一种人源分子改造杀虫蛋白、编码基因及其设计方法和应用与流程
本发明涉及基因工程和生物防治领域,具体涉及一种人源分子改造杀虫蛋白、编码基因及其设计方法与应用。
背景技术
苏云金芽孢杆菌(bacillusthuringiensis,bt)是一种昆虫病原菌,其主要杀虫活性物质为细胞内毒素伴胞晶体蛋白,对多种农业害虫具有特异性毒杀作用(bravoandsoberon,2008);cry1ac是bt毒素的一种,其对鳞翅目昆虫的靶标受体主要包括中肠刷状缘膜囊泡(brushbordermembranevesicles,bbmv)上的碱性磷酸酶(alkalinephosphatase,alp)、氨肽酶(aminopeptidasen,apn)和钙粘蛋白(cadherin)。alp作为cry1a毒素的受体,可促进毒素对膜的插入和孔道形成。诸多从双翅目和鳞翅目昆虫物种中分离出的alp已被鉴定为cry1ac毒素的受体。抗cry1ac毒素独特型单链抗体(anti-idiotypeantibodyscfvs,anti-idscfvs),可以模拟cry1ac毒素的结构与功能。
bt毒素使用量增加使其在害虫抗药性及次生害虫上升等方面存在生态风险(bravoandsoberon,2008;pigottetal,2008),这一现状促使高特异活性和新功能类的bt毒素抗虫资源的积极开发,cry1ab毒素抗独特型单链抗体由于可模拟cry1ab毒素并与其竞争受体蛋白,已成为目前国内外科研的重点,专利号为zl201410037000.9的中国专利公开了一种人源抗虫基因及其编码的抗cry1ab毒素独特型单链抗体b12(以下简称b12),其能模拟bt毒素并用作生物农药防治虫害,取得了积极的效果,但是该抗体的可溶性表达蛋白丧失了与昆虫bbmv结合能力,杀虫效果不甚理想。
基因工程(geneticengineering)又称基因拼接技术和dna重组技术,是以分子遗传学为理论基础,以分子生物学和微生物学的现代方法为手段。将不同来源的基因按预先设计的蓝图,通过对dna分子进行人工“剪切”和“拼接”,对生物的基因进行改造和重新组合,在体外构建重组dna分子,然后导入活细胞,产生出人类所需要的基因产物。如demaagd于2002年针对cry1ab毒素在替换上cry1c毒素的domainiii后,对甜菜夜蛾的毒性较cry1c提高了近10倍。
随着对抗体结构与功能关系的认知不断加深,借助计算机模拟技术进行抗体改造可以在限定范围内有目的地进行设计,根据抗体-抗原结合的氨基酸位点分析进行设计与定向改造,wong等(1995)在已知抗原-抗体复合物三维结构的基础上,对抗苯偶氮基胂酸酯(anti-p-azophenylarsonate)fab重链108位上的phe定点突变为trp之后,相对野生抗体而言,突变体亲和力就提高了10倍;为进一步对抗体分子结合区域改造指明了方向,但目前结合链替换分子改造方法来提高抗bt毒素抗独特型单链抗体亲和力的方法并通过octet技术平台筛选的方法尚未见报道。
技术实现要素:
针对上述问题,本发明提供了一种人源分子改造杀虫蛋白、编码基因及其设计方法和应用,所述人源分子改造杀虫蛋白与小菜蛾(plutellaxylostella)中肠bbmv的亲和力比cry1ab、cry1b毒素高,且与cry1ab、cry1ac和cry1b毒素竞争结合小菜蛾中肠bbmv,为cry1ab、cry1ac和cry1b毒素的模拟物。可有效替代cry1a类毒素用于生物防治虫害,且具有杀虫效果。
本发明提供了一种人源分子改造杀虫蛋白ggcc_scfv,所述杀虫蛋白ggcc_scfv的氨基酸序列如seqidno.1所示。
本发明提供了上述人源分子改造杀虫蛋白ggcc_scfv的设计方法,包括如下步骤:
1)对抗cry1ab毒素独特型单链抗体b12_scfv和cry1a类毒素的氨基酸序列分别进行blast比对分析,得到b12_scfv与cry1a类毒素的相似序列,分别为h-cdr1,l-cdr1,l-cdr2和gs-linker;
2)对抗cry1ab毒素独特型单克隆抗体g1_scfv和cry1b类毒素的氨基酸序列进行blast比对分析,得到g1_scfv与cry1b类毒素的相似序列,分别为h-cdr1’,h-cdr3’和l-cdr2’;
3)将g1_scfv中包含h-cdr1’和h-cdr3’的重链区、gs-linker和b12_scfv中包含l-cdr1和l-cdr2序列的轻链区顺次连接,得到人源分子改造杀虫蛋白ggcc_scfv;所述g1_scfv中包含h-cdr1’和h-cdr3’的重链区的氨基酸序列如seqidno.2所示;所述b12_scfv中包含l-cdr1和l-cdr2序列的轻链区的氨基酸序列如seqidno.3所示;所述gs-linker的氨基酸序列如seqidno.4所示;
所述步骤1)和步骤2)之间没有先后顺序的限定。
优选的,所述g1_scfv的氨基酸序列如seqidno.5所示。
本发明提供了一种杀虫剂,包括上述的人源分子改造杀虫蛋白ggcc_scfv。
本发明提供了一种人源分子改造杀虫蛋白ggcc_scfv的编码基因,所述编码基因的核苷酸序列如seqidno.6所示。
本发明提供了一种含有上述编码基因的重组载体。
本发明提供了一种杀虫重组菌,所述杀虫重组菌含有上述重组载体或编码基因。
本发明还提供了上述人源分子改造杀虫蛋白、杀虫剂、编码基因、重组载体或杀虫重组菌在农作物害虫防治中的应用。
优选的,所述农作物害虫包括小菜蛾。
优选的,所述农作物包括十字花科农作物。
有益效果:
(1)本发明提供了一种人源分子改造杀虫蛋白ggcc_scfv,所述杀虫蛋白ggcc_scfv的氨基酸序列如seqidno.1所示。本发明提供的人源分子改造杀虫蛋白与小菜蛾bbmv的亲和力显著高于b12_scfv、g1_scfv和cry1ab、cry1b毒素;其可与cry1ab、cry1ac和cry1b毒素竞争结合小菜蛾的bbmv,且与cry1b的竞争性更强,为cry1ab、cry1ac和cry1b毒素的模拟物。经小菜蛾室内杀虫生物活性测定,3d致死率可达50%,7d致死率可达95%。可有效的替代cry1a类毒素用于生物防治虫害。
同时,本发明提供的人源分子改造杀虫蛋白ggcc_scfv由b12_scfv基因分子和g1_scfv基因分子改造拼接所获得,抗体骨架区未变,与b12_scfv和g1_scfv同属于人源抗体,因此ggcc_scfv应用于农业虫害防治时,对人体危害小。
(2)本发明提供了上述人源分子改造杀虫蛋白ggcc_scfv的设计方法,对抗cry1ab毒素独特型单链抗体b12_scfv和cry1a类毒素的氨基酸序列进行blast比对分析,得到b12_scfv与cry1a类毒素的相似序列,分别为h-cdr1,l-cdr1,l-cdr2和gs-linker;对抗cry1ab毒素独特型单克隆抗体g1_scfv和cry1b类毒素的氨基酸序列进行blast比对分析,得到g1_scfv与cry1b类毒素的相似序列,分别为h-cdr1’,h-cdr3’,l-cdr2’;将g1_scfv中包含h-cdr1’和h-cdr3’的重链区、gs-linker和b12_scfv中包含l-cdr1和l-cdr2序列的轻链区顺次连接,得到人源分子改造杀虫蛋白ggcc_scfv。本发明提供的方法与传统抗体改造方法(易错pcr等)相比,避免了盲目性,成功率高;且以计算机分子模拟为基础,在已知蛋白三维结构及功能等方面信息的基础上,预测分子间结合区域并确定关键氨基酸残基,更加准确的反应了抗体蛋白作用界面的特征,从而使得分子模拟计算更加准确,大大提高了预测准确度。
附图说明
图1为本发明所述人源分子改造杀虫蛋白ggcc_scfv的设计图;
图2为本发明实施例3中b12_scfv、g1_scfv、改造后的ggcc_scfv、ccgg_scfv和bt毒素cry1ab的三维结构示意图;其中,图2-a为b12_scfv的三维结构示意图(选取scfvmab198的1f3r.1.b作为模板);图2-b为g1_scfv的三维结构示意图(选取sp140phd-bromodomain复合物中结合的scfv6g8r.1.a作为模板);图2-c为ggcc_scfv的三维结构示意图(选选取fabbl3-64q9r.1.a作为模板);图2-d为ccgg_scfv的三维结构示意图(选取sp140phd-bromodomain复合物中结合的scfv6g8r.1.a作为模板);图2-e为cry1ab的三维结构示意图(选取cryia(a)1ciy作为模板);图2-f为pxalp的三维结构图(选取人alp1zef.1.a作为模板);
图3为本发明实施例3中b12_scfv、g1_scfv、改造后的ggcc_scfv、ccgg_scfv、bt毒素cry1ab和pxalp进行分子对接时的三维结构模型示意图;其中,图3-a为b12_scfv的三维结构图;图3-b为g1_scfv的三维结构示意图;图3-c为ggcc_scfv的三维结构示意图;图3-d为ccgg_scfv的三维结构示意图;图3-e为cry1ab的三维结构示意图;图3-f为pxalp的三维结构图;图3-g为b12_scfv与pxalp的分子对接三维结构图;图3-h为g1_scfv与pxalp的分子对接三维结构图;图3-i为ggcc_scfv与pxalp的分子对接三维结构图;图3-j为ccgg_scfv与pxalp的分子对接三维结构图;图3-k为cry1ab与pxalp的分子对接三维结构图。
图4为本发明实施例3中b12_scfv、g1_scfv、改造后的ggcc_scfv、ccgg_scfv、bt毒素cry1ab分别与pxalp受体结合后的复合物中结合热点分析;其中,图4-a、4-b、4-c、4-d和4-e分别为cry1ab、b12_scfv、g1_scfv、ggcc_scfv、ccgg_scfv与alp受体结合区域i结合热点的三维结构图,在其右侧为配体的主要结合结构域;图4-f、4-g、4-h、4-i和4-j分别为cry1ab、b12_scfv、g1_scfv、ggcc_scfv、ccgg_scfv与alp受体结合区域ii结合热点的三维结构图,在其左侧为配体的主要结合结构域;
图5为本发明实施例4中采用生物膜层表面干涉技术(bio-layerinterferometry,bli)fortebiooctet分子相互作用技术平台及酶联免疫法(elisa)以小菜蛾中肠刷状缘膜囊泡蛋白bbmv为靶抗原,对b12_scfv、g1_scfv、ggcc_scfv、ccgg_scfv和bt毒素cry1ab、cry1b的结合活性测定及筛选结果;其中,图5-a为生物膜层表面干涉技术(bio-layerinterferometry,bli)fortebiooctet分子相互作用技术平台对杀虫蛋白活结合活性的测定结果;图5-b为酶联免疫法(elisa)对杀虫蛋白的筛选结果;
图6为本发明实施例4中采用生物膜层表面干涉技术(bio-layerinterferometry,bli)fortebiooctet分子相互作用技术平台,以小菜蛾中肠刷状缘膜囊泡蛋白bbmv为靶抗原,对ggcc_scfv、cry1b和cry1ab亲和力常数的测定结果;其中,图6-a为不同稀释浓度下ggcc_scfv与小菜蛾bbmv的结合曲线,图6-b为不同稀释浓度下bt毒素cry1ab与小菜蛾bbmv的结合曲线,图6-c为不同稀释浓度下bt毒素cry1b与小菜蛾bbmv的结合曲线;图6-d为亲和力常数;
图7为本发明实施例5中以2龄小菜蛾为试虫,利用表达纯化的b12_scfv、g1_scfv、ggcc_scfv、ccgg_scfv和bt毒素cry1ab进行生物活性测定的结果。
图8为本发明实施例6中采用竞争性酶联免疫法(elisa),以小菜蛾中肠刷状缘膜囊泡蛋白bbmv为靶抗原,ggcc_scfv与bt毒素cry1ab、cry1ac和cry1b做竞争性抑制实验的结果;图8-a为ggcc_scfv与bt毒素cry1ab、cry1ac和cry1b竞争结合小菜蛾bbmv的elisa吸光值变化图,图8-b为ggcc_scfv与bt毒素cry1ab、cry1ac和cry1b竞争结合小菜蛾bbmv的抑制率;
具体实施方式
本发明提供了一种人源分子改造杀虫蛋白ggcc_scfv,所述杀虫蛋白ggcc_scfv的氨基酸序列如seqidno.1所示。
在本发明中,所述人源分子改造杀虫蛋白ggcc_scfv由人源抗cry1ab毒素独特型单链抗体b12_phage和g1_phage通过基因分子改造拼接获得。所述单链抗体b12_phage的来源见专利号为zl201410037000.9,专利名称为“一种人源抗虫基因及其编码的抗cry1ab毒素独特型单链抗体与应用”的专利。本发明所述b12_phage为噬菌体展示型scfv,scfv与噬菌体giii蛋白融合表达;b12_scfv为可溶性表达的scfv,不含噬菌体展示外壳。b12_phage中scfv部分的(核酸/蛋白质)序列与b12_scfv的序列完全一致。所述单链抗体b12_scfv的氨基酸序列已由专利zl201410037000.9所公开。所述g1_pahge通过人源噬菌体抗体库(tomlison(i+j)库,从英国剑桥大学引进)筛选获得,g1_scfv为可溶性表达的scfv,不含噬菌体展示外壳,g1_phage中scfv部分的(核酸/蛋白质)序列与g1_scfv的序列完全一致;所述筛选的方法参见专利zl201410037000.9。所述g1_scfv的氨基酸序列如seqidno.5所示。
在本发明中,所述人源分子改造杀虫蛋白ggcc_scfv的设计方法优选包括如下步骤:
1)对抗cry1ab毒素独特型单链抗体b12_scfv和cry1a类毒素的氨基酸序列进行blast比对分析,得到b12_scfv与cry1a类毒素的相似序列,分别为h-cdr1,l-cdr1,l-cdr2和gs-linker;
2)对抗cry1ab毒素独特型单克隆抗体g1_scfv和cry1b类毒素的氨基酸序列进行blast比对分析,得到g1_scfv与cry1b类毒素的相似序列,分别为h-cdr1’,h-cdr3’,l-cdr2’;
3)将g1_scfv中包含h-cdr1’和h-cdr3’的重链区、gs-linker和b12_scfv中包含l-cdr1和l-cdr2序列的轻链区顺次连接,得到人源分子改造杀虫蛋白ggcc_scfv;所述g1_scfv中包含h-cdr1’和h-cdr3’的重链区的氨基酸序列如seqidno.2所示;所述b12_scfv中包含l-cdr1和l-cdr2序列的轻链区的氨基酸序列如seqidno.3所示;所述gs-linker的氨基酸序列如seqidno.4所示;
所述步骤1)和步骤2)之间没有先后顺序的限定。
本发明提供的人源分子改造杀虫蛋白ggcc_scfv为cry1ab、cry1ac和cry1b毒素的模拟物,其可与cry1ab、cry1ac和cry1b毒素竞争结合小菜蛾的bbmv受体位点,所述人源分子改造杀虫蛋白ggcc_scfv与小菜蛾bbmv受体位点的亲和力显著高于b12_scfv、g1_scfv和cry1ab、cry1b毒素。
本发明提供了上述人源分子改造杀虫蛋白ggcc_scfv的编码基因。在本发明中,所述ggcc_scfv的编码基因优选由人源抗cry1ab毒素独特型单链抗体b12_phage和g1_phage基因序列经分子改造拼接设计,由南京锐真生物有限公司人工合成获得。更优选的,所述ggcc_scfv编码基因的核苷酸序列如seqidno.6所示。
在本发明中,所述人源分子改造杀虫蛋白ggcc_scfv的制备方法优选采用重组表达法制备。所述重组表达法制备人源分子改造杀虫蛋白的方法优选包括以下步骤:
(1)将编码人源分子改造杀虫蛋白ggcc_scfv的基因与载体分别进行双酶切,连接,获得重组载体;
(2)将所述重组载体导入表达系统内,诱导培养,得到重组表达的人源分子改造杀虫蛋白ggcc_scfv。
在本发明中,所述表达系统优选为原核细胞表达系统。所述原核表达系统优选为大肠杆菌。所述大肠杆菌的菌株优选为大肠杆菌bl21(de3)。所述载体优选包括pet-26b、pet28a,更优选为pet-26b。所述双酶切用酶优选为ncoi酶和noti酶。双酶切的体系优选为ncoi酶和noti酶各1ul,载体43ul,10×cutsmart缓冲液5ul,37度30min。t4连接体系为双酶切载体1ul,dna片段7ul,t4连接酶1ul,10×t4连接酶反应液1ul。在本发明的实施例中,所述重组载体的构建方法为:在人源分子改造杀虫蛋白ggcc_scfv的核酸序列n端添加ncoi酶切位点,c端添加noti酶切位点,合成基因后经双酶切转入pet-26b载体中。
本发明提供了一种杀虫剂,所述杀虫剂包括上述的人源分子改造杀虫蛋白ggcc_scfv。
在本发明中,所述人源分子改造杀虫蛋白ggcc_scfv在所述杀虫剂中的质量含量优选为0.1~99%,更优选为1~50%。
本发明对所述杀虫剂的制备方法不作特别限定,采用本领域常规方法,以人源分子改造杀虫蛋白ggcc_scfv配合本领域可接受的载体制得即可。
在本发明中,所述杀虫剂在使用时优选采用浸叶法或利用饲料浸泡晾干后喂虫。
本发明还提供了上述人源分子改造杀虫蛋白ggcc_scfv、杀虫剂、基因、重组载体或重组菌在农作物害虫防治方面的应用。
在本发明中,所述在农作物害虫防治方面的应用包括一切以防治农作物害虫为目的的制备本发明所述杀虫蛋白的方法、制备本发明所述杀虫蛋白编码基因的方法、制备本发明所述杀虫重组菌的方法、制备本发明所述杀虫剂的方法、将本发明所述杀虫蛋白、杀虫剂、杀虫重组菌施加到农作物种植区域的方法。
在本发明中,所述农作物害虫优选包括小菜蛾,所述农作物优选包括十字花科农作物,更优选为小白菜(青菜)。
本发明以b12_scfv基因分子和g1_scfv基因分子改造拼接获得的人源分子改造杀虫蛋白ggcc_scfv,其抗体骨架区未变,与b12_scfv和g1_scfv同属于人源抗体。因此,将人源分子改造杀虫蛋白ggcc_scfv应用于农业虫害防治时,对人体危害小。
制得人源分子改造杀虫蛋白ggcc_scfv后,本发明将b12_scfv、g1_scfv、ggcc_scfv、ccgg_scfv和bt毒素cry1ab蛋白用pbs稀释至制定浓度,各取500μl铺于培养皿中饲料表面,晾干;阴性对照为pbs缓冲液,阳性对照为bt毒素cry1ab蛋白。
每个饲料培皿中接入30头已饥饿4h的小菜蛾2龄幼虫,然后置于温度为25±1℃,相对湿度80±5%,光照条件大于14h的培养箱内饲养,3d、7d后分别观察、记录死亡虫数,取出幼虫检查时以小毛笔轻触虫体,无明显反应者为死亡。每处理3次重复。试虫死亡率采用abbott公式对死亡率进行校正,并以平均数±标准误表示(3次重复试验)。
校正死亡率=(处理死亡率-对照死亡率)/(1-对照死亡率)×100%
各处理时间相同时各个样品之间的比较采用单因素方差分析(one-wayanova)和tukey显著性检验,使用spss软件进行数据处理,处理结果表明:3d后,ggcc_scfv对小菜蛾的校正致死率为50%,b12_scfv的校正致死率低于10%,3d后ggcc_scfv较b12_scfv的校正致死率提高了近5倍;7d后ggcc_scfv对小菜蛾的校正致死率为95%,b12_scfv的校正致死率低于40%,ggcc_scfv与cry1ab的校正致死率接近。该结果证明本发明提供的人源分子改造杀虫蛋白可有效的替代cry1ab毒素用于生物防治虫害。
下面结合具体实施例对本发明所述的一种人源分子改造杀虫蛋白ggcc_scfv、编码基因及其设计方法和应用做进一步详细的介绍,本发明的技术方案包括但不限于以下实施例。
实施例中所涉及的实验材料:
载体pet26b载体购于德国novagene公司;ncoi、noti酶、t4dna连接酶均购于neb公司;
pcr试剂购于全式金公司;
质粒提取试剂盒、pcr产物纯化试剂盒、pcr产物凝胶回收试剂盒购于axygen公司;
大肠杆菌bl21(de3)感受态细胞购于全式金公司;
hrpconjugatedanti-histagmousemonoclonalantibodyandhrpconjugategoatanti-rabbitigg购于ge公司;
cry1b、cry1ac、cry1ab毒素购于上海佑隆生物科技有限公司;
96孔板(costar3599)购于美国corning公司;
实施例1
一种人源分子改造杀虫蛋白的设计及制备方法,具体步骤如下:
(1)对抗cry1ab毒素独特型单链抗体b12_scfv和cry1a类毒素(cry1aa、cry1ab、cry1ac)的氨基酸序列进行blast比对分析,分析b12与cry1a类毒素的相似序列,分别为h-cdr1,l-cdr1,l-cdr2和gs-linker。对抗cry1ab毒素独特型单克隆抗体g1_scfv和cry1b类毒素的氨基酸序列进行blast比对分析,分析g1与cry1b类毒素的相似序列,分别为h-cdr1’,h-cdr3’,l-cdr2’。
(2)根据结果设计拼接,以抗cry1ab毒素独特型单链抗体b12_scfv序列的重链区(vh)利用g1_scfv的重链区(vh)进行替换,获得重组抗体ggcc_scfv;或以抗cry1ab毒素独特型单链抗体b12_scfv序列的轻链区(vl)利用g1_scfv的轻链区(vl)进行替换,获得重组抗体ccgg_scfv,并再进行blast比对分析。
(3)对抗cry1ab毒素独特型单链抗体b12_scfv和g1_scfv、人源分子改造蛋白ggcc_scfv和ccgg_scfv的氨基酸序列、cry1ab毒素和小菜蛾中肠bbmv上的alp分别同源建模:抗cry1ab毒素独特型单链抗体b12_scfv选取scfvmab198的1f3r.1.b作为同源建模的模板,g1_scfv选取sp140phd-bromodomain复合物中结合的scfv6g8r.1.a作为同源建模的模板,人源分子改造蛋白ggcc_scfv选取fabbl3-64q9r.1.a作为同源建模的模板,人源分子改造蛋白ccgg_scfv选取sp140phd-bromodomain复合物中结合的scfv6g8r.1.a作为同源建模的模板,cry1ab毒素选择cryia(a)1ciy作为同源建模的模板,pxalp选择人alp1zef.1.a作为同源建模的模板。
(4)利用zdock程序对抗cry1ab毒素独特型单链抗体b12_scfv和g1_scfv、ggcc_scfv和ccgg_scfv以及cry1ab毒素和小菜蛾中肠bbmv上的alp的同源建模进行分子对接,将获得的对接复合物提交至mitchelllab的热点预测服务器kfc2,定义b12_scfv、g1_scfv、ggcc_scfv和ccgg_scfv以及cry1ab毒素为配体,针对与cry1ab相同pxalp结合区域的配体热点进行分析。
(5)ggcc_scfv和ccgg_scfv核酸序列通过n端添加ncoi和c端添加noti酶切位点并基因合成获得,转入pet-26b载体中,利用大肠杆菌bl21(de3)进行表达。目的蛋白经ge的his-trap亲和柱纯化。
(6)利用生物膜层表面干涉技术(bio-layerinterferometry,bli)fortebiooctet分子相互作用技术平台及酶联免疫法(elisa),以小菜蛾中肠刷状缘膜囊泡蛋白bbmv为靶抗原,筛选出高bbmv结合活性的人源分子改造蛋白ggcc_scfv,并测定亲和力常数。
(7)利用酶联免疫法(elisa),以小菜蛾中肠刷状缘膜囊泡蛋白bbmv为靶抗原,人源分子改造杀虫蛋白ggcc_scfv与cry1ab、cry1ac和cry1b做竞争性抑制,以验证ggcc_scfv模拟cry1a类毒素的特性。
实施例2
抗crylab毒素独特型单链抗体b12_scfv、g1_scfv、改造后的ggcc_scfv、ccgg_scfv和cry1a类毒素(cry1aa、cry1ab、cry1ac)与cry1b毒素的氨基酸序列进行blast比对分析:
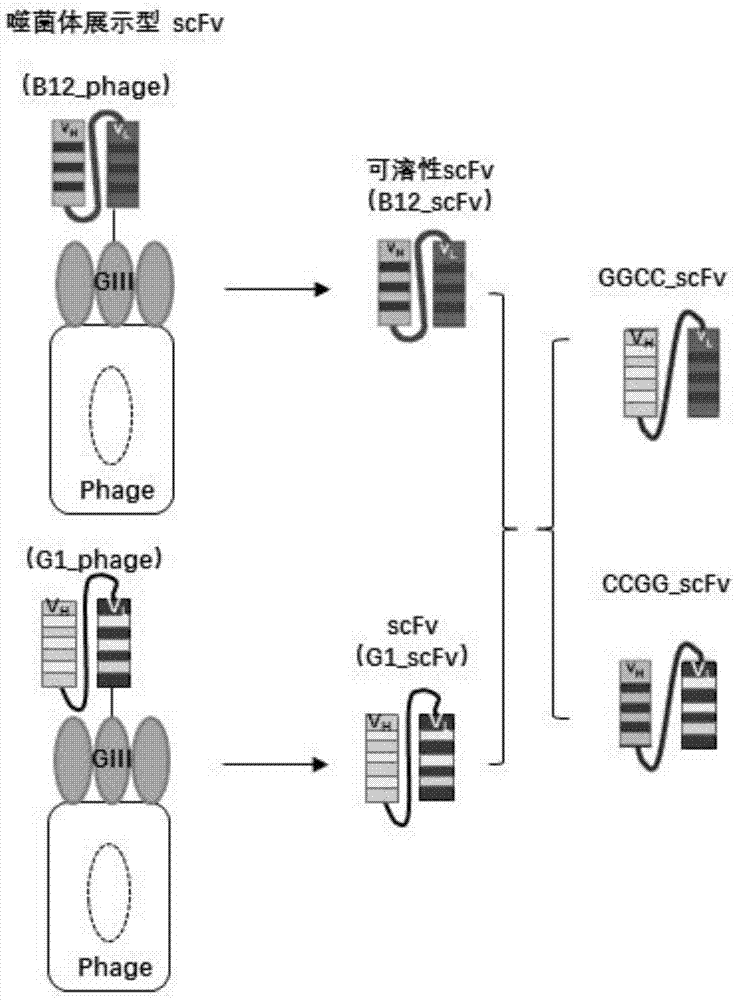
抗crylab毒素独特型单链抗体b12(以下简称b12_phage,来源于噬菌体载体pit2,为噬菌体展示型单链抗体(scfv),即在噬菌体外壳的giii蛋白上共表达的scfv,如图1左侧上方所示)的氨基酸序列已由专利号为zl201410037000.9的中国专利公开,b12_scfv为b12_phage中的抗体序列,重组到原核表达载体pet-26b中,为可溶性表达scfv,如图1中间上方所示。
抗crylab毒素独特型单链抗体g1(以下简称g1_phage,来源于噬菌体载体pit2,为噬菌体展示型单链抗体(scfv),即在噬菌体外壳的giii蛋白上共表达的scfv,如图1左侧下方所示),为本实验室通过人源噬菌体抗体库(tomlison(i+j)库)筛选获得。g1_scfv为g1_phage中的抗体序列,重组到原核表达载体pet-26b中,为可溶性表达scfv,如图1中间下方所示。
bt毒素cry1aa、cry1ab和cry1ac氨基酸序列由ncbi蛋白质数据库获得,登录号为:afk79795.1、alj10947.1和alt07695.1。bt毒素cry1b的氨基酸序列由ncbi蛋白质数据库获得,登录号为:abl60921.1。
b12_scfv和g1_scfv氨基酸序列提交到blast网站上(https://blast.ncbi.nlm.nih.gov/blast.cgi)分别与cry1aa、cry1ab和cry1ac以及cry1b的氨基酸序列进行比对,比对结果如表1所示。
b12_scfv的h-cdr1区与cry1a类、cry1b蛋白的保守结构域endotoxin_n端结构域有相似序列,endotoxin_n端结构域主要是bt杀虫毒素的n端结构,n-端螺旋域涉及到膜插入和孔隙的形成,推测b12_scfv的h-cdr1区有可能具有相似功能。b12_scfv的l-cdr1和l-cdr2区与cry1a类蛋白的保守结构域endotoxin_c端结构域有相似序列,endotoxin_c端结构域主要是bt杀虫毒素的c端结构,主要与受体结合相关。b12_scfv的h-cdr3与cry1b蛋白的保守结构域endotoxin_m端结构域有相似序列,endotoxin_m端结构域主要是bt杀虫毒素的中间结构,主要具有可以特异地结合昆虫肠上皮细胞受体的功能,推测b12_scfv的h-cdr3区有可能具有相似功能。由此,本发明推测b12_scfv包含h-cdr1和h-cdr3的重链区为杀虫活性区,b12_scfv包含l-cdr1和l-cdr2的轻链区为结合活性区。g1_scfv的h-cdr1’和l-cdr2’与cry1b蛋白的保守结构域endotoxin_n端结构域有相似序列,endotoxin_n端结构域主要是bt杀虫毒素的n端结构,n-端螺旋域涉及到膜插入和孔隙的形成。g1_scfv的l-cdr1’与cry1a类蛋白的保守结构域endotoxin_c端结构域有相似序列,endotoxin_c端结构域主要是bt杀虫毒素的c端结构,主要与受体结合相关。g1_scfv的h-cdr3’与cry1b蛋白的保守结构域endotoxin_m端结构域有相似序列,endotoxin_m端结构域主要是bt杀虫毒素的中间结构,主要具有可以特异地结合昆虫肠上皮细胞受体的功能,推测g1_scfv的h-cdr3’区有可能具有相似功能。由此,本发明推测g1_scfv包含h-cdr1’和h-cdr3’的重链区主要为杀虫活性区,g1_scfv包含l-cdr1’和l-cdr2’的的轻链区主要为结合活性区。
根据上述的blast比对分析结果,为提高结合活性,分别通过以抗cry1ab毒素独特型单链抗体b12_scfv序列的轻链区(vl)利用g1_scfv序列的轻链区(vl)进行替换,获得ggcc_scfv,或b12_scfv序列的重链区(vh)利用g1_scfv序列的重链区(vh)进行替换,获得ccgg_scfv(如图1所示)。
改造后的ggcc_scfv、ccgg_scfv的氨基酸序列提交到blast网站上分别与cry1aa、cry1ab和cry1ac以及cry1b的氨基酸序列进行比对,比对结果如表1所示。ggcc_scfv的h-cdr1’和h-cdr3’区分别与cry1b蛋白的保守结构域endotoxin_n端结构域和endotoxin_m端结构域有相似序列,ggcc_scfv的l-cdr1和l-cdr2区分别与cry1a类蛋白的保守结构域endotoxin_c端结构域有相似序列;ccgg_scfv的h-cdr1区与cry1a类和cry1b蛋白的保守结构域endotoxin_n端结构域有相似序列,ccgg_scfv的h-cdr3区与cry1b蛋白的保守结构域endotoxin_m端结构域有相似序列,ccgg_scfv的l-cdr1’区与cry1a类蛋白的保守结构域endotoxin_c端结构域有相似序列,ccgg_scfv的l-cdr2’区与cry1b蛋白的保守结构域endotoxin_n端结构域有相似序列。
表1:blast分析结果
实施例3
b12_scfv、g1_scfv、改造后的ggcc_scfv、ccgg_scfv以及bt毒素cry1ab和小菜蛾(plutellaxyllostella)碱性磷酸酶alp(pxalp)同源建模和分子对接:
(1)同源建模
b12_phage的氨基酸序列已由专利号为zl201410037000.9的中国专利公开。g1_scfv的氨基酸序列如seqidno.2所示。pxalp氨基酸序列由ncbi蛋白质数据库获得,登录号为:ahf20243.2;bt毒素cry1ab氨基酸序列由ncbi蛋白质数据库获得,登录号为:alj10947.1。
改造后的ggcc_scfv、ccgg_scfv氨基酸序列根据b12_scfv、g1_scfv和cry1a类毒素(cry1aa、cry1ab、cry1ac)、cry1b的氨基酸序列的blast比对分析结果,分别通过以抗cry1ab毒素独特型单链抗体b12_scfv序列的轻链区(vl)利用g1_scfv序列的轻链区(vl)进行替换,或b12_scfv序列的重链区(vh)利用g1_scfv序列的重链区(vh)进行替换获得(如图1所示)。
b12_scfv、g1_scfv、cry1ab、改造后的ggcc_scfv和ccgg_scfv以及pxalp同源建模方法为:利用swiss-model网站(https://www.swissmodel.expasy.org/)从pdb(proteindatabank,brookhavennationallaboratory)数据库找到一致性较高的模板,并进行建模;在分别获得三维结构后,再利用saves网站(http://services.mbi.ucla.edu/saves/)的verify3d和errat程序对上述三维结构进行打分,评价三维结构与一级序列的兼容性及其可靠性。
具体为:b12_scfv选取scfvmab198的1f3r.1.b作为同源建模的模板,与模板氨基酸序列一致性为87.28%,其三维结构如图2-a所示,g1_scfv选取sp140phd-bromodomain复合物中结合的scfv6g8r.1.a作为模板,与模板氨基酸序列一致性为89.17%,其三维结构如图2-b所示;ggcc_scfv选取fabbl3-64q9r.1.a作为模板,与模板氨基酸序列一致性为79.58%,其三维结构如图2-c所示;ccgg_scfv选取sp140phd-bromodomain复合物中结合的scfv6g8r.1.a作为模板,与模板氨基酸序列一致性为88.33%,其三维结构如图2-d所示;cry1ab选取cryia(a)1ciy作为模板,与模板氨基酸序列一致性为89.46%,其三维结构如图2-e所示;pxalp选取人alp1zef.1.a作为模板,与模板氨基酸序列一致性为44.93%,其三维结构如图2-f所示。
对同源建模的b12_scfv、g1_scfv、改造后的ccgg_scfv、ggcc_scfv以及bt毒素cry1ab和pxalp三维结构模型的评价结果如表2所示:
表2三维结构模型评价结果
一个好的三维结构模型,errat评价应高于50以及verify3d评价中80%以上的氨基酸残基需大于0.2;如表2所示评价结果,上述5个同源建模的三维结构是合理可信的。
本步骤中蛋白三维结构输出使用pymol软件(thepymolmoleculargraphicssystem,delanoscientific,sancarlos,ca,usa)完成。
(2)分子对接及热点预测
定义pxalp为受体,cry1ab、b12_scfv、g1_scfv、改造后的ggcc_scfv和ccgg_scfv为配体,利用zdock程序(http://zdock.umassmed.edu/)对所构建的pxalp模型和四个配体模型的三维结构进行分子对接,首先针对cry1ab配体,选取前10的预测复合物,根据已知pxalp的结合位点进行筛选,并通过cry1ab配体上pxalp受体结合位点进行二次确定;根据pxalp与cry1ab间的结合热点区域筛选其他4个配体(b12_scfv、g1_scfv、ggcc_scfv和ccgg_scfv)复合物。pymol软件输出的b12_scfv-pxalp、g1_scfv-pxalp,ggcc_scfv-pxalp、ccgg_scfv-pxalp和cry1ab-pxalp对接复合物的空间结构分别如图3-g、3-h、3-i、3-j、3-k所示。
将上述4个对接复合物提交到mitchelllab的热点预测服务器kfc2hotspotpredictionserver(网址为http://kfc.mitchell-lab.org/)(darnelletal.,2007),预测结果如图4所示,热点预测服务器kfc2预测到pxalp受体中与5个配体结合的共有热点区域为trp53(alp受体结合区域i)和ala409、tyr411、tyr433(alp受体结合区域ii);与受体结合的5个配体的热点三维结构图如图4所示,其中图4-a、4-b、4-c、4-d和4-e分别cry1ab、b12_scfv、g1_scfv、ggcc_scfv、ccgg_scfv与alp受体结合区域i结合热点的三维结构图,在其右侧为配体的主要结合结构域;其中图4-f、4-g、4-h、4-i和4-j分别为cry1ab、b12_scfv、g1_scfv、ggcc_scfv、ccgg_scfv与alp受体结合区域ii结合热点的三维结构图,在其左侧为配体的主要结合结构域。
经比较,b12_scfv通过l-cdr1、l-cdr2区和h-cdr3区的热点氨基酸与pxalp受体结合,g1_scfv通过l-cdr1’和h-cdr1’、h-cdr2’、h-cdr3’区的热点氨基酸与pxalp受体结合,改造后的ggcc_scfv可通过h-cdr1’、h-cdr2’、h-cdr3’区和l-cdr2区与pxalp受体结合,改造后的ccgg_scfv仅可通过l-cdr2’区与pxalp受体结合。从分子对接模拟的结果综合来看,改造后的ggcc_scfv与pxalp受体结合热点部位较母体b12_scfv、g1_scfv有所增加,不仅包含g1母体相同的热点结构域h-cdr1’、h-cdr2’和h-cdr3’区外,还包含了b12母体的l-cdr2区,预测ggcc_scfv较b12_scfv、g1_scfv与alp的结合能力更强,而ccgg_scfv较b12_scfv、g1_scfv与alp的结合能力稍弱。
本步骤中涉及的所有程序都是由pymol软件完成并进行图像输出。
实施例4
对杀虫蛋白的筛选及亲和力测定:
(1)小菜蛾中肠bbmv制备
参照wolfersberger的实验方法(wolfersberger,1987),使用mg-egta沉降法制备小菜蛾中肠bbmv,具体做法为:取小菜蛾4龄末期幼虫,提取中肠,在预冷的0.15mol/lnacl中清洗,每500只中肠加入3ml匀浆缓冲液;反复匀浆冰浴后,加入等体积24mmol/lmgcl2混匀,冰浴并低速离心后将上清液转入新的超速离心管中再超速离心;弃上清后倒置离心管待液体流尽后,将沉淀重悬于hepes缓冲液中,分装后于-80℃贮存备用;bbmv蛋白浓度通过bradford法测定。
(2)生物膜层表面干涉技术(bio-layerinterferometry,bli)对杀虫蛋白的筛选
实验操作使用octet分子相互作用技术平台(fortebio),数据分析使用dataanalysis软件(fortebio)。反应过程中所用材料:系统缓冲液pbs(137mmol/lnacl,2.7mmol/lkcl,10mmol/lna2hpo4,and2mmol/lkh2hpo4,ph7.4)、kineticbuffer(1×pbsbuffercontaining0.1%(wt/vol)bsa)和氨丙基(aps)生物传感器(aminopropylsilane-coatedbiosensortips)均为fortebio标配产品。实验所用受体蛋白小菜蛾中肠bbmv使用pbs稀释至10μg/ml,待测样品b12_scfv、g1_scfv、ggcc_scfv、ccgg_scfv和bt毒素cry1ab、cry1b用kineticbuffer稀释到指定浓度。所有试剂均用去离子水配制,用0.22μm滤膜过滤。
将aps生物传感器置于pbs缓冲液中平衡30s,与10μg/ml小菜蛾中肠bbmv固化5min,用kineticbuffer平衡封闭aps生物传感器1min,同时置于表达纯化的b12_scfv、g1_scfv、ggcc_scfv、ccgg_scfv和bt毒素cry1ab、cry1b中结合5min,再置于bsa缓冲液解离5min。
实时监测结果如图5-a所示,ggcc_scfv的结合活性最好,为快结合慢解离,且结合未达到饱和状态;cry1ab结合活性较好,也为快结合慢解离,结合已达到饱和状态;b12_scfv结合效果不佳,表现为快结合快解离;ccgg_scfv、cry1b稍差,表现为慢结合慢解离;g1_scfv结合最差,为慢结合快解离。
(3)常规酶联免疫法(elisa)对杀虫蛋白的筛选
取30μg/ml小菜蛾中肠bbmv包被96微孔板4℃孵育过夜,100μl/孔,次日每孔加入200μl3%mpbs溶液,37℃孵育2h进行封闭;每孔用250μlpbst洗板,然后加入表达纯化的b12_scfv、g1_scfv、ccgg_scfv、ggcc_scfv和bt毒素cry1ab、cry1b(2倍梯度稀释),100μl/孔,37℃孵育2h;每孔用250μlpbst洗板,加入1:5000倍稀释的hrp-anti-histag标签抗体(bt毒素cry1ab/cry1b需加入cry1ab/cry1b的多抗100μl/孔,37℃孵育1h,pbst洗板后,再加入hrp-羊抗兔igg),100μl/孔,37℃孵育1h;pbst洗板后加入四甲基联苯胺(tmb)显色液,100μl/孔,37℃反应10~20min,最后加入浓度为2mol/l的h2so4快速终止反应,并用thermo全自动酶标仪,测定od450值。
结果如图5-b所示,在梯度稀释的样品中只有ggcc_scfv、cry1b和cry1ab有结合活性,其中ggcc_scfv的结合活性最好,od450值最高可达1.3以上。
(4)生物膜层表面干涉技术(bio-layerinterferometry,bli)对ggcc_scfv、cry1b和cry1ab亲和力的测定
实验所用受体蛋白小菜蛾中肠bbmv使用pbs稀释至10μg/ml,待测样品ggcc_scfv和bt毒素cry1b、cry1ab用kineticbuffer分别稀释至4μmol/l、2μmol/l、1μmol/l和0.5μmol/l。
将aps生物传感器置于pbs缓冲液中平衡30s,与10μg/ml小菜蛾中肠bbmv固化5min,用kineticbuffer平衡封闭aps生物传感器1min,同时置于表达纯化的ggcc_scfv、cry1b和cry1ab的梯度稀释溶液中结合5/10min,再置于kineticbuffer解离5/10min。
实时监测结果如图6-a、6-b和6-c所示,图6-a为不同稀释浓度下ggcc_scfv与小菜蛾bbmv的结合曲线,图6-b为不同稀释浓度下bt毒素cry1ab与小菜蛾bbmv的结合曲线,图6-c为不同稀释浓度下bt毒素cry1b与小菜蛾bbmv的结合曲线。亲和力常数如图6-d所示,ggcc_scfv与小菜蛾bbmv亲和力常数为12.8±1.47nm,cry1ab与小菜蛾bbmv亲和力常数为127.1±2.21nm,cry1b与小菜蛾bbmv亲和力常数为2600±930nm。
综上所述,在相同浓度下,改造后ggcc_scfv的bbmv结合活性比bt毒素cry1ab和cry1b强。b12_scfv由于快结合快解离(bli实时监测),在elisa检测(终点检测法)中未表现有结合活性。利用bli技术筛选bbmv结合活性蛋白,具有省时、省样、高通量、实时监测数据的特点,较传统elisa方法(10小时)简便。也较spr法(biacorex100实验平台)使用方便、简单、快捷。
实施例5
对杀虫蛋白的生物活性测定
将b12_scfv、g1_scfv、ggcc_scfv、ccgg_scfv和bt毒素cry1ab蛋白用pbs稀释至制定浓度,各取500μl铺于培养皿中饲料表面,晾干;阴性对照为pbs缓冲液,阳性对照为bt毒素cry1ab蛋白。
每个饲料培皿中接入30头已饥饿4h的小菜蛾2龄幼虫,然后置于温度为25±1℃,相对湿度80±5%,光照条件大于14h的培养箱内饲养,3d、7d后分别观察、记录死亡虫数,取出幼虫检查时以小毛笔轻触虫体,无明显反应者为死亡。每处理3次重复。试虫死亡率采用abbott公式对死亡率进行校正,并以平均数±标准误表示(3次重复试验)。
校正死亡率=(处理死亡率-对照死亡率)/(1-对照死亡率)×100%
各处理时间相同时各个样品之间的比较采用单因素方差分析(one-wayanova)和tukey显著性检验,使用spss软件进行数据处理,处理结果如图7所示。
生物活性测定结果表明,3d后ggcc_scfv对小菜蛾的校正致死率为50%,b12_scfv的校正致死率低于10%,3d后ggcc_scfv较b12_scfv的校正致死率提高了近5倍;7d后ggcc_scfv对小菜蛾的校正致死率为95%,b12_scfv的校正致死率低于40%,ggcc_scfv与cry1ab的校正致死率接近。
实施例6
杀虫蛋白ggcc_scfv与bt毒素的竞争性抑制实验
利用常规酶联免疫法(elisa),以小菜蛾中肠刷状缘膜囊泡蛋白bbmv为靶抗原,ggcc_scfv与bt毒素cry1ab、cry1ac和cry1b做竞争性抑制实验。
bt毒素(500mg/ml)cry1ab、cry1ac和cry1b用pbs缓冲液2倍稀释。稀释后的cry1ab、cry1ac或cry1b蛋白与等体积的ggcc_scfv蛋白混匀,37℃孵育2h.
取30μg/ml小菜蛾中肠bbmv包被96微孔板过夜,100μl/孔,次日每孔加入200μl3%mpbs溶液,37℃孵育2h进行封闭;每孔用250μlpbst洗板,然后加入一系列ggcc_scfv+cry1ab混合物、ggcc_scfv+cry1ac混合物、ggcc_scfv+cry1b混合物,100μl/孔,37℃孵育2h;每孔用250μlpbst洗板,加入1:5000稀释的hrp-anti-histag标签抗体,37℃孵育1h;pbst洗板后加入四甲基联苯胺(tmb)显色液,100μl/孔,37℃反应10-20min,最后加入浓度为2mol/l的h2so4快速终止反应,并用thermo全自动酶标仪,测定od450值。
elisa结果如图8(a)所示,随着bt毒素浓度的增加,ggcc_scfv与小菜蛾bbmv的结合活性被抑制。抑制率如图8(b)所示,cry1ab结合小菜蛾bbmv导致对ggcc_scfv半抑制的浓度(ic50)为89.6μg/ml,cry1ac结合小菜蛾bbmv导致对ggcc_scfv半抑制的浓度(ic50)为49.18μg/ml,cry1b结合小菜蛾bbmv导致对ggcc_scfv半抑制的浓度(ic50)为37.99μg/ml。cry1ab、cry1ac和cry1b蛋白都可以与ggcc_scfv竞争结合小菜蛾bbmv,ggcc_scfv被cry1b蛋白抑制的抑制率可达到73.6%。由此,可以确定ggcc_scfv为cry1b、cry1ab和cry1ac毒素的模拟物。
以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。
序列表
<110>江苏省农业科学院
<120>一种人源分子改造杀虫蛋白、编码基因及其设计方法和应用
<160>6
<170>siposequencelisting1.0
<210>1
<211>245
<212>prt
<213>人工序列(artificialsequence)
<400>1
metalagluvalglnleuleugluserglyglyglyleuvalglnpro
151015
glyglyserleuargleusercysalaalaserglyphethrpheser
202530
sertyralametsertrpvalargglnalaproglylysglyleuglu
354045
trpvalserargilehisthrserglyaspserthrglntyralaasp
505560
servallysglyargphethrileserargaspasnserlysasnthr
65707580
leutyrleuglnmetasnserleuargalagluaspthralavaltyr
859095
tyrcysalalysasptyrargtrppheasptyrtrpglyglnglythr
100105110
leuvalthrvalserserglyglyglyglyserglyglyglyglyser
115120125
glyglyglyglyserthraspileglnmetthrglnserproserser
130135140
leuseralaservalglyaspargvalthrilethrcysargalaser
145150155160
glnserilesersertyrleuasntrptyrglnglnlysproglylys
165170175
alaprolysleuleuiletyralaalaserargleuglnserglyval
180185190
proserargpheserglyserglyserglythraspphethrleuthr
195200205
ileserserleuglnprogluaspphealathrtyrtyrcysglngln
210215220
thrserhisproproleuthrpheglyglnglythrlysvalgluile
225230235240
lysargalaalaala
245
<210>2
<211>118
<212>prt
<213>人(homosapiens)
<400>2
metalagluvalglnleuleugluserglyglyglyleuvalglnpro
151015
glyglyserleuargleusercysalaalaserglyphethrpheser
202530
sertyralametsertrpvalargglnalaproglylysglyleuglu
354045
trpvalserargilehisthrserglyaspserthrglntyralaasp
505560
servallysglyargphethrileserargaspasnserlysasnthr
65707580
leutyrleuglnmetasnserleuargalagluaspthralavaltyr
859095
tyrcysalalysasptyrargtrppheasptyrtrpglyglnglythr
100105110
leuvalthrvalserser
115
<210>3
<211>109
<212>prt
<213>人(homosapiens)
<400>3
thraspileglnmetthrglnserproserserleuseralaserval
151015
glyaspargvalthrilethrcysargalaserglnserileserser
202530
tyrleuasntrptyrglnglnlysproglylysalaprolysleuleu
354045
iletyralaalaserargleuglnserglyvalproserargpheser
505560
glyserglyserglythraspphethrleuthrileserserleugln
65707580
progluaspphealathrtyrtyrcysglnglnthrserhispropro
859095
leuthrpheglyglnglythrlysvalgluilelysarg
100105
<210>4
<211>15
<212>prt
<213>人工序列(artificialsequence)
<400>4
glyglyglyglyserglyglyglyglyserglyglyglyglyser
151015
<210>5
<211>242
<212>prt
<213>人(homosapiens)
<400>5
metalagluvalglnleuleugluserglyglyglyleuvalglnpro
151015
glyglyserleuargleusercysalaalaserglyphethrpheser
202530
sertyralametsertrpvalargglnalaproglylysglyleuglu
354045
trpvalserargilehisthrserglyaspserthrglntyralaasp
505560
servallysglyargphethrileserargaspasnserlysasnthr
65707580
leutyrleuglnmetasnserleuargalagluaspthralavaltyr
859095
tyrcysalalysasptyrargtrppheasptyrtrpglyglnglythr
100105110
leuvalthrvalserserglyglyglyglyserglyglyglyglyser
115120125
glyglyglyglyserthraspileglnmetthrglnserproserser
130135140
leuseralaservalglyaspargvalthrilethrcysargalaser
145150155160
glnserilesersertyrleuasntrptyrglnglnlysproglylys
165170175
alaprolysleuleuiletyrglyalaserthrleuglnserglyval
180185190
proserargpheserglyserglyserglythraspphethrleuthr
195200205
ileserserleuglnprogluaspphealathrtyrtyrcysglngln
210215220
alaserglualaproserthrpheglyglnglythrlysvalgluile
225230235240
lysarg
<210>6
<211>726
<212>dna
<213>人工序列(artificialsequence)
<400>6
atggccgaggtgcagctgttggagtctgggggaggcttggtacagcctggggggtccctg60
agactctcctgtgcagcctctggattcacctttagcagctatgccatgagctgggtccgc120
caggctccagggaaggggctggagtgggtctcaaggattcatacgagtggtgattcgaca180
cagtacgcagactccgtgaagggccggttcaccatctccagagacaattccaagaacacg240
ctgtatctgcaaatgaacagcctgagagccgaggacacggccgtatattactgtgcgaaa300
gattatcggtggtttgactactggggccagggaaccctggtcaccgtctcgagcggtgga360
ggcggttcaggcggaggtggcagcggcggtggcgggtcgacggacatccagatgacccag420
tctccatcctccctgtctgcatctgtaggagacagagtcaccatcacttgccgggcaagt480
cagagcattagcagctatttaaattggtatcagcagaaaccagggaaagcccctaagctc540
ctgatctatgctgcatcccggttgcaaagtggggtcccatcaaggttcagtggcagtgga600
tctgggacagatttcactctcaccatcagcagtctgcaacctgaagattttgcaacttac660
tactgtcaacagacttctcatcctcctctgacgttcggccaagggaccaaggtggaaatc720
aaacgg726
- 还没有人留言评论。精彩留言会获得点赞!