一种针对EGFRL858R基因突变的特异性T细胞受体及其应用的制作方法
本发明涉及基因工程及肿瘤免疫治疗领域,具体涉及一种针对egfrl858r基因突变的特异性t细胞受体及其应用。
背景技术:
肿瘤免疫治疗是通过重新启动并维持肿瘤-免疫循环,恢复机体正常的抗肿瘤免疫反应,从而控制与清除肿瘤的一种治疗方法。tcr(tcellreceptor)疗法即t细胞受体疗法,在该疗法中,分离出内源的t细胞,加以工程化改造,并输注回人体。这样一来,带有靶向癌细胞能力的t细胞数量将大增。这种方法除了能像细胞毒性化疗和靶向治疗快速杀灭肿瘤外,还避免了疫苗和免疫检查点抑制剂疗法的延迟效应。tcr-t是继肿瘤手术、放疗、化疗、靶向治疗之后的最有发展前景的肿瘤治疗技术之一,无论是国际还是国内都是目前肿瘤免疫治疗的研究热点。
t细胞受体(简称tcr)是所有t细胞表面的特征性标志,具有识别抗原呈递细胞(apc)上人类主要组织相容性复合体(mhc)分子-抗原肽复合体的能力。tcr是由α、β两条肽链构成的异二聚体,每条肽链分为可变区(v区)、恒定区(c区)、跨膜区和胞质区等几部分;其胞质区很短,信号传递主要通过与其以非共价键结合的cd3分子进行。tcr分子属于免疫球蛋白超家族,其抗原特异性存在于v区;v区又各有三个高变区cdr1、cdr2、cdr3,其中以cdr3变异最大,直接决定了tcr的抗原结合特异性。在tcr识别mhc-抗原肽复合体时,cdr3可直接与抗原肽相结合。
tcr在识别apc或者靶细胞上的mhc分子所提呈的抗原肽时,既要识别抗原肽,同时也要识别自身mhc分子的多态性部分,此现象即mhc限制性。t细胞通过特异性t细胞受体识别肿瘤细胞表面抗原肽-mhc复合体而被激活,活化后的t细胞可以直接溶解肿瘤细胞,或通过分泌细胞因子如干扰素、肿瘤坏死因子等抑制肿瘤生长。cd8+t细胞介导的特异性mhc-i类分子限制细胞免疫功能,在抗肿瘤免疫中尤其重要。
hla(humanlymphocyteantigen,人类淋巴细胞抗原),受控于mhc的基因簇,是具有高度多态性的同种异体抗原,是目前所知人体最复杂的遗传多态性系统,有几十个基因座位,每个基因座位又有几十个等位基因,且呈共显性表达。hla有a、b、c、d和dr5个位点,分别称为hla-a,hla-b,hla-c,hla-d和hla-dr。
egfr(epidermalgrowthfactorreceptorfamily)人类表皮生长因子受体家族属于酪氨酸激酶受体家族,也被称作her家族或erbb家族。egfr信号通路对细胞的生长、增殖和分化等生理过程发挥重要的作用。egfr等蛋白酪氨酸激酶功能缺失或其相关信号通路中关键因子的活性或细胞定位异常,均会引起肿瘤、糖尿病、免疫缺陷及心血管疾病的发生。egfr诱导癌症至少通过3种机制:egfr配体的过表达、egfr的扩增或egfr的突变活化,其中以egfr的突变活化为主要机制。egfr基因是由28个外显子组成,其中外显子18-21是常见的基因突变位点,也是与易瑞沙等egfr-tki结合的位点。通常来说,egfr的外显子19缺失和外显子21的l858r两个突变对于易瑞沙靶点是敏感的。因此egfr的外显子19缺失和外显子21的l858r突变可以作为肿瘤患者是否适用egfr-tki的一个重要的判断标准,其中l858r基因突变,也就是egfr蛋白的第858个氨基酸从l(亮氨酸)突变成了r(精氨酸)。
肿瘤细胞会发生大量基因突变,有些突变会改变氨基酸编码序列,导致肿瘤细胞表达正常细胞所没有的异常蛋白。这些异常蛋白在细胞内(肿瘤细胞或抗原呈递细胞)被蛋白酶酶解成肽段(抗原表位),与mhc-i类或mhc-ii类分子高亲和力结合,并以复合物形式呈递到细胞表面,与t细胞受体结合,t细胞被激活;激活后的t细胞扩增,浸润肿瘤微环境,识别并杀伤肿瘤细胞。这种肿瘤细胞特异性的异常蛋白称为的新抗原(neoantigen)。
tcr-t细胞治疗包括肿瘤特异性tcr的选取、tcr表达载体构建、tcr-t改造后细胞回输、免疫进程监测等关键技术和治疗手段。与同类型的car-t细胞治疗技术相比,tcr-t具有更广泛抗原选择空间,因此,不仅能扩充其适用肿瘤范围,还有望减少脱靶效应。但由于tcr-t的研发仍在初级阶段,针对特定突变和相应分型患者人群的有效tcr仍然非常匮乏。同时,tcr-t的有效患者人群以及新型构造的tcr-t的治疗效果还有待验证。
技术实现要素:
本发明旨在一定程度上解决上述相关技术中的技术问题之一。因此,本发明的目的在于提供一种针对egfrl858r基因突变的特异性tcr及其应用。
本发明的目的在于提供一种针对egfrl858r基因突变的特异性t细胞受体及其应用。
本发明提供一种针对egfrl858r基因突变的特异性t细胞受体,所述t细胞受体能够与kitdfgrak-hla-a*1101复合物结合。
其中,所述t细胞受体包含α链和β链,所述α链cdr3区为与seqidno:02序列具有至少70%、至少具有75%、至少具有80%、至少具有85%、至少具有90%、至少具有95%、至少具有96%、至少具有97%、至少具有98%或至少具有99%同源性且具有相同功能的氨基酸序列。
进一步,所述α链cdr3区的氨基酸序列为seqidno:02所示的序列。
进一步,所述α链cdr3区的核苷酸序列为seqidno:01所示的序列。
其中,所述α链的可变区序列包含cdr3,所述α链的可变区序列的氨基酸序列为seqidno:06所示的序列。
进一步,所述α链的可变区序列的核苷酸序列为seqidno:05所示的序列。
其中,所述t细胞受体β链cdr3区为与seqidno:04序列具有至少70%、至少具有75%、至少具有80%、至少具有85%、至少具有90%、至少具有95%、至少具有96%、至少具有97%、至少具有98%或至少具有99%同源性且具有相同功能的氨基酸序列。
进一步,β链cdr3区的氨基酸序列为seqidno:04所示的序列。
进一步,β链cdr3区的核苷酸序列为seqidno:03所示的序列。
所述β链的可变区序列包含cdr1、cdr2和cdr3,其中所述β链的cdr3区的核苷酸中,所述β链的可变区序列包含cdr3,所述可变区序列的氨基酸序列为seqidno:08所示的序列。
进一步,所述β链的可变区序列的核苷酸序列为seqidno:07所示的序列。
其中,所述t细胞受体为α链和β链异源二聚体结构,所述α链和β链的恒定区序列,可以为人源或鼠源恒定区序列。
本发明提供一种载体,所述载体含有上述tcr序列的核酸分子或其氨基酸序列对应的密码子优化的核酸分子。
优选的,所述载体为病毒载体。
更优选的,所述载体为慢病毒载体。
本发明还提供一种细胞,所述细胞在细胞膜表达所述的t细胞受体。
优选的,所述细胞为t细胞。
更优选的,所述细胞为cd8阳性t细胞。
其中,所述t细胞对egfrl858r突变的hla-a*1101肿瘤细胞有特异性杀伤作用。
一种t细胞在制备抗肿瘤药物或药物组合领域的应用。
所述的t细胞可用于制备t细胞受体。
所述的t细胞受体、所述的表达tcr的细胞用于制备能够特异性杀伤egfrl858r突变的hla-a*1101肿瘤细胞的药物或药物组合中。
所述抗肿瘤药物或药物组合,包含所述的tcr或表达tcr的t细胞。
所述抗肿瘤药物或药物组合用于治疗hla-a*1101患者。
所述抗肿瘤药物及药物组合,用于治疗涉及表达egfrl858r突变的恶性肿瘤。
所述肿瘤包括肺癌、中枢神经肿瘤、结直肠癌、胃癌、子宫内膜癌等。
一种治疗癌症的方法,对哺乳动物给予治疗上有效量的所述的制备抗肿瘤药物或药物组合。
所述治疗癌症的方法,包括向受试者给予表达tcr的细胞、化疗药物、靶向治疗药物、免疫检查点抑制剂类药物中的一种或几种。
所述一种治疗癌症的方法,可以同时或以任何顺序依次给予表达tcr的细胞、化疗药物、靶向治疗药物、免疫检查点抑制剂类药物中的一种或几种。
本发明的有益效果是:
通过输注能够识别特异靶标的基因修饰t细胞,赋予免疫系统以新的非自然免疫活性。
应用本发明提供的tcr或t细胞制备的抗肿瘤药物及药物组合,具有特异性强,个体化治疗效果好的特点。
附图说明
为了更清楚地说明本发明实施例或现有技术中的技术方案,下面将对实施例或现有技术描述中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图仅仅是本发明的一些实施例,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他的附图。
图1a流式细胞术分析未转染tcr的t细胞;
图1b流式细胞术分析转染tcr的t细胞;
图2a流式细胞术分析未转染hla-a*1101的t2细胞;
图2b流式细胞术分析转染hla-a*1101的t2细胞;;
图3elisa检测tcr-t细胞与靶细胞孵育后ifn-γ表达;
图4elisa检测tcr-t细胞与靶细胞孵育后il-2的表达;
图5荧光素酶法检测特异性tcr-t细胞对靶细胞的杀伤比例。
具体实施方式:
实施例1tcr慢病毒制备
将tcrα链和β链的编码序列通过弗林蛋白酶切割位点、sgsg接头和f2a序列连接,全合成α链和β链基因,应用限制性内切酶ecori和bamhi将tcr基因克隆至慢病毒表达载体pcdh(购自sbi)中,得到重组质粒,该重组质粒能表达序列6和8所示的tcrα链和β链可变区氨基酸序列。
将重组质粒转化xl-10感受态细胞,均匀涂布与含氨苄青霉素的lb固体培养基平板上,37℃培养12h后,挑取单个菌落至含氨苄青霉素的lb液体培养基中,于37℃、220rpm/min震荡培养14-16h,抽提质粒。
重组质粒的包装:取对数生长期293t细胞(购自中国医学科学院基础医学研究所基础医学细胞中心)作为包装细胞,接种到含有培养基(含10%fbs的dmem培养基)的t25细胞培养瓶中,细胞汇合度达到80-90%时进行转染。将重组质粒与混合包装载体质粒ppackh1(购自sbi)混匀,移液枪上下吹打充分混匀后,加入500μl1μg/μl脂质体转染试剂lip2000(invitrogen,11668-027),立即用移液器上下吹打混匀,室温下静置10-15分钟。将上述dna/脂质体复合物逐滴加入培养皿中,充分混匀。将培养皿置于37℃、5%co2培养箱,培养6-8小时后,将含有转染试剂的培养基去掉,更换为新鲜的完全培养基。48小时后,收集培养液至无菌离心管中,4℃、2000g离心10分钟,将上清液通过0.45µm的pes材质的滤膜过滤,取一个新的无菌超速离心管,加入含有病毒的培养基上清(即过滤液),4℃、20,000g离心3小时。小心将离心管中的液体吸去,所得沉淀即为目的慢病毒,加入1mlpbs缓冲液将沉淀重悬,即得到慢病毒悬液,置于-80℃保存。
实施例2tcr-t细胞的制备及流式细胞术分析tcr-t细胞的tcr表达
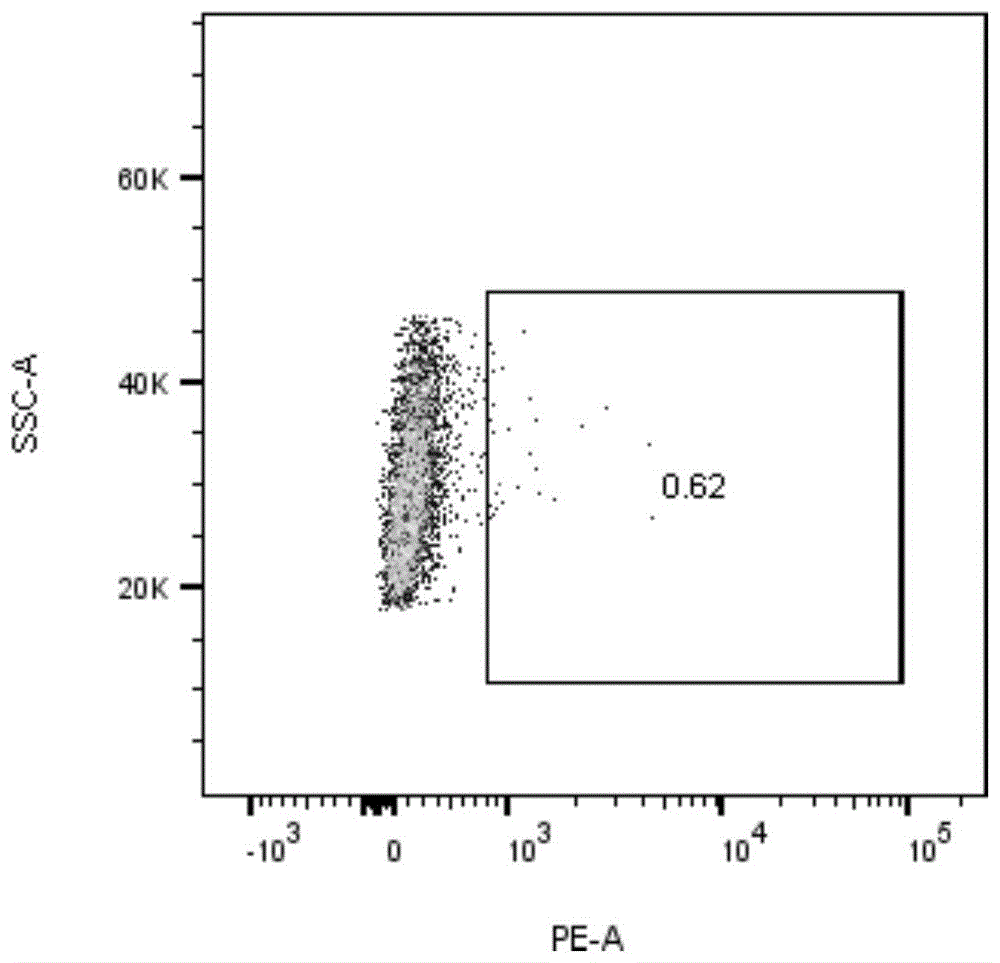
取健康志愿者外周血,使用淋巴细胞分离液(美国stemcell,07861)分离得到人外周血单核细胞(pbmc)。将dynabeads(gibco,11141d)和pbmc混合,室温孵育20min分离活化t细胞。向t细胞中加入3mlx-vivo15培养基(lonza,dl-201),用移液器重悬细胞和dynabeads(thermo,11141d)混合物,并调整细胞密度至0.5~1×106个细胞/ml,得到细胞悬液。将细胞悬液置于37℃、5%co2培养箱中连续培养48h后,将细胞密度调整至1x106个/ml。从-80℃超低温冰箱中取出慢病毒,迅速在37℃水浴锅中解冻,向准备好的t细胞中,加入polybrene(santacruz,sc-134220)至终浓度为6μg/ml,加入上述慢病毒,轻轻吹打充分混匀,800g室温离心1h。然后置于37℃、5%co2的培养箱中,继续培养24小时,去掉含有病毒的培养基上清,用新鲜培养基重悬细胞沉淀,将细胞转移至新的培养器皿中,继续培养,第4天时通过facs检测细胞tcr的表达来指示tcr-t细胞的阳性率,见图1。其中,图1a中为未转染tcr的t细胞,图1b中为转染tcr的t细胞,对比可获得tcr转染效率为15%。
实施例3靶细胞的制备
构建过表达hla-a*1101分子的慢病毒表达载体pcdh-a1101,瞬时转染293t细胞,制备重组慢病毒,方法同实施例1。病毒转导t2细胞系,按照实施例2的方法,构建表达hla-a*1101的t2细胞系。使用hla分子的抗体进行标记后,采用facs检测t2细胞表面hla-a*1101的表达,见图2。图2a中为未转染hla-a*1101的t2细胞,图2b中为转染hla-a*1101的t2细胞,对比可获得hla-a*1101的转染效率为96.6%。
实施例4细胞因子测定法检测特异性tcr-t细胞对靶细胞的杀伤作用
t2细胞系缺乏与抗原处理(tap)相关的转运体,故可以有效地负载外源肽,作为抗原呈递细胞,将加载的抗原肽提呈给t细胞识别。将人工合成的突变型抗原肽kitdfgrak和野生型肽(kitdfglak)与实施例3中得到的t2细胞在37℃、5%co2条件下孵育24h(多肽的浓度为50μg/ml、t2细胞的浓度为1×106个/ml),洗涤去除未结合的抗原肽,然后收集细胞,即为负载抗原肽的t2细胞。
特异性tcr-t细胞与负载抗原肽kitdfgrak的t2细胞在37℃、5%co2条件下孵育24h,靶细胞浓度均为2×104个/ml,效靶比(1:1,2.5:1,5:1,10:1)。对照细胞为未负载抗原肽的t2细胞和负载阴性对照抗原肽(kitdfglak)的t2细胞。elisa检测共培养体系中细胞因子ifn-γ和il-2的表达(humanifn-γelisakit,humanil-2elisakit购自达科为)。评估特异性tcr-t细胞对靶细胞的杀伤作用,见图3,图4。图3显示tcr-t与t2靶细胞共培养后,突变抗原肽孵育的t2细胞可特异性刺激tcr-t分泌ifn-γ,野生型多肽孵育的t2细胞和单独t2细胞不能引起tcr-t分泌ifn-γ。表明该tcr可以特异性识别抗原肽kitdfgrak,转染该tcr的t细胞识别靶细胞后,能够分泌ifn-γ,在效靶比为10:1时,分泌量可达505pg/ml,从而起到对靶细胞的杀伤作用。图4显示tcr-t与t2靶细胞共培养后,突变抗原肽孵育的t2细胞可特异性刺激tcr-t分泌il-2,野生型多肽孵育的t2细胞和单独t2细胞不能引起tcr-t分泌il-2。表明该tcr可以特异性识别抗原肽kitdfgrak,转染该tcr的t细胞识别靶细胞后,能够分泌il-2,在效靶比为10:1时,分泌量可达339pg/ml,从而起到对靶细胞的杀伤作用。
实施例5荧光素酶法检测特异性tcr-t细胞对靶细胞的杀伤作用
以实施例3构建靶细胞株为基础,将带有荧光素酶的慢病毒转染靶细胞,筛选出稳定表达荧光素酶的靶细胞进行体外共培养实验。荧光素酶催化荧光素底物的氧化反应发光,只有活细胞才会产生发光现象,且发光光强度与细胞的数目线性相关。通过荧光强度检测靶细胞数量,反应tcr-t对靶细胞的杀伤作用。
使用上述制备的tcr-t细胞及表达荧光素酶的靶细胞,建立共培养体系,按照不同的效靶比(1:1,2.5:1,5:1,10:1),连续共培养24小时;以未转染的对照t细胞作为对照。使用萤光素酶检测试剂盒(one-glo™luciferaseassaysystempromegae6110)检测检测共培养体系中存活的肿瘤细胞荧光素酶活性,吸除细胞培养液,pbs轻柔的冲洗细胞,加入100μl裂解液。室温条件下,轻摇细胞15min以使细胞完全溶解,cytation3全波长检测荧光强度。计算tcr-t对靶细胞的杀伤百分比。并利用未转染tcr的t细胞作为对照。
如图5所示,结果表明获得的特异性tcr-t细胞可以特异性杀伤负载抗原肽kitdfgrak的t2靶细胞,在效靶比10:1时杀伤效率可达72%。
以上对本发明所提供的一种针对egfrl858r基因突变的特异性tcr及其应用进行了详细介绍。本文中应用了具体个例对本发明的原理及实施方式进行了阐述,以上实施例的说明只是用于帮助理解本发明的方法及其核心思想。应当指出对本领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以对本发明进行若干改进和修饰,这些改进和修饰也落入本发明权利要求的保护范围内。
序列表
<110>天津亨佳生物科技发展有限公司
<120>一种针对egfrl858r基因突变的特异性t细胞受体及其应用
<160>8
<210>1
<211>42
<212>dna
<213>人工序列
<220>
<223>tcrα链cdr3区
<400>1
tgtgctgtggtactatcaggaggaagctacatacctacattt42
<210>2
<211>14
<212>prt
<213>人工序列
<220>
<223>tcrα链cdr3区
<400>2
cysalavalvalleuserglyglysertyrileprothrphe
1510
<210>3
<211>48
<212>dna
<213>人工序列
<220>
<223>tcrβ链cdr3区
<400>3
tgcagtgctagtggggaactagcgggagcagacaatgagcagttcttc48
<210>4
<211>16
<212>prt
<213>人工序列
<220>
<223>tcrβ链cdr3区
<400>4
cysseralaserglygluleualaglyalaaspasngluglnphephe
151015
<210>5
<211>402
<212>dna
<213>人工序列
<220>
<223>tcrα链可变区
<400>5
atgctcctggagcttatcccactgctggggatacattttgtcctgagaactgccagagcc60
cagtcagtgacccagcctgacatccacatcactgtctctgaaggagcctcactggagttg120
agatgtaactattcctatggggcaacaccttatctcttctggtatgtccagtcccccggc180
caaggcctccagctgctcctgaagtacttttcaggagacactctggttcaaggcattaaa240
ggctttgaggctgaatttaagaggagtcaatcttccttcaatctgaggaaaccctctgtg300
cattggagtgatgctgctgagtacttctgtgctgtggtactatcaggaggaagctacata360
cctacatttggaagaggaaccagccttattgttcatccgtat402
<210>6
<211>134
<212>prt
<213>人工序列
<220>
<223>tcrα链可变区
<400>6
metleuleugluleuileproleuleuglyilehisphevalleuarg
151015
thralaargalaglnservalthrglnproaspilehisilethrval
202530
sergluglyalaserleugluleuargcysasntyrsertyrglyala
354045
thrprotyrleuphetrptyrvalglnserproglyglnglyleugln
505560
leuleuleulystyrpheserglyaspthrleuvalglnglyilelys
65707580
glypheglualagluphelysargserglnserserpheasnleuarg
859095
lysproservalhistrpseraspalaalaglutyrphecysalaval
100105110
valleuserglyglysertyrileprothrpheglyargglythrser
115120125
leuilevalhisprotyr
130
<210>7
<211>399
<212>dna
<213>人工序列
<220>
<223>tcrβ链可变区
<400>7
atgctgctgcttctgctgcttctggggccaggctccgggcttggtgctgtcgtctctcaa60
catccgagcagggttatctgtaagagtggaacctctgtgaagatcgagtgccgttccctg120
gactttcaggccacaactatgttttggtatcgtcagttcccgaaaaagagtctcatgctg180
atggcaacttccaatgagggctccaaggccacatacgagcaaggcgtcgagaaggacaag240
tttctcatcaaccatgcaagcctgaccttgtccactctgacagtgaccagtgcccatcct300
gaagacagcagcttctacatctgcagtgctagtggggaactagcgggagcagacaatgag360
cagttcttcgggccagggacacggctcaccgtgctagag399
<210>8
<211>133
<212>prt
<213>人工序列
<220>
<223>tcrβ链可变区
<400>8
metleuleuleuleuleuleuleuglyproglyserglyleuglyala
151015
valvalserglnhisproserargvalilecyslysserglythrser
202530
vallysileglucysargserleuasppheglnalathrthrmetphe
354045
trptyrargglnpheprolyslysserleumetleumetalathrser
505560
asngluglyserlysalathrtyrgluglnglyvalglulysasplys
65707580
pheleuileasnhisalaserleuthrleuserthrleuthrvalthr
859095
seralahisprogluaspserserphetyrilecysseralasergly
100105110
gluleualaglyalaaspasngluglnphepheglyproglythrarg
115120125
leuthrvalleuglu
130
- 还没有人留言评论。精彩留言会获得点赞!