一种黄曲霉无毒株及其防治黄曲霉污染的应用的制作方法
本发明属于微生物学领域,具体涉及一种黄曲霉无毒株及其防治黄曲霉污染的应用。
背景技术:
黄曲霉(aspergillusflavus)是广泛分布于自然界的腐生真菌,生长和扩散能力极强。黄曲霉可以寄生于粮食、食品及饲料中进行生长繁殖,并产生黄曲霉毒素,其中以黄曲霉毒素b1(aflatoxinb1,afb1)的危害最大,具有强肝脏毒性和致癌效应。目前全球每年有25%农作物被霉菌毒素污染,其中最为严重的就是黄曲霉毒素的污染。我国花生和玉米被黄曲霉污染的情况比较普遍,畜禽饲料和水产饲料的黄曲霉污染则更为严重,结果导致黄曲霉毒素通过食物链危害人类健康。由于黄曲霉毒素的理化性质稳定,大规模工业加工、日常烹饪条件或者巴氏灭菌等都无法清除污染。因此黄曲霉污染危害巨大,研究预防和控制黄曲霉污染具有重大的意义。
黄曲霉菌丝生长、产孢,和产毒等的能力与其扩散、繁殖和毒力密切相关,直接影响黄曲霉的致病性,一直以来科学家围绕黄曲霉致病性调控机理展开了大量研究。随着黄曲霉全基因组测序的完成,对其致病性相关的遗传因素筛查和研究日渐增多,然而目前发现即使完全相同遗传背景下,黄曲霉的产毒量也可能差异极大。这就提示黄曲霉对非遗传因素的影响非常敏感。这其中有关外界环境因子如营养、温度、水活度、ph值、氧化状态以及群体感性等影响黄曲霉形态和产毒的研究已开展多年,而关于体内非遗传因素调控黄曲霉致病性研究则仍处在起步阶段。
研究发现真菌的次级代谢相关基因的转录和表达极有可能受到组蛋白乙酰化、dna甲基化等表观遗传因素调控。这些表观遗传修饰被认为可能是联接遗传背景、环境因子两大因素的重要桥梁之一,介导两者的复杂交互作用,最终影响生物的性状。在稻瘟病菌中对ash1样的蛋白-mokmt2h的研究发现该基因直接调控稻瘟病菌的孢子萌发和该菌对水稻的毒力。黄曲霉中还未见关于ash1(aflash1)介导的组蛋白甲基化修饰的研究报道,aflash1介导的表观遗传修饰对黄曲霉的致病性是否有影响也是未知的。
本发明人经过广泛研究,通过生物信息学分析首次在黄曲霉中找到一个假定ash1基因,本发明人通过western-blotting分析,确定该基因确实具有组蛋白甲基化转移酶的生物学功能,于是将之命名为aflash1基因。因此,本发明公开了一种黄曲霉无毒株⊿aflash1菌株,该无毒株中黄曲霉不表达致病相关基因aflash1,本发明还公开了一种通过该无毒菌株防治黄曲霉污染的方法。
技术实现要素:
本发明的目的在于提供一种黄曲霉无毒株(⊿aflash1菌株)及其防治黄曲霉污染的应用。
一种黄曲霉无毒株,所述的无毒株中aflash1基因基本上不表达。
所述的无毒株:
(1)aflash1基因片段缺失;
(2)相对于野生型黄曲霉,所述的无毒株产生的孢子数量在培养基上显著上升;
(3)相对于野生型黄曲霉,所述的无毒株产生的孢子数量在玉米粒等有机体上显著下降;
(4)相对于野生型黄曲霉,所述的无毒株不产菌核。
(5)相对于野生型黄曲霉,所述的无毒株未检测到黄曲霉毒素;所述的黄曲霉毒素包括:黄曲霉毒素b1(aflatoxinb1,afb1),黄曲霉毒素b2(aflatoxinb2,afb2),黄曲霉毒素g1(aflatoxing1,afg1),黄曲霉毒素g2(aflatoxing2,afg2)。
(6)相对于野生型黄曲霉,所述的无毒株中产毒相关基因的表达下降;所述的产毒相关基因包括:aflr基因、afls基因、aflc基因、afld基因、aflk基因,和aflq基因。
(8)相对于野生型黄曲霉,所述的无毒株侵染宿主的能力下降;
(9)所述的宿主包括:粮食及其制品、豆类及其制品、坚果及其制品、植物油及其制品、调味香辛料及其制品和饲料;
(10)所述的侵染宿主的能力包括:在宿主上定殖生成菌丝、产生孢子、产菌核,以及产生黄曲霉毒素afb1、afb2、afg1,和afg2。
所述的无毒株通过同源重组的方法从黄曲霉菌株pts(来源于aspergillusflavusca14菌株,引自aspergillusflavusaswa,agenehomologofaspergillusnidulansoefc,regulatessclerotialdevelopmentandbiosynthesisofsclerotium-associatedsecondarymetabolites)的染色体中敲除aflash1基因获得,所述的aflash1基因具有seqidno.1所示的核苷酸序列,所述的aflash1蛋白具有seqidno.2所示的氨基酸序列。
在本发明的第二方面,提供一种基于⊿aflash1无毒株防治黄曲霉污染的方法,所述方法包括:
(1)利用该菌株不产黄曲霉毒素(图2),在人工培养基中高产孢的特点,其产孢量是野生型菌株的3倍以上(图3),在培养基上人工培养大量的该黄曲霉菌株的孢子;
(2)将步骤(1)中获得的孢子释放到受野生黄曲霉污染的区域(包括上述的花生产区)。
(3)利用⊿aflash1株的无毒(图3),不产菌核(图4),在有机体上低产孢,低致病力的特性(图5和图6),通过步骤(2)大量该菌株的分生孢子与野生产毒黄曲霉菌株在花生主产区等黄曲霉易感区域争夺有限的生态位,有效降低野生产毒黄曲霉菌株的密度,从而有效控制和降低作物受到的黄曲霉感染,黄曲霉毒素污染及其引发的曲霉病感染。
本发明的其它方面由于本文的公开内容,对本领域的技术人员而言是显而易见的。
本发明的优点在于:
目前对黄曲霉污染的研究主要集中在黄曲霉毒素致病机理研究,产毒机理研究,以及黄曲霉毒素污染检测方法的改进等方面,对如何控制黄曲霉对作物的污染方面的研究甚少。本发明能有效控制和降低作物受到的产毒黄曲霉感染,有利于在作物生长和存储时从根本上解除黄曲霉毒素的产生,积累,从而有效控制黄曲霉毒素的污染。
我国的广大亚热带地区常年温暖湿润,农作物受黄曲霉及黄曲霉毒素广泛污染,严重危及了食品安全,和广大人民群众的健康,和社会经济的健康发展。而且afs中的afb1具有强肝脏毒性和致癌效应,是现在发现的毒性和致癌性最强的天然污染物。由于afs热学性质稳定,要加热到268℃-269℃才会分解,因此对己经霉变的上述食品,即使加热煮熟也无法消除其中的黄曲霉毒素。而本发明与其他黄曲霉毒素污染检测,以及毒素污染后解毒等技术相比,从源头上控制黄曲霉的污染,成本显著降低,为抑制和减少黄曲霉菌在粮食等农作物生产和存储过程中产生黄曲霉毒素,为避免其对人畜健康和社会经济造成重大危害提供一条可行的途径。因此,本研究具有潜在的巨大的经济和社会效益。
附图说明
图1.⊿aflash1黄曲霉菌株构建。a.⊿aflash1黄曲霉菌株构建模式图的产毒情况;b.q-pcr鉴定在野生型菌株(wt),⊿aflash1菌株,和aflash1基因回补菌株(com-ash1)中aflash1基因的表达情况。
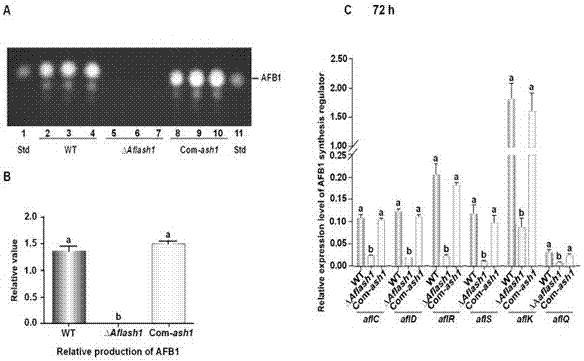
图2.tlc分析⊿aflash1黄曲霉菌株的产毒情况。a.tlc分析结果;b.基于tlc结果柱状图显示⊿aflash1黄曲霉菌株中最主要毒素afb1的合成情况;c.q-pcr分析黄曲霉毒素合成关键基因在各菌株中的表达情况。
图3.⊿aflash1菌株的产孢量显著增加。a.野生型,互补和⊿aflash1菌株在yes培养基上培养4天;b.各菌株孢子梗侧面观(×100);c.柱状图显示⊿aflash1菌株的产孢量显著增加。
图4.⊿aflash1菌株的菌核形成被抑制。a.野生型,互补和⊿aflash1菌株在ypd培养基上培养6天;b.柱状图显示⊿aflash1菌株的菌核形成被彻底抑制。
图5.⊿aflash1菌株在生物体上的产孢和产毒被显著抑制。a.野生型,互补和⊿aflash1菌株在玉米粒上培养7天;b.tlc分析显示⊿aflash1菌株在玉米粒上不产黄曲霉毒素;c.柱状图显示⊿aflash1菌株在玉米粒上的产孢量显著下降;d.柱状图显示⊿aflash1菌株在玉米粒上不产黄曲霉毒素b1。
图6.大田实验中⊿aflash1菌株在花生叶片上的致病力被显著抑制。
图7.western-blotting分析aflash1基因的生物学功能。a.western-blotting分析aflash1基因对h3k9甲基化水平的影响;b.western-blotting分析aflash1基因对h3k36甲基化水平的影响;c.柱状图显示aflash1基因参与了h3k9二甲基化水平的调控;d.柱状图显示aflash1基因主导了h3k36二甲基化水平的调控。
具体实施方式
如本文所用,黄曲霉aflash1基因(seqidno.1)用斜体aflash1表示,黄曲霉的无毒株用⊿aflash1表示,或用⊿aflash1缺失株表示,黄曲霉的回复菌株用com-ash1表示,黄曲霉的野生型菌株用wt表示。黄曲霉aflash1蛋白(seqidno.2)用正体aflash1表示。
如本文所用,“分离的”是指物质从其原始环境中分离出来(如果是天然的物质,原始环境即是天然环境)。如活体细胞内的天然状态下的多聚核苷酸和多肽是没有分离纯化的,但同样的多聚核苷酸或多肽如从天然状态中同存在的其他物质中分开,则为分离纯化的。
如本文所用,所述的“含有”,“具有”或“包括”包括了“包含”、“主要由……构成”、“基本上由……构成”、和“由……构成”;“主要由……构成”、“基本上由……构成”和“由……构成”属于“含有”、“具有”或“包括”的下位概念。
本发明提供了一种黄曲霉的无毒株。该无毒株不表达aflash1基因。其中,所述的黄曲霉无毒株可以用于研究aflash1基因不表达对于黄曲霉中其它基因表达情况的影响,以及研究aflash1基因不表达后黄曲霉的产孢、产菌核、产毒和侵染宿主等致病性情况。
aflash1基因不表达的菌株可以通过各种基因抑制、基因沉默、基因敲除等技术来构建。例如,可以通过基于同源重组的基因敲除技术来将aflash1基因从染色体上敲除,从而使得aflash1基因缺失;可以针对aflash1基因设计干扰性rna或反义核苷酸来使aflash1基因表达抑制或沉默。
作为本发明的一具体实施方式,一种使得aflash1基因缺失的方法是基因敲除技术,所述的aflash1的基因具有seqidno.1所示的核苷酸序列,aflash1蛋白具有seqidno.2所示的氨基酸序列。只要aflash1基因片段在被敲除后可导致aflash1蛋白不表达,丧失aflash1蛋白介导的组蛋白甲基化活性。
在本发明的一具体实施方式中,本发明人通过搜索ncbigenebank数据库,利用生物信息学比较分析,在黄曲霉的基因组序列中发现一个功能未知的新基因-假定的组蛋白赖氨酸甲基转移酶-ash1基因(putativehistone-lysinen-methyltransferase(ash1),该基因编码的产物与烟曲霉的ash1蛋白具有较高的同源性,因此命名为黄曲霉aflash1基因。本发明人体外构建aflash1基因敲除片段,通过同源重组的方法,把黄曲霉染色体aflash1基因中的2.5kbdna同源片段用烟曲霉pyrg基因片段替换,从而敲除染色体上的aflash1基因。
本黄曲霉aflash1基因敲除株在人工培养基(yes培养基)上孢子数量增加,但是在玉米粒等有机体上的产孢子量则显著下降;不产生菌核,使黄曲霉的抗逆境能力显著下降;不产生黄曲霉毒素afb1,afb2等黄曲霉毒素(或产量极低),对生物体的致病力显著下降。在种子侵染实验中,⊿aflash1菌株定殖在玉米子叶上的长出白色菌丝的时间比wt菌株显著要晚,产生的孢子明显减少(在自然界再次扩散的能力极低),tlc分析没有检测到黄曲霉毒素afb1,afb2等黄曲霉毒素的产生。这些结果表明aflash1调控黄曲霉的产孢、产菌核,和产毒,同时也影响该菌的致病性。
鉴于⊿aflash1菌株的第一代(产自人工培养基的高密度孢子,是野生型菌株的3倍以上)能高效地与野生型黄曲霉争夺有限的生态位,而⊿aflash1菌株本生在自然界中(在有机生物体上)传代能力极弱(产孢子量极低,不产生菌核),不产生黄曲霉毒素(或产度量低于可检测水平),致病力弱,本发明利用⊿aflash1菌株在人工培养基(yes培养基)上产生的大量孢子,将其释放到受野生黄曲霉污染的区域。利用⊿aflash1株的不产菌核,无毒,在有机体上低产孢,低致病力的特性,通过大量⊿aflash1菌株的分生孢子与野生产毒黄曲霉菌株在黄曲霉易感区域争夺有限的生态位,有效降低野生产毒黄曲霉菌株的密度,从而有效控制和降低作物受到的黄曲霉感染,黄曲霉毒素污染及其引发的曲霉病感染。
实施例1、黄曲霉中aflash1基因的敲除
为了研究黄曲霉aflash1基因在黄曲霉形态发生和致病性中的功能,首先在黄曲霉中敲除aflash1基因。
图1a显示了基因敲除的策略。体外构建aflash1基因敲除片段,通过同源重组的方法,把染色体中2.5kb的aflash1基因片段用1.9kb的烟曲霉pyrg替换,从而敲除染色体上的aflash1基因。
具体方法如下:
利用5’引物cagtaacgatcctcgcaattactccatg(p1);和3’引物tcgtgcgtgtgcttattgaggtct(p2);从黄曲霉pts菌株基因组dna中用pcr的方法扩增约1.2kb的上游片段;利用5’引物ttttgggctcgttggctacataggaatac(p3);和3’引物agcctctccaggaggatgaacctg(p4);从黄曲霉pts菌株基因组dna中用pcr的方法扩增约1.2kb的下游片段;利用5’引物agacctcaataagcacacgcacgagcctcaaacaatgctcttcaccc(p5)和3’引物gtattcctatgtagccaacgagcccaaaagtctgagaggaggcactgatgc(p6)从烟曲霉基因组dna中用pcr的方法扩增约1.9kb的pyrg基因片段;将上述三个片段用融合pcr法(利用引物p1和p4)连接到一起构建aflash1基因敲除片段,导入黄曲霉pts菌株,在不含尿嘧啶和尿苷的培养基上筛选阳性转化子。通过敲除片段上、下游各1.2kb的两个同源片段与染色体上aflash1基因的上、下游同源片段的同源重组,把染色体上的aflash1基因给替换掉,从而敲除染色体上的aflash1基因(图1a)。本项目进一步利用qrt-pcr验证了这些菌株中aflash1基因的转录水平(图1b),结果表明aflash1基因在⊿aflash1菌株中完全不表达,而在wt和aflash1基因回补菌株中aflash1基因都正常表达,⊿aflash1菌株中已经成功移除aflash1基因。
实施例2、黄曲霉毒素提取及分析
在yes液体培养基中接种各菌株的黄曲霉孢子(浓度为106个/ml),29℃黑暗静置培养6天,然后从中吸取500µl液体培养基,并加入等体积氯仿,振荡混匀,高速离心5min,吸取有机相液体,干燥后用200µl氯仿重新溶解并取10µl点样,进行薄层层析(tlc)分析。图2显示敲除aflash1基因后,⊿aflash1菌株不产afb1,afb2,afg1,和afg2。
而野生型黄曲霉菌株则可大量合成黄曲霉毒素;把aflash1基因回补后(com-ash1菌株)黄曲霉毒素的合成水平回复了。进一步的q-pcr分析发现,敲除aflash1基因后aflr,afls,aflc,afld,aflk基因,和aflq等黄曲霉毒素合成相关产毒基因的表达量显著下降,最终导致⊿aflash1菌株产毒受抑制。
实施例3、aflash1基因的敲除对黄曲霉菌产孢的影响
将wt,⊿aflash1和com-ash1黄曲霉菌株在yes固体培养基上37℃培养4天后,观察各个菌株的产孢情况。结果发现在人工培养基上⊿aflash1菌株产生了大量绿色的孢子,数据统计分析也发现⊿aflash1菌株产生的孢子数量是wt三倍以上,(图3a,b,c)。
该结果表明,aflash1基因的缺失导致黄曲霉在人工培养基上产孢量显著增加。
实施例4、aflash1基因的敲除对黄曲霉菌产菌核的影响
将wt,⊿aflash1和com-ash1黄曲霉菌株在ypd固体培养基上37℃暗培养6天后,用解剖显微镜对形成的菌核计数。图4显示⊿aflash1菌株的菌核形成被彻底抑制。结果表明aflash1基因正调控菌核的形成,aflash1基因的缺失导致⊿aflash1菌株失去形成菌核的能力,使其抵抗高温等不良环境的能力降低。
实施例5、aflash1基因的敲除对黄曲霉致病力的影响
1.aflash1基因对黄曲霉菌侵染玉米粒的影响
用牙签去掉玉米粒的胚芽,然后将玉米粒置于0.05﹪次氯酸钠中浸泡3min,转至无菌水中漂洗30s,70﹪乙醇浸泡5s,再用无菌水中漂洗1min,沥干水后分别用20ml各菌株的孢子溶液(终浓度105个/ml)浸泡玉米粒,空白对照用无菌水浸泡,50rpm,30min,每个培养皿放置重量相近的15颗玉米粒,培养皿中铺三层润湿的滤纸,29℃培养7天;然后将侵染后的玉米粒放入装有10ml0.01﹪吐温20的孢子洗脱液中,涡旋振荡1min,取1ml用于孢子计数,其余液加入5ml丙酮150rpm,10min,室温静置5min,加入5ml氯仿,150rpm,10min,室温静置10min,涡旋混匀,2000rpm离心15min,收集下层有机相液体,干燥后用5ml0.1mnacl甲醇:水(55:45)和2.5ml己烷涡旋混匀1min,2000rpm离心15min,收集己烷层,去掉油脂层(中间),剩余的水相按上述方法再抽提一次,合并收集样,干燥,用500μl氯仿重溶,取10μl点tlc板。
结果显示在生物体上和野生型和互补黄曲霉菌株相比,⊿aflash1菌株的产孢量被显著,通过tlc分析,显示⊿aflash1菌株在生物体上的黄曲霉毒素形成也被抑制了(图5)。
2.在大田中aflash1基因对黄曲霉菌侵染花生小叶片的影响
为了检验大田中⊿aflash1菌株对农作物的致病力,本项目用刀片划伤花生小叶,然后分别接种2µl(1×106个/ml)wt和⊿aflash1黄曲霉菌株的孢子悬液。4天后观察,发现培养4天后观察发现接种了wt黄曲霉孢子的花生小叶的叶片出现明显病斑,而接种⊿aflash1菌株的小叶没有明显病斑出现。结果表明敲除aflash1基因后,黄曲霉菌在大田环境下对作物的致病力显著下降。
实施例6、aflash1是黄曲霉组蛋白甲基转移酶
黄曲霉菌丝加入液氮冷冻后迅速研磨成粉末,按每110mg菌丝粉末加入550μl的ripa裂解液(beyotime),加入pmsf至终浓度1mm,混匀后冰上裂解30min,4℃高速离心10min,收集上清并测定蛋白浓度,进行sds-page电泳,用湿法将蛋白条带转移到尼龙膜上,用含5%脱脂奶粉的tbst缓冲液rt封闭1小时,分别用抗h3k9和h3k36甲基化的单克隆抗体作为一抗4℃杂交过夜,用tbst室温洗膜3次,每次5分钟,用相应的二抗于室温结合1小时,再用tbst洗膜3次,每次5分钟,用ecl试剂显色。结果(图7)显示:aflash1蛋白催化h3k9和h3k36的二甲基化。因此本项目确定在黄曲霉中aflash1蛋白是组蛋白甲基转移酶。
本项目通过同源重组方法扩增aflash1基因的5’utr和3’utr片段,然后通过融合pcr和来自烟曲霉的pyrg基因片段融合到一起。利用该融合片段(5’utr-pyrg-3’utr)转化黄曲霉的原生质体,获得敲除aflash1基因的黄曲霉⊿aflash1菌株。用q-pcr分析野生型菌株(wt),⊿aflash1菌株,和aflash1基因回补菌株中aflash1基因的表达情况发现aflash1基因在⊿aflash1菌株中完全不表达;而在wt和aflash1基因回补菌株中aflash1基因都正常表达。
sequencelisting
<110>福建农林大学
<120>一种黄曲霉无毒株及其防治黄曲霉污染的应用
<130>8
<160>8
<170>patentinversion3.3
<210>1
<211>2533
<212>dna
<213>aflash1全长基因序列
<400>1
atggcctttgagtattcagaacaactcctcacccccgagaacagtcgctcggagacttcc60
agcaacaatgaaaacgcgtctaccgaggaaaagccgcctgtacggcgaagatctacacgt120
gtaacacgcgcctctttacggggggaggcacaactcgatggcgacatggagatagataac180
caagggactttgtctacggctggcgaaaataaccctgtggtctcgggtgagactctagtc240
gataaagctgaaggcggaaagaggtcccaggcctcccacctgcgccacagcatagcggtc300
atggagtcctggagtgaagcaacactggcacaaggcaatatggagactgaaggggaccat360
ccattggccccagatactccagtttctaagtcctcgcaagagctgcaggccagtgacatg420
ggcacgtcactgcaacaacgcactttacggaagcgcgtagagcggatcctgactgaagaa480
gggcacggcaataagggcaaagttactgtaacggctaaagaaatcaagtcgccagtcaga540
cgctcgtctcgtttgagtctgttagagaaggcttcagaccttgttggccgggctagcagc600
gttttgggaaaacgctcgcgtgacgtgatgggaaagggtaaagaacttggccggcgggct660
agtttacgacctcgaactactgcacccaaggaggaaccgaccaaggctgccagcgaggct720
cctgcggcgaaaaagaggcgtgtttctgaaagcgatctacctgtcaaaatccaggaaaat780
gaagaagccgtgcaggaggctcccaagcctgtggttcggtccaggacaaagcgctggtta840
gctcatggattgtacacaggccaggaacacacagagtctcggcccctgcagagtaggagc900
agaaatgcaaagcgcaaaagtcaaggtcccacacaacgcagactgcttcctatgccgatg960
tttgcaggagatcgactcttgaagcagggaagagatttccagctccccttcgatatcttt1020
tctccgctcccttctggtcagcccaaacccaatgagtggagaaagacaaacaagagtatg1080
attgccagcagaaactcgcgatgatggctaacttgtggaatagatgtctttgtcggggaa1140
gcatccagtatatggcgagcaaacaagccattggagctatccaagtgcatgtgtgcagaa1200
gagactggctgtgacgaggaatgtcagaaccgttacatgttctacgaatgtgatgacacc1260
aattgtggtgtgggccccgagtgtggaaatcggaatttcgaggaactgaaacagagaacc1320
aaggctggcggaaaatacaatattggagtcgaagtcatcaagacggaagaccgaggatac1380
ggtgttcgtagcaaccgcacattcgagcctaaccaggtcattgtggagtacacaggggaa1440
atcataacgcaagcggaatgtgagaaacgaatgaggacgatttataagaataacgaggta1500
tgcatcctttgctcacgctttctctggtggacgggcgacttattgaccgctatcagtgct1560
actacctaatgtactttgatcagaacatgatcatcgatgctaccaggggctctatcgctc1620
gtttcgtaaaccattcttgcgagcccaactgtcggatggagaaatggacagtcgcgggga1680
agccgcgtatggcgctttttgctggtgatcgtggaataatgaccggggacgaactgacct1740
atgactataattttgagtaagggtaccttaccgtgtaaatcgggattgtcgatactgacc1800
tgtttcttgcagcccatactctcagaaaaacgttcagcaatgccggtgtggatcggacag1860
atgccgcggtatcctgggaccacggccgcgggagaaagagcaacgttcaaaggagaaaga1920
actgagggctgaaaacgagaagaaatctagttcgaagaataacaatgaaaaggccagcat1980
caccaagcagaaggtattgaatggatcgacttcgcgcgtcaataagcgacagcttcttgg2040
ttcgaagtccatcaagtctggtgtgaagaaagttgtttctaaagcacgcgcttctgcttc2100
taaagcaactacagcatcgagaactagcttgaaaacaactgtcagcacaaaaacctccaa2160
gacgacaaacaagaaagccacggcaactccaacacgacggcaggtgaagaaagataacac2220
caaatccaagaaagaagttaagctgccaaaggtcaaaaccacaaagactaaagccagagc2280
cccggcgagtgccaggaaaccggcacagaaaaataaaacccagtcaacgccattgacatc2340
taaactaagccgtccatctgagaagactaaagcgaaaatcctccaggctgctaaagggac2400
gaacgctcgcagacgtacagtgaaaaaggaagataccaagcccaaaagtcctaccaagcg2460
tacttctaaagccaaagagaccagtcctcgggggaaggcagcaagcaaggctgccaagaa2520
caccaaggcataa2533
<210>2
<211>789
<212>prt
<213>aflash1全长蛋白序列
<400>2
metalapheglutyrsergluglnleuleuthrprogluasnserarg
151015
sergluthrserserasnasngluasnalaserthrgluglulyspro
202530
provalargargargserthrargvalthrargalaserleuarggly
354045
glualaglnleuaspglyaspmetgluileaspasnglnglythrleu
505560
serthralaglygluasnasnprovalvalserglygluthrleuval
65707580
asplysalagluglyglylysargserglnalaserhisleuarghis
859095
serilealavalmetglusertrpserglualathrleualaglngly
100105110
asnmetgluthrgluglyasphisproleualaproaspthrproval
115120125
serlysserserglngluleuglnalaseraspmetglythrserleu
130135140
glnglnargthrleuarglysargvalgluargileleuthrgluglu
145150155160
glyhisglyasnlysglylysvalthrvalthralalysgluilelys
165170175
serprovalargargserserargleuserleuleuglulysalaser
180185190
aspleuvalglyargalaserservalleuglylysargserargasp
195200205
valmetglylysglylysgluleuglyargargalaserleuargpro
210215220
argthrthralaprolysglugluprothrlysalaalasergluala
225230235240
proalaalalyslysargargvalsergluseraspleuprovallys
245250255
ileglngluasngluglualavalglnglualaprolysprovalval
260265270
argserargthrlysargtrpleualahisglyleutyrthrglygln
275280285
gluhisthrgluserargproleuglnserargserargasnalalys
290295300
arglysserglnglyprothrglnargargleuleuprometpromet
305310315320
phealaglyaspargleuleulysglnglyargasppheglnleupro
325330335
pheaspilepheserproleuproserglyglnprolysproasnglu
340345350
trparglysthrasnlysasnvalphevalglyglualaserserile
355360365
trpargalaasnlysproleugluleuserlyscysmetcysalaglu
370375380
gluthrglycysaspgluglucysglnasnargtyrmetphetyrglu
385390395400
cysaspaspthrasncysglyvalglyproglucysglyasnargasn
405410415
pheglugluleulysglnargthrlysalaglyglylystyrasnile
420425430
glyvalgluvalilelysthrgluaspargglytyrglyvalargser
435440445
asnargthrphegluproasnglnvalilevalglutyrthrglyglu
450455460
ileilethrglnalaglucysglulysargmetargthriletyrlys
465470475480
asnasnglucystyrtyrleumettyrpheaspglnasnmetileile
485490495
aspalathrargglyserilealaargphevalasnhissercysglu
500505510
proasncysargmetglulystrpthrvalalaglylysproargmet
515520525
alaleuphealaglyaspargglyilemetthrglyaspgluleuthr
530535540
tyrasptyrasnpheaspprotyrserglnlysasnvalglnglncys
545550555560
argcysglyseraspargcysargglyileleuglyproargproarg
565570575
glulysgluglnargserlysglulysgluleuargalagluasnglu
580585590
lyslysserserserlysasnasnasnglulysalaserilethrlys
595600605
glnlysvalleuasnglyserthrserargvalasnlysargglnleu
610615620
leuglyserlysserilelysserglyvallyslysvalvalserlys
625630635640
alaargalaseralaserlysalathrthralaserargthrserleu
645650655
lysthrthrvalserthrlysthrserlysthrthrasnlyslysala
660665670
thralathrprothrargargglnvallyslysaspasnthrlysser
675680685
lyslysgluvallysleuprolysvallysthrthrlysthrlysala
690695700
argalaproalaseralaarglysproalaglnlysasnlysthrgln
705710715720
serthrproleuthrserlysleuserargproserglulysthrlys
725730735
alalysileleuglnalaalalysglythrasnalaargargargthr
740745750
vallyslysgluaspthrlysprolysserprothrlysargthrser
755760765
lysalalysgluthrserproargglylysalaalaserlysalaala
770775780
lysasnthrlysala
785
<210>3
<211>28
<212>dna
<213>人工序列
<400>3
cagtaacgatcctcgcaattactccatg28
<210>4
<211>24
<212>dna
<213>人工序列
<400>4
tcgtgcgtgtgcttattgaggtct24
<210>5
<211>29
<212>dna
<213>人工序列
<400>5
ttttgggctcgttggctacataggaatac29
<210>6
<211>24
<212>dna
<213>人工序列
<400>6
agcctctccaggaggatgaacctg24
<210>7
<211>47
<212>dna
<213>人工序列
<400>7
agacctcaataagcacacgcacgagcctcaaacaatgctcttcaccc47
<210>8
<211>51
<212>dna
<213>人工序列
<400>8
gtattcctatgtagccaacgagcccaaaagtctgagaggaggcactgatgc51
- 还没有人留言评论。精彩留言会获得点赞!