结合生长抑素受体2的异源二聚抗体的制作方法
本申请要求于2017年4月3日提交的美国临时申请第62/481,065号、于2016年9月20日提交的美国临时申请第62/397,322号、于2016年6月28日提交的美国临时申请第62/355,821号以及于2016年6月28日提交的美国临时申请第62/355,820号的权益,其内容明确地以全文引用的方式整体并入本文中。序列表本申请含有序列表,所述序列表已以ascii格式以电子方式提交且特此以全文引用的方式并入本文中。创建于2017年6月28日的所述ascii复本命名为067461-5194-wo_sl.txt且其大小为2,771,347字节。
背景技术:
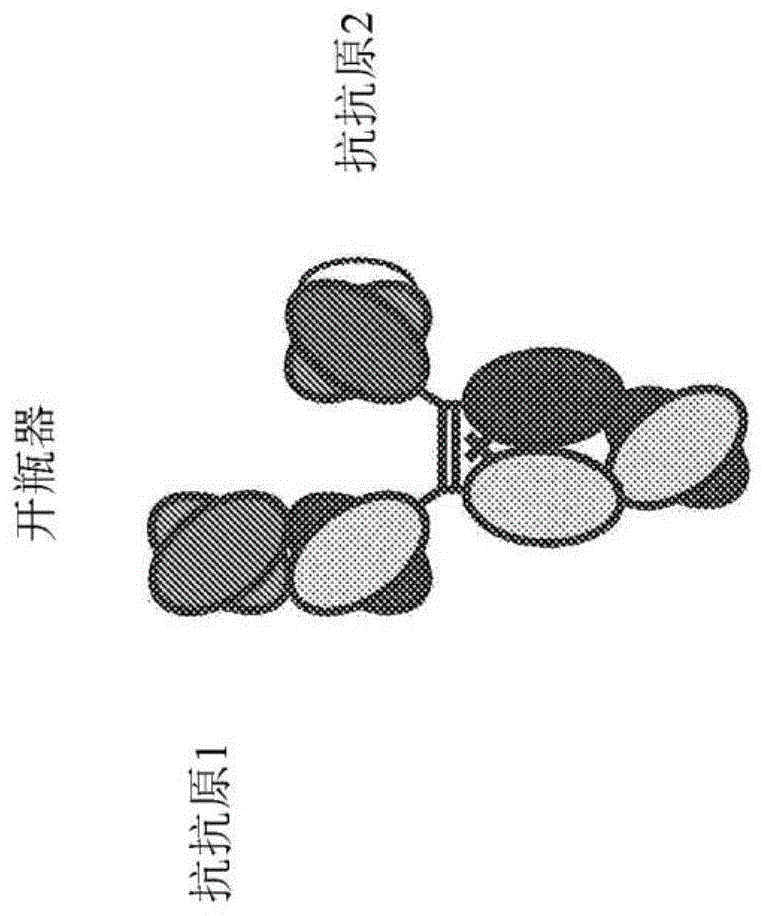
:基于抗体的治疗剂已经成功地用于治疗多种疾病,包含癌症和自体免疫性/发炎性病症。然而,仍需要对这类药物进行改进,具体地说,增强其临床功效。正探索的一种方法是在基于抗体的药物中工程引入另外的新颖抗原结合位点,以便单一免疫球蛋白分子共接合两种不同抗原。接合两种不同抗原的这种非原生或取代抗体形式通常称为双特异性抗体。由于抗体可变区(fv)的可观多样性使得产生识别几乎任何分子的fv成为可能,因此产生双特异性的典型方法是将新可变区引入抗体中。已经探索出针对双特异性目标的多种取代抗体形式(chames及baty,2009,《mabs》1[6]:1-9;holliger及hudson,2005,《自然·生物技术(naturebiotechnology)》23[9]:1126-1136;kontermann,《mabs》4(2):182(2012),所有这些文献明确地均以引用的方式并入本文中)。首先,通过将两种各自产生单一单克隆抗体的细胞系融合来产生双特异性抗体(milstein等人,1983,《自然(nature)》305:537-540)。虽然所得杂交融合瘤或四源杂交瘤确实产生了双特异性抗体,但是其仅仅是较小群体,且分离出所期望的抗体需要进行广泛的纯化。针对此的工程改造方案是使用抗体片段产生双特异性抗体。由于这种片段缺乏全长抗体的复杂四级结构,因此可以将可变的轻链和重链连接成单一基因构筑体。已经产生了许多不同形式的抗体片段,包含双功能抗体、单链双功能抗体、串联scfv和fab2双特异性抗体(chames及baty,2009,《mabs》1[6]:1-9;holliger及hudson,2005,《自然·生物技术》23[9]:1126-1136;这些文献均明确地以引用的方式并入本文中)。虽然这些形式可以在细菌中高水平表达且由于其尺寸小而可以具有有利的渗透好处,但是其在活体内快速清除且会出现与其制备和稳定性有关的制造障碍。造成这些缺点的主要原因是,抗体片段典型地缺乏具有包含较大尺寸、高度稳定性以及结合能在血清中维持长半衰期(即,新生fc受体fcrn)或充当结合位点用于纯化(即,蛋白质a和蛋白质g)的各种fc受体和配体的相关功能特性的抗体恒定区。近期的研究已尝试通过将双重结合工程引入全长抗体样形式中来解决基于片段的双特异性抗体的缺点(wu等人,2007,《自然·生物技术》25[11]:1290-1297;ussn12/477,711;michaelson等人,2009,《mabs》1[2]:128-141;pct/us2008/074693;zuo等人,2000,《蛋白质工程(proteinengineering)》13[5]:361-367;ussn09/865,198;shen等人,2006,《生物化学杂志(jbiolchem)》281[16]:10706-10714;lu等人,2005,《生物化学杂志》280[20]:19665-19672;pct/us2005/025472;这些文献明确地以引用的方式并入本文中)。这些形式克服了抗体片段双特异性抗体的一些障碍,主要是因为其含有fc区。这些形式的一个明显缺点是,因为其在同型二聚恒定链上建立了新的抗原结合位点,所以对新抗原的结合总是二价的。对于在双特异性治疗形式中作为辅助目标而引人注目的多种抗原来说,所期望的结合是单价的,而非二价的。对于多种免疫受体来说,细胞活化是通过单价结合相互作用的交联来实现。交联机制典型地由抗体/抗原免疫复合物介导,或通过效应细胞与目标细胞的接合来介导。举例来说,低亲和力fcγ受体(fcγr),如fcγriia、fcγriib以及fcγriiia,与抗体fc区单价结合。单价结合不能活化表达这些fcγr的细胞;然而,在免疫复合或细胞与细胞接触后,受体发生交联且群集于细胞表面上,引起活化。对于负责介导细胞杀死的受体(例如天然杀手(nk)细胞上的fcγriiia)来说,当效应细胞与目标细胞按高度亲合形式接合时,发生受体交联和细胞活化(bowles及weiner,2005,《免疫学方法杂志(jimmunolmethods)》304:88-99,其明确地以引用的方式并入本文中)。类似地,在b细胞上的抑制性受体fcγriib仅当其与细胞表面b细胞受体(bcr)接合而形成免疫复合物时才下调b细胞活化,这是一种通过可溶性igg与bcr所识别的相同抗原发生免疫复合所介导的机制(heyman,2003,《免疫学通讯(immunollett)》88[2]:157-161;smith和clatworthy,2010,《自然综述免疫学(naturereviewsimmunology)》10:328-343;这些文献明确地以引用的方式并入本文中)。作为另一实例,t细胞的cd3活化仅当其相关t细胞受体(tcr)与抗原呈递细胞上的载抗原mhc按高度亲合的细胞到细胞突触形式接合时才发生(kuhns等人,2006,《免疫(immunity)》24:133-139)。实际上,使用抗cd3抗体发生的cd3非特异性二价交联诱发了细胞因子风暴和毒性(perruche等人,2009,《免疫学杂志(jimmunol)》183[2]:953-61;chatenoud及bluestone,2007,《自然综述免疫学》7:622-632;这些文献明确地以引用的方式并入本文中)。因此在实际临床使用中,用于再导向杀死目标细胞的cd3共接合的优选模式是单价结合,其仅在与共接合的目标接合后才活化。生长激素抑制素是充当内源性抑制调节剂的神经肽。生长激素抑制素具有广泛的细胞功能,如抑制多种分泌物、细胞增殖以及细胞存活(patel,1999,《神经内分泌学前沿(frontneuroendocrinol.)》20:157-198)。生长激素抑制素在中枢神经系统、外周神经系统、胰和肠道中广泛分布(参见,例如watt等人,2008,《分子细胞内分泌学(molcellendocrinol.)》286:251-261;epelbaum,1986,《神经生物学研究进展(prog.neurobiol.)》27:63-100;以及raynor,1992,《神经生物学评论(crit.rev.neurobiol.)》6:273-289)。生长激素抑制素还在神经内分泌肿瘤(net),如甲状腺髓质癌、神经母细胞瘤、神经节瘤、胰升糖素瘤、肾上腺皮质肿瘤以及出现在肺、副神经节、十二指肠中的肿瘤,以及一些其它非net中表达(volante等人,2008,《分子与细胞内分泌学(mol.cell.endocrinol.)》286:219-229)。生长激素抑制素可以通过直接活化生长抑素受体(sstr)对目标细胞造成影响(watt等人,2008,《分子细胞内分泌学》286:251-261;pyronnet等人,2008,《分子细胞内分泌学》286:230-237)。生长抑素受体(sstr)属于g蛋白偶联受体(gpcr)超家族,其各含有由胞外/胞内域和七个跨膜域组成的单个多肽链。sstr在多种培养的肿瘤细胞和原发肿瘤组织,包含net(肺癌、胃肠癌、胰脏癌、垂体癌、髓质癌、前列腺、胰脏肺类癌、骨肉瘤等)以及非net(乳癌、肺癌、结肠直肠癌、卵巢癌、子宫颈癌等)中高度表达(reubi.,2003,《内分泌学综述(endocr.rev.)》24:389-427;volante等人,2008,《分子与细胞内分泌学》286:219-229;以及schulz等人,2003,《妇科肿瘤学(gynecol.oncol.)》89:385-390)。迄今,已鉴别出五种sstr受体子类型(patel等人,1997,《内分泌学与代谢学趋势(trendsendocrinol.metab.)》8:398-405)。尤其sstr2在多种肿瘤细胞上以高浓度表达(volante等人,2008,《分子与细胞内分泌学》286:219-229;以及reubi等人,2003,《欧洲核医学与分子成像杂志(eur.j.nucl.med.mol.imaging)》30:781-793),因此使其成为用于双特异性抗体癌症靶向治疗剂的候选目标抗原。考虑到sstr2在多种肿瘤上高浓度表达,认为抗sstr2抗体适用于,例如,定位针对这种表达sstr2的肿瘤的抗肿瘤治疗剂(例如化疗剂和t细胞)。举例来说,认为sstr2和cd3的双特异性抗体(其能够定位表达sstr2的肿瘤的cd3+效应t细胞)适用于癌症治疗剂。然而由抗体片段产生的双特异性抗体遭受了生物物理学和药物动力学障碍,但是利用全长抗体样形式建构的那些抗体的缺点是,在主要目标抗原不存在的情况下,其与辅助目标抗原以多价方式接合,导致非特异性活化和潜在的毒性。本发明通过引入针对sstr2和cd3的新颖双特异性抗体来解决这一问题。技术实现要素:因此,本文提供生长抑素受体2(sstr2)抗原结合域和抗sstr2抗体(例如双特异性抗体)。一方面,本文提供sstr2“开瓶器”形式抗体,其包含:a)第一重链,其包含i)第一变体fc结构域和ii)单链fv区(scfv),其中scfv区包含第一重链可变域、第一轻链可变域以及带电scfv连接子,其中带电scfv连接子与第一重链可变域和第一轻链可变域共价连接;b)第二重链,其包含vh-ch1-铰链-ch2-ch3单体,其中vh是第二重链可变域且ch2-ch3是第二变体fc结构域;以及c)轻链,其包含第二轻链可变域和轻链恒定域。第二变体fc结构域包含氨基酸取代n208d/q295e/n384d/q418e/n421d,第一变体fc结构域和第二变体fc结构域各包含氨基酸取代e233p/l234v/l235a/g236del/s267k;第一变体fc结构域包含氨基酸取代s364k/e357q且第二变体fc结构域包含氨基酸取代l368d/k370s。另外,第二重链可变域包含seqidno:1071且第二轻链可变域包含seqidno:1076,其中编号根据如kabat中的eu索引。在sstr2“开瓶器”形式抗体的某些实施例中,scfv结合cd3。在一些实施例中,第一重链可变域和第一轻链可变域分别选自其包括以下的组:seqidno:1和seqidno:5;seqidno:10和seqidno:14;seqidno:19和seqidno:23;seqidno:28和seqidno:32;seqidno:37和seqidno:41;以及seqidno:46和seqidno:50。在一些实施例中,第一重链可变域包含seqidno:1且第一轻链可变域包含seqidno:5。在sstr2“开瓶器”形式抗体的某些实施例中,第二重链的ch1-铰链-ch2-ch3组分包含seqidno:1108,第一变体fc结构域包含seqidno:1109,且轻链恒定域包含seqidno:1110。在一些实施例中,第一重链包含seqidno:1080,第二重链包含seqidno:1070,且轻链包含seqidno:1075。另一方面本文中提供2型生长抑素受体(sstr2)抗原结合域,其包含具有seqidno:958的重链可变域和具有seqidno:962的轻链可变域。另一方面,本文中提供一种核酸组合物,其包含编码本文所述的任何异源二聚抗体或抗原结合域的核酸。另一方面,本文中提供一种表达载体,其包含本文所述的任何核酸。一方面,本文中提供经本文所述的任何表达载体或核酸转型的宿主细胞。另一方面,本文中提供一种制备本文所述的主题异源二聚抗体或抗原结合域的方法。所述方法包含如下步骤:在表达抗体或抗原结合域的条件下培养经本文所述的任何表达载体或核酸转型的宿主细胞,并且回收抗体或抗原结合域。一方面,本文中提供一种治疗癌症的方法,其包含向有需要的患者施用本文所述的任一主题抗体。在一些实施例中,癌症是神经内分泌癌。附图说明图1a至ii描绘本发明的几种形式。第一个是“开瓶器”形式,具有第一和第二抗抗原结合域。此外,mab-fv、mab-scfv、中心scfv、中心fv、单臂中心scfv、单scfv-mab、scfv-mab以及双scfv形式均显示出。对于所描绘的所有scfv结构域,其可以为n至c端重链可变-(任选的连接子)-轻链可变,或相反。此外,对于单臂scfv-mab,scfv可以连接于重链单体的n端或轻链的n端。在某些实施例中,图1中的“抗抗原1”是指抗sstr2结合域。在某些实施例中,图1中的“抗抗原1”是指抗cd3结合域。在某些实施例中,图1中的“抗抗原2”是指抗sstr2结合域。在某些实施例中,图1中的“抗抗原2”是指抗cd3结合域。在一些实施例中,图1中的“抗抗原1”是指抗sstr2结合域且图1中的“抗抗原2”是指抗cd3结合域。在一些实施例中,图1中的“抗抗原1”是指抗cd3结合域且图1中的“抗抗原2”是指抗sstr2结合域。图2描绘人类和食蟹猕猴(cynomolgusmonkey/macacafascicularis)sstr2蛋白的氨基酸序列。图3a-3f描绘异源二聚变体组的适用对(包含歪斜变体和pi变体)。在图3f上,存在无对应的“单体2”变体的变体;这些是pi变体,其可,例如,单独用于单体,或包含在开瓶器的fab侧上,且适当带电scfv连接子可用于利用scfv作为第二抗原结合域的第二单体。适合带电连接子显示于图7a和7b中。图4描绘一系列等排变体抗体恒定区和其对应的取代。pi_(-)表示较低pi变体,而pi_(+)表示较高pi变体。这些变体可以任选地且独立地与本发明的其它异源二聚变体(以及其它变体类型,如本文中所概述)组合。图5描绘消融fcγr结合的适用消融变体(有时称为“基因剔除”或“ko”变体)。图6显示本发明的两个特别适用的实施例。图7a和7b描绘多种带电scfv连接子,其应用于增加或降低如本文所述的利用一个或多个scfv作为组分的主题异源二聚抗体的pi。本文中尤其使用(+h)正性连接子,尤其与本文所示的抗cd3vl和vh序列一起使用。根据whitlow等人,《蛋白质工程(proteinengineering)》6(8):989-995(1993),具有单个电荷的单个现有技术scfv连接子称为“whitlow”。应注意,这种连接子用于减少scfv的聚集和增强scfv的蛋白水解稳定性。图8描绘多种可以与本文所述的异源二聚抗体一起使用的异源二聚歪斜变体氨基酸取代。图9a-9e显示几种基于人类igg1且不含fv序列(例如scfv和fab侧的vh和vl)的适用开瓶器形式骨架的序列。开瓶器骨架1基于人类igg1(356e/358m同种异型),且包含s364k/e357q:l368d/k370s歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体以及在两个链上的e233p/l234v/l235a/g236del/s267k消融变体。开瓶器骨架2基于人类igg1(356e/358m同种异型)且包含不同歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体以及在两个链上的e233p/l234v/l235a/g236del/s267k消融变体。开瓶器骨架3基于人类igg1(356e/358m同种异型),且包含不同歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体以及在两个链上的e233p/l234v/l235a/g236del/s267k消融变体。开瓶器骨架4基于人类igg1(356e/358m同种异型)且包含不同歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体以及在两个链上的e233p/l234v/l235a/g236del/s267k消融变体。开瓶器骨架5基于人类igg1(356d/358l同种异型),且包含s364k/e357q:l368d/k370s歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体以及在两个链上的e233p/l234v/l235a/g236del/s267k消融变体。开瓶器骨架6基于人类igg1(356e/358m同种异型),且包含s364k/e357q:l368d/k370s歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体以及在两个链上的e233p/l234v/l235a/g236del/s267k消融变体,以及在两个链上的n297a变体。开瓶器骨架7与开瓶器骨架6一致,不同在于突变是n297s。开瓶器骨架6和7的取代形式可以不包含两个链中的消融变体e233p/l234v/l235a/g236del/s267k。骨架8基于人类igg4,且包含s364k/e357q:l368d/k370s歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体以及在两个链上的e233p/l234v/l235a/g236del/s267k消融变体,以及在两个链上的s228p(eu编号,此为kabat中的s241p)变体,如所属领域中已知,其消融fab臂交换。开瓶器骨架8的取代形式可以不包含两个链上的消融变体e233p/l234v/l235a/g236del/s267k。骨架9基于人类igg2,且包含s364k/e357q:l368d/k370s歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体。骨架10基于人类igg2,且包含s364k/e357q:l368d/k370s歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体以及在两个链上的s267k变体。如将为所属领域的技术人员所了解且在下文中概述,这些序列可以与本文概述的任何vh和vl对一起使用,其中一个单体包含scfv(任选地包含带电scfv连接子)且另一单体包含fab序列(例如vh连接于“fab侧重链”且vl连接于“恒定轻链”)。即,本文概述的用于抗sstr2和抗cd3的任何fv序列,无论作为scfv(同样,任选地具有带电scfv连接子)或是作为fab,均可以任何组合并入到这些图9骨架中。图9a中描绘的恒定轻链可用于图中所有构筑体,尽管κ恒定轻链也可以被取代。应注意,这些开瓶器骨架被应用于图1f的中心scfv形式,其中具有与第一fab相同的抗原结合的另外的第二fab(vh-ch1和vl-恒定轻链)添加到“开瓶器侧”的scfv的n端。这些骨架中的每一个包含如下的序列:与所列举的序列90、95、98以及99%一致(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代(相比于图的“亲本”,如将为所属领域的技术人员所了解,相比于亲本人类igg1(或igg2或igg4,取决于骨架)其已含有多个氨基酸修饰)。即,所列举的骨架除此图的骨架内所含有的歪斜、pi以及消融变体以外可含有另外的氨基酸修饰(通常氨基酸取代)。图10a至10d显示本发明中使用的mab-scfv骨架的序列,其中添加了本发明的fv序列。mab-scfv骨架1基于人类igg1(356e/358m同种异型),且包含s364k/e357q:l368d/k370s歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体以及在两个链上的e233p/l234v/l235a/g236del/s267k消融变体。骨架2基于人类igg1(356d/358l同种异型),且包含s364k/e357q:l368d/k370s歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体以及在两个链上的e233p/l234v/l235a/g236del/s267k消融变体。骨架3基于人类igg1(356e/358m同种异型),且包含s364k/e357q:l368d/k370s歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体以及在两个链上的e233p/l234v/l235a/g236del/s267k消融变体,以及在两个链上的n297a变体。骨架4与骨架3一致,不同在于突变是n297s。mab-scfv骨架3和4的取代形式可以不包含两个链中的消融变体e233p/l234v/l235a/g236del/s267k。骨架5基于人类igg4,且包含s364k/e357q:l368d/k370s歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体以及在两个链上的e233p/l234v/l235a/g236del/s267k消融变体,以及在两个链上的s228p(eu编号,此为kabat中的s241p)变体,如所属领域中已知,其消融fab臂交换。骨架6基于人类igg2,且包含s364k/e357q:l368d/k370s歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体。骨架7基于人类igg2,且包含s364k/e357q:l368d/k370s歪斜变体、在fab侧的n208d/q295e/n384d/q418e/n421dpi变体以及在两个链上的s267k变体。如将为所属领域的技术人员所了解且在下文中概述,这些序列可以与本文概述的任何vh和vl对一起使用,其中一个单体包含fab和scfv(任选地包含带电scfv连接子)且另一单体包含fab序列(例如vh连接于“fab侧重链”且vl连接于“恒定轻链”)。即,本文概述的用于抗sstr2和抗cd3的任何fv序列,无论作为scfv(同样,任选地具有带电scfv连接子)或是作为fab,均可以任何组合并入到此图10骨架中。单体1侧是fab-scfvpi负性侧,且包含异源二聚变体l368d/k370s、等排pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k(均相对于igg1)。单体2侧是scfvpi正性侧,且包含异源二聚变体364k/e357q。然而,其它可以被取代,尤其[s364k/e357q:l368d/k370s];[l368d/k370s:s364k];[l368e/k370s:s364k];[t411t/e360e/q362e:d401k];[l368d/k370s:s364k/e357l]、[k370s:s364k/e357q]、[t366s/l368a/y407v:t366w]以及[t366s/l368a/y407v/y394c:t366w/s354c]。图10a中描绘的恒定轻链可用于图中所有构筑体,尽管κ恒定轻链也可以被取代。应注意,这些mab-scfv骨架被应用于图1h的mab-fv形式(其中一个单体包括位于c端处的vl且另一单体包括位于c端处的vh)以及图1e的scfv-mab形式(其中scfv结构域添加到其中一个单体的c端)。这些骨架中的每一个包含如下的序列:与所列举的序列90、95、98以及99%一致(如本文所定义),和/或含有1、2、3、4、5、6、7、8、9或10个另外的氨基酸取代(相比于图的“亲本”,如将为所属领域的技术人员所了解,相比于亲本人类igg1(或igg2或igg4,取决于骨架)其已含有多个氨基酸修饰)。即,所列举的骨架除此图的骨架内所含有的歪斜、pi以及消融变体以外可含有另外的氨基酸修饰(通常氨基酸取代)。图11a至11g描绘本文所述的示范性的主题抗sstr2抗原结合域的氨基酸序列,其包含:抗sstr2h1.143_l1.30;抗sstr2h1_l1.1;抗sstr2h1.107_l1.30;抗sstr2h1.107_l1.67;抗sstr2h1.107_l1.108;抗sstr2h1.107_l1.111;抗sstr2h1.107_l1.114;抗sstr2h1.107_l1.102;抗sstr2h1.107_l1.110;抗sstr2h1.125_l1.30;抗sstr2h1.125_l1.67;抗sstr2h1.125_l1.108;抗sstr2h1.125_l1.111;抗sstr2h1.125_l1.114;抗sstr2h1.125_l1.102;以及抗sstr2h1.125_l1.10。所描绘的序列包含用于各抗原结合域的重链可变(vh)域和轻链可变(vl)域序列。对于各vh序列,vhcdr1、vhcdr2以及vhcdr3序列带下划线且呈蓝色。对于各vl序列,vlcdr1、vlcdr2以及vlcdr3序列带下划线且呈蓝色。如本文中所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包含带下划线的cdr且还包含使用其它编号系统的vh和vl域内所包含的cdr。此外,至于图中的所有序列,这些vh和vl序列可以scfv形式或以fab形式使用。图12a至12f描绘多种可用于本文所提供的主题抗体的抗cd3抗原结合域(例如抗cd3scfv)。cdr是带下划线的,scfv连接子是带双下划线的(在序列中,scfv连接子是带正电的scfv(gkpgs)4连接子,尽管如将为所属领域的技术人员所了解,此连接子可以替换为其它连接子,包含不带电或带负电连接子,其中一些在图7中描绘。如上,命名规则说明了由n端至c端的scfv的定向;在此图中所列出的序列中,其均以vh-scfv连接子-vl(由n端至c端)方式定向,尽管这些序列还可以相反定向,即(由n端至c端)vl-连接子-vh使用。如本文中所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包含带下划线的cdr且还包含使用其它编号系统的vh和vl域内所包含的cdr。此外,至于图中的所有序列,这些vh和vl序列可以scfv形式或以fab形式使用。图12a描绘了“高cd3”抗cd3_h1.30_l1.47构筑体的序列,其包含重链和轻链可变域(cdr带下划线),以及个别vl和vhcdr,以及具有带电连接子(带双下划线)的scfv构筑体。图中所描绘的所有序列均是如此,这种带电连接子可以在需要时替换为不带电连接子或不同的带电连接子。图12b描绘“高int#l”抗cd3_h1.32_l1.47构筑体的序列,其包含重链和轻链可变域(cdr带下划线),以及个别vl和vhcdr,以及具有带电连接子(带双下划线)的scfv构筑体。图中所描绘的所有序列均是如此,这种带电连接子可以在需要时替换为不带电连接子或不同的带电连接子。图12c描绘“高int#2”抗cd3_h1.89_l1.47构筑体的序列,其包含重链和轻链可变域(cdr带下划线),以及个别vl和vhcdr,以及具有带电连接子(带双下划线)的scfv构筑体。图中所描绘的所有序列均是如此,这种带电连接子可以在需要时替换为不带电连接子或不同的带电连接子。图12d描绘“高int#3”抗cd3_h1.90_l1.47,其包含重链和轻链可变域(cdr带下划线),以及个别vl和vhcdr,以及具有带电连接子(带双下划线)的scfv构筑体。图中所描绘的所有序列均是如此,这种带电连接子可以在需要时替换为不带电连接子或不同的带电连接子。图12e描绘“int”抗cd3_h1.33_l1.47构筑体的序列,其包含重链和轻链可变域(cdr带下划线),以及个别vl和vhcdr,以及具有带电连接子(带双下划线)的scfv构筑体。图中所描绘的所有序列均是如此,这种带电连接子可以在需要时替换为不带电连接子或不同的带电连接子。图12f描绘“低”抗cd3_h1.31_l1.47构筑体的序列,其包含重链和轻链可变域(cdr带下划线),以及个别vl和vhcdr,以及具有带电连接子(带双下划线)的scfv构筑体。图中所描绘的所有序列均是如此,这种带电连接子可以在需要时替换为不带电连接子或不同的带电连接子。图13a-13z描绘可以与本文所述的主题双特异性抗体一起使用的、稳定性优化的人源化抗cd3变体scfv变体的氨基酸序列(例如抗sstr2x抗cd3“开瓶器”抗体)。cdr是带下划线的。对于各重链/轻链组合,列出四个序列:(i)具有c端6xhis标签的scfv,(ii)scfv单独,(iii)vh单独,(iv)vl单独。如本文中所提及且对含有cdr每一序列而言确实如此,取决于所使用的编号方法(如表1中所示),cdr位置的精确标识可能略微不同,且因此本文中不仅包含带下划线的cdr且还包含使用其它编号系统的vh和vl域内所包含的cdr。此外,至于图中的所有序列,这些vh和vl序列可以scfv形式或以fab形式使用。图14a和14b描绘本文所述的示范性的抗sstr2×抗cd3“开瓶器”双特异性抗体xenp018087(sstr2h1.143_l1.30和cd3h1.30_l1.47)的氨基酸序列。对于sstr2fab-fc重链序列,vhcdrl-3是带下划线的且呈蓝色,并且重链可变域与ch1-铰链-ch2-ch3之间的边界通过“/”指示。对于cd3scfv-fc重链序列,各个结构域之间的边界使用“/”指示且如下:scfv重链可变域/scfv连接子/scfv轻链域/fc结构域。vhcdrl-3和vlcdr1-3带蓝色下划线。对于各scfv-fc结构域,vhcdr1-3和vlcdr1-3序列是带下划线的且呈蓝色。对于cd3轻链序列,vlcdr1-3是带下划线的且呈蓝色,并且轻链可变域与轻链恒定域之间的边界通过“/”指示。所描绘的带电连接子是(gkpgs)4,尽管可使用其它带电或不带电连接子,如图7a和7b中所描绘的连接子。此外,本文概述的各序列可在一个fc结构域或优选两个fc结构域中包含或不包含m428l/n434s变体,所述变体引起在血清中半衰期变长。图15a-15r描绘本文所述的另外的示范性的抗sstr2×抗cd3“开瓶器”双特异性抗体,其包含xenp018907(图15a和15b,sstr2h1.143_l1.30和cd3h1.32_l1.47)的氨基酸序列。对于sstr2fab-fc重链序列,vhcdrl-3是带下划线的且呈蓝色,并且重链可变域与ch1-铰链-ch2-ch3之间的边界通过“/”指示。对于cd3scfv-fc重链序列,各个结构域之间的边界使用“/”指示且如下:scfv重链可变域/scfv连接子/scfv轻链域/fc结构域。vhcdrl-3和vlcdr1-3带蓝色下划线。对于各scfv-fc结构域,vhcdr1-3和vlcdr1-3序列是带下划线的且呈蓝色。对于cd3轻链序列,vlcdr1-3是带下划线的且呈蓝色,并且轻链可变域与轻链恒定域之间的边界通过“/”指示。所描绘的带电连接子是(gkpgs)4,尽管可使用其它带电或不带电连接子,如图7a和7b中所描绘的连接子。此外,本文概述的各序列可在一个fc结构域或优选两个fc结构域中包含或不包含m428l/n434s变体,所述变体引起血清中半衰期变长。图16a-16c描绘本文所述的示范性的双特异性抗sstr2×抗cd3抗体的可能组合的矩阵。“a”意指位于矩阵上部的所参考的cd3结合域序列的cdr可以与在矩阵左侧列出的sstr2结合域序列的cdr组合。举例来说,相对于“抗sstr2h1.143_l1.30”和“抗cd3h1.30_l1.47”,“a”表示包含以下的双特异性抗体:a)具有可变重链cd3h1.30序列的vhcdr和可变轻链cd3l1.47序列的vlcdr的cd3结合域,和b)具有sstr2h1.143序列的vhcdr和sstr2l1.30序列的vlcdr的sstr2结合域。“b”意指cd3结合域构筑体的cdr可以与sstr2结合域构筑体的重链可变域和轻链可变域组合。举例来说,相对于“抗sstr2h1.143_l1.30”和“抗cd3h1.30_l1.47”,“b”表示包含以下的双特异性抗体:a)具有可变重链cd3h1.30序列的vhcdr和可变轻链cd3l1.47序列的vlcdr的cd3结合域,和b)具有s重链可变域sstr2h1.143序列和轻链可变域sstr2l1.30序列的sstr2结合域。“c”表示包含以下的双特异性抗体:a)具有抗cd3序列的重链可变域和轻链可变域的cd3结合域,和b)具有抗sstr2序列的cdr的sstr2结合域。“d”表示包含以下的双特异性抗体:具有指定抗sstr2序列的可变重链和可变轻链的sstr2结合域,和具有指定抗cd3序列的可变重链和可变轻链的cd3结合域。“e”表示包含scfv的双特异性抗体,其中cd3的scfv与sstr2的cdr一起使用。“f”表示包含scfv的双特异性抗体,其中cd3的scfv与sstr2抗原结合域的重链可变域和轻链可变域一起使用。所有这些组合可以开瓶器形式进行,例如,具有图9中所示的任何骨架形式,或以取代形式,如图1的mab-fv、mab-scfv、中心scfv、中心fv或双scfv形式,包含图26中所示的骨架形式。举例来说,“a”(cd3cdr和sstr2cdr)可以被添加到开瓶器序列(包含图9的那些序列)或包含有不同的异源二聚变体,或添加到图10的mab-scfv骨架、中心scfv、mab-fv形式或中心fv形式中。然而一般而言,会包含cd3的二价结合的形式是不利的。图16d-16f描绘本文所述的示范性的双特异性抗sstr2×抗cd3开瓶器形式组合的可能组合的矩阵。在这些矩阵中,抗cd3scfv在x轴中列出且抗sstr2fab在y轴中列出。“a”意指位于矩阵上部的所参考的cd3结合域序列的cdr可以与在矩阵左侧列出的sstr2结合域序列的cdr组合。举例来说,相对于“抗sstr2h1.143_l1.30”和“抗cd3h1.30_l1.47”,“a”表示包含以下的双特异性开瓶器形式抗体:a)具有可变重链cd3h1.30序列的vhcdr和可变轻链cd3l1.47序列的vlcdr的抗cd3scfv,和b)具有sstr2h1.143序列的vhcdr和sstr2l1.30序列的vlcdr的抗sstr2fab。“b”意指cd3结合域构筑体的cdr可以与sstr2结合域构筑体的重链可变域和轻链可变域组合。举例来说,相对于“抗sstr2h1.143_l1.30”和“抗cd3h1.30_l1.47”,“b”表示包含以下的双特异性开瓶器抗体:a)具有可变重链cd3h1.30序列的vhcdr和可变轻链cd3l1.47序列的vlcdr的抗cd3scfv,和b)具有s重链可变域sstr2h1.143序列和轻链可变域sstr2l1.30序列的抗sstr2fab。“c”表示包含以下的双特异性开瓶器抗体:a)具有抗cd3序列的重链可变域和轻链可变域的抗cd3scfv,和b)具有抗sstr2序列的cdr的sstr2fab。“d”表示包含以下的双特异性开瓶器抗体:具有指定抗sstr2序列的可变重链和可变轻链的抗sstr2fab,和具有指定抗cd3序列的可变重链和可变轻链的抗cd3scfv。图17a-17p描绘使用人类sstr2转染的cho细胞的示范性抗sstr2抗体和抗sstr2×抗cd3双特异性抗体的细胞表面结合分析。通过流式细胞术使用经藻红蛋白(pe)标记的二级抗体测量结合。图18a-18d描绘使用抗sstr2×抗cd3双特异性抗体和人类sstr2转染的cho细胞的再导向t细胞细胞毒性(redirectedtcellcytotoxicity,rtcc)分析的结果。图19a-19c描绘使用抗sstr2×抗cd3双特异性抗体与tt细胞(人类甲状腺髓质癌细胞株,图19a-19c)的再导向t细胞细胞毒性(rtcc)分析的结果。图20a和20b描绘在食蟹猕猴中抗sstr2×抗cd3双特异性抗体对cd4+和cd8+t细胞活化(图20a)和cd4+和cd8+t细胞分布(图20b)的影响的研究。图21a-21d描绘在食蟹猕猴中抗sstr2×抗cd3双特异性抗体对cd4+和cd8+t细胞活化(图21a)和cd4++和cd8++t细胞分布(图21b)的影响的另外的研究。此外,进行葡萄糖耐量测试(glucosetolerancetest,gtt)(图21c和21d)以评估所测试的受试者分解葡萄糖的能力。图22a-22f描绘示范性抗sstr2×抗cd3双特异性抗体对cd4+和cd8+t细胞活化(图22a和22b)、cd4+和cd8+t细胞分布(图22c和22d)以及血清il-6和tnfα的血清含量(图22e和22f)的另外的研究。图23a-23c描绘xmab18087和xenp13245在人类sstr2转染的cho细胞(图22a)、食蟹猕猴sstr2转染的cho细胞(图22b)以及未转染的亲本cho细胞(图22c)上的细胞表面结合分析。图24a-24c描绘使用xmab18087(方块)和xenp13245(圆圈)与人类sstr2转染的cho细胞(图24a)、tt细胞(人类甲状腺髓质癌细胞株,图28b)或a548细胞(肺腺癌细胞株,图24c)的再导向t细胞细胞毒性(rtcc)分析的结果。图25描绘使用抗sstr2×抗cd3双特异性和对照抗sstr2mah和抗rsv×抗cd3与tt细胞(人类甲状腺髓质癌细胞株)或a548细胞(肺癌)的再导向t细胞细胞毒性(rtcc)分析的结果。图26a-26b描绘在图2中描述的实验中在24小时后在使用人类sstr2转染的cho细胞培育的cd4+和cd8+t细胞(图29a)和tt细胞(图29b)上的cd69的上调。实心数据点显示cd8+t细胞在的cd69mfi且空心数据点显示cd4+t细胞上的cd69mfi。图27描绘用以检验xmab18087的抗肿瘤活性的小鼠研究的设计。图28a-28b描绘通过测量的肿瘤尺寸,其随时间和处理而变。图29描绘生物发光图像(给药#1后第28天)。图30a-30b描绘在食蟹猕猴中xmab18087对cd4+(图30a)和cd8+(图30b)t细胞分布的影响的研究。图31a-31b描绘在食蟹猕猴中xmab18087对cd4+(图31a)和cd8+(图31b)t细胞活化的影响的研究。图32a-32b描绘在食蟹猕猴中xmab18087对血清il-6和tnf的含量的影响。图33描绘如由测量的移植有a549-redfluc肿瘤细胞和人类pbmc的nsg小鼠中的肿瘤尺寸,其随时间和使用多种浓度xmab18087的处理而变。具体实施方式a.材料的并入图和图例所有ussn62/481,065;62/397,322;62/355,821以及62/355,820的图和随附图例明确地独立地以引用的方式整体并入本文中,尤其是其中所描绘的氨基酸序列。序列如下参考随附序列表。适用作abd的抗sstr2序列包含seqidno:958-1069(图11)和seqidno:58至659的抗sstr2重链和轻链序列的重链可变域、轻链可变域以及cdr。适用作abd的抗cd3序列包含seqidno:1-54(图12)和seqidno:835至938中所包含的重链可变域、轻链可变域以及cdr。所述重链可变域、轻链可变域以及cdr可以包含在本文所述的主题抗体和抗原结合域的scfv或fv形式中。示范性双特异性sstr2×cd3抗体的序列包含在seqidno:1070至1088(图14);以及seqidno:1089至1107和660至806(图15)中b.概述本文提供适用于治疗癌症的抗sstr2抗体。因为在神经内分泌肿瘤(net,例如肺癌、胃肠癌、胰脏癌、垂体癌、髓质癌、前列腺、胰脏肺类癌、骨肉瘤等)以及非net(乳癌、肺癌、结肠直肠癌、卵巢癌、子宫颈癌等)中高度表达,认为抗sstr2抗体适用于定位针对这种表达sstr2的肿瘤的抗肿瘤治疗剂(例如化疗剂和t细胞)。特定来说,本文提供抗cd3、抗sstr2双特异性抗体。这种抗体用于使cd3+效应t细胞导向于sstr2+肿瘤,从而允许cd3+效应t细胞攻击并溶解sstr2+肿瘤。已经设计出与cd3和肿瘤抗原目标共接合的抗双特异性抗体且用于再导向t细胞攻击并溶解所靶向的肿瘤细胞。实例包含与cd3和肿瘤抗原单价接合的bite和dart形式。虽然靶向cd3的方法已经显示出可观的前景,但是此类疗法的常见副作用是与其相关地产生细胞因子,通常导致毒性细胞因子释放综合症。由于双特异性抗体的抗cd3结合域与所有t细胞接合,因此募集产生大量细胞因子的cd4t细胞亚群。而且,cd4t细胞亚群包含调节性t细胞,其募集和扩增可以潜在地引起免疫抑制且对长期肿瘤抑制具有负面影响。此外,这些形式不含fc结构域且在患者中显示出非常短的血清半衰期。虽然靶向cd3的方法已经显示出可观的前景,但是此类疗法的常见副作用是与其相关地产生细胞因子,通常导致毒性细胞因子释放综合症。由于双特异性抗体的抗cd3结合域与所有t细胞接合,因此募集产生大量细胞因子的cd4t细胞亚群。而且,cd4t细胞亚群包含调节性t细胞,其募集和扩增可以潜在地引起免疫抑制且对长期肿瘤抑制具有负面影响。减少细胞因子产生且可能减少cd4t细胞活化的一种可行方式是通过减小抗cd3域对cd3的亲和力。因此,在一些实施例中,本发明提供包括抗cd3抗原结合域的抗体构筑体,其是cd3的“强”或“高亲和力”结合子(例如,一个实例是描绘为h1.30_l1.47的重链可变域和轻链可变域(任选地包含带电连接子(适当时)))并且还结合sstr2。在其它实施例中,本发明提供包括抗cd3抗原结合域的抗体构筑体,其是cd3的“弱”或“低亲和力”结合子。另外的实施例提供包括抗cd3抗原结合域的抗体构筑体,其对cd3具有中度或“中等”亲和力且还结合cd38。亲和力通常使用biacore分析测量。应了解,本发明的“高、中、低”抗cd3序列可以多种异源二聚形式使用。虽然本公开的大部分内容使用异源二聚体的“开瓶器”形式,但是这些可变重链和轻链序列以及scfv序列(以及包含这些可变重链和轻链序列的fab序列)可以其它形式使用,如wo公开号2014/145806的图2中所描绘的形式,其中的图式、形式和图例明确地以引用的方式并入本文中。因此,一方面,本文提供结合于两种不同抗原的异源二聚抗体,例如由于其结合两种不同目标抗原,通常sstr2,所以所述抗体是“双特异性”的,如下文所描述。这些异源二聚抗体可以单价(例如存在单个抗原结合域,如重链和轻链可变域对)或双价(存在两个各自独立地结合抗原的抗原结合域)结合这些目标抗原。本文提供的异源二聚抗体是基于使用不同单体,所述不同单体含有“歪斜”形成异源二聚体而非同型二聚体的氨基酸取代,如下文更充分地描述;以及允许简单地从同型二聚体中纯化异源二聚体的“pi变体”,如下文类似地描述。提供的异源二聚双特异性抗体通常依赖于使用工程改造的fc结构域或变体fc结构域,所述fc结构域可以在生产细胞中自组装以产生异源二聚蛋白质;和生成和纯化这种异源二聚蛋白质的方法。c.命名法本发明的双特异性抗体以几种不同形式列出。各多肽被给予唯一的“xenp”编号,但是如在所属领域中将了解的,较长序列可能含有较短编号。举例来说,对于给定序列,开瓶器形式的scfv侧单体的重链将具有第一xenp编号,而scfv结构域将具有不同的xenp编号。一些分子具有三个多肽,因此使用xenp编号与组分作为名称。因此,分子xenp18087,其呈开瓶器形式,包括三个序列:“xenp18087hc-fab”(图14a,称为“sstr2fab-fc重链”);“xenp18087hc-scfv”(图14b,称为“cd3scfv-fc重链”);以及“xenp18087lc”(图14a,称为“sstr2轻链”)或等效物,尽管所属领域的技术人员将能够轻松地经由序列比对来鉴别这些。这些xenp编号出现在序列表以及标识符中,且用于图中。此外,一个包括三个组分的分子产生多个序列标识符。举例来说,fab单体的序列表具有全长序列、可变重链序列以及可变重链序列的三个cdr;轻链具有全长序列、可变轻链序列以及可变轻链序列的三个cdr;且scfv-fc结构域具有全长序列、scfv序列、可变轻链序列、3个轻链cdr、scfv连接子、可变重链序列以及3个重链cdr;应注意具有scfv结构域的本文中所有分子使用单个带电scfv连接子(+h),尽管可使用其它。此外,特定可变域的术语命名使用“hx.xx_ly.yy”类形式,其中编号作为特定可变链序列的唯一标识符。因此,xenp18087的fab侧的可变域是“h1.143_l1.30”,其表示重链可变域h1.143与轻链域l1.30组合。在这些序列按scfv形式使用的情况下,名称“h1.143_l1.30”表示重链可变域h1.143与轻链域luo组合,且由n端至c端呈vh-连接子-vl定向。具有与重链和轻链可变域一致但呈相反顺序的序列的这种分子将命名为“l1.30_h1.143”。类似地,不同构筑体可能“混合并匹配”重链和轻链,如从序列表和图中将显而易见的。d.定义为了能更全面地了解本申请,下文阐述若干定义。此类定义意图涵盖语法等效物。“消融”在本文中意指活性的减少或除去。因此举例来说,“消融fcγr结合”意指相比于不含特定变体的fc区,fc区氨基酸变体具有小于50%起始结合,优选具有大于70-80-90-95-98%活性损失,且一般而言,活性低于biacore、spr或bli分析中的可侦测结合的水平。fcγr结合的消融的特定用途是图5中所展示的那些,其通常被添加到两个单体中。如本文所使用的「adcc」或「抗体依赖性细胞介导的细胞毒性」意指如下的细胞介导的反应,其中表达fcγr的非特异性细胞毒性细胞识别目标细胞上的结合的抗体且随后引起目标细胞溶解。adcc与结合fcγriiia相关;与fcγriiia结合增加引起adcc活性增加。如本文所使用的「adcp」或「抗体依赖性细胞介导的吞噬」意指如下的细胞介导的反应,其中表达fcγr的非特异性吞噬细胞识别目标细胞上的结合的抗体且随后引起目标细胞的吞噬。本文中的“抗原结合域”或“abd”意指六个互补决定区(complementarydeterminingregion,cdr)的组,当作为多肽序列的一部分存在时,特异性结合如本文所论述的目标抗原。因此,“检查点抗原结合域”结合如本文所概述的目标检查点抗原。如所属领域中已知,这些cdr通常以可变重链cdr(vhcdr或vhcdr)的第一组和可变轻链cdr(vlcdr或vlcdr)的第二组的形式存在,对于重链而言各自包括三个cdr:vhcdr1、vhcdr2、vhcdr3,且对于轻链而言各自包括vlcdr1、vlcdr2以及vlcdr3。所述cdr分别存在于重链可变域和轻链可变域中,且一起形成fv区。(cdr编号方案参见表1和上文相关论述)。因此,在一些情况下,抗原结合域的六个cdr由重链可变域和轻链可变域提供。在“fab”形式中,所述组6个cdr由两个不同多肽序列提供:重链可变域(vh或vh;含有vhcdr1、vhcdr2以及vhcdr3)和轻链可变域(vl或vl;含有vlcdr1、vlcdr2以及vlcdr3),其中vh域的c端连接于重链的ch1域的n端且vl域的c端连接于轻链恒定域的n端(且因此形成轻链)。在scfv形式中,vh和vl域共价连接(通常通过使用如本文所概述的连接子)到单个多肽序列中,其可为(由n端开始)vh-连接子-vl或vl-连接子-vh,通常优选前者(包含位于各侧的任选的域连接子,取决于所使用的形式(例如根据图1)。一般来说,在第二单体中scfv结构域的c端连接于铰链的n端。本文中“修饰”意指在多肽序列中的氨基酸取代、插入和/或缺失或化学连接于蛋白质的部分的改变。举例来说,修饰可为连接于蛋白质的改变的碳水化合物或peg结构。本文中“氨基酸修饰”意指在多肽序列中氨基酸取代、插入和/或缺失。出于清楚起见,除非另外说明,否则氨基酸修饰总是针对由dna编码的氨基酸,例如具有dna和rna中的密码子的20种氨基酸。“氨基酸取代”或“取代”在本文中意指用不同氨基酸替换亲本多肽序列中特定位置处的氨基酸。具体来说,在一些实施例中,取代是针对特定位置处的并非天然产生的氨基酸,不是在生物体内或任何生物体内天然产生的。举例来说,取代e272y是指变体多肽,在此情况下是fc变体,其中在位置272处的谷氨酸被酪氨酸替换。为了清楚起见,已经过工程改造以改变核酸编码序列但不改变起始氨基酸(例如将cgg(编码精氨酸)换成cga(仍然编码精氨酸)来提高宿主生物体表达水平)的蛋白质不是“氨基酸取代”;也就是说,尽管创建了编码相同蛋白质的新基因,但是如果蛋白质在特定起始位置处具有相同氨基酸,那么它就不是氨基酸取代。如本文所使用的“氨基酸插入”或“插入”意指在亲本多肽序列中在特定位置处添加氨基酸序列。举例来说,-233e或233e表示在位置233之后且位置234之前插入谷氨酸。此外,-233ade或a233ade表示在位置233之后且位置234之前插入alaaspglu。如本文所使用的“氨基酸缺失”或“缺失”意指在亲本多肽序列中在特定位置处去除氨基酸序列。举例来说,e233-或e233#;e233()或e233del表示233位置处的谷氨酸缺失。此外,eda233-或eda233#表示从位置233开始序列gluaspala缺失。如本文所使用的“变体蛋白质”或“蛋白质变体”或“变体”意指借助于至少一个氨基酸修饰而不同于亲本蛋白质的蛋白质。相比于亲本蛋白质,蛋白质变体具有至少一个氨基酸修饰,但又不过多,否则使得变体蛋白质无法使用如下文所描述的比对程序与亲本蛋白质比对。一般而言,使用下文所描述的比对程序,如blast,变体蛋白质(如本文所概述的变体fc结构域等,通常与亲本蛋白质至少75、80、85、90、91、92、93、94、95、96、97、98或99%一致。如下文所描述,在一些实施例中,亲本多肽,例如fc亲本多肽,是人类野生型序列,如igg1、igg2、igg3或igg4的重链恒定域或fc区,尽管具有变体的人类序列也可以充当“亲本多肽”,例如可以包含us公开案2006/0134105的igg1/2杂合体。本文中的蛋白质变异序列将优选具有与亲本蛋白质序列的至少约80%的一致性,且最优选至少约90%一致性,更优选至少约95-98-99%一致性。因此,如本文所使用的“抗体变体”或“变体抗体”意指借助于至少一个氨基酸修饰而不同于亲本抗体的抗体,如本文所使用的“igg变体”或“变体igg”意指借助于至少一个氨基酸修饰而不同于亲本igg的抗体(同样,在许多情况下,来自人类igg序列),且如本文所使用的“免疫球蛋白变体”或“变体免疫球蛋白”意指借助于至少一个氨基酸修饰而不同于亲本免疫球蛋白序列的免疫球蛋白序列。如本文所使用的“fc变体”或“变体fc”意指相比于人类igg1、igg2或igg4的fc结构域,在fc结构域中包括氨基酸修饰的蛋白质。本发明的fc变体根据构成其的氨基酸修饰来定义。因此,举例来说,n434s或434s是相对于亲本fc多肽在位置434处具有丝氨酸取代的fc变体,其中编号是根据eu索引。同样,m428l/n434s定义相对于亲本fc多肽具有取代m428l和n434s的fc变体。wt氨基酸的身份标识可为非特定的,在此情况下前述变体称为428l/434s。应注意,提供取代的顺序是任意的,也就是说,例如n434s/m428l与m428l/n434s是相同的fc变体,以此类推。关于本发明中所讨论的与抗体有关的所有位置,除非另外指出,否则氨基酸位置编号是根据eu索引。eu索引或如在kabat或eu编号方案中的eu索引是指eu抗体的编号。kabat等人收集了重链和轻链可变区的许多初级序列。根据序列的保守度,其将个别初级序列分类成cdr和框架且制成其列表(参见《免疫关注的序列(sequencesofimmunologicalinterest)》,第5版,nih公开案,第91-3242号,e.a.kabat等人,其以引用的方式整体并入本文中)。还参见edelman等人,1969,《美国国家科学院院刊(procnatlacadsciusa)》63:78-85,特此以引用的方式整体并入本文中。修饰可为添加、缺失或取代。本文中“蛋白质”意指至少两个共价连接的氨基酸,其包含蛋白质、多肽、寡肽和肽。此外,构成本发明的抗体的多肽可以包含一个或多个侧链或末端的合成衍生作用、糖基化、peg化、环形排列、环化、与其它分子的连接子、蛋白质或蛋白质域的融合以及肽标签或标记的添加。如本文所使用的“残基”意指蛋白质中的位置和其相关的氨基酸身份标识。举例来说,天冬酰胺297(亦称为asn297或n297)是人类抗体igg1中位置297处的残基。如本文所使用的“fab”或“fab区”意指包括vh、chi、vl以及cl免疫球蛋白域的多肽,所述域通常位于两个不同的多肽链上(例如vh-ch1位于一个链上而vl-cl位于另一个链上)。fab可能指隔离情况下的这种区或在本发明的双特异性抗体的情况下的这种区。在fab的情况下,fab除ch1域和cl域以外还包括fv区。如本文所使用的“fv”或“fv片段”或“fv区”意指包括abd的vl和vh域的多肽。fv区可被设计成fab和scfv形式(如上文所论述,通常是还包含如上文所概述的恒定区的两个不同的多肽),其中vl域和vh域组合(通常使用如本文所论述的连接子)以形成scfv。本文中“单链fv”或“scfv”意指重链可变域共价连接到轻链可变域,通常使用如本文所论述的scfv连接子,以形成scfv或scfv结构域。scfv结构域由n端至c端可呈任一定向(vh-连接子-vl或vl-连接子-vh)。在序列表和图中所描绘的序列中,vh和vl域的顺序在名称中指示,例如,h.x_l.y意指n-至c端是vh-连接子-vl,l.y_h.x是vl-连接子-vh。如本文所使用的“igg子类修饰”或“同型修饰”意指将igg同型的一个氨基酸转化成不同比对的igg同型中的对应的氨基酸的氨基酸修饰。举例来说,因为在eu位置296处igg1包括酪胺酸且igg2包括苯丙氨酸,在igg2中的f296y取代被视为igg子类修饰。如本文所使用的“非天然存在的修饰”意指不为同型的氨基酸修饰。举例来说,因为人类igg的无一者在位置434处包括丝氨酸,在igg1、igg2、igg3或igg4(或其杂合体)中的取代434s被视为非天然存在的修饰。如本文所使用的“氨基酸”和“氨基酸身份标识”意指经编码用于dna和rna的20种天然存在的氨基酸中的一种。如本文所使用的“效应功能”意指由抗体fc区与fc受体或配体的相互作用引起的生化事件。效应功能包含但不限于adcc、adcp以及cdc。如本文所使用的“iggfc配体”意思指结合于igg抗体的fc区以形成fc/fc配体复合物的来自任何生物体的分子,优选多肽。fc配体包含但不限于fcγri、fcγrii、fcγriii、fcrn、clq、c3、甘露聚糖结合凝集素、甘露糖受体、葡萄球菌(staphylococcal)蛋白a、链球菌(streptococcal)蛋白g以及病毒fcγr。fc配体还包含fc受体同源物(fcrh),其是与fcγr同源的fc受体家族(davis等人,2002,《免疫评论(immunologicalreviews)》190:123-136,其以引用的方式整体并入本文中)。fc配体可能包含未发现的结合fc的分子。特定iggfc配体是fcrn和fcγ受体。如本文所使用的“fc配体”意思指结合于抗体的fc区以形成fc/fc配体复合物的来自任何生物体的分子,优选多肽。如本文所使用的“fcγ受体”、“fcγr”或“fcγr(fcgammar)”意指结合igg抗体fc区并且由fcγr基因编码的蛋白质家族的任何成员。在人类中,这个家族包含但不限于fcγri(cd64),包含同工型fcγria、fcγrib以及fcγric;fcγrii(cd32),包含同工型fcγriia(包含同种异型h131和r131)、fcγriib(包含fcγriib-1和fcγriib-2)以及fcγriic;以及fcγriii(cd16),包含同工型fcγriiia(包含同种异型v158和f158)和fcγriiib(包含同种异型fcγriib-na1和fcγriib-na2)(jefferis等人,2002,《免疫学快报(immunollett)》82:57-65,其以引用的方式整体并入本文中),以及任何未发现的人类fcγr或fcγr同工型或同种异型。fcγr可以来自任何生物体,包含但不限于人类、小鼠、大鼠、兔以及猴。小鼠fcγr包含但不限于fcγri(cd64)、fcγrii(cd32)、fcγriii-1(cd16)和fcγriii-2(cd16-2)以及任何未发现的小鼠fcγr或fcγr同工型或同种异型。如本文所使用的“fcrn”或“新生fc受体”意指结合igg抗体fc区且至少部分地由fcrn基因编码的蛋白质。fcrn可以来自任何生物体,包含但不限于人类、小鼠、大鼠、兔以及猴。如所属领域中已知,功能性fcrn蛋白质包括两个多肽,通常称为重链和轻链。轻链是β-2微球蛋白且重链由fcrn基因编码。除非另外说明,否则本文中fcrn或fcrn蛋白质是指fcrn重链与β-2微球蛋白的复合物。多种fcrn变体用于增强与fcrn受体的结合,且在一些情况下,增加血清半衰期。“fcrn变体”增强与fcrn受体的结合,且适合的fcrn变体在下文中展示。如本文所使用的“亲本多肽”意指随后经过修饰来产生变体的起始多肽。亲本多肽可为天然存在的多肽或天然存在的多肽的变体或工程改造版本。因此,如本文所使用的“亲本免疫球蛋白”意指未修饰的免疫球蛋白多,其经过修饰来产生变体,并且如本文所使用的“亲本抗体”意指未修饰的抗体,其经过修饰来产生变体抗体。应注意“亲本抗体”包含如下文概述的已知的市售、以重组方式产生的抗体。在此上下文中,“亲本fc结构域”将相对于所列举的变体而言;因此,“变体人类igg1fc结构域”是相比于人类igg1的亲本fc结构域,“变体人类igg4fc结构域”是相比于人类igg4亲本fc结构域,诸如此类。如本文所使用的“fc”或“fc区”或“fc结构域”意指包括igg分子的ch2-ch3域,且在一些情况下包含铰链的多肽。在eu编号中,对于人类igg1而言,ch2-ch3域包括氨基酸231至447,且铰链是216至230。因此“fc结构域”的定义包含氨基酸231-447(ch2-ch3)或216-447(铰链-ch2-ch3)或其片段。在此上下文中“fc片段”可能含有较少来自n端和c端中任一个或两个的氨基酸但仍保留与另一fc结构域或fc片段形成二聚体的能力,如可使用通常基于尺寸的标准方法(例如,非变性色谱、尺寸排阻色谱等)检测。在本发明中尤其使用人类iggfc结构域,且可为来自人类igg1、igg2或igg4的fc结构域。相比于亲本fc结构域,“变体fc结构域”含有氨基酸修饰。因此,相比于人类igg1fc结构域,“变体人类igg1fc结构域”含有氨基酸修饰(通常氨基酸取代,尽管在消融变体的情况下包含氨基酸缺失)。一般而言,变体fc结构域具有与对应的亲本人类iggfc结构域的至少约80、85、90、95、97、98或99%一致性(使用下文论述的一致性算法,其中一个实施例利用如所属领域中已知的blast算法使用默认参数)。或者,相比于亲本fc结构域,变体fc结构域可具有1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、11、12、13、14、15、16、17、18、19或20个氨基酸修饰。或者,相比于亲本fc结构域,变体fc结构域可具有至多1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、11、12、13、14、15、16、17、18、19或20个氨基酸修饰。此外,如本文所论述,本文中的变体fc结构域仍保留与另一fc结构域形成二聚体的能力,如使用如本文所描述的已知技术(如非变性凝胶电泳)所测量。本文中“重链恒定区”意指抗体(或其片段)的ch1-铰链-ch2-ch3部分,不包含重链可变域;在人类igg1的eu编号中,此为氨基酸118-447。本文中“重链恒定区片段”意指含有较少来自n端和c端中任一个或两个的氨基酸但仍保留与另一重链恒定区形成二聚体的能力的重链恒定区。如本文中所用,“位置”意指蛋白质序列中的位置。位置可以依序编号,或根据已确立的形式(例如用于抗体编号的eu索引)编号。如本文所使用的“目标抗原”意指通过包括给定抗体的可变区的抗原结合域而特异性结合的分子。如下文所论述,在本发明的情况下目标抗原是检查点抑制剂蛋白质。在本发明的异源二聚抗体中的单体的情况下,“链型”在本文中意指类似于dna中的“匹配”的两股,将异源二聚变体并入各单体中以便保持“匹配”以形成异源二聚体的能力。举例来说,如果将一些pi变体工程引入单体a(例如使得pi更高)中,那么同样可以使用的作为“电荷对”的空间变体不会干扰pi变体,例如将使得pi更高的电荷变体放在相同“链”或“单体”上以保留两种功能。类似地,对于如下文更全面概述的组中成对出现的“歪斜”变体来说,所属领域的技术人员在决定所述一组对将进入哪个链或单体时考虑pi,以使得使用歪斜变体的pi也使pi分离最大化。如本文中所用,“目标细胞”意指表达目标抗原的细胞。在产生根据本发明的双特异性抗体的情况下,本文中的“宿主细胞”意指含有编码双特异性抗体的组分的外源核酸且能够在合适的条件下表达双特异性抗体的细胞。适合的宿主细胞在下文中论述。如本文所使用的“可变区”或“可变域”意指免疫球蛋白的区,其包括基本上由分别构成κ、λ以及重链免疫球蛋白基因基因座的vκ、vλ和/或vh基因中的任一种编码的一个或多个ig域,且含有赋予抗原特异性的cdr。因此,“重链可变域”与“轻链可变域”配对以形成抗原结合域(“abd”)。此外,各可变域包括三个高变区(“互补决定区”,“cdr”)(对于重链可变域而言是vhcdr1、vhcdr2以及vhcdr3,且对于轻链可变域而言是vlcdr1、vlcdr2以及vlcdr3)和四个框架(fr)区,其按照以下顺序由氨基端至羧基端排列:fr1-cdr1-fr2-cdr2-fr3-cdr3-fr4。“野生型或wt”在本文中意指在自然界中发现的氨基酸序列或核苷酸序列,包括等位基因变异。wt蛋白质具有尚未被刻意修饰的氨基酸序列或核苷酸序列。本发明提供多种具有与人类抗体域的序列一致性的抗体域。两个类似序列(例如抗体可变域)之间的序列一致性可通过如以下文献中的算法测量:smith,t.f.和waterman,m.s.(1981)“生物序列对比(comparisonofbiosequences)”,《应用数学进展(adv.appl.math.)》2:482[局部同源算法];needleman,s.b.和wunsch,cd.(1970)“适用于在两个蛋白质的氨基酸序列中搜索类似性的通用方法(ageneralmethodapplicabletothesearchforsimilaritiesintheaminoacidsequenceoftwoproteins)”,《分子生物学杂志(j.mol.biol.)》48:443[同源比对算法];pearson,w.r.和lipman,d.j.(1988)“生物序列比较的改良工具(improvedtoolsforbiologicalsequencecomparison)”《美国国家科学院院刊(proc.natl.acad.sci.(u.s.a.))》85:2444[类似性搜索方法];或altschul,s.f.等人,(1990)“基础局部比对搜索工具(basiclocalalignmentsearchtool)”,《分子生物学杂志》215:403-10,即“blast”算法,参见https://blast.ncbi.nlm.nih.gov/blast.cgi。当使用前述算法中的任一种时,使用默认参数(用于窗口长度、空位罚分等)。在一个实施例中,使用blast算法使用默认参数测定序列一致性。本发明的抗体通常是分离的或重组的。“分离”当用于描述本文中公开的各种多肽时,意指一种多肽,其已经被鉴别且从表达其的细胞或细胞培养物中分离和/或回收。通常,通过至少一个纯化步骤制备经分离的多肽。“经分离的抗体”是指基本上与具有不同抗原特异性的其它抗体脱离的抗体。“重组”意指在外源宿主细胞中使用重组核酸技术产生抗体,且所述抗体也可被分离。“特异性结合”或“特异性结合于”特定抗原或表位,或“对”特定抗原或表位“有特异性”意指结合可测量地不同于非特异性相互作用。特异性结合可以如下测量:例如相较于对照分子的结合来测定一种分子的结合,所述对照分子通常是不具有结合活性的类似结构分子。举例来说,特异性结合可以通过与类似于目标的对照分子竞争来测定。特定抗原或表位的特异性结合可以通过例如抗体来展现,所述抗体针对抗原或表位的kd为至少约10-4m、至少约10-5m、至少约10-6m、至少约10-7m、至少约10-8m、至少约10-9m、或者至少约10-10m、至少约10-11m、至少约10-12m或更大,其中kd是指特定抗体-抗原相互作用的解离速率。典型地,特异性结合抗原的抗体的kd是对照分子相对于抗原或表位的kd的20倍、50倍、100倍、500倍、1000倍、5,000倍、10,000倍或更多倍。另外,针对特定抗原或表位的特异性结合可以通过例如抗体展现,所述抗体对抗原或表位的ka或ka是对照物针对所述表位的ka或ka的至少20倍、50倍、100倍、500倍、1000倍、5,000倍、10,000倍或更多倍,其中ka或ka是指特定抗体-抗原相互作用的结合速率。结合亲和力通常使用biacore、spr或bli分析测量。e.抗体一方面,本文提供结合于sstr2的组合物(例如抗sstr2抗体)。在某些实施例中,抗体结合于人类sstr2(图11)。主题抗sstr2抗体包含单特异性sstr2抗体以及多特异性(例如双特异性)抗sstr2抗体。在某些实施例中,抗sstr2抗体具有如图1中所描绘的任一抗体形式的形式。在一些实施例中,主题组合物包含sstr2结合域。在一些实施例中,组合物包含具有sstr2结合域的抗体。本文所提供的抗体包含一个、两个、三个四个以及五个或更多个sstr2结合域。在某些实施例中,sstr2结合域包含选自由图11中所描绘的sstr2结合域组成的群组的sstr2结合域的vhcdr1、vhcdr2、vhcdr3、vlcdr1、vlcdr2以及vlcdr3序列。在一些实施例中,sstr2结合域包含选自图11中所描绘的sstr2结合域的sstr2结合域的带下划线的vhcdr1、vhcdr2、vhcdr3、vlcdr1、vlcdr2以及vlcdr3序列。在一些实施例中,sstr2结合域包含选自图11中所描绘的sstr2结合域的sstr2结合域的重链可变域和轻链可变域。图11中所描绘的sstr2结合域包含抗sstr2h1.143_l1.30;抗sstr2h1_l1.1;抗sstr2h1.107_l1.30;抗sstr2h1.107_l1.67;抗sstr2h1.107_l1.108;抗sstr2h1.107_l1.111;抗sstr2h1.107_l1.114;抗sstr2h1.107_l1.102;抗sstr2h1.107_l1.110;抗sstr2h1.125_l1.30;抗sstr2h1.125_l1.67;抗sstr2h1.125_l1.108;抗sstr2h1.125_l1.111;抗sstr2h1.125_l1.114;抗sstr2h1.125l1.102;以及抗sstr2h1.125_l1.10。在一个示范性实施例中,抗体包含抗sstr2h1.143_l1.30结合域。在一些实施例中,抗体是结合sstr2和cd3的双特异性抗体。这种抗体包含cd3结合域和至少一个sstr2结合域。任何适合的sstr2结合域均可包含在抗sstr2×抗cd3双特异性抗体中。在一些实施例中,抗sstr2×抗cd3双特异性抗体包含一个、两个、三个四个或更多个sstr2结合域,包含但不限于图11中所描绘的那些。在某些实施例中,抗sstr2×抗cd3抗体包含sstr2结合域,所述sstr2结合域包含选自由图11中描绘的sstr2结合域组成的群组的sstr2结合域的vhcdr1、vhcdr2、vhcdr3、vlcdr1、vlcdr2以及vlcdr3序列。在一些实施例中,抗sstr2×抗cd3抗体包含sstr2结合域,所述sstr2结合域包含选自由图11中所描绘的sstr2结合域组成的群组的sstr2结合域的带下划线的vhcdr1、vhcdr2、vhcdr3、vlcdr1、vlcdr2以及vlcdr3序列。在一些实施例中,抗sstr2×抗cd3抗体包含sstr2结合域,所述sstr2结合域包含选自由图11中所描绘的sstr2结合域组成的群组的sstr2结合域的重链可变域和轻链可变域。在一个示范性实施例中,抗sstr2×抗cd3抗体包含抗sstr2h1.143_l1.30结合域。本文所提供的抗sstr2×抗cd3抗体可包含任何适合的cd3结合域。在某些实施例中,抗sstr2×抗cd3抗体包含cd3结合域,所述cd3结合域包含选自由图12和13中所描绘的cd3结合域组成的群组的cd3结合域的vhcdr1、vhcdr2、vhcdr3、vlcdr1、vlcdr2以及vlcdr3序列。在一些实施例中,抗sstr2×抗cd3抗体包含cd3结合域,所述cd3结合域包含选自由图12或13中所描绘的cd3结合域组成的群组的cd3结合域的vhcdr1、vhcdr2、vhcdr3、vlcdr1、vlcdr2以及vlcdr3序列。在一些实施例中,抗sstr2×抗cd3抗体包含cd3结合域,所述cd3结合域包含选自由图12或13中所描绘的cd3结合域组成的群组的cd3结合域的重链可变域和轻链可变域。在一些实施例中,cd3结合域选自抗cd3h1.30_l1.47;抗cd3h1.32_l1.47;抗cd3h1.89_l1.48;抗cd3h1.90_l1.47;抗cd3h1.33_l1.47;以及抗cd3h1.31_l1.47。如本文中所使用,术语“抗体”普遍使用。用于本发明中的抗体可以采用如本文所述的多种形式,包括本文所述的传统抗体以及抗体衍生物、片段和模拟物。传统抗体结构单元典型地包括四聚体。各四聚体典型地由相同的两对多肽链构成,每对具有一个“轻链”(典型地分子量为约25kda)和一个“重链”(典型地分子量为约50-70kda)。人类轻链分类为κ和λ轻链。本发明针对igg类别,其具有若干子类,包含但不限于igg1、igg2、igg3以及igg4。应注意igg1具有在356(d或e)和358(l或m)处具有多型性的不同同种异型。本文中所描绘的序列使用356d/358m同种异型,然而在本文中包含另一同种异型。即,本文中所包含的包含igg1fc结构域的任何序列可具有356e/358l,替换356d/358m同种异型。此外,本文中的多种抗体具有至少一个位置220处的半胱氨酸替换为丝氨酸;通常对本文中所描绘的大多数序列而言,这是在“scfv单体”侧,尽管其也可以在“fab单体”侧,或两者,以减少二硫化物形成。特别包含在本文中的序列内的是半胱氨酸替换的(c220s)序列中的一个或两个。因此,如本文中所用,“同型”意指根据其恒定区的化学和抗原特征界定的免疫球蛋白的任一子类。应了解治疗抗体还可以包含同型和/或子类的杂合体。举例来说,如us公开案2009/0163699中所示,其以引用的方式并入,本发明包含使用人类igg1/g2杂合体。高变区通常包涵以下氨基酸残基:轻链可变区中的约氨基酸残基24-34(lcdr1;“l”表示轻链)、50-56(lcdr2)以及89-97(lcdr3)以及重链可变区中的约31-35b(hcdr1;“h”表示重链)、50-65(hcdr2)以及95-102(hcdr3);kabat等人,《免疫学所关注的蛋白质序列(sequencesofproteinsofimmunologicalinterest)》第5版,公共卫生署(publichealthservice),美国国家卫生研究院(nationalinstitutesofhealth),马里兰州贝塞斯达(bethesda,md.)(1991);和/或形成高变环的那些残基(例如轻链可变区中的残基26-32(lcdr1)、50-52(lcdr2)以及91-96(lcdr3)以及重链可变区中的26-32(hcdr1)、53-55(hcdr2)以及96-101(hcdr3);chothia和lesk(1987)《分子生物学杂志》196:901-917。本发明的特定cdr描述于下文。如所属领域的技术人员将了解,cdr的精确编号和布置在不同系统中可以不同。然而,应理解,可变重链和/或可变轻链序列的公开包含相关(固有)cdr的公开。因此,各重链可变区的公开是vhcdr(例如vhcdr1、vhcdr2以及vhcdr3)的公开并且各轻链可变区的公开是vlcdr(例如vlcdr1、vlcdr2以及vlcdr3)的公开。cdr编号的可用的比较如下,参见lafranc等人,《发育与比较免疫学(dev.comp.immunol.)》27(l):55-77(2003):表1kabat+chothiaimgtrabatabmchothiacontactxencorvhcdr126-3527-3831-3526-3526-3230-3527-35vhcdr250-6556-6550-6550-5852-5647-5854-61vhcdr395-102105-11795-10295-10295-10293-101103-116vlcdr124-3427-3824-3424-3424-3430-3627-38vlcdr250-5656-6550-5650-5650-5646-5556-62vlcdr389-97105-11789-9789-9789-9789-9697-105在通篇本说明书中,当提及可变域中的残基(大致是轻链可变区中的残基1-107和重链可变区中的残基1-113)时通常使用kabat编号系统且fc区使用eu编号系统(例如kabat等人,前述(1991))。重链中另一类型的ig域是铰链区。本文中“铰链”或“铰链区”或“抗体铰链区”或“铰链域”意指包括抗体的第一恒定域与第二恒定域之间的氨基酸的柔性多肽。结构上,iggch1域结束于eu位置215处,且iggch2域开始于残基eu位置231处。因此本文中对于igg而言定义抗体铰链包含位置216(在igg1中为e216)至230(在igg1中为p230),其中编号是根据如kabat中的eu索引。在一些情况下,使用“铰链片段”,其在铰链域的n端和c端中任一个或两个处含有较少氨基酸。如在本文中所提及,pi变体还可在铰链区中制成。轻链通常包含两个域:轻链可变域(含有轻链cdr且与重链可变域一起形成fv区),和轻链恒定区(通常称为cl或cκ)。所关注的用于另外取代的另一区域是fc区,如下文所概述。本发明提供大量的不同cdr组。在这种情况下,“完整cdr组”包含三个可变轻链cdr和三个可变重链cdr,例如vlcdr1、vlcdr2、vlcdr3、vhcdr1、vhcdr2以及vhcdr3。这些可以相应地是较大轻链可变域或重链可变域的一部分。另外,如本文中更全面地概述,当使用重链和轻链时(例如当使用fab时),重链可变域和轻链可变域可以位于单独的多肽链上,或在scfv序列的情况下位于单个多肽链上。cdr促进了抗体的抗原结合位点,或更具体地说,表位结合位点的形成。“表位”是指与抗体分子可变区中的特定抗原结合位点(称为互补位)相互作用的决定子。表位是如氨基酸或糖侧链的分子的聚组,且通常具有特定的结构特征以及特定的电荷特征。单个抗原可以具有超过一个表位。表位可以包括直接参与结合的氨基酸残基(也称为表位的免疫显性组分)和不直接参与结合的其它氨基酸残基,如被特异性抗原结合肽有效阻断的氨基酸残基;换句话说,氨基酸残基位于特异性抗原结合肽的覆盖区内。表位可以是构象表位或线性表位。构象表位由来自线性多肽链的不同区段的氨基酸在空间上并列而产生。线性表位是由多肽链中的相邻氨基酸残基产生的表位。构象表位和非构象表位的区别可在于,在变性溶剂存在下,与构象表位的结合消失,但与非构象表位的结合不消失。表位典型地包括呈独特空间构象的至少3个且更通常至少5个或8-10个氨基酸。可以在展示一种抗体阻断另一种抗体与目标抗原结合的能力的简单免疫分析(例如“分库(binning)”)中检验识别相同表位的抗体。如下文概述,本发明不仅包含本文中所列举的抗原结合域和抗体,还包含与已结合所列举的抗原结合域的表位竞争结合的那些。因此,本发明提供不同的抗体域。如本文所述以及所属领域中已知,本发明的异源二聚抗体包括重链和轻链内的不同域,其也可以是重叠的。这些域包含但不限于fc结构域、ch1域、ch2域、ch3域、铰链域、重链恒定域(ch1-铰链-fc结构域或ch1-铰链-ch2-ch3)、重链可变域、轻链可变域、轻链恒定域、fab域和scfv结构域。因此,“fc结构域”包含-ch2-ch3域,和任选地铰链域(-h-ch2-ch3)。在本文中的实施例中,当scfv连接于fc结构域时,scfv构筑体的c端连接于fc结构域的全部或部分铰链;例如其通常连接于作为铰链起点的序列epks。重链包括重链可变域和恒定域,其包含ch1-任选的铰链-包括ch2-ch3的fc结构域。轻链包括轻链可变域和轻链恒定域。scfv包括可变重链、scfv连接子以及轻链可变域。在本文概述的大多数构筑体和序列中,可变重链的c端连接于scfv连接子的n端,scfv连接子的c端连接于可变轻链的n端(n-vh-连接子-vl-c),尽管此顺序可以调换(n-vl-连接子-vh-c)。本发明的一些实施例包括至少一个scfv结构域,其虽然不是天然存在的,但是通常包含由scfv连接子连接在一起的重链可变域和轻链可变域。如本文所概述,虽然scfv结构域通常由n端至c端定向为vh-scfv连接子-vl,但是针对任一个scfv结构域(或使用fab的vh和vl序列构筑的那些)这种定向可以逆向,成为vl-scfv连接子-vh,其中取决于形式(大体参见图1)任选的连接子位于一个末端或两个末端。如本文中所示,存在多种可用于共价连接所列举的域的适合的连接子(用作域连接子或scfv连接子),包含传统肽键,其通过重组技术来生成。在一些实施例中,连接肽可能主要包含以下氨基酸残基:gly、ser、ala或thr。连接肽应该具有足以连接两个分子的长度,以便其相对于彼此呈现正确的构象,从而使其保留所期望的活性。在一个实施例中,连接子具有约1至50个氨基酸的长度,优选约1至30个氨基酸的长度。在一个实施例中,可以使用1至20个氨基酸长度的连接子,在一些实施例中发现使用约5至约10个氨基酸。适用的连接子包含甘氨酸-丝氨酸聚合物,包含例如(gs)n、(gsggs)n、(ggggs)n以及(gggs)n(其中n是至少为一(且通常是3至4)的整数)、甘氨酸-丙氨酸聚合物、丙氨酸-丝氨酸聚合物,和其它柔性连接子。或者,可使用多种非蛋白质聚合物,包含但不限于聚乙二醇(peg)、聚丙二醇、聚环氧烷或聚乙二醇和聚丙二醇的共聚物作为连接子。其它连接子序列可以包含cl/ch1域的任何长度的任何序列,但不包含cl/ch1域的所有残基;例如cl/ch1域的前5-12个氨基酸残基。连接子可以衍生自免疫球蛋白轻链,例如cκ或cλ。连接子可以衍生自任何同型的免疫球蛋白重链,包含例如cγ1、cγ2、cγ3、cγ4、cα1、cα2、cδ、cε以及cμ。连接子序列还可以衍生自其它蛋白质,如ig样蛋白质(例如tcr、fcr、kir);为铰链区衍生序列;以及来自其它蛋白质的其它天然序列。在一些实施例中,连接子是“域连接子”,其用于将如本文中概述的任何两个域连接在一起。举例来说,在图1f中,可存在将fab的ch1域的c端连接于scfv的n端的域连接子,且另一任选的域连接子将scfv的c端连接于ch2域(尽管在许多实施例中,使用铰链作为此域连接子)。虽然可使用任何适合的连接子,许多实施例利用甘氨酸-丝氨酸聚合物作为域连接子,包含例如(gs)n、(gsggs)n、(ggggs)n以及(gggs)n,其中n是至少为一(且通常3至4至5)的整数,以及允许两个域的重组连接且具有足够长度和柔性以使得各域保留其生物功能的任何肽序列。在一些情况下,且在关注如下文概述的“链型”的情况下,可以使用如在scfv连接子的一些实施例中所使用的带电域连接子。在一些实施例中,连接子是scfv连接子,用于共价连接如本文所论述的vh域和vl域。在许多情况下,scfv连接子是带电scfv连接子,多种所述带电scfv连接子展示于图7中。相应地,本发明另外提供带电scfv连接子,以辅助第一单体与第二单体之间的pi分离。即,通过并入带正或负(或两者,在使用不同单体上的scfv的支架的情况下)电的scfv连接子,由此允许包括带电连接子的单体改变pi而不会在fc结构域中产生进一步的变化。这些带电连接子可以被取代到含有标准连接子的任何scfv中。此外,如所属领域的技术人员将了解,根据所期望的pi变化,在适当的“链”或单体上使用带电scfv连接子。举例来说,如本文所论述,为了产生三重f形式的异源二聚抗体,计算每个所期望抗原结合域的fv区的初始pi,且选择其一来产生scfv,且依据pi选择正或负电连接子。带电域连接子还可以用于同样增强本发明单体的pi分离,且因此图7中所包含的连接子可以用于本文中利用连接子的任何实施例中。特定来说,图1中所描绘的形式是抗体,通常称为“异源二聚抗体”,意指蛋白质具有至少两种自组装到异源二聚fc结构域中的相关fc序列和至少两个fv区,无论作为fab或作为scfv。f.嵌合和人源化抗体在某些实施例中,本发明的抗体包括来自特定生殖系重链免疫球蛋白基因的重链可变区和/或来自特定生殖系轻链免疫球蛋白基因的轻链可变区。举例来说,这种抗体可以包括如下人类抗体或由其组成,所述人类抗体包括作为特定生殖系序列“的产物”或“衍生自”其的重链或轻链可变区。作为人类生殖系免疫球蛋白序列“的产物”或“衍生自”其的人类抗体可以通过将人类抗体的氨基酸序列与人类生殖系免疫球蛋白的胺基酸序列比较且选择与人类抗体的序列在序列方面最接近(即,最大一致性%)的人类生殖系免疫球蛋白序列(使用本文概述的方法)来鉴别出来。相比于生殖系序列,作为特定人类生殖系免疫球蛋白序列“的产物”或“衍生自”其的人类抗体可含有氨基酸差异,这是由,例如,天然存在的体细胞突变或刻意引入的定点突变造成。然而,人源化抗体典型地在氨基酸序列方面与由人类生殖系免疫球蛋白基因编码的氨基酸序列至少90%一致,且当相比于其它物种(例如鼠类生殖系序列)的生殖系免疫球蛋白胺基酸序列时,含有将所述抗体标识为衍生自人类序列的氨基酸残基。在某些情况下,人源化抗体在氨基酸序列方面可与由生殖系免疫球蛋白基因编码的氨基酸序列至少95、96、97、98或99%或甚至至少96%、97%、98%或99%一致。典型地,衍生自特定人类生殖系序列的人源化抗体相比于由人类生殖系免疫球蛋白基因编码的氨基酸序列将显示不超过10-20个氨基酸差异(在引入本文中的任何歪斜、pi以及消融变体之前;即,在引入本发明的变体之前,变体数目通常较低)。在某些情况下,相比于由生殖系免疫球蛋白基因编码的氨基酸序列,人源化抗体可显示不超过5或甚至不超过4、3、2或1个氨基酸差异(同样,在引入本文中的任何歪斜、pi以及消融变体之前;即,在引入本发明的变体之前,变体数目通常较低)。在一个实施例中,亲本抗体已经发生亲和力成熟,如所属领域中已知。可以利用基于结构的方法进行人源化和亲和力成熟,例如,如ussn11/004,590中所述。可以使用基于选择的方法使抗体可变区发生人源化和/或亲和力成熟,包含但不限于以下文献中所描述的方法:wu等人,1999,《分子生物学杂志》294:151-162;baca等人,1997,《生物化学杂志》272(16):10678-10684;rosok等人,1996,《生物化学杂志》271(37):22611-22618;rader等人,1998,《美国国家科学院院刊》95:8910-8915;krauss等人,2003,《蛋白质工程》16(10):753-759,均以引用的方式整体并入本文中。其它人源化方法可能涉及仅移植cdr的一部分,包含但不限于以下文献中所描述的方法:ussn09/810,510;tan等人,2002,《免疫学杂志》169:1119-1125;depascalis等人,2002,《免疫学杂志》169:3076-3084,均以引用的方式整体并入本文中。g.异源二聚抗体因此,在一些实施例中,主题抗体是依赖于使用两个不同的重链变体fc序列的异源二聚抗体。这种抗体将自组装以形成异源二聚fc结构域和异源二聚抗体。本发明涉及新颖构筑体,其提供允许与多于一种抗原或配体结合,例如允许双特异性结合(例如抗sstr2且抗cd3结合)的异源二聚抗体。异源二聚抗体构筑体是基于抗体的重链中的两个fc结构域的自组装本质,例如两个“单体”组装成“二聚体”。异源二聚抗体是通过改变各单体的氨基酸序列来产生,如下文更充分地论述。因此,本发明大体上涉及建立异源二聚抗体,所述异源二聚抗体可以通过若干方式与抗原(例如sstr2和cd3)共接合,这依赖于各链上不同的恒定区中的氨基酸变体促进异源二聚形成和/或允许异源二聚体的纯化比同型二聚体容易。因此,本发明提供双特异性抗体。在一些实施例中,本发明提供包含sstr2结合域的双特异性抗体。在一些实施例中,双特异性抗体是抗sstr2×抗cd3双特异性抗体。抗体技术中正存在的一个问题是期望“双特异性”抗体同时结合到两种不同抗原,通常由此允许不同抗原接近且从而产生新功能和新疗法。一般来说,这些抗体是通过将每条重链和轻链的基因纳入宿主细胞中来产生。这通常引起所期望的异源二聚体(a-b)以及两种同型二聚体(a-a和b-b(不包含轻链异源二聚问题))的形成。然而,形成双特异性抗体的主要障碍是难以从同型二聚抗体中纯化异源二聚抗体和/或难以偏向于形成异源二聚体(相对于形成同型二聚体)。存在多种机制可以用于产生本发明的异源二聚体。此外,如所属领域的技术人员将了解,这些机制可以组合以确保高度异源二聚。因此,引起异源二聚体产生的氨基酸变体称为“异源二聚变体”。如下文所论述,异源二聚变体可包含空间变体(例如下文所述的“杵和臼”或“歪斜”变体以及下文所述的“电荷对”变体)以及“pi变体”,其允许从异源二聚体中纯化同型二聚体。正如wo2014/145806(以全文引用的方式并入本文中)中大体所述,且如下文关于“异源二聚变体”的论述具体所述,适用的异源二聚机制包含“杵和臼”(“kih”;本文中有时称为“歪斜”变体)(参见wo2014/145806中的论述)、如wo2014/145806中所述的“静电转向”或“电荷对”、如wo2014/145806中所述的pi变体,以及如wo2014/145806和下文中所概述的一般其它fc变体。在本发明中,存在可以容易纯化异源二聚抗体的若干机制;一种依赖于pi变体的使用,以便每种单体具有不同pi,从而允许对a-a、a-b和b-b二聚蛋白质进行等电纯化。或者,一些支架形式(如“三重f”形式)也允许根据尺寸实现分离。如下文进一步概述,还可以“歪斜”形成异源二聚体而非同型二聚体。因此,空间异源二聚变体与pi或电荷对变体的组合特别适用于本发明中。一般而言,在本发明中特定用途的实施例依赖于如下的变体的组:其包含有助于形成异源二聚而形成非同型二聚的歪斜变体,其与pi变体偶合,其增加两个单体之间的pi差异以辅助自同型二聚体中纯化异源二聚体。另外,如下文更全面地概述,取决于异源二聚体抗体的形式,pi变体可以含在单体的恒定域和/或fc结构域内,或可以使用带电连接子(域连接子或scfv连接子)。即,如三重f或“开瓶器”形式的使用scfv的支架可包含带电scfv连接子(正性或负性),其为纯化目的提供进一步pi加强。如所属领域的技术人员将了解,一些三重f形式仅与带电scfv连接子一起使用且不进行另外的pi调节,但是本发明还提供位于一个或两个单体和/或带电域连接子上的pi变体。此外,用于替代功能的另外的氨基酸工程改造也可以赋予pi变化,如fc、fcrn以及ko变体。在使用pi作为分离机制以允许对异源二聚蛋白质纯化的本发明中,可以将氨基酸变体引入一个或两个单体多肽中;即,一个单体(为了简单起见,本文中称为“单体a”)的pi可以加以工程改造以与单体b分离,或单体a和b变化均可以改变,其中单体a的pi增加,而单体b的pi降低。如下文更充分地描述,任一个或两个单体的pi变化可以如下进行:除去或添加带电荷的残基(例如用带正或负电荷的氨基酸残基替换中性氨基酸,例如甘氨酸替换成谷氨酸)、将带电残基从正或负电荷变为相反电荷(天冬氨酸变为赖氨酸)或将带电残基变为中性残基(例如电荷的失去;赖氨酸变为丝氨酸)。多种这些变体显示在图中。相应地,本发明的这个实施例使得至少一个单体的pi产生了足够的变化,从而可以将异源二聚体与同型二聚体分离。如所属领域的技术人员将了解,且如下文进一步所论述,这可通过使用“野生型”重链恒定区和已经工程改造成以增加或降低其pi的变体区进行(wta-+b或wta--b),或通过增加一个区并减少另一区进行(a+-b-或a-b+)。因此,一般来说,本发明的一些实施例的组分是抗体恒定区中的氨基酸变体,其涉及形成“pi抗体”的二聚体蛋白质中至少一个(若非两个)单体的等电点(pi)的改变,这是通过将氨基酸取代(“pi变体”或“pi取代”)并入一个或两个单体中来实现。如本文中所示,如果两个单体的pi相差小到0.1个ph单位(0.2、0.3、0.4和0.5或更大ph单位都能适用于本发明中),那么可以实现异源二聚体与两种同型二聚体的分离。如所属领域的技术人员将了解,为了得到良好分离,在每个或两个单体上包含的pi变体的数目将部分地取决于组分的初始pi,例如在三重f形式中,取决于所关注的scfv和fab的初始pi。即,为了确定对哪种单体进行工程改造或以哪个“方向”(例如更正或更负),计算两种目标抗原的fv序列且据此作出决定。如所属领域中已知,不同的fv具有用于本发明中的不同初始pi。一般来说,如本文中所概述,pi经工程改造而使得每种单体的总pi差异达到至少约0.1log,如本文中所概述优选0.2到0.5。另外,如所属领域的技术人员将了解且如本文中所概述,在一些实施例中,可以根据尺寸将异源二聚体与同型二聚体分离。举例来说,如图1所示,若干种形式允许根据尺寸来分离异源二聚体和同型二聚体。在其中通过使用重链恒定区、使用pi变体实现异源二聚的情况下,提供一种设计和纯化双特异性蛋白质(包含抗体)的更加模块化方法。因此,在一些实施例中,可变区中不包含异源二聚变体(包含歪斜和纯化异源二聚变体),因此必须对每种个别抗体进行工程改造。另外,在一些实施例中,通过引进来自不同igg同型的pi变体以便在不引入显著免疫原性的情况下改变pi来显著降低pi变体导致免疫原性的可能性。因此,待解决的一个另外的问题是低pi恒定域和高人类序列含量的阐明,例如最小化或避免任何特定位置的非人类残基。这种pi工程改造可以得到的一种辅助好处也是血清半衰期延长和fcrn结合增强。即,如ussn13/194,904(以全文引用的方式并入本文中)中所述,降低抗体恒定域(包含抗体和fc融合物中所发现的恒定域)的pi可以延长活体内血清滞留时间。用于延长血清半衰期的这些pi变体还促进了pi变化以便纯化。另外,应注意异源二聚变体的pi变体具有用于双特异性抗体的分析和品质控制方法中的额外好处,因为当存在同型二聚体时,消除、最小化和区分的能力是显著的。类似地,可靠地测试异源二聚抗体产生再现性的能力是重要的。异源二聚变体本发明提供异源二聚蛋白质,包含多种形式的异源二聚抗体,其使用异源二聚变体来实现异源二聚形成和/或从同型二聚体中纯化。异源二聚歪斜变体中存在多个适合对的组。这些变体以成“对”“组”方式出现。即,将所述对的一组并入第一单体中且所述对的另一组并入第二单体中。应注意,这些组不一定表现为“臼包杵”型变体,其中一个单体上的残基与另一个单体上的残基之间存在一对一对应;即,组中的这些对形成了两个单体之间的界面,促进了异源二聚体形成且阻碍了同型二聚体形成,从而使在生物学条件下自发形成异源二聚体的百分比逾90%,而非预期的50%(25%同型二聚体a/a:50%异源二聚体a/b:25%同型二聚体b/b)。空间变体在一些实施例中,通过添加空间变体可以促进异源二聚体的形成。即,通过改变每个重链中的氨基酸,不同重链缔结形成异源二聚体结构的可能性大于与相同fc氨基酸序列形成同型二聚体的可能性。适合的空间变体包含在图12中。一种机制在所属领域中通常称为“杵和臼”,还可以任选地使用所谓的氨基酸工程来产生空间影响以有利于形成异源二聚体而不利于形成同型二聚体;这有时称为“杵和臼”,如以下文献中所述:ussn61/596,846;ridgway等人,《蛋白质工程》9(7):617(1996);atwell等人,《分子生物学杂志》1997270:26;美国专利第8,216,805号,其均特此以全文引用的方式并入本文中。图式标出了依赖于“杵和臼”的“单体a-单体b”对的数目。另外,如merchant等人,《自然·生物技术》16:677(1998)中所述,这些“杵和臼”突变可以与二硫键组合以歪斜形成异源二聚体。一种用于产生异源二聚体的另一种机制有时称为“静电转向”,如gunasekaran等人,《生物化学杂志》285(25):19637(2010)中所述,其特此以全文引用的方式并入本文中。这在本文中有时被称作“电荷对”。在这个实施例中,静电用于歪斜形成异源二聚。如所属领域的技术人员将了解,这些还可以影响pi且从而影响纯化,且因此在一些情况下还可以被视为pi变体。然而,由于这些是为了强制发生异源二聚而产生且不是作为纯化工具使用,因此其被分类为“空间变体”。这些包含但不限于,与d221r/p228r/k409r配对的d221e/p228e/l368e(例如这些是“单体对应组”)和与c220r/e224r/p228r/k409r配对的c220e/p228e/368e。其它单体a和单体b变体可以任选地且独立地以任何量与其它变体(如本文中概述的pi变体或us2012/0149876的图37中所示的其它空间变体,其中的图式和图例和seqidno明确地以引用的方式并入本文中)组合。在一些实施例中,本文中概述的空间变体可以任选地且独立地与任何pi变体(或其它变体,如fc变体、fcrn变体等)一起并入一个或两个单体中,且可以独立地且任选地包含于本发明的蛋白质中或从本发明的蛋白质中排除。一系列适合的歪斜变体见于图3中,其中图8显示了多个实施例中的一些对特定效用。在多个实施例中,特别使用包含但不限于以下的几对组:s364k/e357q:l368d/k370s;l368d/k370s:s364k;l368e/k370s:s364k;t411t/e360e/q362e:d401k;l368d/k370s:s364k/e357l以及k370s:s364k/e357q。就命名法而言,对“s364k/e357q:l368d/k370s”意味着一个单体具有双变体组s364k/e357q且另一个单体具有双变体组l368d/k370s。用于异源二聚体的pi(等电点)变体一般来说,如所属领域的技术人员将了解,pi变体存在两种普通类别:提高蛋白质pi(碱性变化)的变体和降低蛋白质pi(酸性变化)的变体。如本文所述,可以实现这些变体的所有组合:一个单体可以是野生型,或pi与野生型无明显差异的变体;且另一个单体可以更具碱性或更具酸性。或者,改变每个单体,一个更具碱性且一个更具酸性。pi变体的优选组合显示于图4中。如本文中所概述且如图式中所示,这些变化是相对于igg1显示,而且所有同型以及同型杂合体均可以按此方式改变。在重链恒定域是来自igg2-4的情况下,还可以使用r133e和r133q。在一个实施例中,例如在图1a、1e、1f、1g、1h以及1i的形式中,pi变体的优选组合具有一个包括208d/295e/384d/418e/421d变体(当相对于人类igg1时,n208d/q295e/n384d/q418e/n421d)的单体(负性fab侧)和包括带正电的scfv连接子(包含(gkpgs)4(seqidno:818))的第二单体(正性scfv侧)。然而,如所属领域的技术人员将了解,第一单体包含ch1域,其包含位置208。因此,在不包含ch1域的构筑体中(例如,对于不使用例如双scfv形式或“单臂”形式,如图1b、1c或1d中所描绘的形式,中的其中一个域上的ch1域的抗体),优选负性pi变体fc组包含295e/384d/418e/421d变体(当相对于人类igg1时,q295e/n384d/q418e/n421d)。因此,在一些实施例中,一个单体具有来自图4的一组取代且另一单体具有带电连接子(呈带电scfv连接子形式,因为所述单体包括scfv,或呈带电域连接子形式,如所述形式所表明,其可选自图7中所描绘的那些)。同型变体此外,本发明的许多实施例依赖于将来自一种igg同型的特定位置的pi氨基酸“引进”另一种igg同型中,从而减少或排除变体中引入不合需要的免疫原性的可能性。多种这些变体显示于美国公开2014/0370013的图21中,特此以引用的方式并入本文中。即,igg1因各种原因(包括高效应功能)是治疗抗体的常见同型。然而,igg1的重链恒定区具有比igg2的重链恒定区更高的pi(8.10相对于7.31)。通过将特定位置的igg2残基引入igg1骨架中,使得所得单体的pi降低(或提高)且另外展现较长的血清半衰期。举例来说,igg1具有位于位置137处的甘氨酸(pi5.97),且igg2具有谷氨酸(pi3.22);引进谷氨酸将影响所得蛋白质的pi。如下文所述,为了显著影响变异型抗体的pi,通常需要多个氨基酸取代。然而,应注意,如下文所论述,igg2分子中的均一变化使得血清半衰期延长。在其它实施例中,产生非同型氨基酸变化,以降低所得蛋白质的总体电荷状态(例如通过将较高pi氨基酸变为较低pi氨基酸),或允许结构适应稳定性等,如下文更进一步描述。另外,通过对重链和轻链恒定域进行pi工程改造,可以发现异源二聚体中的各个单体发生显著变化。如本文所论述,使两个单体的pi相差至少0.5可以允许通过离子交换色谱或等电点聚焦或对等电点敏感的其它方法来实现分离。计算pi每个单体的pi可以依赖于变体重链恒定域的pi和总单体的pi,包括变体重链恒定域和融合搭配物。因此,在一些实施例中,使用美国公开2014/0370013中的图19的图表,基于变体重链恒定域计算pi变化。如本文所论述,对哪种单体进行工程改造通常根据fv和支架区域的固有pi来决定。或者,可以对每个单体的pi进行比较。也赋予更好的fcrn活体内结合的pi变体在pi变体降低单体pi的情况下,其可以具有改善活体内血清滞留时间的额外好处。虽然仍有待检验,但认为fc区使活体内半衰期延长,原因是在内体中在ph6下结合于fcrn隔绝了fc(ghetie和ward,1997《今日免疫学(immunoltoday)》18(12):592-598,其以引用的方式整体并入本文中)。随后内体隔室使fc再循环到细胞表面。一旦隔室向细胞外空间敞开,则较高ph(约7.4)诱导fc释放回到血液中。dall'acqua等人表明,在小鼠中,在ph6和ph7.4的fcrn结合增强的fc突变体实际上具有减小的血清浓度以及与野生型fc相同的半衰期(dall'acqua等人,2002,《免疫学杂志》169:5171-5180,其以引用的方式整体并入本文中)。fc对fcrn的亲和力在ph7.4下增强被认为是禁止了fc释放回到血液中。因此,延长fc活体内半衰期的fc突变理想地增强了较低ph下的fcrn结合,同时仍允许fc在较高ph下释放。在6.0到7.4的ph范围内,氨基酸组胺酸的电荷状态发生变化。因此,在fc/fcrn复合物中的重要位置找到his残基并不出乎意料。最近,已经提出具有等电点较低的可变区的抗体也可以具有较长血清半衰期(igawa等人,2010peds.23(5):385-392,其以引用的方式整体并入本文中)。然而,对这种抗体的机制的了解尚浅。此外,不同抗体,可变区情况不同。pi降低且半衰期延长的恒定区变体为改进抗体的药物动力学特性提供了更加模块化的方法,如本文所述。用于另外的功能的其它fc变体除pi氨基酸变体之外,可以出于多种原因(包含但不限于,改变与一种或多种fcγr受体的结合、改变与fcrn受体的结合等)而产生多个适用的fc氨基酸修饰。相应地,本发明的蛋白质可以包括氨基酸修饰,包括本文中概述的异源二聚变体,包括pi变体和空间变体。每一组变体可以独立地且任选地包含于任何特定异源二聚蛋白质中或从任何特定异源二聚蛋白质中排除。fcγr变体因此,存在许多适用fc取代可用来改变与fcγr受体中的一个或多个的结合。引起结合增强以及结合减弱的取代都可以是适用的。举例来说,已知与fcγriiia的结合增强通常引起增强的adcc(抗体依赖性细胞介导的细胞毒性;细胞介导的反应,其中表达fcγr的非特异性细胞毒性细胞识别目标细胞上的所结合抗体且随后促使目标细胞溶解)。类似地,在一些情况下,与fcγriib的结合减弱(抑制性受体)也可能是有利的。用于本发明中的氨基酸取代包括ussn11/124,620(尤其是图41)、11/174,287、11/396,495、11/538,406中列出的氨基酸取代,所有这些文献全文且尤其其中所公开的变体明确地以全文引用的方式并入本文中。可以使用的特定变体包含但不限于236a、239d、239e、332e、332d、239d/332e、267d、267e、328f、267e/328f、236a/332e、239d/332e/330y、239d、332e/330l、243a、243l、264a、264v以及299t。此外,存在可以用于增强与fcrn受体的结合以及延长血清半衰期的其它fc取代,如ussn12/341,769中所具体公开(以全文引用的方式并入本文中),包含但不限于434s、434a、428l、308f、259i、428l/434s、259i/308f、436i/428l、436i或v/434s、436v/428l以及259i/308f/428l。消融变体类似地,另一类功能变体是“fcγr消融变体”或“fc基因敲除(fcko或ko)”变体。在这些实施例中,在一些治疗应用中,期望减少或除去fc结构域与一种或多种或全部fcγ受体(例如fcγri、fcγriia、fcγriib、fcγriiia等)的正常结合以避免其它作用机制。即,例如,在许多实施例中,尤其在使用单价结合cd3的双特异性抗体时,通常期望消融fcγriiia结合以消除或显著降低adcc活性,其中fc结构域中的一个包括一种或多种fcγ受体消融变体。这些消融变体描绘于图14中,且各自可以独立地且任选地被包含在内或排除,利用消融变体的优选方面选自由以下组成的群组:g236r/l328r、e233p/l234v/l235a/g236del/s239k、e233p/l234v/l235a/g236del/s267k、e233p/l234v/l235a/g236del/s239k/a327g、e233p/l234v/l235a/g236del/s267k/a327g以及e233p/l234v/l235a/g236del。应注意本文中提及的消融变体消融fcγr结合,但是通常不消融fcrn结合。如所属领域中已知,人类igg1的fc结构域与fcγ受体的结合最强,且因此当异源二聚抗体的主链中的恒定域(或fc结构域)是igg1时可使用消融变体。替代地,或在igg1背景下除消融变体以外,例如,糖基化位置297处的突变(通常成a或s)也能够显著消融与fcγriiia的结合。人类igg2和igg4与fcγ受体的结合天然地降低,且因此可以在具有或不具有消融变体的情况下使用那些骨架。异源二聚变体与fc变体的组合如所属领域的技术人员将了解,所述异源二聚变体(包括歪斜和/或pi变体)全部可以任选地且独立地按任何方式组合,只要其保持其“链型”或“单体分区”。另外,所有这些变体可以按任一种异源二聚形式组合。在pi变体的情况下,虽然图中显示了可以特别使用的实施例,但是可以根据改变两个单体之间的pi差异以辅助纯化的基本规则来产生其它组合。另外,异源二聚变体中的任一种(歪斜变体和pi变体)也独立地且任选地与fc消融变体、fc变体、fcrn变体组合,如本文中所大体概述。h.本发明的适用形式如所属领域的技术人员将了解且下文更充分地论述,本发明的异源二聚融合蛋白可以采用多种多样的构形,如图1中所大体描绘。一些图描绘了“单末端”构形,其中分子的一个“臂”上存在一种类型的特异性且另一个“臂”存在不同特异性。其它图描绘了“双末端”构形,其中分子的“顶部”存在至少一种类型的特异性且分子的“底部”存在一或多种不同特异性。因此,本发明涉及与不同的第一抗原和第二抗原共接合的新颖免疫球蛋白组合物。如所属领域的技术人员将了解,本发明的异源二聚形式可以具有不同的价数且具有双特异性。即,本发明的异源二聚抗体可以为二价且双特异性,其中一种目标肿瘤抗原(例如cd3)通过一种结合域结合且另一目标肿瘤抗原(例如sstr2)通过第二结合域结合。异源二聚抗体也可以为三价且双特异性,其中第一抗原通过两种结合域结合且第二抗原通过第二结合域结合。如本文中所概述,当cd3是其中一个目标抗原时,优选地cd3仅单价结合,以减小潜在副作用。本发明利用抗cd3抗原结合域与抗sstr2结合域的组合。如所属领域的技术人员将了解,可以使用如任一图(具体参见图2到图7以及图18)中所描绘的抗cd3cdr、抗cd3轻链可变域和重链可变域、fab以及scfv的任何集合。类似地,可以使用任一种抗sstr2抗原结合域,可以使用如任一图(例如图8和10)中所描绘的cdr、轻链可变域和重链可变域、fab以及scfv,其任选地且独立地按任何组合形式组合。开瓶器本发明中特别适用的一种异源二聚支架是如图1a中所示的“三重f”或“开瓶器”支架形式。在这个实施例中,抗体的一个重链含有单链fv(“scfv”,如下文所定义)且另一重链是包括可变重链和轻链的“规则”fab形式。在本文中这种结构有时称作“三重f”形式(scfv-fab-fc)或“开瓶器”形式,因为其在视觉上与开瓶器粗略相似。所述两个链通过使用恒定区(例如fc结构域、ch1域和/或铰链区)中的氨基酸变体(其促进异源二聚抗体形成)一起引入,如下文更充分地描述。本发明“三重f”形式存在几个显著优点。如所属领域中已知,依赖于两种scfv构筑体的抗体类似物通常具有稳定性和聚集问题,这些问题在本发明中可以通过添加“规则”的重链与轻链对来缓解。另外,与依赖两个重链和两个轻链的形式相反,不存在重链和轻链配对不当(例如重链1与轻链2配对等)的问题。本文中概述的许多实施例通常依赖于开瓶器形式,其包括含有scfv的第一单体,所述scfv包括使用scfv连接子(在许多情况下带电)共价连接的重链可变域和轻链可变域,其中scfv通常经由域连接子(如本文中所概述,其可以是不带电或带电的)共价连接到第一fc结构域的n端。开瓶器形式中的第二单体是重链,且组合物另外包括轻链。一般来说,在许多优选实施例中,scfv是结合cd3的域,且fab形成sstr2结合域。此外,本发明的fc结构域通常包括歪斜变体(例如,如图3和8中所示的一组氨基酸取代,且特别适用的歪斜变体选自由以下组成的群组:s364k/e357q:l368d/k370s;l368d/k370s:s364k;l368e/k370s:s364k;t411t/e360e/q362e:d401k;l368d/k370s:s364k/e357l以及k370s:s364k/e357q);任选地消融变体(包含图5中所示的那些);任选地带电scfv连接子(包含图7中所示的那些);以及包括pi变体的重链(包含图4中所示的那些)。在一些实施例中,开瓶器形式包含歪斜变体、pi变体以及消融变体。因此,一些实施例包含开瓶器形式,其包括:a)第一单体(“scfv单体”),其包括带电scfv连接子(在一些实施例中,优选图7的+h序列)、歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k以及如本文所概述的结合cd3的fv;b)第二单体(“fab单体”,其包括歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k以及重链可变域,其与轻链可变域一起构成如本文所概述的结合sstr2的fv;以及c)轻链。结合cd3的scfv的示范性重链可变域和轻链可变域包含在图12和13中。结合sstr2的fv的示范性重链可变域和轻链可变域包含在图11中。在一个示范性实施例中,sstr2结合域是h1.143_l1.30sstr2结合域,且结合cd3的scfv包含h1.30_l1.47cd3结合域的重链可变域和轻链可变域。其它特别适用的sstr2和cd3序列组合公开于图16中。在一些实施例中,开瓶器形式包含歪斜变体、pi变体、消融变体以及fcrn变体。因此,一些实施例包含开瓶器形式,其包括:a)第一单体(“scfv单体”),其包括带电scfv连接子(在一些实施例中,优选图7的+h序列)、歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k、fcrn变体m428l/n434s以及如本文所概述的结合cd3的fv;b)第二单体(“fab单体”,其包括歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k、fcrn变体m428l/n434s以及重链可变域,其与轻链可变域一起构成如本文所概述的结合sstr2的fv;以及c)轻链。结合cd3的scfv的示范性重链可变域和轻链可变域包含在图12和13中。结合sstr2的fv的示范性重链可变域和轻链可变域包含在图11中。在一个示范性实施例中,sstr2结合域包含h1.143_l1.30sstr2结合域的重链可变域和轻链可变域,且结合cd3的scfv包含h1.30_l1.47cd3结合域的重链可变域和轻链可变域。其它特别适用的sstr2和cd3序列组合公开于图16中。图9展示一些示范性开瓶器“骨架”序列,其缺失可用于本发明中的fv序列。在一些实施例中,本文中所描绘的任一种vh序列和vl序列(包含图和序列表中所描绘的所有vh序列和vl序列)均可以使用图和序列表中所示的任一种抗cd3scfv序列添加到图9的开瓶器骨架形式中作为“fab侧”。对于图9中的开瓶器骨架1(任选地包含428l/434s变体),在这些实施例中特别适用的cd结合域序列包含但不限于:cd3结合域抗cd3h1.30_l1.47、抗cd3h1.32_l1.47、抗cd3h1.89_l1.47、抗cd3h1.90_l1.47、抗cd3h1.33_l1.47以及抗cd3h1.31_l1.47,以及图12和13中所描绘的那些,其作为图9中所示的骨架的scfv侧连接。对于图9中的开瓶器骨架1(任选地包含428l/434s变体),在这些实施例中特别适用的sstr2结合域序列包含但不限于:抗sstr2h1.143_l1.30;抗sstr2h1_l1.1;抗sstr2h1.107_l1.30;抗sstr2h1.107_l1.67;抗sstr2h1.107_l1.108;抗sstr2h1.107_l1.111;抗sstr2h1.107_l1.114;抗sstr2h1.107_l1.102;抗sstr2h1.107_l1.110;抗sstr2h1.125_l1.30;抗sstr2h1.125_l1.67;抗sstr2h1.125_l1.108;抗sstr2h1.125_l1.111;抗sstr2h1.125_l1.114;抗sstr2h1.125_l1.102;以及抗sstr2h1.125_l1.10。尤其适用于与图9中的开瓶器骨架1(任选地包含428l/434s变体)一起使用的sstr2和cd3序列组合公开于图16中。在一个示范性实施例中,开瓶器抗体包含图9中的开瓶器“骨架”1,sstr2结合域包含h1.143_l1.30sstr2结合域的重链可变域和轻链可变域,且结合cd3的scfv包含h1.30_l1.47cd3结合域的重链可变域和轻链可变域。mab-fv本发明中特别适用的一种异源二聚支架是图1h中所示的mab-fv形式。在这个实施例中,所述形式依赖于使用“额外”重链可变域的c端连接到一个单体且“额外”轻链可变域的c端连接到另一个单体,从而形成第三抗原结合域,其中两个单体的fab部分结合sstr2且“额外”scfv结构域结合cd3。在这个实施例中,第一单体包括第一重链,其包括第一重链可变域和包括第一fc结构域的第一重链恒定域,其中第一轻链可变域使用域连接子共价连接到第一fc结构域的c端(vh1-ch1-铰链-ch2-ch3-[任选的连接子]-vl2)。第二单体包括第二重链可变域,其属于包括第二fc结构域的第二重链恒定域,和第三重链可变域,其使用域连接子共价连接到第二fc结构域的c端(vj1-ch1-铰链-ch2-ch3-[任选的连接子]-vh2)。两个c端连接可变域构成结合cd3的fv(由于其不太优选具有二价cd3结合)。这个实施例进一步使用包括轻链可变域和轻链恒定域的共用轻链,其与重链缔结而形成两个结合sstr2的相同fab。就本文中的许多实施例来说,这些构筑体包含所期望且如本文中所述的歪斜变体、pi变体、消融变体、另外的fc变体等。本发明提供其中cd结合域序列如图12和13以及序列表中所示的mab-fv形式。本发明提供其中sstr2结合域序列如图11和序列表中所示的mab-fv形式。特别适用于与mab-fv形式一起使用的sstr2和cd3序列组合公开于图16中。此外,mab-fv形式的fc结构域包括歪斜变体(例如,如图3和8中所示的一组氨基酸取代,且特别适用的歪斜变体选自由以下组成的群组:s364k/e357q:l368d/k370s;l368d/k370s:s364k;l368e/k370s:s364k;t411t/e360e/q362e:d401k;l368d/k370s:s364k/e357l;k370s:s364k/e357q;t366s/l368a/y407v:t366w;以及t366s/l368a/y407v/y349c:t366w/s354c);任选地消融变体(包含图5中所示的那些);任选地带电scfv连接子(包含图7中所示的那些);以及包括pi变体的重链(包含图4中所示的那些)。在一些实施例中,mab-fv形式包含歪斜变体、pi变体以及消融变体。因此,一些实施例包含mab-fv形式,其包括:a)第一单体,其包括歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k以及第一重链可变域(其与轻链的第一轻链可变域一起构成结合sstr2的fv)和第二重链可变域;b)第二单体,其包括歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k以及第一重链可变域(其与第一轻链可变域一起构成如本文所概述的结合sstr2的fv)和第二可变轻链(其与第二重链可变域一起形成结合cd3的fv(abd);以及c)轻链,其包括第一轻链可变域和轻链恒定域。在一些实施例中,mab-fv形式包含歪斜变体、pi变体、消融变体以及fcrn变体。因此,一些实施例包含mab-fv形式,其包括:a)第一单体,其包括歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k、fcrn变体m428l/n434s以及第一重链可变域(其与轻链的第一轻链可变域一起构成结合sstr2的fv)和第二重链可变域;b)第二单体,其包括歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k、fcrn变体m428l/n434s以及第一重链可变域(其与第一轻链可变域一起构成如本文所概述的结合sstr2的fv),以及第二可变轻链,其与第一单体的第二重链可变域一起形成结合cd3的fv(abd);以及c)轻链,其包括第一轻链可变域和轻链恒定域。对于与图10中的mab-fv骨架1(任选地包含m428l/434s变体)相似的mab-fv序列,在这些实施例中特别适用的cd3结合域序列包含但不限于:抗cd3h1.30_l1.47、抗cd3h1.32_l1.47、抗cd3h1.89_l1.47、抗cd3h1.90_l1.47、抗cd3h1.33_l1.47以及抗cd3h1.31_l1.47,以及图12和13中所描绘的那些。对于与图10中的mab-fv骨架1(任选地包含m428l/434s变体)相似的mab-fv序列,在这些实施例中特别适用的sstr2结合域序列包含但不限于:抗sstr2h1.143_l1.30;抗sstr2h1_l1.1;抗sstr2h1.107_l1.30;抗sstr2h1.107_l1.67;抗sstr2h1.107_l1.108;抗sstr2h1.107_l1.111;抗sstr2h1.107_l1.114;抗sstr2h1.107_l1.102;抗sstr2h1.107_l1.110;抗sstr2h1.125_l1.30;抗sstr2h1.125_l1.67;抗sstr2h1.125_l1.108;抗sstr2h1.125_l1.111;抗sstr2h1.125_l1.114;抗sstr2h1.125_l1.102;以及抗sstr2h1.125_l1.10,以及图11和15和seqidno:68至659中所列出的那些。特别适用于和与图10中的mab-fv骨架1(任选地包含428l/434s变体)相似的mab-fv序列一起使用的sstr2和cd3序列组合公开于图16中。mab-scfv本发明中特别适用的一种异源二聚支架是图1中所示的mab-fv形式。在这个实施例中,所述形式依赖于使用scfv的c端连接到其中一个单体,从而形成第三抗原结合域,其中两个单体的fab部分结合sstr2且“额外”scfv结构域结合cd3。因此,第一单体包括第一重链(包括重链可变域和恒定域),且c端共价连接scfv,其包括scfv轻链可变域、scfv连接子以及scfv重链可变域,呈定向(vh1-ch1-铰链-ch2-ch3-[任选的连接子]-vh2-scfv连接子-vl2或vh1-ch1-铰链-ch2-ch3-[任选的连接子]-vl2-scfv连接子-vh2)。这个实施例进一步使用包括轻链可变域和轻链恒定域的共用轻链,其与重链缔结而形成两个结合sstr2的相同fab。就本文中的许多实施例来说,这些构筑体包含所期望且如本文中所述的歪斜变体、pi变体、消融变体、另外的fc变体等。本发明提供其中cd结合域序列如图12和13以及序列表中所示的mab-scfv形式。本发明提供其中sstr2结合域序列如图11和序列表中所示的mab-scfv形式。特别适用于与mab-scfv形式一起使用的sstr2和cd3序列组合公开于图16中。此外,mab-scfv形式的fc结构域包括歪斜变体(例如,如图3和8中所示的一组氨基酸取代,且特别适用的歪斜变体选自由以下组成的群组:s364k/e357q:l368d/k370s;l368d/k370s:s364k;l368e/k370s:s364k;t411t/e360e/q362e:d401k;l368d/k370s:s364k/e357l;k370s:s364k/e357q;t366s/l368a/y407v:t366w;以及t366s/l368a/y407v/y349c:t366w/s354c);任选地消融变体(包含图5中所示的那些);任选地带电scfv连接子(包含图7中所示的那些);以及包括pi变体的重链(包含图4中所示的那些)。在一些实施例中,mab-scfv形式包含歪斜变体、pi变体以及消融变体。因此,一些实施例包含mab-scfv形式,其包括:a)第一单体,其包括歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k以及重链可变域(其与共用轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv)以及结合cd3的scfv结构域;b)第二单体,其包括歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k以及重链可变域(其与共用轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv);以及c)共用轻链,包括轻链可变域和轻链恒定域。在一些实施例中,mab-scfv形式包含歪斜变体、pi变体、消融变体以及fcrn变体。因此,一些实施例包含mab-scfv形式,其包括:a)第一单体,其包括歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k以及重链可变域(其与共用轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv)以及结合cd3的scfv结构域;b)第二单体,其包括歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k、fcrn变体m428l/n434s以及重链可变域(其与共用轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv);以及c)共用轻链,其包括轻链可变域和轻链恒定域。在图10中的mab-scfv骨架1(任选地包含m428l/n434s)中(任选地包含428l/434s变体cd3),在这些实施例中特别适用的cd3结合域序列包含但不限于:抗cd3h1.30_l1.47;抗cd3h1.32_l1.47、抗cd3h1.89_l1.47、抗cd3h1.90_l1.47、抗cd3h1.33_l1.47以及抗cd3h1.31_l1.47,以及图12和13中所描绘的那些。在图10中的mab-scfv骨架1(任选地包含m428l/n434s)中(任选地包含428l/434s变体),在这些实施例中特别适用的sstr2结合域序列包含但不限于:抗sstr2h1.143_l1.30;抗sstr2h1_l1.1;抗sstr2h1.107_l1.30;抗sstr2h1.107_l1.67;抗sstr2h1.107_l1.108;抗sstr2h1.107_l1.111;抗sstr2h1.107_l1.114;抗sstr2h1.107_l1.102;抗sstr2h1.107_l1.110;抗sstr2h1.125_l1.30;抗sstr2h1.125_l1.67;抗sstr2h1.125_l1.108;抗sstr2h1.125_l1.111;抗sstr2h1.125_l1.114;抗sstr2h1.125_l1.102;以及抗sstr2h1.125_l1.10,以及图11和15和seqidno:68至659中所列出的那些中心scfv本发明中特别适用的一种异源二聚支架是图1中所示的中心scfv形式。在这个实施例中,所述形式依赖于使用所插入的scfv结构域,从而形成第三抗原结合域,其中两个单体的fab部分结合sstr2且“额外”scfv结构域结合cd3。scfv结构域插入到其中一个单体的fc结构域与ch1-fv区之间,从而提供第三抗原结合域。在这个实施例中,一个单体包括第一重链,其包括第一重链可变域、ch1域(和任选的铰链)以及fc结构域的,且scfv包括scfv轻链可变域、scfv连接子以及scfv重链可变域。scfv使用任选的域连接子共价连接在重链恒定域ch1域的c端与第一fc结构域的n端之间(vh1-ch1-[任选的连接子]-vh2-scfv连接子-vl2-[包含铰链的任选的连接子]-ch2-ch3,或scfv相反定向vh1-ch1-[任选的连接子]-vl2-scfv连接子-vh2-[包含铰链的任选的连接子]-ch2-ch3)。另一单体是标准fab侧。这个实施例进一步使用包括轻链可变域和轻链恒定域的共用轻链,其与重链缔结而形成两个结合sstr2的相同fab。就本文中的许多实施例来说,这些构筑体包含所期望且如本文中所述的歪斜变体、pi变体、消融变体、另外的fc变体等。本发明提供其中cd3结合域序列如图12和13以及序列表中所示的中心scfv形式。本发明提供其中抗sstr2序列如图11和序列表中所示的中心scfv形式。特别适用于与中心scfv形式一起使用的sstr2和cd3序列组合公开于图16中。此外,中心scfv形式的fc结构域包括歪斜变体(例如,如图3和8中所示的一组氨基酸取代,且特别适用的歪斜变体选自由以下组成的群组:s364k/e357q:l368d/k370s;l368d/k370s:s364k;l368e/k370s:s364k;t411t/e360e/q362e:d401k;l368d/k370s:s364k/e357l;k370s:s364k/e357q;t366s/l368a/y407v:t366w;以及t366s/l368a/y407v/y349c:t366w/s354c);任选地消融变体(包含图5中所示的那些);任选地带电scfv连接子(包含图7中所示的那些);以及包括pi变体的重链(包含图4中所示的那些)。在一些实施例中,中心scfv形式包含歪斜变体、pi变体以及消融变体。因此,一些实施例包含中心scfv形式,其包括:a)第一单体,其包括歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k以及重链可变域(其与轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv)以及结合cd3的scfv结构域;b)第二单体,其包括歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k以及重链可变域(其与轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv);以及c)轻链,包括轻链可变域和轻链恒定域。在一些实施例中,中心scfv形式包含歪斜变体、pi变体、消融变体以及fcrn变体。因此,一些实施例包含中心scfv形式,其包括:a)第一单体,其包括歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k以及重链可变域(其与轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv)以及结合cd3的scfv结构域;b)第二单体,其包括歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k、fcrn变体m428l/n434s以及重链可变域(其与轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv);以及c)轻链,其包括轻链可变域和轻链恒定域。对于与图9中的开瓶器骨架1相似/使用图9中的开瓶器骨架1的中心scfv序列(任选地包含m428l/434s),在这些实施例中特别适用的cd3结合域序列包含但不限于:抗cd3h1.30_l1.47;抗cd3h1.32_l1.47、抗cd3h1.89_l1.47、抗cd3h1.90_l1.47、抗cd3h1.33_l1.47以及抗cd3h1.31_l1.47,以及图12和13中所描绘的那些。对于与图9中的开瓶器骨架1相似/使用图9中的开瓶器骨架1的中心scfv序列(任选地包含m428l/434s变体),在这些实施例中特别适用的sstr2结合域序列包含但不限于:抗sstr2h1.143_l1.30;抗sstr2h1_l1.1;抗sstr2h1.107_l1.30;抗sstr2h1.107_l1.67;抗sstr2h1.107_l1.108;抗sstr2h1.107_l1.111;抗sstr2h1.107_l1.114;抗sstr2h1.107_l1.102;抗sstr2h1.107_l1.110;抗sstr2h1.125_l1.30;抗sstr2h1.125_l1.67;抗sstr2h1.125_l1.108;抗sstr2h1.125_l1.111;抗sstr2h1.125_l1.114;抗sstr2h1.125_l1.102;以及抗sstr2h1.125_l1.10,以及图11和15和seqidno:68至659中所列出的那些。中心fv本发明中特别适用的一种异源二聚支架是图1g中所示的中心fv形式。在这个实施例中,所述形式依赖于使用插入的fv域(即,中心fv域),从而形成第三抗原结合域,其中两个单体的fab部分结合sstr2且“中心fv”域结合cd3。scfv结构域插入在单体的fc结构域与ch1-fv区域之间,从而提供第三抗原结合域,其中每个单体含有scfv组分(例如一个单体包含重链可变域且另一个单体包含轻链可变域)。在这个实施例中,一个单体包含第一重链,其包含第一重链可变域、ch1域和fc结构域以及另外的轻链可变域。所述轻链域使用域连接子共价连接在重链恒定域的ch1域的c端与第一fc结构域的n端之间(vh1-ch1-[任选的连接子]-vl2-铰链-ch2-ch3)。另一个单体包括第一重链,其包括第一重链可变域、ch1域和fc结构域以及另外的重链可变域(vh1-ch1-[任选的连接子]-vh2-铰链-ch2-ch3)。所述轻链域使用域连接子共价连接在重链恒定域的ch1域的c端与第一fc结构域的n端之间。这个实施例进一步使用包括轻链可变域和轻链恒定域的共用轻链,其与重链缔结而形成两个结合sstr2的相同fab。就本文中的许多实施例来说,这些构筑体包含所期望且如本文中所述的歪斜变体、pi变体、消融变体、另外的fc变体等。本发明提供其中cd3结合域序列如图12和13以及序列表中所示的中心fv形式。本发明提供其中sstr2结合域序列如图11和序列表中所示的中心fv形式。特别适用于与中心fv形式一起使用的sstr2和cd3序列组合公开于图16中。对于中心fv形式,在这些实施例中,特别适用的cd3结合域序列包含但不限于:抗cd3h1.30_l1.47、抗cd3h1.32_l1.47、抗cd3h1.89_l1.47、抗cd3h1.90_l1.47、抗cd3h1.33_l1.47以及抗cd3h1.31_l1.47,以及图12和13中所描绘的那些。对于中心fv形式,在这些实施例中特别适用的sstr2结合域序列包含但不限于:抗sstr2h1.143_l1.30;抗sstr2h1_l1.1;抗sstr2h1.107_l1.30;抗sstr2h1.107_l1.67;抗sstr2h1.107_l1.108;抗sstr2h1.107_l1.111;抗sstr2h1.107_l1.114;抗sstr2h1.107_l1.102;抗sstr2h1.107_l1.110;抗sstr2h1.125_l1.30;抗sstr2h1.125_l1.67;抗sstr2h1.125_l1.108;抗sstr2h1.125_l1.111;抗sstr2h1.125_l1.114;抗sstr2h1.125_l1.102;以及抗sstr2h1.125_l1.10,以及图11和15以及seqidno:68至659中所列出的那些。单臂中心scfv本发明中特别适用的一种异源二聚支架是图1中所示的单臂中心scfv形式。在这个实施例中,一个单体仅包含fc结构域,而另一个单体使用所插入的scfv结构域,从而形成第二抗原结合域。在这种形式中,fab部分结合sstr2且scfv结合cd3或反之亦然。scfv结构域插入在其中一个单体的fc结构域与ch1-fv区之间。在这个实施例中,一个单体包含第一重链,其包括第一重链可变域、ch1域以及fc结构域,且scfv包括scfv轻链可变域、scfv连接子以及scfv重链可变域。scfv使用域连接子共价连接在重链恒定域的ch1域的c端与第一fc结构域的n端之间。第二单体包含fc结构域。这个实施例进一步使用包括轻链可变域和轻链恒定域的轻链,所述轻链与重链缔结而形成fab。就本文中的许多实施例来说,这些构筑体包含所期望且如本文中所述的歪斜变体、pi变体、消融变体、另外的fc变体等。本发明提供其中cd3结合域序列如图12和13以及序列表中所示的中心fv形式。本发明提供其中sstr2结合域序列如图11和序列表中所示的中心fv形式。特别适用于与中心fv形式一起使用的sstr2和cd3序列组合公开于图16中。此外,单臂中心scfv形式的fc结构域大体上包括歪斜变体(例如,如图3和8中所示的一组氨基酸取代,且特别适用的歪斜变体选自由以下组成的群组:s364k/e357q:l368d/k370s;l368d/k370s:s364k;l368e/k370s:s364k;t411t/e360e/q362e:d401k;l368d/k370s:s364k/e357l;k370s:s364k/e357q;t366s/l368a/y407v:t366w以及t366s/l368a/y407v/y349c:t366w/s354c);任选地消融变体(包含图5中所示的那些);任选地带电scfv连接子(包含图7中所示的那些);以及包括pi变体的重链(包含图4中所示的那些)。在一些实施例中,单臂中心scfv形式包含歪斜变体、pi变体以及消融变体。因此,单臂中心scfv形式的一些实施例包括:a)第一单体,其包括:歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k以及重链可变域(其与轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv)以及结合cd3的scfv结构域;b)第二单体,其包含fc结构域,所述fc结构域具有歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k;以及c)轻链,其包括轻链可变域和轻链恒定域。在一些实施例中,单臂中心scfv形式包含歪斜变体、pi变体、消融变体以及fcrn变体。因此,单臂中心scfv形式的一些实施例包括:a)第一单体,其包括歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k、fcrn变体m428l/n434s以及重链可变域(其与轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv)以及结合cd3的scfv结构域;b)第二单体,其包含fc结构域,所述fc结构域具有歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k以及fcrn变体m428l/n434s;以及c)轻链,其包括轻链可变域和轻链恒定域。对于单臂中心scfv形式,特别适用的cd3结合域序列包含但不限于:抗cd3h1.30_l1.47、抗cd3h1.32_l1.47、抗cd3h1.89_l1.47、抗cd3h1.90_l1.47、抗cd3h1.33_l1.47以及抗cd3h1.31_l1.47,以及图12和13中所描绘的那些。对于单臂中心scfv形式,特别适用的sstr2结合域序列包含但不限于:抗sstr2h1.143_l1.30;抗sstr2h1_l1.1;抗sstr2h1.107_l1.30;抗sstr2h1.107_l1.67;抗sstr2h1.107_l1.108;抗sstr2h1.107_l1.111;抗sstr2h1.107_l1.114;抗sstr2h1.107_l1.102;抗sstr2h1.107_l1.110;抗sstr2h1.125_l1.30;抗sstr2h1.125_l1.67;抗sstr2h1.125_l1.108;抗sstr2h1.125_l1.111;抗sstr2h1.125_l1.114;抗sstr2h1.125_l1.102;抗sstr2h1.125_l1.10,以及图11和15以及seqidno:68至659中所列出的那些。单臂scfv-mab本发明中特别适用的一种异源二聚支架是图1d中所示的单臂scfv-mab形式。。在这个实施例中,一个单体仅包括fc结构域,而另一个单体使用连接于重链的n端的scfv结构域,通常通过使用连接子:vh-scfv连接子-vl-[任选的域连接子]-ch1-铰链-ch2-ch3或(呈相反定向)vl-scfv连接子-vh-[任选的域连接子]-ch1-铰链-ch2-ch3。在这一形式中,fab部分各自结合sstr2且scfv结合cd3。这个实施例进一步使用包括轻链可变域和轻链恒定域的轻链,所述轻链与重链缔结而形成fab。就本文中的许多实施例来说,这些构筑体包含所期望且如本文中所述的歪斜变体、pi变体、消融变体、另外的fc变体等。本发明提供其中cd3结合域序列如图12和13以及序列表中所示的单臂scfv-mab形式。本发明提供其中sstr2结合域序列如图11和序列表中所示的单臂scfv-mab形式。特别适用于与单臂scfv-mab形式一起使用的sstr2和cd3序列组合公开于图16中。此外,单臂scfv-mab形式的fc结构域大体上包括歪斜变体(例如,如图3和8中所示的一组氨基酸取代,且特别适用的歪斜变体选自由以下组成的群组:s364k/e357q:l368d/k370s;l368d/k370s:s364k;l368e/k370s:s364k;t411t/e360e/q362e:d401k;l368d/k370s:s364k/e357l;k370s:s364k/e357q;t366s/l368a/y407v:t366w以及t366s/l368a/y407v/y349c:t366w/s354c);任选地消融变体(包含图5中所示的那些);任选地带电scfv连接子(包含图7中所示的那些);以及包括pi变体的重链(包含图4中所示的那些)。在一些实施例中,单臂scfv-mab形式包含歪斜变体、pi变体以及消融变体。因此,单臂scfv-mab形式的一些实施例包括:a)第一单体,其包括:歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k以及重链可变域(其与轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv)以及结合cd3的scfv结构域;b)第二单体,其包含fc结构域,所述fc结构域具有歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k;以及c)轻链,其包括轻链可变域和轻链恒定域。在一些实施例中,单臂scfv-mab形式包含歪斜变体、pi变体、消融变体以及fcrn变体。因此,单臂scfv-mab形式的一些实施例包括:a)第一单体,其包括歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k、fcrn变体m428l/n434s以及重链可变域(其与轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv)以及结合cd3的scfv结构域;b)第二单体,其包含fc结构域,所述fc结构域具有歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k以及fcrn变体m428l/n434s;以及c)轻链,其包括轻链可变域和轻链恒定域。对于单臂scfv-mab形式,特别适用的cd3结合域序列包含但不限于:抗cd3h1.30_l1.47、抗cd3h1.32_l1.47、抗cd3h1.89_l1.47、抗cd3h1.90_l1.47、抗cd3h1.33_l1.47以及抗cd3h1.31_l1.47,以及图12和13中所描绘的那些。对于单臂scfv-mab形式,特别适用的sstr2结合域序列包含但不限于:抗sstr2h1.143_l1.30;抗sstr2h1_l1.1;抗sstr2h1.107_l1.30;抗sstr2h1.107_l1.67;抗sstr2h1.107_l1.108;抗sstr2h1.107_l1.111;抗sstr2h1.107_l1.114;抗sstr2h1.107_l1.102;抗sstr2h1.107_l1.110;抗sstr2h1.125_l1.30;抗sstr2h1.125_l1.67;抗sstr2h1.125_l1.108;抗sstr2h1.125_l1.111;抗sstr2h1.125_l1.114;抗sstr2h1.125_l1.102;以及抗sstr2h1.125_l1.10,以及图11和15以及seqidno:68至659中所列出的那些。scfv-mab本发明中特别适用的一种异源二聚支架是图1e中所示的mab-scfv形式。在这个实施例中,所述形式依赖于使用scfv的n端连接到其中一个单体,从而形成第三抗原结合域,其中两个单体的fab部分结合sstr2且“额外”scfv结构域结合cd3。在这个实施例中,第一单体包括第一重链(包括重链可变域和恒定域),且n端共价连接scfv,其包括scfv轻链可变域、scfv连接子以及scfv重链可变域,呈定向((vh1-scfv连接子-vl1-[任选的域连接子]-vh2-ch1-铰链-ch2-ch3)或(scfv呈相反定向)((vl1-scfv连接子-vh1-[任选的域连接子]-vh2-ch1-铰链-ch2-ch3))。这个实施例进一步使用包括轻链可变域和轻链恒定域的共用轻链,其与重链缔结而形成两个结合sstr2的相同fab。就本文中的许多实施例来说,这些构筑体包含所期望且如本文中所述的歪斜变体、pi变体、消融变体、另外的fc变体等。本发明提供其中cd3结合域序列如图12和13以及序列表中所示的scfv-mab形式。本发明提供其中sstr2结合域序列如图11和序列表中所示的scfv-mab形式。特别适用于与scfv-mab形式一起使用的sstr2和cd3序列组合公开于图16中。此外,scfv-mab形式的fc结构域大体上包括歪斜变体(例如,如图3和8中所示的一组氨基酸取代,且特别适用的歪斜变体选自由以下组成的群组:s364k/e357q:l368d/k370s;l368d/k370s:s364k;l368e/k370s:s364k;t411t/e360e/q362e:d401k;l368d/k370s:s364k/e357l;k370s:s364k/e357q;t366s/l368a/y407v:t366w以及t366s/l368a/y407v/y349c:t366w/s354c);任选地消融变体(包含图5中所示的那些);任选地带电scfv连接子(包含图7中所示的那些);以及包括pi变体的重链(包含图4中所示的那些)。在一些实施例中,scfv-mab形式包含歪斜变体、pi变体以及消融变体。因此,一些实施例包含scfv-mab形式,其包括:a)第一单体,其包括歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k以及重链可变域(其与共用轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv)以及结合cd3的scfv结构域;b)第二单体,其包括歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k以及重链可变域(其与常见轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv);以及c)共用轻链,包括轻链可变域和轻链恒定域。在一些实施例中,scfv-mab形式包含歪斜变体、pi变体、消融变体以及fcrn变体。因此,一些实施例包含scfv-mab形式,其包括:a)第一单体,其包括歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k、fcrn变体m428l/n434s以及重链可变域(其与共用轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv)以及结合cd3的scfv结构域;b)第二单体,其包括歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k、fcrn变体m428l/n434s以及重链可变域(其与共用轻链的轻链可变域一起构成如本文所概述的结合sstr2的fv);以及c)共用轻链,其包括轻链可变域和轻链恒定域。对图10中的于mab-scfv形式骨架1(任选地包含m428l/n434s),特别适用的cd3结合域序列包含但不限于:抗cd3h1.30_l1.47、抗cd3h1.32_l1.47、抗cd3h1.89_l1.47、抗cd3h1.90_l1.47、抗cd3h1.33_l1.47以及抗cd3h1.31_l1.47,以及图12和13中所描绘的那些。对于图10中的mab-scfv形式骨架1(任选地包含m428l/n434s),特别适用的sstr2结合域序列包含但不限于:抗sstr2h1.143_l1.30;抗sstr2h1_l1.1;抗sstr2h1.107_l1.30;抗sstr2h1.107_l1.67;抗sstr2h1.107_l1.108;抗sstr2h1.107_l1.111;抗sstr2h1.107_l1.114;抗sstr2h1.107_l1.102;抗sstr2h1.107_l1.110;抗sstr2h1.125_l1.30;抗sstr2h1.125_l1.67;抗sstr2h1.125_l1.108;抗sstr2h1.125_l1.111;抗sstr2h1.125_l1.114;抗sstr2h1.125_l1.102;以及抗sstr2h1.125_l1.10,以及图11和15以及seqidno:68至659中所列出的那些。双scfv形式本发明还提供如所属领域中已知且如图1b中所示的双scfv形式。在这个实施例中,sstr2×cd3异源二聚双特异性抗体由两个scfv-fc单体构成(两个单体皆呈(vh-scfv连接子-vl-[任选的域连接子]-ch2-ch3)形式或(vl-scfv连接子-vh-[任选的域连接子]-ch2-ch3)形式,或一个单体一种定向且另一个单体另一种定向)。本发明提供其中cd3结合域序列如图12和13以及序列表中所示的双scfv形式。本发明提供其中sstr2结合域序列如图11和序列表中所示的双scfv形式。特别适用于与双scfv形式一起使用的sstr2和cd3序列组合公开于图16中。在一些实施例中,双scfv形式包含歪斜变体、pi变体以及消融变体。因此,一些实施例包含双scfv形式,其包括:a)第一单体,其包括歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k以及结合cd3或sstr2的第一scfv;和b)第二单体,其包括歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k以及结合cd3或sstr2的第二scfv。在一些实施例中,双scfv形式包含歪斜变体、pi变体、消融变体以及fcrn变体。在一些实施例中,双scfv形式包含歪斜变体、pi变体以及消融变体。因此,一些实施例包含双scfv形式,其包括:a)第一单体,其包括歪斜变体s364k/e357q、消融变体e233p/l234v/l235a/g236del/s267k、fcrn变体m428l/n434s以及结合cd3或sstr2的第一scfv;和b)第二单体,其包括歪斜变体l368d/k370s、pi变体n208d/q295e/n384d/q418e/n421d、消融变体e233p/l234v/l235a/g236del/s267k、fcrn变体m428l/n434s以及结合cd3或sstr2的第二scfv。对于双scfv形式,特别适用的cd3结合域序列包含但不限于:抗cd3h1.30_l1.47、抗cd3h1.32_l1.47、抗cd3h1.89_l1.47、抗cd3h1.90_l1.47、抗cd3h1.33_l1.47以及抗cd3h1.31_l1.47,以及图12和13中所描绘的那些。对于双scfv形式,特别适用的sstr2结合域序列包含但不限于:抗sstr2h1.143_l1.30;抗sstr2h1_l1.1;抗sstr2h1.107_l1.30;抗sstr2h1.107_l1.67;抗sstr2h1.107_l1.108;抗sstr2h1.107_l1.111;抗sstr2h1.107_l1.114;抗sstr2h1.107_l1.102;抗sstr2h1.107_l1.110;抗sstr2h1.125_l1.30;抗sstr2h1.125_l1.67;抗sstr2h1.125_l1.108;抗sstr2h1.125_l1.111;抗sstr2h1.125_l1.114;抗sstr2h1.125_l1.102;以及抗sstr2h1.125_l1.10,以及图11和15以及seqidno:68至659中所列出的那些。单特异性单克隆抗体如所属领域的技术人员将了解,本文概述的新颖fv序列也可以用于单特异性抗体(例如“传统”单克隆抗体)或非异源二聚双特异性形式。因此,本发明提供单克隆(单特异性)抗体,其包括图中的6个cdr和/或vh和vl序列,通常具有igg1、igg2、igg3或igg4恒定区,在一些实施例中,igg1、igg2以及igg4(包含包括s228p氨基酸取代的igg4恒定区)特别适用。即,本文中具有“h_l”名称的任何序列可以连接于人类igg1抗体的恒定区。i.目标抗原的抗原结合域本发明的双特异性抗体具有以二价、双特异性形式或三价双特异性形式结合于两个不同的目标检查点抗原(“目标对”)的两个不同的抗原结合域(abd),大体如图1中所示。应注意,对于各对,通常这些双特异性抗体命名为“抗sstr2×抗cd3”或通常简化或简便(且因此互换地)为“sstr2×cd3”等。应注意,除非本文中特别规定,否则名称中抗原列表的顺序不赋予结构;亦即sstr2×cd3开瓶器抗体可具有结合sstr2或cd3的scfv,尽管在一些情况下,顺序规定所指示的结构。如本文更全面地概述,这些abd组合可以呈各种形式,如下文所概述,通常呈其中一个abd在fab形式中且另一个在scfv形式中的组合。如本文所论述且图1中所展示,一些形式使用单个fab和单个scfv(图1a、1c以及1d),并且一些形式使用两个fab和单个scfv(图1e、1f以及1i)。抗原结合域如本文所论述,主题异源二聚抗体包含两个抗原结合域(abd),其中每一个结合sstr2或cd3。如本文所概述,这些异源二聚抗体可为双特异性且二价(例如以图1a中所描绘的形式,每个抗原结合于单个abd)或双特异性且三价(例如如图1f中所描绘,一个抗原结合于单个abd且另一个抗原结合于两个)。此外,一般来说,其中一个abd包括如本文所概述的scfv,所述scfv由n端至c端呈vh-scfv连接子-vl或vl-scfv连接子-vh的定向。其它abd中的一个或两个,根据所述形式,通常是fab,其包括一个蛋白质链上的vh域(通常作为重链的组分)和另一个蛋白质链上的vl(通常作为轻链的组分)。本发明提供多种结合多种不同检查点蛋白的abd,如下文所概述。如所属领域的技术人员将了解,任何一组6个cdr或vh域和vl域可以在scfv形式或fab形式中,其随后添加到重链恒定域和轻链恒定域,其中重链恒定域包括变体(包含在ch1域以及fc结构域内)。含于序列表中的scfv序列使用特定带电连接子,但如本文所概述,可使用不带电连接子或其它带电连接子,包含图7中所描绘的那些。此外,如上文所论述,序列表中所使用用于标识cdr的编号是kabat,然而,可使用不同编号,其将改变如表1中所示的cdr的氨基酸序列。对于本文中所列出的所有重链可变域和轻链可变域,可以制得其它变体。如本文所概述,在一些实施例中,所述组6个cdr可具有0、1、2、3、4或5个氨基酸修饰(特别适用的是氨基酸取代),以及重链可变域和轻链可变域的框架区中的变化,只要框架(不包含cdr)保留与选自美国专利第7,657,380号(其图和图例以全文引用的方式并入本文中)的图1中所列出的序列的人类生殖系序列的至少约80、85或90%一致性。因此,例如,如本文所述的相同的cdr可以与来自人类生殖系序列的不同框架序列组合,只要框架区保留与选自美国专利第7,657,380号的图1中所列出的序列的人类生殖系序列的至少约80、85或90%一致性。或者,cdr可具有氨基酸修饰(例如所述组cdr中的1、2、3、4或5个氨基酸修饰(即,cdr可为经修饰的,只要所述组6个cdr中的变化的总数小于6个氨基酸修饰,其中改变的cdr呈任何组合;例如,vlcdr1中可存在一个变化,vhcdr2中可存在两个变化,vhcdr3中可不存在变化等)),以及具有框架区变化,只要框架区保留与选自美国专利第7,657,380号的图1中所列出的序列的人类生殖系序列的至少80、85或90%一致性。sstr2抗原结合域在一些实施例中,其中一个abd结合sstr2。适合的6个cdr组和/或vh域和vl域,以及scfv序列,描绘在图11和序列表中。特别适用的sstr2结合域序列包含但不限于:抗sstr2h1.143_l1.30;抗sstr2h1_l1.1;抗sstr2h1.107_l1.30;抗sstr2h1.107_l1.67;抗sstr2h1.107_l1.108;抗sstr2h1.107_l1.111;抗sstr2h1.107_l1.114;抗sstr2h1.107_l1.102;抗sstr2h1.107_l1.110;抗sstr2h1.125_l1.30;抗sstr2h1.125_l1.67;抗sstr2h1.125_l1.108;抗sstr2h1.125_l1.111;抗sstr2h1.125_l1.114;抗sstr2h1.125_l1.102;以及抗sstr2h1.125_l1.10,以及图11和15以及seqidno:68至659中所列出的那些。如所属领域的技术人员将了解,适合的sstr2结合域可包括如序列表和图中所描绘的一组6个cdr,其为呈带下划线的形式的,或在使用如本文所述的且如表1中所示的不同编号方案的情况下,所述cdr使用在图11中所描绘的序列的vh和vl序列内的其它比对鉴别出。适合的abd也可以包含如这些序列和图中所描绘的整个vh和vl序列,用作scfv或用作fab。在本文中的许多含有针对sstr2的fv的实施例中,fab单体结合sstr2。除在图和序列表中所公开的形成sstr2的abd的亲本cdr组以外,本发明提供变体cdr组。在一个实施例中,一组6个cdr可具有亲本cdr的1、2、3、4或5个氨基酸变化,只要sstr2abd仍能够结合目标抗原,如由biacore、表面等离子共振(surfaceplasmonresonance,spr)和/或bli(生物膜干涉法,例如octet分析)分析中的至少一个所测量,在许多实施例中,后者特别适用。除本文中所公开的形成sstr2的abd的亲本重链可变域和轻链可变域以外,本发明提供变体vh域和vl域。在一个实施例中,变体vh域和vl域各自可具有来自亲本vh和vl域的1、2、3、4、5、6、7、8、9或10个氨基酸变化,只要abd仍能够结合目标抗原,如biacore、表面等离子共振(spr)和/或bli(生物膜干涉法,例如octet分析)分析中的至少一个所测量,在许多实施例中,后者特别适用。在另一实施例中,变体vh和vl与对应的亲本vh和vl至少90、95、97、98或99%一致,只要abd仍能够结合目标抗原,如biacore、表面等离子共振(spr)和/或bli(生物膜干涉法,例如octet分析)分析中的至少一个所测量,在许多实施例中,后者特别适用。特别优选的实施例包含h1.143_l1.30sstr2抗原结合域,作为“fab”包含在图9中的任一种开瓶器形式骨架内。cd3抗原结合域在一些实施例中,其中一个abd结合cd3。适合的6个cdr组和/或vh域和vl域,以及scfv序列,描绘在图12和13以及序列表中。特别适用的cd3结合域序列包含但不限于:抗cd3h1.30_l1.47、抗cd3h1.32_l1.47、抗cd3h1.89_l1.47、抗cd3h1.90_l1.47、抗cd3h1.33_l1.47以及抗cd3h1.31_l1.47,以及图12和13中所描绘的那些。如所属领域的技术人员将了解,适合的cd3结合域可包括如序列表和图中所描绘的一组6个cdr,其为呈带下划线的形式的,或在使用如本文所述的且如表1中所示的不同编号方案的情况下,所述cdr使用在图11中所描绘的序列的vh和vl序列内的其它比对鉴别出。适合的abd也可以包含如这些序列和图中所描绘的整个vh和vl序列,用作scfv或用作fab。在本文中的许多含有针对cd3的fv的实施例中,scfv单体结合cd3。除在图和序列表中所公开的形成cd3的abd的亲本cdr组以外,本发明提供变体cdr组。在一个实施例中,一组6个cdr可具有亲本cdr的1、2、3、4或5个氨基酸变化,只要cd3abd仍能够结合目标抗原,如由biacore、表面等离子共振(spr)和/或bli(生物膜干涉法,例如octet分析)分析中的至少一个所测量,在许多实施例中,后者特别适用。除本文中所公开的形成cd3的abd的亲本重链可变域和轻链可变域以外,本发明提供变体vh域和vl域。在一个实施例中,变体vh域和vl域各自可具有来自亲本vh和vl域的1、2、3、4、5、6、7、8、9或10个氨基酸变化,只要abd仍能够结合目标抗原,如biacore、表面等离子共振(spr)和/或bli(生物膜干涉法,例如octet分析)分析中的至少一个所测量,在许多实施例中,后者特别适用。在另一实施例中,变体vh和vl与对应的亲本vh和vl至少90、95、97、98或99%一致,只要abd仍能够结合目标抗原,如biacore、表面等离子共振(spr)和/或bli(生物膜干涉法,例如octet分析)分析中的至少一个所测量,在许多实施例中,后者特别适用。特别优选的实施例包含h1.30_l1.47cd3抗原结合域,作为“fab”包含在图9中的任一种开瓶器形式骨架内。j.适用的实施例在一个实施例中,在本发明中适用的歪斜变体与pi变体的特定组合是t366s/l368a/y407v:t366w(任选地包含桥接二硫化物,t366s/l368a/y407v/y349c:t366w/s354c)其中一个单体包括q295e/n384d/q418e/n481d且另一个单体包括带正电的scfv连接子(当所述形式包含scfv结构域时)。如在所属领域中将了解,“臼包杵”型变体不改变pi,且因此可用于任一单体。k.本发明的核酸本发明进一步提供编码本文所提供的抗sstr2抗体的核酸组合物,其包含但不限于抗sstr2×抗cd3双特异性抗体和sstr2单特异性抗体。如所属领域的技术人员将了解,核酸组合物将取决于异源二聚蛋白质的形式和支架。因此,例如,当所述形式需要三种氨基酸序列时,如对于三f形式(例如第一氨基酸单体包括fc结构域和scfv,第二氨基酸单体包括重链和轻链),可在用于表达的一个或更多个表达载体中并入三种核酸序列。类似地,一些形式(例如,如图1中所公开的双scfv形式)仅需要两种核酸;同样,其可以放入一个或两个表达载体中。如所属领域中所知,取决于用于产生本发明的异源二聚抗体的宿主细胞,可以将编码本发明组分的核酸并入如所属领域中已知的表达载体中。一般来说,核酸可操作地连接于任何数目的调节元件(启动子、复制起点、可选标记、核糖体结合位点、诱导剂等)。表达载体可以是额外染色体或整合载体。然后使本发明的核酸和/或表达载体在任何数目的不同类型的如所属领域中熟知的宿主细胞(包含哺乳动物、细菌、酵母、昆虫和/或真菌细胞)中转化,在许多实施例中,适用哺乳动物细胞(例如cho细胞)。在一些实施例中,根据所述形式,编码每种单体的核酸和编码轻链的任选核酸在适当时各自含在单一表达载体内(通常在不同或相同启动子控制下)。在特别用于本发明的实施例中,不同表达载体上均含有这两种或三种核酸中的每一种。如本文中和62/025,931(以引用的方式并入本文中)所示,可以利用不同的载体比率驱动异源二聚体形成。即,出乎意料地,虽然蛋白质包括1:1:2比率的第一单体:第二单体:轻链(在本文中的许多实施例具有构成异源二聚抗体的三种多肽的情况下),但是这些并非是得到最佳结果的比率。本发明的异源二聚抗体是通过培养包括如所属领域中熟知的表达载体的宿主细胞来产生。一经产生,则进行传统的抗体纯化步骤,包括离子交换色谱步骤。如本文所论述,使两个单体的pi相差至少0.5可以允许通过离子交换色谱或等电点聚焦或对等电点敏感的其它方法来实现分离。即,包涵改变每个单体的等电点(pi)的pi取代,使得每个单体具有不同pi且异源二聚体也具有不同pi,从而促进“三重f”异源二聚体的等电纯化(例如阴离子交换柱、阳离子交换柱)。这些取代还有助于测定和监测纯化(例如ief凝胶、cief和分析型iex柱)后受到污染的任何双scfv-fc和mab同型二聚体。l.异源二聚检查点抗体的生物和生化功能总体而言,向癌症患者施用本发明的双特异性sstr2×cd3抗体,以如本文所述的多种方式评估功效。因此,虽然可以进行标准的功效分析,如癌症负荷、肿瘤尺寸、转移的存在或程度的评估等,但是也可以基于免疫状况评估来评定免疫肿瘤学治疗。这可以用许多方式进行,包含体外和体内分析。举例来说,可以与“旧式”测量一起评估免疫状况变化(例如,在ipi治疗后icos+cd4+t细胞的存在),“旧式”测量如肿瘤负荷、尺寸、浸润性、ln参与、转移等。因此,可以评估以下中的任一个或全部:检查点对cd4+t细胞活化或增殖、cd8+t(ctl)细胞活化或增殖、cd8+t细胞介导的细胞毒性活性和/或ctl介导的细胞消除、nk细胞活性以及nk介导的细胞消除的抑制性作用。在一些实施例中,通过使用例如cfse稀释法、免疫效应细胞的ki67胞内染色以及3h-胸苷掺入法评估免疫细胞增殖来评定治疗。在一些实施例中,通过评估基因表达的增加量或活化相关标记的增加的蛋白质水平以及通过cd107a的表面表达测量的细胞脱粒来评定治疗,所述标记包含以下中的一个或多个:cd25、cd69、cd137、icos、pd1、gitr、ox40。一般来说,如本领域中已知地进行基因表达分析。一般来说,还如所属领域中已知地,类似地进行蛋白质表达测量。在一些实施例中,通过评估众多细胞参数来评定通过目标细胞活力检测测量的细胞毒性活性,从而评定治疗,所述细胞参数,如酶活性(包含蛋白酶活性)、细胞膜渗透性、细胞粘附、atp产生、辅酶产生以及核苷酸吸收活性。这些分析的特定实例包含但不限于:台盼蓝(trypanblue)或pi染色、51cr或35s释放法、ldh活性、mtt和/或wst分析、钙黄绿素-am分析、基于发光的分析以及其它分析。在一些实施例中,通过评定通过细胞因子产生测量的t细胞活性,使用细胞因子,使用众所周知的技术,细胞内测量培养物上清液来评定治疗,所述细胞因子包含但不限于:ifnγ、tnfα、gm-csf、il2、il6、il4、il5、il10、il13。因此,可以使用评估以下中的一个或多个的分析来进行评定治疗:(i)增加免疫反应;(ii)增加αβ和/或γδt细胞的活化;(iii)增加细胞毒性t细胞活性;(iv)增加nk和/或nkt细胞活性;(v)缓解αβ和/或γδt细胞抑制;(vi)增加促炎性细胞因子分泌;(vii)增加il-2分泌;(viii)增加干扰素γ产生;(ix)增加th1反应;(x)降低th2反应;(xi)降低或消除至少一种调节性t细胞(treg)的细胞数目和/或活性。测量功效的分析在一些实施例中,使用如所属领域中已知的混合淋巴细胞反应(mixedlymphocytereaction,mlr)分析评定t细胞活化。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量免疫反应的增加或减少,如例如通过不同因子的磷酸化或去磷酸化或通过测量其它翻译后修饰所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量αβ和/或γδt细胞的活化的增加或减少,如例如通过细胞因子分泌或通过增殖或通过如例如cd137、cd107a、pd1等活化标记的表达的变化所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量细胞毒性t细胞活性的增加或减少,如例如通过直接杀死如例如癌细胞的目标细胞或通过细胞因子分泌或通过增殖或通过如例如cd137、cd107a、pd1等活化标记的表达的变化所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量nk和/或nkt细胞活性的增加或减少,如例如通过直接杀死如例如癌细胞的目标细胞或通过细胞因子分泌或通过如例如cd107a等活化标记的表达的变化所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量αβ和/或γδt细胞抑制的增加或减少,如例如通过细胞因子分泌或通过增殖或通过如例如cd137、cd107a、pd1等活化标记的表达的变化所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量促炎性细胞因子分泌的增加或减少,如例如通过elisa或通过基于珠粒的luminex或multiplex方法(luminex/multiplexbeadbasedmethod)或通过胞内染色和facs分析或通过alispot等所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量il-2分泌的增加或减少,如例如通过elisa或通过基于珠粒的luminex或multiplex方法或通过胞内染色和facs分析或通过alispot等所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量干扰素γ的产生的增加或减少,如例如通过elisa或通过基于珠粒的luminex或multiplex方法或通过胞内染色和facs分析或通过alispot等所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量th1反应的增加或减少,如例如通过细胞因子分泌或通过活化标记的表达的变化所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量th2反应的增加或减少,如例如通过细胞因子分泌或通过活化标记的表达的变化所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量调节性t细胞(treg)中的至少一种的细胞数目和/或活性的增加或减少,如例如通过流式细胞术或通过ihc所测量。反应减少指示免疫刺激活性。恰当的减少与增加相同,概述于下文中。在一个实施例中,信号传导路径分析测量m2巨噬细胞的细胞数目的增加或减少,如例如通过流式细胞术或通过ihc所测量。反应减少指示免疫刺激活性。恰当的减少与增加相同,概述于下文中。在一个实施例中,信号传导路径分析测量m2巨噬细胞促致瘤(pro-tumorigenic)活性的增加或减小,如例如通过细胞因子分泌或通过活化标记的表达的变化所测量。反应减少指示免疫刺激活性。恰当的减少与增加相同,概述于下文中。在一个实施例中,信号传导路径分析测量n2嗜中性粒细胞的增加或减少,如例如通过流式细胞术或通过ihc所测量。反应减少指示免疫刺激活性。恰当的减少与增加相同,概述于下文中。在一个实施例中,信号传导路径分析测量n2嗜中性粒细胞促致瘤活性的增加或减小,如例如通过细胞因子分泌或通过活化标记的表达的变化所测量。反应减少指示免疫刺激活性。恰当的减少与增加相同,概述于下文中。在一个实施例中,信号传导路径分析测量t细胞活化的抑制的增加或减少,如例如通过细胞因子分泌或通过增殖或通过如例如cd137、cd107a、pd1等活化标记的表达的变化所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量ctl活化的抑制的增加或减少,如例如通过直接杀死如例如癌细胞的目标细胞或通过细胞因子分泌或通过增殖或通过如例如cd137、cd107a、pd1等活化标记的表达的变化所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量αβ和/或γδt细胞耗竭的增加或减少,如例如通过活化标记的表达的变化所测量。反应减少指示免疫刺激活性。恰当的减少与增加相同,概述于下文中。在一个实施例中,信号传导路径分析测量αβ和/或γδt细胞反应的增加或减少,如例如通过细胞因子分泌或通过增殖或通过如例如cd137、cd107a、pd1等活化标记的表达的变化所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量抗原特异性记忆反应的刺激的增加或减少,如例如通过细胞因子分泌或通过增殖或通过如例如cd45ra、ccr7等活化标记的表达的变化所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量癌细胞的细胞凋亡或溶解的增加或减少,如例如通过细胞毒性分析(如例如mtt、cr释放、calcineam)或通过基于流式细胞术的分析(如例如cfse稀释或碘化丙啶染色等)所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量对癌细胞的细胞毒性或细胞生长抑制作用的刺激的增加或减少,如例如通过细胞毒性分析(如例如mtt、cr释放、calcineam)或通过基于流式细胞术的分析(如例如cfse稀释或碘化丙啶染色等)所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量癌细胞的直接杀死的增加或减少,如例如通过细胞毒性分析(如例如mtt、cr释放、calcineam)或通过基于流式细胞术的分析(如例如cfse稀释或碘化丙啶染色等)所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量th17活性的增加或减小,如例如通过细胞因子分泌或通过增殖或通过活化标记的表达的变化所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,信号传导路径分析测量补体依赖性细胞毒性和/或抗体依赖性细胞介导的细胞毒性的诱导的增加或减少,如例如通过细胞毒性分析(如例如mtt、cr释放、calcineam)或通过基于流式细胞术的分析(如例如cfse稀释或碘化丙啶染色等)所测量。活性增加指示免疫刺激活性。下文概述了恰当的活性增加。在一个实施例中,例如通过直接杀死如例如癌细胞的目标细胞或通过细胞因子分泌或通过增殖或通过如例如cd137、cd107a、pd1等活化标记的表达的变化测量t细胞活化。关于t细胞,增殖、细胞表面活化标记(例如,cd25、cd69、cd137、pd1)、细胞毒性(杀死目标细胞的能力)以及细胞因子产生(例如,il-2、il-4、il-6、ifnγ、tnf-α、il-10、il-17a)的增加将指示与增强的癌细胞杀死一致的免疫调节。在一个实施例中,例如通过直接杀死如例如癌细胞的目标细胞或通过细胞因子分泌或通过如例如cd107a等活化标记的表达的变化测量nk细胞活化。关于nk细胞,增殖、细胞毒性(杀死目标细胞并提高cd107a、颗粒酶和穿孔素表达的能力)、细胞因子产生(例如,ifnγ和tnf)以及细胞表面受体表达(例如,cd25)的增加将指示与增强的癌细胞杀死一致的免疫调节。在一个实施例中,例如通过细胞因子分泌或通过增殖或通过活化标记的表达的变化测量γδt细胞活化。在一个实施例中,例如通过细胞因子分泌或通过活化标记的表达的变化测量th1细胞活化。活性或反应的恰当增加(或恰当减少,如上文所概述的)是相比于在参考样品或对照样品中的信号增加10%、20%、30%、40%、50%、60%、70%、80%、90%、95%或98%到99%的百分比,参考样品或对照样品是例如不含本发明的抗体的测试样品。类似地,相比于参考样品或对照样品增加至少一倍、两倍、三倍、四倍或五倍显示有功效。m.治疗本发明的组合物在制备后可以用于许多应用。在神经内分泌肿瘤(net,例如肺癌、胃肠癌、胰脏癌、垂体癌、髓质癌、前列腺、胰脏肺类癌、骨肉瘤、支气管瘤、胸腺瘤)以及非net(乳癌、肺癌、结肠直肠癌、卵巢癌、子宫颈癌)中sstr2高度表达。相应地,本发明的异源二聚组合物可用于治疗此类sstr2阳性癌症。用于活体内施用的抗体组合物通过将具有期望纯度的抗体与任选的药学上可接受的载剂、赋形剂或稳定剂混合来制备根据本发明使用的抗体的调配物(《雷明顿医药科学(remington'spharmaceuticalsciences)》第16版,osol,a.编[1980]),用于以冻干调配物或水溶液形式储存。可接受的载剂、赋形剂或稳定剂在使用剂量和浓度下对接受者无毒性,并且包含缓冲液,如磷酸盐、柠檬酸盐以及其它有机酸;抗氧化剂,包含抗坏血酸和甲硫氨酸;防腐剂(如十八烷基二甲基苯甲基氯化铵;氯化六羟季铵;苯扎氯铵(benzalkoniumchloride)、苄索氯铵(benzethoniumchloride);酚、丁醇或苯甲醇;对羟基苯甲酸烷酯,如对羟基苯甲酸甲酯或对羟基苯甲酸丙酯;儿茶酚;间苯二酚;环己醇;3-戊醇;以及间甲酚);低分子量(小于约10个残基)多肽;蛋白质,如血清白蛋白、明胶或免疫球蛋白;亲水性聚合物,如聚乙烯吡咯烷酮;氨基酸,如甘氨酸、谷氨酰胺、天冬酰胺、组氨酸、精氨酸或赖氨酸;单糖、二糖以及其它碳水化合物,包括葡萄糖、甘露糖或糊精;螯合剂,如edta;糖,如蔗糖、甘露糖醇、海藻糖或山梨糖醇;成盐相对离子,如钠;金属络合物(例如zn-蛋白质络合物);和/或非离子型表面活性剂,如tweentm、pluronicstm或聚乙二醇(peg)。施用模式根据已知方法,如作为推注静脉内施用或通过连续输注一段时间,来向受试者施用本发明的抗体和化疗剂。治疗模式在本发明的方法中,疗法用于提供相对于疾病或病况而言积极的治疗性反应。“积极治疗性反应”是预期疾病或病况的改善,和/或与疾病或病况相关联的症状的改善。例如,积极治疗性反应将指疾病的以下中的一个或多个改善:(1)赘生性细胞数目减少;(2)赘生性细胞死亡增加;(3)赘生性细胞存活抑制;(5)肿瘤生长抑制(即,在一定程度上减慢,优选地中断);(6)患者生存率增加;以及(7)与疾病或病况相关联的一种或多种症状有一定缓解。任何给定疾病或病况中的积极治疗性反应可通过特定针对于所述疾病或病况的标准化反应指标测定。可以使用筛检技术,如磁共振成像(magneticresonanceimaging,mri)扫描、x-放射照相成像、计算机断层(computedtomographic,ct)扫描、骨骼扫描成像、内窥镜检查以及肿瘤活检取样(包含骨髓穿刺(bonemarrowaspiration,bma)和循环中肿瘤细胞计数),针对肿瘤形态变化(即,总肿瘤负荷、肿瘤尺寸等等)评定肿瘤反应。除这些积极治疗性反应以外,经历疗法的受试者可能受到与疾病相关联的症状得到改善的有利影响。根据本发明的治疗包含使用“治疗有效量”的药剂。“治疗有效量”是指在必需剂量下且在必需时间内有效实现所需治疗结果的量。治疗有效量可能根据如疾病状态、个体的年龄、性别和体重以及药剂在个体体内引发期望反应的能力的因素而不同。治疗有效量还是治疗有利作用超过抗体或抗体部分的任何毒性或有害作用的量。肿瘤疗法的“治疗有效量”也可通过其稳定疾病进展的能力测量。化合物抑制癌症的能力可在可预测对人类肿瘤的功效的动物模型系统中进行评估。或者,组合物的这种特性可以通过有经验的医师所知的活体外分析检查化合物的抑制细胞生长或诱导细胞凋亡的能力来评估。治疗有效量的治疗性化合物能够减小肿瘤尺寸,或以其它方式改善受试者的症状。所属领域的一般技术人员将能够根据如受试者身材、受试者症状的严重度以及所选择的特定组合物或施用途径的因素决定这一量。可以调整剂量方案以提供最佳期望反应(例如一种治疗性反应)。举例来说,如由治疗情况的紧急状态所指示的,可以施用单次推注,可以随着时间施用若干分次剂量,或可以按比例地减少或增加剂量。可以单位剂型形式调配肠胃外组合物以便于施用且使剂量均匀。如本文所使用,剂量单位形式是指适合作为单一剂量用于待治疗受试者的物理分散单元;每个单元含有经计算以与所需药物载剂结合产生期望治疗效果的预定量的活性化合物。本发明的单位剂型的规格由以下各项指定并且直接取决于以下各项:(a)活性化合物的独特特征及欲实现的特定治疗效果;和(b)混配这种治疗性化合物用于治疗个体敏感性在所属领域中固有的限制。本发明中所用的双特异性抗体的高效剂量和剂量方案取决于待治疗的疾病或病况且可以由所属领域的技术人员来决定。本发明中所用的双特异性抗体的治疗有效量的示范性非限制性范围为约0.1至100mg/kg。所有参考文献都明确地以全文引用的方式并入本文中。虽然上文出于说明目的已描述了本发明的特定实施例,所属领域的技术人员应了解,在不脱离如所附权利要求书中所描述的本发明的情况下可以对细节进行多种变化。实例下文提供实例来说明本发明。这些实例不意图将本发明限制于任何特定的应用或操作理论。对于本发明中所论述的所有恒定区位置,根据如kabat中的eu索引(kabat等人,1991,《免疫学所关注的蛋白质序列》第5版,美国公共卫生署,美国国家卫生研究院,贝塞斯达,其以引用的方式整体并入本文中)。抗体领域中的技术人员将了解这个规约由以下组成:免疫球蛋白序列的特定区域的非顺序编号、对免疫球蛋白家族中的保守位置实现标准化的称呼。相应地,如根据eu索引界定的任何指定免疫球蛋白的位置不一定对应于其顺序序列。美国公开2015/0307629、2014/0288275和wo2014/145806概述了通用和特别的科学技术,所有这些文献明确地以全文(且尤其其中概述的技术)引用的方式并入。实例1:抗sstr2×抗cd3双特异性抗体的生成1a:抗sstr2fab臂的生成工程改造抗sstr2抗体的亲本可变区用作本发明的抗sttr2×抗cd3双特异性抗体的组分。如先前在2010年2月2日发布的美国专利第7,657,380号中所描述进行鼠类vh区和vl区的人源化。经由quikchange(stratagene,得克萨斯州雪松溪(cedarcreek,tx.))诱变进行氨基酸取代以尝试凭借改良特性鉴别变体。1b:双特异性抗体产生抗sstr2×抗cd3双特异性形式的卡通示意图显示于图1中。使用如上文所描述的衍生自经工程改造的抗sstr2抗体的抗sstr2fab臂和抗cd3scfv臂产生示范性抗体。示范性抗sstr2×抗cd3开瓶器抗体xenp018087和xenp018907分别显示于图14和15中。编码双特异性表达所需的三种链的dna通过基因合成(华盛顿州博塞尔的蓝鹭生物技术(blueheronbiotechnology,bothell,wash.))且使用ptt5技术将其标准亚克隆到表达载体中来生成或通过quikchange诱变来生成。将dna转染到hek293e细胞中用于表达且使用蛋白质a亲和分析(通用电气医疗集团(gehealthcare))和阳离子交换色谱从上清液中纯化所得蛋白质。使用hitrapsphp色谱柱(通用电气医疗集团)与ph6.0的50mmmes的洗涤/平衡缓冲液和ph6.0的50mmmes且+1mnacl线性梯度的洗脱缓冲液进行阳离子交换色谱纯化。1c:抗sstr2抗体双特异性结合.使用人类sstr2转染的cho细胞评定抗sstr2抗体和示范性抗sstr2×抗cd3双特异性抗体的细胞表面结合。将细胞与指定测试品一起在冰上培育45分钟并离心。将细胞用含有经藻红蛋白(pe)标记的二级抗体(μg/ml;山羊抗人igg)的染色缓冲液再悬浮,且随后在冰上培育45分钟。将细胞离心两次且随后用染色缓冲液再悬浮。通过流式细胞术测量结合(图17a-17p)。实例2:示范性抗sstr2×抗cd3双特异性抗体的表征2a:示范性抗sstr2×抗cd3双特异性抗体的活体外表征针对在sstr2转染的cho细胞(图18a-18d)和sstr2阳性tt细胞(人类甲状腺髓质癌细胞株;图19a-19c)上的再导向t细胞细胞毒性(rtcc)来活体外表征示范性抗sstr2×抗cd3fab-scfv-fc双特异性抗体。rtcc通过测量乳酸脱氢酶(ldh)含量确定。如这些图中所示,相比于对照组,抗sstr2×抗cd3fab-scfv-fc双特异性抗体在sstr2转染的cho细胞中以及人类癌细胞株中展现高rtcc百分比。2b:示范性抗sstr2×抗cd3双特异性抗体的活体内表征在第一个研究中,向食蟹猕猴(n=3)施用两次(在0周和3周时)静脉内(i.v.)给药0.03mg/kgxenp18087或1mg/kgxenp18088。随后评定这些抗sstr2×抗cd3双特异性抗体对cd4+和cd8+t细胞活化的影响(如由cd69表达(图20a)和cd4+和cd8+t细胞分布(图20b)指示)。在第二个研究中,向食蟹猕猴(n=3)施用单次静脉内(i.v.)给药抗sstr2×抗cd3双特异性抗体:0.06mg/kgxenp18087、0.1mg/kgxenp18907、0.5mg/kgxenp18907或2mg/kgxenp18907。评定这些抗sstr2×抗cd3双特异性抗体对cd4+和cd8+t细胞活化(cd69上调,图21a)以及cd4+和cd8+t细胞再分布(细胞计数,图21b)的影响。此外,进行葡萄糖耐量测试(gtt)(图21c和21d)以评定所测试的受试者分解葡萄糖的能力。对于gtt,在8个不同时间点收集血液样品:给药前、施用右旋糖之后5、10、20、30、40、60以及90分钟。如这些研究中所示,在每个处理期间cd4+和cd8+从血液中快速再分布且在给药之后随后恢复并常态化(图21b)。t细胞在给药时立即活化(图21a)且随后消退同时t细胞再分布。在第三个研究中,向食蟹猕猴(n=3)施用两次(在0周和1周时)静脉内(i.v.)给药0.001或0.01mg/kgxenp18087。随后使用cd69表达(即t细胞活化的标记)评定这些抗sstr2×抗cd3双特异性抗体对cd4+和cd8+t细胞活化(图22a-22b)以及cd4+和cd8+t细胞再分布(图22c-22d)的影响。分析这些猴的血清il-6含量和tnf含量(图22e-22f)。如这些研究中所示,在每个处理期间cd4+和cd8+从血液中快速再分布且在给药之后随后恢复并常态化。在每次以剂量依赖性方式给药时t细胞立即活化且随后消退同时t细胞再分布。il-6和tnf细胞因子释放与t细胞活化相关。实例3:xmab18087的评估3a:xmab18087与人类和食蟹猕猴sstr2的特异性结合使用人类sstr2转染的cho细胞和食蟹猕猴sstr2转染的cho细胞评定xmab18087和对照组抗rsv×抗cd3双特异性抗体(xenp13245)的细胞表面结合。还使用亲本cho细胞作为对照组评定细胞表面结合。通过流式细胞术使用如实例1c中所大体描述的经藻红蛋白(pe)标记的二级抗体测量结合。xmab18087不仅结合细胞表面人类sstr2(图23a)还与食蟹猕猴sstr2(图23b)交叉反应,而对照组抗rsv×抗cd3双特异性抗体xenp13245不结合人类sstr2或食蟹猕猴sstr2转染的cho细胞。数据进一步显示,xmab18087不结合未转染的亲本cho细胞(图23c),显示了xmab18087的特异性。3b:通过xmab18087引起的再导向t细胞细胞毒性针对sstr2转染的cho细胞(图24a)、sstr2阳性tt细胞(甲状腺髓质癌细胞株;图24b和25)、a549细胞(肺腺癌细胞株;图24c和25)以及作为对照组的未转染的亲本cho细胞(图24a)的再导向t细胞细胞毒性(rtcc)活体外表征xmab18087。包含抗rsv×抗cd3双特异性抗体(xenp13245)和二价抗sstr2mah作为对照组(图24d)。将目标细胞和人类pbmc与xmab18087或xenp13245以e:t为10或20:1的比率一起培育24小时。通过测量乳酸脱氢酶(ldh)含量确定rtcc。如这些图中所示,相比于对照组抗cd3双特异性抗体xenp13245和对照组二价抗sstr2mah(图25),xmab18087在sstr2转染的cho细胞(图24a)以及人类癌细胞株(图28b-28c和24d)中展现高rtcc百分比。此外,数据显示,xmab18087在未转染的亲本cho细胞中不展现rtcc(图24a)。在实验中还使用sstr2转染的cho细胞和tt细胞通过利用流式细胞术评估cd8+和cd4+t细胞上的cd69的表面表达来研究通过xmab18087引起的t细胞活化(图26a-26b)。如图中所示,xmab18087使cd8+和cd4+t细胞活化达到远高于对照组抗cd3双特异性抗体xenp13245所达到的含量。这一结果表明,xmab18087通过诱导t细胞活化来消除sstr2+目标细胞。3c:xmab18087在移植有a549肺癌细胞和人类pbmc的nsg小鼠中展现抗肿瘤活性在第7天时,将二十五只非肥胖性糖尿病重症联合免疫缺陷γ(nodscidgamma,nsg)小鼠每只移植1×106a549-redfluc肿瘤细胞(0.1ml体积皮下注射)。第0天,将10×106个人类pbmc腹膜内移植到小鼠中。在第0天移植pbmc之后,每周(第0天、第7天以及第14天)通过以3.0mg/kg腹膜内注射给药xmab18087(对照组小鼠给药pbs)。研究设计进一步概述于图27中。通过使用活体外成像系统(luminaiii)测量每只小鼠的总通量来监测肿瘤生长。如图28和29中所示,相比于用pbs处理,用3mg/kgxmab18087处理显著抑制a549局部肿瘤生长。3d:在食蟹猕猴中xmab18087的表征在进一步研究中,向食蟹猕猴(n=3)施用单次静脉内(i.v.)给药xmab18087或对照组抗rsv×抗cd3双特异性抗体(xenp13245)。评定这些双特异性抗体对cd4+和cd8+t细胞活化(cd69上调;图31a-31b)以及细胞因子(il-6和tnf)释放(图32a-32b)的影响。如图中所示,在用xmab18087处理时cd4+和cd8+t细胞从血液中快速再分布(图30a和30b,相比于用xenp13245处理)且在给药之后随后恢复并常态化。在给药xmab18087时t细胞立即活化(相比于给药xenp13245)且随后消退同时t细胞再分布。另外,il-6和tnf细胞因子释放与t细胞活化相关(图32a-32b)。3e:在nsg小鼠中xmab18087的表征在另一研究剂量反应的研究中,在第7天时,对60只nsg小鼠移植1×106a549-redfluc肿瘤细胞(0.1ml体积皮下注射)。在第0天,根据总通量将小鼠分组并腹膜内移植10×106人类pbmc且实施指定浓度下的测试品的给药#1(每个浓度12只小鼠)。第8天和第15天时实施给药#2和#3。如上所述,通过每周两次至三次使用活体外成像系统测量每只小鼠总通量监测肿瘤生长,如图33中所描绘。此外,给药#1之后18天和22天时,通过如图x中所描绘地每周一次至两次用卡尺测量肿瘤体积。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1