涉及经工程改造的CD16的多肽、细胞和方法与流程
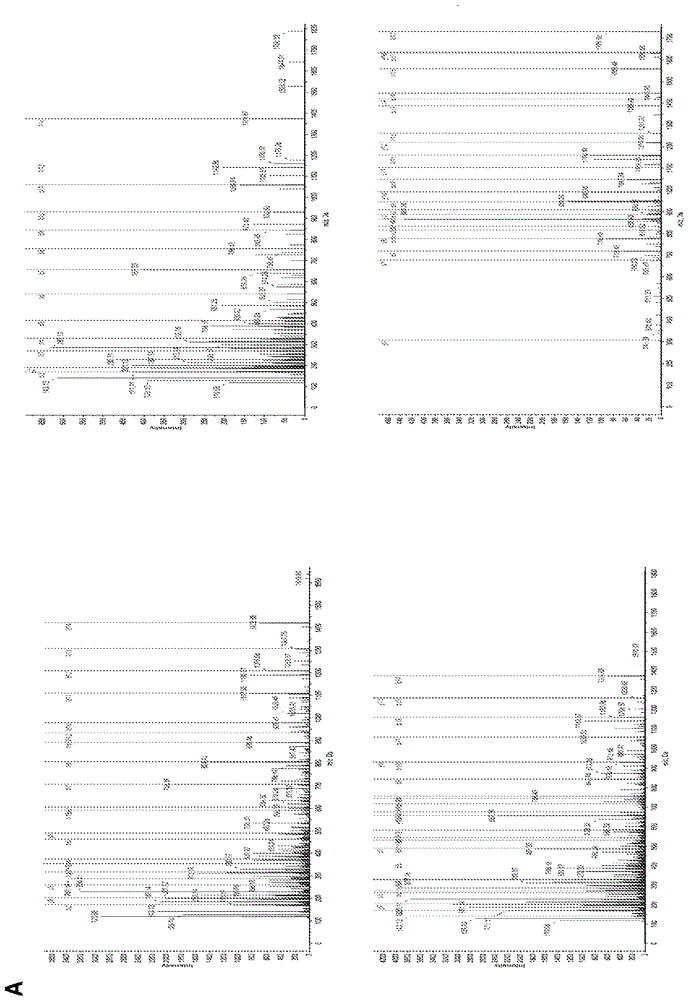
本申请要求于2014年3月28日提交的美国临时专利申请序列号61/971,996的优先权,所述申请通过引用结合到本文中。发明概述本公开内容一般地描述修饰形式的CD16,表达修饰的CD16的遗传修饰的细胞,和涉及遗传修饰的细胞的方法。所述修饰形式的CD16可显示增加的抗肿瘤和/或抗病毒活性,这至少部分上是由于减少的对NK细胞刺激时金属蛋白酶介导的脱落的敏感性。因此,在一个方面,本公开内容描述经遗传修饰以表达具有膜近侧区和膜近侧区中的氨基酸修饰的CD16多肽的细胞。在另一个方面,本公开内容描述包含编码具有膜近侧区和膜近侧区中的氨基酸修饰的CD16多肽的多核苷酸的细胞。在任何一个方面,所述氨基酸药物(medication)反映与CD16膜近侧区野生型氨基酸序列相比一个或多个氨基酸的添加、一个或多个氨基酸的缺失,或一个或多个氨基酸的置换。在这些实施方案的一些中,一个或多个氨基酸的置换包括SEQIDNO:1的197位的丝氨酸残疾的置换。在任何一个方面,细胞可为自然杀伤(NK)细胞、中性粒细胞、单核细胞或T细胞。在任何一个方面,与野生型CD16多肽相比修饰的CD16多肽显示减小的对ADAM17介导的脱落的敏感性。在任何一个方面,与野生型CD1多肽相比修饰的CD16多肽显示减小的对NK细胞刺激时的切割的敏感性。在另一个方面,本公开内容描述一般地涉及给予需要这种处理的患者包括以下的治疗的方法:(a)给予患者治疗性NK效应物,和(b)给予患者上文概述的遗传修饰细胞的任何实施方案。在一些实施方案中,治疗性NK效应物包括治疗剂。在这些实施方案的一些中,治疗剂可包括抗体,或治疗性抗体片段。在这些实施方案的一些中,抗体或抗体片段与病毒抗原特异性结合。在其它实施方案中,抗体或抗体片段与肿瘤抗原特异性结合。在一些实施方案中,治疗剂可包括双特异性杀伤衔接体(BiKE)或三特异性杀伤细胞衔接体(TriKE)。在又一个方面,本公开内容描述用于改善对患者的免疫治疗的方法,其中所述免疫治疗涉及给予患者治疗性NK效应物。一般地所述方法包括进一步给予患者上文概述的遗传修饰细胞的任何实施方案。本发明的以上概述不意在描述本发明的每一公开的实施方案或每一实现。随后的说明书更具体地例示说明性的实施方案。在整个申请的若干位置,指导通过实施例的列表提供,这些实施例可以以不同的组合使用。在每种情况下,叙述的列表仅用作代表性群组,并且不应解释为排他的列表。附图简述图1.人CD16中胞外域切割位点的位置。(A)从PMA-活化人NK细胞或中性粒细胞的细胞上清液免疫沉淀的可溶性CD16的胰蛋白酶肽(trypticpeptides)经受质谱分析。鉴定了四个具有非胰蛋白酶C末端的高置信度肽:1个肽来自NK细胞释放的可溶性CD16(肽#1,左上),3个肽来自中性粒细胞释放的可溶性CD16(肽#2,左下;肽#3,右上;和肽#4,右下)。(B)肽#1-4(加下划线的)以及CD16a(SEQIDNO:1)和CD16b(SEQIDNO:2)中假定切割位点(箭头)的说明。所鉴定的肽中氨基酸176区分CD16a(F)与CD16b(V)。氨基酸1-16指示CD16a和CD16b的预测信号序列。氨基酸210-229指示CD16a的跨膜区域。氨基酸编号从信号序列中的甲硫氨酸开始。CD16a和CD16b的氨基酸序列分别来自NCBI参考序列NM_000569.6和NM_000570.4。图2.CD16胞外域脱落、切割区域和经工程改造的丝氨酸-197至脯氨酸突变的示意图。CD16a和CD16b通过ADAM17在膜近侧区内经受胞外域脱落,如所标明的。膜近侧区内的CD16切割区域基于质谱分析,其显示极为贴近的三个不同的切割位点(箭头)。进行定点突变以将CD16(SEQIDNO:1的氨基酸190-202)中的丝氨酸-197置换为脯氨酸(CD16/S197P)。图3.经工程改造的S197P突变对CD16a和CD16b脱落的作用。转染的HEK293(人胚胎肾)细胞以相似水平分别表达CD16b和CD16b/S197P(A)或CD16a和CD16a/S197P(B),如通过流式细胞术所测定的(左侧面板)。用或不用PMA处理(15ng/ml在37℃持续30分钟)不同转染子并通过ELISA定量培养基上清液中的CD16的可溶性水平(右侧面板)。对于每一实验,每一处理条件重复三次,数据代表三个独立实验。条形图显示平均值±SD。统计显著性表示为***P<0.001。(C)转染的HEK293细胞表达L-选择素(CD62L)或L-选择素和CD16b/S197P。转染和模拟转染细胞上L-选择素和CD16b/S197P的表面水平使用流式细胞术测量(直方图)。在PMA存在或不存在下37℃孵育表达L-选择素或L选择素和CD16b/S197P的转染子30分钟,并测定L-选择素染色的平均荧光强度(MFI)(条形图)。对于每一实验,每一处理条件重复三次,数据代表两个独立实验。条形图显示平均值±SD。统计显著性表示为*P<0.05。对于所有直方图,x-轴=Log10荧光,y-轴=细胞数目。图4.经工程改造的S197P突变对NK细胞中CD16a脱落的作用。不用(Unstim.)或用PMA(100ng/ml)37℃处理用空载体(仅载体)、CD16a或CD16a/S197P转导的NK92细胞30分钟(A),用IL-12和IL-18(分别100ng/ml和400ng/ml)37℃处理24小时(B),或用Raji细胞和利妥昔单抗37℃处理60分钟(C)。CD16a的细胞表面水平通过流式细胞术测定。虚线指示同种型匹配的阴性对照抗体染色。(D)在ADAM17抑制剂BMS566394(5μM)存在或不存在下用Raji细胞和利妥昔单抗37℃处理亲本NK92细胞和表达CD16a或CD16a/S197P的转导细胞60分钟。可溶性CD16a水平通过ELISA测定。每一处理条件重复三次,数据代表三个独立实验。条形图显示平均值±SD。统计显著性表示为***P<0.001。(E)表达CD16a或CD16a/S197P的NK92细胞用抗-ADAM17mAbsM220、623、633或同种型匹配的阴性对照抗体染色,如所标明的。(F)从模拟转导iPSCs(左侧面板)或表达重组CD16a或CD16a/S197P的iPSCs(右侧面板)衍生的CD56+CD45+NK细胞与或不与K562靶细胞在37℃孵育4小时。对于所有直方图,x-轴=Log10荧光,y-轴=细胞数目,并且数据代表至少3个独立实验。图5.经工程改造的S197P突变对CD16a功能的作用。(A)以相等水平表达CD16a或CD16a/S197P的NK92细胞用单体人IgG(0-20μg/ml)处理(左侧面板)。作为对照,细胞也用单体人IgA(20μg/ml)处理并用IgG(20μg/ml)处理NK92亲本细胞(条)。抗体结合通过流式细胞术测定,如材料与方法中所描述的。条形图显示至少三个单独实验的平均值±SD。相对于IgG(0μg/ml)、IgA或NK92亲本细胞+IgG,统计显著性表示为*P<0.05。(B)在Raji细胞不存在(Unstim.)或存在下用或不用抗CD20利妥昔单抗37℃孵育模拟转导的NK92细胞或表达CD16a或CD16a/S197P的NK92细胞达到指定的时间点。NK92细胞活化通过流式细胞术由CD107a染色的上调评估。对于直方图,x-轴=Log10荧光,y-轴=细胞数目。数据代表至少3个独立实验。说明性实施方案的详述本公开内容一般地描述修饰形式的CD16a,表达修饰的CD16a的遗传修饰的细胞,和涉及遗传修饰的细胞的方法。所述修饰形式的CD16a可显示增加的抗肿瘤和/或抗病毒活性,这至少部分上是由于减少的对NK细胞刺激时金属蛋白酶介导的脱落的敏感性。与许多实体肿瘤类型相比,在过去的30年间具有上皮性卵巢癌的妇女的存活率几乎未改变。此外,当前对复发性卵巢癌的标准治疗提供低(<20%)响应率。尽管卵巢癌样品中普遍的HER2过表达,在晚期卵巢癌患者中用抗-HER2抗体曲妥单抗处理仅提供有限的响应。这一对曲妥单抗的抗性可能由功能失调的NK细胞介导的抗体-依赖性细胞毒性引起。因此,存在对创新治疗策略的迫切需求。我们描述了一种用于提供治疗处理策略的新途径。卵巢癌的一个关注为周围环境,其中肿瘤细胞发展可为高度促炎的因此很可能促进浸润NK细胞上CD16a切割从而减少抗体-依赖性细胞毒性。数种抗体已经作为有效的用于治疗人恶性肿瘤的靶向治疗而出现。其功效部分上是由于与自然杀伤(NK)细胞上FcγRIIIa/CD16a的抗体相互作用和通过抗体-依赖性细胞毒性诱导杀死癌细胞。人IgGFc受体CD16(FcγRIII)由两种同种型组成:CD16a(FcγRIIIa)和CD16b(FcγRIIIb)。CD16a由自然杀伤(NK)细胞表达,CD16b由中性粒细胞表达。NK细胞活化通过称为胞外域脱落(涉及金属蛋白酶ADAM17和发生在接近质膜的单一胞外区域的蛋白水解事件)的过程导致两种CD16同种型的表面水平的快速下调(图1A)。如上文所指出的,卵巢癌患者可能抵抗NK细胞介导的免疫治疗——即,肿瘤对NK细胞介导的治疗不敏感。例如,卵巢癌细胞通常表达表皮生长因子受体HER2,但用治疗抗体曲妥单抗靶向其仅提供有限的临床响应。该抗性至少部分上可由胞外域脱落造成——即,细胞因子对NK细胞的活化、靶细胞相互作用和/或肿瘤浸润可导致CD16切割和损伤抗体-依赖性细胞毒性。因此,阻断胞外域脱落过程具有临床意义。我们已经使用质谱法测定CD16a和CD16b的切割位点并从人血白细胞克隆CD16a和CD16b的cDNA。以定向方式突变每一cDNA以诱导单个氨基酸改变。197位的丝氨酸变为脯氨酸(图1B)。该突变阻断CD16a和CD16b的切割,防止细胞活化时其下调。体外扩增的NK细胞中抗切割CD16a的表达维持该IgGFc受体的高表面水平,这增强NK细胞刺激、治疗抗体功效和癌细胞杀伤。ADAM17具有许多细胞表面底物,但不具有可用于预测CD16a切割位点的蛋白质水解共有序列。因此,我们使用LC-MS-MS测定从活化的人外周血白细胞释放的可溶性CD16中的C-端切割位点。我们在CD16的膜近侧区(CD16a与CD16b之间相同的区域)观察到极为接近的三个假定切割位点(图2,箭头)。尽管ADAM17蛋白质水解不需要共有序列,切割区域的二级结构很重要。试图阻断CD16a切割,我们用脯氨酸置换丝氨酸-197(CD16a197P)以引入构象变化。我们通过从活化NK细胞的培养基上清液,并分开地从中性粒细胞的培养基上清液免疫沉淀CD16而鉴定CD16切割的位置。用PNGaseF处理免疫沉淀的CD16以移除N-聚糖,胰蛋白酶消化,对所产生的肽进行质谱分析。鉴定到包含非胰蛋白酶C端的高置信度的四个不同肽谱(peptidepatterns)(图1A)。对于从活化NK细胞的培养基上清液富集的CD16,我们仅观察到一个肽谱,其对应于SEQIDNO:1的氨基酸甘氨酸-174至丙氨酸-195(肽#1,图1A)。CD16a和CD16b的膜近侧区,除了残基176之外,具有相同的氨基酸序列。该位置处苯丙氨酸表示CD16a,其存在于肽#1中(图1A和B)。该肽在丙氨酸-195/缬氨酸-196处显示非胰蛋白酶P1/P1’切割位点(图1B)。对于从活化的中性粒细胞的培养基上清富集的CD16,我们检测到具有非胰蛋白酶C端的三个不同肽谱(肽#2-4,图1A和1B)。肽#2对应于SEQIDNO:2的氨基酸甘氨酸-174至丙氨酸-195,肽#3对应于SEQIDNO:2的氨基酸甘氨酸-174至缬氨酸-196,肽#4对应于SEQIDNO:2的氨基酸天冬酰胺-180至苏氨酸-198。肽#2和肽#3在176位包含缬氨酸,表示CD16b,并在丙氨酸-195/缬氨酸-196和缬氨酸-196/丝氨酸-197处显示P1/P1’位点(图1B)。肽#4在苏氨酸-198/异亮氨酸-199处具有P1/P1’位点(图1B)。尽管该肽从来自富集的中性粒细胞的可溶性CD16衍生,但其不包含176位的氨基酸来鉴定同种型(图1B)。无论如何,高置信度肽显示在CD16中的第三个切割位点。总之,这些发现证明CD16中存在切割区域而不是单个特异性切割位点。我们通过使用定点突变进一步检查CD16的切割区域以确定在基于细胞的测定中是否可破坏CD16a和CD16b切割。ADAM17倾向于优选在底物区域中与其催化位点相互作用的α-螺旋构象。此外,ADAM17切割位点特异性的蛋白质组学研究显示对在P1’、P2’或P3’位置处的脯氨酸残基非常低的偏好性。因此我们用脯氨酸置换CD16a和CD16b切割区域中的丝氨酸-197(S197P,如图2中所标明的)。在人肾细胞系HEK293中分别表达CD16b和CD16b/S197P,该细胞系不表达内源性CD16。HEK293转染子在其表面以相似水平表达CD16b或CD16b/S197P(图3A)。从转染HEK293释放高水平的CD16b,当用PMA处理它们时所述水平进一步增加,如通过ELISA所测定的(图3A)。然而,未处理或PMA处理的HEK293细胞所产生的CD16b/S197P的可溶性水平与CD16b相比显著更低(图3A)。我们还使用相同方法检查了S197P突变对CD16a切割的作用。CD16a的表面表达需要与γ链二聚体联合。我们因此使用稳定表达人γ链的HEK293细胞。比较表达相等表面水平CD16a或CD16a/S197P的HEK293转染子(图3B),我们测定未处理和PMA处理细胞的培养基上清液中每一受体的可溶性水平。再次,当与CD16a相比时观察到显著更低水平的可溶性CD16a/S197P(图3B)。为了评估CD16中经工程改造的S197P突变是否可破坏ADAM17活性,我们还用L-选择素,一个白细胞正常表达的很好地描述的ADAM17底物,转染表达或缺乏CD16b/S197P的HEK293细胞。两种转染子表达相等水平的L-选择素,所述L-选择素水平在用PMA活化后相似地下调(图3C),证明S197P突变影响CD16脱落但不影响ADAM17活性。为了评估S197P突变对NK细胞中CD16a脱落的作用,我们使用人NK细胞系NK92(Gong等人,1994,Leukemia8:652-658)。这些细胞缺乏内源性CD16a的表达,但可稳定表达重组CD16a。我们转导NK92细胞以分别表达CD16a和CD16a/S197P。用PMA活化表达相等水平这些受体的细胞并通过流式细胞术检查细胞表面CD16水平。CD16a,而非CD16a/S197P,经历细胞表面表达的显著下调(图4A)。IL-12和IL-18为NK细胞的生理刺激物,其单独或组合可诱导CD16a脱落。用IL-12和IL-18处理的NK92细胞显示CD16a而非CD16a/S197P在其细胞表面表达中相当可观的下调(图4B)。通过CD16a的结合IgG的细胞的直接参与(engagement)也可诱导其脱落,此处我们通过在抗-CD20mAb利妥昔单抗存在或不存在下将表达CD16a或CD16a/S197P的NK92细胞与CD20阳性Burkitt’s淋巴瘤细胞系Raji孵育来对其进行检查。用利妥昔单抗处理的Raji细胞诱导CD16a而非CD16a/S197P的下调(图4C)。BMS566394为高选择性的ADAM17抑制剂,其具有对于ADAM17比对于其它金属蛋白酶更高的效能数量级。BMS566394以与S197P突变相似的效率阻断CD16a脱落,但对活化的表达CD16a/S197P的NK92细胞没有额外的阻断作用(图4D)。这些发现提供ADAM17是在其切割区域内切割CD16a的主要脱落酶的进一步证据。然而,有可能ADAM17表达水平在表达CD16a或CD16a/S197P的NK92细胞中不相等,导致其不同的脱落。我们因此用多种抗-ADAM17mAbs染色表达CD16a或CD16a/S197P的NK92细胞并观察到相同的细胞表面水平(图4E)。为了确定S197P突变对原代NK细胞的CD16a脱落的作用,我们使用人iPSCs产生经工程改造的NK细胞。我们先前已经报道从iPSCs衍生功能NK细胞及其与外周血NK细胞的相似性(Knorr等人,2013StemCellsTranslMed.2:274-283;Ni等人,2014,StemCells32:1021-1031)。将CD16a和CD16a/S197PcDNA克隆入SleepingBeauty转座子质粒用于基因插入和在iPSC细胞中稳定表达,所述iPSC细胞随后分化为成熟NK细胞。从模拟转导的iPSC细胞衍生的NK细胞表达低水平的内源性CD16a,而转导的CD16a和CD16a/S197P以较高的水平表达(图4F)。当与K562细胞相互作用时NK细胞活化通过多种受体(包括BY55/CD160)发生,导致ADAM17活化和CD16a脱落。我们用K562细胞刺激iPSC衍生的NK细胞,发现CD16a经历显著的细胞表面表达下调,而CD16a/S197P的表达保持稳定(图4F)。内源和重组CD16a具有足够的亲和力以结合单体IgG。为了检查S197P突变对CD16a功能的作用,我们比较CD16a和CD16a/S197P的IgG结合能力。以相等水平表达CD16a或CD16a/S197P的NK92细胞以相似的剂量依赖方式结合IgG(图5A)。对照由结合到表达CD16a或CD16a/S197P的NK92细胞的IgA和结合到NK92亲本细胞的IgG组成。二者均以基本的背景水平发生(图5A)。这些发现证明通过CD16a和CD16a/S197P的特异和相等的IgG结合。CD16a是NK细胞上有效的活化受体,我们检查了当抗体处理的肿瘤细胞参与时经工程改造的S197P突变是否影响CD16a诱导细胞活化的能力。NK92细胞活化通过测量CD107a的上调评估,其在脱粒时非常迅速地发生并且是NK细胞活化的灵敏标志。与用或不用利妥昔单抗处理的Raji细胞孵育的模拟转导NK92细胞显示低水平和相似上调的CD107a(图5B)。当单独与Raji细胞孵育时,以相等水平表达CD16a或CD16a/S197P的NK92细胞也少量上调CD107a,而其与用利妥昔单抗处理的Raji细胞的孵育导致CD107a的相当大上调(图5B)。总之,上述发现表明CD16a中经工程改造的S197P突变不损伤其功能。因此,我们证明CD16a和CD16b中经工程改造的S197P突变在涉及天然ADAM17的基于细胞的测定中有效阻断其脱落。CD16a中的S197P突变还阻断人NK细胞系NK92中的受体脱落,但其不损伤受体功能。表达相等水平CD16a或CD16a/S197P的NK92细胞在一系列抗体浓度中以相似的效率结合单体IgG。另外,当结合到Raji细胞的利妥昔单抗参与时,表达CD16a或CD16a/S197P的NK92细胞以可比较的方式上调活化标志CD107a。多能干细胞允许遗传操作以产生经工程改造的NK细胞。本公开内容描述从表达野生型CD16a或CD16a/S197P的转导iSPCs产生经工程改造的NK细胞。与NK92细胞同样,CD16a在iPSCs衍生的NK细胞中经历脱落,证明当细胞活化时正常的ADAM17活性,而CD16a/S197P不脱落。CD16a和NK细胞细胞毒性功能在癌症患者中可经历相当大的下调。编码CD16a/S197P的cDNA可用于产生稳定的人诱导多能干细胞(iSPCs)和胚胎干细胞(ESCs)。这些干细胞可然后分化为表达CD16a/S197P的原代NK细胞。表达抗切割CD16a/S197P(例如,单核细胞)或CD16b/S197P(例如,中性粒细胞)的其它细胞群也可从hESCs/iPSCs衍生。为了产生用在人类患者中对抗不同形式癌症或感染的NK细胞免疫治疗,CD16a/S197P-表达NK细胞可介导增加的抗体-依赖性细胞毒性(ADCC)活性或其它CD16a介导的活性(例如,IFNγ和TNFα产生)。例如,CD16a/S197P-表达NK细胞可与治疗抗体(例如,曲妥单抗或利妥昔单抗)、双特异性杀伤衔接体(BiKE,例如,CD16×CD33、CD16×CD19或CD16×EP-CAM双特异性杀伤细胞衔接体)或三特异性杀伤细胞衔接体(TriKE)组合。也可产生具有增加的CD16-介导活性的其它治疗细胞群(例如,中性粒细胞、单核细胞、T细胞,等)。CD16a/S197P在人iPSCs或人ESCs中的表达可产生具有增强的抗瘤病况例如,HER2卵巢癌的ADCC活性的NK细胞群。在一些情况下,瘤病况可用治疗抗体例如,曲妥单抗处理。成熟NK细胞可从人胚胎干细胞和iPSCs衍生。可克隆野生型CD16a和/或CD16a/S197P以产生表达单独的CD16a受体的稳定iPSC细胞系或稳定ESC细胞系。可使用任何合适的克隆方法。示例性克隆方法包括,例如,基于病毒的方法、转座子载体(例如,SleepingBeauty)或核转染。在一个实例中,iPSCs可使用SleepingBeauty转座子载体来修饰。载体可包含选择系统,例如,GFP/博来霉素(zeocin)抗性融合蛋白,其允许双选择系统(博来霉素抗性和流式细胞术分选)。iPSCs可分化为成熟NK细胞,如先前所描述的(Ni等人,2011,J.Virol.85:43–50;Knorr等人,2013,StemCellsTranslMed2:274–283;Woll等人,2009,Blood113:6094–6101)。iPSCs中转基因受体的表达可导致衍生NK细胞中高水平表达。未分化iSPCs中CD16表达可破坏NK细胞分化。在这种情况下,CD16表达可使用,例如,CD56或天然CD16a启动子推延,以致CD16a表达更好地与正常NK细胞分化一致。可比较表达相等水平野生型CD16a与CD16a/S197P的NK细胞。CD16构建体的表达水平可通过FACS分选基于GFP表达来匹配,所述GFP表达以与CD16a构建体成比例的方式发生。匹配的CD16a水平可通过FACS验证。NK细胞抗HER2-表达卵巢癌细胞的细胞毒性可在治疗抗体,例如,曲妥单抗存在或不存在下通过标准铬释放测定评估。可评估用非铬标记卵巢癌细胞的抗体-依赖性细胞毒性。可通过ELISA评估NK细胞的细胞因子(例如,IFNγ、TNFα)产生和CD16a的可溶性水平,通过FACS评估CD16a和其它活化标志(例如,CD107a、CD62L)的细胞表面水平。实施例3中描述的人肿瘤异种移植模型可用于体内评估表达不可切割CD16a的NK细胞的抗癌活性。与人CD16不同,当细胞刺激时小鼠CD16不经历胞外域脱落,因此确定CD16a脱落对NK细胞介导的ADCC的作用不能在正常小鼠中建立模型。表1提供实验群组和处理的代表性集合。表1.肿瘤异种移植模型群组n处理#15无处理25仅OVCAR3细胞35OVCAR3+NK细胞/WT-CD16a45OVCAR3+NK细胞/WT-CD16a+曲妥单抗55OVCAR3+NK细胞/CD16a197P65OVCAR3+NK细胞/CD16a197P+曲妥单抗75OVCAR3+NK细胞/仅载体85OVCAR3+NK细胞/载体+曲妥单抗#处理至少进行两次并汇总数据。肿瘤生长和/或消退可通过常规方法每周监测,包括,例如,生物发光成像、超声、CT、MRI、另一种成像技术和/或称重小鼠(Woll等人,2009,Blood113:6094–6101)。也可对小鼠采血(例如,每周)以定量人NK细胞存活。各种效应物功能标志(例如,IFNγ、CD16a)的表达和/或细胞表面水平可使用常规技术例如,通过FACS评估。小鼠可在任何适当时期,例如,60天期间随访。处死时,可检查内脏(例如,脾、肝、肺、肾和/或卵巢)的转移证据(例如,通过生物发光),如先前所描述的(Woll等人,2009,Blood113:6094–6101)。我们的分析允许定义和比较表达野生型CD16a与CD16a/S197P的iPSC-衍生NK细胞的抗体-依赖性细胞毒性活性和体内效能。因此,我们在本文中描述修饰形式的CD16a、表达修饰的CD16a的遗传修饰的细胞(例如,NK细胞、中性粒细胞、单核细胞、T细胞等)和涉及遗传修饰的细胞的方法。例如,表达修饰形式的CD16a,CD16a/S197P的NK细胞显示增加的抗卵巢癌活性,这至少部分上是由于减少的对NK细胞刺激时ADAM17-介导的脱落的敏感性。这继而,在接合(engage)抗体-标记的癌细胞(例如用治疗抗体标记的癌细胞)时,增加抗体-依赖性细胞毒性活性。此外,通过NK细胞的抗体识别增加与肿瘤细胞的接触稳定性并通过其它活化受体(例如NKG2D)支持NK细胞活性。术语“和/或”意指一个或所有所列元素或两个或更多个所列元素的组合;术语“包括”及其变体在说明书和权利要求中这些术语出现处不具有限制性含义;除非另外说明,“一个”、“一种”、“所述”和“至少一个”可交换地使用,意指一个或一个以上;通过端点叙述的数字范围包括该范围内包含的所有数字(例如,1-5包括1、1.5、2、2.75、3、3.80、4、5,等)。在前述说明中,为了清楚起见可能分开描述具体的实施方案。除非另外清楚指明一个具体实施方案的特征与另一个实施方案的特征不相容,否则某些实施方案可包括与一个或多个实施方案有关的本文所描述的相容特征的组合。对于本文所公开的包括不连续步骤的任何方法,所述步骤可以以任何可行次序进行。并且,适当时,可同时进行两个或更多个步骤的任何组合。本发明通过下列实施例说明。应理解的是具体的实施例、材料、量和程序应根据如本文所阐述的本发明的范围和精神广义解读。实施例实施例1质谱法从健康个体采集外周血依照尼苏达大学研究审查委员会(UniversityofMinnesotaInstitutionalReviewBoard)批准的方案根据方案#9708M00134进行。人中性粒细胞和NK细胞分离如先前所描述的进行(Wang等人,2013,BiochimBiophysActa.1833:680-685;Long等人,2010,JLeukocBiol.87:1097-1101;Long等人,2012,JLeukocBiol.92:667-672)。富集的中性粒细胞或NK细胞(在PBS中1×107/ml;Mediatech,Inc.Manassas,VA)用PMA(分别15ng/ml或50ng/ml;Sigma-Aldrich,St.Louis,MO)在37℃活化30分钟。过滤细胞上清液(0.45μm孔径)并使用mAb3G8(BioLegend,Inc.,SanDiego,CA)和Pierce直接免疫沉淀试剂盒(ThermoFisherScientific,Rockford,IL)按照制造商的说明免疫沉淀CD16。纯化的CD16通过靶向几丁质结合结构域的Remove-iTPNGaseF(NewEnglandBioLabs,Inc.,Ipswich,MA)按照制造商的说明脱糖基化。简言之,将10-20μg纯化的CD16在40mMDTT存在下55℃变性10分钟,然后用3μlRemove-iTPNGaseF(NewEnglandBioLabs,inc.,Ipswich,MA)在37℃孵育1小时。然后使用几丁质磁珠从反应中去除Remove-iTPNGaseF。对CD16进行SDS-PAGE并通过氪荧光蛋白染色(ThermoFisherScientific,Rockford,IL)检测可溶性CD16对应的凝胶带,通过CD16免疫印迹分析同一凝胶中邻近的泳道而验证,然后将其切下并用胰蛋白酶对其进行标准凝胶内消化。将从凝胶提取的消化的肽干燥并在98:2:0.01的水:乙腈:甲酸中复溶用于液相色谱-质谱法分析,通过质谱法(VELOSOPBITRAP,ThermoFisherScientific,Rockford,IL)以数据依赖扫描模式分析≤1µg的等分试样,如先前所描述的(Lin-Moshier等人,2013,JBiolChem.288:355-367)。数据库检索用ProteinPilot4.5(ABSciex,Framingham,MA)针对附加污染物数据库(thegpm.org/cRAP/index,109蛋白质)的NCBI参考序列人蛋白质FASTA数据库进行,所述ProteinPilot4.5使用Paragon评分算法(Shilov等人,2007,MolCellProteomics6:1638-1655)。检索参数为:半胱氨酸碘乙酰胺;胰蛋白酶;仪器OrbiMS(1-3ppm)OrbiMS/MS;生物修饰ID焦点,其包括天冬酰胺脱酰胺;彻底的检索努力;和假发现率分析(用相反的数据库)。cDNA表达构建体的产生CD16b作为命名为NA1和NA2的两个等位基因变体存在,其差异为在其胞外区域的N端部分的四个氨基酸。ADAM17以相似的效率切割两个CD16b等位基因变体。对于本研究,我们仅检查NA1变体。CD16a也存在两个等位基因变体,它们在176位具有缬氨酸或苯丙氨酸残基。ADAM17以相似的效率切割CD16a的这两个等位基因变体。对于本研究,我们仅检查缬氨酸等位基因变体CD16a。CD16a和CD16b从人白细胞cDNA扩增,如先前所描述的(Wang等人,2013,BiochimBiophysActa.1833:680-685;Dong等人,2014,ArthritisRheumatol.66:1291-1299)在BamHI和EcoRI限制酶位点单独克隆入pcDNA3.1质粒(Invitrogen,Carlsbad,CA)。构建体然后按照制造商的说明进行Quik-ChangeSite-directedMutagenesis(快速改变定点诱变)(AgilentTechnologies,SantaClara,CA)以将CD16a和CD16b中197位的丝氨酸转换为脯氨酸。所有构建体经测序证实存在预期突变并且不存在任何自发突变。随后将CD16acDNA在BamHI和EcoRI限制酶位点处克隆入Dr.G.Nolan(StanfordUniversity,Stanford,CA)提供的双顺反子逆转录病毒表达载体pBMN-IRES-EGFP。CD16a构建体还如先前所述(Wilber等人,2007,StemCells25:2919-2927;Tian等人,2009,StemCells27:2675-2685)克隆入双顺反子SleepingBeauty转座子质粒(pKT2-IRES-GFP:zeo)中。简言之,使用引物:5'-CCGGAATTCCAGTGTGGCATCATGTGGCAGCTGCTC-3'(正向,SEQIDNO:XX)和5'-CCGGAATTCTCATTTGTCTTGAGGGTCCTTTCT-3'(反向,SEQIDNO:YY)PCR扩增野生型CD16a和CD16a/S197P。有下划线的是EcoRI位点。将EcoRI-消化的CD16a和CD16a/S197PPCR片段单独克隆入pKT2-IRES-GFP:zeo中。正确的CD16a定向和序列通过PCR和测序分析确认。我们先前已经克隆了全长人L选择素(CD62L)cDNA(Feehan等人,1996,JBiolChem.271:7019-7024;Matala等人,2001,JImmunol.167:1617-1623),将所述全长人L选择素(CD62L)cDNA在限制酶位点XbaI处转移至pcDNA3.1载体中。如先前所述(Dong等人,2014,ArthritisRheumatol.66:1291-1299)克隆全长人FcRγcDNA,并修改为使用pcDNA3.1载体。表达重组L-选择素、CD16a和CD16b的细胞系的产生HEK293细胞(人胚胎肾细胞系)和NK92细胞(人NK细胞系)(ATCC,Manassas,VA)按照贮藏所的说明培养。HEK293细胞使用Lipofectamine2000(Invitrogen,Carlsbad,CA)按照制造商的说明用含有或不含CD16b、CD16b/S197P和/或L-选择素的pcDNA3.1瞬时转染。稳定表达人FcRγ的HEK293细胞通过同一方法用含有或不含CD16a或CD16a/S197P的pcDNA3.1瞬时转染。NK92细胞通过先前所描述的逆转录病毒产生和感染程序(Matala等人,2001,JImmunol.167:1617-1623;Walcheck等人,2003,JLeukocBiol.74:389-394;Wang等人,2009,JImmunol.182:2449-2457)用含有或不含CD16a或CD16a/S197P的pBMN-IRES-EGFP稳定转导。构建体表达通过EGFP荧光和CD16染色评估,如通过流式细胞术所测定的。人iPSCs(UCBiPS7,衍生自脐带血CD34细胞)培养(maintained)在小鼠胚胎成纤维细胞上(Knorr等人,2013,StemCellsTranslMed.2:274-283;Ni等人,2014,StemCells32:1021-1031)。CD16a或CD16a/S197P的稳定表达如先前所述使用SleepingBeauty转座子系统来进行(Wilber等人,2007,StemCells25:2919-2927;Tian等人,2009,StemCells27:2675-2685)。简言之,用pKT2-IRES-GFP:zeo与在核转染仪(nucleofector)溶液V中的转座酶DNA的组合(LonzaInc.,Gaithersburg,MD)使用程序设置B16核转染(nucleofect)iPSCs。核转染的细胞立即悬浮在包含博来霉素(50μg/ml)的iPSC生长培养基中并接种到小鼠胚胎成纤维细胞上。从CD16a-hESC和CD16a-iPSC细胞衍生NK细胞hESCs和iPSCs的造血分化如先前所述(Ng等人,2005,Blood106:1601–1603;Ng等人,2008,NatProtoc3:768–776;LeGarff-Tavernier等人,2010,AgingCell9:527–535)进行。简言之,在含有干细胞因子(SCF,40ng/ml)、血管内皮生长因子(VEGF,20ng/ml)和骨形态发生蛋白4(BMP4,20ng/ml)的BPEL培养基中在96孔圆底板的每孔中接种3000个单细胞。BPEL培养基包含Iscove’sModifiedDulbecco’sMedium(IMDM,86ml,Invitrogen,ThermoFisherScientific,Inc.,Waltham.MA)、含GlutmaxI的F12营养混合液(NutrientMixture)(86mL,Invitrogen,ThermoFisherScientific,Inc.,Waltham.MA)、10%去离子牛血清白蛋白(BSA,5ml,Sigma-Aldrich,St.Louis,MO)、5%聚乙烯醇(10ml,Sigma-Aldrich,St.Louis,MO)、亚麻酸(20µl1gm/ml溶液,Sigma-Aldrich,St.Louis,MO)、亚油酸(20µl1gm/ml溶液,Sigma)、Synthecol500x溶液(Sigma-Aldrich,St.Louis,MO)、a-monothioglyceral(3.9µl/100ml,Sigma-Aldrich,St.Louis,MO)、无蛋白杂交瘤混合物II(Invitrogen,ThermoFisherScientific,Inc.,Waltham.MA)、抗坏血酸(5mg/ml,Sigma)、GlutamaxI(Invitrogen,ThermoFisherScientific,Inc.,Waltham.MA)、胰岛素-转铁蛋白-硒100x溶液(Invitrogen,ThermoFisherScientific,Inc.,Waltham.MA)、青霉素/链霉素(Invitrogen,ThermoFisherScientific,Inc.,Waltham.MA)。在造血分化的第11天,将旋转拟胚体(spinembryoidbodies)直接转移至具有或不具EL08-1D2间质细胞的24孔板的提供有细胞因子的NK培养基中(LeGarff-Tavernier等人,2010,AgingCell9:527–535)。培养4-5周后,用APC-、PE-、FITC-和PerCP-cy5.5-偶联的IgG或抗人血液表面抗原的特异性抗体:CD45-PE、CD56-APC、CD56-PE、CD16-PerCP-cy5.5、NKG2D-PE、NKp44-PE、NKp46-PE、CD158b-FITC、CD158e1/2-FITC(BDPharmingen,SanJose,CA)、CD158a/h-PE和CD158i-PE(BeckmanCoulter,Inc.,Pasadena,CA)染色单细胞悬液。抗体染色通过流式细胞术评估。细胞刺激分别用15ng/ml和100ng/mlPMA在37℃活化RPMI1640培养基(Mediatech,Inc.,Manassas,VA)中的HEK293和NK92细胞30分钟。NK92细胞用IL-12(PeproTechInc,RockyHill,NJ)和IL-18(R&DSystems,Inc.,Minneapolis,MN)分别以100ng/ml和400ng/ml活化达到标明的时间点。通过CD16a的NK92细胞活化通过与用抗-CD20mAb利妥昔单抗(1μg/ml)(Genentech,Inc.,SouthSanFrancisco,CA)处理的CD20-阳性Burkitt’s淋巴瘤细胞系Raji(ATCC,按照贮藏所的说明生长)(1:1比率)孵育而介导,如先前所描述的(Romee等人,2013,Blood121:3599-3608)。通过洗涤Raji细胞去除过量的利妥昔单抗。在一些实验中,NK92细胞用选择性ADAM17抑制剂BMS566394(5μM)(Bristol-MyersSquibbCompany,Princeton,NJ)预孵育30分钟。从iPSCs衍生的NK细胞用人红白血病细胞系K562(ATCC,按照贮藏所的说明生长)刺激,如先前所描述的(Romee等人,2013,Blood121:3599-3608)。简言之,与K562靶细胞(2:1比例)在37℃孵育iPSC-衍生的NK细胞4小时。抗体结合测定细胞与单体人IgG和IgA(Sigma-Aldrich,St.Louis,MO)的结合如先前所述(Dong等人,2014,ArthritisRheumatol.66:1291-1299)实施,并进行了一些修改。PBS中5×106/ml的NK92亲本细胞或表达CD16a或CD16a/S197P的转导细胞在4℃用IgG或IgA以标明的浓度一式三份孵育1小时。充分洗涤细胞并用APC-缀合的驴抗人Fc(重链和轻链)抗体(JacksonImmunoresearch,WestGrove,PA)按照制造商的说明进行孵育。洗涤细胞,然后立即通过流式细胞术分析。流式细胞术和ELISA对于细胞染色,阻断非特异性抗体结合位点,细胞用标明的抗体染色并通过流式细胞术检查,如先前所描述的(Wang等人,2013,BiochimBiophysActa.1833:680-685;Romee等人,2013,Blood121:3599-3608)。流式细胞术分析在FACSCanto和LSRII仪器(BDBiosciences,SanJose,CA)上进行。人CD16通过mAbs3G8(BioLegend,Inc.,SanDiego,CA)和DJ130c(SantaCruzBiotech,SantaCruz,CA)检测。CD107a通过mAbH4A3(Biolegend,Inc.,SanDiego,CA)检测。ADAM17通过mAbsM220(Doedens等人,2000,JBiolChem.275:14598-14607)、111633和111623(R&DSystems,Inc.,Minneapolis,MN)检测。人L-选择素通过mAbLAM1-116(AncellCorp.,Stillwater,MN)检测。同种型匹配的阴性对照mAbs用于评估非特异性染色水平。CD16ELISA通过定制的流式微球测定(cytometricbeadassay)进行,如先前所描述的(Wang等人,2013,BiochimBiophysActa.1833:680-685)。统计分析统计分析使用Prism软件(GraphPad,SanDiego,CA)适当时使用ANOVA和学生t检验进行。认为P值<0.05是显著的。实施例2表达相等水平WTCD16a和CD16a197P(CD16a/S197P)的NK细胞的比较CD16构建体的表达水平通过FACS分选基于GFP表达匹配(如上述对于NK92细胞进行的,图2),所述GFP表达以与CD16构建体成比例的方式发生。对于所有测定匹配的CD16a水平通过FACS验证。作为对照,评估用空SleepingBeauty转座子载体修饰的iPSC-衍生NK细胞(仅表达GFP)。iPSC-衍生的NK细胞表达低水平的内源性CD16a(数据未示出)。NK细胞对HER2-表达卵巢癌细胞的细胞毒性在曲妥单抗的存在或不存在下通过标准铬释放测定评估。也进行了用非铬标记卵巢癌细胞的抗体-依赖性细胞毒性。NK细胞的细胞因子(例如,IFNγ、TNFα)产生和CD16a的可溶性水平通过ELISA评估。CD16a和其它活化标志(例如,CD107a、CD62L)的细胞表面水平通过FACS评估。实施例3人肿瘤异种移植模型,其用于检验表达CD16a197P(CD16a/S197P)的iPSC-衍生NK细胞在曲妥单抗存在下是否具有增加的体内抗卵巢癌活性。使用NOD/SCID/γc-/-(NSG)小鼠和经稳定工程改造而表达用于生物发光成像的萤火虫荧光素酶的人卵巢癌细胞系的异种移植模型(Geller等人,2013,Cytotherapy15:1297–1306)用于检验腹膜内(ip)递送NK细胞的抗卵巢癌细胞活性。过表达HER2的OVCAR3卵巢癌细胞系用作体内靶(Hellstrom等人,2001,CancerRes61:2420–2423)。亚致死照射的(225cGY)NSG雌性小鼠腹膜内注射表达荧光素酶用于生物发光成像而产生的OVCAR3(2×105细胞)以定量肿瘤生长或消退(Geller等人,2013,Cytotherapy15:1297–1306)。在小鼠得到单次腹膜内注射20×106NK细胞前,让肿瘤生长7天。然后每隔一天给予小鼠IL-2(5μg/小鼠),持续4周,如先前所描述的(Woll等人,2009,Blood113:6094–6101),以促进NK细胞的体内存活。曲妥单抗以50μg的剂量腹膜内给予,每周一次,持续4周(所述剂量为先前在该模型中使用的剂量)(Warburton等人,2004,Clinicalcancerresearch10:2512–2524)。比较表达相等水平WTCD16或CD16a197P(CDa6a/S197P)的iPSC-衍生NK细胞的体内效能。对照包括仅表达GFP(仅载体)的iPSC-衍生NK细胞,和仅接受卵巢癌细胞的小鼠同龄组。所有小鼠得到相同的IL-2处理。肿瘤生长/消退每周通过生物发光成像和称重小鼠监测,如先前所描述的(Woll等人,2009,Blood113:6094–6101)。每周还对小鼠采血以定量人NK细胞存活。各种效应物功能标志(例如,IFNγ、CD16a)的表达/细胞表面水平通过FACS评估。对小鼠随访达~60天。处死时,通过生物发光检查内脏(例如,脾、肝、肺、肾和/或卵巢)的转移证据,如先前所描述的(Woll等人,2009,Blood113:6094–6101)。示例性实施方案实施方案1.一种经遗传修饰以表达包含膜近侧区和膜近侧区中的氨基酸修饰的CD16多肽的细胞。实施方案2.一种细胞,包含:编码包含膜近侧区和膜近侧区中的氨基酸修饰的CD16多肽的多核苷酸。实施方案3.实施方案1或实施方案2的细胞,其中所述氨基酸药物(medication)反映与CD16膜近侧区野生型氨基酸序列相比一个或多个氨基酸的添加、一个或多个氨基酸的缺失,或一个或多个氨基酸的置换。实施方案4.实施方案3的细胞,其中一个或多个氨基酸的置换包括SEQIDNO:1的197位的丝氨酸残基的置换。实施方案5.任何前述实施方案的细胞,其中所述细胞为自然杀伤(NK)细胞。实施方案6.任何前述实施方案的细胞,其中所述细胞为中性粒细胞。实施方案7.任何前述实施方案的细胞,其中所述细胞为单核细胞。实施方案8.任何前述实施方案的细胞,其中与野生型CD16多肽相比所述修饰的CD16多肽显示减小的对ADAM17介导的脱落的敏感性。实施方案9.任何前述实施方案的细胞,其中与野生型CD16多肽相比所述修饰的CD16多肽显示减小的对NK细胞刺激时的切割的敏感性。实施方案10.一种方法,其包括给予需要这种处理的患者包括以下的治疗:给予患者治疗性NK效应物,和给予患者权利要求1-9中任一项的细胞。实施方案11.实施方案10的方法,其中所述治疗性NK效应物包括治疗剂。实施方案12.实施方案11的方法,其中所述治疗剂特异性识别肿瘤抗原。实施方案13.实施方案12的方法,其中所述治疗剂包括特异性识别肿瘤抗原的抗体或抗体片段。实施方案14.实施方案13的方法,其中所述肿瘤抗原包括HER2。实施方案15.实施方案13或实施方案14的方法,其中所述抗体包括曲妥单抗或利妥昔单抗。实施方案16.实施方案10的方法,其中所述治疗性NK效应物包括双特异性杀伤衔接体(BiKE)。实施方案17.实施方案16的方法,其中所述BiKE包括CD16×CD33BiKE、CD16×CD19BiKE或CD16×EP-CAMBiKE。实施方案18.实施方案10的方法,其中所述治疗性NK效应物包括三特异性杀伤细胞衔接体(TriKE)。实施方案19.实施方案11或16-18中任一项的方法,其中所述治疗剂特异性识别病毒靶。实施方案20.用于改善对患者的治疗的方法,所述治疗包括给予患者治疗性NK效应物,所述方法包括:给予患者权利要求1-9中任一项的细胞。本文所引用的所有专利、专利申请和出版的完整公开内容以及电子可得的材料(包括,例如,核苷酸序列提交(例如GenBank和RefSeq中的)以及氨基酸序列提交(例如SwissProt、PIR、PRF、PDB中的)和来信GenBank和RefSeq中注释的编码区域的翻译)通过引用以其整体结合。如果本申请的公开内容与本文通过引用结合的任何文献的公开内容之间存在任何不一致,应以本申请的公开内容为准。前面的详述和实施例仅为了清楚理解给出。不应从其理解不必要的限制。本发明不限于所显示和描述的确切细节,因为对本领域技术人员显而易见的变更将包含在通过权利要求书界定的本发明内。除非另外说明,说明书和权利要求书中使用的表述组分的量、分子量等的所有数字在所有情况下应理解为被术语“约”修饰。相应地,除非另外指示相反,说明书和权利要求书中列出的数字参数为近似值,其可根据欲通过本发明获得的期需性质而改变。至少,并且不试图将等同物的教义限制在权利要求书的范围中,每一数字参数应至少根据所报道的有效数字的数并通过应用普通舍入技术来解释。尽管阐述本发明的广泛范围的数字范围和参数为近似值,但具体实施例中列出的数值仍尽可能准确地报告。然而,所有数值固有地包含由其各自的检验测量结果中存在的标准偏差所必然得出的范围。所有的标题均为了方便读者并且不应用于限制标题之后的正文的含义,除非如此说明。当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1