酸敏感离子通道调控剂的用途的制作方法
本发明涉及医药技术领域,具体涉及一种酸敏感离子通道调控剂的新用途。
背景技术:
酸敏感离子通道(acid-sensingionchannel,asic)家族包含四条基因编码的六种亚型,即asic1a、1b、2a、2b、3和4等。这些质子门控的通道由三个相同或者不同亚基组成,不同组合形成的离子通道表现出差异的ph敏感性、离子选择性和药理学特性。在中枢神经系统,包含asic1a的通道是最主要的asic成员,asic1a基因缺失的神经元无法表现出酸诱导的电流。asic1a的重要性在啮齿类动物模型上已被充分研究,如缺血性神经元死亡、慢性痛、癫痫终止和神经退行性疾病等。尽管asic1a的神经元表达及其对于病理意义的研究已经取得了重要进展,然而,asic1a在脑神经元中的生理功能仍旧不清楚。
越来越多的证据支持asic1a在突触传递和可塑性中发挥关键性作用。基于asic1a的突触后定位,人们认为asic1a在正常突触传递过程中可被突触活动释放的质子所激活。作为司职于突触内质子受体的asic1a通道,其在调控海马和杏仁核高频刺激或者theta串刺激(thetaburststimulation,tbs)诱导的突触长时程增强(long-termpotentiation,ltp)可塑性过程中发挥作用,并因而贡献于恐惧学习的过程。asic1a在海马区促进树突棘的密度,但是在伏隔核抑制树突棘密度,改变兴奋性突触受体的功能,以及限制可卡因引起的突触可塑性,这些进展提示asic1a在调控突触可塑性和行为的适应性方面可能发挥更加复杂的作用。
asic1a功能的复杂性可能反映在其在不同脑区参与不同形式的可塑性和行为调节。asic1a在岛叶皮层神经元高表达,但是其通道功能并不清楚。岛叶皮层参与了诸多认知和情感调控功能,包括感觉整合、慢性痛、情感处理和味觉识别记忆等。该脑区最为熟知的功能是其在味觉学习中的作用。尤其是,岛叶皮层在一种称之为条件性味觉厌恶(conditionedtasteaversion,cta)的负性情绪驱动的味觉学习中发挥重要功能。在条件性味觉厌恶的学习过程中,学习主体将一种全新味觉(条件刺激,conditionedstimulus,cs)与随后感受到的内脏不适(非条件刺激,unconditionedstimulus,us)建立联系,从而对这种特定味道产生厌恶情绪性记忆。在突触水平,岛叶皮层的长时程突触增强贡献于条件性味觉厌恶记忆的获取。然而,介导条件性味觉厌恶记忆消退的分子和突触机制并不清楚。
厌食症,又称神经性厌食,是一种多见于青少年女性的进食行为异常,特征为故意限制饮食,使体重降至明显低于正常标准,常过分担心发胖,甚至已经明显消瘦仍自认为太胖,即使医生进行解释也无效。目前仍缺乏有效的厌食症治疗药物。
许多有毒瘾或烟瘾或酒瘾的人,常因忍受不了戒毒或戒烟或戒酒后身体上的痛苦,又重新吸毒或吸烟或饮酒。因此,神经系统的治疗药物对于成功戒毒或戒烟或戒酒非常有必要。
焦虑症也是一种神经系统的疾病,其全球发病率为7.3%,而个人一生中的患病率高达11.6%。焦虑症患者在特定环境(如社交环境或陌生环境)中往往表现出过度的恐惧、焦虑或对感知到的危险做过度回避。而这些反应远远超过了实际危险所引起的正常恐惧或焦虑反应。焦虑症的发生可显著降低患者的生活质量,并给患者家庭和整个社会带来了极大的经济负担。然而,目前用于治疗或有效缓解焦虑症的药物还十分匮乏。
技术实现要素:
本发明要解决目前缺乏厌食症、焦虑症、毒瘾、烟瘾等精神障碍疾病治疗药物的技术问题,提供一种酸敏感离子通道调控剂的用途,用于治疗或缓解厌食症、焦虑症、毒瘾、烟瘾等精神障碍疾病。
为了解决上述技术问题,本发明通过如下技术方案实现:
在本发明的一个方面,提供了一种酸敏感离子通道调控剂在制备治疗精神障碍疾病的药物中的用途,所述酸敏感离子通道为包含asic1a亚基的酸敏感离子通道。
所述调控剂包括酸敏感离子通道促进剂或抑制剂。
所述酸敏感离子通道促进剂包括任何增加asic1a亚基蛋白表达和/或活性的化合物或重组载体。优选的,酸敏感离子通道促进剂化合物包括多肽mittx-α/β、和/或精胺。
所述酸敏感离子通道抑制剂包括任何特异性抑制asic1a通道相关蛋白表达和/或活性的化合物或重组载体。优选的,酸敏感离子通道抑制剂化合物包括多肽pctx1、阿米洛利(amiloride)、脒a-317567、萘莫司他(nafamostatmesilate)、非甾体类抗炎药(non-steroidalanti-inflammatorydrugs,nsaids)、compound5b(2-oxo-2h-chromene-3-carboxamidine衍生物)、碱性中和剂、或其组合。所述碱性中和剂包括碳酸氢钠、氯化铵、碱金属盐、碱土金属盐、或其组合。
所述酸敏感离子通道促进剂用于治疗的精神障碍疾病包括:厌食症、焦虑症、成瘾性疾病、神经性贪食及肥胖、和其他任何伴有异常顽固病理性记忆的神经系统疾病。其中,成瘾性疾病包括毒瘾、烟瘾、酒瘾等。
采用本发明的酸敏感离子通道促进剂,并辅助反复消退训练,可明显改善厌食症或成瘾性疾病患者症状;采用本发明的酸敏感离子通道促进剂,并辅助暴露疗法,可有效治疗焦虑症。
所述酸敏感离子通道抑制剂用于治疗的精神障碍疾病包括阿尔茨海默病和其他任何伴有认知障碍和记忆力损害的神经系统疾病。
采用本发明的酸敏感离子通道抑制剂,并辅助认知训练,可改善阿尔茨海默病患者症状。
在本发明中,术语“酸敏感离子通道”(acid-sensingionchannels,asics)是指一类广泛存在于细胞膜上的通透阳离子的蛋白复合体,属于上皮钠通道/退化素蛋白超家族,在感受体液ph值和调控痛觉、机械感受等多项生理机能方面有着重要作用。
分子克隆表明,asic有四条基因编码的至少六种亚基(asic1a,1b,2a,2b,3和4),这些亚基形成同或异三聚体功能复合物。asic亚基表达和分布的不同以及亚基组成和通道门控的变化,使得通道的功能呈现出多样性。
asic1a在中枢和外周神经系统都有分布,主要参与突触传递和可塑性。asic1a的功能异常跟多种神经疾病如缺血性细胞死亡、癫痫、神经退行性疾病等。本发明所指的asic1a通道,是指主要由asic1a亚基组成的同聚体或异聚体。
在本发明中,术语“酸敏感离子通道调控剂”指能够调控酸敏感离子通道的物质,所述调控剂可以是小分子化合物,也可以是大分子化合物。
在本发明中,所述的调控剂包括促进剂(或激动剂)、或抑制剂(或拮抗剂)。
代表性的酸敏感离子通道调控剂包括多肽pctx1、mittx-α/β、精胺、阿米洛利(amiloride)、脒a-317567、萘莫司他(nafamostatmesilate)、非甾体类抗炎药(non-steroidalanti-inflammatorydrugs,nsaids)、compound5b(2-oxo-2h-chromene-3-carboxamidine衍生物)等。
如本发明实施例所用,多肽mittx-α/β,为一种源自于texascoralsnake的毒素,能够直接激活asic1a通道。
如本发明实施例所用,狼蛛毒素pctx1,能有效抑制asic1a通道。
在本发明中,术语“厌食症”是指个体通过节食等手段,有意造成并维持体重明显低于正常标准为特征的一种进食精神障碍,其主要特征是强烈害怕体重增加和发胖,对体重和体型极度关注,体重显著减轻的同时,常有营养不良、代谢和内分泌紊乱,严重患者可因极度营养不良而出现恶病质状态、机体衰竭甚至危及生命,5%~15%的患者最后死于心脏并发症、多器官功能衰竭、继发感染等。
在本发明中,术语“焦虑症”是指患者在特定环境(如社交环境或陌生环境)中往往表现出过度的恐惧、焦虑或对感知到的危险做过度回避。而这些反应远远超过了实际危险所引起的正常恐惧或焦虑反应。焦虑症包括:创伤后应激障碍、分离性焦虑障碍、选择性缄默症、特定恐惧症、社交恐惧症、恐慌症、广场恐怖症、广泛性焦虑症、与另一疾病相关联的焦虑症、药物诱发型焦虑症、疑病症等。
在本发明的另一方面,还提供了一种用于治疗精神障碍疾病的药物组合物,包含安全有效量的酸敏感离子通道调控剂、和药学上可接受的载体,所述酸敏感离子通道为包含asic1a亚基的酸敏感离子通道。
上述可接受的载体是无毒的、能辅助施用并且对酸敏感离子通道调控剂的治疗效果无不利影响。此类载体可以是本领域的技术人员通常能得到的任何固体赋形剂、液体赋形剂、半固体赋形剂或者在气雾剂组合物中可以是气体赋形剂。
本发明药物组合物的各种剂型可以按照药学领域的常规方法制备。例如使该酸敏感离子通道调控剂与一种或者多种载体混合,然后将其制成所需的剂型,如片剂、药丸、胶囊、半固体、粉末、缓释剂型、溶液、混悬液、配剂、气雾剂等。
在本发明的另一方面,还提供了一种筛选治疗精神障碍疾病的候选药物的方法,包括以下步骤:
检测待测化合物对含asic1a亚基酸敏感离子通道的影响,选出对该酸敏感离子通道有调控作用的待测化合物,作为初筛化合物;
检测所述初筛化合物对精神障碍疾病的治疗或缓解效果,选出具有疗效的化合物,作为候选药物。
优选的,获得初筛化合物的检测步骤包括:在待测化合物存在下,培养细胞,并检测该细胞中酸敏感离子通道1a蛋白的表达量、活性和/或asic样电流,并且在不存在所述待测化合物且其他条件相同的对照组中,检测相同细胞中酸敏感离子通道1a蛋白的表达量、活性和/或asic样电流,并进行比较;如果实验组的检测值vt显著高于或显著低于对照组的检测值vc,则表明所述待测化合物为对所述酸敏感离子通道有调控作用的待测化合物。
本发明的酸敏感离子通道调控剂,通过促进或抑制含asic1a亚基酸敏感离子通道的活性,进而促进或抑制条件性记忆的消退,为治疗或缓解厌食症、焦虑症、毒瘾、烟瘾等精神障碍疾病提供新的药物或治疗靶点,应用前景非常广阔。
附图说明
下面结合附图和具体实施方式对本发明作进一步详细的说明。
图1是本发明实施例1的asic1a在野生型和asic1a基因敲除小鼠岛叶皮层的表达结果图;
图2是本发明实施例2的asic1a为低频刺激诱导的长时程突触抑制(ltd)所必需的实验结果图;
图3是本发明实施例3在野生型小鼠干扰细胞外ph动态变化过程阻断低频刺激-长时程突触抑制的实验结果图;
图4是本发明实施例4在asic1a基因敲除小鼠再表达asic1a解救低频刺激-长时程突触抑制的实验结果图;
图5是本发明实施例5的asic1a为岛叶皮层dhpg诱导的长时程突触抑制所必需的实验结果图;
图6是本发明实施例6的asic1a并不贡献于岛叶皮层长时程突触增强(ltp)诱导的实验结果图;
图7是本发明实施例7的岛叶皮层长时程突触抑制选择性被条件性味觉厌恶消退过程所抵消的实验结果图;
图8是本发明实施例8的asic1a的缺失选择性损害条件性味觉厌恶记忆消退的实验结果图;
图9是本发明实施例9的岛叶皮层的功能性asic1a对于条件性味觉厌恶消退具有关键作用的实验结果图;
图10是本发明实施例10的岛叶皮层长时程突触抑制对于条件性味觉厌恶消退是必需的实验结果图;
图11是本发明实施例11的药理学阻断或激活腹侧海马asic1a分别抑制和促进恐惧消退的实验结果图;
图12是本发明实施例12的腹侧海马条件性敲除或过表达asic1a分别抑制和促进恐惧消退的实验结果图。
具体实施方式
本发明利用多电极阵列脑片记录的手段鉴定了asic1a在岛叶皮层长时程突触抑制(long-termdepression,ltd)可塑性中发挥至关重要的作用,同时利用行为学测定的手段分析了其在条件性味觉厌恶学习及其记忆消退中的贡献。跟已知asic1a参与其他脑区的长时程突触增强(long-termpotentiation,ltp)并进而促进联合型学习与记忆的贡献不同,本发明中的asic1a在岛叶皮层是长时程突触抑制(ltd)的关键调控因素,该机制对于已建立味觉厌恶记忆的消退非常重要。
通用方法和材料
1.动物
动物护理和实验步骤得到了上海交通大学医学院动物伦理委员会的批准。为了降低实验的变异性,采用年龄相当的同窝对照小鼠进行实验。所有的行为学测定都是在清醒的、自由活动的、雄性、2–3月龄、c57bl/6j背景的小鼠上进行。常规的asic1a全身基因敲除小鼠为美国爱荷华大学(universityofiowa)michaelj.welsh教授馈赠;条件性asic1a基因敲除小鼠(asic1aflox/flox)由台湾阳明大学cheng-changlien教授提供;在前脑兴奋性神经元中高表达cre重组酶的工具鼠——(camkii-cre)由美国乔治亚摄政大学(georgiaregentsuniversity)joez.tsien教授馈赠。成年c57bl/6j小鼠购于上海斯莱克公司。动物饲养于以12小时为周期的昼夜交替环境内(光照时间:7:00-19:00),饲养室温度恒定于21℃,湿度维持于50-60%,动物在非实验期间可自由摄食饮水。随机选取动物进行双盲实验。在所有行为学实验中,实验者提前至少两周将小鼠转移至行为学饲养室内饲养,实验前1小时将小鼠放置于行为检测室内适应环境,以充分减少环境变化对行为学结果产生的影响。所有的行为学测试都在对基因型或药物处理双盲的条件下开展和评价。
2.条件性味觉厌恶消退模型
条件性味觉厌恶记忆模型的构建是对雄性小鼠进行定时饮水连续训练6~8天,以使其适应每天早上同一个时间点饮水。在条件性味觉厌恶记忆建立当天,给予小鼠以0.5%糖精钠溶液替换水,饮用时间为30分钟,停止饮用后40分钟,对实验小鼠进行腹腔注射氯化锂(licl,0.3摩尔/升)。其中糖精钠溶液为条件刺激,腹腔注射的licl为非条件刺激。味觉厌恶记忆的强度以厌恶指数(aversiveindex)为指标。厌恶指数=饮水量/(饮水量+饮糖量)×100%。在条件性味觉厌恶记忆建立后,通过反复测试,即为味觉厌恶消退学习模型。
3.焦虑症动物模型和焦虑症相关的行为范式
研究恐惧行为最常用的动物模型是恐惧条件反射(fearconditioning),即将无害刺激(条件性刺激,conditionedstimulus,cs)与有害刺激(非条件性刺激,unconditionedstimulus,us)反复偶联,从而使动物对cs产生恐惧反应(条件性反应,conditionalresponse,cr),其表现为躯体和自主神经的反应,包括僵立反应(freezing)、心跳加快、血压升高、痛觉丧失等。其后,若反复只给予动物cs而不给予us,动物对cs的条件性恐惧反应会逐渐降低,此过程称为恐惧消退(fearextinction)。
本发明的行为学测定在清醒的、自由活动的c57bl/6j小鼠或asic1aflox转基因小鼠(均为2–3个月龄,c57bl/6j遗传背景)上进行。采用的焦虑症动物模型是线索依赖的恐惧条件反射。实验开始前对动物进行三天的抚触和实验笼适应。正式实验在连续三天内完成:第一天让动物在a环境(实验笼一面透明,另外三面贴黑白格子相间的壁纸,底部是可导电的铁栅栏,并喷洒75%酒精)中进行恐惧学习,实验中先让动物适应实验笼3分钟,其后给动物30秒、76分贝、4000赫兹的声音(无害刺激或条件性刺激,cs),并在声音结束前2秒给予动物0.5毫安的足底电击(有害刺激或非条件性刺激,us),共给5次联合的声音和足底电击,每次声音间隔的时间20-120s不等;第二天让动物在b环境(实验笼一面透明,另外三面贴灰色的壁纸,底部铺光滑的塑料板,并喷洒4%乙酸)中进行消退学习,实验中连续给动物20次cs,不给us;第三天在b环境中给动物8次cs,检测消退学习后动物对线索的恐惧反应。在三天的实验中通过检测cs期间动物僵立不动(freezing)的时间百分比来反映动物对cs的恐惧反应。恐惧反应的评价是通过分析动物僵立不动(freezing)的时间百分比实现。实施例中的行为学实验在对基因型或药物处理双盲的条件下开展和评价。
4.脑片制备和膜片钳记录
参考文献(kim,j.i.etal.(2011).pi3kgammaisrequiredfornmdareceptor-dependentlong-termdepressionandbehavioralflexibility.natneurosci14,1447-1454;liu,m.g.etal.(2013b).long-termdepressionofsynaptictransmissionintheadultmouseinsularcortexinvitro.eurjneurosci38,3128-3145;qiu,s.etal.(2013).anincreaseinsynapticnmdareceptorsintheinsularcortexcontributestoneuropathicpain.scisignal6,ra34)的方法,分别从不同处理的野生型和突变体小鼠进行脑片制备和膜片钳记录。
5.利用多电极阵列记录和分析系统研究突触可塑性
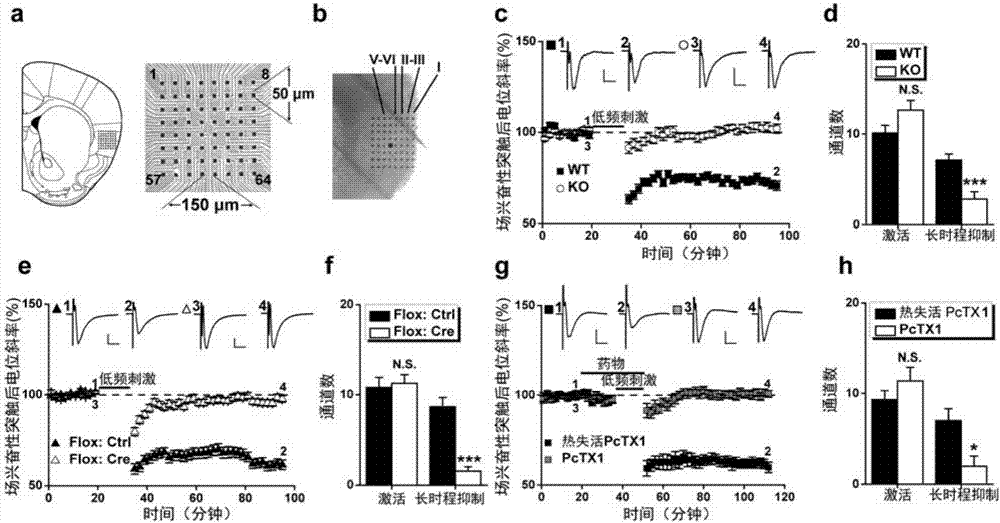
为研究成年小鼠岛叶皮层的突触可塑性,采用平面微电极阵列记录技术(med64多通道记录系统),在急性分离的岛叶脑片上进行多通道兴奋性突触后场电位(fieldexcitatorypostsynapticpotential,fepsp)记录和分析。采用7-10周龄的雄性c57/bl6j背景的野生型或突变体小鼠。麻醉断头后,迅速取出大脑并放入充氧的冰冻人工脑脊液中1-2分钟。然后在冰上修块,将脑组织块转移至冰冻的切片机中切取300μm的岛叶脑片2-3片。切好的岛叶脑片在26-30℃的人工脑脊液中孵育2小时后,选取一片岛叶脑片在显微镜下将其放于med64电极上并移动至指定位置,让电极的8×8阵列尽量完整地覆盖整个岛叶皮层。调好系统噪音,打开mobius操作软件开始基线记录。等基线反应稳定20-30分钟后,给予脑片不同的刺激模式以诱发不同类型的突触可塑性,包括ltp和ltd。根据不同的实验需要,ltp或ltd可记录1-3小时。med64系统采集的数据离线分析主要用mobius软件,着重计算所记录fepsp的斜率在给予不同的突触可塑性诱导参数前后的时程变化。
6.数据分析
所有的结果以平均值±标准误的形式进行表述。统计学比较采用非配对或配对的student’st检验或双向重复测量的方差分析方法。其中,p<0.05,p<0.01,和p<0.001被认为是有显著差异。
实施例1asic1a在小鼠岛叶皮层显著表达
通过逆转录-聚合酶链反应(reversetranscription-polymerasechainreaction,rt-pcr)和免疫印迹(westernblotting)实验确证asic1a在成年小鼠岛叶皮层的表达情况。相比于其他asic亚基,asic1a亚基的mrna在野生型岛叶皮层上最为丰富,但是在asic1a基因敲除小鼠上完全缺失(图1a,b)。在蛋白水平,asic1a在野生型小鼠的岛叶皮层表达丰富,跟前额叶皮层和杏仁核相当,明显高于海马,但是在asic1a基因敲除小鼠上完全缺失(图1c)。这些数据说明,asic1a在岛叶皮层具有显著的表达。
在图1中,(a)rt-pcr显示野生型和asic1a基因敲除小鼠岛叶皮层组织不同asic亚基的表达。(b)实时定量反转录pcr显示的mrna水平的定量统计。n=3。n.s.,利用非配对student’st-检验对野生型(wt)和asic1a基因敲除(asic1ako)两组进行比较后无统计学差异;***p<0.001,利用非配对student’st-检验进行比较后,表明野生型(wt)和asic1a基因敲除(asic1ako)两组间呈现出显著的统计学差异。###p<0.001,利用非配对student’st-检验对两组比较后,表明其他asic亚基分别与asic1a组间呈现出显著的统计学差异。(c)asic1a蛋白在岛叶皮层和其他脑区的水平比较。(上部),asic1a蛋白在前额叶、海马、杏仁核和岛叶皮层的代表性免疫印迹图片。(下部),asic1a蛋白相对量的统计分析。所示数据为相对光密度比值,以海马区数据进行标准化处理。n=4。*p<0.05,**p<0.01,利用配对student’st-检验对各个脑区分别与海马区进行比较后,表明显著的统计学差异。
实施例2asic1a为低频刺激诱导的长时程突触抑制(ltd)所必需
采用64通道多电极阵列记录系统分析多种突触可塑性模型。其中,64通道多电极阵列放置于如图2a,b所示的岛叶皮层脑片上。通过刺激嘴侧(rostral)接近于胼胝体水平的岛叶脑片的深层区域(第v-vi层,如图2b所示),从而记录多电极覆盖整个区域的突触反应。在刺激位点进行电流注入引起64通道电极阵列中约10个通道呈现出显著的场兴奋性突触后电位(fieldexcitatorypostsynapticpotentials,fepsps),将之定义为激活通道(activatedchannels)。跟过去的报道(liu,m.g.etal.(2013b).long-termdepressionofsynaptictransmissionintheadultmouseinsularcortexinvitro.eurjneurosci38,3128-3145)一致,低频刺激(long-frequencystimulation,lfs)在野生型小鼠岛叶皮层脑片大部分激活通道中诱导出场兴奋性突触后电位的降低(图2c),将其定义为低频刺激-长时程突触抑制。这种低频刺激-长时程突触抑制能够被nmda受体的拮抗剂——d-ap5所抑制,因此也称之为nmda受体依赖的长时程突触抑制。长时程突触抑制阳性的通道数占所有激活通道数的比值,被定义为诱导率(inductionratio),在野生型小鼠上为70.5±4.9%(图2d),但是在asic1a基因敲除小鼠上降低为20.9±5.7%(p=1.156e-05,asic1akovs.wt,图2d)。
为进一步建立asic1a在岛叶长时程突触抑制中的作用,采用一种条件性asic1a基因敲除小鼠(asic1aflox/flox)(wu,p.y.etal.(2013).acid-sensingionchannel-1aisnotrequiredfornormalhippocampalltpandspatialmemory.jneurosci33(5),1828-1832)的模型。在进行脑片记录前一个月,在岛叶皮层双侧注射携带cre重组酶或者对照绿色荧光蛋白的腺相关病毒(adeno-associatedvirus,aav)。免疫印迹表明,相比对照病毒,cre病毒注射显著降低了asic1a蛋白的表达。长时程突触抑制的诱导率从注射对照病毒的80.0±4.3%降低至注射cre病毒的15.9±5.5%(p=2.177e-06vs.aav-ctrl,图2e,f)。
接下来检查药理学抑制asic1a通道对于低频刺激-长时程突触抑制的影响。将asic1a同聚体和asic1a/2b异聚体通道抑制剂——狼蛛毒素(psalmotoxin1,pctx1,100纳摩尔/升)从低频刺激前15分钟开始给予直至整个低频刺激过程结束。以热失活的狼蛛毒素(pctx1)作为对照。结果发现,无论是狼蛛毒素或者热失活的狼蛛毒素对于基础兴奋性突触传递都没有影响(图2g);但是给予pctx1后的岛叶皮层长时程突触抑制的诱导率降低为17.9±9.3%,而热失活pctx1处理脑片的长时程突触抑制诱导率为72.4±8.4%(p=0.0018,图2h)。这些结果确认了asic1a在岛叶皮层长时程突触抑制中的关键作用,也排除了asic1a基因敲除小鼠表现出长时程突触抑制诱导异常源自于发育性代偿的可能性。
在图2中,(a)(左)多电极阵列记录电极相当于岛叶皮层冠状切片的位置示意图;(右)多电极阵列记录电极的位置排布(8×8)。(b)多电极阵列记录电极在岛叶皮层脑片上的光镜照片,图示还标注了不同的分层信息(从第i层到v–vi层)。其中红色圆点为刺激位点。(c,e,g)在图示的不同条件下,低频刺激(1赫兹,900次脉冲刺激)引起的岛叶皮层场兴奋性突触后电位斜率的时程依赖改变。插图:图示时间点的代表性场兴奋性突触后电位轨迹。比例尺:纵坐标为100微伏,横坐标为10毫秒。(d,f,h)每张岛叶皮层脑片激活通道数和呈现出长时程突触抑制的通道数总结。n.s.,利用非配对student’st-检验对两组进行比较后无统计学差异;*p<0.05,***p<0.001,利用非配对student’st-检验对两组进行比较后,表明两组呈现出显著的统计学差异。(d)n=7–9张脑片(7–8只小鼠);(f)n=6–7张脑片(每组4只小鼠);(h)n=5–6张脑片(3–5只小鼠)。
实施例3ph动态变化过程为低频刺激诱导的长时程突触抑制(ltd)所必需
为了检测asic1a赖以激活的ph动态变化过程对于长时程突触抑制诱导的影响,增强了低频刺激诱导期间细胞外液的ph缓冲能力,该操作在过去的研究(kreplec.j.etal.(2014).acid-sensingionchannelscontributetosynaptictransmissionandinhibitcocaine-evokedplasticity.natneurosci17(8),1083-1091)中表明能够降低神经元asic的激活。该处理显著地降低了野生型小鼠低频刺激长时程突触抑制的诱导概率(图3)。这些结果说明,asic1a通过感受细胞外ph的改变,从而调节长时程突触抑制的诱导率。
在图3,通过提高细胞外溶液碳酸氢根离子(hco3–,浓度为70毫摩尔/升(mm);常规为25mm)和co2的浓度(浓度为15%;常规为5%)从而增强ph缓冲能力后,阻断了岛叶皮层长时程突触抑制的诱导性。在增强ph缓冲能力的处理条件下,低频刺激引起的场兴奋性突触后电位斜率的时程依赖改变。n=56个激活通道(7张脑片,6只野生型小鼠)。
实施例4在asic1a基因敲除小鼠再表达asic1a解救低频刺激-长时程突触抑制
为了测试asic1a基因敲除小鼠的岛叶皮层上恢复asic1a的表达是否能够解救长时程突触抑制的诱导,准备同时包含绿色荧光蛋白和小鼠asic1a基因序列的aav病毒,这两种功能蛋白序列之间用一种“自剪切”的2a多肽序列进行连接,这种病毒用于过表达asic1a。此外,也准备仅包含绿色荧光蛋白的aav病毒作为对照。为了实现神经元特异性的表达,对照病毒和asic1a过表达病毒都由人源突触蛋白i(humansynapsini)启动子在神经元上进行驱动表达。在脑片记录前1个月,在两侧岛叶皮层分别注射对照或asic1a过表达病毒(图4a-c)。结果发现,注射asic1a过表达病毒,能够成功解救低频刺激-长时程突触抑制(图4d),而对照病毒不具有类似的作用。其中,在岛叶皮层注射对照病毒后,长时程突触抑制的诱导率为14.9±6.3%;而注射asic1a过表达病毒后,长时程突触抑制的诱导率为69.9±5.4%(p=2.513e-05vs.aav-ctrl,图4e)。此外,过表达一种不具有离子通透性的asic1a突变体病毒,即aav-asic1a-hif(也就是第32至34位的组氨酸-异亮氨酸-苯丙氨酸残基都突变为丙氨酸,图4a),不能解救asic1a基因敲除小鼠的长时程突触抑制的异常(图4d),其诱导率为24.0±8.2%,与注射对照病毒的组别没有显著差异(p=0.3976,图4e)。这些结果支持asic1a及其离子通透性对于岛叶皮层长时程突触抑制的关键贡献,同时也进一步排除了发育阶段的代偿对于解释asic1a基因缺陷小鼠长时程突触抑制受损的贡献。
在图4中,(a)aav载体的构成示意图。hsynapsini,代表人源synapsini启动子,主要用于驱动基因选择性地在神经元表达。(b)aav介导的荧光蛋白在岛叶皮层的表达(左)。该脑片用dapi进行负染。为了更清晰展示,右侧放置了相当位置的脑图谱示意图。(c)岛叶皮层单个神经元水平表达荧光蛋白的高放大倍数图片。(d)在图示的不同条件下,低频刺激(1赫兹,900次脉冲刺激)引起的岛叶皮层场兴奋性突触后电位斜率的时程依赖改变。插图:图示时间点的代表性场兴奋性突触后电位轨迹。比例尺:纵坐标为100微伏,横坐标为10毫秒。(e)每张岛叶皮层脑片激活通道数和呈现出长时程突触抑制的通道数总结。n.s.,利用非配对student’st-检验对两组进行比较后无统计学差异;*p<0.05,***p<0.001,利用非配对student’st-检验对两组进行比较后,表明两组呈现出显著的统计学差异。n=6–8张脑片(4–8只小鼠)。
实施例5asic1a对于岛叶皮层dhpg诱导的长时程突触抑制是必需的
为研究asic1a在另外一种突触抑制可塑性形式中的作用,给予i型代谢型谷氨酸受体的激动剂,3,5-二羟苯甘氨酸(3,5-dihydroxyphenylglycine,dhpg,100微摩尔/升),dhpg诱导出的突触抑制对于nmda受体抑制剂不敏感,因此也被称为nmda受体不依赖的长时程突触抑制。显著的是,dhpg诱导的长时程突触抑制在asic1a基因缺陷动物也发生了缺陷(图5a)。在野生型动物,dhpg诱导的长时程突触抑制诱导率为82.2±5.6%;在asic1a基因敲除动物,dhpg诱导的长时程突触抑制诱导率为20.4±9.2%,跟野生型动物相比具有显著差异(p=0.0004,图5b)。重要的是,在asic1a基因敲除小鼠上通过aav病毒再表达asic1a能够恢复dhpg诱导的长时程突触抑制。其中,注射对照病毒后,dhpg-长时程突触抑制的诱导率为15.3±9.4%,过表达asic1a病毒的诱导率为82.2±5.8%,二者相比具有显著差异(p=0.0001,图5c,d)。因此,asic1a对于小鼠岛叶皮层nmda受体依赖与nmda受体不依赖长时程突触抑制的诱导都是必需的。
在图5中,(a,c)在图示条件下,dhpg(100μm,20min,a,c)引起的岛叶皮层场兴奋性突触后电位斜率的时程依赖改变。插图:图示时间点的代表性场兴奋性突触后电位轨迹。比例尺:纵坐标为100微伏,横坐标为10毫秒。(b,d)每张岛叶皮层脑片激活通道数和呈现出长时程突触抑制(b,d)的通道数总结。n.s.,利用非配对student’st-检验对两组进行比较后无统计学差异;*p<0.05,**p<0.01,利用非配对student’st-检验对两组进行比较后,表明呈现出显著的统计学差异。(b)n=5张脑片(每组4只小鼠);(d)n=5–6张脑片(5–6只小鼠)。
实施例6asic1a并不贡献于岛叶皮层长时程突触增强的诱导
检查asic1a基因敲除或抑制对于岛叶皮层另外一种形式的突触可塑性的影响。在野生型岛叶皮层脑片上进行theta串刺激(thetaburststimulation,tbs)后能够在多个位点诱导出快速且持久的突触反应的增强(图6a),即长时程突触增强(long-termpotentiation,ltp)。然而,theta串刺激在野生型和asic1a基因敲除小鼠脑片上诱导出类似水平的长时程突触增强(图6a),其中长时程突触增强的诱导率在野生型小鼠上为75.5±7.4%,在asic1a基因敲除小鼠上为71.2±6.3%,二者之间没有显著性差异(p=0.6711,图6b)。此外,asic1a抑制剂pctx1不影响野生型小鼠theta串刺激引起的长时程突触增强(图6c),其长时程突触增强的诱导率为60.9±6.6%(图6d),跟未处理的野生型脑片之间相比并无差异(p=0.1822)。这些结果表明,asic1a不大可能参与theta串刺激诱导的长时程突触增强,这一点跟前述的asic1a在长时程突触抑制中的重要贡献形成鲜明对比。
在图6中,(a,c)在图示条件下,theta串刺激(以5赫兹的频率共10串刺激,每串刺激包含4次100赫兹频率的脉冲刺激,theta-burststimulation,tbs,a,c)引起的岛叶皮层场兴奋性突触后电位斜率的时程依赖改变。插图:图示时间点的代表性场兴奋性突触后电位轨迹。比例尺:纵坐标为100微伏,横坐标为10毫秒。(b,d)每张岛叶皮层脑片激活通道数和呈现出长时程突触增强(b,d)的通道数总结。n.s.,利用非配对student’st-检验对两组进行比较后无统计学差异;*p<0.05,**p<0.01,利用非配对student’st-检验对两组进行比较后,表明呈现出显著的统计学差异。(b)n=6张脑片(每组6只小鼠);(d)n=5–6张脑片(5–6只小鼠)。其中,图(d)中的对照数据源自于图(b),重复显示是为了更好的比较。
实施例7岛叶皮层长时程突触抑制选择性被条件性味觉厌恶消退过程所抵消
为了诱导条件性味觉厌恶,给予小鼠喂饲糖水,并将其作为条件刺激(conditionedstimulus,cs),随后通过腹腔注射licl引起小鼠腹痛,后者作为非条件刺激(unconditionedstimulus,us)。基于比较小鼠在进行条件性味觉厌恶训练后接下来几天里分别消耗水和糖水的量,计算厌恶指数[饮水量/(饮水量+饮糖量)×100%],从而来评价条件性味觉厌恶的建立。利用这种步骤,大部分动物在经历过训练后获得的厌恶指数在80-90%之间。为了彻底消退条件性味觉厌恶,这些动物在训练后接下来的7天里只被允许喝糖水,而对照动物只被允许喝水以维持对于特定味觉(糖水)的厌恶性记忆(图7a)。如图7b所示,条件性味觉厌恶对照小鼠保持了较高的厌恶指数,然而彻底消退动物的厌恶指数已经显著降低,表明在不与licl注射再次建立偶联的前提下反复消耗糖水将导致条件性味觉厌恶的消退。
通过分析上述小鼠的岛叶皮层脑片,发现条件性味觉厌恶对照动物呈现出正常的长时程突触抑制,其中,低频刺激-长时程突触抑制(图7c)的诱导率为68.5±7.5%(图7d);dhpg-长时程突触抑制(图7e)的诱导率为80.3±2.6%(图7f)。这两个数值跟天然小鼠上获得的数据(图2d和5b)并无差异,表明岛叶皮层长时程突触抑制并不参与条件性味觉厌恶的获取和保留。另一方面,从条件性味觉厌恶彻底消退小鼠获得的脑片对于低频刺激(图7c)或dhpg(图7e)表现出长时程突触抑制诱导率的降低,其中低频刺激-长时程突触抑制的诱导率为19.3±11.4%(图7d,p=0.0049),dhpg-长时程突触抑制的诱导率为11.4±5.5%(图7f,p=1.982e-07),跟条件性味觉厌恶动物相比具有显著的差异。这些结果表明,条件性味觉厌恶的消退过程抵销了导叶皮层的长时程突触抑制。
比较条件性味觉厌恶彻底消退及其对照组织的兴奋性突触后电流的输入-输出曲线后发现,消退过程降低了导叶皮层的兴奋性突触传递(图7g,h)。因此,岛叶皮层长时程突触抑制参与了条件性味觉厌恶记忆的消退过程。味觉厌恶记忆的彻底消退过程导致了一种饱和效应,后者抵消了随后进行的长时程突触抑制诱导过程。
在图7中,(a)行为学操作步骤。在第0天,小鼠通过将糖水与腹腔注射licl引起的腹部不适建立联合性味觉厌恶记忆。在随后的第1–7天,其中一组小鼠将仅被饲喂糖水,以将已建立的条件性味觉厌恶记忆彻底消退;另外一组小鼠将仅被饲喂水,以保留已建立的条件性味觉厌恶记忆。第8天,牺牲小鼠,用于制备脑片,进行电生理记录。(b)通过比较小鼠饮水量和饮糖量计算获得的厌恶指数。厌恶指数=饮水量/(饮水量+饮糖量)×100%。n=5–7。***p<0.001,非配对student’st-检验。(c-f)岛叶皮层长时程突触抑制的诱导在味觉厌恶记忆彻底消退小鼠上消失,但是在保留有味觉厌恶记忆的小鼠依旧存在。(c,e)味觉厌恶记忆彻底消退或依旧保留的小鼠岛叶皮层脑片响应低频刺激(c)或dhpg(g)后引起的场兴奋性突触后电位的时程依赖改变。插图:图示时间点的代表性场兴奋性突触后电位轨迹。比例尺:纵坐标为100微伏,横坐标为10毫秒。(d,f)每张岛叶皮层脑片激活通道数和呈现出长时程突触抑制的通道数总结。n.s.,利用非配对student’st-检验对两组进行比较后无统计学差异;*p<0.05,***p<0.001,利用非配对student’st-检验对两组进行比较后,表明两组呈现出显著的统计学差异。(d)n=6张脑片(每组4只小鼠);(f)n=5–7张脑片(每组4只小鼠)。(g,h)味觉厌恶记忆彻底消退或依旧保留的小鼠岛叶皮层脑片神经元的兴奋性突触后电流的输入-输出曲线。(g)代表性轨迹。(h)汇总数据。n=20–23细胞(每组7只小鼠)。*p<0.05,**p<0.01,***p<0.001利用非配对student’st-检验对两组进行比较后,表明两组呈现出显著的统计学差异。
实施例8asic1a的缺失选择性损害了条件性味觉厌恶记忆的消退
asic1a在岛叶皮层长时程突触抑制诱导中发挥重要作用,预期asic1a基因缺陷小鼠可能表现出条件性味觉厌恶记忆的正常获取和保留,但是在条件性味觉厌恶消退中可能发生障碍。结果表明,野生型和asic1a基因敲除小鼠的确在条件性味觉厌恶记忆的获取方面表现相当。在进行条件化训练的当天,这两株小鼠摄取了类似量的糖水(野生型:1.93±0.11克,n=15;asic1a基因敲除小鼠:1.74±0.15克,n=14;p=0.3020)。在经历过条件化训练后的第一天,两组动物都发展出了类似的厌恶反应(图8a),这表明,asic1a在条件性味觉厌恶记忆的获取和提取过程中并不发挥显著的作用。在随后的3天、7天、14天和28天进行条件性味觉厌恶记忆强度测试,发现野生型和asic1a基因敲除小鼠的记忆保留强度相似(图8b),这表明asic1a基因敲除小鼠跟野生型小鼠一样,能够保留正常的条件性味觉厌恶记忆至少维持1个月。
相比之下,asic1a基因敲除小鼠在随后的条件性味觉厌恶消退测试中却不能表现出显著降低的厌恶程度,而野生型小鼠在味觉选择性测试后第二至三天即表现出显著降低的记忆强度(图8c)。类似地,若对远期的条件性味觉厌恶记忆进行消退训练,asic1a基因敲除小鼠相比野生型小鼠也表现出显著的消退受损(图8d)。这些结果表明,asic1a选择性贡献于条件性味觉厌恶记忆的消退,但是对于记忆获取或保留并没有显著作用。
作为对照,为证实asic1a基因缺陷动物具有完整的味觉区分能力,利用非条件化的双瓶味觉偏好测试(yu,h.,etal..(2009).variantbdnfval66metpolymorphismaffectsextinctionofconditionedaversivememory.jneurosci29,4056-4064),分别检查小鼠对甜味、咸味、苦味和酸味溶液的偏好性,发现野生型和asic1a基因敲除小鼠之间并无明显区别。
在图8中,野生型和asic1a基因敲除小鼠在第0天进行条件性味觉厌恶训练,在第1天进行味觉厌恶记忆的获取测试;利用不同批次的小鼠,分别在进行条件性味觉厌恶训练后第3天、第7天、第14天和第28天,进行味觉厌恶记忆测试,定义为记忆强度保留的测试;关于消退测试,已建立条件性味觉厌恶记忆的小鼠将被给予一瓶糖水和一瓶水进行选择测试,记忆建立后1–7天的连续味觉选择测试,称为近期味觉厌恶记忆的消退;记忆建立后15-21天的连续味觉选择测试,称为远期味觉厌恶记忆的消退。其中,图(c)中第1天的数据跟图(a)数据一样,反复呈现是为了更好的比较。n=8–16。n.s.,利用非配对student’st-检验对两组进行比较后无统计学差异;*p<0.05,**p<0.01,***p<0.001,利用非配对student’st-检验对两组进行比较后,表明两组呈现出显著的统计学差异。
实施例9岛叶皮层asic1a对于条件性味觉厌恶消退的关键作用
为明确岛叶皮层上asic1a对于条件性味觉厌恶消退的作用,再次利用前述的asic1a条件性基因敲除小鼠,即asic1aflox/flox小鼠。相比注射aav对照病毒,往asic1aflox/flox小鼠岛叶皮层注射aav-cre病毒(图9a)导致动物表现出显著降低的条件性味觉厌恶消退(图9b),这结果支持了岛叶表达的asic1a在条件性味觉厌恶消退中发挥特异性作用。
为进一步测试在岛叶皮层对于asic1a进行急性抑制是否影响条件性味觉厌恶的消退,在已建立条件性味觉厌恶记忆的动物进行双瓶选择测试后立即给予asic1a抑制剂——pctx1(10皮摩尔)或者其溶剂对照(图9c)。相比于溶剂对照,给予pctx1(图9d)显著地阻断了条件性味觉厌恶的消退(图9e)。
为测试在asic1a基因敲除小鼠岛叶皮层再表达asic1a是否能够解救基因缺陷动物的消退异常。采用实施例4所示的干预策略,所不同的是,在这里的实验中,在aav病毒注射后1个月对于动物进行条件性味觉厌恶训练,随后在训练后3-6天进行味觉选择性测试(图9f)。正如所料,在asic1a基因缺陷小鼠的岛叶皮层注射asic1a过表达病毒,使得动物的条件性味觉厌恶消退水平接近于野生型小鼠,然而注射对照或者asic1a-hif突变体过表达病毒却没有了类似的效应(图9g)。因此,岛叶皮层上功能性的asic1a是asic1a基因敲除小鼠关键的缺失组分,后者恰好支持了动物的条件性味觉厌恶消退学习过程。
利用另一种消除asic1a表达的策略,构建一种表达靶向asic1a的短发夹rna(shorthairpinrna,shrna)的aav病毒载体,并由u6启动子驱动表达(图9i)。在中国仓鼠卵巢细胞上确证了asic1a-shrna有效性以后,在野生型小鼠岛叶皮层注射aav-asic1a-shrna或阴性对照病毒(图9h)。相比aav阴性对照病毒,aav-asic1a-shrna病毒显著降低了岛叶皮层aisc1a蛋白的表达(p=5.651e-05,n=4–6,图9j),也导致条件性味觉厌恶消退过程被显著阻断(图9k)。在asic1a-shrna的基础上,增加兴奋性神经元类型特异性再表达asic1a的解救策略(图9l)。这里的aav病毒载体除了包含asic1a-shrna元件外,还包括对于shrna不敏感的mcherry-2a-asic1a(记作asic1a*)双flox控制反向(double-floxedinvertedorientation,dio)的表达序列,后者只在存在cre重组内切酶的条件下才能正常表达(图9m)。将这种aav病毒注射至camkii-cre工具小鼠(cre重组酶只表达在前脑的锥体神经元)的岛叶皮层,发现在岛叶皮层的一群神经元里有mcherry(图8n)和asic1a(图9o)的显著表达。但是,将同样的病毒注射至野生型小鼠,mcherry(图9n)或asic1a(图9o)都没有显著的表达。该aav病毒载体包含的asic1a-shrna序列(图9m)也是有效的,表现为注射了该病毒的野生型小鼠条件性味觉厌恶消退显著异常,亦即表现出较高的厌恶指数(图9p)。相比之下,在camkii-cre工具小鼠,aav病毒的注射使得动物的条件性味觉厌恶消退接近于正常对照动物(图9p)。总之,上述结果建立了岛叶皮层asic1a在条件性味觉厌恶消退中的关键作用。
在图9中,(a,c,f,h,l)是行为学步骤。(b,e,g,k,p)是条件性味觉厌恶消退引起厌恶指数的时程依赖改变。*p<0.05,**p<0.01,***p<0.001,利用非配对student’st-检验对两组进行比较后,表明两组呈现出显著的统计学差异。(b)n=7。(e)n=10–11。(g)n=8–13.*p<0.05,***p<0.001,wt+aav-ctrlvs.ko+aav-ctrl;#p<0.05,##p<0.01,ko+aav-asic1avs.ko+aav-ctrl.(k)n=9–12。(p)n=11–12。*p<0.05,**p<0.01,wt+aav-nc-ctrlvs.wt+aav-shrna-dio-asic1a*;#p<0.05,##p<0.01,wtvs.camkii-creforaav-shrna-dio-asic1a*injected,非配对student’st-检验。(d)岛叶皮层定点注射药物后弥散区域的示意图。(i,m)aav载体的构成示意图。(j,o)代表性的免疫印迹图片。(n)岛叶皮层mcherry表达的图片,这里用dapi进行了负染。
实施例10岛叶皮层长时程突触抑制对于条件性味觉厌恶消退是必需的
为进一步建立岛叶皮层asic1a依赖的长时程突触抑制对于条件性味觉厌恶消退的意义,研究在体阻断岛叶皮层长时程突触抑制的行为学后果。ampa受体的内吞是长时程突触抑制的一种普遍机制,该过程可被从细胞内给予的一段glua2模拟短肽——glua2-3y,或者从细胞外给予具有穿模能力的glua2模拟短肽——fitc-tat-glua2-3y(tat-3y,图10a)所阻断。其中,多肽的穿模能力通过包含一段源自于人类免疫缺陷病毒(humanimmunodeficiencyvirus,hiv)的tat短肽实现。其阴性对照,fitc-tat-glua2-3a(tat-3a,图10a)虽然也具有穿模能力,但是不能发挥阻断ampa受体内吞的效应。在进行低频刺激前以及持续期间,给予tat-3y(1微摩尔/升)显著阻断了岛叶皮层长时程突触抑制的诱导,然而对照多肽tat-3a(1微摩尔/升)却没有该效应(图10b)。其中,处理过tat-3y多肽的脑片长时程突触抑制诱导率为16.0±7.1%,而对照多肽tat-3a多肽处理的脑片诱导率为78.0±6.1%,二者相比具有显著性差异(p=5.900e-05,图10c)。这些结果表明,glua2亚基依赖的ampa受体内吞是岛叶皮层长时程突触抑制的关键步骤。
在体给予岛叶皮层tat-3y多肽(100皮摩尔)对条件性味觉厌恶消退的影响见图10d。基于岛叶皮层呈现出的fitc绿色荧光,确证了给予tat-3y多肽的有效性和特异性(图10e)。相比于接受对照多肽tat-3a或者溶剂对照的小鼠,接受了tat-3y多肽的动物表现出显著的条件性味觉厌恶消退的异常(图10f)。该结果支持岛叶皮层的长时程突触抑制对于条件性味觉厌恶消退是必需的。总之,岛叶皮层asic1a在条件性味觉厌恶消退中起着关键作用。
在图10中,(a)阻断ampa受体内吞的干扰肽序列,关键的酪氨酸残基(y)用红色表示;当关键酪氨酸残基被突变为丙氨酸(a)后形成的多肽失去阻断干扰ampa受体内吞的效应,因而作为对照肽。(b)急性给予tat-3y干扰肽(1微摩尔/升,从低频刺激步骤前20分钟开始,共持续35分钟)阻断了低频刺激引起的长时程突触抑制,但是给予对照多肽(tat-3a)无显著作用。(c)每张岛叶皮层脑片激活通道数和呈现出长时程突触抑制的通道数总结。n.s.,利用非配对student’st-检验对两组进行比较后无统计学差异;***p<0.001,利用非配对student’st-检验对两组进行比较后,表明两组呈现出显著的统计学差异。(d)行为学步骤。(e)注射干扰多肽的位置及其侵入细胞内部的示意图。(f)条件性味觉厌恶消退引起厌恶指数的时程依赖改变。n=10–14。***p<0.001,利用非配对student’st-检验对两组进行比较后,表明tat-3a与tat-3y两组间呈现出显著的统计学差异。
实施例11药理学阻断或激活腹侧海马asic1a分别抑制和促进恐惧消退
为了检查asic1a在恐惧消退中的生理意义,第二天消退学习前半小时通过在双侧腹侧海马微注射生理盐水、asic1a激动剂mittx-α/β或asic1a阻断剂pctx1(注射体积均为1微升/侧,图11a,b),观察和定量统计在第二天消退学习及第三天恐惧检测过程中动物对cs的恐惧反应。数据以平均值±标准误的形式进行展示。动物数目为每组8-14只小鼠。
所述实验制剂选自下组:
(i)mittx-α/β,1微摩尔/升,溶于生理盐水。
(ii)pctx1,10微摩尔/升,溶于生理盐水。
结果如下(生理盐水处理组用空白图表示,mittx-α/β或pctx1处理组用填充图表示):
在恐惧学习过程中,动物对cs的恐惧反应逐渐增加,第二天消退学习过程中动物对cs的恐惧反应逐渐降低,而第三天动物能维持对cs的低恐惧反应。与生理盐水组相比,动物消退学习前腹侧海马注射asic1a激动剂mittx-α/β可显著促进恐惧消退,即在第三天mittx-α/β处理组动物对cs维持更低的恐惧反应(图11c,d);相反,动物消退学习前腹侧海马注射asic1a阻断剂pctx1可显著抑制恐惧消退。
实施例12腹侧海马条件性敲除或过表达asic1a分别抑制和促进恐惧消退
在野生型c57bl/6j小鼠腹侧海马定点注射aav-gfp/aav-asic1a(注射体积均为1微升/侧),或在条件性asic1a基因敲除——asic1aflox/flox小鼠腹侧海马定点注射aav-gfp/aav-cre(注射体积均为1微升/侧),并在病毒表达一个月后观察和定量统计动物在恐惧学习、消退学习以及第三天恐惧检测过程中动物对cs的恐惧反应(图12a)。数据以平均值±标准误的形式进行展示。动物数目为每组13-19只小鼠。
所述实验制剂选自下组:
(i)aav-gfp,滴度超过1012病毒基因组(viralgenomes,vg)/毫升(ml),溶于无菌pbs。
(ii)aav-cre,滴度超过1012vg/ml,溶于无菌pbs。
(iii)aav-asic1a,滴度超过1012vg/ml,溶于无菌pbs。
结果如下(aav-gfp处理组用空白图表示,aav-cre或aav-asic1a处理组用填充图表示):
腹侧海马过表达asic1a,在第一天动物恐惧学习不受影响,但随后的消退学习增强(图12b,c);腹侧海马条件性敲除asic1a,在第一天动物恐惧学习同样不受影响,但随后的消退学习受损(图12d,e)。
以上所述实施例仅表达了本发明的实施方式,其描述较为具体和详细,但并不能因此而理解为对本发明专利范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。因此,本发明专利的保护范围应以所附权利要求为准。
- 还没有人留言评论。精彩留言会获得点赞!