控制昆虫害虫的RNA聚合酶II33核酸分子的制作方法
优先权声明
本申请要求2015年3月13日提交的美国临时专利申请序列号62/133,210“rnapolymeraseii33nucleicacidmoleculestocontrolinsectpests”的提交日的权益,该临时专利申请的公开内容据此全文以引用方式并入本文。
本发明整体涉及由昆虫害虫(例如鞘翅目害虫和半翅目害虫)引起的植物损坏的遗传控制。在特定的实施方案中,本发明涉及鉴定靶标编码多核苷酸和非编码多核苷酸,以及使用重组dna技术转录后阻遏或抑制靶标编码多核苷酸和非编码多核苷酸在昆虫害虫细胞中的表达,从而提供植物保护效果。
背景技术:
西方玉米根虫(wcr)(玉米根萤叶甲(diabroticavirgiferavirgiferaleconte))是北美最具破坏性的玉米根虫物种之一,在美国中西部玉米种植区备受关注。北方玉米根虫(ncr)(巴氏根萤叶甲(diabroticabarberismithandlawrence))是与wcr在几乎相同的范围内共栖的近缘物种。还有另外几种叶甲属(diabrotica)的相关亚种是美洲的严重害虫:墨西哥玉米根虫(mcr)(墨西哥玉米根萤叶甲(d.virgiferazeaekrysanandsmith));南方玉米根虫(scr)(十一星根萤叶甲(d.undecimpunctatahowardibarber));黄瓜条根萤叶甲(d.balteataleconte);黄瓜十一星叶甲(d.undecimpunctatatenella);南美叶甲(d.speciosagermar);以及d.u.undecimpunctatamannerheim。美国农业部已估计玉米根虫每年造成10亿美元收益损失,包括8亿美元的产量损失和2亿美元的治理成本。
wcr卵和ncr卵均在夏季期间沉积于土壤中。这些昆虫在整个冬季都停留在卵期。卵为椭圆形、白色,且长度小于0.004英寸。幼虫在五月底或六月初孵出,卵孵出的精确时间由于温度差异和位置不同而每年都有所变化。新孵出的幼虫是白色蠕虫,长度小于0.125英寸。一旦孵出,幼虫便开始以玉米根为食。玉米根虫历经三个幼虫龄期。进食几周后,幼虫蜕皮,进入蛹期。它们在土壤中化蛹,然后在7月和8月以成虫形式从土壤中出现。成体根虫的长度为约0.25英寸。
玉米根虫幼虫在玉米和另外几种禾本科物种上完成发育。在黄色狐尾草上饲养的幼虫较晚出现,并且比起在玉米上饲养的幼虫,成虫的头壳尺寸较小。ellsbury等人,(2005)environ.entomol.34:627-34。wcr成虫以玉米穗丝、花粉和位于暴露的穗尖上的玉米籽为食。如果wcr成虫在玉米生殖组织存在之前出现,则它们可能以叶组织为食,从而减缓植物生长,并且偶尔会杀死寄主植物。然而,一旦有了偏好的穗丝和花粉,成虫便会快速向其转移。ncr成虫还以玉米植物的生殖组织为食,但相比之下很少以玉米叶为食。
玉米中大部分的根虫损害由幼虫进食引起。新孵出的根虫最初以纤细的玉米根毛为食,并钻入根尖内。随着幼虫长得更大,它们以初生根为食并钻入其中。在有大量玉米根虫时,幼虫进食时常导致根修剪,一直到玉米杆的基部。严重的根损伤妨碍根将水和养分运输到植物中的能力、减缓植物生长,并导致籽粒产生减少,从而时常大幅降低总产量。严重的根损伤还时常导致玉米植物倒伏,这使收获变得更加困难,并进一步降低产量。此外,成虫以玉米生殖组织为食可导致穗尖处的穗丝修剪。如果这种“穗丝剪切”在花粉脱落期间足够严重,则传粉可能受到破坏。
可通过作物轮作、化学杀虫剂、生物杀虫剂(例如,形成孢子的革兰氏阳性细菌苏云金芽孢杆菌(bacillusthuringiensis)(bt))、表达bt毒素的转基因植物或这些手段的组合,来尝试控制玉米根虫。作物轮作的缺陷是不必要地限制了农田的用途。另外,一些根虫物种可能在大豆田间产卵,从而降低用玉米和大豆实施的作物轮作的效率。
化学杀虫剂是最为人们所倚重的用于实现玉米根虫控制的策略。尽管如此,使用化学杀虫剂并不是完美的玉米根虫控制策略;如果把化学杀虫剂的成本与使用杀虫剂后仍可能发生的根虫损害所致的损失相加,则美国每年由于玉米根虫可能损失超过10亿美元。大的幼虫群体、暴雨以及杀虫剂应用不当都可导致玉米根虫控制不充分。此外,连续使用杀虫剂可能选择杀虫剂抗性根虫品种,并且由于杀虫剂对非靶标物种存在毒性,所以有严重的影响环境之虞。
椿象和其他半翅目昆虫(异翅亚目)构成了另一大类重要的农业害虫。已知全世界有椿象的50多个近缘物种引起作物损坏。mcpherson&mcpherson(2000)stinkbugsofeconomicimportanceinamericanorthofmexico,crcpress。半翅目昆虫存在于大量的重要作物中,这些作物包括玉蜀黍、大豆、水果、蔬菜和谷类。
椿象在历经多个若虫期后才进入成虫期。这些昆虫在约30至40天内从卵发育为成虫。若虫和成虫均以来自软组织的汁液为食,它们还将消化酶注入软组织中,引起口外组织消化和坏死。然后摄入消化的植物材料和养分。耗尽来自植物维管系统的水和养分造成植物组织损坏。对发育中的籽粒和种子的损害最为显著,因为产量和萌发显著减少。在温暖的气候下会出现多个世代,造成重大的昆虫压力。目前对椿象的管理依赖于在单块田基础上用杀虫剂处理。因此,迫切需要替代性管理策略,来将正在发生的作物损失降至最低限度。
rna干扰(rnai)是一种利用内源性细胞途径的方法,凭借该方法,对整个靶标基因或靶标基因的尺寸足够大的任何部分有特异性的干扰rna(irna)分子(例如dsrna分子)引起由其编码的mrna降解。近年来,在许多物种和实验系统例如秀丽隐杆线虫(caenorhabditiselegans)、植物、昆虫胚胎和组织培养物中的细胞中,已使用rnai执行基因“敲低”。参见例如fire等人,(1998)nature391:806-11;martinez等人,(2002)cell110:563-74;mcmanus和sharp,(2002)naturerev.genetics3:737-47。
rnai通过内源性途径(包括dicer蛋白质复合物)实现mrna的降解。dicer将长的dsrna分子切割成大约20个核苷酸的短片段,称之为小干扰rna(sirna)。sirna解旋成两个单链rna:过客链(passengerstrand)和引导链(guidestrand)。过客链被降解,引导链则被掺入rna诱导的沉默复合物(risc)中。微小核糖核酸(mirna)是从含有与杂交的过客链和引导链相连接的多核苷酸“环”的前体分子切割下来的在结构上非常相似的分子,这些分子可以类似地掺入risc中。当引导链特异性地结合到互补mrna分子并诱导argonaute(risc复合物的催化组分)切割时,发生转录后基因沉默。尽管在一些真核生物诸如植物、线虫类和一些昆虫中,sirna和/或mirna的初始浓度有限,但已知该过程系统性地遍布整个生物体。
只有与sirna和/或mirna互补的转录物被切割和降解,因此mrna表达的敲低是序列特异性的。在植物中,存在dicer基因的几个功能组。rnai的基因沉默效应持续数天,并且在实验条件下,可导致靶向转录物的丰度下降90%或更多,随之发生相应蛋白质的水平降低。在昆虫中,有至少两种dicer基因,其中dicer1促进mirna在argonaute1指引下降解。lee等人,(2004)cell117(1):69-81。dicer2促进sirna在argonaute2指引下降解。
美国专利号7,612,194以及美国专利公布号2007/0050860、2010/0192265和2011/0154545公开了自玉米根萤叶甲(d.v.virgiferaleconte)蛹分离的9112种表达序列标签(est)序列的文库。美国专利号7,612,194和美国专利公布号2007/0050860中提出了将与其中公开的玉米根萤叶甲液泡型h+-atp酶(v-atp酶)的几个特定部分序列之一互补的核酸分子可操作地连接到启动子,以便在植物细胞中表达反义rna。美国专利公布号2010/0192265提出了将启动子可操作地连接到下述核酸分子以便在植物细胞中表达反义rna,该核酸分子与具有未知且未公开的功能的玉米根萤叶甲基因的特定部分序列互补(该部分序列据称与秀丽隐杆线虫(c.elegans)中的c56c10.3基因产物58%相同)。美国专利公布号2011/0154545提出了将启动子可操作地连接到下述核酸分子以便在植物细胞中表达反义rna,该核酸分子与玉米根萤叶甲外被体(coatomer)β亚单位基因的两个特定部分序列互补。另外,美国专利号7,943,819公开了自玉米根萤叶甲幼虫、蛹和切开的中肠分离的906种表达序列标签(est)序列的文库,并且提出了将启动子可操作地连接到下述核酸分子以便在植物细胞中表达双链rna,该核酸分子与玉米根萤叶甲带电的多泡体蛋白4b基因的特定部分序列互补。
除了v-atp酶的几个特定部分序列和具有未知功能的基因的特定部分序列之外,美国专利号7,612,194以及美国专利公布号2007/0050860、2010/0192265和2011/0154545中没有进一步提出使用其中列出的超过9000个序列中的任何特定序列来进行rna干扰。此外,美国专利号7,612,194以及美国专利公布号2007/0050860、2010/0192265和2011/0154545都没有教示其提供的超过9000个序列中的哪些其他序列在用作dsrna或sirna时在玉米根虫物种中会是致命的,甚至没有教示其有任何其他方面的用处。除了带电多泡体蛋白4b基因的特定部分序列之外,美国专利号7,943,819没有提出使用其中列出的超过900个序列中的任何特定序列来进行rna干扰。此外,美国专利号7,943,819没有教示其提供的超过900个序列中的哪些其他序列在用作dsrna或sirna时在玉米根虫物种中会是致命的,甚至没有教示其有任何其他方面的用处。美国专利申请公布号u.s.2013/040173和pct申请公布号wo2013/169923描述了来源于玉米根叶甲(diabroticavirgifera)snf7基因的序列在玉蜀黍中进行rna干扰的用途。(还公开于bolognesi等人,(2012)plosone7(10):e47534.doi:10.1371/journal.pone.0047534中)。
与玉米根虫dna互补的绝大多数序列(诸如前述)在用作dsrna或sirna时,并不提供保护植物免受玉米根虫物种损害的作用。例如,baum等人,(2007)naturebiotechnology25:1322-1326描述了rnai抑制几个wcr基因靶标的作用。这些作者报道,在超过520ng/cm2的极高irna(例如dsrna)浓度下,他们测试的26个靶标基因中有8个不能提供实验上显著的鞘翅目害虫死亡率。
美国专利号7,612,194和美国专利公布号2007/0050860的作者首先报道了玉米植物中的靶向西方玉米根虫的植物界rnai。baum等人,(2007)nat.biotechnol.25(11):1322-6。这些作者描述了用来筛选潜在靶标基因以开发转基因rnai玉蜀黍的高通量活体内饮食rnai系统。在290个靶标的初始基因池中,只有14个表现出控制幼虫的潜力。最有效的双链rna(dsrna)中的一者靶向编码液泡atp酶亚单位a(v-atp酶)的基因,从而以低浓度dsrna引起快速阻抑对应的内源性mrna并触发特异性rnai反应。因此,这些作者首次用文件记载了植物界rnai作为可行的害虫管理工具的潜力,而同时证明了即便从相对小的候选基因组也无法准确地先验鉴定有效靶标。
技术实现要素:
本文公开了用于控制昆虫害虫的核酸分子(例如,靶标基因、dna、dsrna、sirna、mirna、shrna和hprna)及其使用方法,所述昆虫害虫包括例如鞘翅目害虫,诸如玉米根萤叶甲(西方玉米根虫,"wcr”)、巴氏根萤叶甲(北方玉米根虫,"ncr”)、十一星根萤叶甲(南方玉米根虫,"scr”)、墨西哥玉米根萤叶甲(墨西哥玉米根虫,"mcr”)、黄瓜条根萤叶甲、黄瓜十一星叶甲、d.u.undecimpunctata和南美叶甲;以及半翅目害虫,诸如英雄美洲蝽(euschistusheros)(fabr.)(新热带区棕椿象,“bsb”)、褐臭蝽(e.servus(say))(棕椿象)、稻绿蝽(nezaraviridula(l.))(南方绿椿象)、盖德拟壁蝽(piezodorusguildinii(westwood))(红带椿象)、茶翅蝽(halyomorphahalys
在这些和另外的实例中,天然核酸序列可以是靶标基因,其产物可以例如但不限于:参与代谢过程,或参与幼虫或若虫的发育。在一些实例中,通过包含与靶标基因同源的多核苷酸的核酸分子对靶标基因的表达进行转录后抑制对于昆虫害虫可能是致命的,或可能造成昆虫害虫生长减缓和/或生存力下降。在具体的实例中,可选择rna聚合酶ii33kd亚单位(在本文中称为例如rpii33)或rpii33同源物作为用于转录后沉默的靶标基因。在特定的实例中,可用于转录后抑制的靶标基因为rna聚合酶ii33基因,在本文中称为玉米根萤叶甲rpii33-1(例如seqidno:1)、玉米根萤叶甲rpii33-2(例如seqidno:3)的基因,在本文中称为英雄美洲蝽rpii33-1的基因(例如seqidno:76),或在本文中称为英雄美洲蝽rpii33-2的基因(例如seqidno:78)。因此,本文公开了一种经分离的核酸分子,其包含下列多核苷酸:seqidno:1、seqidno:1的互补序列、seqidno:3、seqidno:3的互补序列、seqidno:76、seqidno:76的互补序列、seqidno:78、seqidno:78的互补序列以及/或者前述中任一者的片段(例如seqidno:5-8和seqidno:80-82)。
还公开了包含编码这样的多肽的多核苷酸的核酸分子:该多肽与靶标基因产物(例如rpii33基因的产物)内的氨基酸序列至少约85%相同。例如,核酸分子可包含编码多肽的多核苷酸,所述多肽与seqidno:2(玉米根萤叶甲rpii33-1)、seqidno:4(玉米根萤叶甲rpii33-2)、seqidno:77(英雄美洲蝽rpii33-1)或seqidno:79)英雄美洲蝽rpii33-2)和/或玉米根萤叶甲rpii33-1、玉米根萤叶甲rpii33-2、英雄美洲蝽rpii33-1或英雄美洲蝽rpii33-2的产物中的氨基酸序列至少85%相同。进一步公开了包含这样的多核苷酸的核酸分子:该多核苷酸为编码下述多肽的多核苷酸的反向互补序列,其中该多肽与靶标基因产物内的氨基酸序列至少85%相同。
还公开了可用于产生irna(例如dsrna、sirna、shrna、mirna和hprna)分子的cdna多核苷酸,所述irna分子与昆虫害虫靶标基因(例如rpii33基因)的全部或部分互补。在特定的实施方案中,dsrna、sirna、shrna、mirna和/或hprna可在体外产生或通过基因修饰生物体(诸如植物或细菌)在活体内产生。在特定的实例中,公开了可用于产生下述irna分子的cdna分子:这些irna分子与rpii33基因(例如seqidno:1、seqidno:3、seqidno:76和/或seqidno:78)的全部或部分互补,所述基因例如wcrrpii33基因(例如seqidno:1和/或seqidno:3)或bsbrpii33基因(例如seqidno:76和/或seqidno:78)。
进一步公开了用于抑制鞘翅目害虫中的必需基因表达的构件,以及用于向植物提供鞘翅目害虫防护的构件。一种用于抑制鞘翅目害虫中的必需基因表达的构件是由选自下列的多核苷酸组成的单链或双链rna分子:seqidno:94-97,以及它们的互补序列。用于抑制鞘翅目害虫中的必需基因表达的构件的功能等同物包括与含有seqidno:5、seqidno:6、seqidno:7和/或seqidno:8的鞘翅目rpii33基因的全部或部分基本上同源的单链或双链rna分子。一种用于向植物提供鞘翅目害虫防护的构件是这样的dna分子:该dna分子包含可操作地连接到启动子且编码用于抑制鞘翅目害虫中的必需基因表达的构件的多核苷酸,其中该dna分子能够整合到植物的基因组中。
还公开了用于抑制半翅目害虫中的必需基因表达的构件,以及用于向植物提供半翅目害虫防护的构件。一种用于抑制半翅目害虫中的必需基因表达的构件是由选自下列的多核苷酸组成的单链或双链rna分子:seqidno:100-102,以及它们的互补序列。用于抑制半翅目害虫中的必需基因表达的构件的功能等同物包括与含有seqidno:80、seqidno:81和/或seqidno:82的半翅目rpii33基因的全部或部分基本上同源的单链或双链rna分子。一种用于向植物提供半翅目害虫防护的构件是这样的dna分子:该dna分子包含可操作地连接到启动子且编码用于抑制半翅目害虫中的必需基因表达的构件的多核苷酸,其中该dna分子能够整合到植物的基因组中。
公开了用于控制昆虫害虫(例如鞘翅目或半翅目害虫)群体的方法,包括向昆虫害虫(例如鞘翅目或半翅目害虫)提供irna(例如dsrna、sirna、shrna、mirna和hprna)分子,该irna分子在被所述害虫摄取后发挥作用以抑制所述害虫内的生物功能。
在一些实施方案中,用于控制鞘翅目害虫群体的方法包括向鞘翅目害虫提供irna分子,该irna分子包含选自下列的多核苷酸的全部或部分:seqidno:92、seqidno:92的互补序列、seqidno:93、seqidno:93的互补序列、seqidno:94、seqidno:94的互补序列、seqidno:95、seqidno:95的互补序列、seqidno:96、seqidno:96的互补序列、seqidno:97、seqidno:97的互补序列、与鞘翅目害虫(例如wcr)的天然rpii33多核苷酸杂交的多核苷酸、与鞘翅目害虫的天然rpii33多核苷酸杂交的多核苷酸的互补序列;与叶甲属生物体(例如wcr)的天然编码多核苷酸杂交的多核苷酸,该天然编码多核苷酸包含seqidno:1、3和5-8中任一者的全部或部分;以及与叶甲属生物体的天然编码多核苷酸杂交的多核苷酸的互补序列,该天然编码多核苷酸包含seqidno:1、3和5-8中任一者的全部或部分。
在一些实施方案中,用于控制半翅目害虫群体的方法包括向半翅目害虫提供irna分子,该irna分子包含选自下列的多核苷酸的全部或部分:seqidno:98、seqidno:98的互补序列、seqidno:99、seqidno:99的互补序列、seqidno:100、seqidno:100的互补序列、seqidno:101、seqidno:101的互补序列、seqidno:102、seqidno:102的互补序列、与半翅目害虫(例如bsb)的天然rpii33多核苷酸杂交的多核苷酸、与半翅目害虫的天然rpii33多核苷酸杂交的多核苷酸的互补序列;与半翅目生物体(例如bsb)的天然编码多核苷酸杂交的多核苷酸,该天然编码多核苷酸包含seqidno:76、78和80-82中任一者的全部或部分;以及与半翅目生物体的天然编码多核苷酸杂交的多核苷酸的互补序列,该天然编码多核苷酸包含seqidno:76、78和80-82中任一者的全部或部分。
在特定的实施方案中,从下述dna转录被昆虫害虫摄取后发挥作用以抑制该害虫内的生物功能的irna,该dna包含选自下列的多核苷酸的全部或部分:seqidno:1;seqidno:1的互补序列;seqidno:3;seqidno:3的互补序列;seqidno:76;seqidno:76的互补序列;seqidno:78;seqidno:78的互补序列;叶甲属生物体(例如wcr)的天然编码多核苷酸,其包含seqidno:1、3和5-8中任一者的全部或部分;叶甲属生物体的天然编码多核苷酸的互补序列,所述天然编码多核苷酸包含seqidno:1、3和5-8中任一者的全部或部分;半翅目生物体(例如bsb)的天然编码多核苷酸,其包含seqidno:76、78和80-82中任一者的全部或部分;以及半翅目生物体的天然编码多核苷酸的互补序列,所述天然编码多核苷酸包含seqidno:76、78和80-82中任一者的全部或部分。
本文还公开了下述方法,其中可在基于食料的测定中,或在表达dsrna、sirna、shrna、mirna和/或hprna的基因修饰植物细胞中,向昆虫害虫提供所述dsrna、sirna、shrna、mirna和/或hprna。在这些和另外的实例中,所述dsrna、sirna、shrna、mirna和/或hprna可由害虫摄入。摄入本发明的dsrna、sirna、shrna、mirna和/或hprna随后可引起害虫中的rnai,rnai进而可造成害虫的生存力所必需的基因发生沉默,最终引起害虫死亡。因此,公开了其中向昆虫害虫提供包含可用于亲本控制昆虫害虫的示例性多核苷酸的核酸分子的方法。在特定的实例中,通过使用本发明的核酸分子而控制的鞘翅目和/或半翅目害虫可以是wcr、ncr、scr、十一星根萤叶甲、黄瓜条根萤叶甲、黄瓜十一星叶甲球虫、南美叶甲、d.u.undecimpunctata、、bsb、褐臭蝽、稻绿蝽、盖德拟壁蝽、茶翅蝽、绿椿象、c.marginatum、椿虫、d.furcatus、edessameditabunda、thyantaperditor、horciasnobilellus、taediastigmosa、秘鲁棉红蝽、neomegalotomusparvus、leptoglossuszonatus、niesthreasidae、豆荚草盲蝽,或美国牧草盲蝽。
参考下文结合附图1至2进行的对若干实施方案的详细说明,前述特征和其他特征将变得更加明显。
附图说明
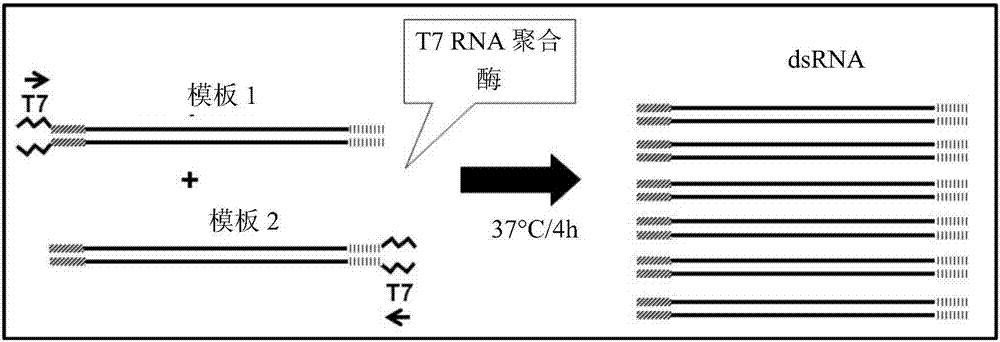
图1包括对用单对引物由单个转录模板提供dsrna所使用的策略的描述。
图2包括对由两个转录模板提供dsrna所使用的策略的描述。
序列表
随附序列表中指定的核酸序列是使用如37c.f.r.§1.822中规定的核苷酸碱基的标准字母缩写示出的。列出的核酸序列和氨基酸序列限定具有以所描述方式布置的核苷酸单体和氨基酸单体的分子(即,分别为多核苷酸和多肽)。列出的核酸序列和氨基酸序列还各自限定包含以所描述方式布置的核苷酸单体和氨基酸单体的一类多核苷酸或多肽。考虑到遗传密码的冗余性,应当理解,包含编码序列的核苷酸序列还描述编码与参考序列所组成的多核苷酸相同的多肽的这类多核苷酸。还应当理解,氨基酸序列描述的是编码该多肽的这类多核苷酸orf。
仅示出了每个核酸序列的一条链,任何对所显示链的提及应当理解为包括互补链在内。由于一级核酸序列的互补序列和反向互补序列必然由该一级序列公开,所以任何对核酸序列的提及也包括该核酸序列的互补序列和反向互补序列在内,除非另有明确说明(或者从其中出现该序列的语境可清楚看出并非如此)。此外,如本领域理解的那样,rna链的核苷酸序列由转录成该rna链的dna的序列决定(但是用尿嘧啶(u)核碱基置换胸腺嘧啶(t)),所以任何对编码rna序列的dna序列的提及都包括该rna序列在内。在随附的序列表中:
seqidno:1示出了示例性wcrrpii33dna,在本文中的一些地方称之为wcrrpii33-1:
gccgatgccatacatacgcttaaaacatcgtatctgctcagttctttaattaacactgaagaaaatcgaattataaaatgccctacgctaacacaccgtcagtacaaatttctgaactaaccgatgaaaatgttaagttcgtcgttgaggacacagaccttagcttggcaaacagtctacgtcgtgttttcatcgctgaaactccaaccctagcaatcgattgggttcaattcgaagccaactccactgtactggcagatgaattccttgcccatcgaattggcttgattccattgatttccgatgaggtagtggacagaatccaaaacactcgtgaatgttcatgcttggacttttgcaccgagtgcagtgtggaatttacattggatgtcaaatgcagcgacgaacatacgcgccacgttaccacggccgatttaaagtccagtgacgcacgagtgctaccagttacgtccagacatcgcgatgacgaggacaacgaatatggagagacgaacgatgaaattctgatcatcaaactgcgcaaaggtcaagagctgaagttgcgagcatacgcgaaaaagggtttcggcaaggaacatgccaaatggaatccaacggctggcgttagctttgaatacgatccagtcaattcgatgagacataccctgtacccgaagccggacgaatggccgaaaagtgagcacaccgaacttgacgatgatcaatacgaagctgaatataactgggaggctaagccgaacaagtttttcttcaacgttgagtcgagtggtgcacttcgaccggaaaacattgtgctgatgggagtcaaagttttgaaaaacaaattgtccaatctacagacgcagttaagtcacgaattgactacaaacgatgcgctcgtgattcagtaaaagcagcgatcccattgaatttcttcaaaatcttgtttttttcctctaag
seqidno:2示出了由示例性wcrrpii33dna(在本文中的一些地方称之为wcrrpii33-1)编码的rpii33多肽的氨基酸序列:
mpyantpsvqiseltdenvkfvvedtdlslanslrrvfiaetptlaidwvqfeanstvladeflahrigliplisdevvdriqntrecscldfctecsveftldvkcsdehtrhvttadlkssdarvlpvtsrhrddedneygetndeiliiklrkgqelklrayakkgfgkehakwnptagvsfeydpvnsmrhtlypkpdewpksehteldddqyeaeynweakpnkfffnvessgalrpenivlmgvkvlknklsnlqtqlshelttndalviq
seqidno:3示出了另外的示例性wcrrpii33dna,在本文中的一些地方称之为wcrrpii33-2:
cgttgacactgttgacagtgacagttgaaattgaaaaccggattagagaagttttcttggaaagttgtttttttaaataactaacattaaatagaagttatttgtttaagggtttaatatgccatatgcaaatcagccatcagttcatataacagatttaacagatgataattgcaaattttatatagaagacactgatttaagtgttgcgaatagcattcgccgcgtccttattgcagaaactcctactctagctatagactgggtaaaattagaagctaactcaactgttctcagtgatgaatttttagcacaccgaattggattgataccattagtttccgatgaagttgtacaaagattacaatatcctagggactgcgtatgtctcgatttttgtcaagaatgcagtgttgaatttactttagatgtaaaatgtacagatgatcaaactcgacatgtaacaactgccgattttaaatctagtgatccacgagtcataccagctacttccaaacatcgtgatgatgaatcctcagagtatggtgaaacagatgaaattcttattattaaactgcgaaagggtcaagagcttaaagttaaagcgtatgccaaaaaaggctttggaaaagagcatgccaaatggaatcctacatgtggtgttgcctttgaatatgatcctgataacgctatgagacatacattatttcctaaaccagacgaatggcctaaaagtgaatacagcgaattagaagatgatcagtatgaagctccatataactgggaattaaaacctaataaattcttctacaatgtggaggctgctggattgttgaaaccagaaaatattgtcatcatgggtgtagctatgttaaaagaaaaactgtcaaatttgcaaacacaactcagccacgaactaacacctgatgttttggccattccaatttaagaagttaattacaatcataggtagagttcattcaaccacagttatacattttttttataatagataagtaagttttacactataggaacaatttttgacatgttgactaaagatcttgttcaaatagactagaaataaaattttgaatccaaaaaaaaaaa
seqidno:4示出了由另外的示例性wcrrpii33dna(即rpii33-2)编码的wcrrpii33多肽的氨基酸序列:
mpyanqpsvhitdltddnckfyiedtdlsvansirrvliaetptlaidwvkleanstvlsdeflahrigliplvsdevvqrlqyprdcvcldfcqecsveftldvkctddqtrhvttadfkssdprvipatskhrddesseygetdeiliiklrkgqelkvkayakkgfgkehakwnptcgvafeydpdnamrhtlfpkpdewpkseyseleddqyeapynwelkpnkffynveaagllkpenivimgvamlkeklsnlqtqlsheltpdvlaipi
seqidno:5示出了在一些实例中用于产生dsrna的示例性wcrrpii33dna,在本文中的一些地方称之为wcrrpii33-1reg1(区域1):
gaattccttgcccatcgaattggcttgattccattgatttccgatgaggtagtggacagaatccaaaacactcgtgaatgttcatgcttggacttttgcaccgagtgcagtgtggaatttacattggatgtcaaatgcagcgacgaacatacgcgccacgttaccacggccgatttaaagtccagtgacgcacgagtgctaccagttacgtccagacatcgcgatgacgaggacaacgaatatggagagacgaacgatgaaattctgatcatcaaactgcgcaaaggtcaagagctgaagttgcgagcatacgcgaaaaagggtttcggcaaggaacatgccaaatggaatccaacggctggcgttagctttgaatacgatccagtcaattcgatgagacataccctgtacccgaagccggacgaatggccgaaaagtgagcacaccgaacttgacgatgatcaatacgaagctgaatataac
seqidno:6示出了在一些实例中用于产生dsrna的另外的示例性wcrrpii33dna,在本文中的一些地方称之为wcrrpii33-2reg1(区域1):
gttctcagtgatgaatttttagcacaccgaattggattgataccattagtttccgatgaagttgtacaaagattacaatatcctagggactgcgtatgtctcgatttttgtcaagaatgcagtgttgaatttactttagatgtaaaatgtacagatgatcaaactcgacatgtaacaactgccgattttaaatctagtgatccacgagtcataccagctacttccaaacatcgtgatgatgaatcctcagagtatggtgaaacagatgaaattcttattattaaactgcgaaagggtcaagagcttaaagttaaagcgtatgccaaaaaaggctttggaaaagagcatgccaaatggaatcctacatgtggtgttgcctttgaatatgatcctgataacgctatgagacatacattatttcctaaaccagacgaatggcctaaaagtgaatacagcgaattagaagatgatcagtatgaagctccatataactggg
seqidno:7示出了在一些实例中用于产生dsrna的另外的示例性wcrrpii33dna,在本文中的一些地方称之为wcrrpii33-2v1(版本1):
ctttagatgtaaaatgtacagatgatcaaactcgacatgtaacaactgccgattttaaatctagtgatccacgagtcataccagctacttccaaacatcgtgatgatgaatcctcagagtatggtgaaacag
seqidno:8示出了在一些实例中用于产生dsrna的另外的示例性wcrrpii33dna,在本文中的一些地方称之为wcrrpii33-2v2(版本2):
gcgtatgccaaaaaaggctttggaaaagagcatgccaaatggaatcctacatgtggtgttgcctttgaatatgatcctgataacgctatgagacatacattatttcctaaaccagacgaatggcc
seqidno:9示出了t7噬菌体启动子的核苷酸序列。
seqidno:10示出了示例性yfp编码序列的片段。
seqidno:11-18示出了用来扩增示例性wcrrpii-33序列的部分的引物,所述序列在一些实例中用于产生dsrna,包括rpii33-1reg1、rpii33-2reg1、rpii33-2v1和rpii33-2v2。
seqidno:19示出了示例性yfp基因。
seqidno:20示出了膜联蛋白区域1的dna序列。
seqidno:21示出了膜联蛋白区域2的dna序列。
seqidno:22示出了β血影蛋白2区域1的dna序列。
seqidno:23示出了β血影蛋白2区域2的dna序列。
seqidno:24示出了mtrp-l4区域1的dna序列。
seqidno:25示出了mtrp-l4区域2的dna序列。
seqidno:26-53示出了用来扩增膜联蛋白、β血影蛋白2、mtrp-l4和yfp的基因区域以合成dsrna的引物。
seqidno:54示出了编码tip41样蛋白的玉蜀黍dna序列。
seqidno:55示出了t20vn引物寡核苷酸的核苷酸序列。
seqidno:56-60示出了用于分析玉蜀黍中的dsrna转录物表达的引物和探针。
seqidno:61示出了用于检测二元载体主干的specr编码区的一部分的核苷酸序列。
seqidno:62示出了用于分析基因组拷贝数的aad1编码区的核苷酸序列。
seqidno:63示出了玉蜀黍转化酶基因的dna序列。
seqidno:64-72示出了用于确定基因拷贝数和检测二元载体主干的dna寡核苷酸的核苷酸序列。
seqidno:73-75示出了用于分析玉蜀黍中的dsrna转录物表达的引物和探针。
seqidno:76示出了示例性bsbrpii33dna,在本文中的一些地方称之为bsbrpii33-1:
gttcggctcgggtgagtgtttaaaccaactacgcatcttgttctcgaacctttgcgaacagtgttcacaaataatgctcggttggtgtaaaggtacctttagagcgtgaccccaacttcttttgactcaccttgcagaaactcgatcactaacaattacgtgtatataatcgattcactacacgaacgatacatggttgtttaggttacattcatgttatctttagtaatgaagttattgagttggcctaattgttgaatgtagttaacagaatgccttatgccaatcaaccttctgttcatgtttcagatttaaccgacgacaatgttaaattccaaatagaagatacagaattaagtgtcgctaacagcctcagaagagtcttcatagctgaaaccccaactttagctattgattgggtgcaattgtctgcaaattctactgttttaagtgatgaatttattgcttctagaatcggacttattcctttaacttctgatgctgcagtcgaaaaattaatctattctagggactgtaattgtactgatttctgcccatcctgtagtgttgagtttactttagatgtcaaatgtgtagatgatcaaactagacatgtgacaactgcagatttaaagactgctgatccatgtgtagttcctgctacatctaaaaatagagatgctgatgccaatgaatatggtgaatcagatgatattttgattgttaaattaagaaaaggacaagagcttaaattgagggcctttgctaagaaaggttttggtaaggaacatgctaagtggaatcctactgctggggtttgttttgagtatgaccctgacaactcaatgaggcatacactgtttccaaaaccagatgagtggccaaaaagtgaatatactgaattagatgaggatcagtatgaagctccatttaattgggaagccaaacctaacaaatttttcttcaatgttgaaagttgtggatctttgcgccccgaaaacatagtattaaaaggagtagaagttctaaaatataaactttctgatttattaattcaattgagtcatgaatcagctggccaagttgatcatatgcctgtttaaccagtttttgtgataaattattatctgaaataattcaattattatatttatattaatgtaaaataaaaagaaatttgataactgaaaaaaaaaaaaaaaaatctattgaaagaatacattcattaatacctttctaaagaaaaattattcaatttaaaattgttgccaaaaagtattcagcatttttttaaaattcaatctaggcatatactactgtaaataaatacaaacaatactttcatttttgtactgttctaaaaattgt
seqidno:77示出了由示例性bsbrpii33dna(即bsbrpii33-1)编码的bsbrpii33多肽的氨基酸序列:
mpyanqpsvhvsdltddnvkfqiedtelsvanslrrvfiaetptlaidwvqlsanstvlsdefiasriglipltsdaavekliysrdcnctdfcpscsveftldvkcvddqtrhvttadlktadpcvvpatsknrdadaneygesddilivklrkgqelklrafakkgfgkehakwnptagvcfeydpdnsmrhtlfpkpdewpkseyteldedqyeapfnweakpnkfffnvescgslrpenivlkgvevlkyklsdlliqlshesagqvdhmpv
seqidno:78示出了示例性bsbrpii33dna,在本文中的一些地方称之为bsbrpii33-2:
tgtaaaacttgttctttaagatctcaagaccttttattagaacatctacaggcttaagagagccctctacaacttctacgtccatgtgcaccgtgtctatttcacaaaggagatctggttcttcctcctcaaccatcggccagtccttcttaagcgtatcttctgtccagtagtttgtggacctagtcttattggttctatcatactcgaacccgacaacagagacaggagaccacttggcatgcatcctccctatccccttcctagcaatacacctaattttcaggctttgattcttcccaagttttgcaattaccggtgtgctttttataaaagtctcgtcactgtcaaattttatgtctttacaagtcacgttaaggggggtctctgaggtgttgctaacatcaagttccatctctacggaacaacgagagcaaagctcatcacagtcacactcttctttatacacaagctctttctttgagtacattgggataagcccaagggactgtgccaatacttcatcggggaggaccgtgttgtttttgatgatttcgacgagatctattgcgatagtaggtacttcagataagaggattctccttagagcattagcataggagactgtaatcccagtgagagtgaatttgatgtgttcgtcgttttgttcgtgaattgtaattttcatgagaaagctggagggcaaaagaaatgaagtaaatttagaagggaacacctgtgaagtatgatcgactacg
seqidno:79示出了由示例性bsbrpii33dna(即bsbrpii33-2)编码的另外的bsbrpii33多肽的氨基酸序列:
mkitiheqndehikftltgitvsyanalrrillsevptiaidlveiiknntvlpdevlaqslglipmyskkelvykeecdcdelcsrcsvemeldvsntsetplnvtckdikfdsdetfikstpviaklgknqslkirciarkgigrmhakwspvsvvgfeydrtnktrstnywtedtlkkdwpmveeeepdllceidtvhmdvevvegslkpvdvlikgleilknkfy
seqidno:80示出了在一些实例中用于产生dsrna的示例性bsbrpii33dna,在本文中的一些地方称之为bsb_rpii33-1reg1(区域1):
ggtgaatcagatgatattttgattgttaaattaagaaaaggacaagagcttaaattgagggcctttgctaagaaaggttttggtaaggaacatgctaagtggaatcctactgctggggtttgttttgagtatgaccctgacaactcaatgaggcatacactgtttccaaaaccagatgagtggccaaaaagtgaatatactgaattagatgaggatcagtatgaagctccatttaattgggaagccaaacctaac
seqidno:81示出了在一些实例中用于产生dsrna的另外的示例性bsbrpii33dna,在本文中的一些地方称之为bsb_rpii33-1v1(版本1):
ttgttttgagtatgaccctgacaactcaatgaggcatacactgtttccaaaaccagatgagtggccaaaaagtgaatatactgaattagatgaggatcagtatgaagctcc
seqidno:82示出了在一些实例中用于产生dsrna的另外的示例性bsbrpii33dna,在本文中的一些地方称之为bsb_rpii33-2reg1(区域1):
cgtcgaaatcatcaaaaacaacacggtcctccccgatgaagtattggcacagtcccttgggcttatcccaatgtactcaaagaaagagcttgtgtataaagaagagtgtgactgtgatgagctttgctctcgttgttccgtagagatggaacttgatgttagcaacacctcagagaccccccttaacgtgacttgtaaagacataaaatttgacagtgacgagacttttataaaaagcacaccggtaattgcaaaacttgggaagaatcaaagcctgaaaattaggtgtattgctaggaaggggatagggaggatgcatgccaagtggtctcctgtctctgttgtcgggttcgagtatgatagaaccaataagactaggtccacaaactactggacag
seqidno:83-88示出了用来扩增示例性bsbrpii-33序列的部分的引物,所述序列在一些实例中用于产生dsrna,包括rpii33-1reg1、rpii33-2reg1和rpii33-1v1。
seqidno:89示出了在一些实例中用于产生dsrna的有义链的示例性yfpv2dna。
seqidno:90和91示出了用于pcr扩增yfp序列yfpv2的引物,所述序列在一些实例中用于产生dsrna。
seqidno:92-102示出了从包含示例性rpii33多核苷酸及其片段的核酸转录的示例性rna。
seqidno:103示出了编码叶甲属rpii33-2v1dsrna的示例性dna;其包含有义多核苷酸、环序列(斜体字)和反义多核苷酸(下划线字体):
ctttagatgtaaaatgtacagatgatcaaactcgacatgtaacaactgccgattttaaatctagtgatccacgagtcataccagctacttccaaacatcgtgatgatgaatcctcagagtatggtgaaacag
seqidno:104示出了编码叶甲属rpii33-2v2dsrna的示例性dna;其包含有义多核苷酸、环序列(斜体字)和反义多核苷酸(下划线字体):
gcgtatgccaaaaaaggctttggaaaagagcatgccaaatggaatcctacatgtggtgttgcctttgaatatgatcctgataacgctatgagacatacattatttcctaaaccagacgaatggcc
seqidno:105-106示出了用于分析dsrna表达的探针。
seqidno:107示出了编码dsrna中的间插环的示例性dna核苷酸序列。
seqidno:108-109示出了从包含示例性rpii33-2多核苷酸片段的核酸转录的示例性dsrna。
seqidno:110-111示出了用于分析玉蜀黍中的dsrna转录物表达的引物。
具体实施方式
i.数个实施方案的概述
我们使用表达dsrna的转基因植物最可能靶向的害虫物种之一西方玉米根虫,将rna干扰(rnai)开发为管理昆虫害虫的工具。迄今为止,被提议作为根虫幼虫中rnai的靶标的大多数基因实际上并没有实现其目的。我们在本文中描述了示例性昆虫害虫西方玉米根虫和新热带区棕椿象中由rnai介导的对rna聚合酶33(rpii33)的敲低,所述亚单位例如在经由摄入或注射rpii33dsrna来递送irna分子时显示出具有致命的表型。在本文的实施方案中,通过进食向昆虫递送rpii33dsrna的能力赋予了对管理昆虫(例如鞘翅目和半翅目)害虫非常有用的rnai效应。通过将rpii33介导的rnai与其他有用的rnai靶标(例如rop(美国专利申请公布号14/577811)、rnapii(美国专利申请公布号14/577854)、如美国专利申请号62/133214中所述的rna聚合酶i1rnai靶标、如美国专利申请号62/133202中所述的rna聚合酶ii215rnai靶标、ncm(美国专利申请号62/095487)、dre4(美国专利申请号14/705,807)、copiα(美国专利申请号62/063,199)、copiβ(美国专利申请号62/063,203)、copiγ(美国专利申请号62/063,192)和copiδ(美国专利申请号62/063,216))组合,影响例如幼虫状态根虫中的多重靶标序列的潜力可增加开发出涉及rnai技术的可持续性昆虫害虫管理方法的机会。
本文公开了用于遗传控制昆虫(例如鞘翅目和/或半翅目)害虫侵染的方法和组合物。还提供了用于鉴定昆虫害虫的生命周期所必需的一个或多个基因以用作rnai介导的昆虫害虫群体控制的靶标基因的方法。可以设计编码rna分子的dna质粒载体,来阻抑生长、存活和/或发育所必需的一个或多个靶标基因。在一些实施方案中,所述rna分子可能能够形成dsrna分子。在一些实施方案中,提供了经由与昆虫害虫中的靶标基因的编码或非编码序列互补的核酸分子来转录后阻遏靶标基因表达或抑制靶标基因的方法。在这些和另外的实施方案中,害虫可摄入一种或多种从与靶标基因的编码或非编码序列互补的核酸分子的全部或一部分转录的dsrna、sirna、shrna、mirna和/或hprna分子,从而提供植物保护效果。
因此,一些实施方案涉及使用与一个或多个靶标基因的编码序列和/或非编码序列互补的dsrna、sirna、shrna、mirna和/或hprna对靶标基因产物的表达进行序列特异性抑制,以实现对昆虫(例如鞘翅目和/或半翅目)害虫的至少部分控制。公开了一组经分离和纯化的核酸分子,其包含例如如以seqidno:1、3、76和78以及它们的片段中的一者示出的多核苷酸。在一些实施方案中,可由这些多核苷酸、其片段或包含这些多核苷酸中的一者的基因表达稳定化的dsrna分子,用于转录后沉默或抑制靶标基因。在某些实施方案中,经分离和纯化的核酸分子包含seqidno:1、3、5-8、76、78和80-82中任一者的全部或部分。
一些实施方案涉及在其基因组中具有编码至少一种irna(例如dsrna)分子的至少一种重组dna的重组宿主细胞(例如植物细胞)。在特定的实施方案中,编码的dsrna分子可以在被昆虫(例如鞘翅目和/或半翅目)害虫摄入时提供,用于转录后沉默或抑制害虫中靶标基因的表达。所述重组dna可包含例如:seqidno:1、3、5-8、76、78和80-82中的任一者;seqidno:1、3、5-8、76、78和80-82中任一者的片段;以及由包含1、3、5-8、76、78和80-82和/或它们的互补序列中的任一者的基因的部分序列组成的多核苷酸。
一些实施方案涉及在其基因组中具有编码至少一种irna(例如dsrna)分子的重组dna的重组宿主细胞,所述irna分子包含下列中任一者的全部或部分:seqidno:92、seqidno:93、seqidno:98或seqidno:99(例如,选自seqidno:94-97和100-102的至少一种多核苷酸),或者它们的互补序列。当被昆虫(例如鞘翅目和/或半翅目)害虫摄入时,所述一种或多种irna分子可沉默或抑制靶标rpii33dna(例如,包含选自seqidno:1、3、5-8、76、78和80-82的多核苷酸的全部或部分的dna)在害虫或害虫后代中的表达,从而造成害虫的生长、发育、生存力和/或进食停止。
在一些实施方案中,在其基因组中具有编码能够形成dsrna分子的至少一种rna分子的至少一种重组dna的重组宿主细胞可以是经转化植物细胞。一些实施方案涉及包含这样的经转化植物细胞的转基因植物。除此类转基因植物之外,还提供了任何转基因植物世代的后代植物、转基因种子和转基因植物产品,它们中的每一者都包含一种或多种重组dna。在特定的实施方案中,可以在转基因植物细胞中表达能够形成dsrna分子的rna分子。因此,在这些和其他实施方案中,可以从转基因植物细胞分离dsrna分子。在特定的实施方案中,所述转基因植物是选自下列的植物:玉米(zeamays)、大豆(glycinemax)、棉花(棉属物种(gossypiumsp.))与禾本科(poaceae)的植物。
其他实施方案涉及用于调控昆虫(例如鞘翅目和/或半翅目)害虫细胞中靶标基因的表达的方法。在这些和其他实施方案中,可提供核酸分子,其中该核酸分子包含编码能够形成dsrna分子的rna分子的多核苷酸。在特定的实施方案中,编码能够形成dsrna分子的rna分子的多核苷酸可以可操作地连接到启动子,并且还可以可操作地连接到转录终止序列。在特定的实施方案中,用于调控昆虫害虫细胞中靶标基因的表达的方法可包括:(a)用包含编码能够形成dsrna分子的rna分子的多核苷酸的载体转化植物细胞;(b)在足以允许包含多个经转化植物细胞的植物细胞培养物发育的条件下培养所述经转化植物细胞;(c)选择已将所述载体整合到其基因组中的经转化植物细胞;以及(d)确定选择的经转化植物细胞包含能够形成由所述载体的多核苷酸编码的dsrna分子的rna分子。可以从基因组中整合有载体并且包含由该载体的多核苷酸编码的dsrna分子的植物细胞再生植物。
还公开了在其基因组中整合有载体的转基因植物,所述载体具有编码能够形成dsrna分子的rna分子的多核苷酸,其中所述转基因植物包含由所述载体的多核苷酸编码的dsrna分子。在特定的实施方案中,在植物中表达能够形成dsrna分子的rna分子足以调控接触所述经转化的植物或植物细胞(例如,通过以所述经转化植物、该植物的一部分(例如根)或植物细胞为食)的昆虫(例如鞘翅目或半翅目)害虫的细胞中靶标基因的表达,使得害虫的生长和/或存活受抑制。本文公开的转基因植物可显示出对昆虫害虫侵染的防护性和/或增强的防护性。特定的转基因植物可显示出对一种或多种选自下列的鞘翅目和/或半翅目害虫的防护性和/或增强的防护性:wcr、bsb、ncr、scr、mcr、黄瓜条根萤叶甲、黄瓜十一星叶甲、d.u.undecimpunctatamannerheim、南美叶甲、油菜露尾甲、英雄美洲蝽、褐臭蝽、稻绿蝽、盖德拟壁蝽、茶翅蝽、绿椿象、c.marginatum(palisotdebeauvois)、椿虫(dallas)、d.furcatus(f.)、edessameditabunda(f.)、thyantaperditor(f.)、horciasnobilellus(berg)、taediastigmosa(berg)、秘鲁棉红蝽、neomegalotomusparvus(westwood)、leptoglossuszonatus(dallas)、niesthreasidae(f.)、豆荚草盲蝽和美国牧草盲蝽。
本文还公开了用于将控制剂(诸如irna分子)递送到昆虫(例如鞘翅目和/或半翅目)害虫的方法。此类控制剂可直接或间接地削弱昆虫害虫群体进食、生长或以其他方式造成宿主损坏的能力。在一些实施方案中,提供了一种方法,包括将稳定化的dsrna分子递送到昆虫害虫以阻抑该害虫中的至少一种靶标基因,从而造成rnai并减轻或消除害虫宿主的植物损坏。在一些实施方案中,抑制昆虫害虫中靶标基因的表达的方法可引起害虫的生长、存活和/或发育停止。
在一些实施方案中,提供了包含irna(例如dsrna)分子的组合物(例如局部用组合物),以供在植物、动物和/或植物或动物的环境中使用,旨在消除或减轻昆虫(例如鞘翅目和/或半翅目)害虫侵染。在特定的实施方案中,该组合物可以是要喂食给昆虫害虫的营养组合物或食物来源,或rnai诱饵。一些实施方案包括使害虫可得到所述营养组合物或食物来源。摄入包含irna分子的组合物可引起该分子被害虫的一个或多个细胞摄取,进而可引起对害虫一个或多个细胞中的至少一种靶标基因表达的抑制。通过在害虫的宿主中提供一种或多种包含irna分子的组合物,可以在任何存在害虫的宿主组织或环境之中或之上,限制或消除由于昆虫害虫侵染造成的对植物或植物细胞的摄入或损害。
本文公开的组合物和方法可与用于控制昆虫(例如鞘翅目和/或半翅目)害虫所致损害的其他方法和组合物一同结合使用。例如,如本文所述的用于保护植物免受昆虫害虫损害的irna分子可在这样的方法中使用:该方法包括额外使用一种或多种对昆虫害虫有效的化学药剂、对这种害虫有效的生物杀虫剂、作物轮作、表现出与rnai介导的方法和rnai组合物的特征不同的特征(例如,在植物中重组产生对昆虫害虫有害的蛋白质(例如bt毒素和pip-1多肽(参见美国专利公布号us2014/0007292a1))的重组基因技术,以及/或者重组表达其他irna分子。
ii.缩写
bsb新热带区棕椿象(英雄美洲蝽)
dsrna双链核糖核酸
est表达序列标签
gi生长抑制
ncbi美国国家生物技术信息中心
gdna基因组脱氧核糖核酸
irna抑制性核糖核酸
orf开放阅读框
rnai核糖核酸干扰
mirna微小核糖核酸
shrna小发夹核糖核酸
sirna小抑制性核糖核酸
hprna发夹核糖核酸
utr非翻译区
wcr西方玉米根虫(玉米根萤叶甲)
ncr北方玉米根虫(巴氏根萤叶甲)
mcr墨西哥玉米根虫(墨西哥玉米根萤叶甲)
pcr聚合酶链式反应
qpcr定量聚合酶链式反应
riscrna诱导的沉默复合物
scr南方玉米根虫(十一星根萤叶甲)
sem平均值标准误差
yfp黄色荧光蛋白
iii.术语
在以下的描述和表中,使用了许多术语。为了提供对说明书和权利要求书的清楚且一致的理解,包括要给予此类术语的范围,提供了以下定义:
鞘翅目害虫:如本文所用,术语“鞘翅目害虫”是指鞘翅目(coleoptera)的害虫昆虫,包括以农作物和作物产品(包括玉米和其他真正的禾本科)为食的叶甲属害虫昆虫。在特定的实例中,鞘翅目害虫选自以下清单:玉米根萤叶甲(wcr)、巴氏根萤叶甲(ncr)、十一星根萤叶甲(scr)、墨西哥玉米根萤叶甲(mcr)、黄瓜条根萤叶甲、黄瓜十一星叶甲、d.u.undecimpunctatamannerheim和南美叶甲。
(与生物体)接触:如本文所用,与生物体(例如鞘翅目或半翅目害虫)“接触”或被生物体“摄取”等术语,就核酸分子而言,包括核酸分子内化到生物体中,例如但不限于:生物体摄入所述分子(例如通过进食);使生物体与包含核酸分子的组合物接触;以及用包含核酸分子的溶液浸泡生物体。
重叠群:如本文所用,术语“重叠群”是指由来源于单个遗传来源的一组重叠dna区段重建的dna序列。
玉米植物:如本文所用,术语“玉米植物”是指物种玉蜀黍(zeamays)的植物。
表达:如本文所用,编码多核苷酸(例如,基因或转基因)的“表达”是指这样的过程,通过该过程,核酸转录单元(包括例如gdna或cdna)的编码信息转化为细胞的操作部分、非操作部分或结构性部分,通常包括蛋白质的合成。基因表达可能受到外部信号的影响,例如细胞、组织或生物体暴露于增加或减少基因表达的药剂。基因表达也可以在从dna到rna再到蛋白质的途径中的任何位置受到调节。调节基因表达例如通过控制对转录、翻译、rna转运和加工、中间分子(诸如mrna)降解的作用,或通过在特异性蛋白分子已产生之后的活化、失活、区室化或降解,或这些的组合而发生。基因表达可通过本领域已知的任何方法,包括但不限于northern印迹法、rt-pcr、western印迹法,或者体外、原位或活体内蛋白质活性测定法,在rna水平或蛋白质水平测量。
遗传物质:如本文所用,术语“遗传物质”包括所有的基因和核酸分子,诸如dna与rna。
半翅目害虫:如本文所用,术语“半翅目害虫”是指半翅目(hemiptera)的害虫昆虫,其以广泛的宿主植物为食并具有刺吸式口器,包括(例如但不限于)蝽科(pentatomidae)、盲蝽科(miridae)、红蝽科(pyrrhocoridae)、缘蝽科(coreidae)、蛛缘蝽科(alydidae)和姬缘蝽科(rhopalidae)的昆虫。在特定的实例中,半翅目害虫选自以下清单:英雄美洲蝽(新热带区棕椿象)、稻绿蝽(南方绿椿象)、盖德拟壁蝽(红带椿象)、茶翅蝽(棕翅蝽)、chinaviahilare(say)(绿椿象)、褐臭蝽(棕椿象)、椿虫(dallas)、dichelopsfurcatus(f.)、edessameditabunda(f.)、thyantaperditor(f.)(新热带区红肩椿象)、chinaviamarginatum(palisotdebeauvois)、horciasnobilellus(berg)(棉花臭虫)、taediastigmosa(berg)、秘鲁棉红蝽、neomegalotomusparvus(westwood)、leptoglossuszonatus(dallas)、niesthreasidae(f.)、豆荚草盲蝽(西部牧草盲蝽)和美国牧草盲蝽。
抑制:如本文所用,术语“抑制”在用来描述对编码多核苷酸(例如基因)的作用时,是指从该编码多核苷酸转录的mrna和/或该编码多核苷酸的肽、多肽或蛋白质产物在细胞水平上可测量地减少。在一些实例中,抑制编码多核苷酸的表达可使得表达近似消失。“特异性抑制”是指在正实现特异性抑制的细胞中对靶标编码多核苷酸的抑制不随之对其他编码多核苷酸(例如基因)的表达产生影响。
昆虫:如本文所用,就害虫而言,术语“昆虫害虫”具体包括鞘翅目昆虫害虫。在一些实例中,术语“昆虫害虫”具体指的是选自以下清单的叶甲属鞘翅目害虫:玉米根萤叶甲(wcr)、巴氏根萤叶甲(ncr)、十一星根萤叶甲(scr)、墨西哥玉米根萤叶甲(mcr)、黄瓜条根萤叶甲、黄瓜十一星叶甲、d.u.undecimpunctatamannerheim和南美叶甲。在一些实施方案中,该术语还包括另一些昆虫害虫,例如半翅目昆虫害虫。
经分离的:“经分离的”生物组分(诸如核酸或蛋白质)已从该组分所天然存在的生物体细胞中的其他生物组分(即其他染色体和染色体外的dna和rna,以及蛋白质)基本上分开、分开产生或纯化出来,同时实现组分中的化学或功能变化(例如,核酸可通过断开将该核酸连接到染色体中的其余dna的化学键而从染色体分离)。已经“经分离的”核酸分子和蛋白质包括通过标准纯化方法纯化的核酸分子和蛋白质。该术语还包括通过在宿主细胞内重组表达而制备的核酸和蛋白质,以及化学合成的核酸分子、蛋白质和肽。
核酸分子:如本文所用,术语“核酸分子”可以指核苷酸的聚合形式,可包括rna、cdna、gdna的有义链和反义链两者,以及上述各项的合成形式和混合聚合物。核苷酸或核碱基可以指核糖核苷酸、脱氧核糖核苷酸,或这两种类型核苷酸中任一者的修饰形式。如本文所用的“核酸分子”与“核酸”和“多核苷酸”是同义词。除非另外指明,核酸分子的长度通常至少为10个碱基。按照惯例,核酸分子的核苷酸序列从该分子的5’端向3’端阅读。核酸分子的“互补序列”是指具有可与该核酸分子的核碱基形成碱基对(即a-t/u和g-c)的核碱基的多核苷酸。
一些实施方案包括含有转录为rna分子的模板dna的核酸,所述rna分子是mrna分子的互补序列。在这些实施方案中,转录为mrna分子的核酸的互补序列以5’至3’取向存在,使得rna聚合酶(其以5’至3’方向转录dna)将从互补序列转录出可与mrna分子杂交的核酸。因此,除非另有明确说明或者从上下文可清楚看出另有所指,术语“互补序列”是指从5’至3’具有可与参考核酸的核碱基形成碱基对的核碱基的多核苷酸。类似地,除非另有明确说明(或者从上下文可清楚看出另有所指),核酸的“反向互补序列”是指取向相反的互补序列。前述内容在以下图解中演示:
atgatgatg多核苷酸
tactactac多核苷酸的“互补序列”
catcatcat多核苷酸的“反向互补序列”
本发明的其他实施方案可包括形成发夹rna的rnai分子。在这些rnai分子中,rna干扰所靶向的核酸的互补序列和所述反向互补序列两者可出现在同一个分子中,使得单链rna分子可以“折叠”到包含互补多核苷酸和反向互补多核苷酸的区域上并在该区域上与自身杂交。
“核酸分子”包括所有的多核苷酸,例如:单链和双链形式的dna;单链形式的rna;和双链形式的rna(dsrna)。术语“核苷酸序列”或“核酸序列”是指作为个别单链或在双链体中的核酸有义链和反义链两者。术语“核糖核酸”(rna)包括irna(抑制性rna)、dsrna(双链rna)、sirna(小干扰rna)、shrna(小发夹rna)、mrna(信使rna)、mirna(微小rna)、hprna(发夹rna)、trna(转移rna,无论是装载有还是卸下了相应的酰化氨基酸)和crna(互补rna)。术语“脱氧核糖核酸”(dna)包括cdna、gdna和dna-rna杂交体。术语“多核苷酸”和“核酸”及其“片段”,本领域技术人员会将其理解为这样的术语:包括两种gdna、核糖体rna、转移rna、信使rna、操纵子,以及较小的工程化改造多核苷酸(其编码或可适于编码肽、多肽或蛋白质)。
寡核苷酸:寡核苷酸是短的核酸聚合物。寡核苷酸可通过切割较长的核酸区段、或通过使单独的核苷酸前体聚合而形成。自动合成仪允许合成长度多达几百个碱基的寡核苷酸。由于寡核苷酸可与互补的核酸结合,所以可用作检测dna或rna的探针。由dna组成的寡核苷酸(寡脱氧核糖核苷酸)可在pcr(一种用于扩增dna的技术)中使用。在pcr中,寡核苷酸通常被称为“引物”,其允许dna聚合酶延伸寡核苷酸并复制互补链。
核酸分子可包括天然存在的核苷酸和/或经修饰的核苷酸,它们通过天然存在的核苷酸连接和/或非天然存在的核苷酸连接而连接在一起。如本领域技术人员易于理解的那样,核酸分子可被化学修饰或生物化学修饰,或者可含有非天然的或衍生化的核苷酸碱基。此类修饰包括例如标记物、甲基化、用类似物置换天然存在的核苷酸中的一者或多者、核苷酸间修饰(例如不带电连接:例如甲基膦酸酯、磷酸三酯、氨基磷酸酯、氨基甲酸酯等;带电连接:例如硫代磷酸酯、二硫代磷酸酯等;悬垂部分:例如肽类;嵌入剂:例如吖啶、补骨脂素等;螯合剂;烷化剂;以及经修饰的连接:例如α异头核酸等)。术语“核酸分子”还包括任何拓扑构象,包括单链的、双链的、部分双链体的、三链体的、发夹形的、圆形的和挂锁形的(padlocked)构象。
如本文所用,就dna而言,术语“编码多核苷酸”、“结构性多核苷酸”或“结构性核酸分子”是指这样的多核苷酸:在被置于适当的调节元件控制下时,经由转录和mrna最终翻译成多肽。就rna而言,术语“编码多核苷酸”是指翻译成肽、多肽或蛋白质的多核苷酸。编码多核苷酸的边界由5’-末端的翻译起始密码子和3’-末端的翻译终止密码子来确定。编码多核苷酸包括但不限于:gdna、cdna、est和重组多核苷酸。
如本文所用,“转录的非编码多核苷酸”是指mrna分子的未翻译成肽、多肽或蛋白质的区段,诸如5'utr、3'utr和内含子区段。另外,“转录的非编码多核苷酸”是指转录成在细胞中起作用的rna的核酸,所述rna例如结构性rna(例如核糖体rna(rrna),举例来说5srrna、5.8srrna、16srrna、18srrna、23srrna和28srrna等)、转移rna(trna),以及snrna诸如u4、u5、u6等。转录的非编码多核苷酸还包括(例如但不限于)小rna(srna),该术语通常用来描述小的细菌非编码rna、小核仁rna(snorna)、微小rna(mirna)、小干扰rna(sirna)、piwi交互作用rna(pirna)和长的非编码rna。还进一步,“转录的非编码多核苷酸”是指这样的多核苷酸:其可天然地在核酸中作为基因内的“间隔序列”存在,并且转录为rna分子。
致命的rna干扰:如本文所用,术语“致命的rna干扰”是指造成向其递送例如dsrna、mirna、sirna、shrna和/或hprna的主体个体死亡或生存力下降的rna干扰。
基因组:如本文所用,术语“基因组”是指存在于细胞核内的染色体dna,也指存在于细胞的亚细胞组分内的细胞器dna。在本发明的一些实施方案中,可将dna分子导入植物细胞中,使得dna分子整合到植物细胞的基因组中。在这些和另外的实施方案中,dna分子可整合到植物细胞的核dna中,或整合到植物细胞的叶绿体或线粒体的dna中。术语“基因组”在应用于细菌时,是指细菌细胞内的染色体和质粒两者。在本发明的一些实施方案中,可将dna分子导入细菌中,使得该dna分子整合到细菌的基因组中。在这些和另外的实施方案中,dna分子可整合于染色体,或者作为稳定的质粒定位或位于稳定的质粒中。
序列同一性:如本文所用,术语“序列同一性”或“同一性”在两个多核苷酸或多肽的语境下,是指在指定比较窗口上以最大对应性比对时,这两个分子的序列中相同的残基。
如本文所用,术语“序列同一性百分比”可以指通过在比较窗口上比较分子的两个最佳比对序列(例如核酸序列或多肽序列)确定的值,其中为了实现这两个序列的最佳比对,该比较窗口中的序列部分相比于参考序列(其不包含添加或缺失)可包含添加或缺失(即空位)。通过确定在两个序列中出现相同的核苷酸或氨基酸残基的位置的数目而产生匹配位置数,用该匹配位置数除以比较窗口中位置的总数,将结果乘以100而产生序列同一性的百分比,从而计算出该百分比。每个位置与参考序列相比均相同的序列被认为与参考序列100%相同,反之亦然。
用于比较的序列比对方法是本领域熟知的。各种程序和比对算法描述于例如以下文献中:smith和waterman(1981)adv.appl.math.2:482;needleman和wunsch(1970)j.mol.biol.48:443;pearson和lipman(1988)proc.natl.acad.sci.u.s.a.85:2444;higgins和sharp(1988)gene73:237-44;higgins和sharp(1989)cabios5:151-3;corpet等人,(1988)nucleicacidsres.16:10881-90;huang等人,(1992)comp.appl.biosci.8:155-65;pearson等人,(1994)methodsmol.biol.24:307-31;tatiana等人,(1999)femsmicrobiol.lett.174:247-50。序列比对方法和同源性计算的详细考虑事项可见于例如altschul等人,(1990)j.mol.biol.215:403-10。
美国国家生物技术信息中心(ncbi)基本局部比对搜索工具(blasttm;altschul等人(1990))可从几个源头(包括美国国家生物技术信息中心(bethesda,md))获得并且可在互联网上获得,以便结合几种序列分析程序使用。怎样使用该程序确定序列同一性的描述可在互联网上blasttm的“帮助”部分获得。为了比较核酸序列,可以采用利用设置为默认参数的默认blosum62矩阵的blasttm(blastn)程序的“blast2序列”功能。在用这种方法评估时,与参考多核苷酸的序列具有甚至更大的序列相似性的核酸将显示同一性百分比增加。
可特异性杂交/特异性互补:如本文所用,术语“可特异性杂交”和“特异性互补”是表明足够程度的互补性的术语,该互补性足以使得在核酸分子与靶标核酸分子之间发生稳定且特异性的结合。两个核酸分子之间的杂交涉及在这两个核酸分子的核碱基之间形成反平行比对。然后,这两个分子能够与相反链上的相应碱基形成氢键以形成双链体分子,如果该双链体分子足够稳定,则可使用本领域熟知的方法检出。多核苷酸与其可特异性杂交的靶标核酸不一定是100%互补的。然而,对于特异性杂交必须存在的互补性的量随所使用的杂交条件而变化。
导致特定严格性程度的杂交条件将根据选择的杂交方法的性质以及杂交核酸的组成和长度而变化。一般说来,杂交的温度和杂交缓冲液的离子强度(尤其是na+和/或mg++浓度)将决定杂交的严格性,但洗涤次数也影响严格性。有关获得特定严格性程度所需要的杂交条件的计算是本领域普通技术人员已知的,并且论述于例如以下文献中:sambrook等人(编辑),molecularcloning:alaboratorymanual,第2版,第1-3卷,coldspringharborlaboratorypress,coldspringharbor,ny,1989年,第9和11章;以及hames和higgins(编辑),nucleicacidhybridization,irlpress,oxford,1985年。有关核酸杂交的进一步详细说明和指导可在以下文献中找到:例如tijssen,"overviewofprinciplesofhybridizationandthestrategyofnucleicacidprobeassays”,载于laboratorytechniquesinbiochemistryandmolecularbiology-hybridizationwithnucleicacidprobes,第i部分,第2章,elsevier,ny,1993年;以及ausubel等人(编辑),currentprotocolsinmolecularbiology,第2章,greenepublishingandwiley-interscience,ny,1995年。
如本文所用,“严格条件”涵盖仅在杂交分子的序列与靶标核酸分子内的同源多核苷酸之间存在小于20%的错配时才发生杂交的条件。“严格条件”包括另外的特定严格性水平。因此,如本文所用,“中等严格性”条件是序列错配超过20%的分子不会杂交的条件;“高严格性”条件是错配超过10%的序列不会杂交的条件;而“极高严格性”条件是错配超过5%的序列不会杂交的条件。
以下是代表性的非限制性杂交条件。
高严格性条件(检测出共享至少90%序列同一性的多核苷酸):在5xssc缓冲液中在65℃下杂交16小时;在2xssc缓冲液中在室温下洗涤两次,每次15分钟;以及在0.5xssc缓冲液中在65℃下洗涤两次,每次20分钟。
中等严格性条件(检测出共享至少80%序列同一性的多核苷酸):在5x-6xssc缓冲液中在65-70℃下杂交16-20小时;在2xssc缓冲液中在室温下洗涤两次,每次5-20分钟;以及在1xssc缓冲液中在55-70℃下洗涤两次,每次30分钟。
非严格对照条件(共享至少50%序列同一性的多核苷酸会杂交):在6xssc缓冲液中在室温至55℃下杂交16-20小时;在2x-3xssc缓冲液中在室温至55℃下至少洗涤两次,每次20-30分钟。
如本文所用,就核酸而言,术语“基本上同源”或“实质性同源性”是指具有在严格条件下与参考核酸杂交的毗连核碱基的多核苷酸。例如,与seqidno:1、3、5-8、76、78和80-82中任一者所代表的参考核酸基本上同源的核酸是在严格条件(例如,前文示出的中等严格性条件)下与所述参考核酸杂交的那些核酸。基本上同源的多核苷酸可具有至少80%的序列同一性。例如,基本上同源的多核苷酸可具有约80%至100%的序列同一性,诸如79%、80%、约81%、约82%、约83%、约84%、约85%、约86%、约87%、约88%、约89%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约98.5%、约99%、约99.5%和约100%。实质性同源性的特性与特异性杂交密切相关。例如,当存在足够程度的互补性时,核酸分子可特异性杂交,以避免核酸在期望特异性结合的情况下(例如,在严格杂交条件下)与非靶标多核苷酸非特异性结合。
如本文所用,术语“直系同源物”是指两种或更多种物种中已经从共同的祖先核酸演变、并且可以在所述两种或更多种物种中保留相同功能的基因。
如本文所用,当以5’至3’方向阅读的多核苷酸的每个核苷酸与以3’至5’方向阅读时的另一多核苷酸的每个核苷酸互补时,两个核酸分子被认为表现出“完全互补性”。与参考多核苷酸互补的多核苷酸将表现出与所述参考多核苷酸的反向互补序列相同的序列。这些术语和描述是本领域中明确定义的,并且是本领域普通技术人员易于理解的。
可操作地连接:当第一多核苷酸与第二多核苷酸具有功能性关系时,则称所述第一多核苷酸与所述第二多核苷酸可操作地连接。以重组方式产生时,可操作地连接的多核苷酸一般是毗连的,并且在必要时可将两个蛋白质编码区连接在同一个阅读框内(例如,在翻译融合的orf中)。然而,可操作地连接的核酸不一定是毗连的。
术语“可操作地连接”在有关调节遗传元件和编码多核苷酸的情况下使用时,意味着该调节元件影响连接的编码多核苷酸的表达。“调节元件”或“控制元件”是指影响转录的时机和水平/量、rna加工或稳定性、或者相关编码多核苷酸的翻译的多核苷酸。调节元件可包括启动子、翻译前导序列、内含子、增强子、茎环结构、阻遏物结合多核苷酸、具有终止序列的多核苷酸、具有多聚腺苷酸化识别序列的多核苷酸,等等。特定的调节元件可位于与其可操作地连接的编码多核苷酸的上游和/或下游。而且,与编码多核苷酸可操作地连接的特定调节元件可位于双链核酸分子的相关互补链上。
启动子:如本文所用,术语“启动子”是指可以在转录起始上游、并且可能参与rna聚合酶和其他蛋白质的识别和结合以启动转录的dna区域。启动子可与用于在细胞中表达的编码多核苷酸可操作地连接,或者启动子可与编码信号肽的多核苷酸可操作地连接,所述多核苷酸可与用于在细胞中表达的编码多核苷酸可操作地连接。“植物启动子”可以是能够启动植物细胞中的转录的启动子。在发育控制下的启动子的实例包括优先启动某些组织中的转录的启动子,所述组织诸如叶、根、种子、纤维、木质部导管、管胞或厚壁组织。此类启动子被称为“组织优先”启动子。仅启动某些组织中的转录的启动子被称为“组织特异性”启动子。“细胞类型特异性”启动子主要驱动一个或多个器官中某些细胞类型(例如,根或叶中的维管细胞)中的表达。“诱导型”启动子可以是可处于环境控制下的启动子。可通过诱导型启动子启动转录的环境条件的实例包括厌氧条件和存在光。组织特异性启动子、组织优先启动子、细胞类型特异性启动子和诱导型启动子构成“非组成型”启动子类别。“组成型”启动子是可以在大多数环境条件下或在大多数组织或细胞类型中有活性的启动子。
在本发明的一些实施方案中可以使用任何诱导型启动子。参见ward等人,(1993)plantmol.biol.22:361-366。利用诱导型启动子,转录速率响应于诱导剂而增大。示例性诱导型启动子包括但不限于:来自acei系统的响应于铜的启动子;来自玉蜀黍的响应于苯磺酰胺除草剂安全剂的in2基因启动子;来自tn10的tet阻遏物;以及来自类固醇激素基因的诱导型启动子,其转录活性可通过糖皮质类固醇激素诱导(schena等人,1991年,proc.natl.acad.sci.usa88:0421)。
示例性组成型启动子包括但不限于:来自植物病毒的启动子,诸如来自花椰菜花叶病毒(camv)的35s启动子、来自水稻肌动蛋白基因的启动子、遍在蛋白启动子、pemu、mas、玉蜀黍h3组蛋白启动子和als启动子,欧洲油菜(brassicanapus)als3结构基因5’的xba1/ncoi片段(或与所述xba1/ncoi片段类似的多核苷酸)(国际pct公布号wo96/30530)。
此外,在本发明的一些实施方案中可以利用任何组织特异性或组织优先启动子。用包含与组织特异性启动子可操作地连接的编码多核苷酸的核酸分子转化的植物可唯一地或优先地在特异性组织中产生所述编码多核苷酸的产物。示例性的组织特异性或组织优先启动子包括但不限于:种子优先启动子,诸如来自菜豆素基因的启动子;叶特异性和光诱导型启动子,诸如来自cab或rubisco的启动子;花药特异性启动子,诸如来自lat52的启动子;花粉特异性启动子,诸如来自zm13的启动子;以及小孢子优先启动子,诸如来自apg的启动子。
大豆植物:如本文所用,术语“大豆植物”是指大豆属(glycine)物种的植物,例如大豆(g.max)。
转化:如本文所用,术语“转化”或“转导”是指将一个或多个核酸分子转移到细胞中。在核酸分子通过将核酸分子并入细胞基因组中、或通过附加型复制而由细胞稳定复制的情况下,细胞被转导到该细胞中的核酸分子“转化”。如本文所用,术语“转化”涵盖可将核酸分子导入这种细胞中的所有技术。实例包括但不限于:用病毒载体转染;用质粒载体转化;电穿孔(fromm等人,1986年,nature319:791-3);脂质体转染(felgner等人,1987年,proc.natl.acad.sci.usa84:7413-7);显微注射(mueller等人,1978年,cell15:579-85);土壤杆菌(agrobacterium)介导的转移(fraley等人,1983年,proc.natl.acad.sci.usa80:4803-7);直接dna摄取;以及微粒轰击(klein等人,1987年,nature327:70)。
转基因:外源性核酸。在一些实例中,转基因可以是编码能够形成dsrna分子的rna的一条或两条链的dna,所述dsrna分子包含与存在于鞘翅目和/或半翅目害虫中的核酸分子互补的多核苷酸。在另外的实例中,转基因可以是反义多核苷酸,其中所述反义多核苷酸的表达抑制靶标核酸的表达,从而产生rnai表型。在又另外的实例中,转基因可以是基因(例如,除草剂耐受性基因、编码工业或药学上有用的化合物的基因,或者编码期望的农业性状的基因)。在这些和其他实例中,转基因可含有与转基因的编码多核苷酸可操作地连接的调节元件(例如启动子)。
载体:导入细胞中例如以产生经转化细胞的核酸分子。载体可包含允许其在宿主细胞中复制的遗传元件,诸如复制起点。载体的实例包括但不限于:质粒、粘粒、噬菌体,或携带外源性dna进入细胞的病毒。载体还可包括一种或多种基因(包括产生反义分子的基因和/或可选择标记基因)以及本领域已知的其他遗传元件。载体可以转导、转化或感染细胞,从而引起细胞表达核酸分子和/或由该载体编码的蛋白质。载体任选地包括辅助核酸分子实现进入细胞的物质(例如脂质体、蛋白质包衣等)。
产量:相对于在相同生长位置在相同时间且在相同条件下生长的检验品种的产量,约100%或更大的稳定化产量。在特定的实施方案中,“提高的产量”或“提高产量”意指相对于在含有相当大密度的对作物有害的鞘翅目和/或半翅目害虫(其为本文的组合物和方法所靶向的害虫)的相同生长位置在相同时间且在相同条件下生长的检验品种的产量,具有105%或更大的稳定化产量的栽培种。
除非特别指明或暗示,如本文所用的术语“一个”、“一种”和“该/所述”表示“至少一个/种”。
除非另外特别地解释,本文使用的全部技术术语和科学术语具有与本公开所属领域的普通技术人员通常所理解的相同的含义。分子生物学中常见术语的定义可在以下出版物中找到:例如lewin’sgenesx,jones&bartlettpublishers,2009(isbn100763766321);krebs等人(编辑),theencyclopediaofmolecularbiology,blackwellscienceltd.,1994(isbn0-632-02182-9);以及meyersr.a.(编辑),molecularbiologyandbiotechnology:acomprehensivedeskreference,vchpublishers,inc.,1995(isbn1-56081-569-8)。除非另外指明,所有百分比均以重量计,所有溶剂混合物比例均以体积计。所有温度均以摄氏度计。
iv.包含昆虫害虫序列的核酸分子
a.概述
本文描述了可用于控制昆虫害虫的核酸分子。在一些实例中,昆虫害虫为鞘翅目(例如叶甲属物种)或半翅目(例如美洲蝽属(euschistus)物种)昆虫害虫。所描述的核酸分子包括靶标多核苷酸(例如,天然基因和非编码多核苷酸)、dsrna、sirna、shrna、hprna和mirna。例如,在一些实施方案中描述了dsrna、sirna、mirna、shrna和/或hprna分子,这些分子可与鞘翅目和/或半翅目害虫中的一种或多种天然核酸的全部或部分特异性互补。在这些和另外的实施方案中,一种或多种天然核酸可以是一种或多种靶标基因,其产物可以例如但不限于:参与代谢过程,或参与幼虫/若虫的发育。本文描述的核酸分子在导入包含与所述核酸分子特异性互补的至少一种天然核酸的细胞中时,可以在细胞中启动rnai,因此降低或消除所述天然核酸的表达。在一些实例中,借助与靶标基因特异性互补的核酸分子降低或消除靶标基因的表达可造成害虫的生长、发育和/或进食减缓或停止。
在一些实施方案中,可以选择昆虫害虫中的至少一种靶标基因,其中该靶标基因包含rpii33多核苷酸。在一些实例中,选择鞘翅目害虫中的靶标基因,其中该靶标基因包含选自seqidno:1、3和5-8的多核苷酸。在一些实例中,选择半翅目害虫中的靶标基因,其中该靶标基因包含选自seqidno:76、78和80-82的多核苷酸。
在其他实施方案中,靶标基因可以是包含这样的多核苷酸的核酸分子:该多核苷酸可以在硅片上(insilico)反向翻译为包含连续氨基酸序列的多肽,所述连续氨基酸序列与rpii33多核苷酸的蛋白质产物的氨基酸序列至少约85%相同(例如,至少84%、85%、约90%、约95%、约96%、约97%、约98%、约99%、约100%或100%相同)。靶标基因可以是昆虫害虫中的任何rpii33多核苷酸,转录后抑制该多核苷酸对害虫的生长、存活和/或生存力造成有害影响,例如旨在为植物提供对抗害虫的保护益处。在特定的实例中,靶标基因是包含这样的多核苷酸的核酸分子,该多核苷酸可以在硅片上反向翻译为包含连续氨基酸序列的多肽,所述连续氨基酸序列与seqidno:2、seqidno:4、seqidno:77或seqidno:79的氨基酸序列至少约85%相同、约90%相同、约95%相同、约96%相同、约97%相同、约98%相同、约99%相同、约100%相同或100%相同。
根据本发明提供了dna,其表达产生包含多核苷酸的rna分子,该多核苷酸与由昆虫(例如鞘翅目和/或半翅目)害虫中的编码多核苷酸编码的天然rna分子的全部或部分特异性互补。在一些实施方案中,在昆虫害虫摄入表达的rna分子之后,可获得害虫细胞中的编码多核苷酸的下调。在特定的实施方案中,可获得害虫细胞中的编码多核苷酸的下调。在特定的实施方案中,昆虫害虫细胞中的编码多核苷酸的下调对害虫的生长、发育和/或存活造成有害影响。
在一些实施方案中,靶标多核苷酸包括转录的非编码rna(诸如5'utr、3'utr)、剪接前导序列、内含子、末端内含子(例如,随后在反式剪接中修饰的5'utrrna)、donatron(例如,提供反式剪接的供体序列所需要的非编码rna)以及靶标昆虫害虫基因的其他非编码转录rna。此类多核苷酸可来源于单顺反子基因和多顺反子基因两者。
本文结合一些实施方案还描述了包含至少一种多核苷酸的irna分子(例如dsrna、sirna、mirna、shrna和hprna),所述多核苷酸与昆虫(例如鞘翅目和/或半翅目)害虫中的靶标核酸的全部或部分特异性互补。在一些实施方案中,irna分子可包含与多个靶标核酸(例如,2、3、4、5、6、7、8、9、10个或更多个靶标核酸)的全部或部分互补的一种或多种多核苷酸。在特定的实施方案中,irna分子可在体外产生,或通过基因修饰生物体(诸如植物或细菌)在活体内产生。还公开了cdna,其可用于产生与昆虫害虫中的靶标核酸的全部或部分特异性互补的dsrna分子、sirna分子、mirna分子、shrna分子和/或hprna分子。进一步描述了在实现特定宿主靶标的稳定转化中使用的重组dna构建体。经转化宿主靶标可从该重组dna构建体表达有效水平的dsrna、sirna、mirna、shrna和/或hprna分子。因此,还描述了一种植物转化载体,其包含与在植物细胞中具有功能的异源性启动子可操作地连接的至少一种多核苷酸,其中所述一种或多种多核苷酸的表达产生包含一串毗连核碱基且与昆虫害虫中的靶标核酸的全部或部分特异性互补的rna分子。
在特定的实例中,可用于控制鞘翅目或半翅目害虫的核酸分子可包括:从叶甲属生物体分离的天然核酸的全部或部分,其包含rpii33多核苷酸(例如,seqidno:1、3和5-8中的任一者);从半翅目生物体分离的天然核酸的全部或部分,其包含rpii33多核苷酸(例如,seqidno:76、78和80-82中的任一者);在表达时产生这样的rna分子的dna,所述rna分子包含与由rpii33编码的天然rna分子的全部或部分特异性互补的多核苷酸;包含与rpii33的全部或部分特异性互补的至少一种多核苷酸的irna分子(例如dsrna、sirna、mirna、shrna和hprna);可用于产生与rpii33的全部或部分特异性互补的dsrna分子、sirna分子、mirna分子、shrna分子和/或hprna分子的cdna;以及在实现特定宿主靶标的稳定转化中使用的重组dna构建体,其中经转化宿主靶标包含前述核酸分子中的一种或多种。
b.核酸分子
本发明尤其提供了抑制昆虫(例如鞘翅目和/或半翅目)害虫的细胞、组织或器官中的靶标基因表达的irna(例如dsrna、sirna、mirna、shrna和hprna)分子;以及能够在细胞或微生物中表达为irna分子以抑制昆虫害虫的细胞、组织或器官中的靶标基因表达的dna分子。
本发明的一些实施方案提供了一种经分离的核酸分子,其包含选自下列的至少一种(例如一种、两种、三种或更多种)多核苷酸:seqidno:1或3;seqidno:1或3的互补序列;seqidno:1或3的至少15个毗连核苷酸的片段(例如seqidno:5-8中的任一者);seqidno:1或3的至少15个毗连核苷酸的片段的互补序列;叶甲属生物体(例如wcr)的天然编码多核苷酸,其包含seqidno:5-8中的任一者;叶甲属生物体的天然编码多核苷酸的互补序列,该天然编码多核苷酸包含seqidno:5-8中的任一者;叶甲属生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段,该天然编码多核苷酸包含seqidno:5-8中的任一者;以及叶甲属生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段的互补序列,该天然编码多核苷酸包含seqidno:5-8中的任一者。
本发明的其他实施方案提供了一种经分离的核酸分子,其包含选自下列的至少一种(例如一种、两种、三种或更多种)多核苷酸:seqidno:76或78;seqidno:76或78的互补序列;seqidno:76或78的至少15个毗连核苷酸的片段(例如seqidno:80-82中的任一者);seqidno:76或78的至少15个毗连核苷酸的片段的互补序列;半翅目生物体(例如bsb)的天然编码多核苷酸,其包含seqidno:80-82中的任一者;半翅目生物体的天然编码多核苷酸的互补序列,该天然编码多核苷酸包含seqidno:80-82中的任一者;半翅目生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段,该天然编码多核苷酸包含seqidno:80-82中的任一者;以及半翅目生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段的互补序列,该天然编码多核苷酸包含seqidno:80-82中的任一者。
在特定的实施方案中,昆虫(例如鞘翅目和/或半翅目)害虫接触或摄取从该经分离的多核苷酸转录的irna抑制了害虫的生长、发育和/或进食。在一些实施方案中,昆虫接触或摄取经由以包含该irna的植物材料或诱饵为食而发生。在一些实施方案中,昆虫接触或摄取经由用包含该irna的组合物喷洒含有该昆虫的植物而发生。
在一些实施方案中,本发明的经分离的核酸分子可包含选自下列的至少一种(例如一种、两种、三种或更多种)多核苷酸:seqidno:92;seqidno:92的互补序列;seqidno:93;seqidno:93的互补序列;seqidno:92或seqidno:93的至少15个毗连核苷酸的片段(例如seqidno:94-97);seqidno:92或seqidno:93的至少15个毗连核苷酸的片段的互补序列;叶甲属生物体的天然编码多核苷酸,其包含seqidno:94-97中的任一者;叶甲属生物体的天然编码多核苷酸的互补序列,该天然编码多核苷酸包含seqidno:94-97中的任一者;叶甲属生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段,该天然编码多核苷酸包含seqidno:94-97中的任一者;以及叶甲属生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段的互补序列,该天然编码多核苷酸包含seqidno:94-97中的任一者。
在其他实施方案中,本发明的经分离的核酸分子可包含选自下列的至少一种(例如一种、两种、三种或更多种)多核苷酸:seqidno:98;seqidno:98的互补序列;seqidno:99;seqidno:99的互补序列;seqidno:98或seqidno:99的至少15个毗连核苷酸的片段(例如seqidno:100-102);seqidno:98或seqidno:99的至少15个毗连核苷酸的片段的互补序列;半翅目(例如bsb)生物体的天然编码多核苷酸,其包含seqidno:100-102中的任一者;半翅目生物体的天然编码多核苷酸的互补序列,该天然编码多核苷酸包含seqidno:100-102中的任一者;半翅目生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段,该天然编码多核苷酸包含seqidno:100-102中的任一者;以及半翅目生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段的互补序列,该天然编码多核苷酸包含seqidno:100-102中的任一者。
在特定的实施方案中,鞘翅目和/或半翅目害虫接触或摄取该经分离的多核苷酸抑制了害虫的存活、生长、发育、繁殖和/或进食。
在某些实施方案中,本发明提供的dsrna分子包含与来自靶标基因的转录物互补的多核苷酸,该靶标基因包含seqidno:1、3、5-8、76、78和80-82以及它们的片段中的任一者,抑制昆虫害虫中的该靶标基因造成害虫的生长、发育或其他生物功能所必需的多肽或多核苷酸剂减少或移除。选择的多核苷酸可与下列各项表现出约80%至约100%的序列同一性:seqidno:1、3、5-8、76、78和80-82中的任一者;seqidno:1、3、5-8、76、78和80-82中任一者的连续片段;以及前述中任一者的互补序列。例如,选择的多核苷酸可与下列各项表现出79%、80%、约81%、约82%、约83%、约84%、约85%、约86%、约87%、约88%、约89%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约98.5%、约99%、约99.5%或约100%的序列同一性:seqidno:1、3、5-8、76、78和80-82中的任一者;seqidno:1、3、5-8、76、78和80-82中任一者的连续片段;以及前述中任一者的互补序列。
在一些实施方案中,能够在细胞或微生物中表达为irna分子以抑制靶标基因表达的dna分子可包含与存在于一个或多个靶标昆虫害虫物种(例如鞘翅目或半翅目害虫物种)中的天然多核苷酸的全部或部分特异性互补的单个多核苷酸,或者dna分子可以是由多个此类特异性互补的多核苷酸构建而成的嵌合体。
在一些实施方案中,核酸分子可包含由“间隔序列”分开的第一多核苷酸和第二多核苷酸。间隔序列可以是包含促进第一多核苷酸和第二多核苷酸之间的二级结构形成(在期望的情况下)的任何核苷酸序列的区域。在一个实施方案中,间隔序列是mrna的有义编码多核苷酸或反义编码多核苷酸的一部分。作为替代,间隔序列可包含能够共价连接到核酸分子的核苷酸或其同源物的任何组合。
例如,在一些实施方案中,dna分子可包含编码一种或多种不同irna分子的多核苷酸,其中所述不同irna分子中的每一种都包含第一多核苷酸和第二多核苷酸,其中第一多核苷酸和第二多核苷酸彼此互补。所述第一多核苷酸和所述第二多核苷酸在rna分子内可通过间隔序列连接。间隔序列可构成所述第一多核苷酸或所述第二多核苷酸的一部分。包含所述第一核苷酸多核苷酸和所述第二核苷酸多核苷酸的rna分子的表达可通过所述第一核苷酸多核苷酸和所述第二核苷酸多核苷酸的特异性分子内碱基配对而引起dsrna分子形成。所述第一多核苷酸或所述第二多核苷酸可与对于昆虫害虫(例如鞘翅目或半翅目害虫)而言天然的多核苷酸(例如,靶标基因或转录的非编码多核苷酸)、其衍生物或其互补多核苷酸基本上相同。
dsrna核酸分子包含聚合核糖核苷酸的双链,并且可包含对磷酸根-糖主干或核苷的修饰。可以适当调整rna结构的修饰以使特异性抑制能够发生。在一个实施方案中,可通过普遍存在的酶促过程来修饰dsrna分子,以便可以生成sirna分子。该酶促过程可利用体外或活体内的rna酶iii酶,诸如真核生物中的dicer。参见elbashir等人,(2001)nature411:494-8;以及hamilton和baulcombe,(1999)science286(5441):950-2。dicer或功能上等同的rna酶iii酶将较大的dsrna链和/或hprna分子切割成较小的寡核苷酸(例如sirna),所述寡核苷酸中每一者的长度为约19-25个核苷酸。由这些酶生成的sirna分子具有2至3个核苷酸3’突出,以及5’磷酸根末端和3’羟基末端。由rna酶iii酶生成的sirna分子在细胞中解旋并分成单链rna。然后,sirna分子与从靶标基因转录的rna特异性杂交,这两种rna分子随后通过固有的细胞rna降解机制得到降解。该过程可引起由靶标生物体中的靶标基因编码的rna有效降解或移除。结果是所靶向的基因发生转录后沉默。在一些实施方案中,通过内源性rna酶iii酶由异源性核酸分子产生的sirna分子可有效介导昆虫害虫中的靶标基因的下调。
在一些实施方案中,核酸分子可包括至少一种可被转录成单链rna分子的非天然存在的多核苷酸,所述单链rna分子能够在活体内通过分子间杂交形成dsrna分子。此类dsrna通常自组装,并且可在昆虫(例如鞘翅目或半翅目)害虫的营养来源中提供,以实现靶标基因的转录后抑制。在这些和另外的实施方案中,核酸分子可包含两种不同的非天然存在的多核苷酸,其中每种多核苷酸与昆虫害虫中的不同靶标基因特异性互补。当向例如鞘翅目和/或半翅目害虫以dsrna分子的形式提供这样的核酸分子时,dsrna分子抑制害虫中至少两种不同靶标基因的表达。
c.获得核酸分子
可使用昆虫(例如鞘翅目和半翅目)害虫中的多种多核苷酸作为靶标来设计核酸分子,诸如irna和编码irna的dna分子。然而,天然多核苷酸的选择并非是直截了当的过程。例如,鞘翅目或半翅目害虫中的天然多核苷酸中只有少数会是有效的靶标。无法肯定地预测特定的天然多核苷酸是否可被本发明的核酸分子有效下调,或者特定的天然多核苷酸的下调是否会对昆虫害虫的生长、生存力、进食和/或存活有不利影响。绝大多数的天然鞘翅目和半翅目害虫多核苷酸,诸如自其分离的est(例如,美国专利号7,612,194中列出的鞘翅目害虫多核苷酸),对害虫的生长和/或生存力没有不利影响。也无法预测,在可以对昆虫害虫有不利影响的天然多核苷酸中,哪些能够在重组技术中使用,从而在宿主植物中表达与此类天然多核苷酸互补的核酸分子,并且在害虫进食后对其造成不利影响,同时不对宿主植物造成危害。
在一些实施方案中,核酸分子(例如,要在昆虫(例如鞘翅目或半翅目)害虫的宿主植物中提供的dsrna分子)被选择为靶向这样的cdna,其编码害虫发育和/或存活所必需的蛋白质或蛋白质部分(诸如涉及代谢或分解代谢生物化学途径、细胞分裂、能量代谢、消化、宿主植物识别等的多肽)。如本文所述,靶标害虫生物体摄入含有一种或多种dsrna(所述dsrna的至少一个区段与靶标害虫生物体的细胞中产生的rna的至少一个基本上相同的区段特异性互补)的组合物,可导致该靶标的死亡或其他抑制作用。可使用来源于昆虫害虫的多核苷酸(dna或rna)来构建免受害虫侵染的植物细胞。例如,可以转化鞘翅目和/或半翅目害虫的宿主植物(例如玉蜀黍或大豆),使其含有来源于鞘翅目和/或半翅目害虫的如本文所提供的一种或多种多核苷酸。转化到宿主中的多核苷酸可编码在经转化宿主内的细胞或生物流体中形成dsrna结构的一种或多种rna,因此,如果害虫与转基因宿主形成营养关系或者当害虫与转基因宿主形成营养关系时,使得dsrna可用。这可引起对害虫细胞中一种或多种基因的表达的阻抑,并最终造成死亡或者对害虫生长或发育的抑制。
在一些实施方案中,靶向实质上参与昆虫(例如鞘翅目或半翅目)害虫的生长和发育的基因。本发明中使用的其他靶标基因可包括例如那些在害虫的生存力、运动、迀移、生长、发育、感染性和进食部位的建立中发挥重要作用的靶标基因。因此,靶标基因可以是管家基因或转录因子。此外,本发明中使用的天然昆虫害虫多核苷酸也可来源于植物、病毒、细菌或昆虫基因的同源物(例如直系同源物),该同源物的功能是本领域技术人员已知的,并且其多核苷酸与靶标害虫的基因组中的靶标基因可特异性杂交。通过杂交鉴定具有已知核苷酸序列的基因的同源物的方法是本领域技术人员已知的。
在其他实施方案中,本发明提供了用于获得核酸分子的方法,该核酸分子包含用于产生irna(例如dsrna、sirna、mirna、shrna和hprna)分子的多核苷酸。一种这样的实施方案包括:(a)分析一种或多种靶标基因在昆虫(例如鞘翅目或半翅目)害虫中,在dsrna介导的基因阻抑后的表达、功能和表型;(b)用探针探查cdna或gdna文库,所述探针包含来自所靶向害虫的多核苷酸的全部或部分或者其同源物,所靶向害虫在dsrna介导的阻抑分析中显示出改变的(例如减弱的)生长或发育表型;(c)鉴定与探针特异性杂交的dna克隆;(d)分离步骤(b)中鉴定的dna克隆;(e)对包含步骤(d)中所分离的克隆的cdna或gdna片段测序,其中测序的核酸分子包含所述rna的全部或大部分或者其同源物;以及(f)化学合成基因或者sirna、mirna、hprna、mrna、shrna或dsrna的全部或大部分。
在另外的实施方案中,用于获得包含用于产生大部分的irna(例如dsrna、sirna、mirna、shrna和hprna)分子的多核苷酸的核酸片段的方法包括:(a)合成第一寡核苷酸引物和第二寡核苷酸引物,它们与来自所靶向的昆虫(例如鞘翅目或半翅目)害虫的天然多核苷酸的一部分特异性互补;以及(b)使用步骤(a)的第一寡核苷酸引物和第二寡核苷酸引物扩增克隆载体中存在的cdna或gdna插入物,其中扩增的核酸分子包含sirna、mirna、hprna、mrna、shrna或dsrna分子的大部分。
核酸可通过多种方法分离、扩增或产生。例如,可通过pcr扩增来源于gdna或cdna文库的靶标多核苷酸(例如,靶标基因或转录的靶标非编码多核苷酸)或其部分来获得irna(例如dsrna、sirna、mirna、shrna和hprna)分子。可从靶标生物体提取dna或rna,并且可使用本领域普通技术人员已知的方法从所述dna或rna制备核酸文库。可使用由靶标生物体生成的gdna文库或cdna文库对靶标基因进行pcr扩增和测序。可使用确认的pcr产物作为体外转录的模板,以在最低限度启动子的情况下生成有义和反义rna。作为替代,核酸分子可通过许多技术中的任一种合成(参见例如ozaki等人,(1992)nucleicacidsresearch,20:5205-5214;以及agrawal等人,(1990)nucleicacidsresearch,18:5419-5423),包括使用自动dna合成仪(例如,p.e.biosystems,inc.(fostercity,calif.)的392或394型dna/rna合成仪)、使用标准化学品(诸如亚磷酰胺化学品)。参见例如beaucage等人,(1992)tetrahedron,48:2223-2311;美国专利号4,980,460、4,725,677、4,415,732、4,458,066和4,973,679。还可采用引起非天然主干基团的替代性化学品,诸如硫代磷酸酯、氨基磷酸酯等。
本发明的rna、dsrna、sirna、mirna、shrna或hprna分子可由本领域技术人员通过手动反应或自动反应以化学或酶促方式产生,或者在包含含有编码rna、dsrna、sirna、mirna、shrna或hprna分子的多核苷酸的核酸分子的细胞中于活体内产生。还可通过部分或完全有机合成产生rna,可通过体外酶促或有机合成导入任何经修饰的核糖核苷酸。可通过细胞rna聚合酶或噬菌体rna聚合酶(例如,t3rna聚合酶、t7rna聚合酶和sp6rna聚合酶)合成rna分子。可用于克隆和表达多核苷酸的表达构建体是本领域已知的。参见例如国际pct公布号wo97/32016,以及美国专利号5,593,874、5,698,425、5,712,135、5,789,214和5,804,693。可以先纯化化学合成的或通过体外酶促合成而合成的rna分子,再将其导入细胞中。例如,可通过用溶剂或树脂提取、沉淀、电泳、色谱法或这些手段的组合从混合物纯化rna分子。作为替代,可以在不纯化或最小程度纯化的情况下使用化学合成的或通过体外酶促合成而合成的rna分子,例如,以避免由于样品加工所致的损失。可将rna分子干燥以贮存,或溶解在水溶液中。该溶液可含有缓冲剂或盐,以促进dsrna分子双链体链退火和/或稳定化。
在特定的实施方案中,可通过单条自身互补的rna链或者由两条互补的rna链形成dsrna分子。dsrna分子可以在活体内或在体外合成。细胞的内源性rna聚合酶可在活体内介导一条或两条rna链的转录,或者可使用克隆的rna聚合酶在活体内或在体外介导转录。转录后抑制昆虫害虫中的靶标基因,通过宿主的器官、组织或细胞类型中的特异性转录(例如,通过使用组织特异性启动子进行);刺激宿主中的环境条件(例如,通过使用响应于感染、应激、温度和/或化学诱导物的诱导型启动子进行);以及/或者工程化改造宿主的某个发育期或发育龄的转录(例如,通过使用发育期特异性启动子进行),可以是宿主靶向的。形成dsrna分子的rna链(不论是体外还是活体内转录的)可以是也可以不是多聚腺苷酸化的,并且可能能够也可能不能够被细胞的翻译装置翻译成多肽。
d.重组载体和宿主细胞转化
在一些实施方案中,本发明还提供了用于导入细胞(例如,细菌细胞、酵母细胞或植物细胞)中的dna分子,其中该dna分子包含这样的多核苷酸,该多核苷酸在表达为rna并被昆虫(例如鞘翅目和/或半翅目)害虫摄取后,可实现对害虫的细胞、组织或器官中的靶标基因的阻抑。因此,一些实施方案提供了重组核酸分子,其包含能够在植物细胞中表达为irna(例如dsrna、sirna、mirna、shrna和hprna)分子以抑制昆虫害虫中的靶标基因表达的多核苷酸。为了启动或增强表达,此类重组核酸分子可包含一种或多种调节元件,所述调节元件可以与能够表达为irna的多核苷酸可操作地连接。在植物中表达基因阻抑分子的方法是已知的,并且可用于表达本发明的多核苷酸。参见例如国际pct公布号wo06/073727;以及美国专利公布号2006/0200878al)。
在具体的实施方案中,本发明的重组dna分子可包含编码可形成dsrna分子的rna的多核苷酸。此类重组dna分子可以编码可形成这样的dsrna分子的rna,所述dsrna分子在被摄入后能够抑制昆虫(例如鞘翅目和/或半翅目)害虫细胞中一种或多种内源性靶标基因的表达。在许多实施方案中,转录的rna可形成这样的dsrna分子,所述dsrna分子可按稳定化形式提供;例如,以发夹和茎环结构的形式提供。
在一些实施方案中,dsrna分子的一条链可通过从多核苷酸转录而形成,所述多核苷酸基本上与选自以下的多核苷酸同源:seqidno:1、3、76和78中的任一者;seqidno:1、3、76和78中的任一者的互补序列;seqidno:1、3、76和78中的任一者的至少15个毗连核苷酸的片段(例如seqidno:5-8和80-82);seqidno:1、3、76和78中的任一者的至少15个毗连核苷酸的片段的互补序列;叶甲属生物体(例如wcr)的天然编码多核苷酸,其包含seqidno:5-8中的任一者;叶甲属生物体的天然编码多核苷酸的互补序列,该天然编码多核苷酸包含seqidno:5-8中的任一者;叶甲属生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段,该天然编码多核苷酸包含seqidno:5-8中的任一者;叶甲属生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段的互补序列,该天然编码多核苷酸包含seqidno:5-8中的任一者;半翅目生物体(例如bsb)的天然编码多核苷酸,其包含seqidno:80-82中的任一者;半翅目生物体的天然编码多核苷酸的互补序列,该天然编码多核苷酸包含seqidno:80-82中的任一者;半翅目生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段,该天然编码多核苷酸包含seqidno:80-82中的任一者;以及半翅目生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段的互补序列,该天然编码多核苷酸包含seqidno:80-82中的任一者。
在其他实施方案中,dsrna分子的一条链可通过从与选自下列的多核苷酸基本上同源的多核苷酸转录而形成:seqidno:5-8和80-82;seqidno:5-8和80-82中任一者的互补序列;seqidno:5-8和80-82中任一者的至少15个毗连核苷酸的片段;以及seqidno:5-8和80-82中任一者的至少15个毗连核苷酸的片段的互补序列。
在特定的实施方案中,编码可形成dsrna分子的rna的重组dna分子可包含这样的编码区:其中至少两个多核苷酸被排列成使得相对于至少一个启动子,一个多核苷酸处于有义取向,并且另一个多核苷酸处于反义取向,其中有义多核苷酸和反义多核苷酸通过例如约五(约5)至约一千(约1000)个核苷酸的间隔序列连接或相连。间隔序列可以在有义多核苷酸和反义多核苷酸之间形成环。有义多核苷酸或反义多核苷酸可与靶标基因(例如,包含seqidno:1、3、5-8、76、78和80-82中任一者的rpii33基因)或其片段基本上同源。然而,在一些实施方案中,重组dna分子可以编码可形成不含间隔序列的dsrna分子的rna。在实施方案中,有义编码多核苷酸和反义编码多核苷酸可具有不同长度。
通过在本发明的重组核酸分子中创建适当的表达盒,可容易地将鉴定为对昆虫害虫存在有害影响或者具有就害虫而言的植物保护效果的多核苷酸掺入表达的dsrna分子中。例如,可通过以下步骤将此类多核苷酸表达为具有茎环结构的发夹:取得对应于靶标基因多核苷酸(例如,包含seqidno:1、3、5-8、76、78和80-82中的任一者,以及前述中任一者的片段的rpii33基因)的第一区段;将该多核苷酸连接到第二区段间隔序列区,所述第二区段间隔序列区与第一区段不是同源或互补的;然后将该连接物连接到第三区段,其中第三区段的至少一部分与第一区段基本上互补。这样的构建体通过第一区段与第三区段的分子内碱基配对而形成茎环结构,其中所述环结构形式包含第二区段。参见例如美国专利公布号2002/0048814和2003/0018993;以及国际pct公布号wo94/01550和wo98/05770。可例如以双链结构诸如茎环结构(例如发夹)的形式生成dsrna分子,由此通过共表达靶标基因的片段(例如在额外的植物可表达盒上的靶标基因的片段)而增强靶向天然昆虫(例如鞘翅目和/或半翅目)害虫多核苷酸的sirna的产生,这造成sirna产生增强,或者减轻甲基化以防止dsrna发夹启动子的转录基因沉默。
本发明的某些实施方案包括将本发明的重组核酸分子导入植物中(即转化),以实现一种或多种irna分子的昆虫(例如鞘翅目和/或半翅目)害虫抑制水平的表达。重组dna分子可以例如是载体,诸如线性或闭合环状质粒。载体系统可以是单一载体或质粒,或者共同含有要导入宿主基因组中的总dna的两个或更多个载体或质粒。另外,载体可以是表达载体。可以例如在合适启动子的控制下将本发明的核酸适当地插入载体中,所述启动子在一种或多种宿主中发挥功能以驱动连接的编码多核苷酸或其他dna元件表达。许多载体可用于该目的,并且对适当载体的选择主要将取决于要插入载体中的核酸的大小和要用载体转化的特定宿主细胞。根据每种载体的功能(例如,扩增dna或表达dna)及与其相容的特定宿主细胞,每种载体含有各种组分。
为了赋予转基因植物对昆虫(例如鞘翅目和/或半翅目)害虫的防护性,例如可以在重组植物的组织或流体内将重组dna转录成irna分子(例如,形成dsrna分子的rna分子)。irna分子可包含与可对宿主植物物种造成损害的昆虫害虫内的相应转录多核苷酸基本上同源且可特异性杂交的多核苷酸。所述害虫可例如通过摄入包含所述irna分子的转基因宿主植物的细胞或流体而接触在转基因宿主植物细胞中转录的所述irna分子。因此,在特定的实例中,靶标基因的表达在侵染转基因宿主植物的鞘翅目和/或半翅目害虫内受到irna分子阻抑。在一些实施方案中,对靶标鞘翅目和/或半翅目害虫中靶标基因表达的阻抑可保护植物免受害虫攻击。
为了使irna分子能够被递送给与已用本发明的重组核酸分子转化的植物细胞处于营养关系的昆虫害虫,需要在植物细胞中表达(即转录)irna分子。因此,重组核酸分子可包含与一种或多种调节元件(诸如在宿主细胞中发挥作用的异源性启动子元件)可操作地连接的本发明的多核苷酸,所述宿主细胞诸如其中要扩增核酸分子的细菌细胞,以及其中要表达核酸分子的植物细胞。
适合用于本发明的核酸分子的启动子包括诱导型启动子、病毒启动子、合成启动子或组成型启动子,它们都是本领域熟知的。描述此类启动子的非限制性实例包括美国专利号6,437,217(玉蜀黍rs81启动子)、5,641,876(水稻肌动蛋白启动子)、6,426,446(玉蜀黍rs324启动子)、6,429,362(玉蜀黍pr-1启动子)、6,232,526(玉蜀黍a3启动子)、6,177,611(玉蜀黍组成型启动子),5,322,938、5,352,605、5,359,142和5,530,196(camv35s启动子),6,433,252(玉蜀黍l3油质蛋白启动子)、6,429,357(水稻肌动蛋白2启动子和水稻肌动蛋白2内含子)、6,294,714(光诱导型启动子)、6,140,078(盐诱导型启动子)、6,252,138(病原体诱导型启动子)、6,175,060(磷缺乏诱导型启动子)、6,388,170(双向启动子)、6,635,806(γ-薏苡醇溶蛋白(coixin)启动子),以及美国专利公布号2009/757,089(玉蜀黍叶绿体醛缩酶启动子)。额外的启动子包括胭脂碱合酶(nos)启动子(ebert等人,(1987)proc.natl.acad.sci.usa84(16):5745-9)和章鱼碱合酶(ocs)启动子(两者都在根癌土壤杆菌(agrobacteriumtumefaciens)的肿瘤诱导质粒上携带);花椰菜花叶病毒组启动子,诸如花椰菜花叶病毒(camv)19s启动子(lawton等人,(1987)plantmol.biol.9:315-24);camv35s启动子(odell等人,(1985)nature313:810-2);玄参花叶病毒35s-启动子(walker等人,(1987)proc.natl.acad.sci.usa84(19):6624-8);蔗糖合酶启动子(yang和russell,(1990)proc.natl.acad.sci.usa87:4144-8);r基因复合体启动子(chandler等人,(1989)plantcell1:1175-83);叶绿素a/b结合蛋白基因启动子;camv35s(美国专利号5,322,938、5,352,605、5,359,142和5,530,196);fmv35s(美国专利号6,051,753和5,378,619);pc1sv启动子(美国专利号5,850,019);scp1启动子(美国专利号6,677,503);以及agrtu.nos启动子(genbanktm登录号v00087;depicker等人,(1982)j.mol.appl.genet.1:561-73;bevan等人,(1983)nature304:184-7)。
在特定的实施方案中,本发明的核酸分子包含组织特异性启动子,诸如根特异性启动子。根特异性启动子驱动唯一地或优先地在根组织中表达可操作连接的编码多核苷酸。根特异性启动子的实例是本领域已知的。参见例如美国专利号5,110,732、5,459,252和5,837,848;opperman等人,(1994)science263:221-3;以及hirel等人,(1992)plantmol.biol.20:207-18。在一些实施方案中,可以在两个根特异性启动子之间克隆根据本发明的用于鞘翅目害虫控制的多核苷酸或片段,所述根特异性启动子相对于所述多核苷酸或片段以相反的转录方向取向、在转基因植物细胞中可操作,并且在转基因植物细胞中表达从而在其中产生rna分子,这些rna分子随后可形成dsrna分子,如前文所述。昆虫害虫可以摄入植物组织中表达的irna分子,从而实现对靶标基因表达的阻抑。
可任选地与核酸可操作地连接的额外的调节元件包括5'utr,5'utr位于启动子元件和编码多核苷酸之间充当翻译前导序列元件。翻译前导序列元件存在于完全加工的mrna中,并且可影响初级转录物的加工和/或rna的稳定性。翻译前导序列元件的实例包括玉蜀黍和矮牵牛花热休克蛋白前导序列(美国专利号5,362,865)、植物病毒外壳蛋白前导序列、植物二磷酸核酮糖羧化酶(rubisco)前导序列等。参见例如turner和foster,(1995)molecularbiotech.3(3):225-36。5'utr的非限制性实例包括gmhsp(美国专利号5,659,122)、phdnak(美国专利号5,362,865)、atant1、tev(carrington和freed,(1990)j.virol.64:1590-7)和agrtunos(genbanktm登录号v00087;bevan等人,(1983)nature304:184-7)。
可任选地与核酸可操作地连接的额外的调节元件还包括3’非翻译元件、3’转录终止区或多聚腺苷酸化区。这些是位于多核苷酸下游的遗传元件,包括提供多聚腺苷酸化信号以及/或者能够影响转录或mrna加工的其他调节信号的多核苷酸。多聚腺苷酸化信号在植物中发挥作用,引起多聚腺苷酸化核苷酸添加至mrna前体的3’端。多聚腺苷酸化元件可以来源于多种植物基因或t-dna基因。3’转录终止区的一个非限制性实例是胭脂碱合酶3’区(nos3’;fraley等人,(1983)proc.natl.acad.sci.usa80:4803-7)。使用不同的3’非翻译区的一个实例在ingelbrecht等人,(1989)plantcell1:671-80中提供。多聚腺苷酸化信号的非限制性实例包括来自豌豆(pisumsativum)rbcs2基因的信号(ps.rbcs2-e9;coruzzi等人,(1984)emboj.3:1671-9)和agrtu.nos(genbanktm登录号e01312)。
一些实施方案可包括植物转化载体,该植物转化载体包含经分离和纯化的dna分子,该dna分子包含与本发明的一种或多种多核苷酸可操作地连接的上述调节元件中的至少一者。所述一种或多种多核苷酸在表达时生成一种或多种包含多核苷酸的irna分子,所述多核苷酸与昆虫(例如鞘翅目和/或半翅目)害虫中的天然rna分子的全部或部分特异性互补。因此,所述一种或多种多核苷酸可包含编码所靶向的鞘翅目和/或半翅目害虫rna转录物内存在的多聚核糖核苷酸的全部或部分的区段,并且可包含所靶向的害虫转录物的全部或部分的反向重复序列。植物转化载体可含有与超过一种靶标多核苷酸特异性互补的多核苷酸,从而允许产生超过一种dsrna以抑制靶标昆虫害虫的一个或多个群体或物种的细胞中两种或更多种基因的表达。可将与不同基因中存在的多核苷酸特异性互补的多核苷酸的区段组合成单个复合核酸分子,以便在转基因植物中表达。这样的区段可以是连续的,或由间隔序列分开。
在其他实施方案中,已含有本发明的至少一种多核苷酸的本发明的质粒可通过在同一质粒中相继插入额外的一种或多种多核苷酸来修饰,其中所述额外的一种或多种多核苷酸与原有的至少一种多核苷酸可操作地连接于相同的调节元件。在一些实施方案中,核酸分子可设计用于抑制多种靶标基因。在一些实施方案中,要抑制的多种基因可获自相同的昆虫(例如鞘翅目或半翅目)害虫物种,这样可增强核酸分子的有效性。在其他实施方案中,基因可来源于不同的昆虫害虫,这样可拓宽一种或多种药剂对其有效的害虫的范围。当靶向多种基因以实现阻抑或者表达和阻抑的组合时,可以工程化改造多顺反子dna元件。
本发明的重组核酸分子或载体可包含赋予经转化细胞(诸如植物细胞)可选择表型的可选择标记。也可使用可选择标记来选择包含本发明的重组核酸分子的植物或植物细胞。所述标记可编码杀生物剂抗性、抗生素抗性(例如,卡那霉素、遗传霉素(g418)、博来霉素、潮霉素等)或除草剂耐受性(例如草甘膦等)。可选择标记的实例包括但不限于:编码卡那霉素抗性并且可以使用卡那霉素、g418等选择的neo基因;编码双丙氨膦抗性的bar基因;编码草甘膦耐受性的突变epsp合酶基因;赋予对溴苯腈的抗性的腈水解酶基因;赋予咪唑啉酮或磺酰脲耐受性的突变乙酰乳酸合酶(als)基因;以及甲氨蝶呤抗性dhfr基因。有多种可选择标记可供使用,其赋予对氨苄青霉素、博来霉素、氯霉素、庆大霉素、潮霉素、卡那霉素、林可霉素、甲氨蝶呤、草丁膦、嘌呤霉素、壮观霉素、利福平、链霉素和四环素等的抗性。此类可选择标记的实例例示于例如美国专利号5,550,318、5,633,435、5,780,708和6,118,047中。
本发明的重组核酸分子或载体还可包含可筛选标记。可使用可筛选标记来监测表达。示例性可筛选标记包括:β-葡萄糖醛酸酶或uida基因(gus),其编码已知各种生色底物的酶(jefferson等人,(1987)plantmol.biol.rep.5:387-405);r-基因座基因,其编码调节植物组织中花色素苷色素(红色)产生的产物(dellaporta等人,(1988)"molecularcloningofthemaizer-njallelebytransposontaggingwithac”,载于第18届斯塔德勒遗传学研讨会(stadlergeneticssymposium),p.gustafson和r.appels编辑,newyork:plenum,第263-82页);β-内酰胺酶基因(sutcliffe等人,(1978)proc.natl.acad.sci.usa75:3737-41);编码已知各种生色底物的酶(例如padac,一种生色头孢菌素)的基因;荧光素酶基因(ow等人,(1986)science234:856-9);xyle基因,其编码可转化生色儿茶酚的儿茶酚双加氧酶(zukowski等人,(1983)gene46(2-3):247-55);淀粉酶基因(ikatu等人,(1990)bio/technol.8:241-2);酪氨酸酶基因,其编码能够将酪氨酸氧化为dopa和多巴醌(其继而缩合成黑色素)的酶(katz等人,(1983)j.gen.microbiol.129:2703-14);以及α-半乳糖苷酶。
在一些实施方案中,在用于创建转基因植物和在植物中表达异源性核酸的方法中,可使用如前文所述的重组核酸分子来制备表现出对昆虫(例如鞘翅目和/或半翅目)害虫的易感性降低的转基因植物。可以例如通过将编码irna分子的核酸分子插入植物转化载体中,然后将这些导入植物中来制备植物转化载体。
用于转化宿主细胞的合适方法包括可将dna导入细胞中的任何方法,诸如通过转化原生质体(参见例如美国专利号5,508,184)、通过干燥/抑制介导的dna摄取(参见例如potrykus等人,(1985)mol.gen.genet.199:183-8)、通过电穿孔(参见例如美国专利号5,384,253)、通过用碳化硅纤维搅拌(参见例如美国专利号5,302,523和5,464,765)、通过土壤杆菌介导的转化(参见例如美国专利号5,563,055、5,591,616、5,693,512、5,824,877、5,981,840和6,384,301)以及通过加速dna包被的粒子(参见例如美国专利号5,015,580、5,550,318、5,538,880、6,160,208、6,399,861和6,403,865),等等。特别可用于转化玉米的技术描述于例如美国专利号7,060,876和5,591,616,以及国际pct公布号wo95/06722中。通过应用诸如这些的技术,可稳定地转化几乎任何物种的细胞。在一些实施方案中,将转化dna整合到宿主细胞的基因组中。在多细胞物种的情况下,可将转基因细胞再生为转基因生物体。可使用这些技术中的任一种来产生转基因植物,例如在其基因组中包含编码一种或多种irna分子的一种或多种核酸的转基因植物。
用于将表达载体导入植物中的最广泛使用的方法以土壤杆菌的天然转化系统为基础。根癌土壤杆菌和发根土壤杆菌(a.rhizogenes)是遗传转化植物细胞的植物致病性土壤细菌。根癌土壤杆菌和发根土壤杆菌的ti质粒和ri质粒分别携带负责遗传转化植物的基因。ti(肿瘤诱导)质粒含有转移到经转化植物的大区段,称为t-dna。ti质粒的另一个区段vir区负责t-dna转移。t-dna区以末端重复序列为边界。在经修饰的二元载体中,肿瘤诱导基因已缺失,vir区的功能用来转移以t-dna边界元件为边界的外来dna。t-区还可含有用于有效回收转基因细胞和植物的可选择标记,以及用于插入转移多核苷酸诸如dsrna编码核酸的多克隆位点。
因此,在一些实施方案中,植物转化载体来源于根癌土壤杆菌的ti质粒(参见例如美国专利号4,536,475、4,693,977、4,886,937和5,501,967,以及欧洲专利号ep0122791)或发根土壤杆菌的ri质粒。额外的植物转化载体包括(例如但不限于)由herrera-estrella等人,(1983)nature303:209-13;bevan等人,(1983)nature304:184-7;klee等人,(1985)bio/technol.3:637-42;以及欧洲专利号ep0120516描述的那些,以及来源于前述载体中的任一者的那些。可以修饰天然地与植物相互作用的其他细菌,诸如中华根瘤菌属(sinorhizobium)、根瘤菌属(rhizobium)和中慢生根瘤菌属(mesorhizobium),以介导到许多各种各样的植物的基因转移。通过获取卸甲ti质粒和合适的二元载体这两者,可使这些植物相关的共生细菌能够胜任基因转移。
在向受体细胞提供外源性dna之后,通常鉴定经转化的细胞以供进一步培养和植物再生。为了提高鉴定经转化细胞的能力,技术人员可能期望采用如前提出的可选择或可筛选标记基因,其中转化载体用来生成转化体。在使用可选择标记的情况下,通过使细胞暴露于一种或多种选择剂而在潜在经转化的细胞群体内鉴定出经转化细胞。在使用可筛选标记的情况下,可针对期望的标记基因性状来筛选细胞。
可将暴露于选择剂后存活的细胞、或者在筛选测定中已被评分为阳性的细胞置于支持植物再生的培养基中培养。在一些实施方案中,可通过包含另外的物质(诸如生长调节剂)来改良任何合适的植物组织培养基(例如ms培养基和n6培养基)。可将组织维持在具有生长调节剂的基础培养基上,直到可得到足够的组织用于启动植物再生工作时为止,或者在重复多轮的手动选择之后,直到组织形态适合于再生时为止(例如,至少2周),然后转移到有益于芽形成的培养基中。定期转移培养物,直到已出现充分的芽形成时为止。一旦形成芽,就将其转移到有益于根形成的培养基中。一旦形成足够的根,就可将植物转移到土壤中,以便进一步生长和成熟。
为了确认再生植物中存在感兴趣的核酸分子(例如,编码一种或多种irna分子的dna,所述irna分子抑制鞘翅目和/或半翅目害虫中的靶标基因表达),可执行多种测定。此类测定包括例如:分子生物学测定,诸如southern印迹和northern印迹、pcr和核酸测序;生物化学测定,诸如检测是否存在蛋白质产物,例如通过免疫学手段(elisa和/或western印迹)或借助酶功能;植物部分测定,诸如叶或根测定;以及对再生全植物的表型的分析。
可例如通过使用例如对感兴趣的核酸分子有特异性的寡核苷酸引物进行pcr扩增来分析整合事件。pcr基因分型应当理解为包括但不限于:来源于经分离的宿主植物愈伤组织的gdna的聚合酶链式反应(pcr)扩增,所述愈伤组织预测含有整合到基因组中的感兴趣核酸分子,接着是pcr扩增产物的标准克隆和序列分析。pcr基因分型的方法已得到充分描述(例如rios等人,(2002)plantj.32:243-53),并且可应用于来源于任何植物物种(例如玉蜀黍或大豆)或组织类型的gdna,包括来源于细胞培养物的gdna。
使用土壤杆菌依赖性转化方法形成的转基因植物通常含有插入一条染色体中的单个重组dna。这单个重组dna的多核苷酸被称为“转基因事件”或“整合事件”。此类转基因植物对于插入的外源性多核苷酸而言是杂合的。在一些实施方案中,通过将含有单个外源性基因的独立的分离转基因植物与自身(例如t0植物)有性交配(自交)以产生t1种子,可获得相对于转基因为纯合的转基因植物。所产生的t1种子的四分之一相对于所述转基因会是纯合的。萌发t1种子产生的植物可用于测试杂合性,所述测试通常使用snp测定或热扩增测定,使得允许区分杂合子和纯合子(即接合性测定)。
在特定的实施方案中,在植物细胞中产生具有昆虫(例如鞘翅目和/或半翅目)害虫抑制效果的至少2、3、4、5、6、7、8、9或10种、或者更多种不同的irna分子。可以从导入不同转化事件中的多种核酸、或从导入单个转化事件中的单个核酸表达irna分子(例如dsrna分子)。在一些实施方案中,在单个启动子的控制下表达多个irna分子。在其他实施方案中,在多个启动子的控制下表达多个irna分子。可以表达包含多个多核苷酸的单一irna分子,所述多核苷酸各自与在相同的昆虫害虫物种的不同群体中或在不同的昆虫害虫物种中的一个或多个昆虫害虫内的不同基因座(例如,由seqidno:1、3、76和78限定的基因座)同源。
除了用重组核酸分子直接转化植物之外,可通过使具有至少一个转基因事件的第一植物与缺乏这种事件的第二植物杂交来制备转基因植物。例如,可将包含编码irna分子的多核苷酸的重组核酸分子导入易于转化的第一植物品系中以产生转基因植物,该转基因植物可与第二植物品系杂交,以使编码irna分子的多核苷酸渗入到第二植物品系中。
在一些方面,包括由来源于经转化植物细胞的转基因植物产生的种子和商品产品,其中所述种子或商品产品包含可检出量的本发明的核酸。在一些实施方案中,可例如通过获得转基因植物并由其制备食物或饲料来生产此类商品产品。包含本发明的多核苷酸中的一者或多者的商品产品包括(例如但不限于):植物的粗粉、油类、碾碎的或完整的籽粒或种子,以及包含含有本发明的核酸中的一者或多者的重组植物或种子的任何粗粉、油或者碾碎的或完整的籽粒的任何食物产品。在一种或多种商品或商品产品中检出本发明的多核苷酸中的一者或多者,实际上证明了该商品或商品产品是由出于控制昆虫(例如鞘翅目和/或半翅目)害虫的目的被设计为表达本发明的irna分子中的一者或多者的转基因植物产生的。
在一些实施方案中,包含本发明的核酸分子的转基因植物或种子也可在其基因组中包含至少一个其他的转基因事件,包括但不限于:自其转录靶向鞘翅目或半翅目害虫中与由下列各项限定的基因座不同的基因座的irna分子的转基因事件:seqidno:1、seqidno:3、seqidno:76和seqidno:78,所述不同的基因座诸如选自下列的一种或多种基因座:caf1-180(美国专利申请公布号2012/0174258)、vatpasec(美国专利申请公布号2012/0174259)、rho1(美国专利申请公布号2012/0174260)、vatpaseh(美国专利申请公布号2012/0198586)、ppi-87b(美国专利申请公布号2013/0091600)、rpa70(美国专利申请公布号2013/0091601)、rps6(美国专利申请公布号2013/0097730)、rop(美国专利申请号14/577,811)、rnapii(美国专利申请号14/577,854)、dre4(美国专利申请号14/705,807)、ncm(美国专利申请号62/095487)、copiα(美国专利申请号62/063,199)、copiβ(美国专利申请号62/063,203)、copiγ(美国专利申请号62/063,192)和copiδ(美国专利申请号62/063,216)、rna聚合酶i1(美国专利申请号62/133214)和rna聚合酶ii215(美国专利申请号62/133202);自其转录靶向与鞘翅目和/或半翅目害虫不同的生物体(例如植物寄生性线虫)中的基因的irna分子的转基因事件;编码杀虫蛋白(例如苏云金芽孢杆菌杀虫蛋白和pip-1多肽)的基因;除草剂耐受性基因(例如,提供对草甘膦的耐受性的基因);以及促成转基因植物中期望的表型(诸如产量增加、脂肪酸代谢改变或细胞质雄性不育性恢复)的基因。在特定的实施方案中,可以在植物中将编码本发明irna分子的多核苷酸与其他昆虫控制性状和疾病控制性状组合,以实现增强对植物疾病和昆虫损害的控制的期望性状。将采用独特作用模式的昆虫控制性状组合由于例如对所述一种或多种性状的抗性会在田间形成的概率降低,可以提供具有优于含单一控制性状的植物的出色耐久性的受保护转基因植物。
v.昆虫害虫中的靶标基因阻抑
a.概述
在本发明的一些实施方案中,可向昆虫(例如鞘翅目和/或半翅目)害虫提供至少一种可用于控制昆虫害虫的核酸分子,其中所述核酸分子在所述害虫中引起rnai介导的基因沉默。在特定的实施方案中,可向鞘翅目和/或半翅目害虫提供irna分子(例如dsrna、sirna、mirna、shrna和hprna)。在一些实施方案中,可通过使可用于控制昆虫害虫的核酸分子与害虫接触,来向所述害虫提供所述核酸分子。在这些和另外的实施方案中,可以在昆虫害虫的进食基质(例如营养组合物)中提供可用于控制所述害虫的核酸分子。在这些和另外的实施方案中,可通过摄入被昆虫害虫摄入的包含可用于控制所述害虫的核酸分子的植物材料,来提供所述核酸分子。在某些实施方案中,核酸分子通过表达导入植物材料中的重组核酸而存在于所述植物材料中,所述表达例如通过用包含重组核酸的载体转化植物细胞,然后从经转化植物细胞再生植物材料或全植物而进行。
在一些实施方案中,害虫通过接触局部用组合物(例如,通过喷洒施用的组合物)或rnai诱饵,而与在该害虫中引起rnai介导的基因沉默的核酸分子接触。将dsrna与食物或引诱剂或这二者混合时,形成rnai诱饵。害虫吃下诱饵时,还会吃掉dsrna。诱饵可采取颗粒、凝胶、可流动粉末、液体或固体的形式。在特定的实施方案中,可将rpii33掺入诱饵制剂中,诸如美国专利号8,530,440中所述,该专利据此以引用方式并入本文。一般说来,在使用诱饵的情况下,把诱饵放置在昆虫害虫的环境中或周围,这样,例如wcr就可接触到诱饵并且/或者被诱饵吸引。
b.rnai介导的靶标基因阻抑
在某些实施方案中,本发明提供了irna分子(例如,dsrna、sirna、mirna、shrna和hprna),可设计此类分子使之靶向昆虫害虫(例如鞘翅目(例如wcr、scr和ncr)或半翅目(例如bsb)害虫)的转录组中的必需天然多核苷酸(例如必需基因),例如通过设计有至少一条链包含与靶标多核苷酸特异性互补的多核苷酸的irna分子。如此设计的irna分子的序列可以与靶标多核苷酸的序列相同,或者可以掺入不阻止irna分子与其靶标多核苷酸之间的特异性杂交的错配。
本发明的irna分子可以在用于昆虫(例如鞘翅目和/或半翅目)害虫中的基因阻抑的方法中使用,从而降低由害虫对植物(例如,包含irna分子的受保护的经转化植物)造成的损害的水平或发生率。如本文所用,术语“基因阻抑”是指用于降低由于基因转录成mrna及随后mrna翻译而产生的蛋白质的水平的任一种熟知方法,包括降低由基因或编码多核苷酸表达的蛋白质,包括转录后抑制表达和转录阻抑。转录后抑制通过从被靶向以受阻抑的基因转录的mrna的全部或部分与用于阻抑的相应irna分子之间的特异性同源性来介导。此外,转录后抑制是指细胞中用于被核糖体结合的可用mrna的量出现实质性及可测量的下降。
在其中irna分子为dsrna分子的特定实施方案中,酶dicer可将dsrna分子切割成短sirna分子(长度大约为20个核苷酸)。借助dicer对dsrna分子的活性而生成的双链sirna分子可分成两个单链sirna:“过客链”和“引导链”。过客链可降解,引导链则可掺入risc中。转录后抑制通过引导链与mrna分子的特异性互补多核苷酸的特异性杂交,随后通过酶argonaute(risc复合物的催化组分)切割而发生。
在本发明的一些实施方案中,可使用任何形式的irna分子。本领域的技术人员会理解,在制备过程中以及在对细胞提供irna分子的步骤过程中,dsrna分子通常比单链rna分子更稳定,并且在细胞中通常也是更稳定的。因此,例如,虽然在一些实施方案中sirna分子和mirna分子可能是同等有效的,但dsrna分子可能由于其稳定性而被选用。
在某些实施方案中,提供了包含多核苷酸的核酸分子,该多核苷酸可在体外表达以产生irna分子,该irna分子与昆虫(例如鞘翅目和/或半翅目)害虫基因组内的多核苷酸所编码的核酸分子基本上同源。在某些实施方案中,体外转录的irna分子可以是包含茎环结构的稳定化dsrna分子。在昆虫害虫接触体外转录的irna分子之后,可发生对该害虫中的靶标基因(例如必需基因)的转录后抑制。
在本发明的一些实施方案中,在用于转录后抑制昆虫(例如鞘翅目和/或半翅目)害虫中的靶标基因的方法中使用包含多核苷酸的至少15个毗连核苷酸(例如,至少19个毗连核苷酸)的核酸分子的表达,其中所述多核苷酸选自:seqidno:1;seqidno:1的互补序列;seqidno:3;seqidno:3的互补序列;seqidno:5;seqidno:5的互补序列;seqidno:6;seqidno:6的互补序列;seqidno:7;seqidno:7的互补序列;seqidno:8;seqidno:8的互补序列;seqidno:1或seqidno:3的至少15个毗连核苷酸的片段;seqidno:1或seqidno:3的至少15个毗连核苷酸的片段的互补序列;叶甲属生物体的天然编码多核苷酸,其包含seqidno:5-8中的任一者;叶甲属生物体的天然编码多核苷酸的互补序列,所述天然编码多核苷酸包含seqidno:5-8中的任一者;叶甲属生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段,所述天然编码多核苷酸包含seqidno:5-8中的任一者;叶甲属生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段的互补序列,所述天然编码多核苷酸包含seqidno:5-8中的任一者;seqidno:76;seqidno:76的互补序列;seqidno:78;seqidno:78的互补序列;seqidno:76或seqidno:78的至少15个毗连核苷酸的片段;seqidno:76或seqidno:78的至少15个毗连核苷酸的片段的互补序列;半翅目生物体的天然编码多核苷酸,其包含seqidno:80-82中的任一者;半翅目生物体的天然编码多核苷酸的互补序列,所述天然编码多核苷酸包含seqidno:80-82中的任一者;半翅目生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段,所述天然编码多核苷酸包含seqidno:80-82中的任一者;以及半翅目生物体的天然编码多核苷酸的至少15个毗连核苷酸的片段的互补序列,所述天然编码多核苷酸包含seqidno:80-82中的任一者。在某些实施方案中,可以使用与前述中任一者至少约80%(例如,79%、约80%、约81%、约82%、约83%、约84%、约85%、约86%、约87%、约88%、约89%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%、约100%和100%)相同的核酸分子的表达。在这些和另外的实施方案中,可以表达与存在于昆虫(例如鞘翅目和/或半翅目)害虫的至少一个细胞中的rna分子特异性杂交的核酸分子。
本文的一些实施方案的重要特征在于,rnai转录后抑制系统能够容忍靶标基因中预期由于遗传突变、株系多态性或进化趋异而可能发生的序列变异。导入的核酸分子可以不必与靶标基因的初级转录产物或完全加工的mrna绝对同源,只要导入的核酸分子与靶标基因的初级转录产物或完全加工的mrna可特异性杂交即可。另外,相对于靶标基因的初级转录产物或完全加工的mrna,导入的核酸分子可以不必是全长的。
使用本发明的irna技术抑制靶标基因是序列特异性的;也就是说,靶向与一种或多种irna分子基本上同源的多核苷酸进行遗传抑制。在一些实施方案中,可以使用包含核苷酸序列与靶标基因的一部分的核苷酸序列相同的多核苷酸的rna分子进行抑制。在这些和另外的实施方案中,可以使用包含相对于靶标多核苷酸具有一个或多个插入、缺失和/或点突变的多核苷酸的rna分子。在特定的实施方案中,irna分子和靶标基因的一部分可共享例如至少从约80%、至少从约81%、至少从约82%、至少从约83%、至少从约84%、至少从约85%、至少从约86%、至少从约87%、至少从约88%、至少从约89%、至少从约90%、至少从约91%、至少从约92%、至少从约93%、至少从约94%、至少从约95%、至少从约96%、至少从约97%、至少从约98%、至少从约99%、至少从约100%以及100%的序列同一性。作为替代,dsrna分子的双链体区可与靶标基因转录物的一部分可特异性杂交。在可特异性杂交的分子中,表现出较大同源性的小于全长的多核苷酸补偿较长的、同源性较小的多核苷酸。dsrna分子的双链体区中与靶标基因转录物的一部分相同的多核苷酸的长度可以是至少约25、50、100、200、300、400、500个或至少约1000个碱基。在一些实施方案中,可以使用大于20个至100个核苷酸的多核苷酸。在特定的实施方案中,可以使用大于约200个至300个核苷酸的多核苷酸。在特定的实施方案中,根据靶标基因的大小,可以使用大于约500个至1000个核苷酸的多核苷酸。
在某些实施方案中,可以将害虫(例如鞘翅目或半翅目)中靶标基因的表达在该害虫的细胞内抑制至少10%、至少33%、至少50%或至少80%,使得发生显著抑制。显著抑制是指高于阈值的抑制,该抑制造成可检出的表型(例如,生长停止、进食停止、发育停止、诱导性死亡等),或与正在抑制的靶标基因对应的rna和/或基因产物出现可检出的减少。虽然在本发明的某些实施方案中,在所述害虫的基本上所有细胞中均发生抑制,但在其他实施方案中,仅在表达靶标基因的细胞子集中发生抑制。
在一些实施方案中,细胞中的转录阻抑由表现出与启动子dna或其互补序列的实质性序列同一性的dsrna分子的存在所介导,从而实现所谓的“启动子反式阻抑”。基因阻抑可以在可摄入或接触此类dsrna分子的昆虫害虫中(例如通过摄入或接触含有所述dsrna分子的植物材料)针对靶标基因起效。在启动子反式阻抑中使用的dsrna分子可以特异性地设计为抑制或阻抑昆虫害虫细胞中的一种或多种同源多核苷酸或互补多核苷酸的表达。美国专利号5,107,065、5,759,829、5,283,184和5,231,020中公开了通过反义或有义取向的rna进行转录后基因阻抑以调节植物细胞中的基因表达。
c.提供给昆虫害虫的irna分子的表达
可按许多体外形式或活体内形式中的任一种表达用于在昆虫(例如鞘翅目和/或半翅目)害虫中进行rnai介导的基因抑制的irna分子。然后可向昆虫害虫提供irna分子,例如通过使irna分子与害虫接触,或通过引起害虫摄入或以其他方式内化irna分子。一些实施方案包括鞘翅目和/或半翅目害虫的经转化宿主植物、经转化植物细胞和经转化植物的后代。经转化植物细胞和经转化植物可工程化改造为例如在异源性启动子的控制下表达所述irna分子中的一者或多者,以提供害虫保护效果。因此,当昆虫害虫在进食期间食用转基因植物或植物细胞时,该害虫可摄入转基因植物或细胞中表达的irna分子。也可将本发明的多核苷酸导入多种多样的原核微生物宿主和真核微生物宿主中以产生irna分子。术语“微生物”包括原核物种和真核物种,诸如细菌和真菌。
调控基因表达可包括对这种表达的部分或完全阻抑。在另一个实施方案中,用于阻抑昆虫(例如鞘翅目和/或半翅目)害虫中的基因表达的方法包括在害虫宿主的组织中提供基因阻抑量的由如本文所述的多核苷酸在转录后形成的至少一种dsrna分子,其至少一个区段与昆虫害虫细胞内的mrna互补。昆虫害虫所摄入的dsrna分子,包括其经修饰形式诸如sirna、mirna、shrna或hprna分子,可以与从例如包含选自seqidno:1、3、5-8、76、78和80-82的多核苷酸的rpii33dna分子转录的rna分子至少约80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或约100%相同。因此,提供了用于制备dsrna分子的经分离和基本上纯化的核酸分子,包括但不限于非天然存在的多核苷酸和重组dna构建体,其在导入昆虫害虫时阻抑或抑制其中的内源性编码多核苷酸或靶标编码多核苷酸的表达。
特定的实施方案提供了用于递送irna分子以便转录后抑制昆虫(例如鞘翅目和/或半翅目)植物害虫中的一种或多种靶标基因并且控制所述植物害虫的群体的递送系统。在一些实施方案中,所述递送系统包括对宿主转基因植物细胞或包含在宿主细胞中转录的rna分子的宿主细胞内容物的摄入。在这些和另外的实施方案中,创建转基因植物细胞或转基因植物,其含有用于提供本发明的稳定化dsrna分子的重组dna构建体。包含编码特定irna分子的核酸的转基因植物细胞和转基因植物可通过下述方式产生:采用重组dna技术(这些基础技术是本领域熟知的)构建包含编码本发明的irna分子(例如,稳定化的dsrna分子)的多核苷酸的植物转化载体,以此转化植物细胞或植物,并以此生成含有转录的irna分子的转基因植物细胞或转基因植物。
为了赋予转基因植物对昆虫(例如鞘翅目和/或半翅目)害虫的防护性,可以例如将重组dna分子转录成irna分子,诸如dsrna分子、sirna分子、mirna分子、shrna分子或hprna分子。在一些实施方案中,从重组dna分子转录的rna分子可以在重组植物的组织或流体内形成dsrna分子。这样的dsrna分子可以包含在多核苷酸的一部分中,所述多核苷酸与从可侵染宿主植物的类型的昆虫害虫内的dna转录的相应多核苷酸相同。所述害虫内靶标基因的表达受到dsrna分子阻抑,并且对所述害虫中靶标基因的表达的阻抑保护转基因植物免受害虫损害。已显示dsrna分子的调控作用适用于害虫中表达的多种基因,包括例如负责细胞代谢或细胞转化的内源性基因,包括管家基因;转录因子;蜕皮相关基因;以及编码涉及细胞代谢或正常生长和发育的多肽的其他基因。
为了从活体内转基因或表达构建体转录,可以在一些实施方案中使用调节区(例如,启动子、增强子、沉默子和多聚腺苷酸化信号)来转录一条或多条rna链。因此,在一些实施方案中,如前文示出的,在产生irna分子中使用的多核苷酸可以与在植物宿主细胞中具有功能的一种或多种启动子元件可操作地连接。启动子可以是通常驻留于宿主基因组中的内源性启动子。在可操作地连接的启动子元件控制下的本发明的多核苷酸可进一步侧接有利地影响其转录和/或所得转录物的稳定性的额外元件。此类元件可位于可操作地连接的启动子上游、表达构建体3’端下游,并且可既存在于所述启动子上游、又存在于表达构建体3’端下游。
一些实施方案提供了用于减轻由以植物为食的昆虫(例如鞘翅目和/或半翅目)害虫引起的对宿主植物(例如玉米植物)的损害的方法,其中所述方法包括在宿主植物中提供表达本发明的至少一种核酸分子的经转化植物细胞,其中所述核酸分子在被所述害虫摄取后发挥作用以抑制所述害虫内靶标多核苷酸的表达,该表达抑制造成所述害虫死亡和/或生长减缓,从而减轻由所述害虫引起的对宿主植物的损害。在一些实施方案中,所述核酸分子包括dsrna分子。在这些和另外的实施方案中,所述核酸分子包括dsrna分子,所述dsrna分子各自包含超过一种与鞘翅目和/或半翅目害虫细胞中表达的核酸分子可特异性杂交的多核苷酸。在一些实施方案中,所述核酸分子由一种多核苷酸组成,所述多核苷酸与昆虫害虫细胞中表达的核酸分子可特异性杂交。
在一些实施方案中,提供了用于提高玉米作物产量的方法,其中所述方法包括将本发明的至少一种核酸分子导入玉米植物中;栽培该玉米植物,以允许表达包含所述核酸的irna分子,其中包含所述核酸的irna分子的表达抑制昆虫(例如鞘翅目和/或半翅目)害虫损害和/或生长,从而降低或消除由于害虫侵染所致的产量损失。在一些实施方案中,所述irna分子为dsrna分子。在这些和另外的实施方案中,所述核酸分子包括dsrna分子,所述dsrna分子各自包含超过一种与昆虫害虫细胞中表达的核酸分子可特异性杂交的多核苷酸。在一些实施方案中,所述核酸分子包含与鞘翅目和/或半翅目害虫细胞中表达的核酸分子可特异性杂交的多核苷酸。
在某些实施方案中,提供了用于调控昆虫(例如鞘翅目和/或半翅目)害虫中靶标基因的表达的方法,所述方法包括:用包含编码本发明的至少一种irna分子的多核苷酸的载体转化植物细胞,其中所述多核苷酸与启动子和转录终止元件可操作地连接;在足以允许包含多个经转化植物细胞的植物细胞培养物发育的条件下培养经转化植物细胞;选择已将所述多核苷酸整合到其基因组中的经转化植物细胞;针对由整合的多核苷酸编码的irna分子的表达,筛选经转化植物细胞;选择表达irna分子的转基因植物细胞;然后用选择的转基因植物细胞饲喂所述昆虫害虫。还可从表达由整合的核酸分子编码的irna分子的经转化植物细胞再生植物。在一些实施方案中,所述irna分子为dsrna分子。在这些和另外的实施方案中,所述核酸分子包括dsrna分子,所述dsrna分子各自包含超过一种与昆虫害虫细胞中表达的核酸分子可特异性杂交的多核苷酸。在一些实施方案中,所述核酸分子包含与鞘翅目和/或半翅目害虫细胞中表达的核酸分子可特异性杂交的多核苷酸。
可将本发明的irna分子以来自掺入植物细胞基因组中的重组基因的表达产物、或者以掺入应用于种植前种子的包衣或种子处理中,而掺入植物物种(例如,玉米或大豆)的种子之内。包含重组基因的植物细胞被视为转基因事件。本发明的实施方案中还包括用于将irna分子递送到昆虫(例如鞘翅目和/或半翅目)害虫的递送系统。例如,可将本发明的irna分子直接导入害虫的细胞中。用于导入的方法可包括将irna与来自昆虫害虫宿主的植物组织直接混合,以及向宿主植物组织应用包含本发明的irna分子的组合物。例如,可将irna分子喷洒到植物表面上。作为替代,可通过微生物表达irna分子,然后可将微生物施用到植物表面上,或通过物理手段诸如注射导入根或茎中。如前文论述的,也可以对转基因植物进行遗传工程改造,使其以足以杀死已知要侵染植物的昆虫害虫的量表达至少一种irna分子。通过化学方法或酶促合成方法产生的irna分子也可按符合常见农业实践的方式配制,并且作为喷雾产品使用以控制昆虫害虫所致的植物损害。配制剂可包含有效叶覆盖所需的适当佐剂(例如粘着剂和湿润剂),以及保护irna分子(例如dsrna分子)免受紫外线损害的紫外线保护剂。此类添加剂常用于生物杀虫剂产业中,并且是本领域技术人员熟知的。此类应用可与其他喷雾杀虫剂应用(基于生物学或其他方面)组合,以增强植物对所述害虫的防护性。
本文引用的全部参考文献(包括出版物、专利和专利申请)据此以引用方式并入,并入程度与本公开的明确细节不冲突,并且以与下述相同的程度这样并入:如同单独地和具体地指明每篇参考文献均以引用方式并入且在本文中全文示出。提供本文论述的参考文献仅仅是为了参考其在本申请提交日之前的公开内容。本文中的任何内容都不应被理解为承认发明人由于在先发明而无权先于这样的公开内容。
提供以下实施例是为了例示某些特定的特征和/或方面。这些实施例不应被理解为将本公开限于所描述的特定特征或方面。
用于实施本发明的模式
i.数个实施方案的概述
转基因植物的发育变得越来越复杂,并且通常需要将多个转基因堆叠成单个基因座。参见xie等人,(2001)nat.biotechnol.19(7):677-9。由于每个转基因通常需要唯一的启动子用于表达,因此需要多个启动子来表达一个基因叠加内的不同转基因。除了增加基因叠加的大小外,这还通常导致同一启动子被重复使用,以获得不同转基因的相似水平的表达模式。该办法经常造成问题,因为由同一启动子驱动多个转基因的表达可导致基因沉默或hbgs。重复启动子中竞争性转录因子(tf)结合位点的过量可导致内源性tf的消耗并引起转录下调。转基因的沉默对所产生的转基因植物表达该转基因的性能而言是不期望的。在转基因内的重复序列经常导致基因座内同源重组,从而导致多核苷酸重排和不合需要的表型或农艺性能。
用于基础研究或生物技术应用的植物启动子一般是单向的,且仅调节在其3’端(下游)融合的一个基因。为了产生具有各种期望性状或特征的转基因植物,减少被部署来驱动编码期望性状和特征的转基因表达的启动子的数目将是有用的。尤其是在需要将多个转基因导入植物用于代谢工程和性状叠加,由此需要多个启动子来驱动多个转基因的表达的应用中。通过开发可驱动启动子侧翼的两个转基因表达的单一稻泛素-3合成双向启动子,可以减少开发转基因作物所需的启动子的总数,由此减少同一启动子的重复施用、减小转基因构建体的大小和/或降低hbgs的可能性。这样的启动子可以通过将已知的顺式元件引入新的或合成的dna段中来产生,或者通过“域交换”来产生,在“域交换”中,一个启动子的结构域被替换为来自其他异源启动子的功能等同的结构域。
本文的实施方案利用这样的过程,其中利用来自水稻(稻)泛素-3基因的单向启动子(例如rubi3)来设计合成的稻泛素-3双向启动子,使得一个启动子可以引导两个基因的表达,在该启动子的每个末端各一个。合成的稻泛素-3双向启动子可允许本领域的技术人员将转基因叠加到植物细胞和植物中,同时减少相同启动子的重复使用,并减小转基因构建体的大小。此外,用单一的合成稻泛素-3双向启动子来调节两个基因的表达还可提供在相同条件下共表达这两个基因的能力,诸如,例如当两个基因各自有助于宿主中的单一性状时可能有用。已在一些情况下报道了在植物中利用启动子的双向功能,包括玉米泛素1启动子(国际专利公布号wo2013101343a1)、camv35启动子(barfield和pua(1991)plantcellrep.10(6-7):308-14;xie等人(2001),同上)和mas启动子(velten等人(1984)emboj.3(12):2723-30;langridge等人(1989)proc.natl.acad.sci.usa86:3219-23)。
植物基因中基因表达的转录起始和调控受共同排列在启动子内的多种dna序列元件的指导。真核生物启动子由最小核心启动子元件(minp)和另外的上游调控序列(urs)组成。核心启动子元件是足以指导准确的转录启动的毗连dna序列的最小区段。植物中的核心启动子还包含与转录起始有关的规范区,诸如caat和tata框。tata框元件通常位于转录起始位点上游的约20至35个核苷酸。
minp的活化依赖于urs,各种蛋白质与之结合并随后与转录起始复合体相互作用。urs包含这样的dna序列,它们决定包含urs的启动子的时空表达模式。启动子的极性通常由minp的取向决定,而urs是双极的(即其功能独立于其取向)。
在一些实施方案的具体实例中,利用最初来源于玉米的经修饰的玉米泛素-1启动子(zmubi1)的最小核心启动子元件(minubi10p)来工程化在植物中起作用的合成稻泛素-3双向启动子,以提供相对于先前描述的双向启动子而言独特的表达控制特征。实施方案包括合成的稻泛素-3双向启动子,该启动子还包含衍生自天然玉米泛素-1启动子的最小核心启动子元件核苷酸序列(minpzmubi1)。
最初来源于玉米栽培种b73的zmubi1启动子包含在玉米基因组内位于转录起始位点5′的约899个碱基内以及转录起始位点3′的约1,093个碱基内的序列。christensen等人,(1992)plantmol.biol.18(4):675-89(描述玉米c.v.b73zmubi1基因)。在一些实例中使用的衍生自b73的经修饰的zmubi1启动子是约2kb的启动子,其含有:tata盒;两个重叠的热休克共有元件;与转录起始位点相邻的82或83个核苷酸(取决于参考链)的前导序列,在本文中称为zmubi1外显子;和1015至-1016个核苷酸的内含子。其他来源于玉米属物种和玉米基因型的玉米泛素启动子变体可在由tata元件和上游热休克共有元件组成的minp元件周围表现出高序列保守性。因此,使用zmubi1启动子的这一短的(~200nt)、高度保守的区域(例如seqidno:2)作为用于构建合成双向植物启动子的最小核心启动子元件,来例示本发明的实施方案。
最初来源于水稻的稻泛素-3启动子包含在水稻基因组内位于转录起始位点5′约1,990个喊基内的序列。esivamani和rqu(2006)expressionenhancementofaricepolyubiquitinpromoter.plantmolecularbiology60:225-239。在一些实例中使用的来源于水稻的经修饰的稻泛素3启动子是2kb启动子,其包含:tata盒、5'utr/内含子序列、和位于稻泛素-3编码序列的开始处的下游增强元件。其他来源于稻属物种和水稻基因型的稻泛素-3启动子变体可在这些启动子元件周围表现出高序列保守性。
ii.缩写
atubi10拟南芥泛素10
bca二辛可宁酸
camv花椰菜花叶病毒
csvmv木薯叶脉花叶病毒
ctp叶绿体转运肽
hbgs基于同源性的基因沉默
minubi1pzmubi1最小核心启动子
ola寡聚物连接扩增
pcr聚合酶链式反应
rca滚环扩增
rubi3稻泛素-3
rt-pcr逆转录酶pcr
snupe单核苷酸引物延伸
urs上游调控序列
zmubi1玉米泛素1
iii.术语
在整个申请中,使用了许多术语。为了提供对说明书和权利要求书的清楚且一致的理解,包括要给予此类术语的范围,提供了以下定义:
内含子:如本文所用,术语“内含子”是指包含在基因中的(或表达感兴趣的多核苷酸序列的)转录但不翻译的任何核酸序列。内含子包括在dna的表达序列内的未翻译核酸序列,以及在从其转录的rna分子中的对应序列。
经分离的:“经分离的”生物组分(诸如核酸或蛋白质)已从该组分所天然存在的生物体细胞中的其他生物组分(即其他染色体和染色体外的dna和rna,以及蛋白质)基本上分开、分开产生或纯化出来,同时实现组分中的化学或功能变化(例如,核酸可通过断开将该核酸连接到染色体中的其余dna的化学键而从染色体分离)。已经“经分离的”核酸分子和蛋白质包括通过标准纯化方法纯化的核酸分子和蛋白质。该术语还包括通过在宿主细胞内重组表达而制备的核酸和蛋白质,以及化学合成的核酸分子、蛋白质和肽。
基因表达:一个过程,通过该过程,核酸转录单元(包括例如基因组dna)的编码信息转化为细胞的操作部分、非操作部分或结构性部分,通常包括蛋白质的合成。基因表达可能受到外部信号的影响,例如细胞、组织或生物体暴露于增加或减少基因表达的药剂。基因表达也可以在从dna到rna再到蛋白质的途径中的任何位置受到调节。调节基因表达例如通过控制对转录、翻译、rna转运和加工、中间分子(诸如mrna)降解的作用,或通过在特异性蛋白分子已产生之后的活化、失活、区室化或降解,或这些的组合而发生。基因表达可通过本领域已知的任何方法,包括但不限于northern印迹法、rt-pcr、western印迹法,或者体外、原位或活体内蛋白质活性测定法,在rna水平或蛋白质水平测量。
基于同源性的基因沉默:如本文所用,“基于同源性的基因沉默”(hbgs)是既包括转录基因沉默也包括转录后基因沉默的遗传术语。非连锁的致沉默基因座对靶基因座的沉默作用可缘于转录抑制(转录基因沉默,tgs)或mrna降解(转录后基因沉默,ptgs),转录抑制或mrna降解分别是由对应于启动子或转录序列的双链rna(dsrna)的产生所导致。在每个过程中牵涉的细胞组分不同,表明dsrna诱导的tgs和ptgs很可能是一种古老的共同机制的多样化的结果。然而,难以实现tgs和ptgs的严格比较,因为这种比较一般依赖于对不同沉默基因座的分析。我们描述既触发tgs又触发ptgs的单个转基因基因座,这是由于对应于不同靶标基因的启动子和转录序列的dsrna的产生。mourrain等人(2007)planta225:365-79。可能的是,sirna是触发同源序列上的tgs和ptgs的实际分子:在此模型中,通过转基因序列的甲基化扩散到内源性启动子中,sirna可以顺式地或反式地触发同源序列的沉默和甲基化。id.
核酸分子:如本文所用,术语“核酸分子”(或“核酸”或“多核苷酸”)可以指核苷酸的聚合形式,可包括rna、cdna、基因组dna的有义链和反义链两者,以及上述各项的合成形式和混合聚合物。核苷酸可以指核糖核苷酸、脱氧核糖核苷酸,或这两种类型核苷酸中任一者的修饰形式。如本文所用的“核酸分子”与“核酸”和“多核苷酸”是同义词。除非另外指明,核酸分子的长度通常至少为10个碱基。该术语可以指不确定长度的rna或dna分子。该术语包括单链和双链形式的dna。核酸分子可包括天然存在的核苷酸和/或经修饰的核苷酸,它们通过天然存在的核苷酸连接和/或非天然存在的核苷酸连接而连接在一起。
如本领域技术人员易于理解的那样,核酸分子可被化学修饰或生物化学修饰,或者可含有非天然的或衍生化的核苷酸碱基。此类修饰包括例如标记物、甲基化、用类似物置换天然存在的核苷酸中的一者或多者、核苷酸间修饰(例如不带电连接:例如甲基膦酸酯、磷酸三酯、氨基磷酸酯、氨基甲酸酯等;带电连接:例如硫代磷酸酯、二硫代磷酸酯等;悬垂部分:例如肽类;嵌入剂:例如吖啶、补骨脂素等;螯合剂;烷化剂;以及经修饰的连接:例如α异头核酸等)。术语“核酸分子”还包括任何拓扑构象,包括单链的、双链的、部分双链体的、三链体的、发夹形的、圆形的和挂锁形的(padlocked)构象。
转录沿着dna链以5′至3′方式进行。这意味着,rna通过在生长链的3′末端依次添加5′-三磷酸核糖核苷酸(随着焦磷酸的必要去除)来合成。在线性或环状核酸分子中,如果独立元件(例如特定的核苷酸序列)在同一核酸中或者会在同一核酸中键合于另一元件的5′方向,则可称它们相对于该另一元件处于“上游”或“5′”。类似地,如果独立元件在同一核酸中或者会在同一核酸中键合于另一元件的3′方向,则可称它们相对于该另一元件处于“下游”或“3′”。
如本文所用,碱基“位置”是指给定的碱基或核苷酸残基在指定核酸内的位置。指定的核酸可通过与参照核酸比对(见下文)来限定。
杂交:寡核苷酸及其类似物通过互补碱基之间的氢键结合而杂交,氢键结合包括watson-crick、hoogsteen或反hoogsteen氢键结合。一般而言,核酸分子由含氮碱基组成,碱基为嘧啶(胞嘧啶(c)、尿嘧啶(u)和胸腺嘧啶(t))或嘌呤(腺嘌呤(a)和鸟嘌呤(g))。这些含氮碱基在嘧啶和嘌呤之间形成氢键,并且嘧啶对嘌呤的结合称为“碱基配对”。更具体地讲,a将与t或u氢键结合,而g将与c结合。“互补”是指在两条不同的核酸序列或同一核酸序列的两个不同区域之间发生的碱基配对。
“可特异性杂交”和“特异性互补”是表明足够程度的互补性的术语,该互补性使得在寡核苷酸与dna或rna靶标之间发生稳定且特异性的结合。寡核苷酸与其可特异性杂交的靶标序列不一定是100%互补的。当寡核苷酸与靶标dna或rna分子的结合干扰靶标dna或rna的正常功能,并且有足够程度的互补性来避免寡核苷酸在期望特异性结合的条件下(例如在体内测定法或系统的情况中在生理学条件下)与非靶标序列发生非特异性结合时,则寡核苷酸是可特异性杂交的。这样的结合成为特异性杂交。
导致特定严格性程度的杂交条件将根据选择的杂交方法的性质以及杂交核酸的组成和长度而变化。一般说来,杂交的温度和杂交缓冲液的离子强度(尤其是na+和/或mg2+浓度)将有助于杂交的严格性,但洗涤次数也影响严格性。有关获得特定严格性程度所需要的杂交条件的计算论述于sambrook等人(编辑),molecularcloning:alaboratorymanual,第2版,第1-3卷,coldspringharborlaboratorypress,coldspringharbor,newyork,1989,第9和11章。
如本文所用,“严格条件”涵盖仅在杂交分子与dna靶标之间存在小于50%的错配时才发生杂交的条件。“严格条件”包括另外的特定严格性水平。因此,如本文所用,“中等严格性”条件是序列错配超过50%的分子不会杂交的条件;“高严格性”条件是错配超过20%的序列不会杂交的条件;而“极高严格性”条件是错配超过10%的序列不会杂交的条件。
在特定的实施方案中,严格条件可包括在65℃下杂交,然后在65℃下用0.1xssc/0.1%sds洗涤40分钟。
以下是代表性的非限制性杂交条件:
极高严格性:在5xssc缓冲液中在65℃下杂交16小时;在2xssc缓冲液中在室温下洗涤两次,每次15分钟;以及在0.5xssc缓冲液中在65℃下洗涤两次,每次20分钟。
高严格性:在5x-6xssc缓冲液中在65-70℃下杂交16-20小时;在2xssc缓冲液中在室温下洗涤两次,每次5-20分钟;以及在1xssc缓冲液中在55-70℃下洗涤两次,每次30分钟。
中等严格性:在6xssc缓冲液中在室温至55℃下杂交16-20小时;在2x-3xssc缓冲液中在室温至55℃下至少洗涤两次,每次20-30分钟。
在特定的实施方案中,可特异性杂交的核酸分子在极高严格性杂交条件下可保持结合。在这些和另外的实施方案中,可特异性杂交的核酸分子在高严格性杂交条件下可保持结合。在这些和另外的实施方案中,可特异性杂交的核酸分子在中等严格性杂交条件下可保持结合。
寡核苷酸:寡核苷酸是短的核酸聚合物。寡核苷酸可通过切割较长的核酸区段、或通过使单独的核苷酸前体聚合而形成。自动合成仪允许合成长度多达几百个碱基对的寡核苷酸。由于寡核苷酸可与互补的核苷酸序列结合,所以可用作检测dna或rna的探针。由dna组成的寡核苷酸(寡脱氧核糖核苷酸)可在pcr(一种用于扩增小dna序列的技术)中使用。在pcr中,寡核苷酸通常被称为“引物”,其允许dna聚合酶延伸寡核苷酸并复制互补链。
序列同一性:如本文所用,术语“序列同一性”或“同一性”在两个核酸或多肽序列的语境下,可指在指定比较窗口上以最大对应性比对时这两个序列中相同的残基。
如本文所用,术语“序列同一性百分比”可以指通过在比较窗口上比较分子的两个最佳比对序列(例如核酸序列和氨基酸序列)确定的值,其中为了实现这两个序列的最佳比对,该比较窗口中的序列部分相比于参考序列(其不包含添加或缺失)可包含添加或缺失(即空位)。通过确定在两个序列中出现相同的核苷酸或氨基酸残基的位置的数目而产生匹配位置数,用该匹配位置数除以比较窗口中位置的总数,将结果乘以100而产生序列同一性的百分比,从而计算出该百分比。
用于比较的序列比对方法是本领域熟知的。各种程序和比对算法描述于例如以下文献中:smith和waterman(1981)adv.appl.math.2:482;needleman和wunsch(1970)j.mol.biol.48:443;pearson和lipman(1988)proc.natl.acad.sci.u.s.a.85:2444;higgins和sharp(1988)gene73:237-44;higgins和sharp(1989)cabios5:151-3;corpet等人,(1988)nucleicacidsres.16:10881-90;huang等人,(1992)comp.appl.biosci.8:155-65;pearson等人,(1994)methodsmol.biol.24:307-31;tatiana等人,(1999)femsmicrobiol.lett.174:247-50。序列比对方法和同源性计算的详细考虑事项可见于例如altschul等人,(1990)j.mol.biol.215:403-10。
美国国家生物技术信息中心(ncbi)基本局部比对搜索工具(blast;altschul等人(1990))可从几个源头(包括美国国家生物技术信息中心(bethesda,md))获得并且可在互联网上获得,以便结合几种序列分析程序使用。怎样使用该程序确定序列同一性的描述可在互联网上blast的“帮助”部分获得。为了比较核酸序列,可以采用利用默认参数的blast(blastn)程序的“blast2序列”功能。在用这种方法评估时,与参考序列具有甚至更大的相似性的核酸序列将显示同一性百分比增加。
可操作地连接:当第一核酸序列与第二核酸序列处于功能性关系中时,第一核酸序列与第二核酸序列可操作地连接。例如,当启动子影响编码序列的转录或表达时,启动子与编码序列可操作地连接。当以重组方式产生时,可操作地连接的核酸序列一般是毗连的,并且在必要时将两个蛋白质编码区连接在同一个阅读框内。然而,可操作地连接的元件不一定是毗连的。
启动子:dna的通常位于下游的(朝向基因的5′区)且需要转录的区域。启动子可允许其控制的基因得到正确活化或受到阻遏。启动子可包含由转录因子识别的特定序列。这些因子可结合到启动子dna序列并导致rna聚合酶的募集,rna聚合酶是一种从基因的编码区合成rna的酶。
转化:在核酸分子通过将核酸分子并入细胞基因组中、或通过附加型复制而由细胞稳定复制的情况下,细胞被转导到该细胞中的核酸分子“转化”。如本文所用,术语“转化”涵盖可将核酸分子导入这种细胞中的所有技术。实例包括但不限于:用病毒载体转染;用质粒载体转化;电穿孔(fromm等人,(1986)nature319:791-3);脂质体转染(felgner等人,(1987)proc.natl.acad.sci.usa84:7413-7);显微注射(mueller等人,(1978)cell15:579-85);土壤杆菌(agrobacterium)介导的转移(fraley等人,(1983)proc.natl.acad.sci.usa80:4803-7);直接dna摄取;晶须介导的转化(whiskers-mediatedtransformation);以及微粒轰击(klein等人,(1987)nature327:70)。
转基因:外源性核酸序列。在一个实例中,转基因是基因序列(例如,除草剂耐受性基因)、编码工业或药学上有用的化合物的基因,或者编码期望的农业性状的基因。在又一个实例中,转基因是反义核酸序列,其中反义核酸序列的表达抑制靶标核酸序列的表达。转基因可控制可操作地连接到该靶基因的调控序列(例如启动子)。在一些实施方案中,感兴趣的核酸序列是转基因。然而,在其他实施方案中,感兴趣的多核苷酸序列是内源性核酸序列,其中期望内源性核酸序列的额外基因组拷贝;或相对于宿主生物体中的目标核酸分子的序列处于反义取向的多核苷酸序列。
转基因事件:通过用异源dna(即包括感兴趣转基因的核酸构建体)转化植物细胞,再生由于该转基因插入到植物的基因组中而产生的植物的群体,并选择以特定的基因组位置中的插入为特征的特定植物,而产生转基因“事件”。术语“事件”是指原始转化体以及该转化体的包含异源dna的后代。术语“事件”还指该转化体与包含基因组/转基因dna的另一品种之间的有性远交产生的后代。即使在与轮回亲本反复回交之后,来自转化的亲本的插入的转基因dna和侧翼基因组dna(基因组/转基因dna)仍存在于杂交后代中相同的染色体位置。术语“事件”还指来自起始转化体及其包含插入的dna和直接邻近于插入的dna的侧翼基因组序列的后代的dna,预期该dna会被转移至这样的后代:所述后代作为一个含有插入的dna的亲本品系(例如起始转化体和其自交所得的后代)和不一个含有该插入的dna的亲本品系有性杂交的结果而接受了含有感兴趣转基因的插入的dna。
载体:一种导入细胞中从而产生经转化的细胞的核酸分子。载体可包含允许其在宿主细胞中复制的核酸序列,诸如复制起点。实例包括但不限于:质粒、粘粒、噬菌体,或携带外源性dna进入细胞的病毒。载体还可包括一种或多种基因、反义分子和/或可选择标记基因和本领域已知的其他遗传元件。载体可以转导、转化或感染细胞,从而引起细胞表达核酸分子和/或由该载体编码的蛋白质。载体可任选地包括辅助核酸分子实现进入细胞的物质(例如脂质体、蛋白质编码等)。
除非另外特别地解释,本文使用的全部技术术语和科学术语具有与本公开所属领域的普通技术人员通常所理解的相同的含义。分子生物学中常见术语的定义可在例如以下出版物中找到:lewin,genesv,oxforduniversitypress,1994(isbn0-19-854287-9);kendrew等人(编辑),theencyclopediaofmolecularbiology,blackwellscienceltd.,1994(isbn0-632-02182-9);以及meyers(编辑),molecularbiologyandbiotechnology:acomprehensivedeskreference,vchpublishers,inc.,1995(isbn1-56081-569-8)。
如本文所用,除非上下文清晰明确地相反指出,否则词语“一个/一种”和“该/所述”包括复数指代物。
iv.合成的双向启动子rubi3,以及包含该启动子的核酸
本公开提供包含可充当双向启动子的合成核苷酸序列的核酸分子。在一些实施方案中,合成双向启动子可以可操作地连接到感兴趣的一个或两个多核苷酸序列。例如,合成的稻泛素3双向启动子可以可操作地连接到编码基因的感兴趣的一个或两个多核苷酸序列。(例如,两个基因,在启动子的每个末端各一个),从而调节至少一个(例如一个或两个)感兴趣核苷酸序列的转录。在一些实施方案中,通过在合成的稻泛素-3双向启动子中并入来自稻泛素-3启动子的urs,可以针对可操作地连接到合成的稻泛素-3双向启动子的感兴趣多核苷酸序列实现特定的表达和调节模式(例如,诸如由稻泛素-3启动子控制下的基因表现的)。
通过将来自单向玉米泛素-1基因(zmubi1)启动子的最小核心启动子元件置于与天然启动子不同的分子环境中以工程化合成的双向启动子,来例示本发明的一些实施方案。该最小核心启动子元件在本文中称为“minubi1p”,其长度为约200nt。对来自多个玉米属物种和玉米基因型的minubi1p元件的测序和分析已揭露:功能性minubi1p元件是高度保守的,使得如果minubi1p与seqidno:2的minubi1p元件共享例如至少约75%、至少约80%、至少约85%、至少约90%、至少约91%、至少约92%、至少约93%、至少约94%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%、和/或至少约100%的序列同一性,则其可以保留作为转录引发物的功能。在本发明的一些实施方案中可能有用的minubi1p元件的特征可以包括例如但不限于上述核苷酸序列的高度保守、存在至少一个tata盒和/或存在至少一个(例如两个)热休克共有元件。在特定的minubi1p元件中,多于一个热休克共有元件可在minubi1p序列内重叠。
在实施方案中,将minubi1p元件并入与天然启动子(即,稻泛素3)不同的分子环境中以工程化合成双向启动子的过程可以包括将minubi1p元件并入稻泛素-3启动子核酸中,同时相对于稻泛素3启动子的其余序列逆转minubi1p元件的取向。因此,合成的稻泛素-3双向启动子可以包含位于稻泛素-3核苷酸序列的3′且与之朝向相反的minubi1p最小核心启动子元件,使得其可以可操作地连接到位于稻泛素-3核苷酸序列3′的感兴趣核苷酸序列。例如,minubi1p元件可以以反向取向并入稻泛素-3启动子的3′末端。
合成的双向稻泛素-3启动子除了minubi1p元件以及天然稻泛素3启动子的元件外,还可以包含一个或多个另外的序列元件。在一些实施方案中,合成的双向稻泛素3启动子可以包含启动子urs、外显子(例如前导或信号肽)、内含子、间隔序列和/或任何前述各者中一个或多个的组合。例如但不限于,合成的双向稻泛素3启动子可以包含来自稻泛素3或zmubi1动子的urs序列、来自稻泛素3或zmubi1基因的内含子、编码来自稻泛素3或zmubi1基因的前导肽的外显子、来自稻泛素3或zmubi1基因的内含子以及这些的组合。
合成的双向稻泛素-3启动子除了minubi1p元件和天然启动子稻泛素3的元件(包括minubi1p)之外还可以包含一个或多个另外的序列元件。在一些实施方案中,合成的双向稻泛素3启动子可以包含启动子urs、外显子(例如前导或信号肽)、内含子、间隔序列和/或任何前述各者中一个或多个的组合。例如但不限于,合成的双向稻泛素3启动子可以包含来自玉米泛素1启动子的urs序列、adh基因的内含子、编码来自玉米泛素基因的前导肽的外显子、来自玉米泛素基因的内含子以及这些的组合。
在包含启动子urs的启动子的一些实施方案中,可选择urs使之赋予合成启动子特定的调节特性。已知的启动子在其施加于可操作地连接的基因上的控制类型方面广泛不同(例如环境应答、发育提示和空间信息),且并入异源启动子中的urs通常维持urs就其天然启动子和可操作地连接的基因而言所展现的控制类型。langridge等人(1989),同上。已表征且根据一些实施方案可以含有包含在合成双向稻泛素3启动子内的urs的真核生物启动子的实例包括例如但不限于:在美国专利号6,437,217(玉米rs81启动子)、5,641,876(稻肌动蛋白启动子)、6,426,446(玉米rs324启动子)、6,429,362(玉米pr-1启动子)、6,232,526(玉米a3启动子)、6,177,611(组成型玉米启动子)、6,433,252(玉米l3油质蛋白启动子)、6,429,357(稻肌动蛋白2启动子和稻肌动蛋白2内含子)、5,837,848(根特异性启动子)、6,294,714(光可诱导的启动子)、6,140,078(盐可诱导的启动子)、6,252,138(病原体可诱导的启动子)、6,175,060(磷缺乏可诱导的启动子)、6,388,170(双向启动子)、6,635,806(醇溶蛋白(γ-coixin)启动子)和美国专利申请序列号09/757,089(玉米叶绿体醛缩酶启动子)中描述的那些启动子。
另外的示例性原核生物启动子包括胭脂碱(nopaline)合酶(nos)启动子(ebert等人(1987)proc.natl.acad.sci.usa84(16):5745-9)、章鱼碱(octopine)合酶(ocs)启动子(其携带在根癌土壤杆菌的肿瘤诱导质粒上)、花椰菜花叶病毒启动子如花椰菜镶嵌病毒(camv)19s启动子(lawton等人(1987)plantmol.biol.9:315-24)、camv35s启动子(odell等人(1985)nature313:810-2)、玄参镶嵌病毒35s启动子(walker等人(1987)proc.natl.acad.sci.usa84(19):6624-8)、蔗糖合酶启动子(yang和russell(1990)proc.natl.acad.sci.usa87:4144-8)、r基因复合物启动子(chandler等人(1989)plantcell1:1175-83)、camv35s(美国专利号5,322,938、5,352,605、5,359,142和5,530,196)、fmv35s(美国专利号6,051,753和5,378,619)、pc1sv启动子(美国专利号5,850,019)、scp1启动子(美国专利号6,677,503)和根癌土壤杆菌nos启动子(genbank登录号v00087;depicker等人(1982)j.mol.appl.genet.1:561-73;bevan等人(1983)nature304:184-7)等。
在一些实施方案中,合成的稻泛素3双向启动子还可以包含外显子。例如,可能期望将可操作地连接到启动子的感兴趣多核苷酸序列所编码的多肽靶向或运输到特定的亚细胞位置和/或区室。在这些和其他实施方案中,编码序列(外显子)可以并入剩余的合成稻泛素3双向启动子序列与编码多肽的核苷酸序列之间的核酸分子中。可以根据熟练从业人员的判断来安排这些元件,以使得合成的稻泛素3双向启动子促进多肽(或可操作地连接到启动子的两个多肽编码序列中的一个或两个)的表达,所述多肽包含与多肽的其余部分处于功能关系的由所并入的编码序列编码的肽。在特定的实例中,可以将编码前导、转运或信号肽(例如玉米ubi1前导肽)的外显子并入。
可以由并入合成稻泛素3双向启动子的外显子编码的肽包括例如但不限于:泛素(例如玉米ubi1)前导序列、叶绿体转运肽(ctp)(例如拟南芥epspsctp(klee等人(1987)mol.gen.genet.210:437-42)和矮牵牛(petuniahybrida)epspsctp(della-cioppa等人(1986)proc.natl.acad.sci.usa83:6873-7)),如对于麦草畏单加氧酶(dmo)的叶绿体靶向在国际pct公布号wo2008/105890中所例示的。
在本发明的一些实施方案中还可以将内含子并入合成的稻泛素-3双向启动子中,例如在剩余的合成稻泛素3双向启动子序列与可操作地连接到启动子的感兴趣多核苷酸序列之间。在一些实例中,并入合成的稻泛素-3双向启动子中的内含子可以为但不限于充当翻译前导序列的5′utr,其存在于翻译起始序列上游的完全加工过的mrna中(这样的翻译前导序列可以影响初始转录物到mrna的加工、mrna稳定性和/或翻译效率)。翻译前导序列的实例包括玉蜀黍和矮牵牛花热休克蛋白前导序列(美国专利号5,362,865)、植物病毒外壳蛋白前导序列、植物二磷酸核酮糖羧化酶(rubisco)前导序列等。参见例如turner和foster,(1995)molecularbiotech.3(3):225-36。5'utr的非限制性实例包括gmhsp(美国专利号5,659,122)、phdnak(美国专利号5,362,865)、atant1、tev(carrington和freed,(1990)j.virol.64:1590-7)和agrtunos(genbank登录号v00087;bevan等人,(1983)nature304:184-7)。在特定的实例中,玉米泛素1内含子可以并入合成的稻泛素-3双向启动子中。
可任选地并入合成的稻泛素-3双向启动子中的另外序列包括例如但不限于:3′非翻译序列、3′转录终止区和多聚腺苷酸化区。这些是位于感兴趣多核苷酸序列(例如可操作地连接到合成稻泛素-3双向启动子的感兴趣序列)下游的遗传元件,且包括提供多聚腺苷酸化信号的多核苷酸,和/或能够影响转录、mrna加工或基因表达的其他调节信号。多聚腺苷酸化信号可在植物中发挥作用,引起多聚腺苷酸化核苷酸添加至mrna前体的3’端。多聚腺苷酸化序列可以来源于天然基因、来源于多种植物基因或来源于t-dna基因。3’转录终止区的非限制性实例是胭脂碱合酶3’区(nos3’;fraley等人,(1983)proc.natl.acad.sci.usa80:4803-7)。使用不同的3’非翻译区的实例在ingelbrecht等人,(1989)plantcell1:671-80中提供。多聚腺苷酸化信号的非限制性实例包括来自豌豆(pisumsativum)rbcs2基因的信号(ps.rbcs2-e9;coruzzi等人,(1984)emboj.3:1671-9)和根癌土壤杆菌nos基因(genbank登录号e01312)。
在一些实施方案中,合成的稻泛素-3双向启动子包含一个或多个促进包含该启动子的核酸靶向目标生物体基因组中的特定基因座的核苷酸序列。例如,可以纳入与宿主中的基因组dna序列(例如稀有或唯一的基因组dna序列)区段同源的一个或多个序列。在一些实例中,这些同源序列可以指导包含合成稻泛素-3双向启动子的核酸在宿主基因组中的同源dna位点处的重组和整合。在特定的实例中,合成的稻泛素-3双向启动子包含一个或多个促进包含该启动子的核酸靶向宿主基因组中的稀有或唯一位置的核苷酸序列,其利用识别在该稀有或唯一位置处的序列并促进在该稀有或唯一位置处的整合的工程化的核酸酶。采用锌指内切核酸酶作为核酸酶的这样的靶向整合系统在美国专利申请号13/011,735中有所描述,其全部内容以引用方式并入本文。
在其他实施方案中,本公开还包括含有某一性状的感兴趣多核苷酸序列作为实施方案。所述性状可以是杀虫剂抗性性状、除草剂耐性性状、氮利用效率性状、水分利用效率性状、营养品质性状、dna结合性状、选择标记物性状及其任意组合。
在另外的实施方案中,性状作为转基因事件整合在转基因植物细胞内。在另外的实施方案中,转基因事件产生商品产品。因此,从本公开的转基因植物细胞衍生组合物,其中所述组合物是选自粕(meal)、面粉(flour)、蛋白质浓缩物或油的商品产品。在另外的实施方案中,包括由来源于经转化植物细胞的转基因植物产生的商品产品,其中所述商品产品包含可检出量的本发明的核酸序列。在一些实施方案中,可例如通过获得转基因植物并由其制备食物或饲料来生产此类商品产品。包含本发明的核酸序列中的一者或多者的商品产品包括(例如但不限于):植物的粗粉、油类、碾碎的或完整的籽粒或种子,以及包含含有本发明的核酸序列中的一者或多者的重组植物或种子的任何粗粉、油或者碾碎的或完整的籽粒的任何食物产品。在一种或多种商品或商品产品中检出本发明的序列中的一者或多者,实际上证明了该商品或商品产品是由被设计为表达一种或多种农业性状的转基因植物产生的。
可以使用本领域已知的任何技术来产生包含合成稻泛素-3双向启动子的核酸,包括例如但不限于:rca、pcr扩增、rt-pcr扩增、ola和snupe。这些和其他等同技术是本领域技术人员熟知的,且进一步详细记载于例如但不限于:sambrook等人,molecularcloning:alaboratorymanual,第3版,coldspringharborlaboratory,2001和ausubel等人,currentprotocolsinmolecularbiology,johnwiley&sons,1998。所有上述参考文献(包括前述两本手册)均整体以引用方式并入本文,包括其中提供的任何附图、图片和/或表格。
v.向细胞递送包含合成双向启动子rubi3的核酸分子
本公开还提供用于用包含合成稻泛素-3双向启动子的核酸分子来转化细胞的方法。根据一些实施方案,可以使用本领域已知用于将核酸分子导入植物中的大量技术中的任一种来将植物用包含合成稻泛素-3双向启动子的核酸分子转化,例如,以将一个或多个合成稻泛素-3双向启动子导入宿主植物基因组,和/或进一步导入可操作地连接到启动子的一个或多个感兴趣多核苷酸。
用于植物转化的合适方法包括可将dna导入细胞的任意方法,例如但不限于:电穿孔(参见例如美国专利5,384,253)、微粒轰击(参见例如美国专利5,015,580、5,550,318、5,538,880、6,160,208、6,399,861和6,403,865)、土壤杆菌介导的转化(参见例如美国专利5,635,055、5,824,877、5,591,616、5,981,840和6,384,301)和原生质体转化(参见例如美国专利5,508,184)。通过应用如前所述的技术,可以稳定地转化基本上任意植物物种的细胞,并且可通过本领域技术人员已知的技术使这些细胞发育成转基因植物。例如,在棉花转化的背景中可能特别有用的技术记载于美国专利5,846,797、5,159,135、5,004,863和6,624,344;具体用于转化芸苔属植物的技术记载于例如美国专利5,750,871;用于转化大豆的技术记载于例如美国专利6,384,301;而用于转化玉米的技术记载于例如美国专利7,060,876和5,591,616以及国际pct公布wo95/06722。
在实现外源核酸到受体细胞的递送后,一般鉴定出经转化的细胞用于进一步培养和植物再生。为了提高鉴定转化体的能力,技术人员可能期望采用可选择或可筛选标记基因,其中转化载体用来生成转化体。在此情况中,可通过将细胞暴露于一种或多种选择剂来测定潜在转化的细胞群体,或可对细胞筛选期望的标记基因性状。
可将暴露于选择剂后存活的细胞、或者在筛选测定中已被评分为阳性的细胞置于支持植物再生的培养基中培养。在一些实施方案中,可通过包含另外的物质(诸如生长调节剂)来改良任何合适的植物组织培养基(例如ms培养基和n6培养基)。可将组织维持在具有生长调节剂的基础培养基上,直到可得到足够的组织用于启动植物再生工作时为止,或者在重复多轮的手动选择之后,直到组织形态适合于再生时为止(例如,至少2周),然后转移到有益于芽形成的培养基中。定期转移培养物,直到已出现充分的芽形成时为止。一旦形成芽,就将其转移到有益于根形成的培养基中。一旦形成足够的根,就可将植物转移到土壤中,以便进一步生长和成熟。
为了确认包含合成的稻泛素-3双向启动子的期望的核酸分子在再生植物中的存在,可以实施多种测定。此类测定包括例如:分子生物学测定,诸如southern和northern印迹及pcr;生物化学测定,诸如检测是否存在蛋白质产物,例如通过免疫学手段(elisa和/或western印迹)或借助酶功能;植物部分测定,诸如叶或根测定;以及对再生全植物的表型的分析。
可例如通过使用例如对感兴趣的核酸分子有特异性的寡核苷酸引物进行pcr扩增来筛选目标整合事件。pcr基因分型应当理解为包括但不限于:来源于经分离的宿主植物愈伤组织的基因组dna的聚合酶链式反应(pcr)扩增,所述愈伤组织预测含有整合到基因组中的感兴趣核酸分子,接着是pcr扩增产物的标准克隆和序列分析。pcr基因分型的方法已得到充分描述(参见例如rios等人,(2002)plantj.32:243-53),并且可应用于来源于任何植物物种或组织类型的基因组dna,包括来源于细胞培养物的基因组dna。结合到靶序列和导入的序列两者的寡核苷酸引物的组合可在pcr扩增反应中按顺序使用或复用。可以产生被设计为与靶位点、导入的核酸序列和/或这两者的组合退火的寡核苷酸引物。因此,pcr基因分型策略可包括例如但不限于:植物基因组中特定序列的扩增、植物基因组中多个特定序列的扩增、植物基因组中非特定序列的扩增和前述任何扩增的组合。本领域的技术人员可设计出引物和扩增反应的额外组合来查询基因组。例如,一组正向和反向寡核苷酸引物可被设计为与特异于所导入的核酸序列边界外的靶标的核酸序列退火。
正向和反向寡核苷酸引物可被设计为与导入的核酸分子特异性退火,例如在对应于其中包含的感兴趣多核苷酸序列内的编码区的序列处,或核酸分子的其他部分。这些引物可连同上文所述的引物一起使用。寡核苷酸引物可根据期望的序列而合成,并且可商购获得(例如购自integrateddnatechnologies,inc.,coralville,ia)。扩增之后可以是克隆和测序,或扩增产物的直接序列分析。本领域技术人员可想到用于分析在pcr基因分型期间生成的扩增产物的备选方法。在一个实施方案中,在pcr扩增中采用特异于基因靶标的寡核苷酸引物。
vi.包含合成的双向启动子rubi3的细胞、细胞培养物、组织和生物体
本发明的一些实施方案还提供包含合成稻泛素-3双向启动子(例如可以存在于核酸构建体中的)的细胞。在特定的实例中,根据一些实施方案的合成稻泛素-3双向启动子可用作调控序列来调控转基因在植物细胞和植物中的表达。在一些这样的实例中,使用可操作地连接到感兴趣多核苷酸序列(例如转基因)的合成双向rubi3启动子可以减少调控给定数目的感兴趣多核苷酸序列的表达所需的异源启动子的数目,和/或减小导入给定数目的感兴趣核苷酸序列所需的核酸构建体的大小。此外,使用合成稻泛素-3双向启动子可以允许两个可操作连接的感兴趣核苷酸序列在相同条件(即该rubi3启动子有活性的条件)下的共表达。这样的实例在例如以下情况中可能特别有用:两个可操作地连接的感兴趣多核苷酸序列各自有助于包含该感兴趣核苷酸序列的转基因宿主中的单一性状,且感兴趣核苷酸序列的共表达有利地影响该性状在转基因宿主中的表达。
在一些实施方案中,包含一个或多个合成稻泛素-3双向启动子和/或感兴趣核苷酸序列的转基因植物可以具有由感兴趣核苷酸序列在植物中的表达所赋予(例如引入、增强或有助于)的一种或多种期望的性状。此类性状可以包括例如但不限于:对昆虫、其他害虫和病原的抗性;对除草剂的耐受性;提高的稳定性、产量或货架期;环境耐受性;药物产生;工业产物产生;和营养增强。在一些实例中,可通过以下方式赋予期望的性状:用包含可操作地连接到感兴趣多核苷酸序列的合成稻泛素-3双向启动子的核酸分子来转化植物。在一些实例中,可对经由育种产生作为后代植物的植物赋予期望的性状,该性状可以是由可操作地连接到合成稻泛素-3双向启动子的一个或多个感兴趣多核苷酸序列赋予的,所述感兴趣核苷酸序列是从包含可操作地连接到合成稻泛素-3双向启动子的感兴趣核苷酸序列的亲本植物传递给该植物的。
根据一些实施方案的转基因植物可以是任何能够用本发明的核酸分子转化,或能够用经本发明的核酸分子转化的植物育种产生的植物。因此,所述植物可以是双子叶或单子叶植物。用于一些实例的双子叶植物的非限制性实例包括:苜蓿、豆类、椰菜、甘蓝、卡诺拉油菜、胡萝卜、花椰菜、芹菜、大白菜、棉花、黄瓜、茄子、莴苣、甜瓜、豌豆、胡椒、花生、马铃薯、南瓜、萝卜、油菜籽、菠菜、大豆、瓜类、甜菜、向日葵、烟草、番茄和西瓜。用于一些实例的单子叶植物的非限制性实例包括:短柄草属、玉米、洋葱、稻、高粱、小麦、黑麦、小米、甘蔗、燕麦、黑小麦、柳枝稷和草坪草。
在一些实施方案中,可以任意方式来使用或栽培转基因植物,其中期望存在合成的稻泛素-3双向启动子和/或可操作地连接的感兴趣多核苷酸序列。因此,此类转基因植物可通过用根据本发明的核酸分子转化而工程化,以尤其具有一种或多种期望的性状或者转基因事件,并且此类转基因植物可通过本领域技术人员已知的任意方法收获和/或栽培。
提供以下实施例是为了例示某些特定的特征和/或实施方案。这些实施例不应被理解为将本公开限于所举例说明的特定特征或实施方案。
实施例
实施例1:材料和方法
样品制备和生物测定
使用
在使用饲喂人工昆虫食料的新生昆虫幼虫进行的生物测定中测试样品的昆虫活性。wcr卵获自cropcharacteristics,inc.(farmington,mn)。
生物测定在特别为昆虫生物测定设计的128孔塑料托盘(c-dinternational,pitman,nj)中进行。每个孔装有约1.0ml设计用于支持鞘翅目昆虫生长的人工食料。用移液管将60μl等份的dsrna样品递送到每个孔的食料的表面上(40μl/cm2)。dsrna样品浓度作为孔中每平方厘米表面积(1.5cm2)的dsrna量(ng/cm2)来计算。将经处理的托盘保持在通风橱中,直到食料表面上的液体蒸发或吸收到食料中为止。
在孵化后的几个小时内,用湿润的骆驼毛刷拾取幼虫个体,将其放置在经处理的食料上(每孔一个或两个幼虫)。然后用透明塑料粘合片密封128孔塑料托盘上带虫的孔,并通气以允许气体交换。使生物测定托盘在受控环境条件(28℃,约40%相对湿度,16:8(光照:黑暗))下保持9天,之后记录暴露于每个样品的昆虫总数、死亡昆虫数以及存活昆虫的重量。计算每个处理的平均死亡率百分比和平均生长抑制。生长抑制(gi)如下计算:
gi=[1–(twit/tnit)/(twibc/tnibc)],
其中twit是处理中的活昆虫的总重量;
tnit是处理中的昆虫的总数;
twibc是背景检查(缓冲液对照)中的活昆虫的总重量;
tnibc是背景检查(缓冲液对照)中的昆虫的总数。
使用jmptm软件(sas,cary,nc)进行统计分析。
lc50(致死浓度)定义为50%的测试昆虫被杀死时的剂量。gi50(生长抑制)定义为测试昆虫的平均生长(例如活体重)为背景检查样品中观察到的平均值的50%时的剂量。
重复的生物测定证明,摄入特定的样品造成了玉米根虫幼虫的令人惊讶且意想不到的死亡率和生长抑制。
实施例2:鉴定候选靶标基因
选择来自多个wcr(玉米根萤叶甲)发育期的昆虫进行汇集的转录组分析,以提供通过rnai转基因植物昆虫防护技术控制的候选靶标基因序列。
在一个范例中,从约0.9g的完整第一龄wcr幼虫(孵化后4到5天;保持在16℃)分离总rna,并使用下述基于苯酚/tri
室温下将幼虫置于装有10mltri
小心取出并弃去上清液,然后通过用75%乙醇涡旋将rna沉淀洗涤两次,每次洗涤后通过以7,500xg(4℃或25℃)离心5分钟加以回收。小心地除去乙醇,让沉淀风干3至5分钟,然后溶解在无核酸酶的无菌水中。通过在260nm和280nm处测量吸光度(a)来测定rna浓度。从约0.9g幼虫的典型提取过程产生超过1mg的总rna,其中a260与a280的比值为1.9。如此提取的rna在-80℃储存,直到进一步加工。
通过使等份跑过1%琼脂糖凝胶来测定rna质量。在经高温灭菌的容器中,使用由经depc(焦碳酸二乙酯)处理的水稀释的经高温灭菌10xtae缓冲液(tris乙酸盐edta;1x浓度为0.04mtris乙酸盐、1mmedta(乙二胺四乙酸钠盐),ph8.0)制成琼脂糖凝胶溶液。使用1xtae作为运行缓冲液。使用前,用rnaseawaytm(invitrogeninc.,carlsbad,ca)清洁电泳槽和造孔梳。取2μlrna样品与8μlte缓冲液(10mmtrishcl,ph7.0;1mmedta)和10μlrna样品缓冲液(
由商业服务提供商(eurofinsmwgoperon,huntsville,al)利用随机引发自幼虫总rna制备标准化的cdna文库。在eurofinsmwgoperon通过gsflx454titaniumtm系列化学品以1/2板规模对标准化的幼虫cdna文库进行测序,产生平均读段长度为348bp的超过600,000个读段。将350,000个读段组装成超过50,000个重叠群。使用公众可得到的程序formatdb(可获自ncbi)将未组装的读段和重叠群这两者都转换成可blast的数据库。
类似地由在其他wcr发育期收获的材料制备了总rna和标准化的cdna文库。合并代表各发育期的cdna文库成员,由此构建用于筛选靶标基因的汇集的转录组文库。
假设用于rnai靶向的候选基因对于害虫昆虫的存活和生长是必需的。对于选择的靶标基因,在转录组序列数据库中鉴定其同源物,如下所述。通过pcr扩增靶标基因的全长或部分序列,来制备用于产生双链rna(dsrna)的模板。
使用候选蛋白质编码序列针对含有未组装的叶甲属序列读段或已组装的重叠群的可blast数据库运行tblastn搜索。使用blastx针对ncbi非冗余数据库确认对叶甲属序列的显著命中(定义为:对于重叠群同源性,优于e-20;对于未组装的序列读段同源性,优于e-10)。此blastx搜索的结果确认,在tblastn搜索中鉴定的叶甲属同源物候选基因序列确实包含叶甲属基因,或者是针对叶甲属序列中存在的非叶甲属候选基因序列的最佳命中。在少数情况下可明显看出,一些基于与非叶甲属候选基因的同源性选择的叶甲属重叠群或未组装的序列读段有重叠,并且对重叠群的组装未能连接起这些重叠。在这些情况下,使用sequenchertmv4.9(genecodescorporation,annarbor,mi)将序列组装成更长的重叠群。
编码叶甲属rpii33的几种候选靶标基因(seqidno:1和seqidno:3)被鉴定为可造成鞘翅目害虫死亡,wcr生长抑制、发育抑制和/或进食抑制的基因。
seqidno:1和seqidno:3的多核苷酸是新颖的。这些序列未在公共数据库中提供,也未在pct国际专利公布号wo/2011/025860、美国专利申请号20070124836、美国专利申请号20090306189、美国专利申请号us20070050860、美国专利申请号20100192265、美国专利号7,612,194和美国专利申请号2013192256中公开。叶甲属rpii33-1(seqidno:1)与来自果蝇(ceratitiscapitata)的序列的片段(genbank登录号xm_002064757.1)在一定程度上相关。不存在与在genbank中发现的叶甲属rpii33-2(seqidno:3)显著同源的核苷酸序列。叶甲属rpii33-1氨基酸序列(seqidno:2)的最接近的同源物为埃和伊蚊(aedesaegypti)蛋白,其genbank登录号为xp_001659470.1(94%相似;在同源区上87%相同)。叶甲属rpii33-2氨基酸序列(seqidno:4)的最接近的同源物为中欧山松大小蠹(dendroctonusponderosae)蛋白,其genbank登录号为aae63493.1(96%相似;在同源区上91%相同)。
rpii33dsrna转基因可与其他dsrna分子组合,以提供冗余的rnai靶向和协同的rnai效应。表达靶向rpii33的dsrna的转基因玉米事件可用于防止玉米根虫引起的噬根性损害。rpii33dsrna转基因代表新的作用模式,可用来与苏云金芽孢杆菌杀虫蛋白技术在昆虫抗性管理基因累加(insectresistancemanagementgenepyramid)中结合,以减缓根虫群体对这些根虫控制技术中的任一者产生抗性。
实施例3:扩增靶标基因以产生dsrna
使用rpii33候选基因的序列的全长或部分克隆来生成pcr扩增子以供dsrna合成。设计引物,以便通过pcr扩增每个靶标基因的编码区部分。参见表1。在适当情况下,将t7噬菌体启动子序列(ttaatacgactcactatagggaga;seqidno:9)掺入经扩增的有义链或反义链的5’端。参见表1。使用
表1.用来扩增示例性rpii-33靶标基因和yfp阴性对照基因的编码区部分的引物和引物对。
实施例4:rnai构建体
通过pcr制备模板以及dsrna合成。
图1中示出了用于提供特异性模板以产生rpii33和yfpdsrna的策略。使用表1中的引物对,以及由分离自wcr卵、第一龄幼虫或成虫的总rna制备的第一链cdna(作为pcr模板),通过pcr制备了意图在rpii33dsrna合成中使用的模板dna。对于每个选定的rpii33和yfp靶标基因区,pcr扩增在经扩增的有义链和反义链的5’端导入了一个t7启动子序列(yfp区段扩增自yfp编码区的dna克隆)。然后将每个靶标基因区的两个经pcr扩增的片段以大致相等的量混合,并将混合物用作产生dsrna的转录模板。参见图1。用特定引物对扩增的dsrna模板的序列为:seqidno:5(rpii33-1reg1)、seqidno:6(rpii33-2reg1)、seqidno:7(rpii33-2ver1)、seqidno:8(rpii33-2v2)和yfp(seqidno:10)。使用
构建植物转化载体。
使用化学合成片段(dna2.0,menlopark,ca)的组合以及标准分子克隆方法,组装了入门载体,该入门载体包含具有rpii33的区段(seqidno:1、seqidno:3、seqidno:76或seqidno:78)的用于形成发夹的靶标基因构建体。通过(在单个转录单位内)以彼此相反的取向排布rpii33靶标基因区段的两个拷贝来易化rna初级转录物形成分子内发夹,所述两个区段被一个接头多核苷酸(例如seqidno:107和st-ls1内含子(vancanneyt等人,(1990)mol.gen.genet.220(2):245-50))分开。因此,初级mrna转录物含有被所述内含子序列分开的两个rpii33基因区段序列,这两个区段序列互为大的反向重复序列。使用启动子(例如,玉蜀黍遍在蛋白1,美国专利号5,510,474;来自花椰菜花叶病毒(camv)的35s;甘蔗杆状病毒(scbv)启动子;来自水稻肌动蛋白基因的启动子;遍在蛋白启动子、pemu、mas、玉蜀黍h3组蛋白启动子、als启动子、菜豆素基因启动子、cab、rubisco、lat52、zm13和/或apg)的拷贝来驱动初级mrna发夹转录物的产生,并用一个包含3’非翻译区(例如,玉蜀黍过氧化物酶5基因(zmper53'utrv2;美国专利号6,699,984)、atubi10、atef1或stpinii)的片段来终止表达发夹rna的基因的转录。
入门载体pdab126158和pdab126159包含rpii33-rna构建体(分别为seqidno:103和104),该构建体含有rpii33的区段(分别为seqidno:7和8)。
在标准的
二元目标载体包含处于植物可操作启动子(例如,甘蔗杆状病毒(scbv)启动子(schenk等人,(1999)plantmol.biol.39:1221-30)和zmubi1(美国专利号5,510,474))的调节之下的除草剂耐受性基因(芳氧基链烷酸酯加双氧酶;aad-1v3)(美国专利号7,838,733(b2),以及wright等人,(2010)proc.natl.acad.sci.u.s.a.107:20240-5)。5'utr和内含子定位在所述启动子区段的3’端与aad-1编码区的起始密码子之间。使用一个包含来自玉蜀黍脂肪酶基因的3’非翻译区(zmlip3'utr;美国专利号7,179,902)的片段来终止aad-1mrna的转录。
借助使用典型的二元目标载体与入门载体的标准
实施例5:筛选候选靶标基因
在基于食料的测定中,将设计为抑制实施例2中鉴定的靶标基因序列的合成dsrna施用于wcr时,引起了死亡和生长抑制。
重复的生物测定证明,摄入来源于rpii33-2reg1、rpii33-2v1和rpii33-2v2的dsrna制备物造成了西方玉米根虫幼虫的死亡和生长抑制。表2和表3示出了在wcr幼虫暴露于这些dsrna9天之后的基于食料的饲喂生物测定的结果,以及由黄色荧光蛋白(yfp)编码区(seqidno:10)制备的dsrna阴性对照样品获得的结果。
表2。利用西方玉米根虫幼虫进食9天之后获得的rpii33dsrna食料饲喂测定的结果。anova分析发现了平均死亡率%和平均生长抑制(gi)%上的显著差异。使用tukey-kramer检验分离平均值。
*sem=平均值标准误差括号中的字母指明统计水平。不是由同一字母连接的水平存在显著差异(p<0.05)。
**te=trishcl(1mm)加edta(0.1mm)缓冲液,ph7.2。
***yfp=黄色荧光蛋白
表3。rpii33dsrna对wcr幼虫的口服效力(ng/cm2)汇总。
以前有人提出,某些叶甲属物种基因可用于rnai介导的昆虫控制。参见美国专利公布号2007/0124836(其公开了906个序列)和美国专利号7,612,194(其公开了9,112个序列)。然而,技术人员确定,许多被提出对rnai介导的昆虫控制有效用的基因并不能有效地控制叶甲属。技术人员还确定,与被提出对rnai介导的昆虫控制有效用的其他基因相比,序列rpii33-2v1、rpii33-2v2和rpii33-2reg1提供了令人惊讶且意想不到的对叶甲属的出色控制。
例如,美国专利号7,612,194中提出,膜联蛋白、β血影蛋白2和mtrp-l4在rnai介导的昆虫控制中全都有效。seqidno:20为膜联蛋白区域1(reg1)的dna序列,而seqidno:21为膜联蛋白区域2(reg2)的dna序列。seqidno:22为β血影蛋白2区域1(reg1)的dna序列,而seqidno:23为β血影蛋白2区域2(reg2)的dna序列。seqidno:24为mtrp-l4区域1(reg1)的dna序列,而seqidno:25为mtrp-l4区域2(reg2)的dna序列。还使用yfp序列(seqidno:10)来产生作为阴性对照的dsrna。
使用前述序列中的每一个通过实施例3的方法来产生dsrna。图2中示出了用于提供特异性模板以产生dsrna的策略。使用表4中的引物对,以及由分离自wcr第一龄幼虫的总rna制备的第一链cdna(作为pcr模板),通过pcr制备了意图在dsrna合成中使用的模板dna。(yfp从dna克隆扩增。)对于每个选择的靶标基因区,执行两次单独的pcr扩增。第一次pcr扩增在经扩增有义链的5’端导入t7启动子序列。第二次反应在反义链的5’端掺入t7启动子序列。然后将每个靶标基因区的两个经pcr扩增的片段以大致相等的量混合,并将混合物用作产生dsrna的转录模板。参见图2。使用
表4.用来扩增基因的编码区部分的引物和引物对。
表5.利用9天之后的西方玉米根虫幼虫获得的食料饲喂测定的结果。
*te=trishcl(10mm)加edta(1mm)缓冲液,ph8。
**yfp=黄色荧光蛋白
实施例6:产生包含杀虫dsrna的
转基因玉蜀黍组织
土壤杆菌介导的转化。在土壤杆菌介导的转化之后,制成了通过表达稳定地整合到植物基因组中的嵌合基因而产生一种或多种杀虫dsrna分子(例如,至少一种dsrna分子,包括靶向包含rpii33的基因(例如seqidno:1和seqidno:3))的dsrna分子)的转基因玉蜀黍细胞、组织和植物。采用超二元转化载体或二元转化载体的玉蜀黍转化方法是本领域已知的,如例如美国专利号8,304,604中所述,该专利全文以引用方式并入本文。根据在含有吡氟氯禾灵的培养基上生长的能力来选择经转化组织,并酌情筛选经转化组织来产生dsrna。将一部分这样的经转化组织培养物提供给新生玉米根虫幼虫以执行生物测定,大体上如实施例1中所述。
土壤杆菌培养启动。将包含上述(实施例4)二元转化载体的土壤杆菌菌株dat13192细胞(pct国际公布号wo2012/016222a2)的甘油储备液划线接种在含有适当抗生素的ab基本培养基平板(watson等人,(1975)j.bacteriol.123:255-264)上,并在20℃下生长3天。然后将培养物划线接种到含有相同抗生素的yep平板(酵母提取物,10g/l;蛋白胨,10g/l;nacl,5g/l)上,并在20℃下温育1天。
土壤杆菌培养。实验当天,以适合于实验中的构建体数目的体积制备接种培养基和乙酰丁香酮的储备液,并将其吸移到250ml一次性无菌烧瓶中。接种培养基(frame等人,(2011)genetictransformationusingmaizeimmaturezygoticembryos,载于plantembryoculturemethodsandprotocols:methodsinmolecularbiology。t.a.thorpe和e.c.yeung(编辑),springerscienceandbusinessmedia,llc.,第327-341页)含有:2.2g/lms盐,1xisu改良的ms维生素(frame等人,出处同上),68.4g/l蔗糖,36g/l葡萄糖,115mg/ll-脯氨酸,以及100mg/l肌醇;ph为5.4。将乙酰丁香酮从100%二甲基亚砜中的1m储备液添加到装有接种培养基的烧瓶中,达到200μm的最终浓度,并充分混合溶液。
对于每种构建体,将来自yep平板的1或2个满接种环的土壤杆菌悬浮在50ml一次性无菌离心管内的15ml接种培养基/乙酰丁香酮储备液中,然后在分光光度计中测量溶液在550nm处的光密度(od550)。然后使用额外的接种培养基/乙酰丁香酮混合物将悬浮液稀释到0.3至0.4的od550。然后将装有土壤杆菌悬浮液的管水平放置在平台摇床(设置在室温下,约75rpm)上,在进行胚切开的同时振摇1至4小时。
穗消毒和胚分离。从玉米近交系b104(hallauer等人,(1997)cropscience37:1405-1406)植物获得未成熟玉蜀黍胚,所述植物在温室中培养,并进行自花授粉或近缘授粉以产生穗。在授粉后大约10至12天收获穗。实验当天,将穗脱壳,通过浸没在市售漂白剂(ultra
土壤杆菌共培养。分离之后,将胚在摇动平台上放置5分钟。然后将管的内容物倾倒在共培养培养基的平板上,所述培养基含有4.33g/lms盐、1xisu改良的ms维生素、30g/l蔗糖、700mg/ll-脯氨酸、3.3mg/l麦草畏(3,6-二氯-o-茴香酸或3,6-二氯-2-甲氧基苯甲酸)的koh溶液、100mg/l肌醇、100mg/l酪蛋白酶水解物、15mg/lagno3、200μm乙酰丁香酮的dmso溶液和3g/lgelzantm,ph为5.8。用一次性无菌移液管移除液态土壤杆菌悬浮液。然后借助显微镜,使用无菌镊子使胚取向为盾片面朝上。盖上平板,用3mtmmicroporetm医用胶带密封,然后放置在具有大约60μmolm-2s-1光合有效辐射(par)的连续光照的25℃培养箱中。
选择愈伤组织和再生转基因事件。在共培养期之后,将胚转移到静息培养基上,静息培养基的组成为:4.33g/lms盐、1xisu改良的ms维生素、30g/l蔗糖、700mg/ll-脯氨酸、3.3mg/l麦草畏的koh溶液、100mg/l肌醇、100mg/l酪蛋白酶水解物、15mg/lagno3、0.5g/lmes(2-(n-吗啉代)乙磺酸一水合物;phytotechnologieslabr.;lenexa,ks)、250mg/l羧苄青霉素和2.3g/lgelzantm,ph为5.8。将不超过36个胚移到每个平板上。将平板放置在透明的塑料盒中,在27℃和大约50μmolm-2s-1par的连续光照下温育7至10天。然后将愈伤的胚转移(<18个/板)到选择培养基i上,该培养基由具有100nmr-吡氟氯禾灵酸(0.0362mg/l;用于选择包含aad-1基因的愈伤组织)的静息培养基(如上文)组成。将平板放回透明盒中,在27℃和大约50μmolm-2s-1par的连续光照下温育7天。然后将愈伤的胚转移(<12个/板)到选择培养基ii上,该培养基由具有500nmr-吡氟氯禾灵酸(0.181mg/l)的静息培养基(如上文)组成。将平板返回到透明盒中,在27℃和大约50μmolm-2s-1par的连续光照下温育14天。这一选择步骤允许转基因愈伤组织进一步增殖和分化。
将增殖中的胚性愈伤组织转移(<9个/板)到预再生培养基上。预再生培养基含有4.33g/lms盐、1xisu改良的ms维生素、45g/l蔗糖、350mg/ll-脯氨酸、100mg/l肌醇、50mg/l酪蛋白酶水解物、1.0mg/lagno3、0.25g/lmes、0.5mg/l萘乙酸的naoh溶液、2.5mg/l脱落酸的乙醇溶液、1mg/l6-苄基氨基嘌呤、250mg/l羧苄青霉素、2.5g/lgelzantm和0.181mg/l吡氟氯禾灵酸,ph为5.8。将平板保存在透明盒中,在27℃和大约50μmolm-2s-1par的连续光照下温育7天。然后将再生中的愈伤组织转移(<6个/板)到phytatraystm(sigma-aldrich)中的再生培养基上,在28℃下以每天16小时光照/8小时黑暗(大约160μmolm-2s-1par)温育14天,或直到发出芽和根为止。再生培养基含有4.33g/lms盐、1xisu改良的ms维生素、60g/l蔗糖、100mg/l肌醇、125mg/l羧苄青霉素、3g/lgellantm胶和0.181mg/lr-吡氟氯禾灵酸,ph为5.8。然后分离具有初生根的小芽,不经选择直接转移到伸长培养基上。伸长培养基含有4.33g/lms盐、1xisu改良的ms维生素、30g/l蔗糖和3.5g/lgelritetm,ph为5.8。
根据在含有吡氟氯禾灵的培养基上生长的能力来选择经转化的植物芽,将这些芽从phytatraystm移栽到填充生长培养基(promixbx;premiertechhorticulture)的小盆中,小盆用杯子或humi-domes(arcoplastics)覆盖,然后在conviron生长室中炼苗(白天27℃/夜晚24℃,16小时光周期,50-70%rh,200μmolm-2s-1par)。在一些情况下,分析推定的转基因小植物的转基因相对拷贝数,这使用被设计为检测整合到玉蜀黍基因组中的aad1除草剂耐受性基因的引物通过定量实时pcr测定来完成。另外,利用rt-qpcr测定来检测推定的转化体中是否存在接头序列和/或靶标序列。然后将选择的经转化小植物移到温室中,以便进一步生长和测试。
转移t0植物并在温室中定植,以进行生物测定和产生种子。当植物达到v3-v4期时,将其移栽到iecustomblend(profile/metromix160)土壤混合物中,在温室中(光暴露类型:光合或同化;高光限值:1200par;16小时昼长;白天27℃/夜晚24℃)培植到开花。
将要用于昆虫生物测定的植物从小盆移栽到tinustm350-4
通过对t0转基因植物的穗丝授粉(其中花粉从非转基因近交系b104植物或其他适当的花粉供体采集而来)并种植所得的种子而获得t1世代植物。在可能时执行互交。
实施例7:转基因玉蜀黍组织的分子分析
对在评估噬根性损害前一天或同一天从温室培植的植物采集的叶的样品执行了玉蜀黍组织的分子分析(例如rt-qpcr)。
使用靶标基因的rt-qpcr测定的结果来验证转基因的表达。作为替代,使用表达的rna中重复序列之间的间插序列(为形成dsrna发夹分子必不可少的)的rt-qpcr测定结果来验证是否存在发夹转录物。测量了相对于内源性玉蜀黍基因的rna水平的转基因rna表达水平。
通过dnaqpcr分析检测gdna中aad1编码区的一部分,用来估计转基因插入拷贝数。从培植在环境室中的植物采集样品用于这些分析。将结果与被设计为检测单拷贝天然基因的一部分的测定法的dnaqpcr结果进行比较,并且将简单事件(具有rpii33转基因的一个或两个拷贝)推进到温室中的进一步研究。
此外,使用被设计为检测壮观霉素抗性基因(specr;包含在t-dna外部的二元载体质粒上)的一部分的qpcr测定来确定转基因植物是否含有外来的整合质粒主干序列。rna转录物表达水平:靶向qpcr。通过靶标序列的实时定量pcr(qpcr)来分析愈伤组织细胞事件或转基因植物以确定转基因的相对表达水平,与内部玉蜀黍基因(seqidno:54,genbank登录号bt069734)的转录物水平比较,所述内部玉蜀黍基因编码tip41样蛋白(即,genbank登录号at4g34270的玉蜀黍同源物;tblastx得分为74%同一性)。使用norgenbiotektm总rna分离试剂盒(norgen,thorold,on)分离rna。按照试剂盒建议的方案对总rna进行柱上dnase1处理。然后在nanodrop8000分光光度计(thermoscientific)上将rna定量,并将浓度归一化为50ng/μl。基本上按照制造商推荐的方案,使用高容量cdna合成试剂盒(invitrogen)制备第一链cdna,反应体积为10μl,含有5μl变性rna。略微修改该方案,包括将10μl的100μmt20vn寡核苷酸(idt)(ttttttttttttttttttttvn,其中v是a、c或g,n是a、c、g或t;seqidno:55)添加到随机引物储备混合物的1ml管中,以制备随机引物和寡dt混合的工作储备液。
在cdna合成后,用无核酸酶的水将样品以1:3稀释,并储存在-20℃直到测定时为止。
在lightcyclertm480(rochediagnostics,indianapolis,in)上以10μl反应体积分别执行对靶标基因和tip41样转录物的实时pcr测定。对于靶标基因测定,反应用引物rpii33v1fwdset2(seqidno:56)和rpii33v1revset2(seqidno:57)以及idt定制寡核苷酸探针rpii33v1prbset2运行、用fam标记且用zen和iowablack淬灭剂(seqidno:105)予以双重淬灭;或用引物rpii33v2fwdset2(seqidno:111)和rpii33v2revset2(seqidno:112)以及idt定制寡核苷酸探针rpii33v2prbset2运行、用fam标记且用zen和iowablack淬灭剂(seqidno:106)予以双重淬灭。对于tip41样参考基因测定,使用引物tipmxf(seqidno:58)和tipmxr(seqidno:59),以及用hex(六氯荧光素)标记的探针hxtip(seqidno:60)。
所有测定都包括没有模板的阴性对照(仅有混合物)。为制备标准曲线,在源板中还包括空白(源孔中加水),以检查样品交叉污染。引物和探针的序列在表6中示出。用于检测各种转录物的反应组分配方在表7中公开,pcr反应条件汇总于表8中。在465nm处激发fam(6-羧基荧光素亚磷酰胺)荧光部分,并测量510nm处的荧光;hex(六氯荧光素)荧光部分的对应值为533nm和580nm。
表6.用于转基因玉蜀黍中转录物水平的分子分析的寡核苷酸序列。
*tip41样蛋白。
表7.用于检测转录物的pcr反应配方。
表8.用于rnaqpcr的热循环仪条件。
使用lightcyclertm软件v1.5,根据供应商的推荐利用二阶导数最大算法计算cq值,通过相对定量分析数据。对于表达分析,使用δδct方法(即2-(cqtarget–cqref))计算表达值,该方法依赖于比较两个靶标之间的cq值的差异,其中假定对于优化的pcr反应,每个循环产物都加倍,基值选择为2。
转录物大小和完整性:northern印迹测定。在一些情况下,使用northern印迹(rna印迹)分析来确定在表达rpii33发夹dsrna的转基因植物中rpii33发夹dsrna的分子大小,从而获得转基因植物的额外的分子表征。
所有的材料和设备在使用之前用rnasezap(ambion/invitrogen)处理。将组织样品(100mg至500mg)采集到2mlsafelockeppendorf管中,用配备三颗钨珠的kleckotm组织粉碎器(garciamanufacturing,visalia,ca)在1mltrizol(invitrogen)中破碎5分钟,然后在室温(rt)下温育10分钟。任选地,将样品在4℃下以11,000rpm离心10分钟,然后将上清液转移到新鲜的2mlsafelockeppendorf管中。在将200μl氯仿添加到匀浆中之后,通过翻转该管2至5分钟进行混合,在rt下温育10分钟,然后在4℃下以12,000xg离心15分钟。将上层相转移到无菌的1.5mleppendorf管中,添加600μl的100%异丙醇,在rt下温育10分钟至2小时之后,在4℃至25℃下以12,000xg离心10分钟。弃去上清液,用1ml的70%乙醇将rna沉淀洗涤两次,在两次洗涤之间,在4℃至25℃下以7,500xg离心10分钟。弃去乙醇,将沉淀短暂风干3至5分钟,然后重悬在50μl无核酸酶的水中。
使用nanodrop
电泳之后,在2xssc中将凝胶漂洗5分钟,然后在geldoc工作站(biorad,hercules,ca)上成像,接着在rt下过夜,使rna被动地转移到尼龙膜(millipore)上,其中使用10xssc作为转移缓冲液(由3m氯化钠和300m梓檬酸三钠组成的20xssc,ph7.0)。转移后,在2xssc中将膜漂洗5分钟,利用紫外线使rna与膜交联(agilent/stratagene),然后使膜在室温下干燥最多2天。
使膜在ultrahybtm缓冲液(ambion/invitrogen)中预杂交1至2小时。探针由含有感兴趣序列(例如,seqidno:5-8或103-104的反义序列部分,视情况而定)的pcr扩增产物组成,其借助rocheappliedsciencedig程序用地高辛配基标记。在杂交管中推荐的缓冲液中,于60℃的温度下杂交过夜。杂交后,对印迹进行dig洗涤,包装,暴露于胶片1至30分钟,然后将胶片显影,所有这些操作都通过dig试剂盒的供应商所推荐的方法来进行。
确定转基因拷贝数。将大约等同于2个叶冲孔块(punch)的玉蜀黍叶片收集在96孔的收集平板(qiagentm)中。用配备一颗不锈钢珠的kleckotm组织粉碎器(garciamanufacturing,visalia,ca)在biosprint96ap1裂解缓冲液(与biosprint96plantkit一起提供;qiagen)中进行组织破碎。组织浸软之后,使用biosprint96plantkit和biosprint96提取机器人以高通量形式分离gdna。在设立qpcr反应之前,以1:3的dna:水稀释gdna。
qpcr分析。通过使用
使用拟合点算法(
表9.用于确定基因拷贝数和检测二元载体质粒主干的引物和探针(具有荧光缀合物)的序列。
cy5=青色素-5
表10.用于分析基因拷贝数和检测质粒主干的反应组分。
*na=不适用
**nd=未确定
表11.用于dnaqpcr的热循环仪条件。
实施例8:转基因玉蜀黍的生物测定
昆虫生物测定。通过生物测定方法来证实在植物细胞中产生的本主题发明的dsrna的生物活性。参见例如baum等人,(2007)nat.biotechnol.25(11):1322-1326。技术人员能够例如通过在受控的饲喂环境下给靶标昆虫喂食来源于产生杀虫dsrna的植物的各种植物组织或组织片来证实效力。作为替代,由来源于产生杀虫dsrna的植物的各种植物组织制备提取物,并将提取的核酸分配在用于如本文之前描述的生物测定的人工食料上面。将这样的饲喂测定的结果与类似进行的采用来自不产生杀虫dsrna的宿主植物的适当对照组织、或其他对照样品的生物测定进行比较。相比于对照组,测试食料上的靶标昆虫的生长减缓且存活率降低。
使用转基因玉蜀黍事件的昆虫生物测定。选择从经洗涤的卵孵化的两条西方玉米根虫幼虫(1至3日龄),并将其置于生物测定托盘的每个孔中。然后将这些孔用“拉-揭”(pulln'peel)护盖(bio-cv-16,bio-serv)覆盖,并置于具有18小时/6小时的光照/黑暗周期的28℃培养箱中。在初始侵染9天之后,评估幼虫死亡率,其按照每个处理中死亡昆虫在昆虫总数中所占的百分比来计算。将昆虫样品在-20℃冷冻两天,然后汇集来自每个处理的昆虫幼虫并称重。生长抑制百分比按照实验处理的平均重量除以两个对照孔处理的平均重量的均值来计算。数据表示为(阴性对照的)生长抑制百分比。将超过对照平均重量的平均重量归一化为零。
温室中的昆虫生物测定。从cropcharacteristics(farmington,mn)收到带有西方玉米根虫(wcr,玉米根萤叶甲)卵的土壤。在28℃下温育wcr卵10至11天。洗掉卵上的土壤,将卵置于0.15%琼脂溶液中,把浓度调节至每0.25ml等份大约75至100个卵。将一份卵悬浮液加入培养皿中以设置孵化平板,用于监测孵化速率。
用150至200个wcr卵侵染
转基因阴性对照植物通过用包含被设计为产生黄色荧光蛋白(yfp)的基因的载体转化而生成。非转化的阴性对照植物由产生转基因植物的亲代玉米品种的种子培植而来。进行生物测定,其中每一组植物材料中均包括阴性对照。
实施例9:包含鞘翅目害虫序列的转基因玉米
如实施例6中所述生成10至20株转基因t0玉米植物。获得另外的10至20个表达rnai构建体的发夹dsrna的t1玉米独立品系用于玉米根虫攻击。发夹dsrna包含seqidno:1或seqidno:3的一部分(例如,从seqidno:103和seqidno:104转录的发夹dsrna)。额外的发夹dsrna来源于例如鞘翅目害虫序列,诸如caf1-180(美国专利申请公布号2012/0174258)、vatpasec(美国专利申请公布号2012/0174259)、rho1(美国专利申请公布号2012/0174260)、vatpaseh(美国专利申请公布号2012/0198586)、ppi-87b(美国专利申请公布号2013/0091600)、rpa70(美国专利申请公布号2013/0091601)、rps6(美国专利申请公布号2013/0097730)、rop(美国专利申请公布号14/577811)、rnapii(美国专利申请公布号14/577854)、dre4(美国专利申请号14/705,807)、ncm(美国专利申请号62/095487)、copiα(美国专利申请号62/063,199)、copiβ(美国专利申请号62/063,203)、copiγ(美国专利申请号62/063,192)或copiδ(美国专利申请号62/063,216)。这些通过rt-pcr或其他分子分析方法得到确认。
来自选择的独立t1品系的总rna制备物任选地用于rt-pcr,其中引物被设计为在每个rnai构建体中的发夹表达盒的接头中结合。另外,用于rnai构建体中每个靶标基因的特异性引物任选地用于扩增在植物内产生sirna所需要的预加工mrna,并用于确认产生了所述预加工的mrna。对于每个靶标基因的期望条带的扩增确认每个转基因玉米植物中表达了发夹rna。随后任选地利用rna印迹杂交在独立的转基因品系中确认靶标基因的dsrna发夹加工成了sirna。
另外,具有与靶标基因存在80%以上序列同一性的错配序列的rnai分子影响玉米根虫的方式类似于使用与靶标基因具有100%序列同一性的rnai分子时所见的方式。错配序列与天然序列的配对在同一个rnai构建体中形成发夹dsrna,由此递送能够影响进食中的鞘翅目害虫的生长、发育和生存力的经植物加工的sirna。
在植物内递送对应于靶标基因的dsrna、sirna或mirna,随后被鞘翅目害虫通过进食而摄取,造成鞘翅目害虫中的靶标基因由于rna介导的基因沉默而下调。当靶标基因在发育的一个或多个阶段发挥重要作用时,鞘翅目害虫的生长和/或发育受到影响,并且就wcr、ncr、scr、mcr、黄瓜条根萤叶甲、黄瓜十一星叶甲、南美叶甲和d.u.undecimpunctatamannerheim中的至少一者而言,造成鞘翅目害虫无法成功地侵染、进食、发育和/或造成鞘翅目害虫死亡。然后通过选择靶标基因和成功应用rnai来控制鞘翅目害虫。
转基因rnai品系和非转化玉米的表型比较。选择用于创建发夹dsrna的靶标鞘翅目害虫基因或序列与任何已知的植物基因序列都没有相似性。因此,预期由靶向这些鞘翅目害虫基因或序列的构建体产生或活化(系统性)rnai不会对转基因植物产生任何有害影响。然而,将转基因品系的发育和形态特征与非转化植物、以及用没有发夹表达基因的“空”载体转化的那些转基因品系进行比较。比较了植物的根、芽、叶和生殖特征。记录植物的芽特征,诸如高度、叶片数和大小、开花时间、花的大小和外观。一般说来,当在体外和在温室土壤中培养时,在转基因品系和未表达靶标irna分子的那些品系之间没有可观察到的形态差异。
实施例10:包含鞘翅目害虫序列和额外的rnai构建体的转基因玉米
转基因玉米植物在其基因组中包含异源性编码序列,该异源性编码序列被转录成靶向鞘翅目害虫之外的生物体的irna分子,对这样的转基因玉米植物经由土壤杆菌或whiskerstm方法(参见petolino和arnold,(2009)methodsmol.biol.526:59-67)进行二次转化,以产生一种或多种杀虫dsrna分子(例如,至少一种dsrna分子,包括靶向包含seqidno:1和/或seqidno:3的基因的dsrna分子)。大体上如实施例4中所述制备植物转化质粒载体,经由土壤杆菌或whiskerstm介导的转化方法递送到获自转基因hiii或b104玉米植物的玉蜀黍悬浮细胞或未成熟玉蜀黍胚中,所述玉米植物在其基因组中包含异源性编码序列,所述异源性编码序列被转录成靶向鞘翅目害虫之外的生物体的irna分子。
实施例11:包含rnai构建体和额外的鞘翅目害虫控制序列的转基因玉米
转基因玉米植物在其基因组中包含被转录成靶向鞘翅目害虫生物体的irna分子的异源性编码序列(例如,至少一种dsrna分子,包括靶向包含seqidno:1和/或seqidno:3的基因的dsrna分子),对这样的转基因玉米植物经由土壤杆菌或whiskerstm方法(参见petolino和arnold,(2009)methodsmol.biol.526:59-67)进行二次转化,以产生一种或多种杀虫蛋白分子,例如cry3、cry6、cry34和cry35杀虫蛋白。大体上如实施例4中所述制备植物转化质粒载体,经由土壤杆菌或whiskerstm介导的转化方法递送到获自转基因b104玉米植物的玉蜀黍悬浮细胞或未成熟玉蜀黍胚中,所述玉米植物在其基因组中包含异源性编码序列,所述异源性编码序列被转录成靶向鞘翅目害虫生物体的irna分子。获得产生用于控制鞘翅目害虫的irna分子和杀虫蛋白的双重转化植物。
实施例12:筛选新热带区棕椿象(英雄美洲蝽)中的
候选靶标基因
新热带区棕椿象(bsb;英雄美洲蝽)集落。bsb在27℃的培养箱中,在65%相对湿度和16小时:8小时的光照:黑暗周期下饲养。将历经2至3天收集的1克卵接种在底部有滤纸盘的5l容器中,用18目网覆盖容器以便通风。每个饲养容器产生约300至400条bsb成虫。在所有阶段,每周向昆虫饲喂新鲜青豆三次,每周更换一次装有向日葵种子、大豆和花生(重量比3:1:1)的种子混合物的小袋。在小瓶中补充水,用棉塞作为捻子。在最初的两周后,每周一次将昆虫转移到新的容器中。
bsb人工食料。如下制备bsb人工食料。在
bsb转录组的组装。选择六个bsb发育期用于制备mrna文库。从冷冻在-70℃的昆虫提取总rna,然后将其置于
bsbrpii33直系同源物鉴定。使用查询序列果蝇属rpii-33(蛋白质序列genbank登录号abi30983)执行bsb汇集的转录组的tblastn搜索。bsbrpii33-1(seqidno:76)和bsbrpii33-2(seqidno:78)被鉴定为英雄美洲蝽候选靶标基因,其产物具有预测的肽序列,分别为seqidno:77和seqidno:79。
模板制备和dsrna合成。使用
cdna扩增。使用针对rt-pcr的superscriptiiifirst-strandsynthesissystemtm(invitrogen),遵循供应商的推荐方案,从5μg的bsb总rna模板和寡dt引物反转录出cdna。用无核酸酶的水将转录反应的最终体积调节为100μl。
使用如表12中所示的引物来扩增bsb_rpii33-1、bsb_rpii33-2、bsb_rpii33-3。以1μlcdna(如上文)作为模板,通过触地(touch-down)pcr(退火温度从60℃降至50℃,以1℃/循环降低)扩增dna模板。在35个pcr循环期间生成了包含bsb_rpii33-1(seqidno:80)的255bp区段、bsb_rpii33-1v1(seqidno:81)的111bp区段以及bsb_rpii33-2(seqidno:82)的398bp区段的片段。还使用上述程序,利用yfpv2-f(seqidno:90)和yfpv2-r(seqidno:91)引物来扩增301bp阴性对照模板yfpv2(seqidno:89)。bsb_rpii33-1、bsb_rpii33-1v1、bsb_rpii33-2和yfpv2引物在其5’端含有t7噬菌体启动子序列(seqidno:9),因此使得yfpv2和bsb_rpii33dna片段能够用于dsrna转录。
表12.用来扩增示例性rpii33靶标基因和yfp阴性对照基因的编码区部分的引物和引物对。
dsrna合成。利用megascripttmt7rnai试剂盒(ambion),根据制造商的说明,使用2μlpcr产物(如上文)作为模板来合成dsrna。参见图1。在nanodroptm8000分光光度计上将dsrna定量,并在无核酸酶的0.1xte缓冲液(1mmtrishcl,0.1mmedta,ph7.4)中稀释到500ng/μl。
注射dsrna到bsb血腔中。bsb在27℃的培养箱中,在65%相对湿度和16小时:8小时的光照:黑暗光周期下,以集落形式饲养在青豆和种子食料上。用小刷子轻轻操作第二龄若虫(每只重1至1.5mg)以防损伤,并将它们置于冰上的培养皿中,以使昆虫感到寒冷从而固定不动。给每只昆虫注射55.2nl的500ng/μldsrna溶液(即,27.6ngdsrna;18.4至27.6μg/g体重的剂量)。使用配备有由drummond3.5英寸#3-000-203-g/x玻璃毛细管拉制而成的注射针的nanojecttmii注射器(drummondscientific,broomhall,pa)进行注射。把针尖打破,用轻矿物油装填毛细管,然后填充2至3μl的dsrna。将dsrna注射到若虫的腹部中(每次试验每份dsrna注射10只昆虫),在不同的三天重复试验。将经注射的昆虫(每孔5只)转移到装有人工bsb食料小丸并覆盖有pull-n-peeltm盖片(bio-cv-4;bio-serv)的32孔托盘(bio-rt-32饲养托盘(bio-rt-32rearingtray);bio-serv,frenchtown,nj)中。借助带棉捻子且装有1.25ml水的1.5ml微量离心管提供水分。将这些托盘在26.5℃、60%湿度和16小时:8小时的光照:黑暗光周期下温育。在注射7天之后,获取生存力计数和重量。
bsbrpii33为致命的dsrna靶标。如表13和表14中所汇总,在每个平行测定中,将55.2nlbsb_rpii33-1、bsb_rpii33-2和bsb_rpii33-1v1dsrna(500ng/μl)注射到至少10只第二龄bsb若虫(每只1至1.5mg)的血腔中,达到大约18.4-27.6μgdsrna/g昆虫的最终浓度。针对这些dsrna所确定的致死率明显不同于相同量的注射yfpv2dsrna(阴性对照)所见的致死率,p<0.05(学生t检验)
表13.bsb_rpii33dsrna注射到第二龄新热带区棕椿象若虫的血腔中七天之后的结果。
*每种dsrna每次试验注射10只昆虫。
**平均值标准误差。
***使用学生t检验时,与yfpv2对照存在显著差异。(p<0.05)。
表14.bsb_rpii33-1v1dsrna注射到第二龄新热带区棕椿象若虫的血腔中七天之后的结果。
*每种dsrna每次试验注射10只昆虫。
**平均值标准误差。
实施例13:包含半翅目害虫序列的转基因玉米
如实施例4中所述生成10至20株包含核酸的表达载体的转基因t0玉米植物,所述核酸含有seqidno:76和/或seqidno:78的任何部分(例如seqidno:80-82)。获得另外的10至20个表达rnai构建体的发夹dsrna的t1玉米独立品系用于bsb攻击。衍生的发夹dsrna包含seqidno:76和/或seqidno:78或者它们的区段(例如seqidno:80-82)的一部分。这些通过rt-pcr或其他分子分析方法得到确认。来自选择的独立t1品系的总rna制备物任选地用于rt-pcr,其中引物被设计为在每个rnai构建体中的发夹表达盒的接头内含子中结合。另外,用于rnai构建体中每个靶标基因的特异性引物任选地用于扩增在植物内产生sirna所需要的预加工mrna,并用于确认产生了所述预加工的mrna。对于每个靶标基因的期望条带的扩增确认每个转基因玉米植物中表达了发夹rna。随后任选地利用rna印迹杂交在独立的转基因品系中确认靶标基因的dsrna发夹加工成了sirna。
另外,具有与靶标基因存在80%以上序列同一性的错配序列的rnai分子影响半翅目昆虫的方式类似于使用与靶标基因具有100%序列同一性的rnai分子时所见的方式。错配序列与天然序列的配对在同一个rnai构建体中形成发夹dsrna,由此递送能够影响进食中的半翅目害虫的生长、发育和生存力的经植物加工的sirna。
在植物内递送对应于靶标基因的dsrna、sirna、shrna、hprna或mirna,随后被半翅目害虫通过进食而摄取,造成半翅目害虫中的靶标基因由于rna介导的基因沉默而下调。当靶标基因在发育的一个或多个阶段发挥重要作用时,半翅目害虫的生长、发育和/或存活受到影响,并且就英雄美洲蝽、褐臭蝽、稻绿蝽、盖德拟壁蝽、茶翅蝽、chiraviahilare、c.marginatum、椿虫、d.furcatus、edessameditabunda、thyantaperditor、horciasnobilellus、taediastigmosa、neomegalotomusparvus、leptoglossuszonatus、秘鲁棉红蝽、niesthreasidae、豆荚草盲蝽和美国牧草盲蝽中的至少一者而言,造成半翅目害虫无法成功地侵染、进食、发育,并且/或者造成半翅目害虫死亡。然后通过选择靶标基因和成功应用rnai来控制半翅目害虫。
转基因rnai品系和非转化玉米的表型比较。选择用于创建发夹dsrna的靶标半翅目害虫基因或序列与任何已知的植物基因序列都没有相似性。因此,预期由靶向这些半翅目害虫基因或序列的构建体产生或活化(系统性)rnai不会对转基因植物产生任何有害影响。然而,将转基因品系的发育和形态特征与非转化植物、以及用没有发夹表达基因的“空”载体转化的那些转基因品系进行比较。比较了植物的根、芽、叶和生殖特征。记录植物的芽特征,诸如高度、叶片数和大小、开花时间、花的大小和外观。一般说来,当在体外和在温室土壤中培养时,在转基因品系和未表达靶标irna分子的那些品系之间没有可观察到的形态差异。
实施例14:包含半翅目害虫序列的转基因大豆
生成10至20株包含核酸的表达载体的转基因t0大豆植物,所述核酸含有seqidno:76和/或seqidno:78或它们的区段(例如seqidno:80-82)的一部分,所述植物按照本领域已知的方式生成,例如通过如下所述的土壤杆菌介导的转化。用氯气将成熟大豆(glycinemax)种子消毒十六小时过夜。用氯气消毒之后,将种子置于laminartm层流罩内的开放容器中以驱散氯气。接着,使用黑盒,在黑暗中24℃下使消过毒的种子吸收无菌h2o十六小时。
制备分割种子大豆。包含部分胚轴的大豆种子的分割方案需要制备纵向切开的大豆种子材料,纵向切开使用附连在解剖刀上的10号刀片,沿种子的种脐分离并除去种皮,然后把种子分割为两个子叶部分。小心地移除部分胚轴,其中约1/2–1/3的胚轴保持附着于子叶的节端。
接种。然后将包含部分胚轴的分割大豆种子置于根癌土壤杆菌(例如,菌株eha101或eha105)的溶液中浸没约30分钟,所述根癌土壤杆菌携带包含seqidno:76和/或seqidno:78和/或它们的区段(例如seqidno:80-82)的二元质粒。在浸入带胚轴的子叶之前,将根癌土壤杆菌溶液稀释到λ=0.6od650的最终浓度。
共培养。接种后,让分割的大豆种子与根癌土壤杆菌菌株在共培养培养基(agrobacteriumprotocols,第2卷,第2版,wang,k.(编辑),humanapress,newjersey,2006年)上共培养5天,共培养培养基置于培养皿中,培养皿用一片滤纸覆盖。
芽诱导。在共培养5天之后,将分割的大豆种子置于由b5盐、b5维生素、28mg/l二价铁、38mg/lna2edta、30g/l蔗糖、0.6g/lmes、1.11mg/lbap、100mg/ltimentintm、200mg/l头孢噻肟和50mg/l万古霉素组成的液体芽诱导(si)培养基(ph5.7)中洗涤。然后将分割的大豆种子置于由b5盐、b5维生素、7g/l诺布尔琼脂、28mg/l二价铁、38mg/lna2edta、30g/l蔗糖、0.6g/lmes、1.11mg/lbap、50mg/ltimentintm、200mg/l头孢噻肟和50mg/l万古霉素组成的芽诱导i(sii)培养基(ph5.7)上培养,其中子叶的平坦一侧面朝上,而子叶的节端埋入培养基中。在培养2周之后,将来自经转化的分割大豆种子的外植体转移到芽诱导ii(siii)培养基中,该培养基含有补充了6mg/l草铵膦
芽伸长。在siii培养基上培养2周之后,将子叶从外植体去除,通过在子叶的基部切口而切下含有胚轴的齐平的芽垫。将分离自子叶的芽垫转移到芽伸长(se)培养基上。该se培养基由ms盐、28mg/l二价铁、38mg/lna2edta、30g/l蔗糖和0.6g/lmes、50mg/l天冬酰胺、100mg/ll-焦谷氨酸、0.1mg/liaa、0.5mg/lga3、1mg/l玉米素核苷、50mg/ltimentintm、200mg/l头孢噻肟、50mg/l万古霉素、6mg/l草铵膦和7g/l诺布尔琼脂组成,ph为5.7。每2周将培养物转移到新鲜的se培养基上。培养物在convirontm生长室中在24℃下生长,使用18h光周期,光强度为80至90μmol/m2s。
生根。从子叶芽垫发出的伸长芽,通过在子叶芽垫基部切割伸长芽加以分离,并将伸长芽浸入1mg/liba(吲哚3-丁酸)中1至3分钟以促进生根。接着,将伸长芽转移到phyta托盘中的生根培养基(ms盐、b5维生素、28mg/l二价铁、38mg/lna2edta、20g/l蔗糖和0.59g/lmes、50mg/l天冬酰胺、100mg/ll-焦谷氨酸、7g/l诺布尔琼脂,ph5.6)中。
栽培。在convirontm生长室中,在24℃和18小时光周期下培养1至2周后,将已生根的芽转移到带盖的圣代杯内的土壤混合物中,放到convirontm生长室(cmp4030和cmp3244型,controlledenvironmentslimited,winnipeg,manitoba,canada)内,置于长日照条件下(16小时光照/8小时黑暗),光强度为120至150μmol/m2s,温度(22℃)和湿度(40-50%)恒定,以驯化小植物。生根的小植物在圣代杯中驯化数周之后,转移到温室中以便进一步驯化并定植健壮的转基因大豆植物。
获得另外的10至20个表达rnai构建体的发夹dsrna的t1大豆独立品系用于bsb攻击。衍生的发夹dsrna可包含seqidno:76和/或seqidno:78,或者它们的区段(例如seqidno:80-82)。这些通过rt-pcr或如本领域已知的其他分子分析方法得到确认。来自选择的独立t1品系的总rna制备物任选地用于rt-pcr,其中引物被设计为在每个rnai构建体中的发夹表达盒的接头内含子中结合。另外,用于rnai构建体中每个靶标基因的特异性引物任选地用于扩增在植物内产生sirna所需要的预加工mrna,并用于确认产生了所述预加工的mrna。对于每个靶标基因的期望条带的扩增确认每个转基因大豆植物中表达了发夹rna。随后任选地利用rna印迹杂交在独立的转基因品系中确认靶标基因的dsrna发夹加工成了sirna。
具有与靶标基因存在80%以上序列同一性的错配序列的rnai分子影响bsb的方式类似于使用与靶标基因具有100%序列同一性的rnai分子时所见的方式。错配序列与天然序列的配对在同一个rnai构建体中形成发夹dsrna,由此递送能够影响进食中的半翅目害虫的生长、发育和生存力的经植物加工的sirna。
在植物内递送对应于靶标基因的dsrna、sirna、shrna或mirna,随后被半翅目害虫通过进食而摄取,造成半翅目害虫中的靶标基因由于rna介导的基因沉默而下调。当靶标基因在发育的一个或多个阶段发挥重要作用时,半翅目害虫的生长、发育、生存力和进食受到影响,并且就英雄美洲蝽、盖德拟壁蝽、茶翅蝽、稻绿蝽、chinaviahilare、褐美洲蝽、椿虫、dichelopsfurcatus、edessameditabunda、thyantaperditor、chinaviamarginatum、horciasnobilellus、taediastigmosa、neomegalotomusparvus、leptoglossuszonatus、niesthreasidae、秘鲁棉红蝽和美国牧草盲蝽中的至少一者而言,造成半翅目害虫无法成功地侵染、进食、发育,并且/或者造成半翅目害虫死亡。然后通过选择靶标基因和成功应用rnai来控制半翅目害虫。
转基因rnai品系和非转化大豆的表型比较。选择用于创建发夹dsrna的靶标半翅目害虫基因或序列与任何已知的植物基因序列都没有相似性。因此,预期由靶向这些半翅目害虫基因或序列的构建体产生或活化(系统性)rnai不会对转基因植物产生任何有害影响。然而,将转基因品系的发育和形态特征与非转化植物、以及用没有发夹表达基因的“空”载体转化的那些转基因品系进行比较。比较了植物的根、芽、叶和生殖特征。记录植物的芽特征,诸如高度、叶片数和大小、开花时间、花的大小和外观。一般说来,当在体外和在温室土壤中培养时,在转基因品系和未表达靶标irna分子的那些品系之间没有可观察到的形态差异。
实施例15:以人工食料为食的英雄美洲蝽的生物测定。
在使用人工食料的dsrna饲喂测定中,与注射实验(参见实施例12)相同,设置具有一颗约18mg的人工食料小丸和水的32孔托盘。将浓度为200ng/μl的dsrna添加到食物小丸和水样品中,向两个孔中的每一个添加100μl。向每个孔中加入5只第二龄的英雄美洲蝽若虫。将水样品和靶向yfp转录物的dsrna用作阴性对照。在三个不同的日期重复实验。对存活昆虫称重,并在处理8天后测定死亡率。在具有rpii33dsrna的孔内相比于对照孔观察到显著的死亡和/或生长抑制。
实施例16:包含半翅目害虫序列的转基因拟南芥(arabidopsisthaliana)
使用与实施例4相似的标准分子方法生成含有用于形成发夹的靶标基因构建体的拟南芥属(arabidopsis)转化载体,该靶标基因构建体包含rpii33的区段(seqidno:76或seqidno:78)。使用标准的基于土壤杆菌的方法执行拟南芥属转化。用草铵膦耐受性可选择标记来选择t1种子。生成转基因t1拟南芥属植物,并生成纯合的单拷贝t2转基因植物用于昆虫研究。对具有花序的生长中的拟南芥属植物执行生物测定。取五至十只昆虫放置在每株植物上,并在14天内监测存活情况。
构建拟南芥属转化载体。使用化学合成片段(dna2.0,menlopark,ca)的组合,以及标准的分子克隆方法,组装基于入门载体的入门克隆,这些入门克隆含有用于形成发夹的靶标基因构建体,该靶标基因构建体包含rpii33的区段(seqidno:76或seqidno:78)。通过(在单个转录单位内)以相反的取向排布靶标基因区段的两个拷贝来易化rna初级转录物形成分子内发夹,所述两个区段被一个接头序列(seqidno:107)分开。因此,初级mrna转录物含有被所述接头序列分开的两个rpii33基因区段序列,这两个区段序列互为大的反向重复序列。使用启动子(例如,拟南芥遍在蛋白10启动子(callis等人,(1990)j.biologicalchem.265:12486-12493))的拷贝来驱动初级mrna发夹转录物的产生,并使用包含来自根癌土壤杆菌开放阅读框23的3’非翻译区(atuorf233'utrv1;美国专利号5,428,147)的片段来终止表达发夹rna的基因的转录。
将入门载体内的发夹克隆用于标准的
二元目的载体包含在木薯叶脉花叶病毒启动子(csvmv启动子v2,美国专利号7,601,885;verdaguer等人,(1996)plantmol.biol.31:1129-39)调节下的除草剂耐受性基因dsm-2v2(美国专利公布号2011/0107455)。使用包含来自根癌土壤杆菌开放阅读框1的3’非翻译区(atuorf13'utrv6;huang等人,(1990)j.bacteriol.172:1814-22)的片段来终止dsm2v2mrna的转录。
借助使用典型的二元目标载体与入门载体的标准
产生包含杀虫rna的转基因拟南芥属:土壤杆菌介导的转化。将含有发夹dsrna序列的二元质粒电穿孔到土壤杆菌菌株gv3101(pmp90rk)中。通过对重组土壤杆菌菌落的质粒制备物的限制性分析确认重组土壤杆菌克隆。使用qiagen质粒大提试剂盒(plasmidmaxkit)(qiagen,目录号12162),遵循制造商推荐的方案从土壤杆菌培养物提取质粒。
拟南芥属转化和t1选择。取12至15株拟南芥属植物(columbia栽培变种)在温室中的4英寸盆中生长,温室中的光强度为250μmol/m2,温度为25℃,采用18小时:6小时的光照:黑暗条件。转化前一周修剪主花茎。通过将10μl重组土壤杆菌甘油储备液置于以225rpm振荡的28℃下的100mllb肉汤(sigmal3022)+100mg/l壮观霉素+50mg/l卡那霉素中温育72小时来制备土壤杆菌接种物。收获土壤杆菌细胞,将其悬浮于5%蔗糖+0.04%silwet-l77(lehleseeds目录号vis-02)+10μg/l苯甲氨基嘌呤(ba)溶液中,至od6000.8~1.0,然后进行花序浸蘸。将植物的地上部分浸入土壤杆菌溶液5至10分钟,轻柔搅动。然后将植物转移到温室中进行正常生长,并定期浇水施肥,直到种子成形。
实施例17:转基因拟南芥属的生长和生物测定
选择用dsrna构建体转化的t1拟南芥属。将来自每次转化的多达200mg的t1种子置于0.1%琼脂糖溶液中分层。把种子种植在装有5号阳光培养基(sunshinemedia)的发芽托盘(10.5英寸×21英寸×1英寸;t.o.plasticsinc.,clearwater,mn.)中。在种植后6天和9天,选择对280g/ha的
英雄美洲蝽植物饲喂生物测定。为每个构建体选择至少四个低拷贝(1至2个插入)事件、四个中拷贝(2至3个插入)事件和四个高拷贝(≥4个插入)事件。使植物生长至生殖期(植物有花和长角果)。土壤表面覆盖有约50ml体积的白色沙子,以便于鉴别昆虫。将5至10只第二龄的英雄美洲蝽若虫引到每株植物上。植物用直径为3英寸、高16英寸、壁厚0.03英寸的塑料管(产品号484485,visipackfentonmo)覆盖,用尼龙网盖住管子以隔离昆虫。在conviron培养箱中将植物保持在正常的温度、光照和浇水条件下。在14天中,收集昆虫并称重,计算死亡率百分比以及生长抑制(1–处理重量/对照重量)。用表达yfp发夹的植物作为对照。在转基因bsb_rpii33dsrna植物上的若虫进食方面相比于对照植物上的若虫,观察到显著的死亡和/或生长抑制。
生成t2拟南芥属种子和t2生物测定。对于每个构建体,由选择的低拷贝(1至2个插入)事件产生t2种子。如上所述,对植物(纯合的和/或杂合的)进行英雄美洲蝽饲喂生物测定。从纯合子收获t3种子并储存用于将来分析。
实施例18:转化额外的作物物种
用rpii33dsrna转基因转化棉花,来提供对半翅目昆虫的控制,该过程利用了本领域技术人员已知的方法。例如,与以前在美国专利号7,838,733的实施例14、或pct国际专利公布号wo2007/053482的实施例12中所述基本上相同的技术。
实施例19:昆虫管理中的rpii33dsrna
将rpii33dsrna转基因与转基因植物中的其他dsrna分子组合,以提供冗余的rnai靶向和协同的rnai效应。表达靶向rpii33的dsrna的转基因植物包括(例如但不限于)玉米、大豆和棉花,这些转基因植物可用于预防鞘翅目昆虫和半翅目昆虫造成的进食损害。rpii33dsrna转基因还在植物中与苏云金芽孢杆菌杀虫蛋白技术和/或pip-1杀虫多肽结合,以代表昆虫抗性管理基因累加中的新作用模式。当在转基因植物中与靶向昆虫害虫的其他dsrna分子和/或杀虫蛋白结合时,观察到协同的杀虫效应,该效应还减缓了抗性昆虫群体壮大的速度。
序列表
<110>dowagrosciences,llc
fime
narva,kene.
worden,sarahe.
frey,meghan
rangasamy,murugesan
veeramani,balaji
gandra,premchand
lo,wendy
fishilevich,elane
vilcinskas,andreas
knorr,eileen
<120>控制昆虫害虫的rna聚合酶33核酸分子
<130>2971-p12623.1us(76745)
<150>62/133,210
<151>2015-03-13
<160>111
<170>patentin3.5版
<210>1
<211>961
<212>dna
<213>玉米根萤叶甲
<400>1
gccgatgccatacatacgcttaaaacatcgtatctgctcagttctttaattaacactgaa60
gaaaatcgaattataaaatgccctacgctaacacaccgtcagtacaaatttctgaactaa120
ccgatgaaaatgttaagttcgtcgttgaggacacagaccttagcttggcaaacagtctac180
gtcgtgttttcatcgctgaaactccaaccctagcaatcgattgggttcaattcgaagcca240
actccactgtactggcagatgaattccttgcccatcgaattggcttgattccattgattt300
ccgatgaggtagtggacagaatccaaaacactcgtgaatgttcatgcttggacttttgca360
ccgagtgcagtgtggaatttacattggatgtcaaatgcagcgacgaacatacgcgccacg420
ttaccacggccgatttaaagtccagtgacgcacgagtgctaccagttacgtccagacatc480
gcgatgacgaggacaacgaatatggagagacgaacgatgaaattctgatcatcaaactgc540
gcaaaggtcaagagctgaagttgcgagcatacgcgaaaaagggtttcggcaaggaacatg600
ccaaatggaatccaacggctggcgttagctttgaatacgatccagtcaattcgatgagac660
ataccctgtacccgaagccggacgaatggccgaaaagtgagcacaccgaacttgacgatg720
atcaatacgaagctgaatataactgggaggctaagccgaacaagtttttcttcaacgttg780
agtcgagtggtgcacttcgaccggaaaacattgtgctgatgggagtcaaagttttgaaaa840
acaaattgtccaatctacagacgcagttaagtcacgaattgactacaaacgatgcgctcg900
tgattcagtaaaagcagcgatcccattgaatttcttcaaaatcttgtttttttcctctaa960
g961
<210>2
<211>277
<212>prt
<213>玉米根萤叶甲
<400>2
metprotyralaasnthrproservalglnilesergluleuthrasp
151015
gluasnvallysphevalvalgluaspthraspleuserleualaasn
202530
serleuargargvalpheilealagluthrprothrleualaileasp
354045
trpvalglnpheglualaasnserthrvalleualaaspglupheleu
505560
alahisargileglyleuileproleuileseraspgluvalvalasp
65707580
argileglnasnthrargglucyssercysleuaspphecysthrglu
859095
cysservalgluphethrleuaspvallyscysseraspgluhisthr
100105110
arghisvalthrthralaaspleulysserseraspalaargvalleu
115120125
provalthrserarghisargaspaspgluaspasnglutyrglyglu
130135140
thrasnaspgluileleuileilelysleuarglysglyglngluleu
145150155160
lysleuargalatyralalyslysglypheglylysgluhisalalys
165170175
trpasnprothralaglyvalserpheglutyraspprovalasnser
180185190
metarghisthrleutyrprolysproaspglutrpprolysserglu
195200205
histhrgluleuaspaspaspglntyrglualaglutyrasntrpglu
210215220
alalysproasnlysphephepheasnvalgluserserglyalaleu
225230235240
argprogluasnilevalleumetglyvallysvalleulysasnlys
245250255
leuserasnleuglnthrglnleuserhisgluleuthrthrasnasp
260265270
alaleuvalilegln
275
<210>3
<211>1110
<212>dna
<213>玉米根萤叶甲
<400>3
cgttgacactgttgacagtgacagttgaaattgaaaaccggattagagaagttttcttgg60
aaagttgtttttttaaataactaacattaaatagaagttatttgtttaagggtttaatat120
gccatatgcaaatcagccatcagttcatataacagatttaacagatgataattgcaaatt180
ttatatagaagacactgatttaagtgttgcgaatagcattcgccgcgtccttattgcaga240
aactcctactctagctatagactgggtaaaattagaagctaactcaactgttctcagtga300
tgaatttttagcacaccgaattggattgataccattagtttccgatgaagttgtacaaag360
attacaatatcctagggactgcgtatgtctcgatttttgtcaagaatgcagtgttgaatt420
tactttagatgtaaaatgtacagatgatcaaactcgacatgtaacaactgccgattttaa480
atctagtgatccacgagtcataccagctacttccaaacatcgtgatgatgaatcctcaga540
gtatggtgaaacagatgaaattcttattattaaactgcgaaagggtcaagagcttaaagt600
taaagcgtatgccaaaaaaggctttggaaaagagcatgccaaatggaatcctacatgtgg660
tgttgcctttgaatatgatcctgataacgctatgagacatacattatttcctaaaccaga720
cgaatggcctaaaagtgaatacagcgaattagaagatgatcagtatgaagctccatataa780
ctgggaattaaaacctaataaattcttctacaatgtggaggctgctggattgttgaaacc840
agaaaatattgtcatcatgggtgtagctatgttaaaagaaaaactgtcaaatttgcaaac900
acaactcagccacgaactaacacctgatgttttggccattccaatttaagaagttaatta960
caatcataggtagagttcattcaaccacagttatacattttttttataatagataagtaa1020
gttttacactataggaacaatttttgacatgttgactaaagatcttgttcaaatagacta1080
gaaataaaattttgaatccaaaaaaaaaaa1110
<210>4
<211>276
<212>prt
<213>玉米根萤叶甲
<400>4
metprotyralaasnglnproservalhisilethraspleuthrasp
151015
aspasncyslysphetyrilegluaspthraspleuservalalaasn
202530
serileargargvalleuilealagluthrprothrleualaileasp
354045
trpvallysleuglualaasnserthrvalleuseraspglupheleu
505560
alahisargileglyleuileproleuvalseraspgluvalvalgln
65707580
argleuglntyrproargaspcysvalcysleuaspphecysglnglu
859095
cysservalgluphethrleuaspvallyscysthraspaspglnthr
100105110
arghisvalthrthralaaspphelysserseraspproargvalile
115120125
proalathrserlyshisargaspaspgluserserglutyrglyglu
130135140
thraspgluileleuileilelysleuarglysglyglngluleulys
145150155160
vallysalatyralalyslysglypheglylysgluhisalalystrp
165170175
asnprothrcysglyvalalapheglutyraspproaspasnalamet
180185190
arghisthrleupheprolysproaspglutrpprolysserglutyr
195200205
sergluleugluaspaspglntyrglualaprotyrasntrpgluleu
210215220
lysproasnlysphephetyrasnvalglualaalaglyleuleulys
225230235240
progluasnilevalilemetglyvalalametleulysglulysleu
245250255
serasnleuglnthrglnleuserhisgluleuthrproaspvalleu
260265270
alaileproile
275
<210>5
<211>483
<212>dna
<213>玉米根萤叶甲
<400>5
gaattccttgcccatcgaattggcttgattccattgatttccgatgaggtagtggacaga60
atccaaaacactcgtgaatgttcatgcttggacttttgcaccgagtgcagtgtggaattt120
acattggatgtcaaatgcagcgacgaacatacgcgccacgttaccacggccgatttaaag180
tccagtgacgcacgagtgctaccagttacgtccagacatcgcgatgacgaggacaacgaa240
tatggagagacgaacgatgaaattctgatcatcaaactgcgcaaaggtcaagagctgaag300
ttgcgagcatacgcgaaaaagggtttcggcaaggaacatgccaaatggaatccaacggct360
ggcgttagctttgaatacgatccagtcaattcgatgagacataccctgtacccgaagccg420
gacgaatggccgaaaagtgagcacaccgaacttgacgatgatcaatacgaagctgaatat480
aac483
<210>6
<211>496
<212>dna
<213>玉米根萤叶甲
<400>6
gttctcagtgatgaatttttagcacaccgaattggattgataccattagtttccgatgaa60
gttgtacaaagattacaatatcctagggactgcgtatgtctcgatttttgtcaagaatgc120
agtgttgaatttactttagatgtaaaatgtacagatgatcaaactcgacatgtaacaact180
gccgattttaaatctagtgatccacgagtcataccagctacttccaaacatcgtgatgat240
gaatcctcagagtatggtgaaacagatgaaattcttattattaaactgcgaaagggtcaa300
gagcttaaagttaaagcgtatgccaaaaaaggctttggaaaagagcatgccaaatggaat360
cctacatgtggtgttgcctttgaatatgatcctgataacgctatgagacatacattattt420
cctaaaccagacgaatggcctaaaagtgaatacagcgaattagaagatgatcagtatgaa480
gctccatataactggg496
<210>7
<211>132
<212>dna
<213>玉米根萤叶甲
<400>7
ctttagatgtaaaatgtacagatgatcaaactcgacatgtaacaactgccgattttaaat60
ctagtgatccacgagtcataccagctacttccaaacatcgtgatgatgaatcctcagagt120
atggtgaaacag132
<210>8
<211>125
<212>dna
<213>玉米根萤叶甲
<400>8
gcgtatgccaaaaaaggctttggaaaagagcatgccaaatggaatcctacatgtggtgtt60
gcctttgaatatgatcctgataacgctatgagacatacattatttcctaaaccagacgaa120
tggcc125
<210>9
<211>24
<212>dna
<213>人工序列
<220>
<223>合成的启动子寡核苷酸
<400>9
ttaatacgactcactatagggaga24
<210>10
<211>503
<212>dna
<213>人工序列
<220>
<223>合成的部分编码区
<400>10
caccatgggctccagcggcgccctgctgttccacggcaagatcccctacgtggtggagat60
ggagggcaatgtggatggccacaccttcagcatccgcggcaagggctacggcgatgccag120
cgtgggcaaggtggatgcccagttcatctgcaccaccggcgatgtgcccgtgccctggag180
caccctggtgaccaccctgacctacggcgcccagtgcttcgccaagtacggccccgagct240
gaaggatttctacaagagctgcatgcccgatggctacgtgcaggagcgcaccatcacctt300
cgagggcgatggcaatttcaagacccgcgccgaggtgaccttcgagaatggcagcgtgta360
caatcgcgtgaagctgaatggccagggcttcaagaaggatggccacgtgctgggcaagaa420
tctggagttcaatttcaccccccactgcctgtacatctggggcgatcaggccaatcacgg480
cctgaagagcgccttcaagatct503
<210>11
<211>46
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>11
ttaatacgactcactatagggagagaattccttgcccatcgaattg46
<210>12
<211>48
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>12
ttaatacgactcactatagggagagttatattcagcttcgtattgatc48
<210>13
<211>49
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>13
ttaatacgactcactatagggagagttctcagtgatgaatttttagcac49
<210>14
<211>49
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>14
ttaatacgactcactatagggagacccagttatatggagcttcatactg49
<210>15
<211>48
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>15
ttaatacgactcactatagggagactttagatgtaaaatgtacagatg48
<210>16
<211>44
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>16
ttaatacgactcactatagggagactgtttcaccatactctgag44
<210>17
<211>46
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>17
ttaatacgactcactatagggagagcgtatgccaaaaaaggctttg46
<210>18
<211>44
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>18
ttaatacgactcactatagggagaggccattcgtctggtttagg44
<210>19
<211>705
<212>dna
<213>人工序列
<220>
<223>合成的人工序列
<400>19
atgtcatctggagcacttctctttcatgggaagattccttacgttgtggagatggaaggg60
aatgttgatggccacacctttagcatacgtgggaaaggctacggagatgcctcagtggga120
aaggttgatgcacagttcatctgcacaactggtgatgttcctgtgccttggagcacactt180
gtcaccactctcacctatggagcacagtgctttgccaagtatggtccagagttgaaggac240
ttctacaagtcctgtatgccagatggctatgtgcaagagcgcacaatcacctttgaagga300
gatggcaacttcaagactagggctgaagtcacctttgagaatgggtctgtctacaatagg360
gtcaaactcaatggtcaaggcttcaagaaagatggtcatgtgttgggaaagaacttggag420
ttcaacttcactccccactgcctctacatctggggtgaccaagccaaccacggtctcaag480
tcagccttcaagatctgtcatgagattactggcagcaaaggcgacttcatagtggctgac540
cacacccagatgaacactcccattggtggaggtccagttcatgttccagagtatcatcac600
atgtcttaccatgtgaaactttccaaagatgtgacagaccacagagacaacatgtccttg660
aaagaaactgtcagagctgttgactgtcgcaagacctacctttga705
<210>20
<211>218
<212>dna
<213>玉米根萤叶甲
<400>20
tagctctgatgacagagcccatcgagtttcaagccaaacagttgcataaagctatcagcg60
gattgggaactgatgaaagtacaatmgtmgaaattttaagtgtmcacaacaacgatgaga120
ttataagaatttcccaggcctatgaaggattgtaccaacgmtcattggaatctgatatca180
aaggagatacctcaggaacattaaaaaagaattattag218
<210>21
<211>424
<212>dna
<213>玉米根萤叶甲
<220>
<221>misc_feature
<222>(393)..(393)
<223>n为a、c、g或t
<220>
<221>misc_feature
<222>(394)..(394)
<223>n为a、c、g或t
<220>
<221>misc_feature
<222>(395)..(395)
<223>n为a、c、g或t
<400>21
ttgttacaagctggagaacttctctttgctggaaccgaagagtcagtatttaatgctgta60
ttctgtcaaagaaataaaccacaattgaatttgatattcgacaaatatgaagaaattgtt120
gggcatcccattgaaaaagccattgaaaacgagttttcaggaaatgctaaacaagccatg180
ttacaccttatccagagcgtaagagatcaagttgcatatttggtaaccaggctgcatgat240
tcaatggcaggcgtcggtactgacgatagaactttaatcagaattgttgtttcgagatct300
gaaatcgatctagaggaaatcaaacaatgctatgaagaaatctacagtaaaaccttggct360
gataggatagcggatgacacatctggcgactannnaaaagccttattagccgttgttggt420
taag424
<210>22
<211>397
<212>dna
<213>玉米根萤叶甲
<400>22
agatgttggctgcatctagagaattacacaagttcttccatgattgcaaggatgtactga60
gcagaatagtggaaaaacaggtatccatgtctgatgaattgggaagggacgcaggagctg120
tcaatgcccttcaacgcaaacaccagaacttcctccaagacctacaaacactccaatcga180
acgtccaacaaatccaagaagaatcagctaaacttcaagctagctatgccggtgatagag240
ctaaagaaatcaccaacagggagcaggaagtggtagcagcctgggcagccttgcagatcg300
cttgcgatcagagacacggaaaattgagcgatactggtgatctattcaaattctttaact360
tggtacgaacgttgatgcagtggatggacgaatggac397
<210>23
<211>490
<212>dna
<213>玉米根萤叶甲
<400>23
gcagatgaacaccagcgagaaaccaagagatgttagtggtgttgaattgttgatgaacaa60
ccatcagacactcaaggctgagatcgaagccagagaagacaactttacggcttgtatttc120
tttaggaaaggaattgttgagccgtaatcactatgctagtgctgatattaaggataaatt180
ggtcgcgttgacgaatcaaaggaatgctgtactacagaggtgggaagaaagatgggagaa240
cttgcaactcatcctcgaggtataccaattcgccagagatgcggccgtcgccgaagcatg300
gttgatcgcacaagaaccttacttgatgagccaagaactaggacacaccattgacgacgt360
tgaaaacttgataaagaaacacgaagcgttcgaaaaatcggcagcggcgcaagaagagag420
attcagtgctttggagagactgacgacgttcgaattgagagaaataaagaggaaacaaga480
agctgcccag490
<210>24
<211>330
<212>dna
<213>玉米根萤叶甲
<400>24
agtgaaatgttagcaaatataacatccaagtttcgtaattgtacttgctcagttagaaaa60
tattctgtagtttcactatcttcaaccgaaaatagaataaatgtagaacctcgcgaactt120
gcctttcctccaaaatatcaagaacctcgacaagtttggttggagagtttagatacgata180
gacgacaaaaaattgggtattcttgagctgcatcctgatgtttttgctactaatccaaga240
atagatattatacatcaaaatgttagatggcaaagtttatatagatatgtaagctatgct300
catacaaagtcaagatttgaagtgagaggt330
<210>25
<211>320
<212>dna
<213>玉米根萤叶甲
<400>25
caaagtcaagatttgaagtgagaggtggaggtcgaaaaccgtggccgcaaaagggattgg60
gacgtgctcgacatggttcaattagaagtccactttggagaggtggaggagttgttcatg120
gaccaaaatctccaacccctcatttttacatgattccattctacacccgtttgctgggtt180
tgactagcgcactttcagtaaaatttgcccaagatgacttgcacgttgtggatagtctag240
atctgccaactgacgaacaaagttatatagaagagctggtcaaaagccgcttttgggggt300
ccttcttgttttatttgtag320
<210>26
<211>47
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>26
ttaatacgactcactatagggagacaccatgggctccagcggcgccc47
<210>27
<211>23
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>27
agatcttgaaggcgctcttcagg23
<210>28
<211>23
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>28
caccatgggctccagcggcgccc23
<210>29
<211>47
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>29
ttaatacgactcactatagggagaagatcttgaaggcgctcttcagg47
<210>30
<211>46
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>30
ttaatacgactcactatagggagagctccaacagtggttccttatc46
<210>31
<211>29
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>31
ctaataattcttttttaatgttcctgagg29
<210>32
<211>22
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>32
gctccaacagtggttccttatc22
<210>33
<211>53
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>33
ttaatacgactcactatagggagactaataattcttttttaatgttcctgagg53
<210>34
<211>48
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>34
ttaatacgactcactatagggagattgttacaagctggagaacttctc48
<210>35
<211>24
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>35
cttaaccaacaacggctaataagg24
<210>36
<211>24
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>36
ttgttacaagctggagaacttctc24
<210>37
<211>48
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>37
ttaatacgactcactatagggagacttaaccaacaacggctaataagg48
<210>38
<211>47
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>38
ttaatacgactcactatagggagaagatgttggctgcatctagagaa47
<210>39
<211>22
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>39
gtccattcgtccatccactgca22
<210>40
<211>23
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>40
agatgttggctgcatctagagaa23
<210>41
<211>46
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>41
ttaatacgactcactatagggagagtccattcgtccatccactgca46
<210>42
<211>46
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>42
ttaatacgactcactatagggagagcagatgaacaccagcgagaaa46
<210>43
<211>22
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>43
ctgggcagcttcttgtttcctc22
<210>44
<211>22
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>44
gcagatgaacaccagcgagaaa22
<210>45
<211>46
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>45
ttaatacgactcactatagggagactgggcagcttcttgtttcctc46
<210>46
<211>51
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>46
ttaatacgactcactatagggagaagtgaaatgttagcaaatataacatcc51
<210>47
<211>26
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>47
acctctcacttcaaatcttgactttg26
<210>48
<211>27
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>48
agtgaaatgttagcaaatataacatcc27
<210>49
<211>50
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>49
ttaatacgactcactatagggagaacctctcacttcaaatcttgactttg50
<210>50
<211>50
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>50
ttaatacgactcactatagggagacaaagtcaagatttgaagtgagaggt50
<210>51
<211>25
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>51
ctacaaataaaacaagaaggacccc25
<210>52
<211>26
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>52
caaagtcaagatttgaagtgagaggt26
<210>53
<211>49
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>53
ttaatacgactcactatagggagactacaaataaaacaagaaggacccc49
<210>54
<211>1150
<212>dna
<213>玉米
<400>54
caacggggcagcactgcactgcactgcaactgcgaatttccgtcagcttggagcggtcca60
agcgccctgcgaagcaaactacgccgatggcttcggcggcggcgtgggagggtccgacgg120
ccgcggagctgaagacagcgggggcggaggtgattcccggcggcgtgcgagtgaaggggt180
gggtcatccagtcccacaaaggccctatcctcaacgccgcctctctgcaacgctttgaag240
atgaacttcaaacaacacatttacctgagatggtttttggagagagtttcttgtcacttc300
aacatacacaaactggcatcaaatttcattttaatgcgcttgatgcactcaaggcatgga360
agaaagaggcactgccacctgttgaggttcctgctgcagcaaaatggaagttcagaagta420
agccttctgaccaggttatacttgactacgactatacatttacgacaccatattgtggga480
gtgatgctgtggttgtgaactctggcactccacaaacaagtttagatggatgcggcactt540
tgtgttgggaggatactaatgatcggattgacattgttgccctttcagcaaaagaaccca600
ttcttttctacgacgaggttatcttgtatgaagatgagttagctgacaatggtatctcat660
ttcttactgtgcgagtgagggtaatgccaactggttggtttctgcttttgcgtttttggc720
ttagagttgatggtgtactgatgaggttgagagacactcggttacattgcctgtttggaa780
acggcgacggagccaagccagtggtacttcgtgagtgctgctggagggaagcaacatttg840
ctactttgtctgcgaaaggatatccttcggactctgcagcgtacgcggacccgaacctta900
ttgcccataagcttcctattgtgacgcagaagacccaaaagctgaaaaatcctacctgac960
tgacacaaaggcgccctaccgcgtgtacatcatgactgtcctgtcctatcgttgcctttt1020
gtgtttgccacatgttgtggatgtacgtttctatgacgaaacaccatagtccatttcgcc1080
tgggccgaacagagatagctgattgtcatgtcacgtttgaattagaccattccttagccc1140
tttttccccc1150
<210>55
<211>22
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<220>
<221>misc_feature
<222>(22)..(22)
<223>n为a、c、g或t
<400>55
ttttttttttttttttttttvn22
<210>56
<211>25
<212>dna
<213>人工序列
<220>
<223>引物rpii33-2v1fwdset2
<400>56
gatcaaactcgacatgtaacaactg25
<210>57
<211>23
<212>dna
<213>人工序列
<220>
<223>引物rpii33-2v1revset2
<400>57
ggattcatcatcacgatgtttgg23
<210>58
<211>21
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>58
tgagggtaatgccaactggtt21
<210>59
<211>24
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>59
gcaatgtaaccgagtgtctctcaa24
<210>60
<211>32
<212>dna
<213>人工序列
<220>
<223>合成的探针寡核苷酸
<400>60
tttttggcttagagttgatggtgtactgatga32
<210>61
<211>151
<212>dna
<213>大肠杆菌
<400>61
gaccgtaaggcttgatgaaacaacgcggcgagctttgatcaacgaccttttggaaacttc60
ggcttcccctggagagagcgagattctccgcgctgtagaagtcaccattgttgtgcacga120
cgacatcattccgtggcgttatccagctaag151
<210>62
<211>69
<212>dna
<213>人工序列
<220>
<223>合成的部分编码区
<400>62
tgttcggttccctctaccaagcacagaaccgtcgcttcagcaacacctcagtcaaggtga60
tggatgttg69
<210>63
<211>4233
<212>dna
<213>玉米
<400>63
agcctggtgtttccggaggagacagacatgatccctgccgttgctgatccgacgacgctg60
gacggcgggggcgcgcgcaggccgttgctcccggagacggaccctcgggggcgtgctgcc120
gccggcgccgagcagaagcggccgccggctacgccgaccgttctcaccgccgtcgtctcc180
gccgtgctcctgctcgtcctcgtggcggtcacagtcctcgcgtcgcagcacgtcgacggg240
caggctgggggcgttcccgcgggcgaagatgccgtcgtcgtcgaggtggccgcctcccgt300
ggcgtggctgagggcgtgtcggagaagtccacggccccgctcctcggctccggcgcgctc360
caggacttctcctggaccaacgcgatgctggcgtggcagcgcacggcgttccacttccag420
ccccccaagaactggatgaacggttagttggacccgtcgccatcggtgacgacgcgcgga480
tcgtttttttcttttttcctctcgttctggctctaacttggttccgcgtttctgtcacgg540
acgcctcgtgcacatggcgatacccgatccgccggccgcgtatatctatctacctcgacc600
ggcttctccagatccgaacggtaagttgttggctccgatacgatcgatcacatgtgagct660
cggcatgctgcttttctgcgcgtgcatgcggctcctagcattccacgtccacgggtcgtg720
acatcaatgcacgatataatcgtatcggtacagagatattgtcccatcagctgctagctt780
tcgcgtattgatgtcgtgacattttgcacgcaggtccgctgtatcacaagggctggtacc840
acctcttctaccagtggaacccggactccgcggtatggggcaacatcacctggggccacg900
ccgtctcgcgcgacctcctccactggctgcacctaccgctggccatggtgcccgatcacc960
cgtacgacgccaacggcgtctggtccgggtcggcgacgcgcctgcccgacggccggatcg1020
tcatgctctacacgggctccacggcggagtcgtcggcgcaggtgcagaacctcgcggagc1080
cggccgacgcgtccgacccgctgctgcgggagtgggtcaagtcggacgccaacccggtgc1140
tggtgccgccgccgggcatcgggccgacggacttccgcgacccgacgacggcgtgtcgga1200
cgccggccggcaacgacacggcgtggcgggtcgccatcgggtccaaggaccgggaccacg1260
cggggctggcgctggtgtaccggacggaggacttcgtgcggtacgacccggcgccggcgc1320
tgatgcacgccgtgccgggcaccggcatgtgggagtgcgtggacttctacccggtggccg1380
cgggatcaggcgccgcggcgggcagcggggacgggctggagacgtccgcggcgccgggac1440
ccggggtgaagcacgtgctcaaggctagcctcgacgacgacaagcacgactactacgcga1500
tcggcacctacgacccggcgacggacacctggacccccgacagcgcggaggacgacgtcg1560
ggatcggcctccggtacgactatggcaagtactacgcgtcgaagaccttctacgaccccg1620
tccttcgccggcgggtgctctgggggtgggtcggcgagaccgacagcgagcgcgcggaca1680
tcctcaagggctgggcatccgtgcaggtacgtctcagggtttgaggctagcatggcttca1740
atcttgctggcatcgaatcattaatgggcagatattataacttgataatctgggttggtt1800
gtgtgtggtggggatggtgacacacgcgcggtaataatgtagctaagctggttaaggatg1860
agtaatggggttgcgtataaacgacagctctgctaccattacttctgacacccgattgaa1920
ggagacaacagtaggggtagccggtagggttcgtcgacttgccttttcttttttcctttg1980
ttttgttgtggatcgtccaacacaaggaaaataggatcatccaacaaacatggaagtaat2040
cccgtaaaacatttctcaaggaaccatctagctagacgagcgtggcatgatccatgcatg2100
cacaaacactagataggtctctgcagctgtgatgttcctttacatataccaccgtccaaa2160
ctgaatccggtctgaaaattgttcaagcagagaggccccgatcctcacacctgtacacgt2220
ccctgtacgcgccgtcgtggtctcccgtgatcctgccccgtcccctccacgcggccacgc2280
ctgctgcagcgctctgtacaagcgtgcaccacgtgagaatttccgtctactcgagcctag2340
tagttagacgggaaaacgagaggaagcgcacggtccaagcacaacactttgcgcgggccc2400
gtgacttgtctccggttggctgagggcgcgcgacagagatgtatggcgccgcggcgtgtc2460
ttgtgtcttgtcttgcctatacaccgtagtcagagactgtgtcaaagccgtccaacgaca2520
atgagctaggaaacgggttggagagctgggttcttgccttgcctcctgtgatgtctttgc2580
cttgcatagggggcgcagtatgtagctttgcgttttacttcacgccaaaggatactgctg2640
atcgtgaattattattattatatatatatcgaatatcgatttcgtcgctctcgtggggtt2700
ttattttccagactcaaacttttcaaaaggcctgtgttttagttcttttcttccaattga2760
gtaggcaaggcgtgtgagtgtgaccaacgcatgcatggatatcgtggtagactggtagag2820
ctgtcgttaccagcgcgatgcttgtatatgtttgcagtattttcaaatgaatgtctcagc2880
tagcgtacagttgaccaagtcgacgtggagggcgcacaacagacctctgacattattcac2940
tttttttttaccatgccgtgcacgtgcagtcaatccccaggacggtcctcctggacacga3000
agacgggcagcaacctgctccagtggccggtggtggaggtggagaacctccggatgagcg3060
gcaagagcttcgacggcgtcgcgctggaccgcggatccgtcgtgcccctcgacgtcggca3120
aggcgacgcaggtgacgccgcacgcagcctgctgcagcgaacgaactcgcgcgttgccgg3180
cccgcggccagctgacttagtttctctggctgatcgaccgtgtgcctgcgtgcgtgcagt3240
tggacatcgaggctgtgttcgaggtggacgcgtcggacgcggcgggcgtcacggaggccg3300
acgtgacgttcaactgcagcaccagcgcaggcgcggcgggccggggcctgctcggcccgt3360
tcggccttctcgtgctggcggacgacgacttgtccgagcagaccgccgtgtacttctacc3420
tgctcaagggcacggacggcagcctccaaactttcttctgccaagacgagctcaggtatg3480
tatgttatgacttatgaccatgcatgcatgcgcatttcttagctaggctgtgaagcttct3540
tgttgagttgtttcacagatgcttaccgtctgctttgtttcgtatttcgactaggcatcc3600
aaggcgaacgatctggttaagagagtatacgggagcttggtccctgtgctagatggggag3660
aatctctcggtcagaatactggtaagtttttacagcgccagccatgcatgtgttggccag3720
ccagctgctggtactttggacactcgttcttctcgcactgctcattattgcttctgatct3780
ggatgcactacaaattgaaggttgaccactccatcgtggagagctttgctcaaggcggga3840
ggacgtgcatcacgtcgcgagtgtaccccacacgagccatctacgactccgcccgcgtct3900
tcctcttcaacaacgccacacatgctcacgtcaaagcaaaatccgtcaagatctggcagc3960
tcaactccgcctacatccggccatatccggcaacgacgacttctctatgactaaattaag4020
tgacggacagataggcgatattgcatacttgcatcatgaactcatttgtacaacagtgat4080
tgtttaatttatttgctgccttccttatccttcttgtgaaactatatggtacacacatgt4140
atcattaggtctagtagtgttgttgcaaagacacttagacaccagaggttccaggagtat4200
cagagataaggtataagagggagcagggagcag4233
<210>64
<211>20
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>64
tgttcggttccctctaccaa20
<210>65
<211>22
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>65
caacatccatcaccttgactga22
<210>66
<211>24
<212>dna
<213>人工序列
<220>
<223>合成的探针寡核苷酸
<400>66
cacagaaccgtcgcttcagcaaca24
<210>67
<211>18
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>67
tggcggacgacgacttgt18
<210>68
<211>19
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>68
aaagtttggaggctgccgt19
<210>69
<211>26
<212>dna
<213>人工序列
<220>
<223>合成的探针寡核苷酸
<400>69
cgagcagaccgccgtgtacttctacc26
<210>70
<211>19
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>70
cttagctggataacgccac19
<210>71
<211>19
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>71
gaccgtaaggcttgatgaa19
<210>72
<211>21
<212>dna
<213>人工序列
<220>
<223>合成的探针寡核苷酸
<400>72
cgagattctccgcgctgtaga21
<210>73
<211>20
<212>dna
<213>人工序列
<220>
<223>引物环-f
<400>73
ggaacgagctgcttgcgtat20
<210>74
<211>20
<212>dna
<213>人工序列
<220>
<223>引物环-r
<400>74
cacggtgcagctgattgatg20
<210>75
<211>18
<212>dna
<213>人工序列
<220>
<223>探针环-p
<400>75
tcccttccgtagtcagag18
<210>76
<211>1368
<212>dna
<213>英雄美洲蝽
<400>76
gttcggctcgggtgagtgtttaaaccaactacgcatcttgttctcgaacctttgcgaaca60
gtgttcacaaataatgctcggttggtgtaaaggtacctttagagcgtgaccccaacttct120
tttgactcaccttgcagaaactcgatcactaacaattacgtgtatataatcgattcacta180
cacgaacgatacatggttgtttaggttacattcatgttatctttagtaatgaagttattg240
agttggcctaattgttgaatgtagttaacagaatgccttatgccaatcaaccttctgttc300
atgtttcagatttaaccgacgacaatgttaaattccaaatagaagatacagaattaagtg360
tcgctaacagcctcagaagagtcttcatagctgaaaccccaactttagctattgattggg420
tgcaattgtctgcaaattctactgttttaagtgatgaatttattgcttctagaatcggac480
ttattcctttaacttctgatgctgcagtcgaaaaattaatctattctagggactgtaatt540
gtactgatttctgcccatcctgtagtgttgagtttactttagatgtcaaatgtgtagatg600
atcaaactagacatgtgacaactgcagatttaaagactgctgatccatgtgtagttcctg660
ctacatctaaaaatagagatgctgatgccaatgaatatggtgaatcagatgatattttga720
ttgttaaattaagaaaaggacaagagcttaaattgagggcctttgctaagaaaggttttg780
gtaaggaacatgctaagtggaatcctactgctggggtttgttttgagtatgaccctgaca840
actcaatgaggcatacactgtttccaaaaccagatgagtggccaaaaagtgaatatactg900
aattagatgaggatcagtatgaagctccatttaattgggaagccaaacctaacaaatttt960
tcttcaatgttgaaagttgtggatctttgcgccccgaaaacatagtattaaaaggagtag1020
aagttctaaaatataaactttctgatttattaattcaattgagtcatgaatcagctggcc1080
aagttgatcatatgcctgtttaaccagtttttgtgataaattattatctgaaataattca1140
attattatatttatattaatgtaaaataaaaagaaatttgataactgaaaaaaaaaaaaa1200
aaaatctattgaaagaatacattcattaatacctttctaaagaaaaattattcaatttaa1260
aattgttgccaaaaagtattcagcatttttttaaaattcaatctaggcatatactactgt1320
aaataaatacaaacaatactttcatttttgtactgttctaaaaattgt1368
<210>77
<211>276
<212>prt
<213>英雄美洲蝽
<400>77
metprotyralaasnglnproservalhisvalseraspleuthrasp
151015
aspasnvallyspheglnilegluaspthrgluleuservalalaasn
202530
serleuargargvalpheilealagluthrprothrleualaileasp
354045
trpvalglnleuseralaasnserthrvalleuseraspglupheile
505560
alaserargileglyleuileproleuthrseraspalaalavalglu
65707580
lysleuiletyrserargaspcysasncysthraspphecysproser
859095
cysservalgluphethrleuaspvallyscysvalaspaspglnthr
100105110
arghisvalthrthralaaspleulysthralaaspprocysvalval
115120125
proalathrserlysasnargaspalaaspalaasnglutyrglyglu
130135140
seraspaspileleuilevallysleuarglysglyglngluleulys
145150155160
leuargalaphealalyslysglypheglylysgluhisalalystrp
165170175
asnprothralaglyvalcyspheglutyraspproaspasnsermet
180185190
arghisthrleupheprolysproaspglutrpprolysserglutyr
195200205
thrgluleuaspgluaspglntyrglualapropheasntrpgluala
210215220
lysproasnlysphephepheasnvalglusercysglyserleuarg
225230235240
progluasnilevalleulysglyvalgluvalleulystyrlysleu
245250255
seraspleuleuileglnleuserhisgluseralaglyglnvalasp
260265270
hismetproval
275
<210>78
<211>758
<212>dna
<213>英雄美洲蝽
<400>78
tgtaaaacttgttctttaagatctcaagaccttttattagaacatctacaggcttaagag60
agccctctacaacttctacgtccatgtgcaccgtgtctatttcacaaaggagatctggtt120
cttcctcctcaaccatcggccagtccttcttaagcgtatcttctgtccagtagtttgtgg180
acctagtcttattggttctatcatactcgaacccgacaacagagacaggagaccacttgg240
catgcatcctccctatccccttcctagcaatacacctaattttcaggctttgattcttcc300
caagttttgcaattaccggtgtgctttttataaaagtctcgtcactgtcaaattttatgt360
ctttacaagtcacgttaaggggggtctctgaggtgttgctaacatcaagttccatctcta420
cggaacaacgagagcaaagctcatcacagtcacactcttctttatacacaagctctttct480
ttgagtacattgggataagcccaagggactgtgccaatacttcatcggggaggaccgtgt540
tgtttttgatgatttcgacgagatctattgcgatagtaggtacttcagataagaggattc600
tccttagagcattagcataggagactgtaatcccagtgagagtgaatttgatgtgttcgt660
cgttttgttcgtgaattgtaattttcatgagaaagctggagggcaaaagaaatgaagtaa720
atttagaagggaacacctgtgaagtatgatcgactacg758
<210>79
<211>229
<212>prt
<213>英雄美洲蝽
<400>79
metlysilethrilehisgluglnasnaspgluhisilelysphethr
151015
leuthrglyilethrvalsertyralaasnalaleuargargileleu
202530
leusergluvalprothrilealaileaspleuvalgluileilelys
354045
asnasnthrvalleuproaspgluvalleualaglnserleuglyleu
505560
ilepromettyrserlyslysgluleuvaltyrlysgluglucysasp
65707580
cysaspgluleucysserargcysservalglumetgluleuaspval
859095
serasnthrsergluthrproleuasnvalthrcyslysaspilelys
100105110
pheaspseraspgluthrpheilelysserthrprovalilealalys
115120125
leuglylysasnglnserleulysileargcysilealaarglysgly
130135140
ileglyargmethisalalystrpserprovalservalvalglyphe
145150155160
glutyraspargthrasnlysthrargserthrasntyrtrpthrglu
165170175
aspthrleulyslysasptrpprometvalglugluglugluproasp
180185190
leuleucysgluileaspthrvalhismetaspvalgluvalvalglu
195200205
glyserleulysprovalaspvalleuilelysglyleugluileleu
210215220
lysasnlysphetyr
225
<210>80
<211>255
<212>dna
<213>英雄美洲蝽
<400>80
ggtgaatcagatgatattttgattgttaaattaagaaaaggacaagagcttaaattgagg60
gcctttgctaagaaaggttttggtaaggaacatgctaagtggaatcctactgctggggtt120
tgttttgagtatgaccctgacaactcaatgaggcatacactgtttccaaaaccagatgag180
tggccaaaaagtgaatatactgaattagatgaggatcagtatgaagctccatttaattgg240
gaagccaaacctaac255
<210>81
<211>111
<212>dna
<213>英雄美洲蝽
<400>81
ttgttttgagtatgaccctgacaactcaatgaggcatacactgtttccaaaaccagatga60
gtggccaaaaagtgaatatactgaattagatgaggatcagtatgaagctcc111
<210>82
<211>398
<212>dna
<213>英雄美洲蝽
<400>82
cgtcgaaatcatcaaaaacaacacggtcctccccgatgaagtattggcacagtcccttgg60
gcttatcccaatgtactcaaagaaagagcttgtgtataaagaagagtgtgactgtgatga120
gctttgctctcgttgttccgtagagatggaacttgatgttagcaacacctcagagacccc180
ccttaacgtgacttgtaaagacataaaatttgacagtgacgagacttttataaaaagcac240
accggtaattgcaaaacttgggaagaatcaaagcctgaaaattaggtgtattgctaggaa300
ggggatagggaggatgcatgccaagtggtctcctgtctctgttgtcgggttcgagtatga360
tagaaccaataagactaggtccacaaactactggacag398
<210>83
<211>49
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>83
ttaatacgactcactatagggagaggtgaatcagatgatattttgattg49
<210>84
<211>50
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>84
ttaatacgactcactatagggagagttaggtttggcttcccaattaaatg50
<210>85
<211>49
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>85
ttaatacgactcactatagggagattgttttgagtatgaccctgacaac49
<210>86
<211>53
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>86
ttaatacgactcactatagggagaggagcttcatactgatcctcatctaattc53
<210>87
<211>49
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>87
ttaatacgactcactatagggagacgtcgaaatcatcaaaaacaacacg49
<210>88
<211>48
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>88
ttaatacgactcactatagggagactgtccagtagtttgtggacctag48
<210>89
<211>301
<212>dna
<213>人工序列
<220>
<223>合成的人工序列
<400>89
catctggagcacttctctttcatgggaagattccttacgttgtggagatggaagggaatg60
ttgatggccacacctttagcatacgtgggaaaggctacggagatgcctcagtgggaaagg120
ttgatgcacagttcatctgcacaactggtgatgttcctgtgccttggagcacacttgtca180
ccactctcacctatggagcacagtgctttgccaagtatggtccagagttgaaggacttct240
acaagtcctgtatgccagatggctatgtgcaagagcgcacaatcacctttgaaggagatg300
g301
<210>90
<211>47
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>90
ttaatacgactcactatagggagagcatctggagcacttctctttca47
<210>91
<211>46
<212>dna
<213>人工序列
<220>
<223>合成的引物寡核苷酸
<400>91
ttaatacgactcactatagggagaccatctccttcaaaggtgattg46
<210>92
<211>961
<212>rna
<213>玉米根萤叶甲
<400>92
gccgaugccauacauacgcuuaaaacaucguaucugcucaguucuuuaauuaacacugaa60
gaaaaucgaauuauaaaaugcccuacgcuaacacaccgucaguacaaauuucugaacuaa120
ccgaugaaaauguuaaguucgucguugaggacacagaccuuagcuuggcaaacagucuac180
gucguguuuucaucgcugaaacuccaacccuagcaaucgauuggguucaauucgaagcca240
acuccacuguacuggcagaugaauuccuugcccaucgaauuggcuugauuccauugauuu300
ccgaugagguaguggacagaauccaaaacacucgugaauguucaugcuuggacuuuugca360
ccgagugcaguguggaauuuacauuggaugucaaaugcagcgacgaacauacgcgccacg420
uuaccacggccgauuuaaaguccagugacgcacgagugcuaccaguuacguccagacauc480
gcgaugacgaggacaacgaauauggagagacgaacgaugaaauucugaucaucaaacugc540
gcaaaggucaagagcugaaguugcgagcauacgcgaaaaaggguuucggcaaggaacaug600
ccaaauggaauccaacggcuggcguuagcuuugaauacgauccagucaauucgaugagac660
auacccuguacccgaagccggacgaauggccgaaaagugagcacaccgaacuugacgaug720
aucaauacgaagcugaauauaacugggaggcuaagccgaacaaguuuuucuucaacguug780
agucgaguggugcacuucgaccggaaaacauugugcugaugggagucaaaguuuugaaaa840
acaaauuguccaaucuacagacgcaguuaagucacgaauugacuacaaacgaugcgcucg900
ugauucaguaaaagcagcgaucccauugaauuucuucaaaaucuuguuuuuuuccucuaa960
g961
<210>93
<211>1110
<212>rna
<213>玉米根萤叶甲
<400>93
cguugacacuguugacagugacaguugaaauugaaaaccggauuagagaaguuuucuugg60
aaaguuguuuuuuuaaauaacuaacauuaaauagaaguuauuuguuuaaggguuuaauau120
gccauaugcaaaucagccaucaguucauauaacagauuuaacagaugauaauugcaaauu180
uuauauagaagacacugauuuaaguguugcgaauagcauucgccgcguccuuauugcaga240
aacuccuacucuagcuauagacuggguaaaauuagaagcuaacucaacuguucucaguga300
ugaauuuuuagcacaccgaauuggauugauaccauuaguuuccgaugaaguuguacaaag360
auuacaauauccuagggacugcguaugucucgauuuuugucaagaaugcaguguugaauu420
uacuuuagauguaaaauguacagaugaucaaacucgacauguaacaacugccgauuuuaa480
aucuagugauccacgagucauaccagcuacuuccaaacaucgugaugaugaauccucaga540
guauggugaaacagaugaaauucuuauuauuaaacugcgaaagggucaagagcuuaaagu600
uaaagcguaugccaaaaaaggcuuuggaaaagagcaugccaaauggaauccuacaugugg660
uguugccuuugaauaugauccugauaacgcuaugagacauacauuauuuccuaaaccaga720
cgaauggccuaaaagugaauacagcgaauuagaagaugaucaguaugaagcuccauauaa780
cugggaauuaaaaccuaauaaauucuucuacaauguggaggcugcuggauuguugaaacc840
agaaaauauugucaucauggguguagcuauguuaaaagaaaaacugucaaauuugcaaac900
acaacucagccacgaacuaacaccugauguuuuggccauuccaauuuaagaaguuaauua960
caaucauagguagaguucauucaaccacaguuauacauuuuuuuuauaauagauaaguaa1020
guuuuacacuauaggaacaauuuuugacauguugacuaaagaucuuguucaaauagacua1080
gaaauaaaauuuugaauccaaaaaaaaaaa1110
<210>94
<211>483
<212>rna
<213>玉米根萤叶甲
<400>94
gaauuccuugcccaucgaauuggcuugauuccauugauuuccgaugagguaguggacaga60
auccaaaacacucgugaauguucaugcuuggacuuuugcaccgagugcaguguggaauuu120
acauuggaugucaaaugcagcgacgaacauacgcgccacguuaccacggccgauuuaaag180
uccagugacgcacgagugcuaccaguuacguccagacaucgcgaugacgaggacaacgaa240
uauggagagacgaacgaugaaauucugaucaucaaacugcgcaaaggucaagagcugaag300
uugcgagcauacgcgaaaaaggguuucggcaaggaacaugccaaauggaauccaacggcu360
ggcguuagcuuugaauacgauccagucaauucgaugagacauacccuguacccgaagccg420
gacgaauggccgaaaagugagcacaccgaacuugacgaugaucaauacgaagcugaauau480
aac483
<210>95
<211>496
<212>rna
<213>玉米根萤叶甲
<400>95
guucucagugaugaauuuuuagcacaccgaauuggauugauaccauuaguuuccgaugaa60
guuguacaaagauuacaauauccuagggacugcguaugucucgauuuuugucaagaaugc120
aguguugaauuuacuuuagauguaaaauguacagaugaucaaacucgacauguaacaacu180
gccgauuuuaaaucuagugauccacgagucauaccagcuacuuccaaacaucgugaugau240
gaauccucagaguauggugaaacagaugaaauucuuauuauuaaacugcgaaagggucaa300
gagcuuaaaguuaaagcguaugccaaaaaaggcuuuggaaaagagcaugccaaauggaau360
ccuacaugugguguugccuuugaauaugauccugauaacgcuaugagacauacauuauuu420
ccuaaaccagacgaauggccuaaaagugaauacagcgaauuagaagaugaucaguaugaa480
gcuccauauaacuggg496
<210>96
<211>132
<212>rna
<213>玉米根萤叶甲
<400>96
cuuuagauguaaaauguacagaugaucaaacucgacauguaacaacugccgauuuuaaau60
cuagugauccacgagucauaccagcuacuuccaaacaucgugaugaugaauccucagagu120
auggugaaacag132
<210>97
<211>125
<212>rna
<213>玉米根萤叶甲
<400>97
gcguaugccaaaaaaggcuuuggaaaagagcaugccaaauggaauccuacaugugguguu60
gccuuugaauaugauccugauaacgcuaugagacauacauuauuuccuaaaccagacgaa120
uggcc125
<210>98
<211>1368
<212>rna
<213>英雄美洲蝽
<400>98
guucggcucgggugaguguuuaaaccaacuacgcaucuuguucucgaaccuuugcgaaca60
guguucacaaauaaugcucgguugguguaaagguaccuuuagagcgugaccccaacuucu120
uuugacucaccuugcagaaacucgaucacuaacaauuacguguauauaaucgauucacua180
cacgaacgauacaugguuguuuagguuacauucauguuaucuuuaguaaugaaguuauug240
aguuggccuaauuguugaauguaguuaacagaaugccuuaugccaaucaaccuucuguuc300
auguuucagauuuaaccgacgacaauguuaaauuccaaauagaagauacagaauuaagug360
ucgcuaacagccucagaagagucuucauagcugaaaccccaacuuuagcuauugauuggg420
ugcaauugucugcaaauucuacuguuuuaagugaugaauuuauugcuucuagaaucggac480
uuauuccuuuaacuucugaugcugcagucgaaaaauuaaucuauucuagggacuguaauu540
guacugauuucugcccauccuguaguguugaguuuacuuuagaugucaaauguguagaug600
aucaaacuagacaugugacaacugcagauuuaaagacugcugauccauguguaguuccug660
cuacaucuaaaaauagagaugcugaugccaaugaauauggugaaucagaugauauuuuga720
uuguuaaauuaagaaaaggacaagagcuuaaauugagggccuuugcuaagaaagguuuug780
guaaggaacaugcuaaguggaauccuacugcugggguuuguuuugaguaugacccugaca840
acucaaugaggcauacacuguuuccaaaaccagaugaguggccaaaaagugaauauacug900
aauuagaugaggaucaguaugaagcuccauuuaauugggaagccaaaccuaacaaauuuu960
ucuucaauguugaaaguuguggaucuuugcgccccgaaaacauaguauuaaaaggaguag1020
aaguucuaaaauauaaacuuucugauuuauuaauucaauugagucaugaaucagcuggcc1080
aaguugaucauaugccuguuuaaccaguuuuugugauaaauuauuaucugaaauaauuca1140
auuauuauauuuauauuaauguaaaauaaaaagaaauuugauaacugaaaaaaaaaaaaa1200
aaaaucuauugaaagaauacauucauuaauaccuuucuaaagaaaaauuauucaauuuaa1260
aauuguugccaaaaaguauucagcauuuuuuuaaaauucaaucuaggcauauacuacugu1320
aaauaaauacaaacaauacuuucauuuuuguacuguucuaaaaauugu1368
<210>99
<211>758
<212>rna
<213>英雄美洲蝽
<400>99
uguaaaacuuguucuuuaagaucucaagaccuuuuauuagaacaucuacaggcuuaagag60
agcccucuacaacuucuacguccaugugcaccgugucuauuucacaaaggagaucugguu120
cuuccuccucaaccaucggccaguccuucuuaagcguaucuucuguccaguaguuugugg180
accuagucuuauugguucuaucauacucgaacccgacaacagagacaggagaccacuugg240
caugcauccucccuauccccuuccuagcaauacaccuaauuuucaggcuuugauucuucc300
caaguuuugcaauuaccggugugcuuuuuauaaaagucucgucacugucaaauuuuaugu360
cuuuacaagucacguuaaggggggucucugagguguugcuaacaucaaguuccaucucua420
cggaacaacgagagcaaagcucaucacagucacacucuucuuuauacacaagcucuuucu480
uugaguacauugggauaagcccaagggacugugccaauacuucaucggggaggaccgugu540
uguuuuugaugauuucgacgagaucuauugcgauaguagguacuucagauaagaggauuc600
uccuuagagcauuagcauaggagacuguaaucccagugagagugaauuugauguguucgu660
cguuuuguucgugaauuguaauuuucaugagaaagcuggagggcaaaagaaaugaaguaa720
auuuagaagggaacaccugugaaguaugaucgacuacg758
<210>100
<211>255
<212>rna
<213>英雄美洲蝽
<400>100
ggugaaucagaugauauuuugauuguuaaauuaagaaaaggacaagagcuuaaauugagg60
gccuuugcuaagaaagguuuugguaaggaacaugcuaaguggaauccuacugcugggguu120
uguuuugaguaugacccugacaacucaaugaggcauacacuguuuccaaaaccagaugag180
uggccaaaaagugaauauacugaauuagaugaggaucaguaugaagcuccauuuaauugg240
gaagccaaaccuaac255
<210>101
<211>111
<212>rna
<213>英雄美洲蝽
<400>101
uuguuuugaguaugacccugacaacucaaugaggcauacacuguuuccaaaaccagauga60
guggccaaaaagugaauauacugaauuagaugaggaucaguaugaagcucc111
<210>102
<211>398
<212>rna
<213>英雄美洲蝽
<400>102
cgucgaaaucaucaaaaacaacacgguccuccccgaugaaguauuggcacagucccuugg60
gcuuaucccaauguacucaaagaaagagcuuguguauaaagaagagugugacugugauga120
gcuuugcucucguuguuccguagagauggaacuugauguuagcaacaccucagagacccc180
ccuuaacgugacuuguaaagacauaaaauuugacagugacgagacuuuuauaaaaagcac240
accgguaauugcaaaacuugggaagaaucaaagccugaaaauuagguguauugcuaggaa300
ggggauagggaggaugcaugccaaguggucuccugucucuguugucggguucgaguauga360
uagaaccaauaagacuagguccacaaacuacuggacag398
<210>103
<211>437
<212>dna
<213>人工序列
<220>
<223>编码叶甲属rpii33-2v1dsrna的dna
<400>103
ctttagatgtaaaatgtacagatgatcaaactcgacatgtaacaactgccgattttaaat60
ctagtgatccacgagtcataccagctacttccaaacatcgtgatgatgaatcctcagagt120
atggtgaaacaggaagctagtaccagtcatcacgctggagcgcacatataggccctccat180
cagaaagtcattgtgtatatctctcatagggaacgagctgcttgcgtatttcccttccgt240
agtcagagtcatcaatcagctgcaccgtgtcgtaaagcgggacgttcgcaagctcgtccg300
cggtactgtttcaccatactctgaggattcatcatcacgatgtttggaagtagctggtat360
gactcgtggatcactagatttaaaatcggcagttgttacatgtcgagtttgatcatctgt420
acattttacatctaaag437
<210>104
<211>423
<212>dna
<213>人工序列
<220>
<223>编码叶甲属rpii33-2v2dsrna的dna
<400>104
gcgtatgccaaaaaaggctttggaaaagagcatgccaaatggaatcctacatgtggtgtt60
gcctttgaatatgatcctgataacgctatgagacatacattatttcctaaaccagacgaa120
tggccgaagctagtaccagtcatcacgctggagcgcacatataggccctccatcagaaag180
tcattgtgtatatctctcatagggaacgagctgcttgcgtatttcccttccgtagtcaga240
gtcatcaatcagctgcaccgtgtcgtaaagcgggacgttcgcaagctcgtccgcggtagg300
ccattcgtctggtttaggaaataatgtatgtctcatagcgttatcaggatcatattcaaa360
ggcaacaccacatgtaggattccatttggcatgctcttttccaaagccttttttggcata420
cgc423
<210>105
<211>27
<212>dna
<213>人工序列
<220>
<223>探针rpii33-2v1prbset2
<400>105
agtgatccacgagtcataccagctact27
<210>106
<211>29
<212>dna
<213>人工序列
<220>
<223>探针rpii33-2v2prbset2
<400>106
tgtggtgttgcctttgaatatgatcctga29
<210>107
<211>173
<212>dna
<213>人工序列
<220>
<223>dsrna环多核苷酸
<400>107
gaagctagtaccagtcatcacgctggagcgcacatataggccctccatcagaaagtcatt60
gtgtatatctctcatagggaacgagctgcttgcgtatttcccttccgtagtcagagtcat120
caatcagctgcaccgtgtcgtaaagcgggacgttcgcaagctcgtccgcggta173
<210>108
<211>437
<212>rna
<213>人工序列
<220>
<223>rpii33-2v1dsrna
<400>108
cuuuagauguaaaauguacagaugaucaaacucgacauguaacaacugccgauuuuaaau60
cuagugauccacgagucauaccagcuacuuccaaacaucgugaugaugaauccucagagu120
auggugaaacaggaagcuaguaccagucaucacgcuggagcgcacauauaggcccuccau180
cagaaagucauuguguauaucucucauagggaacgagcugcuugcguauuucccuuccgu240
agucagagucaucaaucagcugcaccgugucguaaagcgggacguucgcaagcucguccg300
cgguacuguuucaccauacucugaggauucaucaucacgauguuuggaaguagcugguau360
gacucguggaucacuagauuuaaaaucggcaguuguuacaugucgaguuugaucaucugu420
acauuuuacaucuaaag437
<210>109
<211>423
<212>rna
<213>人工序列
<220>
<223>rpii33v2dsrna
<400>109
gcguaugccaaaaaaggcuuuggaaaagagcaugccaaauggaauccuacaugugguguu60
gccuuugaauaugauccugauaacgcuaugagacauacauuauuuccuaaaccagacgaa120
uggccgaagcuaguaccagucaucacgcuggagcgcacauauaggcccuccaucagaaag180
ucauuguguauaucucucauagggaacgagcugcuugcguauuucccuuccguagucaga240
gucaucaaucagcugcaccgugucguaaagcgggacguucgcaagcucguccgcgguagg300
ccauucgucugguuuaggaaauaauguaugucucauagcguuaucaggaucauauucaaa360
ggcaacaccacauguaggauuccauuuggcaugcucuuuuccaaagccuuuuuuggcaua420
cgc423
<210>110
<211>19
<212>dna
<213>人工序列
<220>
<223>引物rpii33-2v2fwdset2
<400>110
aaagagcatgccaaatgga19
<210>111
<211>19
<212>dna
<213>人工序列
<220>
<223>引物rpii33-2v2revset2
<400>111
ggccattcgtctggtttag19
- 还没有人留言评论。精彩留言会获得点赞!