由β-葡聚糖中产生龙胆二糖或葡萄糖的新型β-1,6-内切葡聚糖酶的制作方法
本发明涉及一种由β-葡聚糖产生龙胆二糖或葡萄糖的新型β-1,6-内切葡聚糖酶。
背景技术:
β-葡聚糖(β-glucan)是由通过糖苷键(glycosidicbond)连接的葡萄糖残基构成的多糖(polysaccharide),且是构成酵母、蘑菇等的细胞壁的极重要组分。β-葡聚糖已长期广泛用于例如抗氧化剂效果、抗癌功能、皮肤保护剂等的应用。
根据构成聚合物的键类型将β-葡聚糖划分成β-1,3-葡聚糖、β-1,4-葡聚糖以及β-1,6-葡聚糖,且已知β-1,6-葡聚糖相较于其它β-1,3-键或β-1,4-键以较小量存在于自然界中。衍生自疱脐衣属(lasalliapustulata)的石耳素是β-1,6-葡聚糖的熟知代表性实例,且昆布糖也熟知为构成棕色海藻的β-1,3-1,6-葡聚糖。
已知β-1,6-葡聚糖酶随机裂解β-葡聚糖的β-1,6-糖苷键(β-1,6-glycosidiclinkages),且根据碳水化合物活性酶数据库(carbohydrateactiveenzymesdatabase;cazy;http://www.cazy.org/),已知β-1,6-葡聚糖酶属于糖苷水解酶(glycosidehydrolase,gh)家族5和糖苷水解酶家族30。衍生自真菌的β-1,6-葡聚糖酶主要报道为β-1,6-葡聚糖酶,且到目前为止无细菌β-1,6-葡聚糖酶的报道。
技术实现要素:
技术问题
本发明的目标是提供一种能够由β-葡聚糖产生龙胆二糖或葡萄糖的新型β-1,6-内切葡聚糖酶的用途。
技术解决方案
为实现上述目标,本发明提供一种用于产生龙胆二糖或葡萄糖的组合物,所述组合物包含β-1,6-内切葡聚糖酶(β-1,6-endoglucanase)(包含seqidno:1的氨基酸序列),其中β-1,6-内切葡聚糖酶使用选自昆布糖(laminarin)和石耳素(pustulan)组成的组中的一或多个作为基质。
本发明还提供一种产生龙胆二糖或葡萄糖的方法,所述方法包含使包含seqidno:1的氨基酸序列的β-1,6-内切葡聚糖酶(β-1,6-endoglucanase)与作为基质的选自昆布糖(laminarin)和石耳素(pustulan)组成的组中的一或多个进行反应。
有利效果
本发明提供一种关于β-葡聚糖展现β-1,6-内切葡聚糖酶活性的β-1,6-内切葡聚糖酶。
β-1,6-内切葡聚糖酶可使用昆布糖或石耳素作为基质,由此提供高以产率产生龙胆二糖或葡萄糖的效果。
附图说明
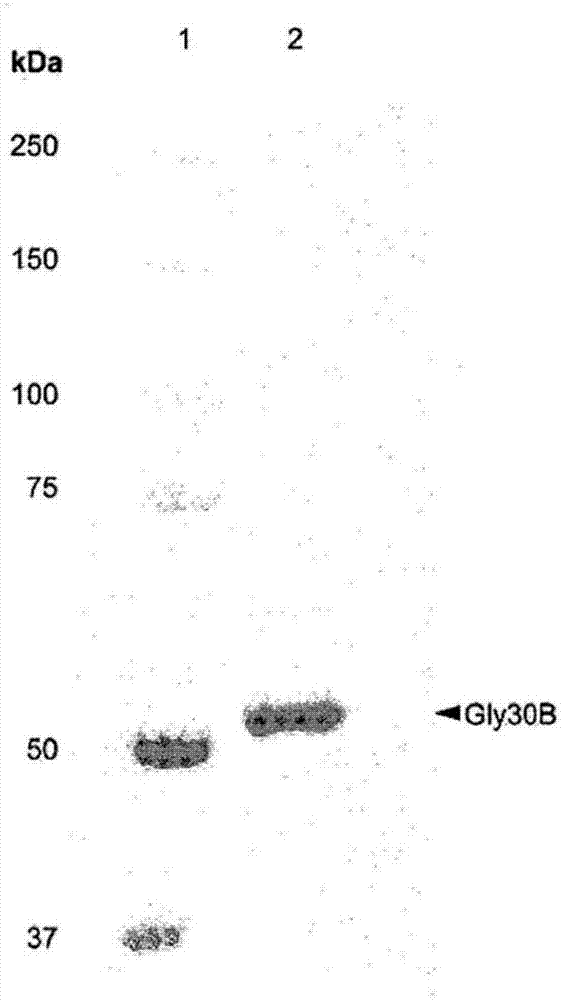
图1是确认本发明的β-1,6-内切葡聚糖酶的表达的凝胶电泳照片。
图2示出确认本发明的β-1,6-内切葡聚糖酶的最优活性ph值的结果。
图3示出确认本发明的β-1,6-内切葡聚糖酶的最优活性温度的结果。
图4示出确认本发明的β-1,6-内切葡聚糖酶的热稳定性的结果。
图5是本发明的β-1,6-内切葡聚糖酶关于石耳素的水解的双倒数绘图(lineweaver-burkplot)。
图6示出本发明的β-1,6-内切葡聚糖酶的水解产物关于石耳素的tlc(a)分析结果和hplc(b)分析结果。
图7示出本发明的β-1,6-内切葡聚糖酶的水解产物关于昆布糖的tlc(a)分析结果和hplc(b)分析结果。
图8是本发明的β-1,6-内切葡聚糖酶的系统发生树。
具体实施方式
本发明的发明人确认属于gh30家族的gly30b蛋白质的β-葡聚糖降解活性,假定其具有β-糖苷酶活性。因此,gly30b蛋白质使用昆布糖作为基质来裂解昆布糖的β-1,6-糖苷键,由此产生龙胆二糖或葡萄糖,且使用石耳素作为基质来裂解石耳素的β-1,6-糖苷键,由此产生龙胆二糖或葡萄糖,所述昆布糖是由β-1,3-连接的葡萄糖主链和β-1,6-连接的葡萄糖支链构成的多糖,所述石耳素是由β-1,6连接的葡萄糖构成的多糖。
因此,本发明提供一种用于产生龙胆二糖或葡萄糖的组合物,所述组合物包含具有seqidno:1的氨基酸序列的β-1,6-内切葡聚糖酶,其中β-1,6-内切葡聚糖酶使用选自昆布糖和石耳素组成的组中的一或多个作为基质。
本发明还提供一种龙胆二糖或葡萄糖的方法,所述方法包含使具有seqidno:1的氨基酸序列的β-1,6-内切葡聚糖酶与选自昆布糖和石耳素组成的组中的一或多个基质反应。
β-1,6-内切葡聚糖酶关于β-葡聚糖展现β-1,6-内切葡聚糖酶活性。
β-1,6-内切葡聚糖酶在范围介于约20℃到约45℃的温度下维持热稳定性,且关于昆布糖或石耳素展现最优降解活性。更具体地说,β-1,6-内切葡聚糖酶可在约20℃到约40℃的温度下展现最优活性。
另外,缓冲剂中的β-1,6-内切葡聚糖酶的最优ph值可根据缓冲剂的类型而变化,且可范围介于约4到约10,更具体地说约6到约8,且更具体地说约7。
β-1,6-内切葡聚糖酶可使用昆布糖、石耳素等作为基质。
酶的反应产物可以是聚合度为2的龙胆二糖或葡萄糖。
β-1,6-内切葡聚糖酶可衍生自变形菌株(saccharophagusdegradans)2-40t,但本发明不受其特定限制。
另外,β-1,6-内切葡聚糖酶可通过写码基因转录和转译,所述写码基因是与多肽的产生相关联的dna片段,包含酶的写码区的上游区域和下游区域以及个别写码片段之间的插入序列。举例来说,β-1,6内切葡聚糖酶可由seqidno:2的序列来转录和转译,但本发明不特别受限于此。另外,由一或多个取代、缺失、易位、添加等的前述酶不同且具有低聚糖或葡萄糖水解活性的突变蛋白也涵盖在本发明的酶的范围内,且其优选地具有与seqidno:1的氨基酸序列至少80%同源性、至少85%同源性、至少90%同源性、至少93%同源性、至少94%同源性、至少95%同源性、至少96%同源性、至少97%同源性、至少98%同源性以及至少99%同源性的氨基酸序列。
β-1,6-内切葡聚糖酶可从变形菌株2-40t培养液的上清液分离且纯化,且可使用基因重组技术产生且从除变形菌株2-40t以外的菌株分离或通过人工化学合成方法产生和分离。
当使用重组技术时,可使用用于有助于重组蛋白表达的典型因素,例如耐抗生素基因和可用于亲和性管柱色谱法的报道体蛋白或肽,且此技术属于本发明的范围内,其可由本发明涉及的一般技术人员中的任一个容易地进行。举例来说,β-1,6-内切葡聚糖酶可从转染为包含核酸的重组载体的宿主细胞或其培养产物获得,所述核酸编码β-1,6-内切葡聚糖酶,即seqidno:2的核苷酸序列。大肠杆菌也可用作宿主细胞,但本发明不限于此。
可在范围介于20℃到45℃的温度和5到10的ph下使β-1,6-内切葡聚糖酶与基质的反应进行5分钟到1天。更具体地说,当将昆布糖或石耳素用作基质时,可在范围介于30℃到40℃的温度和6到8的ph下使反应进行5分钟到5小时。
酶的降解产物可依序经历硅胶色谱(其是吸附色谱)和生物凝胶p2色谱(其是凝胶渗透色谱),由此分离和纯化具有约95%高纯度的低聚糖或葡萄糖。
如本文中所使用,术语“蛋白质”和“多肽”可互换地使用。
在本发明中,多肽具有关于另一序列的某些百分比(例如,80%、85%、90%、95%或99%)的序列一致性意味着当两个序列关于彼此比对且进行比较时,两个序列在氨基酸残基中具有所提及百分比的一致性。可使用所属领域中已知的任何适当软件程序,例如,[现代分子生物学实验技术(currentprotocolsinmolecularbiology)(f.m.奥斯贝(f.m.ausubel)等人(编)1987副刊30章节7.7.18)]中描述的一种软件程序来测定比对和同源性或一致性百分比。优选程序的实例包含:gcg叠加程序(gcgpileupprograms)、fasta(皮尔逊(pearson)等人,1988美国科学院院刊(proc.natlacad.sciusa)85:2444-2448)以及blast(blast手册(blastmanual),阿尔丘尔(altschul)等人,国立生物技术信息中心(natl.cent.biotechnol.inf.)、国立医学图书馆(natllib.med.)(ncibnlmnih),贝塞斯达、md以及阿尔丘尔等人,1997nar25:3389-3402)。另一优选比对程序是alignplus(科学和教育软件(scientificandeducationalsoftware),宾夕法尼亚州(pa)),优选地使用基本参数。又一合适的序列软件程序是序列软件包版本6.0(sequencesoftwarepackageversion6.0)(遗传学计算机组(geneticscomputergroup),威斯康星大学,麦迪逊(madison),威斯康星州(wi))中可获得的tfasta数据搜索程序(tfastadatasearchingprogram)。
在本发明中,结合细胞、核酸、蛋白或载体使用的术语“重组”意味着通过导入非均相核酸或蛋白,或更改固有核酸或蛋白来修饰细胞、核酸、蛋白或载体,或意味所述细胞衍生自这些修饰的细胞。也就是说,例如,重组细胞表达将不会在呈其原始(非重组)形式的细胞中发现的基因,或表达将非正常表达或将完全不表达的原始基因。
在本说明书中,术语“核酸”涵盖单股或双股dna、rna以及其化学变体的所有种类。术语“核酸”和“多核苷酸”可在本文中互换地使用。由于基因代码简并,一或多个密码子可用于编码某一氨基酸,且本发明涵盖编码某些氨基酸序列的多核苷酸。
用于描述将核酸序列插入到细胞中的术语“导入”是指“转染(transfection)”、“转化”或“转导(transduction)”,且涵盖对将核酸序列整合到真核或原核细胞中的提及。在这种情况下,将核酸序列整合到细胞的基因组(例如,染色体、质体、色素体或粒线体dna)中且将其转化为自发的复制子或按时序表达。
在下文中,将参考以下实例更详细地描述本发明。然而,这些实例并不意欲限制本发明的范围。
具体实施方式
<实例1>通过克隆获得gly30b基因
将衍生自变形菌株2-40t(atcc43961)的推定β-1,6-内切葡聚糖酶(sde_2994)克隆到大肠杆菌(escherichiacoli)dh5α中。更具体描述如下。
在30℃下将变形菌株2-40t(atcc43961)在含有23g/l即溶海水盐、50mmtris-hcl、2g/l葡萄糖、2g/l酵母提取物以及0.5g/l氯化铵的基本培养基中培养12小时。
使用可商购的dna分离试剂盒(美国加利福尼亚州凯杰巴伦西亚市(qiagen,valencia,ca,usa))获得变形菌株2-40t(atcc43961)的基因组dna。
使用solg2×taqpcr智能混合2(solgent,daejeon,韩国)放大靶基因gly30b(genbankidabd82251.1)。所使用引子如下:
正向引子:5'-gcgggatcccaccaccaccaccaccaccaatactggttaaccagcggtgatctaagt-3'(seqidno:3);以及
反向引子:5'-gcgctcgaggtggtggtggtggtggtgatctataactagcgttacaacgctctgtgc-3′(seqidno:4)。
这些引子在5'端处具有限制酶位点bamhi和限制酶位点xhoi。另外,为增加histrap管柱的亲和性,添加编码组胺酸的基因的核苷酸序列。
pcr产物和载体pet28a用bamhi和xhoi进行双重消化,且接合最终dna片段。将携带gly30b的质体转化到大肠杆菌dh5α。
<实例2>gly30b蛋白质的过度表达和纯化
为过度表达根据实例1获得的基因,将所述基因转化到大肠杆菌bl21(de3),其为用于蛋白质表达的宿主。
使用补充有50mg/l卡那霉素(kanamycin)的卢里亚-贝尔塔尼(luria-bertani;lb)培养液(美国马里兰州斯帕克斯bd(bd,sparks,md,usa))在37℃下培养细胞直到吸光率在600nm下达到0.6。使用0.1mmiptg诱导蛋白质表达且将诱导温度设定为16℃来以水溶性形式表达重组蛋白。
为分离所表达gly30b蛋白质,细胞通过超声波处理断裂且和离心,且接着使用histrap管柱(美国皮斯卡塔威市通用电气医疗集团(gehealthcare,piscataway,usa))来纯化上清液。用amiconultra离心滤波器(美国马萨诸塞州比尔里卡密理博公司)来浓缩纯化的蛋白质。通过sds-page来将所表达gly30b蛋白质的分子量测量为大约52kda(见图1)。使用二喹啉甲酸(bicinchoninicacid;bca)蛋白质分析试剂盒(美国伊利诺伊州罗克福德皮尔斯公司(pierce,rockford,il,usa))测量蛋白质的浓度。
<实例3>验证gly30b蛋白质的基质特异性和阳离子作用
为确认gly30b蛋白质的酶促活性,允许1.89nmgly30b蛋白质在40℃下以2%在含有例如石耳素、昆布糖(日本大阪和光株式会社(wako,osaka,japan))等的基质的100μl的20mmtris-hcl(ph6.0)中反应30分钟。另外,为确认gly30b蛋白质的基质特异性,允许10.5μmgly30b蛋白质在40℃下在含有例如石耳素、昆布糖、凝胶多糖(日本大阪和光株式会社)、羧甲基纤维素(美国密苏里州圣路易斯市西格玛-奥德里奇公司(sigma-aldrich,stlouis,mo,usa))、木聚糖(美国密苏里州圣路易斯市西格玛-奥德里奇公司)等的各种葡聚糖的100μl的20mmtris-hcl(ph6.0)中反应30分钟。通过dns方法来测量所产生的还原糖。
如下表1中所示出,gly30b蛋白质示出关于石耳素的最高活性,且当将昆布糖用作基质时,相较于当将石耳素单独用作基质时,所述gly30b蛋白质展现约22%的相对活性。确认gly30b蛋白质不水解凝胶多糖,且这确认gly30b酶选择性地裂解β-1,6-葡聚糖键。另外,发现gly30b蛋白质不与β-1,4-糖苷键(例如微晶纤维素)、cm-纤维素以及木聚糖反应。
由于检验gly30b蛋白质的阳离子作用,确认通过例如ni2+、cu2+、fe2+以及mg2+的阳离子来抑制gly30b蛋白质的反应性,且其中,通过cu2+抑制最多(参看表2)。
[表1]
gly30b蛋白质的基质特异性
[表2]
确认gly30b蛋白质的阳离子作用
<实例4>确认gly30b蛋白质的最优活性温度和ph
为找出针对gly30b蛋白质的活性的最优温度和ph,允许10.5μmgly30b蛋白质和2%(w/v)昆布糖的混合物在各种温度范围(20℃到70℃)和ph条件(2.0到10.0)中反应。
图2示出在范围介于2.0到10.0的ph下gly30b的相对活性。发现gly30b蛋白质在ph7.0下展现最大活性且酶促活性在小于或大于ph7.0的ph下快速降低。gly30b蛋白质在ph2.0(20mm甘氨酸-hcl缓冲剂)下展现约40%相对活性且在ph10.0(20mm甘氨酸-naoh缓冲剂)下展现约50%相对活性。
图3示出gly30b蛋白质在范围介于20℃到70℃的温度下的相对活性。gly30b蛋白质的酶促活性随着20℃到40℃的温度范围中的温度的增加而逐渐增加,在40℃下达到其最大值。gly30b蛋白质在20℃和60℃的反应温度下的酶促活性低于在40℃下观测到的酶促活性。由于检验gly30b蛋白质的热稳定性,确认gly30b蛋白质在40℃或小于40℃的反应温度下是稳定的且在更高温度下经历相对活性的显著减小(参看图4)。因此,40℃被测定为gly30b蛋白质的最佳反应温度且用于所有后续实验中。
<实例5>确认gly30b蛋白质的酶反应速率
为确认gly30b蛋白质关于石耳素和昆布糖的酶反应速率,允许gly30b蛋白质以范围介于0.45%到9.1%的各种浓度在ph7.0和40℃下与含有每种基质的20mmtris-hcl缓冲剂反应。
因此,从双倒数绘图(参看图5)确认,在石耳素基质的情况下,km值、vmax值以及kcat值分别为100.8g/l、32.8u/mg以及28.9s-1,且在昆布糖基质的情况下,km值、vmax值以及kcat值分别为24.2g/l、153.8u/mg以及135.6s-1。
<实例6>通过tlc和hplc特征化gly30b蛋白质的酶反应
为根据反应时间分析gly30b蛋白质的酶反应特征,使用薄层色谱法(thinlayerchromatography;tlc)和高效液相色谱法(highperformanceliquidchromatography;hplc)。
tlc分析的反应产物使用由正丁醇、乙酸以及水(3:2:2,v/v/v)组成的混合性溶剂系统产生于硅胶60培养盘(默克公司(merck))上,用10%(v/v)硫酸处理以用于观测,且接着在130℃下热处理5分钟。
使用配备有凝胶穿透和配体交换管柱(ks-802;昭和(shodex))的安捷伦(agilent)1100hplc(安捷伦)进行hplc分析,且使用折射率检测器(安捷伦)进行检测。使用无菌水作为溶剂以用于hplc分析,将流动速率设置为0.5ml/min,且将管柱温度设置为80℃。作为分析的标准材料,使用昆布二糖(laminaribiose)(聚合度(degreeofpolymerization):dp2)、昆布三糖(laminaritriose)(dp3)、昆布四糖(laminaritetraose)(dp4)、昆布五糖(laminaripentose)(dp5)以及昆布六糖(laminarihexose)(dp6)。
由于gly30b蛋白质与石耳素之间的反应的产物的tlc分析(参看图6a),确认龙胆二糖和葡萄糖在反应5分钟时开始产生且龙胆二糖随时间转化为葡萄糖。通过进行反应120分钟,确认葡萄糖作为主要反应产物且确认产生少量龙胆二糖。由于通过hplc的相同反应产物的分析(参看图6b),确认随着反应时间流逝,对应于龙胆二糖的峰减小,且对应于葡萄糖的峰产生。
在gly30b蛋白质与昆布糖之间的反应的情况下,确认不同于使用石耳素基质的情况,初始昆布糖和由其衍生的低聚糖未完全降解且即使在反应后仍存在(参看图7a),且产生葡萄糖和龙胆二糖(参看图7b)。根据这些结果,确认gly30b蛋白质选择性地分解β-1,6-键而非β-1,3-键。
<实例7>gly30b蛋白质的系统发生分析
为确认gly30b蛋白质的新颖性,从cazy数据库(http://www.cazy.org)中获得先前已知微生物衍生的β-1,6-内切葡聚糖酶的基因信息并绘制其系统发生树。
因此,确认gly30b蛋白质与其它先前已知β-1,6-内切葡聚糖酶系统发生学上分离,且gly30b蛋白质是先前从未报道的第一细菌衍生的酶(参看图8)。
工业实用性
本发明可应用于葡萄糖生产领域。
<110>高丽大学校产学协力团
<120>由β-葡聚糖中产生龙胆二糖或葡萄糖的新型β-1,6-内切葡聚糖酶
<130>x17u13c0108
<150>kr10-2016-0085868
<151>2016-07-07
<160>4
<170>kopatentin2.0
<210>1
<211>481
<212>prt
<213>变形菌2-40t(saccharophagusdegradans2-40t)
<400>1
metthrleuleulysasnileasnlysglnargleualaarglysval
151015
lysleuvalcysseralaileserleuvalvalmetglyphesercys
202530
alaalaseralaasnglntyrtrpleuthrserglyaspleuserala
354045
alapheglugluglnglyglulystyralavalalaproserproglu
505560
metproleuilethrileasplysserglnalapheglnthrmetglu
65707580
glypheglytyrthrleuasnglyglyseralathrhisleualaasn
859095
metseraspalaalaargalaargleuleuglngluilepheglygln
100105110
seraspglyalaasnthrasnlysproserileglyvalsertyrleu
115120125
argleuserleuglyalaseraspleuaspproalaprophesertyr
130135140
asnaspleuproproglygluvalaspleulysleuglulysphethr
145150155160
ilealaglnaspglulysthrleuileproileleulysglnileleu
165170175
alaileasnproasnilethrphemetalaserprotrpserpropro
180185190
valtrpmetlysthrasnglyserthrileglyglygluleuasnpro
195200205
glutyrtyrservaltyralaglntyrphevallystyrvalglnala
210215220
metalagluhisglyileasnileaspalailethrileglnasnglu
225230235240
promethisproglyasnasnproserleuleumethisalalysasp
245250255
glnalaasppheilealaasnhisleuglyproalaphelysglnala
260265270
gluleulysthrlysileilevaltrpasphisasnalaasplyspro
275280285
glutyrproilegluvalleuasnhisprovalalaasnglntyrile
290295300
hisglyseralaphehisleutyrglyglyaspvalasnalaileser
305310315320
glnvalhisasnalahisproasplyshisleutyrphethrglugln
325330335
trpvalglyalaasnserasnphetrpglyaspvalalatrphisval
340345350
gluasnleuilevalglyalathrargasntrpcyslysthrvalleu
355360365
glutrpasnleualaalaaspserasnleuglnprohisthrleugly
370375380
glycysaspalacysleuglyalaleuthrileaspglyaspasnval
385390395400
lysargasnalaalatyrtyrileilealahisalaalalyshisval
405410415
proproglyservalargilehisserhisargvalalaglyleupro
420425430
asnvalalapheleuthrproglnlyslysvalvalvalvalvalleu
435440445
asnasnthrthrglnleuglnserphethrleuvalhisasparggln
450455460
lysphealatyrsermetproalaglnservalvalthrleuvalile
465470475480
asp
<210>2
<211>1446
<212>dna
<213>变形菌2-40t(saccharophagusdegradans2-40t)
<400>2
atgacgttactaaaaaatataaataagcaacgcctagcacgtaaagttaaactcgtctgt60
agcgcgataagcctagtggttatggggttcagctgtgccgcgtcagccaatcaatactgg120
ttaaccagcggtgatctaagtgcggcgttcgaagagcagggcgaaaagtacgcggtagct180
cccagccccgaaatgccccttattaccatagataaaagccaagcatttcaaaccatggaa240
ggctttggctatacccttaatggcggcagtgcaactcacctagccaatatgagtgacgca300
gctagagcgcggctattacaagaaatatttggtcaaagtgatggtgcgaatactaataag360
cccagtattggcgtgtcttatttgcgtttaagcctaggcgcatcagatttagaccccgcc420
ccgtttagctacaacgatttgccgcctggcgaagtggatttaaagctagaaaaatttact480
atcgcccaagatgaaaaaactcttatccccatacttaagcaaatattagctattaaccca540
aatattacatttatggctagcccttggtctccgcctgtatggatgaaaacaaacggctct600
accattggtggtgagctaaacccagaatactacagcgtatatgcacaatattttgttaaa660
tatgttcaagcaatggctgagcacggcataaacatagatgccattactattcaaaatgaa720
cctatgcacccgggtaataacccaagcttgctcatgcatgcaaaagatcaagccgacttt780
attgctaatcacttaggccccgcgtttaagcaggcagagctaaaaacaaaaatcattgtg840
tgggatcacaacgcagacaaacccgaataccccatagaggtactgaatcaccccgttgcc900
aatcaatatattcacggctcggcattccatttatatggcggcgatgtaaatgccataagc960
caagtgcacaatgctcacccagataagcacttatattttactgagcagtgggtaggcgca1020
aattccaacttttggggcgatgtagcttggcatgtagaaaatttaattgttggggcaacc1080
cgcaattggtgcaaaacggtattggagtggaatttagccgcagacagtaacttacagcct1140
cacactcttggtggatgcgacgcctgcttaggcgcgctaactattgatggcgataacgtg1200
aagcgcaatgccgcgtattacattattgcccacgcagctaaacatgtaccgccaggctcg1260
gtgcgtatccattcgcaccgtgtggcgggtttacctaatgttgcttttcttacgccgcag1320
aaaaaggttgttgtagtagtgcttaataatactactcaattacagtcatttacattggtg1380
cacgaccgtcaaaagtttgcctattccatgcccgcacagagcgttgtaacgctagttata1440
gattaa1446
<210>3
<211>57
<212>dna
<213>人工序列(artificialsequence)
<220>
<223>顺向引物
<400>3
gcgggatcccaccaccaccaccaccaccaatactggttaaccagcggtgatctaagt57
<210>4
<211>57
<212>dna
<213>人工序列(artificialsequence)
<220>
<223>反向引物
<400>4
gcgctcgaggtggtggtggtggtggtgatctataactagcgttacaacgctctgtgc57
- 还没有人留言评论。精彩留言会获得点赞!
- 一种含葡糖酰胺基的三硅氧烷的制备方法
- 以氨基葡聚糖为还原剂和保护剂制备胶态金属分散体的制作方法
- 葡糖胺基聚糖(gag)模拟物的制作方法
- 1-(β-D-吡喃葡糖基)-4-甲基-3-[5-(4-氟苯基)-2-噻吩基甲基]苯的结晶方法
- 1-(β-D-吡喃葡糖基)-4-甲基-3-[5-(4-氟苯基)-2-噻吩基甲基]苯的晶型B及其制备方法
- 天然产物葡聚五糖的高效合成方法
- 1-(β-D-吡喃葡糖基)-4-甲基-3-[5-(4-氟苯基)-2-噻吩基甲基]苯的新晶型B及其制备方法
- 1-(β-D-吡喃葡糖基)-4-甲基-3-[5-(4-氟苯基)-2-噻吩基甲基]苯与L-苯丙氨酸共晶物 ...的制作方法
- 解聚海参糖胺聚糖在制药中的应用的制作方法
- 1→6连接的氨基葡聚寡糖和糖苷及其合成和应用的制作方法