花生株型相关基因位点的分子标记方法及其应用与流程
技术领域:
本发明属于分子遗传学和分子育种技术领域,涉及花生侧枝角度、生长习性和株型相关基因的分子标记方法,用于栽培花生侧枝角度的分子标记辅助改良以及分子标记辅助的花生株型育种和种质资源的利用。
背景技术:
花生(arachishypogaeal.2n=4x=40,aabb异源四倍体)富含食用油脂(约38~60%)和蛋白质(约24~36%),同时含有多种维生素及钙、铁等微量元素,是半干旱温热地区许多国家的重要食品和油料作物。分子生物技术的进步,推动花生的分子遗传研究从最初重点关注的含油量、品质、抗病等方面,发展向多种农艺性状方面(janila,nigametal.2013)。其中花生的株型是一个值得重点关注的方面,它关系着花生的群体产量、耕作方式以及收获机械化的难易等多方面问题。花生的株型可以直观地分为:匍匐型、直立型和半匍匐型三种,这一划分取决于直立的主茎之外的侧枝的生长角度。野生花生(大多数为二倍体,仅monticola为异源四倍体)均为完全匍匐生长,各级侧枝贴伏地面,且末端不上翘。常见的栽培花生(异源四倍体)为直立或半匍匐生长,普遍认为是从匍匐生长的二倍体野生花生天然异交加倍后的异源四倍体野生种arachismonticola直接驯化而来(seijo,laviaetal.2007),其株型的改变是一个主要的驯化改良事件。我国早期栽培花生多数属于半匍匐型(亦被称为半蔓型),典型的代表是我国特有的龙生型花生以及早期北方主要种植区的姜格庄半蔓、蓬莱一窝猴等(半匍匐型)。后来,随着密植和收获简易化的要求,逐渐发展为以伏花生为代表的直立型为主,直立型伏花生是从半蔓型花生中民间选育而成。当下国内审定的品种绝大多数为直立型,直立型花生的适当密植可以充分发挥肥沃土地的承载能力,同时荚果相对集中便于人工收获。与中国不同,美国花生的主栽品种多数为半匍匐型,代表性品种包括florunner和tifrunner(albaghdadi,durhametal.2007),这种匍匐型株型便于其采用分段式机械收获时的反扣晾晒,虽然其单产低于我国,但半匍匐花生的用种量较少和机械化收获的低廉成本使美国花生的成本一直低于我国。我国根据自身的实际情况,在理论上,也提出了花生高产的理想株型,其中强调了直立型是花生群体产量保证的理想株型(李新国,郭峰等.2013),开发研制出一系列直立花生的收获机械,大大提高了花生生产的机械化程度。在实际生产中,为了增加植株密度导致花生的用种量较高,每亩地平均需要近20kg的籽仁,成本超过200元人民币。近年,花生的种植向新疆、辽宁、吉林等大片机械化农田发展,也对收获期的机械化和收获后的及时晾晒提出了要求。根据不同的生产情况,选育适当株型的品种是花生生产的重要方面。鉴于此,发掘花生株型中侧枝角度的关键遗传位点,将为分子遗传手段改良花生的株型和选育适合的花生品种提供理论依据,同时可以为双子叶植物的侧枝生长角度的基础研究提供新的参考。
侧枝的生长角度决定着整个植株的株型,对于多数植物来说,侧枝一般是非垂直的,便于群体内获得更多的阳光、空气等资源。关于侧枝角度的遗传控制在模式植物中已经有十分丰富的研究(roychoudhryandkepinski2015)。早期,已经有人研究将重力决定的侧枝角度(gravitropicset-pointangle,gsa)与向地性生长区分开(digbyandfirn1995)。向(背)重力性决定侧枝角度是一类由生长素的非对称性分布调节的,生长素的合成相关基因以及其信号通路tir1/afb-aux/iaa-arf在对侧枝角度的调控中已被比较详细的诠释(roychoudhry,delbiancoetal.2013)。另外,通过突变体等手段另外有多个相关的基因被发现和验证,其中包括三个转录因子idd14、idd15和idd16,协调控制生长素的合成和运输,其中idd15的失活能够显著的增加侧枝的角度(dayong,jingboetal.2013)。除了模式植物拟南芥,在多种作物中也通过正向遗传学的手段,获得了多个控制侧枝或分蘖角度的基因。水稻中利用一个散生型的突变体与直立生长的水稻品种回交多次获得的导入系la1-zf802与多个普通水稻杂交构建的f2精细定位并克隆了水稻散生基因lazy1,这一基因是一个单子叶草本植物中特异的基因,它在生长素的极性运输中发挥负调节作用,去功能化的lazy1基因背景下,生长素的极性运输增强,降低了分蘖的向重力性,进而导致分蘖角度增大(li,wangetal.2007)。玉米中分蘖角度的图位克隆工作也发现一个位点,其对应的功能基因与水稻lazy1同源的基因zmlazy1具有类似的功能(zhaobin,chuanetal.2013)。玉米中还发现了一个控制雄蕊分支角度的基因bad1,它是一个tcp转录因子,在侧枝基部的叶枕形成中发挥作用(fang,renataetal.2012)。水稻中另外一个控制分蘖角度的主效qtl位点是tac1,tac1表达量的降低可以使水稻的分蘖接近垂直生长,它在高纬度地区的粳稻密植中发挥了重要的作用(yu,linetal.2007)。另外,它的同源基因在其他单子叶草本植物芒草中也发挥具有类似的功能(zhao,huaietal.2014)。对于野生匍匐水稻与栽培水稻株型差异进行的遗传定位也发现了一个控制水稻分蘖角度的主效qtl位点prog1,它是匍匐野生水稻驯化为直立栽培水稻的一个关键位点,编码一个c2h2型锌指蛋白,去功能化的prog1可以使水稻从匍匐生长走向直立生长(jin,huangetal.2008,tan,lietal.2008)。独角金内酯作为新发现的一种植物内源激素,在侧枝角度的调节中也发挥了重要的作用,其合成基因突变体sols可以通过降低生长素的合成,抑制水稻分蘖的向地性,部分地恢复lazy1突变体的松散的表型(dajun,dongqinetal.2014)。最近在油菜的侧枝角度研究中,利用关联分析和连锁群体定位的方式也发现了多个控制侧枝角度的位点(liu,wangetal.2016,sun,wangetal.2016),其中利用极端群体法和关键区域连锁定位的策略定位到了一个候选基因bnayucca6,功能预测是一个生长素合成途径中的一个关键基因,在侧枝角度大小不同的材料间存在多个snp差异,进一步研究推断是该基因表达量的差异在发挥作用(hui,chengetal.2016)。
栽培花生是由两个不同基因组类型的二倍体野生种天然异交加倍后驯化而来,普遍认为异源四倍体野生种a.monticola是其直接原始种(seijo,laviaetal.2007)。monticola及其二倍体供体野生种均是完全匍匐生长型。早期对于花生株型的遗传研究由于取材不同,所得的结果也不尽相同。较早的对花生株型的研究是balaiah,c.等研究认为直立型对匍匐型为显性(balaiah,reddyetal.1977)。但后来较多的研究认为匍匐型对直立型为显性或不完全显性,例如:蒋复和张俊武研究发现匍匐与直立品种杂交,f1为不完全显性的半匍匐,后代f2分离为直立、半匍匐和匍匐三种类型(蒋复和张俊武1982)。甘信民和曹玉良等研究认为,匍匐相对于直立为显性,f2代的分离符合3匍匐:1直立的单基因控制模型(甘信民,曹玉良等.1984)。已有的研究仅限于此,还未见对以花生侧枝角度为代表的花生株型的较为系统及分子遗传方面的研究。
技术实现要素:
本发明的目的是提供一种对花生侧枝角度、生长习性和株型相关基因位点的分子标记方法,通过检测与花生侧枝角度相关基因位点连锁的分子标记,可以确定控制花生侧枝角度等位基因位点的选择;提供花生株型育种的分子标记辅助选择的室内检测和表型鉴定筛选的可操作性;提高性状的选择效率、加快育种进度。
为了实现上述目的,本发明所采用的技术方案是:
本发明提供一种用于鉴别花生侧枝角度、生长习性和株型相关基因位点的匍匐型和直立型等位变异的caps分子标记lba5b-caps,所述分子标记lba5b-caps匍匐型等位变异的核苷酸序列如seqidno:1所示;所述分子标记lba5b-caps直立型等位变异的核苷酸序列如seqidno:2所示;分子标记lba5b-caps的引物对为:
上游lba5b-caps-f:5`-aggaaagattgagatgaaaa-3`如seqidno:3所示;
下游lba5b-caps-r:5`-attgatgactaattgatggc-3`如seqidno:4所示。
本发明还提供一种花生侧枝角度、生长习性和株型相关基因位点的caps分子标记lba5b-capsd的方法:以花生基因组dna为模板,利用上下游引物进行pcr扩展,对扩增产物进行限制性内切酶scai的消化,之后通过琼脂糖电泳分离检测消化后的产物;如电泳分离检测结果显示pcr产物被切成两个小片段,则说明该个体存在lba5b增效等位变异基因位点,如果pcr产物不能被切开,则说明该个体不存在lba5b增效变异基因,而存在降低侧枝与主茎间角度的减效等位变异lba5b。
本发明所述的pcr扩展反应为:pcr体系为25ul,其中2×taqmastermix(dyeplus)12.5ul,模板1ul,10um上下游引物各1ul,加水至25ul;pcr反应程序为dna95℃预变性5min;95℃变性30s,55℃退火15s,72℃延伸30s,循环35次;最后72℃延伸10min。
本发明所述的dna限制性内切酶为saci。
本发明所述的限制性内切酶消化反应为:体系为25ul,其中pcr产物5ul,10×hbuffer2.5ul,scaⅰ1ul,加水至25ul,pcr仪37℃保温1h后加入2.5ul10×loadingbuffer终止反应。
本发明所述的琼脂糖电泳检测方法为:利用加热或微波炉将1%的琼脂糖tae溶液融化后,制作水平电泳凝胶,再在凝胶中加入核酸燃料;将消化后的产物加入适量loadingbuffer后混匀加入上样孔中,在120v的电压下进行电泳。
本发明还提供一种鉴别花生侧枝角度、生长习性和株型相关基因位点两侧连锁的indel标记,包括上游indel标记pm15-id1对应的序列如seqidno:5所示,下游indel标记pm15-id2对应的序列如seqidno.6所示;
indel标记pm15-id1的引物对为:
上游pm15-id1-f:5`-gttgagggttagaatcgaaactg-3`如seqidno:7所示;
下游pm15-id1-r:5`-ggagaagaaggtcccaaaggtg-3`如seqidno:8所示;
indel标记pm15-id2的引物对为:
上游pm15-id2-f:5`-atccaaacctatcgaagctg-3`如seqidno:9所示;
下游pm15-id2-r:5`-aagagggagttgaaccgtaa-3`如seqidno:10所示。
本发明提供的鉴别花生侧枝角度、生长习性和株型相关基因位点两侧连锁的indel标记方法,用indel标记pm15-id1或pm15-id2的上下游引物扩增匍匐型花生(小红毛)与直立型花生(南阳花生)的杂交后代的基因组dna,对于indel标记pm15-id1如果能扩增出281bp则标志着匍匐型花生的增效等位变异位点的存在,否则扩增出293bp则标志着存在直立型花生的减效等位变异(见图3);对于indel标记pm15-id2如果能扩增出209bp的扩增片段,则标志着匍匐型花生的增效等位变异位点的存在,否则扩增出179bp则标志着存在直立型花生的减效等位变异;
本发明所述的用indel分别标记pm15-id1或pm15-id2对花生侧枝角度、生长习性和株型相关基因位点的选择效率均在95%以上,用indel同时标记pm15-id1和pm15-id2对花生侧枝角度、生长习性和株型相关基因位点的选择效率超过99%。
本发明具有以下有益效果:
本发明所提供的花生侧枝角度、生长习性和株型相关基因位点lba5b的分子标记方法,具有以下优点:
(1)本发明通过多个ril群体,在15号染色体末端得到花生侧枝角度主效qtl位点,并获得了共分离或高度紧密连锁的indel标记pm15-id1或pm15-id2以及共分离caps标记lba5b-caps;
(2)通过本发明分子标记定位的花生侧枝角度相关基因位点lba5b的位置明确、精细、鉴定方法快速简便,选择效率高,只需检测这些标记的扩增条带扩增,可以判断花生侧枝角度相关基因位点lba5b的增效变异存在与否,由此预测花生侧枝角度大小的表型,可以快速有目的的筛选花生侧枝角度较小的品种或品系,用于花生改良株型和增产的育种工作;这些标记与侧枝角度相关基因位点lba5b共分离或高度紧密连锁;
(3)辅助育种选择目标明确,节约成本,不受环境影响。在传统育种方法中,首先要收集具有侧枝角度较大的亲本与骨干亲本进行一系列杂交,回交,而且要对侧枝角度进行单株选择。对花生侧枝角度进行表型鉴定要在花生开花下针期之前,且尽量选择互相没有遮挡的单株为准。通过标记检测花生侧枝角度主效基因位点,可以在苗期就鉴定出侧枝角度较大的单株或角度较小的单株,选择保留下来需要的株型结构单株,淘汰其他单株,可以有效的控制育种规模、提高花生侧枝角度选择育种的进程和选择效率。
附图说明:
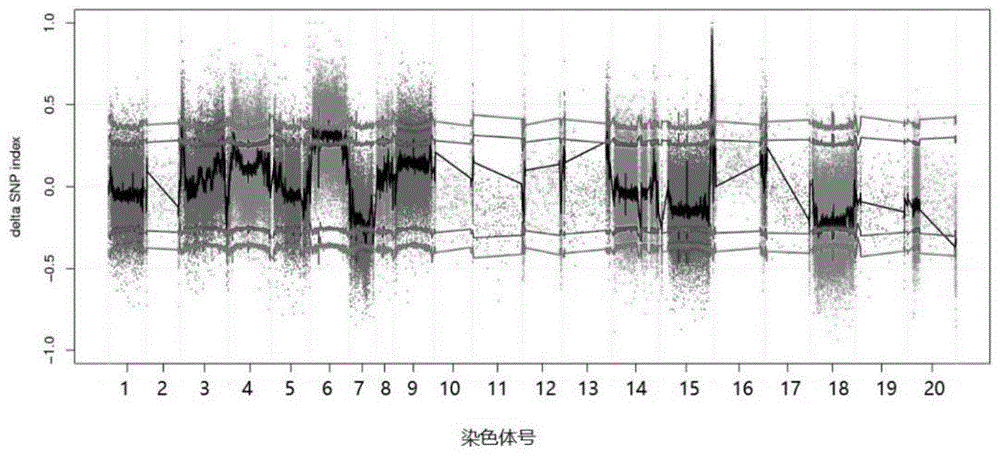
图1为本发明涉及的花生b05染色体(或称第15染色体)控制花生侧枝角度、生长习性和株型相关基因位点的bsa定位。
图2为本发明涉及的花生侧枝角度、生长习性和株型相关基因位点lba5b的连锁定位分析;所示j15-11和j15-12标记区别与pm15-id1和pm15-id2。
图3为本发明涉及的新开发的与lba5b紧密连锁的indel标记pm15-id1、pm15-id2聚丙烯酰胺凝胶分析。
图4为本发明涉及的lba5b-caps标记琼脂糖电泳条带,其中双带为lba5b等位变异,单条带为lba5b型等位变异。
具体实施方式:
下面通过实施例对本发明技术做进一步说明。
实施例1:花生侧枝角度、生长习性和株型相关基因位点的遗传定位
(一)材料与方法:
(1)植物材料和遗传群体的构建:匍匐型栽培花生小红毛与直立型栽培花生河南南阳花生杂交,种植杂交种,自交获得f2分离群体,连续多代株系自交繁育,最终形成f6代重组自交系hn-f6。用于主效与侧枝间角度相关基因位点的检测与定位。
(2)用ctab法提取个体dna(具体参照《分子克隆实验指南》)。
(3)多态性indel标记开发:根据亲本基因组的重测序数据与参考基因组的比较(tifrunner基因组数据:https://www.peanutbase.org/peanut_genome),开发了花生b05染色体的31对indel引物对,以小红毛与河南南阳花生的dna为模板,进行pcr扩增,筛选条带清晰,亲本间条带大小有差异的多态性标记,用于后续研究。
(4)indel标记的pcr反应体系及产物检测:pcr体系为25μl,其中2×taqmastermix(dyeplus)12.5μl,模板1μl,10μm上下游引物各1μl,补充加超纯水至25μl。pcr反应程序为dna95℃预变性5min;95℃变性30s,55℃退火15s,72℃延伸30s,循环35次;最后72℃延伸10min。在pcr仪上进行pcr扩增,扩增产物在8%非变性聚丙烯酰胺凝胶上进行电泳,凝胶通过硝酸银染色显影。
(5)利用hn-f6全群体个体的基因型资料构建花生的b05染色体的遗传图谱,所用的软件为qtlicimappingv4.1中的遗传图谱构建模块,设置lod值=3.0为阈值进行遗传图谱构建。利用该软件中定位模块进行基因定位。
(6)初定位采用bsa-seq的方法。
(二)结果与分析
(1)花生侧枝角度、生长习性和株型相关基因位点的初定位
从花生品种小红毛(匍匐型)与河南南阳花生(直立型)杂交、连续多代自交获得f6代重组自交系群体(hn-f6)中挑选30个匍匐家系和26个直立家系,对每个家系分别进行dna提取,对dna进行定量测定,将30个匍匐家系和26个直立家系的dna分别等量混合,构成两个极端个体池。将亲本小红毛和河南南阳花生,以及两个极端个体池交给公司进行基因组重测序。通过计算两个极端个体池的snp_index值的差(δsnp-index),将控制花生侧枝角度、生长习性和株型的相关位点定位在了花生b05染色体的末端153.8m-159.6m之间(见图1)。
(2)花生侧枝角度、生长习性和株型相关基因位点的精细定位和候选基因预测
根据亲本基因组重测序数据开发双亲本间可能有多态的31个indel标记,通过多态性分析获得了19个有多态的indel标记。利用相对均匀分布于花生b05(15号)染色体的12个indel多态性标记和hn-f6群体的192个家系,建立了花生b05染色体的分子标记遗传图谱(见图2)。对花生侧枝角度、生长习性和株型相关性状进行qtl分析发现:控制花生侧枝角度的qtl位于花生b05(15号)染色体末端,位于分子标记j15-11和j15-12之间,将其定名为lba5b(见图2)。
通过亲本转录组和后代代表个体的转录组分析发现:在j15-11和j15-12标记区间内存在一个符合表型差异的差异表达基因,将该基因暂时定为lba5b的候选基因。该基因在亲本及群体中表达差异显著,在匍匐亲本和子代中均高水平表达,在直立亲本和子代中均低水平表达。通过双亲间的比较发现:亲本间在该基因内部存在一个ta插入缺失差异。
实施例2:
(一)花生侧枝角度、生长习性和株型相关基因位点共分离标记lba5b-caps的开发与应用
根据实施例1的结果,将候选基因lba5b上的双亲间ta插入与否的差异开发为caps标记,将其命名为lba5b-caps(见图4)。利用caps标记lba5b-caps的上下游引物,扩增hn-f6群体各个个体基因组dna,经限制性内切酶sacⅰ消化后,在1%琼脂糖凝胶电泳上进行电泳检测,如果pcr产物被切成两个小片段,则说明该个体存在lba5b增效等位变异基因位点(来自小红毛),如果pcr产物不能被切开,则说明该个体不存在lba5b增效变异基因,而存在降低侧枝与主茎间角度的减效等位变异lba5b(来自南阳花生)。通过该分子标记对hn-f6群体进行等位变异分析和表型分析,发现该标记与后代侧枝角度、生长习性和株型表型共分离;
本实施例涉及的caps标记lba5b-caps的上下游引物为:
上游lba5b-caps-f:5`-aggaaagattgagatgaaaa-3`如seqidno:3所示;
下游lba5b-caps-r:5`-attgatgactaattgatggc-3`如seqidno:4所示;
本实施例涉及的caps标记lba5b-caps的pcr反应为:pcr体系为25ul,其中2×taqmastermix(dyeplus)12.5ul,模板1ul,10um上下游引物各1ul,加水至25ul;pcr反应程序为dna95℃预变性5min;95℃变性30s,55℃退火15s,72℃延伸30s,循环35次;最后72℃延伸10min;
本实施例涉及的限制性内切酶消化反应为:体系为25ul,其中pcr产物5ul,10×hbuffer2.5ul,scaⅰ1ul,加水至25ul,pcr仪37℃保温1h后加入2.5ul10×loadingbuffer终止反应;
本实施例涉及的琼脂糖电泳检测为:1%的琼脂糖tae溶液,利用加热或微波炉融化后,制作水平电泳凝胶,凝胶中加入核酸燃料。将消化后的产物加入适量loadingbuffer后混匀加入上样孔中,在120v的电压下进行电泳(该过程可以根据需要适当调整参数)。
(2)花生侧枝角度、生长习性和株型相关基因位点辅助选择标记的开发
根据亲本基因组重测序数据,在距离候选基因上游120kb和下游285kb左右各开发了一个相对更近的indel标记,分别命名为pm15-id1和pm15-id2;该两个标记在双亲间存在插入缺失差异;用indel标记pm15-id1或pm15-id2的上下游引物扩增匍匐型花生(小红毛)与直立型花生(南阳花生)的杂交后代的基因组dna,对于indel标记pm15-id1在匍匐亲本小红毛中扩增产物为281bp,在直立亲本南阳花生中的扩出条带大小为293bp。对于indel标记pm15-id2在匍匐亲本小红毛中扩增产物为209bp,在直立亲本南阳花生中能扩增出179bp的条带(见图3);利用该两个标记对hn-f6群体后代的进行标记型鉴定,同时结合表型分析,发现:标记pm15-id1与表型间的重组率低于1.0%,标记pm15-id2与表型间的重组率低于3.0%;
本实施涉及的indel标记pm15-id1的上下游引物为:
上游pm15-id1-f:5`-gttgagggttagaatcgaaactg-3`如seqidno:7所示;
下游pm15-id1-r:5`-ggagaagaaggtcccaaaggtg-3`如seqidno:8所示;
本实施涉及的indel标记pm15-id2的上下游引物为
上游pm15-id2-f:5`-atccaaacctatcgaagctg-3`如seqidno:9所示;
下游pm15-id2-r:5`-aagagggagttgaaccgtaa-3`如seqidno:10所示。
综合以上结果,通过检测与花生侧枝角度、生长习性和株型相关基因位点连锁或共分离的分子标记,能够确定控制花生侧枝角度、生长习性和株型的等位变异在育种后代中的分布,实现对花生株型改良中后代的分子标记辅助选择,提高花生株型改良的目的性和准确性,提高花生侧枝角度、生长习性和株型选择育种的效率、加快育种进度。
序列表
<110>青岛农业大学
<120>花生株型相关基因位点的分子标记方法及其应用
<130>20191001
<160>11
<170>siposequencelisting1.0
<210>1
<211>943
<212>dna
<213>人工序列(artificialsequence)
<400>1
aggaaagattgagatgaaaaggatagagaatgcaacaagccgtcaagtaacgttttcaaa60
gaggaggagtgggcttctaaagaaagcacatgagctttctgttctttgtgatgcacaaat120
tgctcttataatcttttcacaaagtgggaggctctttgaatactcaagtacttcagagta180
agcccttacttaatcactttattttcttgcttcatcaacattcatgttagtgttagtttc240
attctttaatttttattcttctcctaattaattagtacaatactactacttattattgaa300
ctattatatatatggtttaatcttggagaaactaaaggtttactttaatttgctcaaact360
atgatacatgtgttttgttgtgagttgtgctaagtcttcctcaaactacctagatttgat420
gatcaaagaacatggatctcattgaaaaaaaagtgtgtggctttccgtttatggtaagct480
aactactactggtaattacaaaagtatatacactaatttagaaatactcaacacttgcta540
aattgtagtagtagcatttggttgctgaataaattggaacttgaaatactctgctatgta600
attatttcaatgctctccgtatcaaataaggtaaaaaaaaaatggttttttaaaattttt660
tctaagatattatatagtgtatttttcattatacatgttttaagtgagattaagaaaaaa720
atatgtgaagagcaaagatatattgtagtattacaaaagaccatcaccttttgcaacatt780
acttaagaaaaaaaaaaacctaagattacctatttttatttgatgttgagtttagttctg840
caatctgctctgtactaattatatctttatcctaaatataacaaagaggagtgctaaggg900
gccagcagattttgtgagttgtagccatcaattagtcatcaat943
<210>2
<211>945
<212>dna
<213>人工序列(artificialsequence)
<400>2
aggaaagattgagatgaaaaggatagagaatgcaacaagccgtcaagtaacgttttcaaa60
gaggaggagtgggcttctaaagaaagcacatgagctttctgttctttgtgatgcacaaat120
tgctcttataatcttttcacaaagtgggaggctctttgaatactcaagtatacttcagag180
taagcccttacttaatcactttattttcttgcttcatcaacattcatgttagtgttagtt240
tcattctttaatttttattcttctcctaattaattagtacaatactactacttattattg300
aactattatatatatggtttaatcttggagaaactaaaggtttactttaatttgctcaaa360
ctatgatacatgtgttttgttgtgagttgtgctaagtcttcctcaaactacctagatttg420
atgatcaaagaacatggatctcattgaaaaaaaagtgtgtggctttccgtttatggtaag480
ctaactactactggtaattacaaaagtatatacactaatttagaaatactcaacacttgc540
taaattgtagtagtagcatttggttgctgaataaattggaacttgaaatactctgctatg600
taattatttcaatgctctccgtatcaaataaggtaaaaaaaaaatggttttttaaaattt660
tttctaagatattatatagtgtatttttcattatacatgttttaagtgagattaagaaaa720
aaatatgtgaagagcaaagatatattgtagtattacaaaagaccatcaccttttgcaaca780
ttacttaagaaaaaaaaaaacctaagattacctatttttatttgatgttgagtttagttc840
tgcaatctgctctgtactaattatatctttatcctaaatataacaaagaggagtgctaag900
gggccagcagattttgtgagttgtagccatcaattagtcatcaat945
<210>3
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>3
aggaaagattgagatgaaaa20
<210>4
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>4
attgatgactaattgatggc20
<210>5
<211>601
<212>dna
<213>人工序列(artificialsequence)
<400>5
gagagaagatactatttacgaaaataatattaatccttgaaataaggaaaaattaaaatt60
agagataagtattgttttggttcctaacgttgagggttagaatcgaaactgtctccaacg120
taattttttatttagaatcatctttaacgtttttttttatattaaaattatttttttaat180
aaaatatttttattttattactaaatttatccctatcttaataaaaaaaagaaaaggacc240
cgaggaaaaggggaaaaaaaaaggggaaaggacgtgcacgcatgtggaggaaaaggagaa300
aggggagctccggcagccgtcgtcgtgaattccagtgtcgtcgccgccacctttgggacc360
ttcttctccacctaacacgagccagccagagatcagagagaaagagagagagagtcggtg420
gaggggaggaggccgctgccatagccgccgccaccatcgcaacacctgcatcgtcgccgg480
tctgcccacgagccgccgccagaatcgcgaacagaggaagacgttgctgccgttcatgca540
tgcttctgccgccgtggagtgcgcactataacaatatagattattttttagggttataat600
g601
<210>6
<211>614
<212>dna
<213>人工序列(artificialsequence)
<400>6
acaagagaattaattggcttgtttggtatcaaactcttgaatacatctttgtgaattgca60
acttctgaaaatactatattctccttatcgctgccaaagataaattaacataaataaaaa120
tagctaaacaaatccacaacaccattaaaaaataatatacttattaagtttatttactta180
tccaccaatagattatcatacaagtatgttgtaattgtaagtttaatattatccaaacct240
atcgaagctgttagtttaaaagacatgtaatccactaaatataaaaaaaaaactgttact300
ataataataataataataataataataataataataataaaaactaaactacttattata360
tagtactcataatcaatgttgacaaaattgcatgcaaattcgtaaaaacgtacattttta420
cggttcaactccctcttttcgttgcgcttctgctgtacgagtgtgtagagtaggaattat480
acaaaagtgctgtgtaaaatgtaaactcttcagaatcgtgactttgccagaaaaaaaaat540
atctcaaaaagggtggtctaaaaccttgcttttcacttaaaaaaatgtacatactatata600
catactattatttt614
<210>7
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>7
gttgagggttagaatcgaaactg23
<210>8
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>8
ggagaagaaggtcccaaaggtg22
<210>9
<211>0
<212>dna
<213>人工序列(artificialsequence)
<400>9
<210>10
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>10
aagagggagttgaaccgtaa20
<210>11
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>11
atccaaacctatcgaagctg20
- 还没有人留言评论。精彩留言会获得点赞!