一种植物花粉特异性启动子PSP1及其应用的制作方法
技术领域:
本发明属于分子生物学和基因工程领域,涉及一种植物花粉特异性启动子psp1及其应用。
背景技术:
:
转基因技术被认为是解决土地资源锐减和人口急剧膨胀这一矛盾最有效的方法之一。利用转基因技术改良和培育新品种可以带来巨大经济、社会效益和显著的生态效益,因此受到各国政府和科学家的重点关注。
转基因植物所携带的外源基因可能随花粉漂移到近缘物种或野生种,出现“基因漂移”,这种基因漂移可能会导致附近近缘物种或野生种发生内在的基因变化,产生新的性状或形成新的物种,从而引起生态系统的变化。特别是具有抗病虫害、抗除草剂功能或对环境胁迫具有耐受性的基因的漂移,可能会增强受体近缘物种或野生种在生长势、越冬性、种子活力等方面的特性,提高其入侵其他植物栖息地的能力,破坏自然种群的平衡,影响生物多样性,最后很有可能会演变无法控制的“超级杂草”,严重影响的生态环境和经济社会发展。
植物花粉是“基因漂移”现象中最直接的“肇事者”,通过风媒或虫媒进行花粉传播是最常见的方式。因此,为了减少或避免基因漂移,只有通过阻断花粉传播或者改变花粉活性使其丧失活力,而通过物理阻断花粉漂移的难度较大、可行性低。因此,改变花粉活性使其丧失活力是阻断花粉漂移的重要手段。要改变花粉活力,最直接的方式就是针对花粉进行基因调控。花粉作为植物的雄配子体,其发育包括花粉母细胞形成期、花粉母细胞减数分裂期、单细胞花粉期和二、三细胞花粉期等,整个过程非常复杂,需要大量基因在不同启动子的精确调控下协同表达,而花粉特异启动子对于花粉的正常发育调控起着重要作用。
目前在农业生物技术领域被广泛应用的主要是一些组成型的强启动子,比如ubiquitin类型启动子和camv35s启动子,但是在利用这些启动子诱导目的基因转化水稻等植物进行植物改良时,往往会由于目的基因表达的时空特异性不好控制导致改良效果不显著,或者由于组成型启动子诱导基因表达量太高从而影响植物的生长发育。因此,为了改变花粉的活力,同时避免或减少对植物生长带来过多的负面影响,鉴定和研究花粉特异性启动子具有重要意义。
技术实现要素:
:
本发明目的是提供一种可驱动外源目的基因在植物花粉中特异性表达的启动子及其应用。
为实现上述目的,本发明提供一种植物花粉特异性启动子psp1,具有(a)、(b)或(c)所示的序列:
(a)seqidno:1或图2所示的核苷酸序列。该序列来源于水稻品种日本晴(oryzasativalcv.nipponbare)序列,本文中称为psp1或启动子psp1或psp1启动子。
(b)在(a)所限定的核苷酸序列中添加和/或取代和/或缺失一个或多个核苷酸而生成的具有同等花粉特异性启动功能的突变基因、等位基因或衍生物;其包含与(a)所述的核苷酸序列相比具有70%以上同源性,且具有同等花粉特异性启动子功能的核苷酸序列。本领域技术人员根据相同目的能够容易地获得对应序列,以实现相同功能。
(c)与(a)或(b)所示的核苷酸序列互补的核苷酸序列。本领域技术人员根据相同目的能够容易地鉴定并利用与植物花粉特异性启动子psp1核苷酸序列互补的核苷酸序列,因此,具有启动子活性并在严格条件下与本发明启动子序列或其片段杂交的分离序列包括在本发明中。
其中,所述核苷酸序列互补,是指在严格条件下能与psp1杂交。严格条件是指探针将与其靶序列杂交至可探测程度超过与其它序列杂交(如至少2倍于背景)的条件。严格条件具有序列依赖性,且因环境的不同而不同。通过控制杂交和/或洗涤条件的严格性,可以鉴定与探针100%互补的靶序列(同源探测)。可选择地,可以调节严格条件以允许一些序列错配,使得探测到较低程度的相似性(异源探测)。通常,探针长度短于大约1000个核苷酸,优选地短于500个核苷酸。
本发明提供了含有上述花粉特异性启动子psp1的表达盒。
作为进一步地改进,所述表达盒中,在花粉特异性启动子psp1的下游连接有结构基因、调节基因、结构基因的反义基因或调节基因的反义基因。
本发明提供了含有上述花粉特异性启动子psp1的重组表达载体。在所述的重组表达载体中,所述花粉特异性启动子psp1连接于目的基因或待表达的基因序列的上游;优选地,所述目的基因或待表达的基因为gus基因,所述重组表达载体为psp1-gus-pcambia1305,该重组表达载体将seqidno:1所示的核苷酸序列即psp1或启动子psp1或psp1启动子构建于pcambia1305中得到的重组表达载体,本文中称为psp1-gus-pcambia1305。或者:所述目的基因或待表达的基因为任何对植物的花粉性状具有改良能力的基因,用于增加花粉活性。或者:所述目的基因或待表达的基因为任何对植物的花粉性状具有抑制功能的基因,用于降低花粉活性,减少或避免基因飘移。
本发明提供了含有上述表达盒的重组表达载体或工程菌。优选地,所述的重组表达载体是由上述表达盒与质粒、病毒或表达载体所构建的重组载体。优选地,重组表达载体为双元载体或共合载体。
本发明还提供了含有上述花粉特异性启动子psp1、表达盒、重组表达载体或工程菌的宿主细胞。优选地,该细胞为大肠杆菌细胞、农杆菌细胞或植物细胞。
本发明还提供了扩增上述花粉特异性启动子psp1的引物对,所述引物对的核苷酸序列为seqidno:2-3所示的核苷酸序列。
本发明还提供了一种在植物的花粉中驱动目的基因表达的方法,将上述花粉特异性启动子psp1和目的基因引入植物基因组,得到目的基因在花粉中特异表达的转基因植物。所述方法优选可包括:(a)将前述花粉特异性启动子psp1连接于目的基因的上游构建植物表达载体;(b)将含有启动子psp1与目的基因融合产物的植物表达载体转入到根瘤农杆菌中;(c)利用转入了植物表达载体的根瘤农杆菌对特定植株的成熟胚愈伤组织进行农杆菌浸染转化;(d)利用转化后的成熟胚愈伤组织筛选、分化、培养相应的转基因植株,在所述的转基因植株中包含有上述植物表达载体。需要特别说明的是:(a)上面所述的目的基因是指任何一段核酸序列,具有编码某种蛋白质或其他活性物质的功能,包括rna或dna序列;(b)所述核酸序列包括不同于启动子植物种类的异源核酸序列,以及从相同于启动子植物种类中获得的同源核酸序列,但这些序列与野生型(非转基因)植物的启动子无关;(c)所述的表达载体是指现有技术中已知的、能够在植物中进行表达的任何载体;(d)所述的在植物的花粉中驱动特定的目的基因表达的方法,是指已知的、能够将目的基因导入植物细胞或植物组织的任何一种植物转化方法,如基因枪转化法等。
本发明还提供了上述花粉特异性启动子psp1、表达盒、重组表达载体、宿主细胞或引物对在以下(a)至(d)中任一项中的应用:
(a)培育植物品种或品系;
(b)培育授粉受精能力增强植物品种或品系;
(c)培育授粉受精能力消弱的植物品种或品系;
(d)培育雄性不育或恢复系植物品种或品系。
本发明中的启动子等可以应用于禾本科植物,例如水稻、玉米、小麦、大麦、高粱等,优选为水稻。
本发明以花粉特异性启动子psp1构建的表达载体通过农杆菌介导的水稻愈伤转化获得了转基因水稻植株,转基因植株鉴定及组织化学染色分析显示该启动子可驱动目的基因在花粉中高效特异性表达。本发明提供的花粉特异性启动子psp1及其表达载体可用于外源目的基因在花粉中特异性表达,避免基因在植物其他组织组成型或持续性表达带来的不利影响;可用于植物花粉生长发育相关基因的功能分析和鉴定;可用于雄性不育系和恢复系的创建,为杂种优势利用提供关键材料基础;可用于花粉败育,避免转基因植物基因漂移或逃逸带来的生物安全问题;特别是在水稻等禾本科植物的基因工程应用中,具有广阔应用前景和农业经济价值。
术语“启动子”是指rna聚合酶或一些反式作用因子识别并与之结合从而正确有效地起始基因转录的一段dna序列。
术语“花粉特异性启动子”是指可驱动目的基因在花粉中特异性表达的启动子,而在其他组织无表达活性或表达活性很弱。
术语“表达盒”是指基因表达需要的最小核苷酸序列框架,包括三个部件启动子、目的基因和终止子。
附图说明:
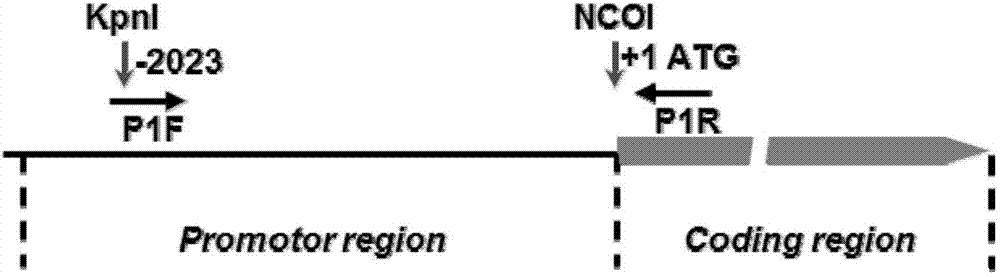
图1显示的是水稻花粉特异性启动子克隆流程图。p1f和p1r为扩增启动子序列的特异性引物,kpni和ncoi为启动子序列两端酶切位点。
图2显示的是经过测序鉴定的水稻花粉特异性启动子psp1核苷酸序列。
图3显示的是psp1-gus-pcambia1305表达载体图谱。载体骨架是pcambia1305载体,表达盒包括psp1启动子、gus基因和nos终止子。
图4显示的是转化psp1-gus-pcambia1305载体的转基因过程,包括(a)水稻愈伤诱导,(b)浸染后抗性愈伤筛选,(c)抗性愈伤分化和(d)生根筛选。
图5显示的是转化psp1-gus-pcambia1305载体的转基因阳性植株的鉴定。p2f和p2r是鉴定转基因植株的特异性引物。m为dl2000标准分子量。上样孔5和12为转基因阳性植株;上样孔13为阴性对照非转基因植株;上样孔14为阴性对照双蒸水;上样孔15为阳性对照psp1-gus-pcambia1305质粒。
图6显示的是转化psp1-gus-pcambia1305载体的转基因阳性植株抽穗期表型。
图7显示的是转基因阳性植株花粉gus染色图。a:花药,标尺为100μm;b:花粉,标尺为5μm。箭头指示着色后花粉粒。
具体实施例方式:
以下实施例进一步说明本发明的内容,但不应该理解为对本发明的限制。在不背离本发明精神和实质的情况下,对本发明方法、步骤或条件所作的修改或替换,均属于本发明的范围。若未特别指明,实施例中所用的技术手段为本领域技术人员所熟知的常规手段。若未特别指明,实施例中所用的生化试剂、载体、耗材等均为市售购买产品。
实施例1.水稻花粉特异性启动子psp1的克隆
根据ncbi中提供的水稻品种日本晴(oryzasativalcv.nipponbare)全基因组序列,设计psp1序列的特异性扩增引物,并根据选用的pcambia1305表达载体及psp1序列的特征,在特异性引物两端添加特异性酶切位点(图1)。具体设计的引物为:正向引物(p1f)5’端添加kpni酶切位点(ggtacc),反向引物(p1r)5’端添加ncoi酶切位点(ccatgg),引物序列如下:
p1f正向引物:5’-cggggtaccaaccaaacctgtggctgc-3’kpni
p1r反向引物:5’-catgccatggcgctctgtgacgccat-3’ncoi
然后提取水稻品种日本晴基因组dna,并以日本晴基因组dna为模板,利用上面设计的引物(p1f和p1r)扩增启动子psp1序列,采用如下扩增程序:94℃预变性4分钟;98℃变性30秒,55℃退火30秒,68℃延伸2分钟,35个循环;最后72℃再延伸10分钟。回收pcr扩增的目的片段,目的片段长度为2258bp,将其连接zeroblunttopo载体到(invitrogen),转化大肠杆菌dh5a感受态细胞,然后经过菌落pcr鉴定筛选阳性克隆,并将阳性克隆送invitrogen公司测序。测序验证正确的克隆即为所获得的psp1启动子序列,其核苷酸序列如图2或seqidno:1所示。
实施例2.启动子psp1驱动的植物表达载体的构建
将实施1中获得的、经过测序鉴定的阳性克隆进行质粒提取,提取的质粒用kpni和ncoi双酶切,回收psp1启动子片段。同时利用kpni和ncoi对pcambia1305进行双酶切线性化,并回收pcambia1305骨架,将酶切后回收的psp1启动子片段与酶切后回收的pcambia1305骨架用t4连接酶(购于neb公司)进行连接,得到psp1启动子与gus基因融合的植物表达载体psp1-gus-pcambia1305(图3),采用电击转化法将psp1-gus-pcambia1305表达载体转入根瘤农杆菌(agrobacteriumtumefaciens)eha105中。
实施例3.将启动子psp1驱动gus基因的表达载体转化水稻
采用农杆菌介导的水稻成熟胚愈伤浸染转化方法,将重组表达载体psp1-gus-pcambia1305转入水稻成熟胚中,转化方法如下:(1)水稻成熟胚愈伤组织的诱导:将成熟的日本晴种子去壳,然后用75%酒精表面消毒2min,然后用30%naclo溶液浸泡30min,并重复一次,然后用灭菌水清洗4-5次。然后将种子置于诱导培养基上培养,26度避光培养诱导愈伤组织用于转化(图4a)。(2)水稻愈伤组织与农杆菌的共培养:将实施例2中鉴定含有psp1-gus-pcambia1305表达载体的eha105菌株进行活化、富集、重悬,调整od600=0.3-0.5。将愈伤组织收集于50ml无菌离心管中,倒入重悬好的农杆菌悬浮液,浸染愈伤。浸泡10-30min后,倒掉悬浮液,将浸染过的愈伤置于无菌滤纸上吸干多余的农杆菌菌液。然后将愈伤置于铺有无菌滤纸的培养皿中,26度避光培养2-3天。(3)抗性愈伤的筛选:共培养完成后,将愈伤转移至含有50mg/ml的潮霉素的筛选培养基中,26-28度条件下抗性筛选(图4b)。(4)抗性愈伤的分化:将筛选培养基中生长状态良好的愈伤置于分化培养基中,置于16小时光照/8小时黑暗、环境温度在26-28度之间的条件下,分化培养,直至分化长出小苗(图4c)。(5)分化小苗的生根:待分化小苗月2cm左右时,将小苗转移到生根培养基中,进行生根培养(图4d)。长出根系的小苗移栽于温室或转基因圃中进行生长。
实施例4.对实施例3中获得的转基因水稻植株进行分子检测
对实施例3中获得的转基因水稻植株进行分子检测,以便筛选出转基因阳性植株,检测方法主要为特异性引物的pcr检测。pcr检测根据psp1启动子和gus基因设计检测引物p2f和p2r,以t0代转基因水稻植株的基因组dna为模板,并设置阳性和阴性对照组,进行pcr检测,pcr程序同实施例1。结果如图5所示,12株转基因水稻植株中有2株为阳性转基因植株。转基因阳性植株抽穗期表型如图6所示。
引物序列如下:
p2f正向引物:5’-tgagaagacgagggggttta-3’
p2r反向引物:5’-aggaatccgcccttgtgct-3’
实施例5.对实施例4中鉴定的转基因水稻进行gus组织化学检测
对实施例4中获得的转基因水稻植株进行gus染色处理。x-gluc反应液配制需要试剂:100mm磷酸钠缓冲液,ph=7.5-7.7;05medta溶液(mw:372.24);100mm的k3fe(cn)6(mw:329.3);100mm的k4fe(cn)6(mw:422.4);1mg/10μlx-gluc染料。具体染色处理过程:首先将待处理的样品用90%丙酮溶液-20℃处理1小时。用0.1m磷酸缓冲液和5mmk3fe(cn)6的混合液在真空洗2次,每次5分钟。向清洗过的样品中加gus染液,37℃避光处理过夜。反应完全的样品在70%无水乙醇中脱色直到叶绿素消失。结果如图7所示,在转基因水稻的花粉中特异性的呈现明显蓝色,表明psp1启动子可以驱动gus基因在水稻花粉中特异性表达。
本发明以花粉特异性启动子psp1构建的表达载体通过农杆菌介导的水稻愈伤转化获得了转基因水稻植株,转基因植株鉴定及组织化学染色分析显示该启动子可驱动目的基因在花粉中高效特异性表达。可用于(a)培育植物品种或品系;(b)培育授粉受精能力增强植物品种或品系;(c)培育授粉受精能力消弱的植物品种或品系;(d)培育雄性不育或恢复系植物品种或品系等。
序列表
<110>中国水稻研究所
<120>一种植物花粉特异性启动子psp1及其应用
<160>5
<170>siposequencelisting1.0
<210>1
<211>2023
<212>dna
<213>稻属水稻(oryzasativa)
<400>1
aaccaaacctgtggctgcgaattaaggtggggtaattaacctgcgtcctctccgctctct60
gggcttgatgcccccaccattttctccaactaaccactgctccccgctgatccgctgcgg120
gggccccgaacaccgagcagcccgtgcgtgtggcgtttctggccgttggatcgtgtgggg180
ccgcgtggatggacggcggatggaggcgtccccgcgtggctcacccgccgcagcggatcg240
gatcggatgcgcgccagcacggagcgagaacatcgctctcgtccgcgagcacccagagtg300
gaatttcgctttttaatcggagtagatgtgtttttgttttacacctaggtcctctctatt360
ttttctttctacatcaaccatttgttttgttaggtatttatgtttattactattattatt420
gcctctttttgttaatttcttctgtttttgtagtatcatctatgatgaaatttcgacgtg480
cgatttctacatataatcatatactccgtataagataagcccatccatgtactagcgcat540
agcgttgtcattggaattgaccctaaaatgggacacataatgtatggaaagaaatgtgcg600
gaataagggttatgttccataatggaatccataaggtatacagatactctcttctactcg660
aaaggaacatggaagattcttattattaaaagcaaaacaaagtggataaccacaataaaa720
gttagctatagggcaccaacaattggaaattaattaatacatgccacttaaatcttgatc780
tataccatatatagttacatatcaatgctaattacttggcatataattatctctttaatg840
tgcatatataataattgattcgtcatactcatacataaaggccccagaggcacatgctca900
tccagttaaattactccctccgtttcaaaatgtttgacaccgttgactttttagcacgtg960
tttgaccattcgtcttattcaaaaaattttgtgaaatatgtaaaactatatgtgtacatg1020
aaagtatatttaacaatgaatcaaatgatatgaaaagaataaataattacttaaattttt1080
tgaataagacgaatggtcaaacacatactaaaaagtcaacagtgtcaaacattttgaaac1140
ggagggagtataataagatacgacataaatatatgacttagtagataaccggcttgcgga1200
tctttttctaacatgacaggtgccagctaagtatgtaagtactatagtatcataccgtgt1260
ctttctccatattttatcaaacagaacatggtcgcacaaatttggcataagagagaatat1320
tgctgctataattaaaaaagaaatattatcttgtaataagtatggaatagtagtatcact1380
taatagaatatggaatattctcctacagtaaacatgtaatatattctcttatataagaaa1440
atattgacataaagtgcataagcatgttgtaatttcattacttaatgaaatcaaccccat1500
tatccaaaaaagcaaataatccttccgtttcataccatatataagacgttttgaatttct1560
aaaattttaacttattaaagtttgacctaatttataggaaaaatattgtttatagcatca1620
gattggttttattaaatttaacattgaatatattctctgatagtttccttatttatgtta1680
aaaatataaaatgacttataaaaaattaaaataatttataatataaaacagatgtggtat1740
tatatccaagaaacactcacaaatcaatcaaatactactaggtgtactcctaacgtacaa1800
acataaatgtacaagtacgcgtagttgtttaaaaattgagaagacgagggggtttactgc1860
gagaaggcaaaacagaaggaaaaaaaaaacagcgaaactctactggagtcgcgagcagcg1920
tactctcctccaacctccttcccacttctcaacctcgcctcgccatctccgcgccgcggc1980
ccagatccctccggcgagcgccgccgccgccaccgcctccgcc2023
<210>2
<211>27
<212>dna
<213>人工序列(p1f正向引物)
<400>2
cggggtaccaaccaaacctgtggctgc27
<210>3
<211>26
<212>dna
<213>人工序列(p1f反向引物)
<400>3
catgccatggcgctctgtgacgccat26
<210>4
<211>20
<212>dna
<213>人工序列(p2f正向引物)
<400>4
tgagaagacgagggggttta20
<210>5
<211>19
<212>dna
<213>人工序列(p2f反向引物)
<400>5
aggaatccgcccttgtgct19
- 还没有人留言评论。精彩留言会获得点赞!