一种前列腺癌的标志物SPOP及其在指导肿瘤治疗中的应用的制作方法
本发明属于癌症指导治疗领域,具体涉及spop作为标志物对前列腺癌进行分子分型以及指导前列腺癌患者应用bet抑制剂进行治疗的应用。
背景技术:
前列腺癌(prostaticcancer,pca)是一种严重威胁男性健康的恶性肿瘤,占全球肿瘤发病率第二位、死亡率第六位。我国pca的发病率近年来一直处于显著的上升趋势。晚期前列腺癌激素治疗敏感性的差异。雄激素阻断治疗是晚期前列腺癌的主要治疗方式,但是,许多学者在临床实践中发现,尽管在首次治疗时运用持续性雄激素阻断治疗对近90%的患者有效,但几乎所有患者在2~3年内均不可避免地进展为雄激素非依赖性前列腺癌,且发生上述转归的患者预后差,中位生存时间仅为18个月。晚期不同前列腺癌患者对内分泌治疗的反应差别迥异,这种现象临床上称之为前列腺癌对内分泌治疗的敏感性不同,综合国际国内研究,这种敏感性的差异尚无有效方法准确预测,这便导致了众多晚期前列腺癌的患者无奈地接受了不合理,不规范的内分泌治疗。溴结构域与额外末端结构域蛋白(bromodomainandextraterminaldomain,bet)逐渐成为乳腺癌和白血病中新的治疗靶点,应用bet小分子抑制剂可以有效抑制肿瘤的生长。而e3泛素酶亚层基结合配接器斑点型poz蛋白(e3ubiquitinligasesubstrate‐bindingadaptorspeckle‐typepozprotein,spop)在前列腺癌中发生高频的突变。
技术实现要素:
本发明的目的在于提供spop作为标志物对前列腺癌进行分子分型以及指导前列腺癌患者应用bet抑制剂进行治疗的应用,通过spop的检测,提高对前列腺癌进行bet抑制剂治疗的成功率。本发明发现野生型spop蛋白通过识别降解子模体能够结合并诱导bet蛋白家族(brd2/3/4)发生泛素化降解。spop为泛素连接酶e3家族成员cul3与底物结合的桥梁蛋白(adaptor),通过介导许多核蛋白的泛素化而促进它们的降解,从而参与调控细胞的多种功能。本发明中,作为前列腺癌的标志物可以是spop的dna、mrna或由其编码的蛋白质。所述spopdna基因pubmedgeneid为8405,在ncbi中的序列号(ncbireferencesequence):ng_041815.1,序列见https://www.ncbi.nlm.nih.gov/nuccore/ng_041815.1。所述spopmrna的核苷酸序列如seqidno.1所示,见https://www.ncbi.nlm.nih.gov/nuccore/aj000644.1;所述spop蛋白质序列如seqidno.2所示,见https://www.ncbi.nlm.nih.gov/protein/np_003554.1。本发明中,可通过spopdna是否发生突变,以及spop的mrna或由其编码的蛋白质的表达水平变化,确定基因的表达水平变化,从而对前列腺癌进行分子分型以及指导前列腺癌患者应用bet抑制剂进行治疗。
本发明还提供了spopdna的扩增引物或检测试剂在制备对前列腺癌进行分子分型的产品中的应用,所述spopdna的检测试剂可以但不限于是特异性检测spopdna的核酸探针。通过spopdna是否突变进行定量,来对前列腺癌进行分子分型。
本发明还提供了spopmrna的扩增引物或检测试剂在制备对前列腺癌进行分子分型的产品中的应用,所述spopmrna的检测试剂可以但不限于是特异性检测spopmrna的核酸探针。通过对所述基因spopmrna的表达水平进行定量,来对前列腺癌进行分子分型。所述mrna的表达水平可通过以下方法确定:微阵列技术、rna印迹和定量pcr;所述定量pcr为实时定量pcr或多重pcr等。
本发明还提供了spop蛋白质的检测试剂在制备对前列腺癌进行分子分型的产品中的应用,所述蛋白质的检测试剂可以但不限于是特异性检测spop蛋白质的抗体。通过对spop蛋白质的表达水平进行定量,来对前列腺癌进行分子分型。所述spop蛋白质的表达水平可通过以下方法确定:免疫组织化学、蛋白质印迹、elisa、ria和质谱法等。
本发明还提供了一种前列腺癌体外诊断产品,所述体外诊断产品包括特异性检测spopdna的试剂,和/或特异性检测spopmrna的试剂,和/或特异性检测spop蛋白质的试剂。所述前列腺癌体外诊断产品可用于对前列腺癌进行分子分型。所述特异性检测spopdna的试剂可以但不限于是核酸探针,所述核酸探针能特异性识别所述spopdna;所述特异性检测spopmrna的试剂可以但不限于是核酸探针,所述核酸探针能特异性识别所述spopmrna;所述特异性检测spop蛋白质的试剂可以但不限于是抗体,所述抗体能特异性识别所述spop蛋白质。
其中,所述前列腺癌体外诊断产品包括试剂盒、基因芯片、固体支持体等。所述固体支持体包括阵列、微阵列、蛋白质阵列等。
本发明还提供了spop在指导bet小分子抑制剂用于前列腺癌治疗中的应用。
本发明还提供了bet蛋白家族小分子抑制剂的应用,若相对于正常人群,患有前列腺癌的患者中所述spopdna发生突变,即为dna突变组,否则为dna正常组;或spopmrna或由其编码的蛋白质的表达量降低,则为低表达组,否则为正常表达组;dna正常组和mrna或蛋白正常表达组前列腺癌患者可采用bet小分子抑制剂进行治疗;而dna突变组和mrna或蛋白低表达组均不推荐应用bet小分子抑制剂进行治疗;其中,所述bet小分子抑制剂包括但不限于jq1。
本发明还提供了一种对已确诊为前列腺癌的患者进行分子分型以及指导前列腺癌患者应用bet抑制剂进行治疗的方法,所述方法包括以下步骤:
a)检测前列腺癌患者的病理样本中spop的dna拷贝数、或mrna或由其编码的蛋白质的表达量;
b)将患者分为dna正常组和突变组,mrna或蛋白质正常表达组和低表达组,若相对于正常人群,所述spopdna发生突变,即为dna突变组,否则为dna正常组;或spopmrna或由其编码的蛋白质的表达量降低,则为低表达组,否则为正常表达组。dna正常组和mrna或蛋白正常表达组前列腺癌患者可采用bet抑制剂进行治疗;而dna突变组和mrna或蛋白低表达组均不推荐应用bet抑制剂进行治疗。
本发明还提供了spop作为e3泛素连接酶cul‐3的衔接蛋白在制备促进bet蛋白的多聚泛素化以及蛋白降解的药物中的应用;其中,所述bet蛋白优选为brd2、brd3、brd4蛋白。
本发明还提供了spop在制备降低dek/erg/src‐3蛋白表达水平的药物中的应用。
本发明还提供了spop在制备降低rac1基因mrna和蛋白表达水平的药物中的应用。
本发明还提供了spop突变体在制备上调bet蛋白水平的药物中的应用;其中,所述bet蛋白优选为brd2、brd3、brd4蛋白;其中,所述spop突变体包括y87c,y87n,f102c,s119n,f125v,k129e,w131c,w131g,f133l,f133v和k134n。本发明中,所述y87c,y87n,f102c,s119n,f125v,k129e,w131c,w131g,f133l,f133v和k134n对应的突变位点和突变信息,之前文献均已报道,为公开信息,可参见http://www.pnas.org/content/110/17/6997.full.pdf。
本发明还提供了spop在制备降低jq1的耐药性的药物中的应用。
本发明还提供了brd小分子抑制剂在制备降低jq1的耐药性的药物中的应用;其中所述bet蛋白包括brd2、brd3、brd4蛋白,优选为brd4蛋白。
本发明还提供了usp9x/usp49的抑制剂在制备促进bet蛋白的多聚泛素化以及蛋白降解的药物中的应用;其中,所述bet蛋白优选为brd2、brd3、brd4蛋白。
本发明还提供了usp9x/usp49的抑制剂在制备降低jq1的耐药性的药物中的应用。
本发明还提供了一种降低jq1的耐药性的方法,所述方法包括提高spop的mrna和蛋白质的表达水平,或加入brd蛋白抑制剂,或加入usp9x/usp49抑制剂等。
本发明的有益效果在于,本发明提出的指导前列腺癌治疗的新标志物,即spop基因,该标志物能对前列腺癌进行分子分型以及指导前列腺癌患者应用bet抑制剂进行治疗。
附图说明
图1为酵母双杂交系统筛选鉴别的bet蛋白家族brd2/3/4:垂直虚线所示区域为bet蛋白与spop结合的区域,箭头所示区域为sbc模体。
图2为免疫共沉淀实验(co‐ip)应用抗brd2/3/4抗体进行共沉淀,发现brd2/3/4蛋白能够与spop蛋白发生结合。
图3为外源性高表达spop蛋白后,下调了brd2/3/4蛋白的表达水平;加入蛋白抑制剂(mg132)后逆转了这一现象。
图4为干扰spop蛋白表达后,brd2/3/4蛋白表达水平发生上调:sic:sirnacontrol阴性对照,sispop:spop干扰rna。
图5为293t细胞转染野生型spop和spop突变体spop‐△btb(btb结构域缺失)进行泛素化实验。野生型spop能够促进brd2/3/4的多聚泛素化,异源性表达的spop突变体spop‐△btb(btb结构域缺失)均无法促进brd2/3/4蛋白的多聚泛素化
图6为brd2/3/4蛋白中存在spop结合序列(spop‐bindingconsensus,sbc),即φ‐π‐s/t‐s/t‐s/t,φ为非极性残基,π为极性残基;a为brd2/3/4、macroh2a和dek的sbc;b为野生型以及sbc缺失型brd2/3/4蛋白。
图7为免疫共沉淀实验分析显示spop只与野生型brd2/3/4结合,但却不与sbc缺失的突变体结合。
图8为293t细胞转染brd2/3/4和brd2/3/4‐△sbc突变的质粒,发现spop不能促进brd2/3/4‐△sbc突变体的降解。
图9为293t细胞转染brd2/3/4和brd2/3/4‐△sbc突变的质粒后进行泛素化实验,想spop无法促进brd2/3/4‐△sbc突变体的降解。
图10为与myc标记的与前列腺癌相关的spop突变体,包括y87c,y87n,f102c,s119n,f125v,k129e,w131c,w131g,f133l,f133v和k134n,突变后的spop无法与brd2/3/4反应;进一步体内泛素化分析实验显示,所有spop突变体失去促进brd2/3/4蛋白多聚泛素化的能力。异源性表达的spop导致内源性brd2/3/4蛋白水平上升,提示前列腺癌相关的spop突变不能促进brd2/3/4蛋白多聚泛素化以及蛋白降解。a:spop突变体,包括y87c,y87n,f102c,s119n,f125v,k129e,w131c,w131g,f133l,f133v和k134n的免疫共沉淀实验;b:spop突变体失去促进brd2/3/4蛋白多聚泛素化的能力;c:异源性表达的spop导致内源性brd2/3/4蛋白水平上升。
图11为野生型及突变型spop前列腺癌患者肿瘤组织的brd2/3/4免疫组化;spop‐wt:spop野生型前列腺癌患者,spop‐mut:spop突变型前列腺癌患者。
图12为brd2/3/4mrna水平在野生型spop和突变型spop前列腺癌组织样本中无明显差异。
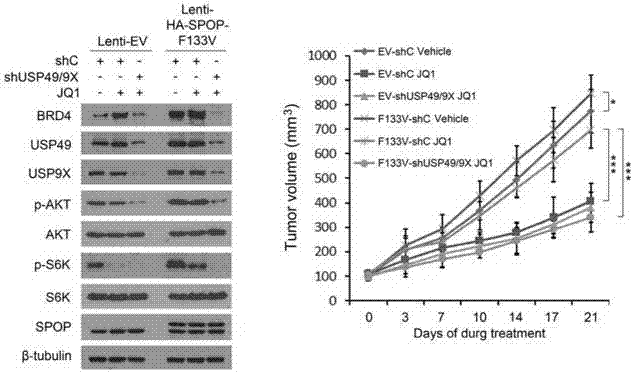
图13为体内实验证实spop突变f133v细胞系应用brd2/3/4干扰慢病毒和jq1抑制剂在前列腺癌中的作用。ev‐shc:空载对照;ev‐shbrd2/3/4:稳定干扰brd2/3/4对照;f133v‐shc:spopf133v突变空载对照;f133v‐shbrd2/3/4:稳定干扰brd2/3/4的spopf133v对照;jq1:bet蛋白抑制剂。*:p<0.05;***:p<0.001;n.s.:无统计学意义。
图14为spop突变的前列腺癌患者中发生上调的jq1耐药相关基因的韦恩图。
图15为野生型和spopf133v突变型c4‐2细胞在加入jq1后brd4蛋白在rac1基因位点的chip‐seq数据。
图16为体内实验证实spop突变f133v细胞系应用usp49/9x干扰慢病毒和jq1抑制剂在前列腺癌中的作用。ev‐shc:空载对照;ev‐shusp49/9x:稳定干扰usp49/9x4对照;f133v‐shc:spopf133v突变空载对照;f133v‐shusp49/9x:稳定干扰usp49/9x的spopf133v对照;jq1:bet蛋白抑制剂。
具体实施方式
结合以下具体实施例和附图,对本发明作进一步的详细说明。实施本发明的过程、条件、实验方法等,除以下专门提及的内容之外,均为本领域的普遍知识和公知常识,本发明没有特别限制内容。
实施例1spop结合并识别brd2/3/4蛋白的sbc模体降解子进而促进其泛素化降解
(1.1)酵母双杂交系统筛选鉴别spop的降解靶蛋白
本发明通过对酵母双杂交系统筛选spop的靶蛋白,寻找到包括已知dek和src-3在内的246个阳性克隆菌株,并在其中筛选靶蛋白,其中命中数最高的是bet蛋白家族,52个克隆为brd2,2个克隆为brd3,6个克隆为brd4(如图1所示)。
在lncap细胞中进行免疫共沉淀反应(co-ip),应用igg或brd2/3/4抗体进行westernblot发现spop蛋白能够与brd2/3/4蛋白发生结合(图2)。因此,spop蛋白可与bet蛋白家族成员发生结合。
(1.2)spop促进brd2/3/4蛋白降解
如上所述,spop蛋白可与bet蛋白家族成员发生结合,提示brd2/3/4可能为蛋白降解通路的靶点。通过本发明的研究发现,在293t细胞中通过myc-spop质粒异源性表达spop蛋白发现spop与brd2/3/4蛋白发生反应,加入蛋白抑制剂mg132后,发现spop对brd2/3/4蛋白的降解作用消失,brd2/3/4蛋白水平增加(图3)。在lncap、22rv1前列腺癌细胞系和bph-1前列腺正常细胞系中,应用sirna敲减spop表达水平后发现,相应细胞系中内源性brd2/3/4蛋白含量上调,src作为已知的spop下游靶蛋白也发生了蛋白水平相应的上调(图4)。将spop具有酶活性的相应序列进行缺失得到spop突变体spop-△btb(btb结构域缺失),进行泛素化检测发现,野生型spop能够促进brd2/3/4的多聚泛素化,而异源性高表达的spop突变体spop-△btb(btb结构域缺失)无法促进brd2/3/4蛋白的多聚泛素化(图5)。以上数据均提示spop能促进brd2/3/4蛋白的多聚泛素化和降解。
综上所述,本发明的研究表明,在不同类型的细胞中(包括前列腺癌细胞),spop作为e3泛素连接酶cul-3的衔接蛋白可促进brd2/3/4蛋白的多聚泛素化以及蛋白降解。
(1.3)spop特异性识别结合brd2/3/4蛋白中的sbc结构域
spop若想发挥作用,底物蛋白必须要具备特异的sbc域(spop-bingconsensusmotif,sbcmotif)。本发明发现brd2中的260~313氨基酸,brd中的3148~572氨基酸以及brd中的4241~335氨基酸分别为需要与spop反应的最小区域,且在这些区域中存在一相同的sbc结构域(图6a)。分别构建了缺失sbc结构域的brd2/3/4突变体(brd2-△sbc,brd3-△sbc,brd4-△sbc)(图6b)。转染缺失sbc结构域的brd2/3/4突变体(brd2-△sbc,brd3-△sbc,brd4-△sbc)至293t细胞中,加入myc-spop质粒,应用免疫共沉淀实验分析显示spop只与野生型brd2/3/4结合,但却不与sbc缺失的突变体结合(图7-8)。应用泛素化实验检测293t细胞中brd2/3/4蛋白及其突变体的表达水平,发现spop无法促进brd2/3/4-△sbc突变体的降解,仅能促进野生型brd2/3/4蛋白降解(图9)。
(1.4)spop突变无法促进brd2/3/4蛋白降解
本发明建立了一系列与myc标记的与前列腺癌相关的spop突变体,包括y87c,y87n,f102c,s119n,f125v,k129e,w131c,w131g,f133l,f133v和k134n。免疫共沉淀实验显示,在293t前列腺癌细胞中,突变后的spop无法与brd2/3/4反应(图10a)。进一步体内泛素化分析实验显示,所有spop突变体失去促进brd4蛋白多聚泛素化的能力,而仅有野生型spop能够发挥促进brd4蛋白泛素化的作用(图10b)。慢病毒转染spop及相应突变体spop-y87c/f102c/f125v/f133l至前列腺癌细胞系c4-2中,高表达相应spop蛋白后发现,野生型spop能够降低brd2/3/4及dek/erg/src-3蛋白水平,而spop突变体蛋白无法发挥相应的作用,brd2/3/4等蛋白表达水平发生上调(图10c)。上述结果提示,前列腺癌相关的spop突变不能促进brd2/3/4蛋白多聚泛素化以及蛋白降解。
实施例2spop对bet蛋白抑制剂抗肿瘤作用的影响
(2.1)前列腺癌患者肿瘤组织bet蛋白表达水平上调
本发明通过对两组样本含量共为99例原发性前列腺癌队列中进行全基因组测序、rna测序、sanger测序后发现,有13例前列腺癌样本包含spop突变。免疫组织化学(ihc)分析实验显示,绝大多数spop突变的前列腺组织中brd2/3/4蛋白染色强阳性(brd2:85%(11/13),brd3:92%(12/13),brd4:85%(11/13))(图11)。相反,在野生型spop的前列腺癌组织样本中,仅有一部分brd2/3/4蛋白染色强阳性(brd2:29%(25/86),brd3:34%(29/86),brd4:41%(35/86)),大多数brd2/3/4染色呈弱阳性或阴性(brd2:71%(61/86),brd3:66%(57/86),brd4:59%(51/86))(图11)。说明发生spop突变的前列腺癌患者肿瘤组织中brd2/3/4蛋白表达水平发生上调。
(2.2)前列腺癌患者肿瘤组织中brd2/3/4mrna水平检测
本发明通过对32例前列腺癌组织样本进行rna测序发现(26例野生型spop,6例突变型spop),brd2/3/4mrna水平在野生型spop和突变型spop前列腺癌组织样本中无明显差异(图12),说明发生spop突变的前列腺癌患者肿瘤组织中brd2/3/4的mrna表达水平并未发生变化,上述结果提示spop突变的前列腺癌中brd2/3/4蛋白水平的改变发生在brd2/3/4mrna翻译后水平。
综上,前列腺癌相关的spop突变可在翻译后水平导致bet蛋白含量上升。
(2.3)spop对bet抑制剂应用于抗肿瘤的影响
bet小分子抑制剂是目前潜在的肿瘤靶向治疗方法。本发明发现分别用bet抑制剂jqi处理实验组(ev-shcjq1)、干扰brd2/3/4表达组(ev-shbrd2/3/4jq1)和阴性对照组(ev-shcvehicle),发现jq1可明显抑制肿瘤增长。而spopf133v突变组(f133v-shcjq1)中,jq1对肿瘤的抑制作用消失。在spopf133v突变组对brd2/3/4进行敲低后的细胞系(f133v-shbrd2/3/4jq1)中,jq1又可以显著抑制肿瘤的生长(图13),说明spop突变后brd2/3/4蛋白表达水平上调时,应用jq1抑制肿瘤生长的效果较差,可能是产生jq1耐药的原因。
spop突变的前列腺癌患者中发生上调的jq1耐药相关基因与brd4chip-seq相重合的基因包括rac1、fdft1、dhcr24等(图14),进而在chip-seq公开数据对于brd4蛋白是否作用于其启动子区域进行验证。野生型和spopf133v突变型c4-2细胞在加入jq1后brd4蛋白在rac1基因位点的chip-seq数据发现brd4结合到rac1基因启动子区域,在jq1耐药的spopf133v突变细胞系中rac1基因mrna和蛋白表达上调(图15),说明在spop突变后brd4蛋白能够促进rac1基因转录,应用jq1不能阻碍这一进程,提示这是spop突变患者应用jq1产生耐药的机制。本发明发现分别用bet抑制剂jqi处理单纯干扰usp9x/usp49组(ev-shusp9x/usp49jq1)、spopf133v单纯突变组(f133v-shcjq1)和对spopf133v突变组进行usp9x/usp49敲低后的细胞系(f133v-shusp9x/usp49jq1),发现jq1在spopf133v突变组中不能够抑制肿瘤生长,但在干扰usp9x/usp49蛋白表达后又可以显著抑制肿瘤的生长(图16)。上述结果提示,brd4与usp9x/usp49发生相互作用,干扰usp9x/usp49后能够促进brd4多聚泛素化,能够部分阻断spopf133v突变引起的brd4蛋白水平上升,进而能够逆转f133v引起的jq1耐药。
此外,本发明研究发现brd4蛋白水平的下降可以引起细胞对jq1敏感,同时spop敲低后可以引起细胞对jq1的耐受。
以上研究均提示brd4蛋白为去势抵抗性前列腺癌细胞(如c4-2细胞)中决定jq1敏感性的关键性分子,spop突变细胞中bet蛋白水平升高与jq1抵抗性有关。
本发明的保护内容不局限于以上实施例。在不背离发明构思的精神和范围下,本领域技术人员能够想到的变化和优点都被包括在本发明中,并且以所附的权利要求书为保护范围。
<110>上海长海医院
<120>一种前列腺癌的标志物spop及其在指导肿瘤治疗中的应用
<160>2
<170>patentinversion3.3
<210>1
<211>1642
<212>spopmrna
<213>人工序列
<400>1
1gaatcggcggtcccgcaggtcccggatgttgcggacagtatgaggcaagcgcagggggac
61ggggaccagcagctgtcgccgccgctctcagggtgaagagggaacagaaatctttgcccc
121ctgactttggaaatctcgtttaaccttcaaactggcgatgtcaagggttccaagtcctcc
181acctccggcagaaatgtcgagtggccccgtagctgagagttggtgctacacacagatcaa
241ggtagtgaaattctcctacatgtggaccatcaataactttagcttttgccgggaggaaat
301gggtgaagtcattaaaagttctacattttcatcaggagcaaatgataaactgaaatggtg
361tttgcgagtaaaccccaaagggttagatgaagaaagcaaagattacctgtcactttacct
421gttactggtcagctgtccaaagagtgaagttcgggcaaaattcaaattctccatcctgaa
481tgccaagggagaagaaaccaaagctatggagagtcaacgggcatataggtttgtgcaagg
541caaagactggggattcaagaaattcatccgtagagattttcttttggatgaggccaacgg
601gcttctccctgatgacaagcttaccctcttctgcgaggtgagtgttgtgcaagattctgt
661caacatttctggccagaataccatgaacatggtaaaggttcctgagtgccggctggcaga
721tgagttaggaggactgtgggagaattcccggttcacagactgctgcttgtgtgttgccgg
781ccaggaattccaggctcacaaggctatcttagcagctcgttctccggtttttagtgccat
841gtttgaacatgaaatggaggagagcaaaaagaatcgagttgaaatcaatgatgtggagcc
901tgaagtttttaaggaaatgatgtgcttcatttacacggggaaggctccaaacctcgacaa
961aatggctgatgatttgctggcagctgctgacaagtatgccctggagcgcttaaaggtcat
1021gtgtgaggatgccctctgcagtaacctgtccgtggagaacgctgcagaaattctcatcct
1081ggccgacctccacagtgcagatcagttgaaaactcaggcagtggatttcatcaactatca
1141tgcttcggatgtcttggagacctctgggtggaagtcaatggtggtgtcacatccccactt
1201ggtggctgaggcataccgctctctggcttcagcacagtgcccttttctgggacccccacg
1261caaacgcctgaagcaatcctaagatcctgcttgttgtaagactccgtttaatttccagaa
1321gcagcagccactgttgctgccactgaccaccaggtagacagcgcaatctgtggagctttt
1381actctgttgtgaggggaagagactgcattgtggccccagacttttaaaacagcactaaat
1441aacttgggggaaacggggggagggaaaatgaaatgaaaaccctgttgctgcgtcactgtg
1501ttccctttggcctgtctgagtttgatactgtggggattcagtttaggcgctggcccgagg
1561atatcccagcggtggtacttcggagacacctgtctgcatctgactgagcagaacaaatcg
1621tcaggtgcctggagcaaaaagg
<210>2
<211>374
<212>spopprotein
<213>人工序列
<400>2
1msrvpsppppaemssgpvaeswcytqikvvkfsymwtinnfsfcreemgeviksstfssg
61andklkwclrvnpkgldeeskdylslylllvscpksevrakfkfsilnakgeetkamesq
121rayrfvqgkdwgfkkfirrdflldeangllpddkltlfcevsvvqdsvnisgqntmnmvk
181vpecrladelgglwensrftdcclcvagqefqahkailaarspvfsamfehemeeskknr
241veindvepevfkemmcfiytgkapnldkmaddllaaadkyalerlkvmcedalcsnlsve
301naaeililadlhsadqlktqavdfinyhasdvletsgwksmvvshphlvaeayrslasaq
361cpflgpprkrlkqs
- 还没有人留言评论。精彩留言会获得点赞!