膳食产品的制作方法
发明领域
本发明总体上涉及用于延缓或抑制患有癌症的患者中的转移并增加对治疗的响应性的治疗领域。更特别地,本发明涉及通过降低膳食中的天冬酰胺或通过其他手段改变受试者的血清中的天冬酰胺水平,以便延缓或抑制转移和/或防止上皮向间充质转变。本发明还涉及对患有癌症的、处于癌症转移的特定风险的患者群体进行鉴定和治疗。
发明背景
癌症是一种细胞经历不受控制的生长、生长和分裂超过正常细胞生长极限的疾病。这些细胞可以侵袭并破坏周围的组织。此外,癌细胞可以转移,其中它们可以通过血液或淋巴系统扩散到身体的其他区域。
癌症治疗可以包括切除肿瘤的手术、缩小肿瘤尺寸的放射治疗或者药物治疗/化学治疗,使用药物或其他药品(medicine)治疗癌症。癌症的存活率因癌症类型而异;然而,对于具有转移率(metasizedrate)的癌症来说,存活率特别低。
例如,大多数死于乳腺癌的女性不是死于原发性肿瘤,而是死于最初病变被切除后趋于明显的转移。为了促成细胞转移,它们必须离开原发部位,进入脉管系统,在血液中存活,并且然后外渗并在继发部位定殖(vanharanta等人2013)。先前对乳腺肿瘤异质性模型的研究鉴定到两个克隆的4t1亚系(4t1-e和4t1-t),它们具有通过非侵袭性机制进入脉管系统的能力,该能力需要血管模拟(wagenblast等人2015和miller等人1983)。然而,这两个克隆在其对继发性病变的贡献方面差异极大。
因此,需要鉴定转移驱动因子并且需要提供能够延缓或抑制转移的药物。
公开内容的简述
本发明人令人惊讶地发现,原发性肿瘤中的天冬酰胺合成酶(asns)表达与晚期转移性复发密切相关,并且降低患有癌症的受试者的血清天冬酰胺水平可以延缓或抑制转移。此外,本发明人令人惊讶地证明,通过膳食或其他方式降低胞外天冬酰胺可用性有利地延缓或抑制转移。延缓或抑制转移的膳食手段可以有利地补充现有癌症治疗方案。
因此,在本发明的第一方面,本发明提供了一种包含多种氨基酸的膳食产品,其中该膳食产品包含所有必需氨基酸并且其中该膳食产品基本上不含天冬酰胺。合适地,该膳食产品可以包含至少12种氨基酸。
合适地,该膳食产品可以基本上不含选自由以下组成的组的至少一种或多于一种另外的非必需氨基酸:谷氨酰胺、甘氨酸、丝氨酸、半胱氨酸、酪氨酸和精氨酸。
合适地,该膳食产品还可以包含一种或更多种大量营养元素和/或一种或更多种微量营养元素。
合适地,该膳食产品还可包含例如水平少于25mg/kg/日或少于20mg/kg/日或少于18mg/kg/日或少于16mg/kg/日的甲硫氨酸。
合适地,该产品可以被配制成至少提供基于平均每日总蛋白消耗的每日推荐摄入量的必需氨基酸。
合适地,该膳食产品可以是固体或流体的形式。其可以被配制成用于作为膳食替代品(mealreplacement)口服施用。可选地,其可被静脉内递送。
在另一方面,本发明提供了制备本发明的膳食产品的方法,其中组分被溶解或分散在水中并喷雾干燥。
在另外的方面,本发明提供了一种药物组合物,所述药物组合物包含本发明的膳食产品或根据本发明的方法产生的膳食产品以及药学上可接受的运载体、赋形剂或稀释剂。
合适地,该药物组合物还可以包含选自以下的治疗剂:癌细胞生长抑制剂、抗转移剂、免疫检查点抑制剂、放射治疗剂和化学治疗剂。合适地,治疗剂可以降低血液中的天冬酰胺水平。
合适地,治疗剂可以是天冬酰胺合成酶抑制剂或l-天冬酰胺酶。
本发明还提供了用于在治疗中使用的本发明的膳食产品或根据本发明产生的膳食产品或本发明的药物组合物。
在另外的方面,本发明提供了一种用于在延缓或抑制患有癌症的受试者中的转移中使用的药物,其中所述药物降低患有癌症的受试者的血液中的天冬酰胺水平(即,降低血液中天冬酰胺的生物可利用度,例如胞外水平)。
合适地,该药物可以选自由以下组成的组:
a.本发明的膳食产品;
b.根据本发明的方法产生的膳食产品;
c.本发明的药物组合物;
d.天冬酰胺合成酶抑制剂;或者
e.l-天冬酰胺酶。
合适地,癌症可以选自由以下组成的组:乳腺癌、结肠癌、头颈鳞癌(squamousheadandneckcancer)、肾透明细胞癌(renalclearcellcancer)和子宫内膜癌。
合适地,药物可以是本发明的膳食产品或药物组合物,其可以被配制成与l-天冬酰胺酶共施用或顺序施用。
合适地,药物(例如膳食产品)可以与选自以下的治疗剂组合使用:癌细胞生长抑制剂、抗转移剂、免疫检查点抑制剂、放射治疗剂和化学治疗剂。
合适地,受试者可能已经被确定为具有高于对照或预定水平的天冬酰胺合成酶表达水平。合适地,可以确定在肿瘤样品中的天冬酰胺合成酶表达水平。
合适地,受试者可能已被确定为具有高于对照或预定水平的血清天冬酰胺水平。
合适地,受试者可能具有实体瘤。
在另一方面,本发明涉及化合物或组合物在制备用于延缓或抑制患有癌症的受试者中的转移的药物中的用途,其中所述化合物或组合物降低受试者的血液中的天冬酰胺水平(即降低血液中天冬酰胺的生物可利用度,例如胞外水平)。
合适地,化合物可以是以下或组合物可以包括以下:
a.本发明的膳食产品;
b.根据本发明的方法产生的膳食产品;
c.本发明的药物组合物;
d.天冬酰胺合成酶抑制剂;或者
e.l-天冬酰胺酶。
合适地,癌症可以选自由以下组成的组:乳腺癌、结肠癌、头颈鳞癌、肾透明细胞癌和子宫内膜癌。
合适地,膳食产品或药物组合物可以被配制成与l-天冬酰胺酶共施用或顺序施用。
合适地,所述化合物或组合物(例如膳食产品)可以与选自以下的治疗剂组合使用:癌细胞生长抑制剂、抗转移剂、免疫检查点抑制剂、放射治疗剂和化学治疗剂。
合适地,受试者可能已被确定为具有高于对照或预定水平的天冬酰胺合成酶表达水平。
合适地,受试者可能已被确定为具有高于对照或预定水平的血清天冬酰胺水平。
合适地,受试者可能具有实体瘤。
在另外的方面,本发明提供了一种治疗受试者中的癌症的方法,所述方法包括向所述受试者施用治疗有效量的药物,其中所述药物降低患有癌症的受试者的血液中的天冬酰胺水平(即降低血液中天冬酰胺的生物可利用度,例如胞外水平)。
合适地,药物可以包括:
a.本发明的膳食产品;
b.根据本发明的方法产生的膳食产品;
c.本发明的药物组合物;
d.天冬酰胺合成酶抑制剂;或者
e.l-天冬酰胺酶。
合适地,癌症可以选自由以下组成的组:乳腺癌、结肠癌、头颈鳞癌、肾透明细胞癌和子宫内膜癌。
合适地,治疗有效量的药物(例如本发明的膳食产品或药物组合物)可以与治疗有效量的l-天冬酰胺酶共施用或顺序施用。
合适地,治疗有效量的药物(例如膳食产品)可以与选自以下的治疗剂组合施用:癌细胞生长抑制剂、抗转移剂、免疫检查点抑制剂、放射治疗剂和化学治疗剂。
合适地,受试者可能已被确定为具有高于对照或预定水平的天冬酰胺合成酶表达水平。
合适地,受试者可能已被确定为具有高于对照或预定水平的血清天冬酰胺水平。
合适地,受试者可能具有实体瘤。
合适地,膳食产品可以是受试者的唯一营养来源。
合适地,治疗可以在至少24小时的时间内施用或者施用至直到观察到治疗终点。
合适地,药物可以每日施用1次和6次之间。
合适地,通过每日施用膳食产品的方案至少可以满足每日推荐量的必需氨基酸。
在另一方面,本发明提供了血液(血清)天冬酰胺水平作为生物标记鉴定具有肿瘤的、处于增加的转移风险的患者或患者群体的用途。
在另外的方面,本发明提供了天冬酰胺合成酶表达作为标记鉴定具有肿瘤的、处于增加的转移风险的患者或患者群体的用途。合适地,可以确定原发性肿瘤中的表达水平。
本发明还提供了一种鉴定患有癌症的、具有增加的转移可能性的受试者的方法,所述方法包括:
a)确定从受试者分离的生物样品(例如血清)中的天冬酰胺水平;
b)将生物样品中的天冬酰胺水平与对照样品或天冬酰胺的预定参考水平进行比较,
其中生物样品中的天冬酰胺水平与对照样品相比或与预定参考水平相比的增加指示转移可能性增加。
合适地,该方法还可以包括向所述受试者施用治疗有效量的药物,其中所述药物降低患有癌症的受试者的血液中的天冬酰胺水平。
合适地,药物可以是以下或组合物可以包括以下:
a.本发明的膳食产品;
b.根据本发明的方法产生的膳食产品;
c.本发明的药物组合物;
d.天冬酰胺合成酶抑制剂;或者
e.l-天冬酰胺酶。
本发明还提供了一种鉴定在进食基本上不含天冬酰胺的膳食或施用l-天冬酰胺酶时对癌症治疗具有增加响应性或敏感性的可能性的受试者的方法,所述方法包括:
a)确定从受试者分离的生物样品中的天冬酰胺水平;
b)将生物样品中的天冬酰胺水平与对照样品或天冬酰胺的预定参考水平进行比较,
其中生物样品的天冬酰胺水平与对照样品相比或与预定参考水平相比的增加指示在所述癌症治疗结合基本上不含天冬酰胺的膳食或与l-天冬酰胺酶一起施用时,对所述癌症治疗的响应性或敏感性。
合适地,该方法还可以包括在受试者被鉴定为在进食基本上不含天冬酰胺的膳食或被施用l-天冬酰胺酶时对癌症治疗具有增加的响应性或敏感性的可能性时,施用治疗有效量的本发明的膳食产品或根据本发明产生的膳食产品或根据本发明的药物组合物或l-天冬酰胺酶与化学治疗剂的组合。
合适地,生物样品可以是血液样品(例如血清)。
本发明还提供了一种确定具有癌症的受试者中的转移性复发的可能性的方法,所述方法包括:
a)确定从受试者分离的生物样品中的天冬酰胺合成酶水平;
b)将生物样品中的天冬酰胺合成酶水平与对照样品或天冬酰胺的预定参考水平进行比较,
其中生物样品中的天冬酰胺合成酶水平与对照样品相比或与预定参考水平相比增加指示转移性复发的可能性增加。
在另外的方面,本发明提供了一种逆转受试者中上皮向间充质转变或防止患有癌症的受试者中上皮向间充质转变的方法,所述方法包括向有相应需要的受试者施用治疗有效量的本发明的膳食产品或根据本发明方法产生的膳食产品或本发明的药物组合物或l-天冬酰胺酶。
这些方面及其他方面在详述中进行扩展。
贯穿本说明书的描述和权利要求书,词语―包括(comprise)”和―含有(contain)”及它们的变化形式意指―包括但不限于”,并且它们不意图(并且不)排除其他部分、添加物、组分、整数或步骤。
除非上下文另外要求,否则贯穿本说明书的描述和权利要求书,单数涵盖复数。特别地,除非上下文另外要求,否则在使用不定冠词的情况下,本说明书应被理解为预期多于一个以及单个。
结合本发明的特定方面、实施方案或实例所描述的特征、整数、特性、化合物、化学部分或基团应理解为可应用于本文所述的任何其他方面、实施方案或实例,除非与该方面、实施方案或实例不相容。
本文提到的专利、科学和技术文献确立了本领域技术人员在申请日时可获得的知识。本文引用的已颁发的专利、公布的和未决的专利申请以及其他出版物的全部内容通过引用特此并入,其程度等同于各自被具体地且单独地指明通过引用并入的相同程度。在任何不一致的情况下,以本公开内容为准。
下文将进一步详细描述本发明的各个方面。
附图简述
下文参考以下附图进一步描述本发明的实施方案,其中:
图1示出了对转移驱动因子的鉴定。a)从nsg小鼠的肺中提取的4t1-e细胞和4t1-t细胞的相对比例,所述nsg小鼠被通过尾静脉引入了不同浓度的细胞混合物。每个条代表一个样品或一只独立的小鼠。b)在患有不同疾病亚型的患者的原发性肿瘤中,与4t1-e相比,在4t1-t中被鉴定为过表达的基因的表达水平(盒的边缘是第25和第75百分位数,并且误差条延伸到值q3+w(q3-q1)和q1-w(q3-q1),其中w是1.5,并且q1和q3是第25和第75百分位数,并且对于c也是如此,anovap值<0.0001)。c)相同基因在无疾病存活者和具有肺部复发的患者中的表达水平(秩和p值<0.01)。d)鉴定体外侵袭和体内外渗并定殖的驱动因子的rnai筛选方案(n=5只小鼠/shrna池或n=2个6孔基质胶侵袭室/shrna池,基因水平击中用经验贝叶斯调整的t-检验判定,fdr<0.05)。e)在d中描绘的rnai筛选的每个组(arm)中鉴定的基因之间的重叠(超几何检验p值<0.0001)。
图2示出了对天冬酰胺合成酶作为侵袭和转移驱动因子的验证。a)对静脉内注射了asns-沉默的4t1-t细胞或表达asns的4t1-t细胞的小鼠的肺部转移的定量(n=10只小鼠/细胞系,盒的边缘是第25和第75百分位数,并且误差条延伸到值q3+w(q3-q1)和q1-w(q3-q1),其中w是1.5,并且q1和q3是第25和第75百分位数,并且对于e、f和g也是如此,秩和检验p值<0.001)。b)a中描述的肺的代表性图像。c)在应用asns-沉默的细胞和表达asns的细胞24小时后,基质胶测定的收集孔的代表性图像。d)原位注射a中描述的细胞产生的肿瘤体积(n=10只小鼠/细胞系,误差条是第25和第75百分位数)。e)对应于d中描述的肿瘤的动物中ctc的相对丰度(n=4只小鼠/细胞系,秩和p值<0.05)。ctc丰度通过针对mcherry的qpcr从全血基因组dna测量,mcherry由逆转录病毒shrna递送载体表达。f)对来自e中描述的小鼠的h&e染色的肺部切片中的转移的定量(秩和p值<0.0002)。g)f中描述的转移的直径。
图3示出胞外天冬酰胺可用性驱动侵袭和转移。a)基于hplc对asns-沉默的细胞和表达asns的细胞的细胞游离天冬酰胺水平的定量(n=3个重复品/细胞系)。b)通过基质胶侵袭测定测量对亲本4t1细胞在补充有所指示的非必需氨基酸的培养培养基中的侵袭率的定量(n=5个侵入室,秩和p值<0.01)。c)对注射了asns-沉默的4t1-t细胞或表达asns的4t1-t细胞的动物中的肺转移的定量。对于每条线,一半的动物被施用pbs,而另一半被施用l-天冬酰胺酶(n=10只小鼠/条件,误差条是第25和第75百分位数,并且对于e和f也是如此,对于每条线的l-天冬酰胺酶与对照以及对于处于每种药物条件的asns-沉默的细胞与asns-未沉默的细胞,秩和p值<0.0005)。d)来自c中呈现的每种实验条件的代表性h&e染色的肺切片。e)对注射了asns-沉默的4t1-t细胞和表达asns的4t1-t细胞的动物中的肺转移的定量。在实验持续期间,向动物施用具有0%、0.6%或4%天冬酰胺含量的膳食(n=10只小鼠/条件,对于所有膳食,asns-沉默的细胞与表达asns的细胞、伴随0%对比0.6%对比4%膳食的shrenilla细胞,并且对于伴随4%对比0%膳食的shasns-1和shasns-2,秩和p值<0.0005,对于伴随0.6%对比0%或0.6%对比4%膳食的shasns-1和shasns-2,秩和p值<0.05)。f)对施用l-天冬酰胺酶或pbs的动物的乳腺、血清和肺中的天冬酰胺水平的质谱定量(通过总代谢物峰面积归一化的相对丰度,n>8个组织切片/条件,对于所有组织,pbs对比l-天冬酰胺酶,秩和p值<0.005,对于乳腺对比肺,秩和p值<0.05,并且对于血清对比肺和血清对比乳腺,秩和p值<0.0005)。
图4示出天冬酰胺可用性调节上皮向间充质的转变。a)由asns-沉默诱导的蛋白与rna水平表达改变分析氨基酸富集。在蛋白水平的改变低于通过对应的rna水平改变所预期的情况下,在蛋白中富集具有负相关性的氨基酸。正相关指示,在蛋白水平改变超过通过rna量的改变预测的改变时富集在蛋白中的氨基酸。b)asns-沉默的4t1-t细胞和表达asns的4t1-t细胞中emt-上调蛋白的相对蛋白丰度(n=3个重复品/细胞系,盒的边缘是第25和第75百分位数,并且误差条延伸到值q3+w(q3-q1)和q1-w(q3-q1),其中w是1.5,并且q1和q3是第25和第75百分位数,并且对于c、d和h也是如此,秩和p值<0.05)。c)当在正常培养基和补充了天冬酰胺的培养基中生长时,亲本4t1细胞中emt-上调蛋白的相对蛋白丰度(n=3个重复品/细胞系,秩和p值<0.05)。d)从原位注射亲本4t1细胞获得的肺转移与原发性肿瘤中emt-上调和emt-下调基因的相对表达水平(n=4只小鼠,对于emt-上调基因,符号秩(sign-rank)p值0.001)。e)针对来源于asns-沉默的4t1-t细胞和表达asns的4t1-t细胞的原位肿瘤中的twist1和cdh1的ihc染色的代表性图像。f)对e中描述的所有twist1染色的定量(n=5个肿瘤切片和n>5个肺转移灶,误差条代表1个标准偏差,并且对于g也是如此,对于asns-沉默的肿瘤对比表达asns的肿瘤和转移灶,秩和p值分别<0.01和<0.05)。g)对e中描述的所有cdh1染色的定量(n=5个肿瘤切片和n=9个肺转移灶,对于asns-沉默的肿瘤与表达asns的肿瘤和转移灶,秩和p值分别<0.01和<0.05)。h)从来源于asns-沉默的4t1-t细胞和表达asns的4t1-t细胞的肿瘤和肺分离的细胞中的twist1、cdh1、emt-上调基因和emt-下调基因的相对表达水平(n=2个重复品/条件,对于肿瘤中的emt-下调基因,符号秩p-值<0.05,并且对于肺部的emt-上调基因,符号秩p-值<0.001,cdh1和twist1在两种组织中差异表达,deseqfdr<0.05)。
图5示出了对患者数据中asns表达水平的初步分析。对于在4t1-t与4t1-e中被鉴定为上调的每个基因,使用三个不同的数据集计算预后值。一个由三个患者匹配的基础肿瘤(basaltumour)和转移灶对(患者a1、a7和a11)的基因表达测量值组成。在这里,如果在每个转移灶中表达较高,则基因被分类为与进展相关,而如果在每个原代肿瘤中表达较高,则基因被分类为与进展负相关。另外两个数据集由具有匹配结果的原发性肿瘤基因表达谱组成。对于unc254患者数据集,复发部位不可获得,并且如果基因具有>1的显著的无复发存活率风险比(hazardratio),则认为该基因与进展正相关,而如果这些比是显著的(coxp值<0.05)并且<1,则认为该基因与进展负相关。由于unc855数据集也具有复发部位信息,此处使用无复发存活率和无肺部复发存活率两者(rfs和lrfs)的危险比来基于对unc254数据使用的相同的标准将基因分类为与进展正相关或负相关。示出了在不同数据集中测量的具有人类直向同源物的基因。
图6示出了对患者数据中asns表达水平的二次分析。a)具有不同疾病亚型的患者的原发性肿瘤中的天冬酰胺合成酶表达水平(盒的边缘是第25和等75百分位数,并且误差条延伸到值q3+w(q3-q1)和q1-w(q3-q1),其中w是1.5,并且q1和q3是第25和第75百分位数,并且对于b也是如此,anovap值<0.0001)。b)具有非特异性复发且复发至淋巴结、骨、脑、肝或肺的患者的原发性肿瘤中的天冬酰胺合成酶表达水平与未复发至每个对应部位的患者中的表达水平的比较(秩和p值<0.005)。c)对另外3个乳腺癌患者组中的asns的分析(mdacc、metrabic和tcga)。示出了存活率图和相关统计学数据(coxp值<0.001)。d)对tcga泛癌表达数据中asns的分析。示出了数据集中呈现的10种非乳腺实体肿瘤的存活率图和相关统计学数据(对于结肠癌、头颈鳞癌、肾透明细胞癌和子宫内膜癌,coxp值<0.05)。e)对tcga泛癌数据集中呈现的所有肿瘤中的asns的分析(coxp值<0.001)。
图7示出了对asns作为侵袭和转移驱动因子的初步验证。a)对asns-沉默的4t1-t细胞和表达asns的4t1-t细胞的基质胶侵袭能力的定量(n=3个重复品/细胞系)。b)在约50%的细胞被携带靶向renilla萤光素酶和天冬酰胺合成酶的shrna的表达mcherry的构建体感染后,对mcherry阳性的4t1-t细胞的定量。使细胞在图2c中描述的基质胶侵袭测定进行的24小时期间生长(n=3个重复品/细胞系)。c)相对于初始群体,asns-沉默的4t1-t细胞和表达asns的4t1-t细胞的紫色细胞标记强度。使细胞在图2c中描述的基质胶侵袭测定进行的24小时期间生长(n=3个重复品/细胞系)。d)图2d中描述的肿瘤的代表性h&e染色切片,其中asns-沉默的4t1-t细胞和表达asns的4t1-t细胞被原位注射到nsg小鼠中。e)来自携带d中显示的肿瘤的小鼠的肺的代表性h&e染色切片。
图8示出了对asns作为侵袭和转移驱动因子的二次验证。a)对由原位注射asns-沉默的亲本4t1细胞和表达asns的亲本4t1细胞产生的肿瘤的体积测量(n=10只小鼠/细胞系,盒的边缘是第25和第75百分位数,并且误差条延伸到值q3+w(q3-q1)和q1-w(q3-q1),其中w是1.5,并且q1和q3是第25和第75百分位数,并且对于b也是这种情况。b)对应于a中描述的肿瘤的肺转移的定量(秩和p值<0.002)。c)对由原位注射具有asns基础(empty)表达或强化(asns)表达的亲本4t1细胞产生的肿瘤的体积测量(n=10只小鼠/细胞系,盒的边缘是第25和第75百分位数,并且误差条延伸到值q3+w(q3-q1)和q1-w(q3-q1),其中w是1.5,并且q1和q3是第25和第75百分位数,并且对于d、f和g也是如此)。d)对应于c中描述的肿瘤的肺转移的定量(秩和p值<0.02)。e)d中描述的肺的代表性h&e染色切片。f)由原位注射具有asns基础(empty)表达或强化表达的mda-mb-231细胞产生的肿瘤的体积测量(n=每次重复10只小鼠)。g)对对应于f中描述的肿瘤的肺转移的定量(秩和p值<0.02)。h)对f中描述的mda-mb-231来源的细胞系的基质胶侵袭的定量(n=3个侵袭室/细胞系)。i)用于h中描述的侵袭测定的收集孔的代表性图像。
图9示出了对胞外天冬酰胺可用性影响侵袭和转移的初步验证。a)用hplc测量asns-沉默的细胞和表达asns的细胞的细胞游离氨基酸的百分比组成。示出了这些中的每种氨基酸的百分比的log2倍数改变(n=3个重复品/细胞系,经验贝叶斯调整的t-检验fdr<0.05)。b)在约50%的细胞被携带靶向renilla萤光素酶和asns的shrna的表达mcherry的构建体感染后,对mcherry阳性的4t1-t细胞的定量。在感染后,使细胞在补充了l-天冬酰胺或d-天冬酰胺的培养基中生长,并且在48小时和96小时测量mcherry百分比(n=3个重复品/细胞系,误差条代表1个标准偏差,并且对于c也是如此。c)当在补充有l-天冬酰胺和未补充l-天冬酰胺的培养基中测定时,对asns-沉默的细胞和表达asns的细胞的基质胶侵袭的定量。d)当培养基中补充有dmem培养培养基中不含的每一种非必需氨基酸时,对亲本4t1细胞的细胞游离氨基酸百分比的hplc定量(n=3个重复品/细胞系)。e)在与图3b中描述的相同条件下对mda-mb-231基质胶侵袭率的定量(n=5个侵袭室/条件,秩和p-值<0.001)。f)当在d中描述的培养基条件下培养时,对mda-mb-231细胞的细胞游离氨基酸百分比的hplc定量(n=3个重复品/细胞系)。g)当亲本4t1细胞持续图3b中描述的基质胶侵袭测定进行的24小时时间段在不含天冬酰胺和补充了天冬酰胺的培养基中生长时,该亲本4t1细胞的紫色细胞标记强度(n=3个重复品/细胞系)。h)当mdamb-231细胞持续e中描述的基质胶侵袭测定所进行的24小时时间段在不含天冬酰胺和补充天冬酰胺的培养基中生长时,mdamb-231细胞的紫细胞标记强度(n=3个重复品/细胞系)。
图10示出了对胞外天冬酰胺可用性影响侵袭和转移的二次验证。a)由原位注射亲本4t1细胞产生的肿瘤体积。一半的小鼠接受l-天冬酰胺酶,而另一半以相同注射速率接受相同体积的pbs(n=10只小鼠/条件,盒的边缘是第25和第75百分位数,并且误差条延伸值q3+w(q3-q1)和q1-w(q3-q1),其中w是1.5,并且q1和q3是第25和第75百分位数,并且对于b、d、e和f也是如此。b)对在a)中描述的动物中检测到的肺转移的定量(秩和p值<0.0002)。c)如a中描述的肺的代表性h&e染色切片。d)对应于图3c&图3d中描述的肺转移灶的肿瘤体积,其中asns-沉默的4t1-t细胞和表达asns的4t1-t细胞被注射到小鼠中。一半的小鼠接受l-天冬酰胺酶,而另一半以相同注射速率接受相同体积的pbs(n=10只小鼠/条件,对于l-天冬酰胺酶治疗的小鼠中的asns-沉默的细胞与表达asns的细胞,秩和p值<0.05)。e)由原位注射asns沉默的mda-mb-231细胞和表达asns的mda-mb-231细胞产生肿瘤并随后用l-天冬酰胺酶或pbs治疗被注射的动物的肿瘤体积(n=10只小鼠/细胞系)。f)对应于e中描述的原位肿瘤的肺转移(对于处于两种治疗的asns-沉默的细胞与表达asns的细胞以及对于pbs与l-天冬酰胺酶治疗的动物的每种细胞系,秩和p值<0.05)。
图11示出了对胞外天冬酰胺可用性影响侵袭和转移的三次验证。a)喂食0%、0.6%或4%天冬酰胺膳食的小鼠的血清游离氨基酸池中的天冬酰胺含量(%)(n=5只小鼠/膳食,盒的边缘是第25和第75百分位数,并且误差条延伸到值q3+w(q3-q1)和q1-w(q3-q1),其中w是1.5,并且q1和q3是第25和第75百分位数,并且对于b、d、e和h也是如此,每种膳食之间的秩和p值<0.05)。b)对应于图3e中描述的肺转移的原位肿瘤(orthotropictumour)的体积,其中asns-沉默的4t1-t细胞和表达asns的4t1-t细胞被原位注射到喂食0%、0.6%和4%天冬酰胺膳食的小鼠中(n=10只小鼠/条件)。c)针对图3e描述的肺转移的代表性图像,其也对应于b中描述的小鼠。d)由将亲本4t1细胞原位注射到喂食0%、0.6%或4%天冬酰胺膳食的小鼠中产生的肿瘤的体积(n=10只小鼠/膳食)。e)对在d中描述的动物的肺部转移的定量(每种膳食之间的秩和p值<0.05)。f)e中描述的肺的h&e染色切片的代表性图像。g)用l-天冬酰胺酶或pbs治疗的小鼠的乳腺、血清和肺中的asns的相对表达,通过用两个引物对的qpcr测量(n=4/条件,误差条延伸到1个标准偏差,组织之间的秩和p值<0.05)。h)对asns在人类乳腺、肺和全血样品中的表达的测量,每千个碱基的转录每百万映射读取的读段(readsperkilobaseoftranscriptpermillionmappedreads)(rpkm)(对于每种组织,n>90,组织之间的秩和p值<0.05)。
图12示出了对天冬酰胺可用性调节emt的初步验证。a)当对基因按转录水平改变(基于asns-沉默的细胞的改变水平,分别被基因上调和基因下调的前10%和后10%的基因)和天冬酰胺含量(基于天冬酰胺含量,分别为asp-高和asp-低的前10%和后10%的基因,盒的边缘分别是第25和第75百分位数,并且误差条延伸到值q3+w(q3-q1)和q1-w(q3-q1),其中w是1.5,并且q1和q3是第25和第75百分位数,对于两个个体变量,秩和p值<0.001,并且对于交互变量,秩和p值<0.05)分级时,asns-沉默的细胞和表达asns的细胞之间的蛋白水平改变。b)基于由asns-沉默诱导的rna和蛋白水平表达改变,对基因的氨基酸富集的分析。负相关的氨基酸是其丰度与由沉默诱导的对应rna或蛋白水平改变呈负相关的那些氨基酸,而正相关的氨基酸与这些表达改变呈正相关。c)鼠emt-上调蛋白中的氨基酸富集。d)人类emt-上调蛋白中的氨基酸富集。e)对携带至少10个直向同源物的直向同源物matrix数据库中列出的126种物种中的促进上皮向间充质转变的蛋白直向同源物的天冬酰胺富集分析(对于所有物种,符号秩p值<1.0x10-13,并且对于哺乳动物对比其他物种,符号秩p值<9.0x10-9)。
图13示出了对天冬酰胺可用性调节emt的二次验证。a)在4t1-t细胞中响应于asns-沉默而发生的emt-上调基因和emt-下调基因的转录水平改变(n=2个重复品/条件,盒的边缘是第25和第75百分位数,并且误差条延伸到值q3+w(q3-q1)和q1-w(q3-q1),其中w是1.5,并且q1和q3是第25和第75百分位数,并且对于b、c、f和g也是如此,对于emt-上调基因,符号秩p值<0.001)。b)响应于亲本4t1细胞的培养基被补充天冬酰胺而发生的emt-上调基因和emt-下调基因的转录水平改变(n=2个重复品/条件,对于emt-上调基因,符号秩p值<0.005)。c)由原位注射tgf-β-沉默的4t1-t细胞和表达tgf-β的4t1-t细胞产生的肿瘤的体积(n=10只小鼠/细胞系)。d)基于对来自c中描述的肿瘤的切片的ihc染色的twist1阳性区域百分比(对于asns-沉默的细胞和表达asns的细胞,n=5个肿瘤切片/细胞系,误差条延伸到1个标准偏差,并且对于e也是如此,秩和p值<0.01)。e)基于对来自c中描述的肿瘤的切片的ihc染色的cdh1阳性区域百分比(对于asns-沉默的细胞与表达asns的细胞,n=5个肿瘤切片/细胞系,秩和p值<0.01)。f)对由c中描述的肿瘤产生的转移的定量(秩和p值<0.05)。g)对由静脉内注射tgf-β-沉默的细胞和表达tgf-β的细胞产生的转移的定量(每细胞系10只小鼠,秩和p值<0.05)。
图14示出了对天冬酰胺可用性调节emt的三次验证。a)对来自图4f&图4g中描述的肺的切片上的twist1和cdh1进行ihc染色的代表性图像,其中小鼠被原位注射了asns沉默的4t1-t细胞和表达asns的4t1-t细胞。b)通过qpcr测量的a中描述的肿瘤和肺中的相对twist1表达(n=2个肿瘤和肺/细胞系,误差条延伸至1个标准偏差,并且对于c、d、e、f和g也是如此)。c)通过qpcr测量的a中描述的肿瘤和肺中的相对cdh1表达(n=2个肿瘤和肺/细胞系)。d)对由向用pbs或l-天冬酰胺酶治疗的动物原位注射表达asns的细胞和asns-沉默的细胞产生的肿瘤中的twist1阳性区域的定量(n=5个肿瘤切片/条件,秩和p值<0.01)。e)对d中描述的由向喂食0%、0.6%或4%天冬酰胺的动物原位注射表达asns的细胞和asns-沉默的细胞而产生的肿瘤中的cdh1阳性区域的定量(n=5个肿瘤切片/条件,秩和p值<0.01)。f)对由对被喂食0%、0.6%或4%天冬酰胺膳食的小鼠原位注射表达asns的细胞和asns-沉默的细胞而产生的肿瘤中的twist1阳性区域的定量(n=5个肿瘤切片/条件,在asns-沉默的细胞和表达asns的细胞之间以及在膳食之间,秩和p值<0.01)。g)对f中描述的肿瘤中的cdh1阳性区域的定量(n=5个肿瘤切片/条件,在asns-沉默的细胞和表达asns的细胞之间以及膳食之间,秩和p值<0.01)。h)在细胞从a中描述的肿瘤和转移灶被分离后培养的该细胞的图像。
详细描述
本发明人令人惊讶地发现,基本上不含天冬酰胺的膳食可在延缓或抑制患有增殖性紊乱诸如癌症的受试者中的转移方面具有效用。具体地,从小鼠膳食中去除天冬酰胺显著减少来自原位肿瘤的转移。不希望受理论束缚,本发明人已经表明天冬酰胺限制减少促进上皮向间充质转变的蛋白的产生,这可能是天冬酰胺的可用性调节转移进展的一种可能机制。此外,天冬酰胺限制可以减少受试者中的癌症干细胞表型。
―生物可利用度”意指进入循环的天冬酰胺的比例。
合适地,本发明可以涉及用基本上不含天冬酰胺的处方膳食部分或完全替代患有癌症的受试者的正常膳食。这样的膳食可以潜在地通过提供本文详述的膳食产品,或者通过两种或更多种可以同时地或顺序施用的膳食补充剂来实现。潜在地,这样的膳食可以通过适当的食物选择来进一步补充,使用目前可获得的成分使得膳食保持基本上不含天冬酰胺。
合适地,膳食产品可以被配制成使得其提供所需的每日摄入量的必需氨基酸。本领域普通技术人员将容易认识到,这可以通过各种方式实现,并且将取决于膳食产品的施用方案。例如,如果膳食产品以每日单次剂量施用,则该剂量将至少包括每日推荐量的必需氨基酸,而如果每日施用3次,则组合的三次施用将至少提供每日推荐量的必需氨基酸。
合适地,可以在癌症治疗之前使用降低血清天冬酰胺水平的膳食或其他药物,以使癌细胞对进一步的治疗敏感。本发明人已经令人惊讶地证明,膳食天冬酰胺含量与原发性肿瘤中的上皮向间充质特征(signature)正相关。通过降低癌细胞中的天冬酰胺水平,可以防止或逆转上皮细胞向间充质转变,增加癌细胞对治疗的敏感性。
膳食产品
在本发明的第一方面,提供了一种包含多种氨基酸的膳食产品,其中膳食产品包含所有必需氨基酸并且其中膳食产品至少基本上不含天冬酰胺。
―必需氨基酸”是指甲硫氨酸、亮氨酸、苯丙氨酸、异亮氨酸、缬氨酸、赖氨酸、苏氨酸、组氨酸和色氨酸。
―膳食产品”是指包含一种或更多种必需氨基酸或其盐或酯的组合物,其被用于食物产品中,或与食物产品组合使用或消耗,以向消耗膳食产品的受试者提供期望水平的氨基酸或其盐或酯。这些产品中的膳食成分可以包括:维生素、矿物质、草药或其他植物材料、氨基酸以及诸如酶、器官组织、腺体和代谢物的物质。在本发明中,膳食产品可以被配制成在单一组合物中或在膳食补充剂的组合中包含所有必需氨基酸,所述膳食补充剂组合地提供例如一日时程内的所有必需氨基酸。合适地,膳食补充剂基本上不含天冬酰胺。
在一些实施方案中,膳食产品是受试者作为其膳食的一部分消耗的外源性氨基酸的唯一来源。合适地,在一些方面,膳食产品可以旨在基本上或完全替代受试者的膳食。因此,在一些方面,膳食产品可以是受试者的完全膳食替代物。
有利地,用本发明的膳食产品代替对通常的氨基酸来源诸如蛋白的消耗将产生基本上不含天冬酰胺的膳食。这可以为癌症受试者提供治疗益处。
如本文使用的,根据本发明的所有方面,术语―受试者”优选地指哺乳动物,包括人类、兽医或农场动物、家养动物或宠物,以及临床研究通常使用的动物,包括非人类灵长类动物、犬和小鼠。更具体地,本发明的受试者可以是人类。
合适地,受试者可以具有带癌症干细胞表型的细胞群体。癌症干细胞是本领域已知的,并且可能不仅涉及肿瘤复发,而且还涉及致瘤性、转移性(metastization)和耐药性。
合适地,膳食产品可以包含至少9种氨基酸。合适地,膳食产品可以包含至少10种或至少11种或至少12种或至少13种或至少14种或至少15种或至少16种或至少17种或18种氨基酸。合适地,膳食产品可包含例如9种至18种氨基酸或12-18种氨基酸,或12-17种氨基酸或13-17种氨基酸或14-17种氨基酸。
合适地,膳食产品可以基本上不含一种或更多种另外的非必需氨基酸。膳食产品中可能基本上不含的另外的氨基酸可以包括(或主要由其组成或由其组成)以下氨基酸中的两种或更多种:甘氨酸、丝氨酸、半胱氨酸、酪氨酸、脯氨酸和精氨酸。可选地,除了天冬酰胺之外,膳食产品可以不含以下氨基酸中的至少两种或至少三种或至少四种或至少五种或至少六种:甘氨酸、丝氨酸、半胱氨酸、酪氨酸、脯氨酸、精氨酸、丙氨酸、天冬氨酸、谷氨酸和谷氨酰胺。
在本文中,―主要由其组成”意指,膳食产品可以不缺少对本发明的膳食产品具有重要影响(materialeffect)的另外氨基酸。―重要影响”意指显著的治疗效果,其可以被测量为以下之一:a)对癌症相对于健康细胞的特异性的显著效果;b)对延缓或抑制转移的显著效果;c)对癌细胞毒性的显著效果或d)a)-c)的任何组合。在一些方面,这可以通过比较含有和不含有特定氨基酸的膳食产品并确定该氨基酸的缺少是否具有重要影响来测量。
wo2017/144877公开了基本上不含丝氨酸和/或甘氨酸的膳食或膳食产品可用于减少增殖和/或癌细胞存活率。该专利申请还表明:基本上不含甘氨酸、丝氨酸和半胱氨酸的膳食产品在许多癌细胞系中有效抑制癌细胞增殖并增加癌细胞死亡,所述癌细胞系包括结肠直肠癌(诸如在hct116和rko中)、肝癌(hepg2)、骨肉瘤(u2os)和乳腺癌(mdamb231);基本上不含甘氨酸、丝氨酸和精氨酸的膳食产品令人惊讶地有效抑制癌结肠直肠细胞系(诸如rko和hct116)的细胞增殖和/或增加癌细胞死亡;基本上不含甘氨酸、丝氨酸和酪氨酸的膳食产品令人惊讶地有效抑制结肠直肠细胞系(诸如rko和hct116)的癌细胞增殖和/或增加癌细胞死亡,且当膳食基本上不含半胱氨酸时会产生进一步的有益效果。因此,除了基本上不含天冬酰胺之外,本发明的膳食产品基本上不含任何一种或更多种这些非必需氨基酸将是有益的。
膳食产品还可包含水平少于25mg/kg受试者体重/日或少于20mg/kg/日或少于18mg/kg/日或少于16mg/kg/日的的甲硫氨酸。
除非本文另外说明,否则本发明的膳食产品可以被配制成至少提供基于平均每日总蛋白消耗的每日推荐摄入量的必需氨基酸。
如基于平均每日总蛋白消耗,医学研究所推荐的必需氨基酸的每日摄入量为:组氨酸18mg/g消耗的蛋白;异亮氨酸25mg/g消耗的蛋白;亮氨酸55mg/g消耗的蛋白、赖氨酸51mg/g消耗的蛋白、甲硫氨酸和半胱氨酸的组合25mg/g消耗的蛋白;苯丙氨酸和酪氨酸的组合47mg/g消耗蛋白、苏氨酸27mg/g消耗的蛋白、色氨酸7mg/g消耗的蛋白和缬氨酸32mg/g消耗的蛋白。酪氨酸和半胱氨酸是非必需氨基酸。在本发明的膳食产品基本上不含酪氨酸和/或半胱氨酸的情况下,调整该膳食产品使得该膳食产品被配制成提供基于平均每日蛋白消耗至少25mg/g消耗的蛋白的量的甲硫氨酸和至少47mg/g蛋白的量的苯丙氨酸。
合适地,半胱氨酸―有限”的膳食产品是一种被配制成提供基于平均每日蛋白消耗少于推荐的每日摄入量的半胱氨酸的膳食产品。例如,半胱氨酸有限的膳食产品可以是提供少于20mg/g消耗的蛋白或少于15mg/g消耗的蛋白或少于10mg/g消耗的蛋白或少于5mg/g消耗的蛋白的膳食产品。
合适地,膳食产品可以被配制成提供有限水平的总非必需氨基酸/克蛋白消耗。例如,与推荐的每日摄入量的总非必需氨基酸/克消耗的蛋白相比,非必需氨基酸的组合的每日摄入量可以与基本上不含至少一种或至少两种或至少三种或至少四种或至少五种或至少六种或至少七种非必需氨基酸的膳食相当。
例如,医学研究所推荐对于成年人,蛋白的消耗速率为每日每公斤体重0.8克。膳食产品可以被配制成在推荐每日消耗该产品期间提供每千克体重至少0.8克蛋白。
合适地,本发明的膳食产品可以被配制成提供上述这些推荐的水平。例如,一种或更多种氨基酸可以被配制在膳食产品中,以提供基于平均每日总蛋白消耗的每日平均摄入量的至少2倍、3倍、4倍、5倍或6倍。
合适地,存在于本发明的膳食产品中的氨基酸可以是游离形式的氨基酸、前药形式的氨基酸、盐或氨基酸酯。也可以设想具有一个或更多个n-末端或c-末端修饰的氨基酸,以及同聚物、同二聚体、异聚物和异二聚体形式。
合适地,膳食产品可以被配制成每日施用一次至八次。优选地,每日一次至四次。因此,膳食产品可以被配制成适当的单位剂型。
本发明的膳食产品还可以包含一种或更多种大量营养元素和/或微量营养元素。
关于大量营养元素的指南和建议的推荐每日摄入量可见于医学研究所2002年9月发布的对于能量、碳水化合物、纤维、脂肪、脂肪酸、胆固醇、蛋白和氨基酸的膳食参考摄入(dietaryreferenceintakesforenergy,carbohydrate,fiber,fat,fattyacids,cholesterol,proteinandaminoacids)。
可以是膳食产品的另外组分的大量营养元素的非详尽列表包括:碳水化合物、纤维和脂肪(诸如n-6多不饱和脂肪酸、n-3多不饱和脂肪酸、饱和脂肪酸和反式脂肪酸以及胆固醇)。
微量营养元素的非详尽列表包括维生素a、维生素c、维生素d、维生素e、维生素k、硫胺素、核黄素、烟酸、维生素b6、叶酸、维生素b12、泛酸、生物素、胆碱、钙、铬、铜、氟化物、碘、铁、镁、钼、磷、硒、锌、钾、钠和氯化物。合适地,膳食产品可以被配制成以可接受的或推荐的每日摄入量提供这些微量营养元素,如出版物"dietaryreferenceintakes:rdaandalforvitaminsandelements”,nas.iom.foodandnutritionboard中详述的。
本文描述的膳食产品含有氨基酸失衡,通常形式为不含两种或更多种非必需氨基酸的形式,任选地补充了过剩的一种或更多种其他氨基酸。例如,基本上不含的氨基酸与其他氨基酸的平均丰度相比,可以是至少10倍、15倍、20倍、30倍、45倍、50倍、100倍或1000倍低。可以遵循膳食学家的推荐摄取低蛋白但富含其他营养元素的食物,诸如水果、蔬菜和某些坚果,确保膳食氨基酸的摄入量比率保持在预期的比率。这种膳食(包括本发明的膳食产品以及任选地基本上不含天冬酰胺的其他营养源)旨在被单独摄取或与药物治疗诸如具有抗癌活性的药物治疗组合摄取。
在一些实施方案中,本发明的膳食产品配制在两种或更多种膳食补充剂中,所述膳食补充剂一起提供本发明的膳食产品。这些可以同时或顺序施用至所述受试者,使得由膳食补充剂提供的组合的平均膳食提供本发明的膳食产品。这对增加受试者的膳食多样性可能是有利的。
膳食产品可以粉末、凝胶、溶液、悬浮液、糊状物、固体、液体、液体浓缩物、可重构的粉末、摇瓶(shake)、浓缩物、丸剂、条(bar)、片剂、胶囊或即用型产品的形式提供。设想了当补充剂为片剂、丸剂、胶囊、液体、气雾剂、可注射溶液或其他药学上可接受的制剂形式时,膳食产品也可以是药物组合物。合适地,膳食产品可以是饮料。合适地,饮料可以一日施用2次至6次。
合适地,膳食产品可以不是天然存在的食物。
合适地,膳食产品可以包含特定氨基酸之外的另外化合物。合适地,这样的另外化合物可以无助于基本上不含的氨基酸的从头合成。
如本文使用的,述及氨基酸的―基本上不含”意指完全或几乎不含(诸如痕量的)该氨基酸。
任选地,施用可以通过静脉内途径。任选地,肠胃外施用可以以弹丸(bolus)或通过输注来提供。
合适地,膳食产品可以是:
a)管喂的肠营养产品(tubefedenteralnutritionalproduct)(诸如鼻-胃营养产品,其可以通过ng管施用;鼻-空肠营养产品,其可以通过nj管施用;或者peg(经皮内窥镜胃造口术)管营养产品);
b)肠胃外营养产品(其可以通过中心静脉施用,例如通过静脉导管上的专用管腔施用);或者
c)iv输注产品。
优选地,施用可以作为食物或饮料。
在某些实施方案中,本发明的膳食或膳食产品在至少1天、2天、3天、4天、5天、6天、7天,至少2周、3周、4周、5周、6周、7周、8周、9周、10周、11周、12周、13周、14周、15周、16周、17周、18周、19周、20周、21周、22周、23周、24周、25周、26周的时间段内施用或施用至直到观察到治疗终点(例如,血清中的天冬酰胺水平有统计学上的显著降低,或者观察到癌细胞的上皮至间充质(emt)表型的减少)。
本发明还提供了制备本发明的膳食产品的方法,其中氨基酸被溶解或分散在水中并喷雾干燥。
合适地,氨基酸可以与另外的组分诸如大量营养元素和微量营养元素混合。可以根据需求加入粘合剂、乳化剂或其他适合用于人类或动物摄取的成分。
药物组合物
在另一方面,本发明提供了一种药物组合物,所述药物组合物包含本发明的膳食产品或根据本发明产生的膳食产品以及药学上可接受的运载体、赋形剂或稀释剂。
合适的药物制剂的选择和制备的常规程序在例如―pharmaceuticals-thescienceofdosageformdesigns”,m.e.aulton,churchilllivingstone,1988中描述。
本发明的组合物可以是以适合用于口服使用的形式(例如作为片剂、锭剂、硬胶囊或软胶囊、水性或油性悬浮液、乳液、可分散的粉末或颗粒、糖浆或酏剂)。
本发明的组合物可以使用本领域中熟知的常规药物赋形剂通过常规的程序获得。因此,意图口服使用的组合物可以包含例如一种或更多种着色剂、甜味剂、调味剂和/或防腐剂。
合适地,药物组合物被配制成提供治疗有效量的本发明的膳食产品。
用于在状况的治疗中使用的“有效量”是足以在温血动物,特别是人类中在症状上缓解状况的症状或减慢状况进展的量。在一些方面,―有效量”是足以降低受试者血清中的天冬酰胺水平和/或延缓或抑制转移的量。
术语―治疗有效量”涵盖化合物或组合物的量,当施用时,所述量足以防止受试者中转移的发展,或在某种程度上减轻受试者中的转移。术语―治疗有效量”还涵盖足以引起细胞、组织、器官、系统、动物或人类的生物或医学应答的化合物或组合物的量,这是研究者、医学医生或临床医生正在寻求的。在任何个例中,适当的―有效”量可以由本领域中普通技术人员使用常规实验确定。应当理解,对于任何特定患者的具体剂量水平和施用频率可以不同,并且将取决于多种因素,包括所用具体化合物的活性;该化合物的生物可利用度、代谢稳定性、排出速率和作用时长;化合物施用的方式和时间;患者的年龄、体重、一般健康状况、性别和膳食;以及正被治疗的特定状况的严重程度。
术语―治疗”("treat”)、―治疗”("treating”)和―治疗”("treatment”)涵盖减轻或消除状况、紊乱或疾病,或与该状况、紊乱或疾病相关的一种或更多种症状,并且涵盖减轻或根除该状况、紊乱或疾病本身的病因。在某些实施方案中,术语―治疗”("treat”)、―治疗”("treating”)和―治疗”("treatment”)是指为了减轻、消除或预防状况、紊乱或疾病或与其相关的症状或其病因向受试者施用化合物、药物组合物或药物剂型。
合适地,本发明的药物组合物还可以包含选自以下的治疗剂:癌细胞生长抑制剂、放射治疗剂、抗转移剂、免疫检查点抑制剂、化学治疗剂、氨基酸代谢/周转/相互转化抑制剂、非必需氨基酸生物合成抑制剂、氨基酸转运抑制剂、促进氨基酸降解的酶或药物或者螯合氨基酸的物质。
癌症治疗
在一方面,本发明提供了用于在药物中使用的本发明的膳食产品或根据本发明的方法产生的膳食产品或本发明的药物组合物。
例如,本发明提供了用于在延缓或抑制转移中使用的本发明的膳食产品或根据本发明的方法产生的膳食产品或本发明的药物组合物。
如本文使用的,术语―转移”是指癌性肿瘤在未直接与原始癌性肿瘤的器官相连的器官或身体部分中的生长。转移也可以被定义为一个过程的几个步骤,诸如癌细胞从原始肿瘤部位离开,以及癌细胞迁移和/或侵袭到身体的其他部分。因此,本发明设想了延缓或抑制一种或更多种癌性肿瘤在未直接与原始癌性肿瘤的器官相连的器官或身体部分中的进一步生长和/或导致该生长的过程中的任何步骤。
本发明令人惊讶地证明,通过降低患有癌症的患者的天冬酰胺水平(例如血清中的天冬酰胺水平),可以延缓或抑制转移。在这方面,本发明人已经证明使天冬酰胺合成酶沉默降低了体内转移潜能和体外侵袭潜能两者—例如参见图2。用l-天冬酰胺酶降低胞外天冬酰胺水平(即天冬酰胺的生物可利用度)的治疗被证明导致4t1细胞注射的nsg小鼠中的转移负荷的显著降低,并且此外,消耗膳食中的天冬酰胺也被证明导致动物中的转移负荷的降低。
因此,本发明提供了一种用于在延缓或抑制患有癌症的受试者中的转移中使用的药物,其中所述药物降低患有癌症的受试者的血液中的胞外天冬酰胺水平。在另外的方面,本发明提供了化合物或组合物在制备用于延缓或抑制患有癌症的受试者中的转移的药物中的用途,其中所述化合物或组合物降低受试者血液中的胞外天冬酰胺水平。本发明还提供了一种治疗受试者中的癌症的方法,所述方法包括向所述受试者施用治疗有效量的药物,其中所述药物降低患有癌症的受试者的血液中的胞外天冬酰胺水平。
本领域的普通技术人员容易知晓能够降低患有癌症的受试者的血液中的天冬酰胺水平的各种药物、化合物和组合物。这些包括天冬酰胺合成酶抑制剂、l-天冬氨酸酶以及如本文详述的膳食产品。
天冬酰胺合成酶抑制剂是本领域已知的,并且包括:胍基琥珀酸;草酰乙酸;l-半胱氨酸亚磺酸;氨基丙二酸二乙酯;含有l-天冬氨酸的二肽(l-天冬氨酰甘氨酸、l-天冬氨酰-l-亮氨酸、l-天冬氨酰-l-苯丙氨酸、l-天冬氨酰-l-脯氨酸、l-α-天冬氨酰-l-丝氨酸和l-α-天冬氨酰-l-缬氨酸);n-邻硝基苯基亚磺酰基-l-天冬氨酸;n-邻硝基苯基亚磺酰基-l-谷氨酰胺;s-腺苷-l-甲硫氨酸;l-高丝氨酸-β-腺苷酸;依他尼酸;莫匹罗星、phosmidosine、β-天冬氨酰腺苷酸(β-asparaginyladenylate)、βaspamp中间体的磺酰胺类似物。任何适合用于在治疗中使用的天冬酰胺合成酶都可以用于本发明。
对于所有方面,示例性癌症包括但不限于肾上腺皮质癌、aids相关癌症、aids相关淋巴瘤、肛门癌、肛门直肠癌、肛管癌、阑尾癌、儿童小脑星形细胞瘤(childhoodcerebellarastrocytoma)、儿童大脑星形细胞瘤(childhoodcerebralastrocytoma)、基底细胞癌、皮肤癌(非黑色素瘤)、胆管癌、肝外胆管癌、肝内胆管癌、膀胱癌(bladdercancer)、泌尿膀胱癌(urinarybladdercancer)、骨和关节癌、骨肉瘤和恶性纤维组织细胞瘤、脑癌、脑肿瘤、脑干胶质瘤、小脑星形细胞瘤、大脑星形细胞瘤/恶性胶质瘤、室管膜瘤、成神经管细胞瘤、幕上原始神经外胚层肿瘤(supratentorialprimitiveneuroectodermaltumors)、视觉通路和下丘脑神经胶质瘤、乳腺癌、支气管腺瘤/类癌、类癌肿瘤、胃肠道、神经系统癌、神经系统淋巴瘤、中枢神经系统癌、中枢神经系统淋巴瘤、宫颈癌、儿童癌症、慢性淋巴细胞白血病、慢性骨髓性白血病、慢性骨髓增生性紊乱、结肠癌、结肠直肠癌、皮肤t细胞淋巴瘤、淋巴肿瘤、蕈样肉芽肿、seziary综合征、子宫内膜癌、食管癌、颅外生殖细胞瘤、性腺外生殖细胞瘤、肝外胆管癌、眼癌、眼内黑色素瘤、视网膜母细胞瘤、胆囊癌(gallbladdercancer)、胃(胃)癌(gastric(stomach)cancer)、胃肠道类癌肿瘤、胃肠道间质肿瘤(gist)、生殖细胞肿瘤、卵巢生殖细胞肿瘤、妊娠滋养层肿瘤胶质瘤、头和颈癌、肝细胞(肝)癌、霍奇金淋巴瘤(hodgkinlymphoma)、下咽癌、眼内黑色素瘤、眼部癌症、胰岛细胞肿瘤(内分泌胰腺)、卡波西肉瘤(kaposisarcoma)、肾癌(kidneycancer)、肾癌(renalcancer)、肾癌(kidneycancer)、喉癌(laryngealcancer)、急性成淋巴细胞白血病、急性骨髓性白血病、慢性淋巴细胞白血病、慢性髓细胞白血病、毛细胞白血病、唇和口腔癌症、肝癌、肺癌、非小细胞肺癌、小细胞肺癌、aids相关淋巴瘤、非霍奇金淋巴瘤(non-hodgkinlymphoma)、原发性中枢神经系统淋巴瘤、waldenstram巨球蛋白血症、髓母细胞瘤、黑色素瘤、眼内(眼)黑色素瘤、默克尔细胞癌、间皮瘤恶性肿瘤、间皮瘤、转移性鳞状颈部癌、口癌、舌癌、多内分泌瘤形成综合征、蕈样肉芽肿、骨髓发育不良综合征、骨髓增生异常/骨髓增生性疾病、慢性骨髓性白血病、急性髓细胞白血病、多发性骨髓瘤、慢性骨髓增生性紊乱、鼻咽癌、神经母细胞瘤、口癌(oralcancer)、口腔癌(oralcavitycancer)、口咽癌(oropharyngealcancer)、卵巢癌、卵巢上皮癌、卵巢低恶性潜能肿瘤、胰腺癌、胰岛细胞胰腺癌、副鼻窦和鼻腔癌、甲状旁腺癌、阴茎癌、咽癌(pharyngealcancer)、嗜铬细胞瘤、成松果体细胞瘤和幕上原始神经外胚层肿瘤、垂体肿瘤、浆细胞瘤/多发性骨髓瘤、胸膜肺母细胞瘤、前列腺癌、直肠癌、肾盂和输尿管、移行细胞癌(transitionalcellcancer)、视网膜母细胞瘤、横纹肌肉瘤、涎腺癌、尤文肉瘤家族肿瘤(ewingfamilyofsarcomatumor)、卡波西肉瘤(kaposisarcoma)、软组织肉瘤、子宫癌、子宫肉瘤、皮肤癌(非黑色素瘤)、皮肤癌(黑色素瘤)、默克尔细胞皮肤癌、小肠癌、软组织肉瘤、鳞状细胞癌、胃(胃)癌(stomach(gastric)cancer)、幕上原始神经外胚层肿瘤、睾丸癌、喉癌、胸腺瘤、胸腺瘤和胸腺癌、甲状腺癌、肾盂和输尿管和其他泌尿器官的移行细胞癌、妊娠滋养层肿瘤、尿道癌、子宫内膜子宫癌、子宫肉瘤、子宫体癌、阴道癌、外阴癌和威尔姆肿瘤(wilm'stumor)。合适地,癌症可以选自由以下组成的组:乳腺癌、结肠癌、头颈鳞癌、肾透明细胞癌和子宫内膜癌。
膳食产品可以基本上不含半胱氨酸。合适地,基本上不含半胱氨酸的膳食可以在大量消耗外源性半胱氨酸的癌症诸如肺癌、结肠直肠癌和乳腺癌中具有效用。合适地,基本上不含半胱氨酸的膳食可以在mtap表达被下调的癌症中具有效用。
膳食产品可以基本上不含丝氨酸和/或甘氨酸。合适地,基本上不含丝氨酸和/或甘氨酸的膳食可以在依赖大量消耗外源性丝氨酸和/或甘氨酸的癌症诸如肺癌、结肠直肠癌和乳腺癌、淋巴瘤、结肠直肠癌、肝癌、骨肉瘤和乳腺癌中具有效用。
膳食产品可以基本上不含精氨酸和/或酪氨酸。合适地,基本上不含精氨酸的膳食可以在癌症诸如结肠直肠癌中具有效用。
组合治疗
降低受试者血液中的天冬酰胺水平的药物、化合物和组合物(诸如本发明的膳食产品或药物组合物)可以单独使用以提供治疗效果(例如延缓或抑制转移)。合适地,本发明的膳食产品或药物组合物也可以与癌细胞生长抑制剂、抗转移剂、免疫检查点抑制剂、放射治疗剂和/或化学治疗中的一种或更多种组合使用。
此类化学治疗可以包括以下类别的抗癌剂中的一种或更多种:
(i)抗增殖药/抗肿瘤药及其组合,诸如,烷基化剂(例如顺铂、奥沙利铂、卡铂、环磷酰胺、氮芥、尿嘧啶氮芥、苯达莫司汀、美法仑、苯丁酸氮芥、盐酸氮芥、白消安、替莫唑胺、亚硝基脲、异环磷酰胺(ifosamide)、美法仑、哌泊溴烷、三乙撑蜜胺、三乙撑硫代磷酰胺(triethylenethiophoporamine)、卡莫司汀、洛莫司汀、链佐星和达卡巴嗪);抗代谢物(例如,吉西他滨和叶酸拮抗物诸如氟嘧啶如5-氟尿嘧啶和替加氟、雷替曲塞、氨甲蝶呤、培美曲塞、胞嘧啶阿拉伯糖苷、氟尿苷、阿糖孢苷、6-巯基嘌呤、6-硫代鸟嘌呤、氟达拉滨磷酸、喷司他丁(pentostatin)、和吉西他滨和羟基脲);抗生素(例如,蒽环霉素如阿霉素、博来霉素、多柔比星、柔红霉素、表柔比星、伊达比星、丝裂霉素-c、更生霉素和光辉霉素);抗有丝分裂剂(例如长春花生物碱如长春新碱、长春花碱、长春地辛和长春瑞滨,以及紫杉烷如紫杉醇和泰索帝(taxotere),以及polo激酶(polokinase)抑制剂);蛋白酶体抑制剂例如卡非佐米和硼替佐米;干扰素治疗;和拓扑异构酶抑制剂(例如,表鬼臼毒素如依托泊苷和替尼泊苷、安吖啶、拓扑替康、伊立替康、米托蒽醌以及喜树碱);博来霉素、更生霉素、柔红霉素、多柔比星、表柔比星、伊达比星、ara-c、紫杉醇(taxoltm)、白蛋白结合型紫杉醇(abpaclitaxel)、多西紫杉醇(docetaxel)、光神霉素、脱氧助间型霉素(deoxyco-formycin)、丝裂霉素-c、l-天冬酰胺酶、干扰素(特别是ifn-a)、依托泊苷、替尼泊苷、dna脱甲基化剂(例如,阿扎胞苷或地西他滨);以及组蛋白去乙酰化酶(hdac)抑制剂(例如伏立诺他(vorinostat)、ms-275、帕比司他(panobinostat)、罗米地(romidepsin)、丙戊酸、莫西汀(mgcd0103)和普拉西诺司他sb939(prasinostatsb939));
(ii)细胞生长抑制剂诸如抗雌激素(例如他莫昔芬、氟维司群、托瑞米芬、雷洛昔芬、屈洛昔芬和艾多昔芬(iodoxyfene))、抗雄激素(例如比卡鲁胺、氟他胺、尼鲁米特和醋酸环丙孕酮)、lhrh拮抗剂或lhrh激动剂(例如戈舍瑞林、亮丙瑞林和布舍瑞林)、孕激素(例如醋酸甲地孕酮)、芳香化酶抑制剂(例如阿那曲唑、来曲唑、伏氯唑(vorazole)和依西美坦)以及5*还原酶抑制剂诸如非那雄胺;和诺维本(avelbene)、cpt-ll、阿那曲唑、来曲唑、卡培他滨、reloxafme、环磷酰胺、异环磷酰胺和屈洛昔芬(droloxafine);
(iii)抗侵入剂例如达沙替尼和博舒替尼(ski-606),以及金属蛋白酶抑制剂、尿激酶纤维蛋白溶酶原激活剂受体功能(urokinaseplasminogenactivatorreceptorfunction)的抑制剂或乙酰肝素酶(heparanase)的抗体;
(iv)生长因子功能的抑制剂:例如,这样的抑制剂包括生长因子抗体和生长因子受体抗体例如抗erbb2抗体曲妥珠单抗(trastuzumab)[赫塞汀tm]、抗egfr抗体帕尼单抗、抗erbb1抗体西妥昔单抗,酪氨酸激酶抑制剂例如表皮生长因子家族的抑制剂(例如,egfr家族酪氨酸激酶抑制剂例如吉非替尼、埃罗替尼和6-丙烯酰胺基-n-(3-氯-4-氟苯基)-7-(3-吗啉代丙氧基)-喹唑啉-4-胺(ci1033)、阿法替尼、范德塔尼、奥西默蒂尼和罗基列替尼),erbb2酪氨酸激酶抑制剂诸如拉帕替尼)和共刺激分子诸如ctla-4、4-lbb和pd-1的抗体,或细胞因子抗体(il-10、tgf-β);肝细胞生长因子家族的抑制剂;胰岛素生长因子家族的抑制剂;细胞凋亡的蛋白调节物的调节剂(例如bcl-2抑制剂);血小板衍生的生长因子家族的抑制剂,例如伊马替尼和/或尼罗替尼(amn107);丝氨酸/苏氨酸激酶的抑制剂(例如,ras/raf信号传送抑制剂例如法尼基转移酶抑制剂,索拉非尼、替吡法尼和洛那法尼),通过mek和/或akt激酶的细胞信号传送的抑制剂、c-kit抑制剂、abl激酶抑制剂、pi3激酶抑制剂、plt3激酶抑制剂、csf-1r激酶抑制剂、igf受体激酶抑制剂;极光激酶抑制剂和周期蛋白依赖性激酶抑制剂诸如cdk2抑制剂和/或cdk4抑制剂;ccr2、ccr4或ccr6拮抗剂;和raf激酶抑制剂,如在wo2006043090、wo2009077766、wo2011092469或wo2015075483中所述的那些。
(v)抗血管生成剂,例如抑制血管内皮生长因子的效应的那些抗血管生成剂,[例如抗血管内皮细胞生长因子抗体贝伐单抗(avastintm)];沙利度胺;来那度胺;和例如vegf受体酪氨酸激酶抑制剂诸如凡德他尼、瓦他拉尼(vatalanib)、舒尼替尼、阿西替尼和帕唑帕尼;
(vi)基因治疗方法,包括例如替换畸变基因例如畸变p53或畸变的brca1或brca2的方法;
(vii)免疫治疗方法,包括例如抗体治疗剂,诸如阿仑单抗、利妥昔单抗、替伊莫单抗(ibritumomabtiuxetan)
(viii)细胞毒性剂例如氟达拉滨(fludaribine)(福达华(fludara))、克拉屈滨(cladribine)、喷司他丁(pentostatin)(nipenttm);
(ix)靶向治疗剂,例如pi3k抑制剂,例如艾代拉里(idelalisib)和培福辛(perifosine);smac(胱天蛋白酶的第二线粒体驱动的活化剂)模拟物,也称为凋亡蛋白抑制剂(iap)拮抗剂(iap拮抗剂)。这些剂起到抑制lap例如xiap、clap1和clap2的作用,并且从而重建细胞凋亡途径。特定的smac模拟物包括birinapant(tl32711,tetralogicpharmaceuticals)、lcl161(novartis)、aeg40730(aegeratherapeutics)、sm-164(universityofmichigan)、lbw242(novartis)、ml101(sanford-burnhammedicalresearchinstitute)、at-406(ascentatherapeutics/universityofmichigan)、gdc-0917(genentech)、aeg35156(aegeratherapeutic)和hgs1029(humangenomesciences);以及靶向泛素蛋白酶体系统(ups)的剂,例如硼替佐米、卡菲佐米、马里佐米(npi-0052)和mln9708;以及
(xii)嵌合抗原受体、抗癌疫苗和精氨酸酶抑制剂。
术语―免疫检查点”是指cd4+t细胞和/或cd8+t细胞的细胞表面上的通过下调或抑制抗肿瘤免疫应答来对免疫应答进行微调的一组分子。免疫检查点蛋白是本领域熟知的,并且包括但不限于,ctla-4、pd-1、vista、b7-h2、b7-h3、pd-ll、b7-h4、b7-h6、2b4、icos、hvem、pd-l2、cd160、gp49b、pir-b、kir家族受体、tim-1、tim-3、tim-4、lag-3、btla、sirpα(cd47)、cd48、2b4(cd244)、b7.1、b7.2、ilt-2、ilt-4、tigit、hhla2、嗜乳脂蛋白(butyrophilin)和a2ar(参见,例如,wo2012/177624)。该术语还涵盖生物活性蛋白片段,以及编码全长免疫检查点蛋白及其生物活性蛋白片段的核酸。在一些实施方案中,该术语还涵盖根据本文提供的同源性描述的任何片段。―免疫检查点抑制剂”是指抑制免疫检查点核酸和/或蛋白的剂。对一种或更多种免疫检查点的抑制可以阻断或以其他方式中和抑制性信号,从而上调免疫应答,以更有效地治疗癌症。可用于抑制免疫检查点的示例性剂包括抗体、小分子、肽、肽模拟物、天然配体和天然配体的衍生物(其可以结合和/或灭活或抑制免疫检查点蛋白或其片段);以及rna干扰剂、反义核酸、核酸适配体等(其可以下调免疫检查点核酸或其片段的表达和/或活性)。用于上调免疫应答的示例性剂包括针对一种或更多种免疫检查点蛋白的阻断这些蛋白和其天然受体之间的相互作用的抗体;一种或更多种免疫检查点蛋白的非激活形式(例如,显性失活(dominantnegative)多肽);阻断一种或更多种免疫检查点蛋白和其天然受体之间的相互作用的小分子或肽;与其天然受体结合的融合蛋白(例如,免疫检查点抑制蛋白的胞外部分与抗体或免疫球蛋白的fc部分融合);阻断免疫检查点核酸转录或翻译的核酸分子等。这样的剂可以直接阻断一种或更多种免疫检查点和其天然受体(例如抗体)之间的相互作用,以防止抑制性信号传导并上调免疫应答。可选地,剂可以间接阻断一种或更多种免疫检查点蛋白和其天然受体之间的相互作用,以阻止抑制性信号传导并上调免疫应答。例如,免疫检查点蛋白配体的可溶性形式,诸如稳定的胞外结构域可以与其受体结合,以间接降低受体与适当配体结合的有效浓度。合适地,抗pd-1抗体、抗pd-ll抗体和抗ctla-4抗体(单独地或组合地)可以用于抑制免疫检查点。
术语―抗转移剂”意指抑制、降低或减少癌细胞的转移的物质。抗转移剂包括抑制或降低血管生成、组织因子活性、因子vlla活性或组织因子/因子vlla复合物活性的物质。实例包括veg抑制剂、抗vegf抗体(即贝伐单抗,avastin)、非抗凝性肝素、低分子量肝素(lmwh,诸如lovenox)、抗组织因子抗体、cra-5(抗fvlla)、bms262084、ttp889、蛋白c、apc(drotrecogin)、stm、艾卓肝素(idraparinux)、dx9065a、bay59-7939、ly-51,7717、bms-562247、du-176b、奥米沙班、雷扎沙班和nap蛋白。
合适地,本发明的组合物可以与一种或更多种消耗氨基酸的治疗性酶组合使用。可将这样的治疗性酶与本发明的组合物关联。例如,对于基本上不含精氨酸的组合物,可以使用诸如精氨酸酶的治疗性酶。
另外,或者在替代方案中,本发明的组合物可以与参与抑制氨基酸从头合成的一种或更多种化合物组合使用。可将这样的化合物与本发明的组合物关联。例如,基本上不含天冬酰胺的膳食产品可以与天冬酰胺合成酶抑制剂组合使用。
本发明方法中使用的治疗剂可以是单一的剂或剂的组合。优选的组合将包括具有不同作用机制的剂。
在本文中,在使用术语―组合”的情况下,应理解的是,这是指同时、分开或顺序施用。在本发明的一个方面,―组合”是指同时施用。在本发明的另一个方面,―组合”是指分开施用。在本发明的另外的方面,―组合”是指顺序施用。在施用是顺序性的或分开的情况下,施用第二组分的延缓不应当使得损失组合的有益效果。
在一方面,降低血液中的天冬酰胺水平的药物、化合物或组合物(即剂)以在降低血清天冬酰胺水平或降低emt表型方面获得期望的治疗终点的给药方案施用,并且随后向受试者施用选自癌细胞生长抑制剂、抗转移剂、免疫检查点抑制剂、放射治疗剂和化学治疗剂的治疗剂,直到达到癌细胞生长或增殖方面的期望治疗终点。
例如,给药方案的治疗终点可能导致:
·血清中的天冬酰胺降低至少10%或至少20%或至少25%或至少30%或至少40%或至少45%或至少50%;
·血清游离氨基酸池中的天冬酰胺含量(%)少于0.4%或少于0.38%或少于0.36%或少于0.32%或少于0.3%;
·肿瘤细胞群体中的twist、上皮细胞钙粘蛋白和snail中的至少一种或更多种(例如,至少两种或全部三种)降低至少20%或至少30%或至少40%或至少50%或至少60%或至少70%或至少80%或至少90%。
如本文使用的,术语―与...组合施用”以及语法等同形式等意在涵盖向单个患者施用选择的治疗剂,且意图包括其中通过相同或不同的施用途径或在相同或不同的时间施用这些剂的治疗方案。在一些实施方案中,本文描述的化合物将与其他剂共同施用。这些术语涵盖向动物施用两种或更多种剂使得剂和/或其代谢物都同时存在于动物中。其包括以分开的组合物同时施用,以分开的组合物在不同时间施用,和/或以其中存在这两种剂的组合物施用。
本文公开的剂可以通过任何途径施用,包括皮内、皮下、口服、动脉内或静脉内施用。
在使用组合治疗的一些实施方案中,当组合时,本发明的膳食产品或药物组合物的量和其他药物活性剂的量对于治疗患者中的目标紊乱是治疗有效的。在这种情况下,当组合时,如果组合的量足以降低或完全减轻紊乱的症状或其他有害影响;治愈紊乱;逆转、完全停止或减慢紊乱的进展;延缓或降低紊乱的转移或者降低紊乱变恶化的风险,则组合的量是―治疗有效量”。通常,本领域技术人员可以通过例如从本说明书对于本发明化合物描述的剂量范围和其他药物活性化合物的已批准或以其他方式公布的剂量范围出发来确定这样的量。
根据本发明的另外的方面,提供了用于在癌症的联合治疗中使用的如上文定义的本发明的膳食产品或药物组合物和如上文定义的另外的抗癌剂。
根据本发明的另外的方面,提供了一种治疗患有癌症的人类或动物受试者的方法,所述方法包括与如本文前面定义的另外的抗癌剂同时地、顺序地或分开地向受试者施用治疗有效量的本发明的膳食产品或药物组合物。
根据本发明的另外的方面,提供了用于治疗癌症的本发明的膳食产品或药物组合物,其与本文前面定义的另外的抗癌剂同时地、顺序地或分开使用。
本发明的膳食产品或药物组合物也可以与放射治疗组合使用。合适的放射疗法治疗包括例如x射线治疗、质子束治疗或电子束治疗。放射治疗还可以涵盖使用放射性核素剂,例如131i、32p、90y、89sr、153sm或223ra。这样的放射性核素治疗剂是熟知的,并且可商购。
根据本发明的另外的方面,提供了用于与放射治疗联合地在癌症治疗中使用的如本文前面定义的本发明的膳食产品或药物组合物或其药学上可接受的盐。
根据本发明的另外的方面,提供了一种治疗患有癌症的人类或动物受试者的方法,所述方法包括与放射治疗同时地、顺序地或分开地向受试者施用治疗有效量的本发明的膳食产品或药物组合物或其药学上可接受的盐。
在一方面,每种化学治疗剂的剂量(或化学治疗剂的总组合剂量)可以相当于至少0.1g/kg患者体重/日,优选地至少0.2g/kg/日或0.3g/kg/日或0.4g/kg/日或0.5g/kg/日。合适地,化学治疗剂(或组合的化学治疗剂的组合)的剂量可以相当于至少1g/kg/日,优选地2g/kg/日。
此外,在另一方面,本发明提供了一种治疗受试者中的癌症的方法,所述方法包括施用:a)本发明的膳食产品以及b)化学治疗剂的协同有效的组合。
在某些实施方案中,基本上不含天冬酰胺的膳食包括膳食产品或由膳食产品组成。
根据本发明待施用的治疗剂的浓度将根据几个因素而变化,所述因素包括待施用的化合物的剂量、所用化合物的药代动力学特征和施用途径。剂可以单剂量或重复剂量施用。根据许多因素,包括患者的总体健康状况,以及选择的化合物的制剂和施用途径,治疗可以每日施用或更频繁地施用。
优选地,所述癌症治疗还包括施用治疗有效量的所述治疗剂。如本文使用的,术语―治疗有效量”是指被施用的至少一种剂或化合物的足以治疗或预防特定疾病或状况的量。结果可以是疾病的病征、症状或病因的减少和/或减轻,或任何其他期望的对生物系统的改变。例如,用于治疗用途的―有效量”是包含如本文公开的化合物的组合物提供疾病的临床上显著减轻所需要的量。在任何个体的情况下,可以使用诸如剂量递增研究(doseescalationstudy)的技术来确定合适的―有效”量。
在某些实施方案中,所述膳食在至少1天、2天、3天、4天、5天、6天、7天,至少2周、3周、4周、5周、6周、7周、8周、9周、10周、11周、12周、13周、14周、15周、16周、17周、18周、19周、20周、21周、22周、23周、24周、25周、26周的时间段内或施用至直到观察到治疗终点。
在基本上不含天冬酰胺的膳食包括膳食产品或由膳食产品组成的情况下,膳食产品被每日施用1次至10次。
分级(stratification)
本发明令人惊讶地证明,受试者的原发性肿瘤中升高的天冬酰胺合成酶水平与晚期转移性复发强烈相关,并且患有癌症的受试者的血液中的天冬酰胺水平升高有助于转移。
因此,血清天冬酰胺水平或天冬酰胺合成酶表达(例如在原发性肿瘤中)可以被用作生物标记来鉴定具有处于增加的转移风险的肿瘤的患者或患者群体。
因此,本发明提供了一种鉴定患有具有增加的转移可能性的癌症的受试者的方法,所述方法包括:
a)确定从受试者分离的生物样品的天冬酰胺水平;
b)将生物样品的天冬酰胺水平与对照样品或天冬酰胺的预定参考水平进行比较,
其中生物样品的天冬酰胺水平与对照样品相比或与预定参考水平相比增加指示转移可能性增加。该方法还可以包括向所述受试者施用治疗有效量的药物,其中所述药物降低患有癌症的受试者的血液中的天冬酰胺水平。
本发明还提供了一种鉴定在进食基本上不含天冬酰胺的膳食或被施用l-天冬酰胺酶时对癌症治疗具有增加的响应性或敏感性的可能性的受试者的方法,所述方法包括:
a)确定从受试者分离的生物样品的天冬酰胺水平;
b)将生物样品的天冬酰胺水平与对照样品或天冬酰胺的预定参考水平进行比较,
其中生物样品中的天冬酰胺水平与对照样品相比或与预定参考水平相比增加指示在所述癌症治疗结合基本上不含天冬酰胺的膳食或与l-天冬酰胺酶一起施用时,对所述癌症治疗的响应性或敏感性。
本发明还提供了一种确定具有癌症的受试者中的转移性复发可能性的方法,所述方法包括:
a)确定从受试者分离的生物样品中的天冬酰胺合成酶水平;
b)将生物样品中的天冬酰胺合成酶水平与对照样品或天冬酰胺合成酶的预定参考水平进行比较,
其中生物样品中的天冬酰胺合成酶水平与对照样品相比或与预定参考水平相比增加指示转移性复发可能性增加。
这样的方法还可以包括向所述受试者施用治疗有效量的药物,其中所述药物降低患有癌症的受试者的血液中的天冬酰胺水平。
如本文使用的,术语―生物样品”和―从受试者分离的样品”可互换使用,指从患者分离的组织、细胞和生物流体,以及存在于患者体内的组织、细胞和流体。样品可以是尿液样品、血液样品、血清样品、痰样品、粪便样品、身体组织的活检物例如移植肾组织的活检物、脑脊液样品、精液样品或涂片样品。优选的样品是血清或血浆。
如本文使用的,―参考水平”或―对照”是指具有正常水平的天冬酰胺/日冬酰胺合成酶表达的样品,例如来自不患有或怀疑不患有癌症的健康受试者的样品,或者关于天冬酰胺合成酶表达,是来自不受癌症影响的同一受试者组织的样品。可选地,参考水平/预定水平可以是来自参考数据库的水平,其可以用于生成预定的截止值(cutoffvalue),即统计学上预测具有一种症状或疾病或没有该症状或疾病的诊断评分,或者可以是基于标准群体样品的预定参考水平。
除非本文另外定义,否则本文使用的所有技术和科学术语具有与本发明所属领域内的普通技术人员通常理解的相同的含义。例如,singleton和sainsbury,dictionaryofmicrobiologyandmolecularbiology,第2版,johnwiley和sons,ny(194);以及hale和marham,theharpercollinsdictionaryofbiology,harperperennial,ny(1991)为本领域技术人员提供了本发明中使用的许多术语的通用词典。尽管类似于或等同于本文描述那些方法和材料的任何方法和材料可以在实践本发明时使用,但是本文描述了优选的方法和材料。因此,下文即将定义的术语通过参考说明书整体而被更充分地描述。此外,除非上下文另外清楚地指示,如本文使用的单数术语―一(a)”、―一(an)”和―该(the)”包括复数指示物。除非另外指示,分别地,核酸以5'至3'方向从左到右书写;氨基酸序列以氨基至羧基方向从左至右书写。应理解本发明不限于所描述的特定方法学、方案和试剂,因为这些可根据本领域技术人员使用它们的背景而变化。
本发明的各方面通过以下非限制性实施例来说明。
实施例
研究使用4t1-t与其他41t克隆进行比较,以鉴定转移驱动因子,其在肿瘤细胞进入血流后发挥作用。差异表达和集中的体外和体内rnai筛选的组合揭示了区分这些克隆的候选转移驱动因子,然后在来自乳腺癌患者的基因表达数据集中对这些候选转移驱动因子进行了评估。在这些之中,患者的原发性肿瘤中的天冬酰胺合成酶(asns)的表达与晚期转移性复发最强烈相关。asns沉默降低了体内转移潜能和体外侵袭潜能。相反,增加胞外天冬酰胺的可用性增加了小鼠和人类乳腺癌细胞的侵袭潜能,并且增强asns表达促进的转移。通过用l-天冬酰胺酶治疗或甚至通过膳食限制来降低小鼠的天冬酰胺可用性,强烈减少了来自原位肿瘤的转移。天冬酰胺可用性因组织而异,这可能解释了对肿瘤进展特定步骤的选择性影响。天冬酰胺限制减少了促进上皮向间充质转变的蛋白的产生,这为单一氨基酸的可用性如何能够调节转移进程提供了一种可能的机制。
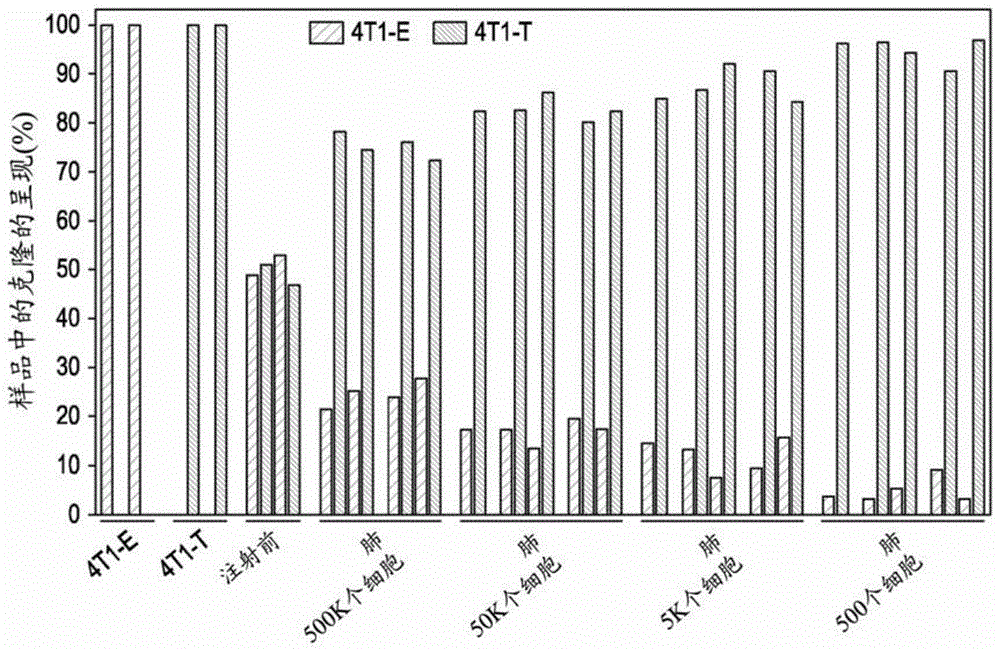
为了验证在含有ctc(ctcproficient)的克隆中4t1-t具有更大转移潜能的观察结果,本发明人将等数目的4t1-e细胞和4t1–t细胞组合,并通过尾静脉注射将它们直接引入免疫受损的接受者(nod-scid-il2rg-/-(nsg)小鼠)的血流中。对初始池的测序表明,两个克隆以相等的丰度存在(图1a)。然而,当从肺中收获细胞时,克隆t占优势,其相对呈现(relativerepresentation)与注射的细胞总数呈负相关。
差异基因表达分析鉴定出与4t1-e相比在4t1-t中具有更高表达的192个基因(数据未示出,倍数改变>2,fdr<0.05)。富集了它们对应的基因本体项目(geneontologyterm)以获得对转移扩散重要的过程(数据未示出,例如对上皮细胞迁移的正调节和对运动的调节)(ashburner等人,2000年)。对乳腺癌患者数据的回顾性分析显示,该组中的基因在侵袭性肿瘤亚型中表达更高(图1b,基础(basal)和密封蛋白(claudin)-低,anovap值<0.0001,harrell等人,2012)。此外,与无复发存活者相比,它们在后期复发至骨、脑和肺的患者的原发性肿瘤中被更高表达(图1c,对于肺,秩和p值<0.01)。
为了确定这组差异表达的基因是否含有转移驱动因子,本发明人用两个平行的组进行了rnai筛选(图1d)。将总计靶向192个基因的~50种shrna的26个池(累计每个基因占约~6种shrna)引入4t1-t细胞(knott等人2014)。将每个池放置在两个分开的6孔基质胶侵袭室上,或者通过尾静脉注射引入到5只nsg重复小鼠体内。在24小时后,收集已经侵袭通过基质胶的细胞,并且7天后,收获小鼠的肺并对肺进行灌注以排除来自脉管系统的残留ctc。使用高通量测序,本发明人鉴定了在侵袭的细胞群体或肺转移灶中被消耗的shrna(经验贝叶斯调整的t-检验fdr<0.05),可能是因为它们靶向了对于这些过程重要的基因。当比较体外和体内候选物时,观察到强烈重叠(图1e,超几何检验p值<0.0001)。如果在筛选的每一组中至少两个对应的shrna被消耗,则基因被分类为候选物(数据未示出)。
在体外和体内测定中被评分的候选基因中,天冬酰胺合成酶(asns)都具有最强有力的临床证据,支持其与疾病进展的相关性(图5)。在两个乳腺癌患者数据集中,人类直向同源物asns的表达水平预示全身复发(generalrelapse)和肺特异性复发(coxp值<0.001)。此外,当对一小部分匹配的肿瘤和肺转移灶进行转录分析时,发现asns在继发性病灶中被更高表达。asns在侵袭性肿瘤亚型中被更高表达(基础、密封蛋白-低和her2+,图6a,anovap值<0.0001),并且与无复发存活者相比,asns在复发至淋巴结、脑、肝和肺的患者中被更高表达(图6b,秩和p值<0.005)。随后对另外三个乳腺癌患者数据集的分析鉴定出asns预测存活率(扩展的数据,图2c,coxp值<0.001)。除了乳腺之外,asns与tcga泛癌数据集中呈现的10种其他实体肿瘤中的4种肿瘤中的存活率呈负相关(图6d,coxp值<0.05)。最后,asns也是实体肿瘤的整体预测性生物标志物(图6e,coxp值<0.001)。
为了验证asns是转移驱动因子,本发明人用两种靶向asns的shrna或一种靶向renilla萤光素酶的对照shrna感染4t1-t细胞,并且将这些细胞静脉内注射到nsg小鼠中(数据未示出)。当在注射后9天评估肺时,接受asns-沉默的细胞的那些动物显示显著降低的转移负荷(图2a&图2b,秩和检验p值<0.001)。asns-沉默的细胞也显示出弱的对基质胶的侵袭(图2c&图7a)。使asns沉默确实影响体外生长;然而,这种缺陷与在侵袭测定中观察到的缺陷相比较小(扩展的数据,图7b&图7c)。
当将asns-沉默的细胞原位注射到乳腺脂肪垫中时,没有观察到原发性肿瘤形成的显著改变(图2d&图7d)。然而,对于来源于asns-沉默的细胞的肿瘤,对应的ctc和肺转移减少(图2e、图2f&图7e,秩和p值分别<0.05和<0.0002)。由沉默的细胞引发的转移明显较小(图2g)。尽管这种差异在统计学上被认为是不显著的,但它确实暗示了转移部位处的生长被影响。用asns-沉默的亲本4t1细胞获得了类似的结果,表明asns依赖性不是该单克隆系的特质(图8a&图8b,秩和p值<0.002)。在亲本4t1群体中增强asns表达不影响原发性肿瘤生长,但是对应的转移的数目显著增加(图8c-图8e,秩和p值0.02)。在人类乳腺癌细胞系mda-mb-231中增强asns表达后观察到类似的结果(图8f-8i,对于asns表达增强的转移与正常转移,秩和p值<0.02)。
在使4t1-t细胞中的asns沉默后,检测到胞内游离天冬酰胺的显著减少(图3a&图9a,经验贝叶斯调整的t-检验fdr<0.05)。此外,当为培养基补充天冬酰胺时,asns-沉默的细胞的侵袭和生长能力增加(图9b&图9c)。因此,本发明人想知道促进侵袭的能力是否是dmem培养基中缺乏的非必需氨基酸(丙氨酸、天冬氨酸、谷氨酸和脯氨酸)特有的。本发明人分开地用这些氨基酸中的每一种来补充培养基并且使用甘氨酸作为阴性对照,并且测定在每种条件生长的细胞的侵袭性。hplc确认了,除了天冬氨酸和谷氨酸之外,这些氨基酸中的每一种的摄入水平类似(图9d)。4t1细胞对天冬酰胺补充有独特的响应,侵袭性增加约2倍(图3b,秩和p值<0.01)。用mda-mb-231细胞观察到类似的结果(图9e&图9f)。任一细胞系的生长都不受天冬酰胺补充的影响(图9g&图9h,秩和p值<0.01)。
这些体外测定指示,侵袭受天冬酰胺可用性的影响,天冬酰胺可用性受生物合成能力和局部环境两者的影响。因此,本发明人想知道转移是否可被胞外天冬酰胺可用性的降低影响。使用细菌天冬酰胺水解酶,l-天冬酰胺酶来减少胞外天冬酰胺,限制癌细胞对天冬酰胺的获取是被成功采用的治疗患有急性淋巴细胞白血病(all)患者的策略(richards等人1998,richards等人2006)。l-天冬酰胺酶已被证明对治疗实体瘤无效(tallal等人1970),这与指示asns表达不是原发位点处生长的必要条件的结果一致。为了确定疾病进展是否受全身天冬酰胺水平影响,本发明人将亲本4t1细胞原位注射到nsg小鼠中,并且用60u的l-天冬酰胺酶治疗一半的动物,每周5次,持续19天,将血清天冬酰胺降低到hplc检测不到的水平(数据未示出)。虽然未检测到原发性肿瘤体积的显著差异,但在l-天冬酰胺治疗的小鼠中观察到转移负荷的显著降低(图10a-图10c,秩和p值<0.0002)。
所有患者通常被归类为telaml1融合基因阳性或阴性。当用含有l-天冬酰胺酶的化学治疗混合物治疗时,那些阳性患者具有高的应答率。2000,stams等人2005)。然而,telaml1阴性患者更易于产生耐药性,并且正是在这些患者中,在治疗后观察到高asns水平,指示耐药性可能是通过生物合成产生实现的。在本模型中,当将asns-沉默的4t1细胞原位注射到l-天冬酰胺治疗的小鼠中时,所导致的转移几乎检测不到(图3c&图3d,秩和p值<0.0005)。在这种情况下,还观察到原发性肿瘤体积的降低(扩展的数据,图6d,秩和p值<0.05)。用asns-沉默的mda-mb-231细胞获得了类似的结果(图10e&图10f,秩和p值<0.05)。
胞外天冬酰胺的可用性不仅可以通过用l-天冬酰胺酶治疗来控制,还可以通过消耗膳食中的天冬酰胺来控制。将shrna感染的4t1-t细胞原位注射到接受对照、低天冬酰胺或高天冬酰胺膳食(分别为0.6%、0%和4%)的小鼠中。hplc确认了,血清天冬酰胺水平根据膳食摄入量而显著改变(图11a)。不管asns表达状态如何,天冬酰胺限制不会影响原发性肿瘤生长(图11b)。相比之下,不管asns表达状态如何,转移负荷在喂食低天冬酰胺膳食的动物中降低,而在给予高天冬酰胺膳食的动物中增加(图3e&图11c,秩和p值<0.0005)。在注射asns-沉默的细胞并喂食低天冬酰胺膳食的小鼠中,几乎检测不到转移。当将亲本4t1细胞原位注射到喂食这些相同天冬酰胺控制膳食的动物中时,获得了类似的结果(图11d-图11f,秩和p值<0.05)。
通过质谱对乳腺、血清和肺的代谢组学分析揭示,在正常生理条件下,天冬酰胺水平在乳腺中最高,但在血清中最低(图3f,秩和p值<0.0005)。在l-天冬酰胺酶治疗的动物血清中几乎检测不到天冬酰胺。qpcr分析确定天冬酰胺丰度与那些组织中的asns表达水平相关(图11g,秩和p值<0.05)。乳腺中的高天冬酰胺可用性可能会缓冲asns-沉默的影响或整体天冬酰胺水平的改变,从而维持肿瘤生长速率。然而,血清中的低水平使细胞对这些改变敏感。最后,肺中的中等水平可以解释由asns沉默产生的较小转移。asns表达水平跨人类组织遵循相似的模式,然而此处肺中的表达略高于乳腺中的表达(图11h)。
为了理解天冬酰胺可用性可能影响侵袭和转移的潜在机制,本发明人在rna和蛋白水平上都检查了由asns沉默诱导的表达改变。rna测量是蛋白水平改变的最强预测因子(图12a,秩和p值<0.001)。根据先前关于l-天冬酰胺酶处理的细胞中天冬酰胺残基翻译暂停的报道,本发明人还发现天冬酰胺含量预测对应的蛋白水平改变(图12a,秩和p值<0.001,loayza-puch等人2016)。对于未归一化的差异和不同于由对应的rna测量预测的改变都是如此(图12b&图4a,秩和p值<0.001)。
在asns沉默后表达减少的富含天冬酰胺的蛋白中,本发明人发现多个基因过度表达,所述基因的人类直向同源物先前被鉴定为在诱导上皮向间充质转变(emt)时被上调(taube等人2010)(图4a、图4b和图12c,emt-上调蛋白,超几何p值<0.005)。被消耗的蛋白具有比所分析的蛋白池高18%的天冬酰胺含量,而emt-上调蛋白的含量高20%。当对亲本4t1细胞的培养基补充天冬酰胺时,这些相同的蛋白的表达增加(图4c,符号秩p值<0.05)。人类直向同源物也富含天冬酰胺(图12d,秩和p值<0.001),第二最多富集的氨基酸是天冬氨酸,天冬氨酸是胞内天冬酰胺生物合成的底物。此外,天冬酰胺富集是emt-上调蛋白的整体保守的特性(图12e,符号秩p-值<1.0×10-13),其在哺乳动物中水平最高(秩和p-值<9.0×10-9)。
在asns-沉默的细胞中,emt-上调基因也在转录水平被下调(图13a,符号秩p值<0.001)。此外,两种典型emt标志物(twist1和上皮钙黏蛋白(e-cadherin))的mrna水平被改变,指示emt程序受到干扰(deseq,fdr<0.05)。当使亲本4t1细胞在补充了天冬酰胺的培养基中生长时,emt-上调基因在其mrna水平上也增加(图13b,符号秩p值<0.005)。对现有数据的重新分析还显示,当调节asns转录的atf4在来自l-天冬酰胺酶治疗的atf4敲除小鼠的单倍体细胞和肝细胞中缺失时,emt-上调基因的表达降低对其emt程序的干扰比相似治疗的wt小鼠更多。综合考虑,这些数据表明天冬酰胺生物可利用度可能至少部分地通过调节emt而影响转移。
虽然emt强烈地牵涉转移,但其重要程度尚不清楚。当比较匹配的亲本4t1肿瘤和肺转移的基因表达谱时,发现emt-上调基因在继发性病变中显著增加(图4d,符号秩p值<0.001)。相比之下,当emt被诱导时被下调的基因(emt下调基因)没有改变。为了在功能上验证emt在该模型中的作用,本发明人原位注射了4t1-t细胞,在该细胞中本发明人已使tgf-β(emt的一种关键驱动因子)的表达沉默。原位部位的生长不受tgf-β沉默影响(图13c)。然而,肿瘤切片的ihc揭示,twist1和上皮钙黏蛋白(两种典型emt标志物)在tgf-β-沉默的肿瘤中发生改变(图13d&图13e,秩和p值<0.01)。与emt的重要作用一致,在携带具有emt受损的肿瘤的小鼠中观察到较少的转移(扩展的数据,9f,秩和p值<0.05)。当静脉内注射tgf-β-沉默的细胞时,这些细胞也产生较少的转移,指示emt在肿瘤细胞进入血流后的转移中起作用(图13g,秩和p值<0.05)。
虽然在来源于asns-沉默的细胞的肿瘤中没有检测到形态学差异,但是对twist和上皮钙黏蛋白的ihc染色表明在这些病变中emt受到干扰,并且在对应的转移灶中也观察到这种相同的模式(图4e-4g&图14a)。当对来自这些病灶的恶性细胞进行facs分离并通过qpcr分析时,发现twist1和上皮钙黏蛋白表达水平以与染色一致的方式改变(图14b&图14c,秩和p值<0.05)。在用l-天冬酰胺酶治疗的小鼠的原发性肿瘤中观察到类似的模式(图14d&图14e)。在那里,沉默具有最大的影响,但是使用l-天冬酰胺酶消耗胞外天冬酰胺确实引起了显著的表达改变(秩和p值<0.01)。膳食天冬酰胺也影响表达,天冬酰胺含量与原发性肿瘤中的emt信号呈正相关(图14f和图14g,秩和p值<0.01)。
为了更密切地检查这些病灶内的恶性细胞,本发明人通过6-tg选择从肿瘤和肺转移灶中分离了shrna感染的4t1-t细胞。不管asns-表达状态如何,转移的细胞显示出间充质形态(被拉长且为纺锤形)(图14h)。然而,从原发性肿瘤中分离出的asns-沉默的细胞显示出更多的上皮形态。在这些细胞中,emt-下调基因被上调,并且twist和上皮钙黏蛋白表达水平也以与emt程序被干扰一致的方式被改变(图4h,符号秩p-值<0.05,且deseqfdr<0.05)。在asns-沉默的转移性细胞中,emt-上调基因被下调,并且twist和上皮钙黏蛋白的表达也被改变(秩和p值<0.001,且deseqfdr<0.05)。
我们的乳腺肿瘤异质性模型强烈暗示天冬酰胺生物可利用度是转移性进展的调节因子。这也可能与人类癌症相关,因为高asns表达是许多肿瘤类型预后不良的标记。我们发现的一个潜在机制可能是天冬酰胺生物可利用度和emt之间的联系,可以在体外和体内被观察到该联系。在我们的乳腺癌模型中,门控步骤可能发生在血液循环中,其中天冬酰胺水平是低的并且受l-天冬酰胺酶治疗或膳食限制的强烈影响。尽管如此,我们确实观察到了对原发部位和继发部位的上皮样与间充质样肿瘤细胞的比率的影响,这也可能影响肿瘤进展和对治疗的响应二者。
实验方法
细胞培养
将小鼠乳腺肿瘤细胞系4t1(atcc)和任何衍生的克隆细胞系培养在补充有5%胎牛血清、5%小牛血清、mem非必需氨基酸(neaa)和青霉素链霉素(thermofisherscientific)的dmem高葡萄糖培养基中。将人类乳腺肿瘤细胞系mdamb-231(atcc)培养在补充有10%胎牛血清、neaa和青霉素链霉素(thermofisherscientific的dmem高葡萄糖培养基中。atcc测试和验证了4t1细胞系和mda-mb-231细胞系。将用于产生病毒的铂-a(cellbiolabs)和293ft(thermofisherscientific)包装细胞系培养在补充有10%胎牛血清和青霉素链霉素的dmem高葡萄糖中。所有细胞系都已常规测试了支原体污染。
病毒产生
如wagenblast等人2015先前描述的,将逆转录病毒载体使用铂-a(cellbiolabs)细胞系包装,并且将慢病毒载体用293ft细胞系(thermofisherscientific)包装。
动物研究
所有的小鼠实验都得到冷泉港动物保护和使用委员会(thecoldspringharboranimalcareandusecommittee)批准。从未在任何方向超过被允许的最大肿瘤尺寸20mm。所有的小鼠注射都是用6-8周龄的雌性nod-scid-il2rg-/-(nsg)小鼠(jax)进行的。balb/c小鼠未用于本研究,因为由于慢病毒条形码载体造成不同克隆细胞系具有不同的gfp水平。使用5×105个小鼠乳腺肿瘤细胞进行尾静脉注射,所述细胞被重悬在100ul的pbs中,并通过尾静脉注射。使用1×105个小鼠乳腺肿瘤细胞或5×105个mda-mb-231细胞进行原位注射。对此,将细胞重悬在1:1的pbs与生长因子减少的基质胶(bdbiosciences)的混合物中。对于小鼠乳腺肿瘤细胞,将20ul体积注射到乳腺#4中,并且对于mda-mb-231细胞,注射40ul体积。使用式v=1/2(lxw2)测量原发性肿瘤体积,其中l是原发性肿瘤的长度,且w是原发性肿瘤的宽度。对于l-天冬酰胺酶研究,将小鼠通过腹膜内注射每周施用200ul的60u的l-天冬酰胺酶5x。对于调整了l-天冬酰胺的膳食,对小鼠给予对照氨基酸膳食(0.6%天冬酰胺)、不含天冬酰胺的膳食(0%天冬酰胺)或富含天冬酰胺的膳食(4%天冬酰胺)。所有的膳食都是等氮的(isonitrogenous),并且含有类似的卡路里密度。样品尺寸被选择为提供足以用标准统计学检验判定显著性的能力。以每种条件10只动物进行小鼠实验,以考虑在该体内实验中观察到的可变性。通过随机笼选择将动物分配到治疗组。
条形码分析
如先前在wagenblast等人2015中描述的,对4t1-e细胞和4t1-t细胞的条形码进行扩增和测序。
体内shrna肺筛选和体外侵袭筛选
shrna是基于knott等人2014中描述的sherwood算法预测的。~50种shrna的池被包装在铂-a细胞中。对于每个池,将1000万个4t1-t细胞以0.3的感染复数(moi)进行感染。用500ug/ml潮霉素对感染的细胞进行持续5天的选择,并且将每个池通过尾静脉分别注射到5只小鼠中。在注射时收集注射前的池,以验证每种shrna的相等呈现。在7天后,处死小鼠并用pbs灌注以去除肺中的血液和非外渗细胞(non-extravasatedcell)。收获肺并使用苯酚氯仿提取分离基因组dna。注射前的池的基因组dna使用qiaampdna血液迷你试剂盒(qiagen)进行分离。
平行地进行体外侵袭测定。将每个池铺板在两个6孔biocoat基质胶侵袭板(corning)上。将在没有血清的细胞培养基中的6×105个细胞铺板在每个孔的上部。允许细胞侵袭通过基质胶,进入含有5%胎牛血清和5%小牛血清的培养基中。将侵袭的细胞刮掉,用pbs洗涤,并且使用qiaampdna血液迷你试剂盒分离基因组dna。
使用knott等人2014中先前描述的关于下一代测序的两步pcr方案对shrna进行扩增。
第一pcr正向引物1:5-cagaatcgttgcctgcacatcttggaaac-3[seqidno:1]和反向引物1:5-ctgctaaagcgcatgctccagactgc-3[seqidno:2]。
第二pcr正向引物2:5-aatgatacggcgaccaccgagatctacactagcctgcgcacgtagtgaagccacagatgta-3[seqidno:3]和反向引物2:5-caagcagaagacggcatacgagatnnnnnngtgactggagttcagacgtgtgctcttccgatctctgctaaagcgcatgctccagactgc-3[seqidno:4]。反向引物含有能够实现多重化的条形码(nnnnnn)。
对筛选数据的分析
如knott等人2014中描述的对每个筛选池进行分析。
基因本体富集分析
基因本体富集分析使用gorilla门户网站进行。与4t1-e细胞相比,在4t1-t细胞中被鉴定为过表达的基因的参考序列(refseq)标识符被用作前景(foreground),并且整个参考序列基因列表被用作背景。
表达亚型和复发分析
所有的临床数据分析都使用universityofnorthcarolina的“855患者集”进行,该数据集作为已公布的数据可在https://genome.unc.edu获得。所有的数据都来源于以横轴为患者并且纵轴为基因布置初始矩阵。初始归一化包括分位数标准化,以确保每个患者的整体表达谱是相似的。此后,对所有患者的每个基因的z评分归一化。对于图1b,对于每个患者亚型,计算每个基因的平均表达水平。对于图1c,对于具有和没有向各继发部位复发的患者计算每个基因的平均表达水平。对于图6a,绘制了按亚型分级的每个患者的asns水平。对于图6b,每个盒形图代表在具有和没有向各继发部位复发的每个患者的asns水平。
个体体外侵袭测定
细胞的体外侵袭能力使用6孔biocoat基质胶侵袭板测量。对于亲本4t1细胞,将1×106个细胞铺板在个体孔上,对于4t1-t细胞,将8×105个细胞铺板在个体孔上,并且对于mda-mb-231细胞,每个个体孔使用5×105个细胞。将细胞重悬在没有血清的培养基中,并且细胞侵袭到具有5%胎牛血清和5%小牛血清的培养基中。对于图3b和图9e,将4t1细胞和mda-mb-231细胞在含有100x浓度的特定氨基酸(相对于1xneaa浓度)的培养基中分别培养2日或3日,然后开始侵袭测定。在24h后,去除未侵袭的细胞,并在pbs中洗涤侵袭孔,在2%戊二醛中固定2min,并且用0.5%结晶紫染色10min。将这些孔在蒸馏h2o中洗涤,风干并使用odyssey红外扫描仪扫描。信号使用imagej(nih)定量。
竞争和增殖测定
对于mcherry竞争测定,将shrna转导的mcherry阳性细胞与未转导的细胞混合。在lsrii流式细胞仪(bdbiosciences)上对mcherry萦光定量。增殖测定使用celltraceviolet细胞增殖试剂盒(thermofisherscientific)进行。对于图3b和图9e,将4t1细胞和mda-mb-231细胞在含有100x浓度的特定氨基酸(相对于1xneaa浓度)的培养基中分别培养2日或3日,然后开始增殖测定。将细胞用celltraceviolet染色,并且然后胰酶消化并重悬在培养基中。在24小时后,收集细胞,以便使用sh800流式细胞仪(sony)对紫色萦光强度定量。
分离肿瘤和肺转移性细胞
如wagenblast等人2015中先前报道的收获肿瘤和肺组织、切碎并消化成单细胞。使细胞在含有60μm6-硫鸟嘌呤的4t1细胞培养基中生长以消耗基质细胞,或者使用facsariaiii细胞分选仪(bdbiosciences)基于mcherry表达直接对细胞进行分选。
rnaseq文库制备
如wagenblast等人2015中先前描述的从培养的4t1-t细胞一式两份地制备rnaseq文库。在illuminahiseq测序仪上对每个样品进行测序,产生76nt单末端(se)读段。
rnaseq分析
使用bowtie2用默认参数将illumina测序读段与小鼠基因组(mm10)进行比对(langmead等人2012)。使用htseqcount对基因分配计数(anders等人2015)。差异表达分析使用deseq进行(anders等人2010)。
shrna敲低和cdna过表达研究
分别用表达shrna的逆转录病毒或慢病毒构建体转导小鼠细胞系和人类细胞系。在感染后,用500ug/ml潮霉素对4t1-t细胞进行5天的选择,并且用2ug/ml嘌呤霉素对mda-mb-231进行4天的选择。用g418对被过表达cdna的逆转录病毒构建体感染的细胞系进行一周的选择。用600ug/mlg418对亲本4t1细胞系进行选择,并且用1500ug/mlg418对mda-mb-231细胞进行选择。
cdna过表达的基因:
小鼠asns:nm_012055.1
人类asns:nm_001673.2
shrna敲低序列:
小鼠shasns-1[seqidno:5]:
小鼠shasns-2[seqidno:6]:
小鼠shtgfb1-1[seqidno:7]:
小鼠shtgfb1-2[seqidno:8]:
人类shasns-1[seqidno:9]:
人类shasns-2[seqidno:10]:
qrt-pcr
使用rneasy迷你试剂盒(qiagen)从细胞纯化总rna并且进行dna酶处理。对于全部组织,使用trizol加rna纯化试剂盒(thermoscientific)分离rna。将组织裂解物使用dounce匀浆器匀质化,并通过柱匀浆器(thermofisherscientific)以降低黏度。在agilentbioanalyzer(rnanano试剂盒)上测量rna完整性(rna完整性评分>9)。使用superscriptiii逆转录酶(sigma)合成cdna。在eppendorfmastercyclereprealplex上进行定量pcr分析。所有信号都使用δct方法进行定量,并且针对gapdh水平进行归一化。对于流式细胞仪分选的mcherry阳性肿瘤和肺转移性细胞,使用taqmangnenexpressioncells-to-ct试剂盒(thermofisherscientific)直接从裂解的细胞产生cdna。使用taqman引物/探针组在cfx96(bio-rad)上进行定量pcr分析,并且如以上描述的对所有信号进行定量。
qrt-pcr引物
小鼠asns(外显子1-2)[seqidno:11]:
5′-cctctgctccaccttctct-3′5′-gatcttcatcgcactcagaca-3′
小鼠asns(外显子6-7)[seqidno:12]:
5′-ccaagttcagtatcctctccag-3′5′-cttcatgatgctcgcttcca-3′
小鼠tgfb1(外显子1-2)[seqidno:13]:
5′-ccgaatgtctgacgtattgaaga-3′5′-gcggactactatgctaaagagg-3′
小鼠tgfb1(外显子3-4)[seqidno:14]:
5′-gttatctttgctgtcacaagagc-3′5′-cccactgatacgcctgag-3′
小鼠gapdh(外显子2-3)[seqidno:15]:
5′-aatggtgaaggtcggtgtg-3′5′-gtggagtcatactggaacatgtag-3′
人类asns(外显子8-9)[seqidno:16]:
5′-gagtcagacctttgtttaaagca-3′5′-ggagtgcttcaatgtaacaagac-3′
人类asns(外显子12-13)[seqidno:17]:
5′-ctggatgaagtcatattttccttgg-3′5′-cagagaagatcaccacgctatc-3′
人类gapdh(外显子2-3)[seqidno:18]:
5′-acatcgctcagacaccatg-3′5′-tgtagttgaggtcaatgaaggg-3′
taqman探针:
小鼠asns:mm00803785_ml
小鼠上皮钙黏蛋白(cdh1):mm01247357_ml
小鼠twist1:mm00442036_ml
小鼠gapdh:mm99999915_gl
循环肿瘤细胞(ctc)的qpcr:
如wagenblast等人2015中先前描述的对ctc进行定量。从血液中分离基因组dna,并使用针对mcherry的qpcr测定进行定量,mcherry是由逆转录病毒shrna递送载体表达的。
mcherry探针和引物:
引物1:5′-gactacttgaagctgtccttcc-3′[seqidno:19]
引物2:5′-cgcagcttcaccttgtagat-3′[seqidno:20]
hex探针:5′-/56-fam/ttcaagtgg/zen/gagcgcgtgatgaa/3iabkfq//-3′[seqidno:21]
持家基因探针和引物:
引物1:5′-gacttgtaacgggcaggcagattgtg-3′[seqidno:22]
引物2:5′-gaggtgtgggtcacctcgacatc-3′[seqidno:23]
hex探针:
5′-/5hex/ccgtgtcgc/zen/tctgaagggcaatat/3iabkfq/-3′[seqidno:24]
对肺转移的定量
对于每个肺,制备5微米切片,并且用标准的h&e方案染色。通过对一个切片上的单独肺结节计数来确定肺转移负荷。
上皮钙黏蛋白和twist1分析:
对于免疫组织化学,如wagenblast等人2015中先前描述的对原发性肿瘤和肺进行处理。将上皮钙黏蛋白(24e10)兔mab(3195,cellsignaling)以1∶400稀释度使用,并且将twist1(twist2c1a)小鼠mab(ab50887,abcam)以1∶100稀释度使用。使用imagej(nih)对上皮钙黏蛋白和twist1的二氨基联苯胺(dab)染色以及苏木精染色进行定量。对此,根据ruifrok等人2001的研究,对图像进行颜色解卷积,并且测量上皮钙黏蛋白和twist1阳性染色的面积百分比。
使用hplc对游离氨基酸定量
对培养的细胞和血清中的游离氨基酸进行定量。对于培养的细胞,将4t1细胞和mda-mb-231细胞在含有100x浓度的特定氨基酸(相对于1xneaa浓度)的培养基中分别培养2日或3日。使用dounce匀浆器对所有培养的细胞进行均质,并且随后过滤裂解物。使用高效液相色谱(hplc)和萦光检测器一式三份地对每个样品进行定量。对于每个重复品,测量每种氨基酸的纳摩尔数。使用每三份的平均值计算每种氨基酸的摩尔百分比组成。
使用等量异位素标签(isobarictags)相对和绝对定量(itraq)的蛋白组学分析
如ross等人2004中先前描述的,将细胞在冰冷的pbs中洗涤,并且收获用于itraq定量蛋白组学。对于每个细胞系使用三个重复品。
使用液相色谱串联质谱(lc-ms/ms)的代谢物分析
如先前描述的(26)进行代谢物提取。将器官组织样品放置在2ml裂解管中,所述裂解管预填充有1.4mm陶瓷珠(针对乳腺)或2.8mm陶瓷珠(针对肺)和1ml预冷却的80%甲醇。用precellys24匀浆器(bertininstruments)对样品进行均质,程序设置为6500hz,30s,3个循环和4min暂停时间。在每个循环结束时,将样品在液氮中速冻并放置在干冰上。使用在-80℃的200ul的80%甲醇对血清样品(50ul)进行代谢物提取。离心10min(13.2krpm,4℃)后,将上清液蒸发至干燥,并在-80℃储存,直到lc-ms/ms分析。
将干燥(dried-down)的提取物重悬在25ulhplc级别的水中,并使用与串联质谱分析(lc-ms/ms)偶联的亲水作用色谱(hilic)对1ul进行分析。分析仪器由与配备有电喷雾离子源的qtrap6500混合三重四极杆/线性离子阱质谱仪(sciex)偶联的nexerax2(shimadzu)液相色谱组成。使用analyst1.6.2(sciex)获取原始lc-srm-ms数据,并使用multiquant1.1软件(sciex)对每种代谢物的lc-srm-ms迹线的峰面积进行积分。代谢物差异通过将样品针对总峰面积进行归一化并使用单向anova与多重比较对每个组的重复品进行比较来分析。
氨基酸组成分析
对于图4a,转录水平和蛋白水平的log-倍数改变被分位数归一化为相同的分布。然后将蛋白水平的改变减去rna水平的改变。然后使用秩和检验对后续值最高的10%和最低的10%的基因的氨基酸呈现进行比较,以鉴定其丰度无法由转录改变解释的而与蛋白水平改变相关的氨基酸。对于图12b,使用相同的秩和检验对显示出rna和蛋白表达最大增加/减少的基因之间的氨基酸呈现进行比较。对于图12c&图12d,进行了相同的分析,这次比较了在emt期间相比所有其他基因被检测为上调的基因的蛋白。在图12c的情况下,emt-上调基因是emt-上调的人类基因的小鼠直向同源物。
对于图12e,分析了携带图12d中描述的促emt(pro-emt)人类基因的最少10个直向同源物的基因的每种生物体。对于每种生物体,计算了每种蛋白的天冬酰胺百分比。然后,通过计算促emt蛋白与剩余的生物体特异性蛋白的中值天冬酰胺百分比的比率,计算每种生物体的天冬酰胺富集水平。如关于图12c和图12d描述的计算富集的统计学显著性。
核糖体谱(profiling)分析
为便于分析,使用cutadapt19进行质量微调和接头去除。使用bowtie2去除映射到污染性rna(例如rrna和trna序列)14的读段。随后,使用star将长度29-33的读段映射到人类转录组20。基于读段长度对每次读段的偏移进行校正,并且将下游的12个和15个核苷酸标记为p位点和a位点。然后我们对于每个基因计算了所有位置处的事件数,并且汇总了每个密码子的计数并随后汇总了氨基酸的计数。
参考文献
greaves,m.&maley,c.c.clonalevolutionincancer.nature481,306-313(2012).
kang,y.etal.amultigenicprogrammediatingbreastcancermetastasistobone.cancercell3,537-549(2003).
lawson,d.a.etal.single-cellanalysisrevealsastem-cellprograminhumanmetastaticbreastcancercells.nature526,131-135(2015).
lohr,j.g.etal.widespreadgeneticheterogeneityinmultiplemyeloma:implicationsfortargetedtherapy.cancercell25,91-101(2014).
meacham,c.e.&morrison,s.j.tumourheterogeneityandcancercellplasticity.nature501(2013).
mullighan,c.g.etal.genomicanalysisoftheclonaloriginsofrelapsedacutelymphoblasticleukemia.science(newyork,n.y.)322,1377-1380(2008).
navin,n.etal.tumourevolutioninferredbysingle-cellsequencing.nature472(2011).
nguyen,a.,yoshida,m.,goodarzi,h.&tavazoie,s.f.highlyvariablecancersubpopulationsthatexhibitenhancedtranscriptomevariabilityandmetastaticfitness.naturecommunications7,11246(2016).
shackleton,m.,quintana,e.,fearon,e.r.&morrison,s.j.heterogeneityincancer:cancerstemcellsversusclonalevolution.cell138,822-829(2009).
tirosh,l.etal.dissectingthemulticellularecosystemofmetastaticmelanomabysingle-cellrna-seq.science(newyork.n.y.)352,189-196(2016).
wang,y.etal.clonalevolutioninbreastcancerrevealedbysinglenucleusgenomesequencing.nature512,155-160,doi:10.1038/nature13600(2014).
yu,m.etal.rnasequencingofpancreaticcirculatingtumourcellsimplicateswntsignallinginmetastasis.nature487,510-513(2012).
wagenblast,e.etal.amodelofbreastcancerheterogeneityrevealsvascularmimicryasadriverofmetastasis.nature520,358-362(2015).
vanharanta,s.&massague,j.originsofmetastatictraits.cancercell24,410-421(2013).
miller,f.r.,miller,b.e.&heppher,g.h.characterizationofmetastaticheterogeneityamongsubpopulationsofasinglemousemammarytumor:heterogeneityinphenotypicstability.invasion&metastasis3,22-31(1983).
ashburner,m.etal.geneontology:toolfortheunificationofbiology.thegeneontologyconsortium.naturegenetics25,25-29(2000).
harrell,j.c.etal.genomicanalysisidentifiesuniquesignaturespredictiveofbrain,lung,andliverrelapse.breastcancerresearchandtreatment132,523-535(2012).
knott,s.r.etal.acomputationalalgorithmtopredictshrnapotency.molecularcell56,796-807(2014).
richards,n.g.&schuster,s.m.mechanisticissuesinasparaginesynthetasecatalysis.advancesinenzymologyandrelatedareasofmolecularbiology72,145-198(1998).
richsrds,n.g.&kilberg,m.s.asparaginesynthetasechemotherapy.annualreviewofbiochemistry75,629-654(2006).
tallal,l.etal.e.colil-asparaginaseinthetreatmentofleukemiaandsolidtumorsin131children.cancer25,306-320(1970).
ramakers-vanwoerden,n.l.etal.tel/aml1genefusionisrelatedtoinvitrodrugsensitivityforl-asparaginaseinchildhoodacutelymphoblasticleukemia.blood96,1094-1099(2000).
stams,w.a.etal.asparaginesynthetaseexpressionislinkedwithlasparaginaseresistanceintel-aml1-negativebutnottel-aml1-positivepediatricacutelymphoblasticleukemia.blood105,4223-4225(2005).
loayza-puch,f.etal.tumour-specificprolinevulnerabilityuncoveredbydifferentialribosomecodonreading.nature530,490-494(2016).
taube,j.h.etal.coreepithelial-to-mesenchymaltransitioninteractomegene-expressionsignatureisassociatedwithclaudin-lowandmetaplasticbreastcancersubtypes.proceedingsofthenationalacademyofsciencesoftheunitedstatesofamerica107,15449-15454(2010).
langmead,b.&salzberg,s.l.fastgapped-readalignmentwithbowtie2.nat.methods9,357-359(2012).
anders,s.,pyl,p.t.&huber,w.htseq--apythonframeworktoworkwithhigh-throughputsequencingdata.bioinformatics31,166-169(2015).
anders,s.&huber,w.differentialexpressionanalysisforsequencecountdata.genomebiol.11,r106(2010).
ruifrok,a.c.&johnston,d.a.quantificationofhistochemicalstainingbycolordeconvolution.anal.quant,cytol.histol.23,291-299(2001).
martin,m.cutadaptremovesadaptersequencesfromhigh-throughputsequencingreads.embnet.journal17,10(2011).
dobin,a.etal.star:ultrafastuniversalrna-seqaligner.bioinformatics29,15-21(2013).
ross,p.l.etal.multiplexedproteinquantitationinsaccharomycescerevisiaeusingamine-reactiveisobarictaggingreagents.mol.cellproteomics3,1154-1169(2004).
- 还没有人留言评论。精彩留言会获得点赞!