对酶活。但y199c的ee值只有94.3%。综合考虑活力及对映体选择性,为了得到一个更好的突变子,我们将该4个位点进行了饱和突变,利用全细胞催化测定所有的突变子在酶活和ee值方面的表现。
表2腈水解酶bcj2315单点饱和突变体酶活及对映体选择性比较
在筛选有益突变体的过程中,活力及ee值都是我们考虑的因素。ee值大于95%而活力没有下降的突变体将会被优先挑选出来。在t49所有突变体中,只有t49n表现出了活力的微弱提高,但其ee值相比野生型bcj2315下降了很多,只有76.1%。高于95%的突变体有8个(t49a,t49e,t49f,t49g,t49i,t49m,t49s,t49y),但这8个突变体的相对活力都下降了,最高的只有野生型bcj2315的87%(t49s)。t310所有突变体中,ee值高于95%的只有t310p,t310p表现出了很好的对映体选择性,ee值为98.1%。但相对活力却只有野生型的32%。在t49和t310位点的饱和突变中我们没有发现有益突变。绝大多数i113突变体都表现出了优秀的对映体选择性,其中7个突变体的ee值都大于99.9%。遗憾的是大部分突变体都表现出了相对酶活的降低,这7个突变的相对活力只有野生型的1%-6%,表现出了极低的相对酶活。两个i113突变体表现出了相对较高的酶活力,i113m和i113l。相对酶活分别为138%和188%。且i113m的对映体选择性很好,产物ee值为97.2%。因此i113m突变体被选定用于进一步研究。与i113突变体不同的是,y199所有突变体绝大多数都表现出了相对酶活的提高,但对映体选择性却提高的不大。ee值高于95%只有3个,y199a,y199e和y199g,ee值分别为95.6%,95.1%和96.7%。因此这几个突变体也被挑选出来做进一步的研究。
考虑到突变体的ee值及相对酶活力,共四个有益突变体被挑选出来。它们是i113m,y199a,y199e和y199g。四个突变体都表现出了活力和ee值的提高,产物ee值最高为97.6%。在4个位点的饱和突变体中,我们观察到y199绝大部分都表现出了相对酶活力的提高和ee值的提高,而在另外三个位点的饱和突变中,只有i113m被挑选出来,表现出了ee值和相对酶活的双重提高。位点的联合突变通常会引起有益结果的叠加效应。因此我们选取i113m突变体作为模板,通过对i113m突变体的199位进行饱和突变,考察一下两个位点的联合突变是否会进一步提高酶的相对酶活和对映体选择性。结果如表6.6所示。通过两点的联合突变,我们得到了很多有益的突变。所有的突变体ee值都高于97%,最高的ee值为98.7%,由突变体i113m/y199g产生。突变体中最高的相对酶活力也由突变体i113m/y199g产生,为野生型的3.76倍,因此i113m/y199g被选作进一步的研究。
目前已有很多研究报道了在基因水平上改造腈水解酶,通过蛋白质工程改变腈水解酶的底物特异性、对映体选择性、ph耐受性、底物耐受性及提高酶的活力等。kiziak发现去除pseudomonasfluorescensebc191腈水解酶c末端47-67个氨基酸导致酶活降低,酰胺产物的增多和对映体选择性的改变。kiziak鉴定了一个保守的氨基酸位点h296,该位点与pseudomonasfluorescensebc191腈水解酶对映体选择性和酰胺的形成有关。desantis对通过宏基因组方法得到的一个腈水解酶进行全基因饱和突变,得到的突变体可以耐受高浓度的3-羟基戊二腈(3m)且对映体选择性提高到了98.1%。来源于敏捷食酸菌的脂肪族腈水解酶,一个单一的突变(t210a)使其对3-羟基戊腈的活性提高了7.3倍。两个点的联合突变(f168v+l201n)使其对羟基乙腈的活性提高了15.3倍,且突变体酶活,体积产率,产物收率都满足工业上的需求。来源于紫红红球菌的腈水解酶,y142残基被证明对底物特异性起作用。来源于neurosporacrassaor74a的腈水解酶,w168a突变体导致其在水解扁桃腈和2-苯基丙腈时产生更多的酰胺。且对2-苯基丙腈的对映体选择性完全反转(由r型反转为s型)。迄今为止,通过基因工程改造腈水解酶使之能够高度对映体选择性水解外消旋邻氯扁桃腈生成(r)-(-)-邻氯扁桃酸尚未见报道。schreiner通过定向进化技术得到了来源于alcaligenesfaecalis的腈水解酶nitaf的突变体phnit45,该突变体在ph4.5时具有较高的稳定性。phnit45在ph4.5的反应条件下可以水解r型的邻氯扁桃腈生成(r)-(-)-邻氯扁桃酸,产物ee值大于99%。但在ph7.0时,nitaf和phnit45的选择性都很差,分别为76%和19%。在我们的研究中,通过对腈水解酶bcj2315的定向进化,得到的有益突变体在活力和ee方面都有了很大的提高,可以在中性及偏碱性反应条件下水解外消旋邻氯扁桃腈生成(r)-(-)-邻氯扁桃酸,理论产率为100%,ee值为98.7%。
表3腈水解酶bcj2315突变体i113m199位单点饱和突变体酶活及对映体选择性比较
在i113m/y199g突变体基础上,针对催化活性位点的氨基酸继续进行迭代饱和突变,在筛选有益突变体的过程中,筛选到一株有益突变株,经过测序发现g59a突变体在i113m/y199g突变体基础上,酶活有了进一步提高,因此在后续r-邻氯扁桃酸制备工艺中,采用g59a/i113m/y199g(mga)突变体作为生物催化剂。
2腈水解酶催化合成r-邻氯扁桃酸工艺研究
2.1.1摇瓶培养
如无特殊说明,以下培养过程中,lb培养基中均需添加kan0.05mm,amp0.1mm。
(1)取保藏于-80℃下的e.coli工程菌菌种,接种于lb培养基中,37℃培养过夜。
(2)过夜菌0.5%(v/v)接种于400mllb培养基中,37℃培养至od600=0.6。
(3)培养物中加入iptg至终浓度0.1mm,30℃培养16h后收集菌体。
2.1.2催化剂制备
(1)摇瓶培养所获得的培养物于4℃下8000rpm离心5min。收集菌体。
(2)菌体重悬于100mmph8.0的na2hpo4/nah2po4缓冲液中,4℃8000rpm离心5min,收集菌体。
(3)重复步骤(2)一次。
(4)缓冲液洗涤后的菌体于-20℃保存备用。
2.1.3e.coli/mga工程菌发酵
对于工业级制备r-邻氯扁桃酸,需要大批量的工程菌作为催化剂,因此有必要建立一条廉价稳定的e.coli/mga工程菌发酵工艺。高密度发酵是指在一定条件和培养体系下,获得最多的细胞量,由此更多地或更高效地获得目的产物。实现高密度发酵不仅可以减少培养体积,强化下游分离提取,还可以缩短生产周期,减少设备投资,从而降低生产成本。
利用一般的发酵工艺生产大肠杆菌或以其构建的基因工程菌的表达产物,大肠杆菌的生物量,蛋白表达量,代谢产物在菌体内的发酵液中的浓度都比较低,难以获得理想的生产效率。重组大肠杆菌进行高密度发酵可获得较高的生物量,但对发酵条件有非常高的要求。影响高密度发酵的因素非常多,如细胞生长所需要的营养物质、发酵过程中生长抑制物的积累,溶氧浓度,诱导方式,发酵液的ph值,补料方式及发酵液流变学特性等。
2.1.3.1
培养基
培养基按组成成分可分为合成培养基、半合成培养基、复合培养基。其组成元素包括:c、h、o、n、s、p、fe、mg、k等。大肠杆菌高密度发酵所使用的培养基一般为半合成培养基,它是在合成培养基的基础上添加能促进细胞生长及代谢产物形成的不同物质,如适量无机盐,氨基酸,维生素等。常用的碳源有糖类,油脂,有机酸和低碳醇。葡萄糖是碳源中最易利用的糖,常被用作培养基的主要成分。常用的氮源可分为有机氮源和无机氮源。培养基各组分的浓度和比例要适当,尤其是碳源与氮源的比例。如果碳源与氮源浓度过高,苏安然比例合适,发酵起始会导致菌体的大量繁殖,但发酵后期菌体生长缓慢,代谢废物过多,增大发酵粘度,影响溶解氧浓度,容易引起菌体的代谢异常。因此设计一种平衡的培养基有利于产物的表达。
在培养基组分的设计过程中,我们采用蛋白胨与酵母膏作为基础碳氮源,因为两者不仅可以提供工程菌生长过程中所需的碳氮源,而且提供微量元素与生长因子等微量物质。在此基础上,添加少量甘油作为速效碳源有利于发酵初期菌体的生长。磷酸钾盐的添加有利维持发酵液的ph平衡。mg2+的额外添加有利于提高酶的活力。
表4发酵基础培养基组分
表5发酵补料培养基组分
2.1.3.2ph值
稳定的ph值是使菌体保持最佳生长状态的必要条件。由于外界的ph值变化会通过弱酸或弱碱的变化而改变菌体细胞内的ph值,从而影响细胞的代谢反应。因此在发酵过程中ph值的改变会影响细胞的生物量和基因产物的表达。另一方面,微生物的生长可引起培养液ph值的变化。在基本培养基中培养微生物,通常ph值会发生较大的变化。铵盐中氨的消耗,或者微生物利用碳源产生的代谢产物即有机酸的积累,都会造成ph值的下降,而有机酸的消耗则会造成ph值的上升。在复合培养基中培养,当消耗碳源时,培养基中的氨基酸被用于异化代谢时,因氨的释放,会造成ph值升高。发酵过程中跟踪ph值的变化,可以为了解微生物代谢的变化提供十分重要的信息。在发酵工艺控制中,必须及时使用低浓度的碱或氨水调节ph值使之处于适宜的ph范围内,避免ph值剧烈变化对细胞生长和代谢造成的不利影响。
在e.coli/mga工程菌的发酵过程中,我们控制ph在7.2左右,在该ph条件下有利于工程菌的生长和mga腈水解酶的积累。由于大肠杆菌产酸的积累,会造成发酵液ph值的降低,ph值的维持最重要的手段是碱性物质的添加。在本发酵中,氨水与naoh溶液均在ph调控手段的考虑范围之内。naoh溶液用于调控ph优势在于灭菌简便,不挥发,且可配制成高浓度溶液,其缺点在于由于naoh为强碱性物质,在滴入过程中会造成局部发酵液瞬间产生强碱性环境,对菌体造成破坏。氨水用于调控ph的优势在于氨水相对碱性较弱,调节相对较温和,但是氨水灭菌相对麻烦,需过滤灭菌,且由于氨水极易挥发,对操作人员危害较大。
2.1.3.3溶解氧
溶解氧浓度是好氧微生物高密度发酵过程中影响菌体生长的重要因素之一,而且它往往会成为发酵的控制因素。这主要是由于氧在水中溶解度很小所致。在细胞的对数生长期若停止供氧,则发酵液中的溶解氧会在几分钟内耗尽。在发酵后期,反应器中生物量很大,而且发酵液粘度增大,此时受反应器供氧能力的限制,发酵液中的溶解氧浓度会低于临界氧浓度。这样,氧的溶解和传递便成了该细胞反应的限制步骤。
发酵开始阶段,控制溶氧在20%以上。溶氧的控制主要通过逐步提高搅拌速度以及改变通气量的方式控制。初期搅拌速度设定为300rpm,搅拌转速的最终速度为700rpm。
2.1.3.4温度与诱导条件
培养温度是影响菌体生长和调控细胞代谢的重要因素。在发酵初期,温度设定为37℃,有利于细胞的快速生长与繁殖。当进入对数生长期后,通过iptg进行诱导以调节工程菌的基因表达水平。此时以获得较大量的目标蛋白即mga腈水解酶为主要目标。由于在较高温度下工程菌内mga的表达容易形成较多无活性的包涵体形式,因此在诱导开始后,需适当降低工程菌的代谢水平,此时将温度降低至30℃进行发酵。在较低的温度下有利于mga的可溶性表达,降低包涵体的形成。
2.1.3.5发酵结果
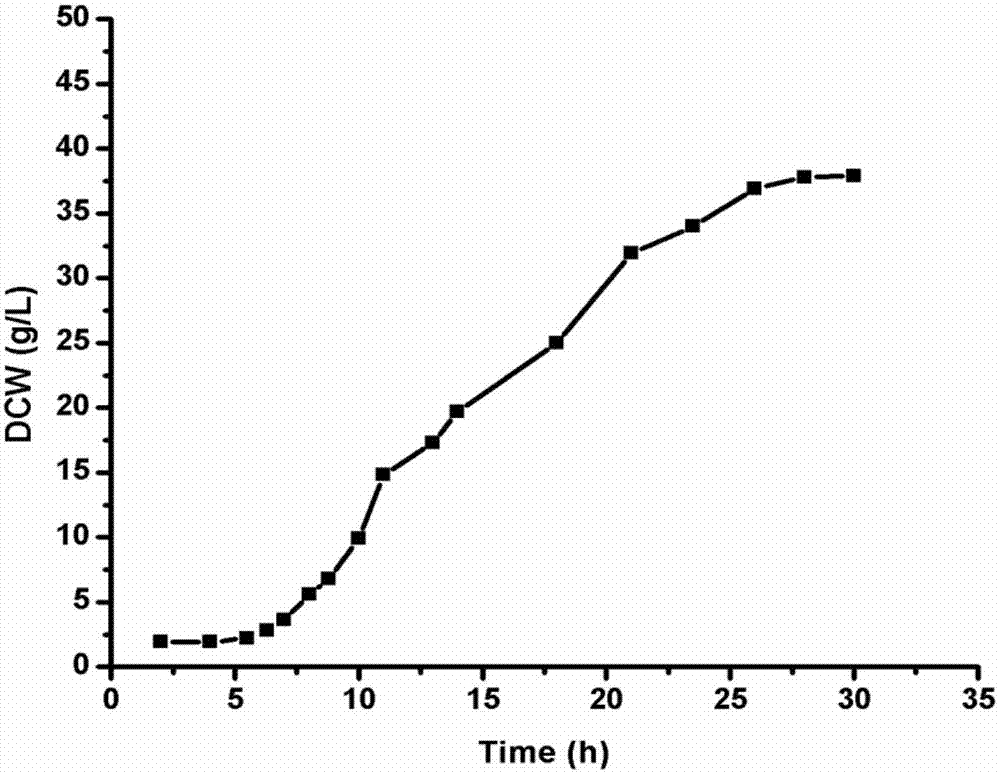
综合上述条件,对工程菌e.coli/mga进程了发酵,在发酵过程中不同时间点进行取样分析,检测发酵过程中od600,dcw以及酶产量的增长情况。具体结果见图1,图2,图3结果所示。
由上述三图可以看到,通过对发酵培养基配方的设计,以及在发酵过程中ph,温度以及溶氧的调控。以酶产量为主要目标同时兼顾菌体浓度的条件下,工程菌的发酵浓度达到了od600在75左右,dcw在35g/l左右,酶产量达到35000ku/l。酶产量在腈水解酶发酵领域处于领先水平。
2.2催化反应
腈水解酶催化水解邻氯扁桃腈制备r-邻氯扁桃酸的反应采用连续流加补料的反应模式,通过维持底物浓度在较低的水平以减缓酶失活的速率,通过连续流加补料,以达到r-邻氯扁桃酸的高浓度积累。
2.2.1转化液制备
腈水解酶催化水解邻氯扁桃腈制备r-邻氯扁桃酸的转化反应于100mmph8.0的na2hpo4/nah2po4缓冲液中进行。
其中,缓冲液中添加乙醇10%(v/v)作为助溶剂。
20mg/ml终浓度的静息细胞。
配制后的转化液保温至30℃用于后续反应。
2.2.2流加底物的制备
用于流加反应的邻氯扁桃腈溶于乙醇中配置成终浓度为6m的流加液,流加液于30℃条件下保温。
2.2.3连续补料流加反应
高浓度的邻氯扁桃腈对腈水解酶mga的抑制作用非常明显,更容易造成酶的失活。为了得到高浓度的产物(r)-(-)-邻氯扁桃酸,我们通过采用连续流加底物模式来调整反应中的底物浓度,使反应始终保持在一个较高的速率下进行,以达到高浓度(r)-(-)-邻氯扁桃酸的累积生产。在流加底物的过程中,将底物溶解在有机溶剂中,可以同时提高反应体系的有机助溶剂,以同时达到提高产物对映体选择性的目的。向反应体系中加入有机溶剂还可以提高反应的速率,减少反应时间。我们选定乙醇作为助溶剂。底物邻氯扁桃腈溶于乙醇形成高浓度的母液(6m),在流加底物的同时不断提高反应体系中乙醇的含量,使反应的活力和ee值都处于一个增长的过程弥补反应过程中部分酶的失活。起始反应乙醇的添加量为20%,在此浓度下,生成产物的ee值为95.9%。通过tlc监测反应过程,调整底物补料速度。
300ml的反应于500ml的三口圆底烧瓶中进行,水浴保温30℃,双桨叶搅拌桨机械搅拌转速350rpm。14#软管流加底物,流加速度60mm/h,其中软管与流加液需30℃保温。
ph电极监控反应ph的变化,6m氨水维持ph在7.95-8.10之间,其中ph的调节与监控电极联动。
2.2.4检测方法
反应进程的监控采用tlc在线监控,hplc定量检测的方法。检测方法具体细节见分析方法一节。
2.2.5反应结果
底物以60mm/h的流加速度连续流加10h后停止流加,继续延伸反应2h后结束反应。反应进程结果见图4。
在12h反应时间里,共600mm的底物被完全水解,由于底物流加和ph调控的稀释作用,累计产生了112g/l的产物邻氯扁桃酸。整个反应过程中反应转化率保持在90%以上。反应10h后停止流加,继续反应2h后底物全部转化完。
目前,关于利用腈水解酶大规模生产邻氯扁桃酸的文献报道并不多。张陈胜利用含有来源于labrenziaaggregata的重组腈水解酶细胞通过水/甲苯两相体系在8h内将300mm的邻氯扁桃腈完全水解,产率为94.5%,产物ee值为96.5%。含有腈水解酶的节杆菌(arthrobactersp.)f-73通过水/乙酸乙酯两相体系,通过分批添加邻氯扁桃腈(终浓度100mm),9h内共转化400mm的邻氯扁桃腈,产率为92%,产物ee值为98.5%。产物经回收并重结晶后ee值可提高到99.3%。
2.3产物分离纯化
r-邻氯扁桃酸的纯化采用结晶法进行。经过离心处理后的反应液,通过浓hcl进行酸化至ph1,通过旋转蒸发浓缩至1/3体积,在10℃条件下冷却结晶,收集晶体,重复浓缩结晶过程2次后,将收集到的晶体溶于乙醇中进行重结晶。重结晶晶体烘干后装袋。
2.4产品检测
纯化后的r-邻氯扁桃酸检测结果如下:
1hnmr(400mhz,d2o)δ/ppm:5.530(s,1h),7.262–7.300(m,2h),7.322–7.356(m,1h),7.382–7.415(m,1h).
[α]d25=-153(c=0.5,ethanol)
ee>98%
经检测,产品符合企业标准。
最后应说明的是:以上各实施例仅用以说明本发明的技术方案,而非对其限制;尽管参照前述各实施例对本发明进行了详细的说明,但本领域的普通技术人员应当理解:其依然可以对前述各实施例所记载的技术方案进行修改,或者对其中部分或者全部技术特征进行等同替换;而这些修改或者替换,并不使相应技术方案的本质脱离本发明各实施例技术方案的范围。
sequencelisting
<110>枣庄市杰诺生物酶有限公司
<120>一种腈水解酶及其应用
<160>2
<170>patentinversion3.3
<210>1
<211>334
<212>prt
<213>人工序列
<400>1
metglnaspthrlysphelysvalalavalvalglnalaalaproval
151015
phemetaspalaproalaservalalalysalaileglypheileala
202530
glualaglyalaalaglyalalysleuleualapheprogluvaltrp
354045
ileproglytyrprotrptrpleutrpleualathrproalatrpgly
505560
metglnphevalproargtyrhisalaasnserleuargalaaspgly
65707580
proaspileleualaleucysalaalaalaalaglualalysileasn
859095
valvalmetglyphesergluileaspglyglythrleutyrleuser
100105110
metvalpheileseraspalaglygluileilephelysargarglys
115120125
leulysprothrhisvalgluargthrleutyrglygluglyaspgly
130135140
serasppheargvalvalgluserservalglyargleuglyalaleu
145150155160
cyscysalagluhisileglnproleuserlystyralamettyrser
165170175
metasngluglnvalhisvalalasertrpproserphethrleutyr
180185190
argasplysalatyralaglyglyhisgluvalasnleualaalaser
195200205
glniletyralaleugluglyglycysphevalleuhisalaserala
210215220
ilethrglyglnaspmetpheaspmetleucysaspthrproglulys
225230235240
alaaspleuleuasnalagluglyalalysproglyglyglytyrser
245250255
metilepheglyproaspglyglnprometcysgluhisleuprogln
260265270
asplysgluglyileleutyralaaspvalaspleusermetileala
275280285
ilealalysalaalatyraspprothrglyhistyralaargglyasp
290295300
valvalargleumetvalasnargserproargargthrservalser
305310315320
phesergluaspgluasnalaalavalthrphethrgluthr
325330
<210>2
<211>1005
<212>dna
<213>人工序列
<400>2
atgcaggacacgaaattcaaagtcgcggtcgttcaggctgcgccggtattcatggatgcg60
ccagcttccgtggccaaggcgattggtttcattgcggaggcgggtgcagcgggggcgaag120
ctgctggcgttcccggaggtctggattccgggctatccttggtggctttggctcgggacg180
ccggcttggggaatgcagtttgtaccccgctatcacgccaattcgctgcgtgctgatggg240
cccgacatcctcgcactgtgtgcggccgccgccgaagcgaaaatcaatgtggtgatgggc300
ttctccgaaatcgacggaggaacgctctacctaagtcaggtctttattagcgatgcggga360
gagatcatcttcaagcgccgaaagctcaagccgacacacgtcgaacgtacgctctatggc420
gaaggagatgggtctgatttccgcgtcgtcgaaagcagcgtcggacgtctcggagccttg480
tgctgcgccgagcacattcagccgttgtcgaaatacgccatgtactcgatgaacgagcaa540
gttcacgtggcgtcgtggccatcttttacgctgtatcgcgacaaggcctacgctttgggc600
catgaggtgaatctcgccgccagccagatctacgcgctagagggaggttgcttcgtcctg660
catgcctcggcaattactggtcaagatatgttcgacatgctttgcgatactccggaaaag720
gccgatttgctgaacgcggagggagcgaagccgggtggaggctattcgatgatcttcggt780
cccgatggtcagccgatgtgcgagcatctgccgcaggacaaggaaggcatcctctatgct840
gacgtggacctgtcaatgatcgcaatcgccaaagcggcctacgaccccacggggcattac900
gcccgcggcgatgtcgtccgtctcatggtcaatcgcagcccgcgtcgcacgagcgtcagc960
ttcagcgaagacgagaacgcggcggtcactttcaccgagacttga1005
- 还没有人留言评论。精彩留言会获得点赞!