植物花药花粉发育后期特异性表达启动子pOsLPS3的鉴定和应用的制作方法
本发明属于植物生物技术种领域,具体而言,本发明涉及分离的dna,其能够指导可操作地连接在其下游的核酸在植物花药花粉中特异转录和/或表达。另外,本发明还涉及包含该dna的表达盒和植物等,并涉及该dna的应用。
背景技术:
植物基因调控主要是在转录水平上进行的,受多种顺式作用元件和反式作用因子的相互协调。启动子是重要的顺式作用元件,它是位于结构基因5’端上游区域调控基因转录的一段dna序列,能活化rna聚合酶,使之与模板dna准确地结合,确保转录精确而有效地起始,在转录调控中起关键作用。根据其驱动基因表达的不同特点,启动子分为组成型启动子和特异性启动子:组成型启动子能在所有细胞或组织中,不分时间和空间地启动转录;特异性启动子又可分为组织特异性启动子和诱导型启动子,其中组织特异性启动子可启动下游基因在某些特定的器官或组织部位表达;而诱导型启动子平时不启动转录或转录活性很低,但在某些特定的逆境信号的刺激下,转录活性能够显著地提高。
外源dna序列可以通过连接到特定的启动子从而启动其在植物宿主中的表达,启动子类型的选择决定基因的表达时间和部位。目前在农业生物技术领域广泛应用的主要是一些组成型的强启动子,比如花椰菜花叶病毒的camv35s启动子和玉米的ubiquitin-1启动子,然而在利用这些启动子诱导目的基因转化水稻等作物以期改良作物的品质时,往往会由于目的基因表达的时间(发育阶段特异性)或空间(组织器官特异性)不能很好地控制而导致改良效果不明显,或者由于这些组成型启动子诱导基因表达量太高而对植物的生长发育造成影响,这些都是目前利用组成型强启动子结合功能基因来改良作物品质时遇到的障碍。
此外,在研究某些代谢过程或调节途径时,常常需要将同一途径上的两个以上基因转化到同一个株系中去,如果采用转化其中一个基因得到转基因植株后再转化另外一个基因,或者两个基因分别转化完成后再进行杂交,都需要等待较长的时间。为了提高效率以缩短多个基因转化的时间,最近有报道可以利用新的载体同时进行多个基因的转化,但是在多基因转化时如果重复使用同一个启动子,也会由于启动子序列的高同源性可能导致基因沉默。
近年来,通过基因工程调控植物花粉育性、创造植物雄性不育系及其恢复系已在一些作物上获得成功,为作物杂种优势的利用开创了新的前景。目前利用基因工程创造雄性不育的策略主要是利用花粉发育的特异启动子与外源基因嵌合,构建表达载体,转化植物来阻断花粉发育的过程从而达到雄性不育的目的。植物花粉或花药启动子的驱动活性和特异性决定了通过基因工程手段调控花粉育性、创造植物不育系及恢复系的成败。目前已知的驱动活性高且特异性良好的植物花粉或花药特异启动子还相对较少,因此,对重要农作物--水稻的花粉特异表达启动子的克隆和功能分析对于在水稻中利用基因工程调控花粉育性、创造植物雄性不育系,从而为水稻杂种优势资源在水稻育种中的充分利用打下基础。
技术实现要素:
本发明的目的是提供一种植物花药花粉发育后期特异表达的启动子序列及克隆并应用该启动子的方法。
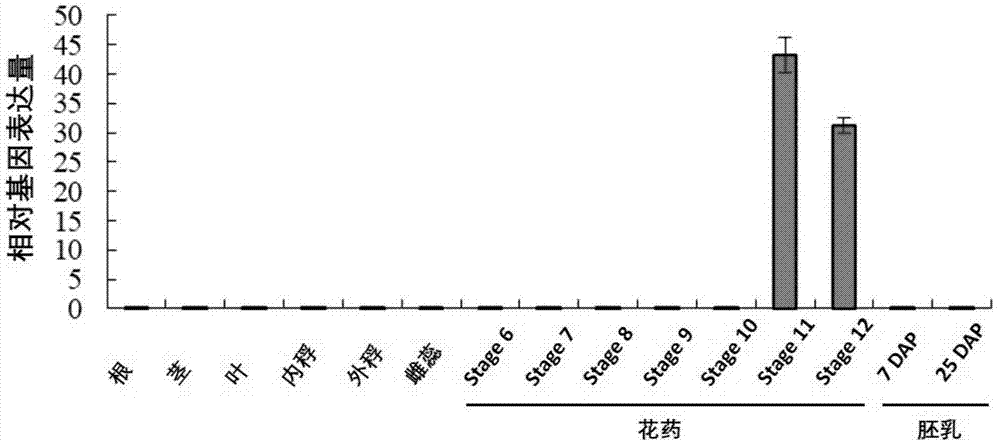
发明人在研究中偶然发现,水稻oslps3(latestagepollen-specific)基因在不同组织器官和不同发育时期的花药中表达量明显不同。分别取水稻植株的根、茎、叶、外稃、內稃、雌蕊、花药(stage6-12)和胚乳(dap7和dap25),提取rna,反转录为cdna作为模板,以水稻actin基因作为内参照,利用quantitativerealtime-pcr(qrt-pcr)方法,分析了oslps3基因在水稻各个组织中的表达情况。
通过对不同发育时期水稻oslps3基因的表达量分析,发现oslps3基因只在花药发育后期(stage11-12)特异性表达,且在stage11期表达量最高,在根、茎、叶或同时期的其他花器官等组织器官中均不表达(如图1所示)。这表明,oslps3基因是花药特异性表达,且只在花药发育后期(stage11-12)中特异性表达的基因。
本发明提供了一种poslps3启动子,所述启动子具有在花药花粉发育后期中特异表达的功能,其位于oslps3基因自翻译起始位点atg往上-2501bp的片段,序列如seqidno:1所示。为了进一步验证该启动子的功能,将poslps3启动子与报告基因egfp(enhancedgreenfluorescentprotein,序列如seqidno:2所示)相连并转化植物,在转基因植株的根、茎、叶、外稃、內稃、雌蕊、胚乳(dap7和dap25)等器官中均检测不到egfp基因的表达,在花药stage6-10时期中也检测不到egfp基因的表达,poslps3启动子只能驱动egfp基因在水稻的花药花粉发育后期中表达,更具体地只在花药花粉发育的stage11-12期特异性的高表达。说明本发明所提供的poslps3启动子(seqidno:1)是一个花药花粉特异性表达的启动子。
本发明所提供的植物花药花粉特异表达启动子,含有如seqidno:1所示的核苷酸序列,或与seqidno:1所示序列互补的dna分子,或包含与seqidno:1中所列核苷酸序列具有90%以上相似性的核苷酸序列,或包含来源于seqidno:1序列上的100个及100以上连续的核苷酸片段,并且可以驱动与该启动子操作性连接的核苷酸序列在植物花药花粉中的特异性表达。本领域技术人员根据相同目的能够容易地鉴定并利用与植物花粉花药特异性启动子poslps3核苷酸序列互补的dna分子,因此,具有启动子活性并在严格条件下与本发明启动子序列或其片段杂交的分离序列包括在本发明中。含有上述序列的表达载体、转基因细胞系以及宿主菌等均属于本发明的保护范围。扩增本发明所公开的seqidno:1启动子的任一核苷酸片段的引物对也在本发明的保护范围之内。
本发明所提供的启动子核苷酸序列还可用于从水稻以外的其它植物中分离相应序列,尤其是从其他单子叶植物中进行同源克隆。根据这些相应序列与本文所列启动子序列间的序列同源性,或与本启动子基因的同源性,使用如pcr、杂交等技术来鉴别分离这些相应序列。因此,根据它们与本发明所列的seqidno:1启动子序列(或其片段)间的序列相似性而分离的相应片段,也包括在实施方案中。
本发明所述的“启动子”是指一种dna调控区域,其通常包含能指导rna聚合酶ii在特定编码序列的合适转录起始位点起始rna合成的tata盒。启动子还可包含其它识别序列,这些识别序列通常位于tata盒的上游或5’端,通常被称为上游启动子元件,起调控转录效率的作用。本领域技术人员应该知晓,虽然已经鉴定了针对本发明公开的启动子区域的核苷酸序列,但是分离和鉴定处于本发明鉴定的特定启动子区域的tata盒上游区域的其它调控元件也在本发明的范围内。因此,本文公开的启动子区域通常被进一步界定为包含上游调控元件,例如用于调控编码序列的组织表达性和时间表达功能的那些元件、增强子等。以相同的方式,可以鉴定、分离出使得能在目标组织(例如雄性组织)中进行表达的启动子元件,将其与其它核心启动子一起使用,以验证雄性组织优先的表达。核心启动子指起始转录所需的最小限度的序列,例如被称为tata盒的序列,这是编码蛋白质的基因的启动子通常都具有的。因此,可选地,poslps3启动子可与其自身的或来自其它来源的核心启动子关联使用。
核心启动子可以是任何一种已知的核心启动子,例如花椰菜花叶病毒35s或19s启动子(美国专利no.5,352,605)、泛素启动子(美国专利no.5,510,474)、in2核心启动子(美国专利no.5,364,780)或玄参花叶病毒启动子。
所述基因启动子的功能可以通过以下方法进行分析:将启动子序列与报告基因可操作性连接,形成可转化的构建体,再将该构建体转入植株中,在获得转基因后代中,通过观察报告基因在植物各个组织器官中的表达情况来确认其表达特性;或者将上述构建体亚克隆进用于瞬时表达实验的表达载体,通过瞬时表达实验来检测启动子或其调控区的功能。
用来测试启动子或调控区域功能的适当表达载体的选择将取决于宿主和将该表达载体引入宿主的方法,这类方法是本领域普通技术人员所熟知的。对于真核生物,在载体中的区域包括控制转录起始和控制加工的区域。这些区域被可操作地连接到报告基因,所述报告基因包括gfp、uida、gus基因或荧光素酶。包含位于基因组片段中的推定调控区的表达载体可以被引入完整的组织,例如阶段性花药,或引入愈伤组织,以进行功能验证。
启动子的活性和强度可以根据其驱动的报告基因的mrna或蛋白质的表达量来测定。报告基因(reportergene)是一种编码可被检测的蛋白质或酶的基因,也就是说,是一个其表达产物非常容易被鉴定的基因。将报告基因的编码序列和基因表达调节序列相融合形成嵌合基因,或与其它目的基因相融合,在调控序列控制下进行表达,从而利用它的表达产物来确定目的基因的表达调控特性。常用的报告基因有绿色荧光蛋白基因gfp,红色荧光蛋白rfp和β-葡萄糖苷酸酶基因gus。
本发明通过egfp报告基因来检测启动子的表达特性。egfp蛋白在488nm的激发光下能发出绿色荧光,用激光共聚焦显微镜或荧光显微镜可检测到,在一定程度下可根据荧光亮度来反映出egfp基因表达的强弱。因此利用该方法可观察到外源基因在特定器官、组织,甚至单个细胞内的表达情况。
此外,本发明的启动子可与并非oslps3基因的核苷酸序列相连,以表达其它异源核苷酸序列。本发明的启动子核苷酸序列及其片段和变体可与异源核苷酸序列一起组装在一个表达盒中,用于在目的植株中表达,更具体地,在该植株的雄性器官中表达。所述表达盒有合适的限制性酶切位点,用于插入所述启动子和异源核苷酸序列。这些表达盒可用于对任何植株进行遗传操作,以获得想要的相应表型。
本发明所公开的poslps3启动子,可用于驱动下列异源核苷酸序列的表达,以使转化的植株获得雄性不育的表型。所述异源核苷酸序列可编码促使碳水化合物降解的酶或修饰酶、淀粉酶、脱支酶和果胶酶,更具体的如α-淀粉酶基因、生长素(auxin)、rotb、细胞毒素基因、白喉毒素、dam甲基化酶、亲和素,或者可选自原核调控系统,还可以是显性的雄性不育基因。
在某些实施方式中,本发明中所提到的可操作地连接在本发明启动子下游的核酸,其中所述的“核酸”可以是操作性连接于本文所公开的启动子之上的结构基因、调节基因、结构基因的反义基因、调节基因的反义基因或者能够干扰内源基因表达的小rna。
本发明所提供的启动子序列可分离自任何植物,包括但不限于芸苔属、玉米、小麦、高粱、两节荠属、白芥、蓖麻子、芝麻、棉籽、亚麻子、大豆、拟南芥属、菜豆属、花生、苜蓿、燕麦、油菜籽、大麦、燕麦、黑麦(rye)、粟、蜀黍、小黑麦、单粒小麦、斯佩尔特小麦(spelt)、双粒小麦、亚麻、格兰马草(grammagrass)、摩擦禾、假蜀黍、羊茅、多年生麦草、甘蔗、红莓苔子、番木瓜、香蕉、红花、油棕、香瓜、苹果、黄瓜、石斛、剑兰、菊花、百合科、棉花、桉、向日葵、芸苔、甜菜、咖啡、观赏植物和松类等。优选地,植物包括水稻、玉米、大豆、红花、芥菜、小麦、大麦、黑麦、棉花和高粱。
本发明还包括含有poslps3启动子的构建体,所述构建体包括通常所说的载体或表达盒。上述构建体中还可包括其它组分,这主要取决于载体构建的目的和用途,例如可进一步包括选择标记基因、靶向或调控序列、稳定序列或引导序列、内含子等。表达盒还将在目标异源核苷酸序列的3’端包括在植物中具有功能的转录和翻译终止子。终止子可以是本发明所提供基因的终止子,也可以是来自外源的终止子。更具体地,上述终止子可以是胭脂氨酸合酶或章鱼碱合酶终止区域。
在希望将异源核苷酸序列的表达产物引向特定细胞器,例如质体、造粉体,或者引向内质网,或在细胞表面或细胞外分泌的情况下,表达盒还可包含用于编码转运肽的核苷酸序列。此类转运肽是本领域所公知的,其包括但不限于rubisco的小亚基、植物epsp合酶、玉米brittle-1叶绿体转运肽等。
在制备表达盒的过程中,可对多种dna片段加以操作,以提供处于合适方向,或是处于正确读码框中的dna序列。为达到此目的,可使用衔接子或接头,将dna片段连起来,或者进一步包括其它操作,以提供方便的限制性酶切位点等。
进一步地,本发明所提供的构建体中还可包括选择标记基因,用于选择经转化的细胞或组织。所述选择标记基因包括赋予抗生素抗性或对除草剂抗性的基因。合适的选择标记基因包括但不限于:氯霉素抗性基因、潮霉素抗性基因、链霉素抗性基因、奇霉素抗性基因、磺胺类抗性基因、草甘磷抗性基因、草丁嶙抗性基因。所述选择标记基因还可以是红色荧光基因、青色荧光蛋白基因、黄色荧光蛋白基因、荧光素酶基因、绿色荧光蛋白基因、花青甙p1等基因。
本发明所提供的表达盒或载体可被插入质粒、粘粒、酵母人工染色体、细菌人工染色体或其他适合转化进宿主细胞中的任何载体中。优选的宿主细胞是细菌细胞,尤其是用于克隆或储存多核苷酸、或用于转化植物细胞的细菌细胞,例如大肠杆菌、根瘤土壤杆菌和毛根土壤杆菌。当宿主细胞是植物细胞时,表达盒或载体可被插入被转化的植物细胞的基因组中。插入可以是定位的或随机的插入。优选地,插入通过诸如同源重组来实现。另外,表达盒或载体可保持在染色体外。本发明的表达盒或载体可存在于植物细胞的核、叶绿体、线粒体和/或质体中。优选地,本发明的表达盒或载体被插入植物细胞核的染色体dna中。
本发明还提供了扩增所述花粉花药特异性启动子poslps3的引物对,所述引物对的核苷酸序列为seqidno:9-10所示。
一种扩增花粉花药特异性启动子poslps3的方法,其为通过seqidno:9-10引物对pcr扩增花粉花药特异性启动子poslps3的核苷酸序列。
本发明还包括所公开的poslps3启动子的应用,在某些应用的实施方式中,可以应用本发明所提供的poslps3启动子来实现一些育性相关基因突变所获得的雄性不育系的繁殖和保持,所述育性相关基因包括但不限于osnp1、zmms7、ms26、ms45和msca1等。
本发明的所提供的花药花粉特异表达启动子可用于外源基因在花药中的特异性表达,从而避免该外源基因在植物其他组织中持续表达所带来的不利影响,还可以用于植物花药生长发育相关基因的功能分析和鉴定;可用于雄性不育系、保持系和恢复系的创建;并可应用于花粉败育实验中,从而避免由植物转基因漂移或花粉逃逸所带来的生物安全问题,对植物雄性不育系和恢复系的创造具有重要意义。
本发明的转基因植物使用植物生物技术领域技术人员已知的转化方法制备。任何方法可被用于将重组表达载体转化进植物细胞中,以产生本发明的转基因植物。转化方法可包括直接和间接的转化方法。合适的直接方法包括聚乙二醇诱导的dna摄入、脂质体介导的转化、使用基因枪导入、电穿孔、以及显微注射等。在本发明的具体实施方式中,本发明使用了基于土壤杆菌的转化技术(seiichitoki,nahohara,kazukoono,etal.(2006)earlyinfectionofscutellumtissuewithagrobacteriumallowshigh-speedtransformationofrice.theplantjournal.47,969–976;苏益,黄善金,蔺万煌等(2008).根癌农杆菌介导的水稻快速转化方法研究.中国农学通报(j),24(5):83~86)。土壤杆菌菌株(例如根瘤土壤杆菌或毛根土壤杆菌)包含质粒(ti或ri质粒)和t-dna元件,所述质粒和元件在用土壤杆菌转染后被转移至植物,而t-dna被整合进植物细胞的基因组中。t-dna可位于ri-质粒或ti-质粒上,或独立地包含在所谓的双元载体中。土壤杆菌介导的转化方法描述于例如中。土壤杆菌介导的转化最适合双子叶植物,但是也适合单子叶植物。土壤杆菌对植物的转化描述于例如中。转化可导致瞬时或稳定的转化和表达。尽管本发明的核苷酸序列可被插入落入这些广泛种类中的任何植物和植物细胞中,但是其尤其适用于作物植物细胞。
本发明提供的花粉花药特异启动子poslps3优点如下:
(1)花粉花药特异启动子poslps3为水稻内源基因,对水稻基因工程十分有利;
(2)花药特异启动子poslps3驱动报告基因特异性表达实验表明,花粉花药特异启动子poslps3驱动外源基因在花粉花药中特异性表达的表达水平精确;
(3)本发明提供了一种驱动外源基因在花粉花药中特异性表达的新方法。
下面通过具体实施方式,结合附图对本发明做进一步详细描述,但不以任何方式限制本发明的范围。
附图说明
图1是oslps3基因在水稻不同组织器官和不同发育时期的花药中的real-timepcr表达分析。分别取根、茎、叶、外稃、內稃、雌蕊、花药(stage6-12)和胚乳(dap7和dap25)材料进行检测。stage6-12分别指取自该材料中不同时期的花药组织,dap(dayafterpollination)指授粉后的天数。
图2是poslps3重组表达载体pszyjy-04的构建流程图。先由原始载体pcambia1300插入egfp基因后获得中间载体pszyjy-03,然后再插入poslps3序列后得到pszyjy-04载体。
图3是szyjy-04转基因水稻的组织器官的gfp荧光检测。其中,a为转基因水稻的根;b为转基因水稻的茎;c为转基因水稻的叶片;d-j分别为转基因水稻的stage6-12期花药;k为转基因水稻的种子外稃;l为转基因水稻的种子内稃;m为转基因水稻的雌蕊;n为转基因水稻的早期种子胚乳(授粉后7天);o为转基因水稻晚期种子胚乳(授粉后25天)。dap(dayafterpollination)指授粉后的天数。白色箭头指向观察到的绿色荧光。a-c、k-l、n-o的标尺为2mm;d-j和m的标尺为100μm。
图4是szyjy-04转基因水稻的成熟花药和花粉的gfp荧光检测。其中,a和b分别为野生型和转基因水稻的stage11-12期花药中的gfp荧光;c和d分别为野生型和转基因水稻的stage11-12期花粉中的gfp荧光。白色箭头指向能观察到的绿色荧光。标尺为100μm。
具体实施方式
下述实施例中所用方法如无特别说明均为常规方法,所用引物均由北京六合华大基因科技股份有限公司合成,测序由生工生物工程(上海)股份有限公司完成,kodfxneo高保真酶购自东洋纺(上海)生物科技有限公司(toyobo)、载体构建过程中的核酸内切酶、重组连接酶in-fusion和实时荧光定量pcr试剂盒均购自宝生物工程(北京)有限公司(takara),大肠杆菌感受态trans1-t1和反转录试剂盒购自北京全式金生物技术有限公司(transgenbiotech)。实验中所用的载体pcambia1300可购自于cambia公司。
实施例1.oslps3基因的组织表达模式分析
发明人在研究中偶然发现,水稻oslps3基因在不同组织器官和不同发育时期的花药中表达量明显不同。分别取水稻植株的根、茎、叶、外稃、內稃、雌蕊、花药(stage6-12)和胚乳(dap7和dap25),提取rna,反转录为cdna作为模板,以水稻actin基因作为内参照,分别设计针对oslps3和actin基因的引物,利用quantitativerealtime-pcr(qrt-pcr)方法,分析了oslps3基因在水稻各个组织中的表达情况。
oslps3基因的qrt-pcr引物为:
引物1:5'-ccagaacaacgaggactaccg-3'(seqidno:3);
引物2:5'-gtcaccaccgacttgagcc-3'(seqidno:4);
actin基因的qrt-pcr引物为:
引物3:5'-gctatgtacgtcgccatcca-3'(seqidno:5);
引物4:5'-ggacagtgtggctgacaccat-3'(seqidno:6);
qrt-pcr检测体系为:
加ddh2o补齐至10ul。
qrt-pcr反应条件:95℃,30s;95℃,5s,60℃,34s,40个循环。
在real-timepcr仪上进行反应,反应结束后,分析软件输出的数据,得到的real-timepcr检测结果。结果如图1所示,oslps3基因只在花药发育后期(stage11-12)特异性表达,且在stage11期表达量最高,在根、茎、叶或同时期的其他花器官等组织器官中均不表达。这表明,本发明的oslps3基因是花药特异性表达,且只在花药发育后期(stage11-12)中特异性表达的基因。
实施例2.荧光报告基因egfp和启动子poslps3的克隆
克隆荧光报告基因egfp所需引物为:
引物5:5'-ccggggatcctctagaatggtgagcaagggcgaggagctgt-3'(seqidno:7);
引物6:5'-ggccagtgccaagcttttacttgtacagctcgtccatgccg-3'(seqidno:8);
克隆启动子poslps3所需引物为:
引物7:5'-ccatgattacgaattcgggtgacggcaggtactgatatata-3'(seqidno:9);
引物8:5'-tgctcaccattctagattacttgtacagctcgtccatgccg-3'(seqidno:10);
其中,引物5和8中下划线序列为xbai的酶切位点,引物6中下划线序列为hindiii的酶切位点,引物7中下划线序列为ecori的酶切位点。引物5、6、7和8酶切位点左侧的10个碱基为骨架载体重组片段,用于连接载体时进行同源重组连接。
利用荧光报告基因的引物(seqidno:7和8),以实验室保存的egfp基因载体质粒为模板,进行pcr扩增,反应条件是:94℃预变性3min;98℃变性30s,61℃退火30s,68℃延伸1min,30个循环;68℃延伸5min。扩增产物长度约700bp,为带有限制性内切酶位点及骨架载体重组片段的序列(seqidno:2所示)。pcr产物经1%琼脂糖凝胶电泳检测回收。
同时,利用poslps3启动子的引物(seqidno:9和10),以水稻日本晴基因组dna模板,进行扩增,反应条件是:94℃预变性3min;98℃变性30s,61℃退火30s,68℃延伸3min,30个循环;68℃延伸7min。扩增产物长度约2500bp,为带有限制性内切酶位点及骨架载体重组片段的启动子序列(seqidno:1所示)。pcr产物经1%琼脂糖凝胶电泳检测回收。
实施例3.表达载体pszyjy-04的构建
如图2所示,将实施例2中pcr胶回收的报告基因egfp序列在同源重组酶in-fusion的作用下连入用xbai和hindiii双酶切的载体pcambia1300中,挑取菌落进行pcr检测,选取pcr结果为阳性的菌落进行测序,egfp报告基因的序列如seqidno:2,测序验证正确后,提取相应阳性克隆质粒,命名为pszyjy-03。
然后,将实施例2中胶回收的poslps3启动子序列在同源重组酶in-fusion的作用下连入再次用ecori和xbai双酶切的载体pszyjy-03中,挑取菌落进行pcr检测,选取pcr结果为阳性的菌落进行测序,poslps3启动子的序列如seqidno:1,测序验证正确后,提取相应阳性克隆质粒,命名为pszyjy-04。其中,菌落pcr检测所需引物为pcambia1300载体上引物(seqidno:11和12),位于所克隆的报告基因和启动子片段两侧。
菌落pcr的检测引物如下所示:
引物9:5'-ccaggctttacactttatgc-3'(seqidno:11);
引物10:5'-gcgattaagttgggtaacgc-3'(seqidno:12);
pcr检测体系为:
加ddh2o补齐至20ul。
pcr反应条件为:94℃预变性3min;94℃变性30s,61℃退火30s,72℃延伸3min,30个循环;72℃延伸7min。
实施例4.农杆菌介导的水稻遗传转化及转基因植株的功能鉴定
利用电击法将质粒pszyjy-04转入农杆菌agl0菌株,采用农杆菌介导法对水稻进行共转化,获得阳性转基因植株szyjy-04。并且,从上述阳性转基因植株中分离各组织器官,即根、茎、叶、外稃、內稃、雌蕊、花药(stage6-12)和胚乳(dap7和dap25),分别采用激光共聚焦进行gfp荧光检测。
结果如图3所示,egfp基因只在转基因水稻的花药发育后期(stage11-12)中有很强的表达,显现出肉眼可明显观测到的较强的绿色;而在其它各器官(叶片、茎、和根)及花药的其它时期中,基本没有检测到egfp基因的表达。进一步的观察发现,egfp基因只在pszyjy-04转基因植株的成熟花粉(stage11-12)中表达,结果如图4所示,而在其他组织或器官中没有表达,即没有产生荧光。这表明,本发明的启动子poslps3能够在转基因植株stage11-12期的花药花粉中指导其下游的egfp蛋白表达,而且这种表达具有花药花粉组织表达特异性。
序列表
<110>深圳广三系农业科技有限公司
<120>植物花药花粉发育后期特异性表达启动子poslps3的鉴定和应用
<160>12
<170>siposequencelisting1.0
<210>1
<211>2501
<212>dna
<213>oryzasativa
<400>1
gggtgacggcaggtactgatatatagagtttagggtcgtgcaaccaaccattgacgcaac60
cctaatgggttccaacacgatacacggcccaaaggcccaaatacggtgacgcaacgccag120
gacagattctagacgtcaacttgcgagaaggctagttagcgatcagtcgcgcgcgcccct180
ccccgcgcgactggacacgtgtcgcgcgcggagaggggacggcggacgcgccgcgctaag240
aggagagagagagagagagcaagttttccctttttttgttttttccctttttttggtttc300
tgtttggttggtttttccttttttttctggtttctctttttaaggtgtaggaatacaaag360
tttgtattcgtatgtataccaagtttgtatttgtacgtatataaactttgtatacgcgta420
tgcaaattttgtatacgttgtgtattttctagttttttattttttaatacgtatgtatat480
aaagtttatattcatcgtatacaaagtttgtacgtcctatataaactttgtatatgtata540
tatttttcatgcattaaaaatatattcatatatatatatatatatatatatgaataccat600
ggtatctaatgcataaaatcactcaaattttttaaaattttcaaattgtgagtatatacc660
tagttataataattcgattcatacctatacctatcaacaaaacaactcaaatttaacttt720
atttcacaatccatgattacatacctaactaattatatgtatggatgctatatacctaga780
atcaagatctaacaaaaacaagttcaagttcagttcaaacttgctttgaactagttagta840
aagtagttaatgttaatacctaagtaattgaaaaagtttcatactcatttgaattgggta900
tatactcaatttcaacaatggtgcccgggcaccatggagcaccaaacaaatttaatatat960
atatatatatatatatatatatatatatatatatatatatatatataggtaaattaactc1020
gtgctacatggagtattggctccaagattcaaattgagtatataaccaatttgaatgagt1080
atgaattttttttaaatgttttaggtatgaacattaacttctttactaacaagtttgaac1140
tgagtttgaacttttttttttgttagattttgattctaggtatataacatccaaacatat1200
aattcgttaggtatgtaataatgaattgtgaaataaagttgagtttgagttattttgttg1260
ttaggtattggtatgaatcaaatttatagacctaggtatatactcacaatttaaattttt1320
taaaaaatttgagtgattttgtgtgttttataccttagagcccgggcaccatagagtccc1380
atatgggtatctatatatatatatacatatatatatatatacacattggatatgttatct1440
tcttagctttccatccatacaaagaaaaccccaattcagtattaccttcaatgtttaagt1500
agttttgtgattttaccaaatagaattgctaaaagttttttttgtttggattctatctat1560
ttctgtgtcatcagatgcacaccgagcgtaaataaaatcacgcatgggggcgggtgatta1620
aactcatgaccatgcctaccaagggagcaaaattacccagatttcataataattacatgc1680
aagttgcatataagttataaaatttagtttaaaatataattacactatatttgcactata1740
acgttatatttaatatatttccacccaaaaagttatcgcagcacttttagagtggtcttt1800
actataagtcatcgggatcgagtgacaccaatgttaattgttaattgttagcctgtcaat1860
atagatccacccctaaacataagaccaaacttgccaagaatatatctcgttttaacacac1920
tcttaatagccgcgacttatggtatactgacaaacctcacaaaattcttactgaaagcaa1980
atcctccatgtgcgtgtgtacactaggatttaaactagatactgtgtacccaacgccacg2040
gctaccccaacgatgttatcccctcactatactaatgttattctgggaccacctaacttt2100
ctgcaaatatgaataatagacatccaaatttaactcaaaattaaatcttcaatatttatt2160
ttgcaactaaacccttgttcattgaacgcagcaacagagaaatctacggatcttaccatt2220
ctatttctctgtgaactaattgcccagtacatgtctataaataacatcactcacctgcat2280
gcgtccactcatccagcaactgtttctctgtgtaccgtttctctcgccgattgtgtaccg2340
attcaattcaaaatatcgaaaacaacgcaccatttttcgtaaatcataattagtgctagc2400
taagtcaatcctgcaagcctatatatattcgcttctctgcataattagctgaatcgtctt2460
cgttagcagtgagagtgacagacgacacatagaaggttttc2501
<210>2
<211>720
<212>dna
<213>aequoreavictoria
<400>2
atggtgagcaagggcgaggagctgttcaccggggtggtgcccatcctggtcgagctggac60
ggcgacgtaaacggccacaagttcagcgtgtccggcgagggcgagggcgatgccacctac120
ggcaagctgaccctgaagttcatctgcaccaccggcaagctgcccgtgccctggcccacc180
ctcgtgaccaccctgacctacggcgtgcagtgcttcagccgctaccccgaccacatgaag240
cagcacgacttcttcaagtccgccatgcccgaaggctacgtccaggagcgcaccatcttc300
ttcaaggacgacggcaactacaagacccgcgccgaggtgaagttcgagggcgacaccctg360
gtgaaccgcatcgagctgaagggcatcgacttcaaggaggacggcaacatcctggggcac420
aagctggagtacaactacaacagccacaacgtctatatcatggccgacaagcagaagaac480
ggcatcaaggtgaacttcaagatccgccacaacatcgaggacggcagcgtgcagctcgcc540
gaccactaccagcagaacacccccatcggcgacggccccgtgctgctgcccgacaaccac600
tacctgagcacccagtccgccctgagcaaagaccccaacgagaagcgcgatcacatggtc660
ctgctggagttcgtgaccgccgccgggatcactctcggcatggacgagctgtacaagtaa720
<210>3
<211>21
<212>dna
<213>人工合成(artificialsynthesis)
<400>3
ccagaacaacgaggactaccg21
<210>4
<211>19
<212>dna
<213>人工合成(artificialsynthesis)
<400>4
gtcaccaccgacttgagcc19
<210>5
<211>20
<212>dna
<213>人工合成(artificialsynthesis)
<400>5
gctatgtacgtcgccatcca20
<210>6
<211>21
<212>dna
<213>人工合成(artificialsynthesis)
<400>6
ggacagtgtggctgacaccat21
<210>7
<211>41
<212>dna
<213>人工合成(artificialsynthesis)
<400>7
ccggggatcctctagaatggtgagcaagggcgaggagctgt41
<210>8
<211>41
<212>dna
<213>人工合成(artificialsynthesis)
<400>8
ggccagtgccaagcttttacttgtacagctcgtccatgccg41
<210>9
<211>41
<212>dna
<213>人工合成(artificialsynthesis)
<400>9
ccatgattacgaattcgggtgacggcaggtactgatatata41
<210>10
<211>41
<212>dna
<213>人工合成(artificialsynthesis)
<400>10
tgctcaccattctagattacttgtacagctcgtccatgccg41
<210>11
<211>20
<212>dna
<213>人工合成(artificialsynthesis)
<400>11
ccaggctttacactttatgc20
<210>12
<211>20
<212>dna
<213>人工合成(artificialsynthesis)
<400>12
gcgattaagttgggtaacgc20
- 还没有人留言评论。精彩留言会获得点赞!