流感病毒蛋白与宿主蛋白CPSF30的相互作用在抑制癌细胞增殖中的应用的制作方法
本发明涉及生物技术和免疫学领域。更具体地,本发明涉及流感病毒蛋白与宿主蛋白CPSF30的相互作用在抑制癌细胞增殖中的应用。
背景技术:
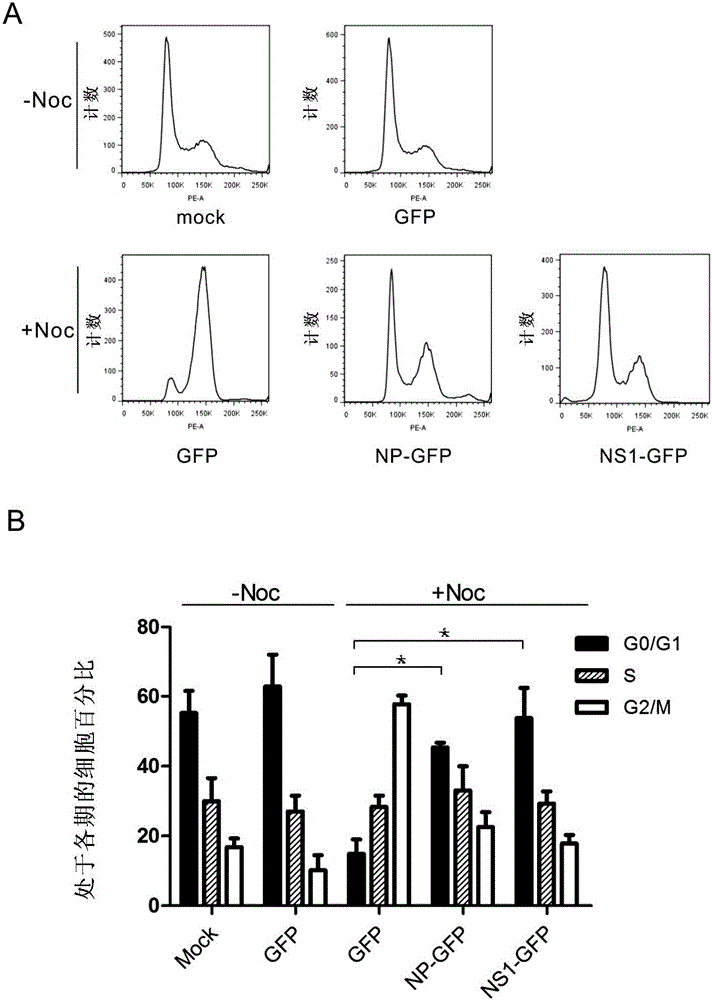
:癌症是威胁人类健康的重要疾病,全球每年死于癌症的人数比死于艾滋病、疟疾、结核的人数总和还要多,大概13%的人类死亡源于癌症,而这一比例正呈逐年上升趋势。在癌症类型、患病人口增多的情况下,研发新型有效的抗癌药物的重要性毋庸置疑。包括癌细胞在内的所有体细胞想要实现细胞增殖,都需要经历细胞周期到达有丝分裂,将1个细胞变成2个,实现细胞数量的增加。癌症的诱因有很多种,发生的部位也不尽相同,但是所有的癌症都具有一个共同的特征,就是癌细胞相比于正常细胞大多处于细胞分裂的活跃时期,表现为不受限制的、紊乱的细胞生长。因此,不论是传统的放射线治疗和化学治疗方法还是其他免疫治疗方法,都是为了抑制细胞分裂和增殖,乃至杀死癌细胞。病毒感染对正常机体来说是不利于健康的,但是人们对病毒学的研究发现,病毒通过和宿主的相互作用可以影响很多宿主信号通路和生理反应,这一点使得病毒也有被加以利用的可能,利用病毒的特性用来治疗病变的细胞。在癌症治疗中,利用病毒感染导致细胞死亡的原理杀死癌细胞的想法早在1940年左右出现,但直到1991年,才发现第一个溶癌病毒(Oncolyticvirus)疱疹病毒可以裂解脑胶质瘤细胞,诞生了第一个病毒疗法(Virotherapy)。随后,很多不同的病毒被用来研究其溶癌特性。然而,这种病毒疗法主要还是利用病毒感染最终导致细胞裂解的原理,虽然可以实现癌细胞感染特异性,但需要具有复制能力的活病毒,其安全性导致很大的应用限制。此外,目前一些常用的肿瘤治疗方法的副作用仍很大,导致病人甚至不得不中止治疗。综上,本领域尚未开发出令人满意的抑制癌细胞的药物,因此迫切需要开发安全性更好的抑制癌细胞的方法和药物。技术实现要素:本发明的目的就是提供一种新颖的、安全性更好的抑制癌细胞的方法和药物。在本发明的第一方面,提供了一种病毒蛋白、或其表达载体、或其激动剂的用途,用于制备(a)用于将细胞的细胞周期停滞于G0/G1期的组合物;和/或(b)用于抑制癌细胞生长的组合物。在另一优选例中,所述的病毒蛋白来自选自下组的病毒:流感病毒、副流感病毒。在另一优选例中,所述的病毒为流感病毒,更佳地为甲型流感病毒。在另一优选例中,所述的病毒蛋白选自下组:NP蛋白、NS1蛋白、或其组合。在另一优选例中,所述的细胞包括哺乳动物的细胞。在另一优选例中,所述的组合物包括药物组合物、膳食补充剂、保健品组合物、食品组合物。在另一优选例中,所述的组合物包括注射剂、口服制剂、透皮制剂。在另一优选例中,所述的癌细胞选自下组:肾癌细胞、肺癌细胞、肝癌细胞、乳腺癌细胞、大肠癌细胞、胃癌细胞、食管癌细胞、卵巢癌。在另一优选例中,所述的载体包括病毒载体、质粒。在本发明的第二方面,提供了一种表达外源的流感病毒蛋白的载体的用途,用于制备(a)用于将细胞的细胞周期停滞于G0/G1期的组合物;和/或(b)用于抑制癌细胞生长的组合物。在另一优选例中,所述的载体包括病毒载体、质粒。在另一优选例中,所述的病毒载体包括慢病毒载体、黄热病毒载体、腺病毒载体。在另一优选例中,所述的外源的流感病毒蛋白选自下组:NP蛋白、NS1蛋白、或其组合。在本发明的第三方面,提供了一种CPSF30抑制剂的用途,用于制备将细胞的细胞周期停滞于G0/G1期的组合物。在另一优选例中,所述的CPSF30抑制剂为病毒蛋白。在另一优选例中,所述的CPSF30抑制剂选自下组:流感病毒的NP蛋白、NS1蛋白、或其组合。在另一优选例中,所述的CPSF30抑制剂包括小分子化合物、反义核酸、或siRNA。在另一优选例中,所述的CPSF30抑制剂包括抗体、配体、或短肽。在另一优选例中,所述的组合物还用于抑制癌细胞或用于治疗肿瘤。在本发明的第四方面,提供了一种药物组合物或药物组合,其特征在于,所述的药物组合物或药物组合含有(i)药学上可接受的载体;(ii)病毒蛋白或其激动剂;和(iii)任选的不同于组分(ii)的其他抗肿瘤活性成分。在另一优选例中,所述的组分(ii)和(iii)可以位于同一制剂、或位于不同的制剂中。在另一优选例中,所述的组分(iii)为抑制癌细胞的有丝分裂的药物。在另一优选例中,所述的组分(iii)选自下组:raf酶抑制剂(如SORAFENIB(索拉非尼)或其类似药物)、有丝分裂酶抑制剂(如吉非替尼或其类似药物)。在另一优选例中,所述的组分(iii)为CPSF30抑制剂。在另一优选例中,所述的CPSF30抑制剂选自下组:小分子化合物、反义核酸、、siRNA、抗体、配体、短肽或其组合。在本发明的第五方面,提供了一种体外非治疗性的将细胞的细胞周期停滞于G0/G1期的方法,包括步骤:在(a)病毒蛋白或其激动剂,和/或(b)CPSF30抑制剂存在下,培养所述的细胞,从而使得所述细胞的细胞周期停滞于G0/G1期,其中,所述的病毒蛋白选自下组流感病毒蛋白:NP蛋白、NS1蛋白、或其组合。在另一优选例中,所述的细胞是表达CPSF30的细胞。在另一优选例中,所述的细胞来自哺乳动物(如灵长动物),更佳地人。在另一优选例中,所述的细胞包括正常细胞、或癌细胞。在另一优选例中,所述的癌细胞选自下组:肾癌细胞、肺癌细胞、肝癌细胞、乳腺癌细胞、大肠癌细胞、胃癌细胞、食管癌细胞、卵巢癌。在另一优选例中,所述的CPSF30抑制剂是反义核酸(如siRNA)。在本发明的第六方面,提供了一种筛选抗肿瘤候选药物的方法,包括步骤:(i)在实验组中,在所述测试物存在下,并且在病毒蛋白存在下,体外培养哺乳动物的细胞,并测试实验组中所述细胞的细胞周期情况;并且在对照组中,在测试条件相同但无所述测试物存在下,体外培养哺乳动物的细胞,并测试对照组中所述细胞的细胞周期,其中,所述的病毒蛋白选自下组:NP蛋白、NS1蛋白、或其组合;(ii)将实验组中细胞周期处于G0/G1期的细胞比例数值Rs与对照组中细胞周期处于G0/G1期的细胞比例数值Rc进行比较,如果Rs显著高于Rc,则表明该测试物为潜在的抗肿瘤候选药物(即与所述病毒蛋白存在相互作用关系的潜在的抗肿瘤候选药物);如果Rs不显著高于Rc,则不将该测试物视为与所述病毒蛋白存在相互作用关系的潜在的抗肿瘤候选药物;(iii)任选地,对于上一步骤中选出的潜在的抗肿瘤候选药物,测试其在无所述病毒蛋白存在情况下,对肿瘤细胞的抑制或杀伤能力。在另一优选例中,所述的“显著高于”指Rs/Rc之比≥1.20,较佳地≥1.50,更佳地≥2.0。另一种本发明的筛选方法包括步骤:(i)在实验组中,在所述测试物存在下,并且在病毒蛋白和CPSF30存在下,并测试实验组中所述病毒蛋白与CPSF30结合形成复合物的情况;并且在对照组中,在测试条件相同但无所述测试物存在下,测试实验组中所述病毒蛋白与CPSF30结合形成复合物情况,其中,所述的病毒蛋白选自下组:NP蛋白、NS1蛋白、或其组合;(ii)将实验组中所述病毒蛋白与CPSF30结合形成复合物的数量Vs与对照组中所述病毒蛋白与CPSF30结合形成复合物的数量Vc进行比较,如果Vs显著高于Vc,则表明该测试物为潜在的抗肿瘤候选药物(即与所述病毒蛋白存在相互作用关系的潜在的抗肿瘤候选药物),并可进行步骤(iii);如果Vs不显著高于Vc,则不将该测试物视为潜在的抗肿瘤候选药物;(iii)任选地,对于上一步骤中选出的潜在的抗肿瘤候选药物,测试其在无所述病毒蛋白存在情况下,对肿瘤细胞的抑制或杀伤能力。在另一优选例中,所述的测试物包括小分子化合物、反义核酸、抗体、或其组合。在本发明的第七方面,提供了一种复合物,其特征在于,所述的复合物是CPSF30与病毒蛋白形成的复合物,并且所述复合物选自下组:CPSF30/NP二元复合物、或CPSF30/NP/NS1三元复合物。在另一优选例中,所述的复合物是CPSF30/NP/NS1三元复合物。在本发明的第八方面,提供了本发明第七方面所述的复合物的用途,它用于筛选影响细胞周期的药物。在另一优选例中,所述的“影响细胞周期”指将细胞的细胞周期停滞于G0/G1期。在本发明的第九方面,提供了一种治疗方法,所述的治疗是将细胞的细胞周期停滞于G0/G1期,所述方法包括步骤:给需要的对象施用(a)病毒蛋白或其激动剂,和/或(b)CPSF30抑制剂。在另一优选例中,所述的病毒蛋白选自下组:NP蛋白、NS1蛋白、或其组合。在另一优选例中,所述的对象包括哺乳动物(如灵长动物),较佳地人。在另一优选例中,所述的施用病毒蛋白包括直接施用蛋白和间接施用蛋白(如施用表达所述蛋白的载体)。应理解,在本发明范围内中,本发明的上述各技术特征和在下文(如实施例)中具体描述的各技术特征可以互相组合,从而构成新的或优选的技术方案。限于篇幅,在此不再一一累述。附图说明图1显示了甲型流感病毒感染人肺癌细胞A549引起细胞周期停滞在G0/G1期。A549细胞以MOI=2感染1h,感染后24h收细胞,流式细胞术分析细胞周期。图中,(A)未感染的细胞(对照组,mock)和感染的细胞(感染组,infected)分别用NP的抗体染色,阳性细胞表明感染效率。(B)未感染的细胞(对照组)和感染的细胞(感染组)分别用PI染色,分析细胞周期。(C)使用ModFitLT软件统计B中处于不同细胞周期的细胞比例。图2显示了甲型流感病毒的NS1和NP基因的表达引起细胞周期停滞在G0/G1期。HEK293细胞不转染(mock),或者分别转染可表达GFP或者NP-GFP和NS1-GFP融合蛋白的质粒,转染后24h使用nocodazole(Noc)处理细胞16h,将细胞停滞在G2/M期,收细胞,流式细胞术分析。图中,(A)各组细胞分别用PI染色,分析细胞周期。(B)使用ModFitLT软件统计B中处于不同细胞周期的细胞比例。图3显示了NS1蛋白抑制细胞周期的能力与其结合CPSF30的能力正相关。用GST-CPSF30蛋白pulldownH5N1NS1和其F103S/M106I突变以及pH1N1NS和其三突变(TripleMut)108K/125D/189D,其中H5N1NS1,pH1N1NS1及其它们的突变蛋白是通过外翻译试剂盒(Promega)合成的。按图2同样的方法分别转染H5N1株和pH1N1株NS1基因和相应的突变基因,收取细胞然后检测细胞周期结果。图4显示了NP蛋白抑制细胞周期的能力与其结合CPSF30的能力正相关。上图为用GST-CPSF30蛋白pulldownH5N1NP和其17Gdel/K400R/A451T突变以及pH1N1NP和其三突变17Gdel/K400R/A451T的结果,其中H5N1NP,pH1N1NP及其它们的突变蛋白是通过外翻译试剂盒(Promega)合成的。下图为按图2同样的方法分别转染H5N1株和pH1N1株NP基因和相应的突变基因,收取细胞然后检测细胞周期结果。图5显示了抑制CPSF30表达水平导致细胞周期停滞在G0/G1期。图中,(A-B)HEK293细胞分别转染CPSF30的siRNA1,siRNA2和NegativecontrolsiRNA(NC),48小时后收集细胞,(A)RT-PCR检测CPSF30基因的转录情况;(B)做免疫蛋白印迹实验检测CPSFF30蛋白的表达情况。(C)将验证有效的siRNA1, siRNA2和NC转染HEK293细胞,24小时后加入Nocodazole,再经24小时后收集细胞检测细胞周期状态。(D)为ModFitLT软件分析细胞周期图以得到细胞周期中每一周期的细胞所占百分比,结果用柱状图显示,显示的是三次独立实验的平均值和标准差。图6显示了NS1和NP基因的表达可以抑制癌细胞的增殖。分别用表达NS1基因和NP基因的慢病毒或者不表达任何基因的对照病毒以MOI=20感染人肺癌细胞A549细胞(A和B)以及人子宫颈癌细胞Hela细胞(C和D),在感染后24、48、72、96、120小时分别向各组细胞中加入MTT反应液,检测OD值。图7显示了抑制CPSF4基因的表达可以抑制癌细胞的增殖。分别用表达siRNA1的慢病毒或者不表达任何基因的对照病毒以MOI=20感染人子宫颈癌细胞Hela细胞,在感染后24、48、72、96、120小时分别向各组细胞中加入MTT反应液,检测OD值。具体实施方式本发明人经过广泛而深入的研究,发现了流感病毒感染可以导致细胞周期停滞在G0/G1期的现象。令人意外的是,两个流感病毒蛋白NS1和NP能够有效起到了细胞周期的抑制作用,并且都是通过抑制一个共同的宿主蛋白,即mRNA剪切和多聚腺苷酸化因子CPSF30的活性,实现对细胞周期的抑制。在此基础上完成了本发明。具体地,本发明人利用流感病毒蛋白(NS1和/或NP)抑制细胞周期停滞在G0/G1期的原理,通过在癌细胞中外源表达流感病毒的NS1和NP蛋白,或者用siRNA的方法抑制癌细胞内源性CPSF30的表达来抑制癌细胞的增殖。术语如本文所用,术语“停滞于G0/G1期”指对于一细胞群而言,处于G0/G1期的细胞比例显著提高。例如,与对照组的细胞群相比,实验组处于G0/G1期的细胞比例显著提高。通常,这种相对提高幅度为≥20%,更佳地≥50%或更高。本发明蛋白及其制备如本文所用,术语“本发明蛋白”指NS1蛋白、NP蛋白或其组合。如本文所用,术语“本发明的多核苷酸”指用于编码NS1蛋白、NP蛋白或其组合的核苷酸序列。本发明的多核苷酸可以是DNA形式或RNA形式。DNA形式包括cDNA、基因 组DNA或人工合成的DNA。DNA可以是单链的或是双链的。DNA可以是编码链或非编码链。编码成熟多肽的编码区序列包括简并序列。本发明中的多肽和多核苷酸优选以分离的形式提供,更佳地被纯化至均质。本发明的NS1或NP的核苷酸全长序列或其片段通常可以用PCR扩增法、重组法或人工合成的方法获得。对于PCR扩增法,可根据本发明所公开的有关核苷酸序列,尤其是开放阅读框序列来设计引物,通过扩增而得有关序列。当序列较长时,常常需要进行两次或多次PCR扩增,然后再将各次扩增出的片段按正确次序拼接在一起。一旦获得了有关的序列,就可以用重组法来大批量地获得有关序列。这通常是将其克隆入载体,再转入细胞,然后通过常规方法从增殖后的宿主细胞中分离得到有关序列。此外,还可用人工合成的方法来合成有关序列,尤其是片段长度较短时。通常,通过先合成多个小片段,然后再进行连接可获得序列很长的片段。目前,已经可以完全通过化学合成来得到编码本发明蛋白(或其片段,或其衍生物)的DNA序列。然后可将该DNA序列引入本领域中已知的各种现有的DNA分子(或如载体)和细胞中。此外,还可通过化学合成将突变引入本发明蛋白序列中。本发明也涉及包含本发明的多核苷酸的载体,以及用本发明的载体或本发明蛋白编码序列经基因工程产生的宿主细胞,以及经重组技术产生本发明所述多肽的方法。通过常规的重组DNA技术(Science,1984;224:1431),可利用本发明的多聚核苷酸序列可用来表达或生产重组的本发明蛋白。一般来说有以下步骤:(1).用本发明的编码本发明蛋白的多核苷酸(或变异体),或用含有该多核苷酸的重组表达载体转化或转导合适的宿主细胞;(2).在合适的培养基中培养的宿主细胞;(3).从培养基或细胞中分离、纯化蛋白质。mRNA剪切和多聚腺苷酸化因子CPSF30如本文所用,术语“CPSF30”和“CPSF-30”以及“CPSF4”可互换使用,指剪切和多聚腺苷酸化特异性因子30(cleavageandpolyadenylationspecificityfactor30)。应理解,该术语CPSF30不仅包括野生型CPSF30,还包括突变型CPSF30。此外,CPSF30基因或核酸序列包括基因组、cDNA或RNA形式,而CPSF30蛋白或多肽包括全长CPSF30或其活性片段。在另一优选例中,所述的CPSF30是来源于哺乳动物,尤其是人的CPSF30。人CPSF30的核苷酸序列和氨基酸序列的登录号为GenBankNM_006693.2,它是剪切和多聚腺苷酸化特异性因子家族蛋白中的一员,蛋白大小约30KD,因此被称为CPSF30,因其是4个家族蛋白中最小的一个;CPSF30与该家族的其他蛋白CPSF73,CPSF100及CPSF160组成复合体负责剪切前体mRNA的3’端并加入多聚腺苷酸尾巴,促进mRNA的成熟。流感病毒蛋白NP如本文所用,术语“流感病毒蛋白NP”和“蛋白NP”可互换使用,指来自于流感病毒的核蛋白。应理解,该术语NP不仅包括野生型NP,还包括突变型NP。此外,NP基因或核酸序列包括DNA或RNA形式,而NP蛋白或多肽包括全长NP或其活性片段。此外,流感病毒蛋白NP不仅包括来自各型或各亚型流感病毒的NP蛋白或其活性片段,还包括来自于类似病毒(如副流感病毒)的NP蛋白或其活性片段,只要该NP蛋白或其活性片段同样具有与CPSF30蛋白(尤其是人CPSF30蛋白)结合并抑制CPSF30活性的功能。代表性的各型流感病毒包括(但并不限于):甲型流感病毒、乙型流感病毒、丙型流感病毒等。应理解,不同的流感病毒的基因存在一定的变异。因此,基于本发明,术语NP包括这些变异的或突变的NP基因和NP蛋白。作为一种代表性的NP蛋白,甲型流感病毒A/WSN/33(H1N1)的NP的核苷酸序列和氨基酸序列如下所示:甲型流感病毒A/WSN/33(H1N1)的NP的核苷酸序列(SEQIDNO.:1)atggcgaccaaaggcaccaaacgatcttacgaacagatggagactgatggagaacgccagaatgccactgaaatcagagcatctgtcggaaaaatgattgatggaattggacgattctacatccaaatgtgcaccgaacttaaactcagtgattatgagggacggctgattcagaacagcttaacaatagagagaatggtgctctctgcttttgacgagaggaggaataaatatctagaagaacatcccagtgcggggaaagatcctaagaaaactggaggacctatatacaggagagtagatggaaagtggaggagagaactcatcctttatgacaaagaagaaataagacgaatctggcgccaagctaataatggtgacgatgcaacggctggtctgactcacatgatgatctggcactccaatttgaatgatgcaacttaccagaggacaagagctcttgttcgcacaggaatggatcccaggatgtgctcactgatgcagggttcaaccctccctaggaggtctggggccgcaggtgctgcagtcaaaggagttggaacaatggtgatggaattgatcagaatgatcaaacgtgggatcaatgatcggaacttctggaggggtgagaatggacggagaacaaggattgcttatgaaagaatgtgcaacattctcaaagggaaatttcaaacagctgcacaaagaacaatggtggatcaagtgagagagagccggaatccaggaaatgctgagttcgaagatctcatctttttagcacggtctgcactcatattgagagggtcagttgctcacaagtcctgcctgcctgcctgtgtgtatggatctgccgtagccagtggatacgact ttgaaagagagggatactctctagtcggaatagaccctttcagactgcttcaaaacagccaagtatacagcctaatcagaccaaatgagaatccagcacacaagagtcaactggtgtggatggcatgccattctgctgcatttgaagatctaagagtatcaagcttcatcagagggacgaaagtggtcccaagagggaagctttccactagaggagttcaaattgcttccaatgaaaacatggagactatggaatcaagtacccttgaactgagaagcagatactgggccataaggaccagaagtggagggaacaccaatcaacagagggcttcctcgggccaaatcagcatacaacctacgttctcagtacagagaaatctcccttttgacagaccaaccattatggcagcattcactgggaatacagaggggagaacatctgacatgagaaccgaaatcataaggctgatggaaagtgcaagaccagaagatgtgtctttccaggggcggggagtcttcgagctctcggacgaaaaggcaacgagcccgatcgtgccctcctttgacatgagtaatgaaggatcttatttcttcggagacaatgcagaggagtacgacaattaa甲型流感病毒A/WSN/33(H1N1)的NP的蛋白序列(SEQIDNO.:2)MATKGTKRSYEQMETDGERQNATEIRASVGKMIDGIGRFYIQMCTELKLSDYEGRLIQNSLTIERMVLSAFDERRNKYLEEHPSAGKDPKKTGGPIYRRVDGKWRRELILYDKEEIRRIWRQANNGDDATAGLTHMMIWHSNLNDATYQRTRALVRTGMDPRMCSLMQGSTLPRRSGAAGAAVKGVGTMVMELIRMIKRGINDRNFWRGENGRRTRIAYERMCNILKGKFQTAAQRTMVDQVRESRNPGNAEFEDLIFLARSALILRGSVAHKSCLPACVYGSAVASGYDFEREGYSLVGIDPFRLLQNSQVYSLIRPNENPAHKSQLVWMACHSAAFEDLRVSSFIRGTKVVPRGKLSTRGVQIASNENMETMESSTLELRSRYWAIRTRSGGNTNQQRASSGQISIQPTFSVQRNLPFDRPTIMAAFTGNTEGRTSDMRTEIIRLMESARPEDVSFQGRGVFELSDEKATSPIVPSFDMSNEGSYFFGDNAEEYDN在本发明中,术语“NP蛋白”指具有NP蛋白活性、以SEQIDNO:2序列为代表的多肽。该术语还包括具有与NP蛋白相同功能的、SEQIDNO:2序列的变异形式。这些变异形式包括(但并不限于):一个或多个(通常为1-50个,较佳地1-30个,更佳地1-20个,最佳地1-10个)氨基酸的缺失、插入和/或取代,以及在C末端和/或N末端添加一个或数个(通常为20个以内,较佳地为10个以内,更佳地为5个以内)氨基酸。例如,在本领域中,用性能相近或相似的氨基酸进行取代时,通常不会改变蛋白质的功能。又比如,在C末端和/或N末端添加一个或数个氨基酸通常也不会改变蛋白质的功能。该术语还包括NP蛋白的活性片段和活性衍生物。该多肽的变异形式包括:同源序列、保守性变异体、等位变异体、天然突变体、诱导突变体、在高或低的严紧度条件下能与NPDNA杂交的DNA所编码的蛋白、以及利用抗NP多肽的抗血清获得的多肽或蛋白。本发明还提供了其他多肽,如包含NP多肽或其片段的融合蛋白(如与GFP形成的融合蛋白)。除了几乎全长的多肽外,本发明还包括了NP多肽的可溶性片段。通常,该片段具有NP多肽序列的至少约10个连续氨基酸,通常至少约30个连续氨基酸,较佳地至少约50个连续氨基酸,更佳地至少约80个连续氨基酸,最佳地至少约100个连续氨基酸。在本发明中,“NP保守性变异多肽”指与SEQIDNO:2的氨基酸序列相比,有至多10个,较佳地至多8个,更佳地至多5个,最佳地至多3个氨基酸被性质相似或相近的氨基酸所替换而形成多肽。这些保守性变异多肽最好根据表1进行氨基酸替换而产生。表1最初的残基代表性的取代优选的取代Ala(A)Val;Leu;IleValArg(R)Lys;Gln;AsnLysAsn(N)Gln;His;Lys;ArgGlnAsp(D)GluGluCys(C)SerSerGln(Q)AsnAsnGlu(E)AspAspGly(G)Pro;AlaAlaHis(H)Asn;Gln;Lys;ArgArgIle(I)Leu;Val;Met;Ala;PheLeuLeu(L)Ile;Val;Met;Ala;PheIleLys(K)Arg;Gln;AsnArgMet(M)Leu;Phe;IleLeuPhe(F)Leu;Val;Ile;Ala;TyrLeuPro(P)AlaAlaSer(S)ThrThrThr(T)SerSerTrp(W)Tyr;PheTyrTyr(Y)Trp;Phe;Thr;SerPheVal(V)Ile;Leu;Met;Phe;AlaLeu流感病毒蛋白NS1如本文所用,术语“流感病毒蛋白NS1”和“蛋白NS1”可互换使用,指来自于甲型流感病毒的非结构蛋白1。应理解,该术语NS1不仅包括野生型NS1,还包括突变型NS1。此外,NS1基因或核酸序列包括DNA或RNA形式,而NS1蛋白或多肽包括全长NS1或其活性片段。此外,流感病毒蛋白NS1不仅包括来自各型或各亚型流感病毒的NS1蛋白或其活性片段,还包括来自于类似病毒(如副流感病毒)的NS1蛋白或其活性片段,只要该NS1蛋白或其活性片段同样具有与CPSF30蛋白(尤其是人CPSF30蛋白)结合并抑制CPSF30活性的功能。代表性的各型流感病毒包括(但并不限于):甲型流感病毒、乙型流感病毒、 丙型流感病毒等。应理解,不同的流感病毒的基因存在一定的变异。因此,基于本发明,术语NS1包括这些变异的或突变的NS1基因和NS1蛋白。作为一种代表性的NS1蛋白,甲型流感病毒A/WSN/33(H1N1)的NS1的核苷酸序列和氨基酸序列如下所示:甲型流感病毒A/WSN/33(H1N1)的NS1的核苷酸序列(SEQIDNO.:3)ATGGATCCAAACACTGTGTCAAGCTTTCAGGTAGATTGCTTTCTTTGGCATGTCCGCAAAAGAGTTGCAGACCAAGAACTAGGTGATGCCCCATTCCTTGATCGGCTTCGCCGAGATCAGAAGTCCCTAAGAGGAAGAGGCAGCACTCTTGGTCTGGACATCGAAACAGCCACCCGTGCTGGAAAGCAAATAGTGGAGCGGATTCTGAAGGAAGAATCTGATGAGGCACTCAAAATGACCATGGCCTCTGTACCTGCATCGCGCTACCTAACTGACATGACTCTTGAGGAAATGTCAAGGCACTGGTTCATGCTCATGCCCAAGCAGAAAGTGGCAGGCCCTCTTTGTATCAGAATGGACCAGGCGATCATGGATAAGAACATCATACTGAAAGCGAACTTCAGTGTGATTTTTGACCGGCTGGAGACTCTAATATTACTAAGGGCCTTCACCGAAGAGGGGACAATTGTTGGCGAAATTTCACCACTGCCCTCTCTTCCAGGACATACTGATGAGGATGTCAAAAATGCAGTTGGGGTCCTCATCGGAGGACTTGAATGGAATAATAACACAGTTCGAGTCTCTGAAACTCTACAGAGATTCGCTTGGAGAAGCAGTAATGAGAATGGGAGACCTCCACTCACTCCAAAACAGAAACGGAAAATGGCGGGAACAATTAGGTCAGAAGTTTGA甲型流感病毒A/WSN/33(H1N1)的NS1的蛋白序列(SEQIDNO.:4)MDPNTVSSFQVDCFLWHVRKRVADQELGDAPFLDRLRRDQKSLRGRGSTLGLDIETATRAGKQIVERILKEESDEALKMTMASVPASRYLTDMTLEEMSRHWFMLMPKQKVAGPLCIRMDQAIMDKNIILKANFSVIFDRLETLILLRAFTEEGTIVGEISPLPSLPGHTDEDVKNAVGVLIGGLEWNNNTVRVSETLQRFAWRSSNENGRPPLTPKQKRKMAGTIRSEV在本发明中,术语“NS1蛋白”指具有NS1蛋白活性、以SEQIDNO:4序列为代表的多肽。该术语还包括具有与NS1蛋白相同功能的、SEQIDNO:4序列的变异形式。这些变异形式包括(但并不限于):一个或多个(通常为1-50个,较佳地1-30个,更佳地1-20个,最佳地1-10个)氨基酸的缺失、插入和/或取代,以及在C末端和/或N末端添加一个或数个(通常为20个以内,较佳地为10个以内,更佳地为5个以内)氨基酸。例如,在本领域中,用性能相近或相似的氨基酸进行取代时,通常不会改变蛋白质的功能。又比如,在C末端和/或N末端添加一个或数个氨基酸通常也不会改变蛋白质的功能。该术语还包括NS1蛋白的活性片段和活性衍生物。该多肽的变异形式包括:同源序列、保守性变异体、等位变异体、天然突变体、诱导突变体、在高或低的严紧度条件下能与NS1DNA杂交的DNA所编码的蛋白、以及利用抗NS1多肽的抗血清获得的多肽或蛋白。本发明还提供了其他多肽, 如包含NS1多肽或其片段的融合蛋白(如与GFP形成的融合蛋白)。除了几乎全长的多肽外,本发明还包括了NS1多肽的可溶性片段。通常,该片段具有NS1多肽序列的至少约10个连续氨基酸,通常至少约30个连续氨基酸,较佳地至少约50个连续氨基酸,更佳地至少约80个连续氨基酸,最佳地至少约100个连续氨基酸。在本发明中,“NS1保守性变异多肽”指与SEQIDNO:4的氨基酸序列相比,有至多10个,较佳地至多8个,更佳地至多5个,最佳地至多3个氨基酸被性质相似或相近的氨基酸所替换而形成多肽。这些保守性变异多肽最好根据表1进行氨基酸替换而产生。药物组合物和治疗本发明还提供了一种药物组合物或药物组合,所述的药物组合物含有(i)药学上可接受的载体;(ii)本发明的病毒蛋白或其激动剂;和(iii)任选的不同于组分(ii)的其他抗肿瘤活性成分。这类药学上可接受的载体包括(但并不限于):盐水、缓冲液、葡萄糖、水、甘油、乙醇、及其组合。药物制剂应与给药方式相匹配。本发明的药物组合物可以被制成针剂形式,例如用生理盐水或含有葡萄糖和其他辅剂的水溶液通过常规方法进行制备。诸如片剂和胶囊之类的药物组合物,可通过常规方法进行制备。药物组合物如针剂、溶液、片剂和胶囊宜在无菌条件下制造。此外,本发明的药物活性成分还可与其他抗肿瘤治疗剂一起使用。使用药物组合物时,是将安全有效量的(a)本发明病毒蛋白或其激动剂,和/或(b)CPSF30抑制剂施用于哺乳动物,其中该安全有效量通常至少约10微克/千克体重,而且在大多数情况下不超过约8毫克/千克体重,较佳地该剂量是约10微克/千克体重-约1毫克/千克体重。当然,具体剂量还应考虑给药途径、病人健康状况等因素,这些都是熟练医师技能范围之内的。本发明还提供了一种治疗方法,所述的治疗是将细胞的细胞周期停滞于G0/G1期,所述方法包括步骤:给需要的对象施用(a)病毒蛋白或其激动剂,和/或(b)CPSF30抑制剂。在另一优选例中,所述的病毒蛋白选自下组:NP蛋白、NS1蛋白、或其组合。在另一优选例中,所述的对象包括哺乳动物(如灵长动物),较佳地人。在另一优选例中,所述的施用病毒蛋白包括直接施用蛋白和间接施用蛋白(如施用表达所述蛋白的载体)。本发明的所述治疗方法可以用于通过将肿瘤细胞停滞于G0/G1期,从而有效抑制和治疗各种不同的肿瘤。药物筛选本发明还提供了一种抗肿瘤药物的筛选方法,该方法可以更高效地筛选出与本发明的NP蛋白或NS1蛋白存在相互作用关系的潜在抗肿瘤药物。典型地,一种筛选方法包括以下步骤:(i)在实验组中,在所述测试物存在下,并且在病毒蛋白存在下,体外培养哺乳动物的细胞,并测试实验组中所述细胞的细胞周期情况;并且在对照组中,在测试条件相同但无所述测试物存在下,体外培养哺乳动物的细胞,并测试对照组中所述细胞的细胞周期,其中,所述的病毒蛋白选自下组:NP蛋白、NS1蛋白、或其组合;(ii)将实验组中细胞周期处于G0/G1期的细胞比例数值Rs与对照组中细胞周期处于G0/G1期的细胞比例数值Rc进行比较,如果Rs显著高于Rc,则表明该测试物为潜在的抗肿瘤候选药物(即与所述病毒蛋白存在相互作用关系的潜在的抗肿瘤候选药物),并可进行步骤(iii);如果Rs不显著高于Rc,则不将该测试物视为潜在的抗肿瘤候选药物;(iii)任选地,对于上一步骤中选出的潜在的抗肿瘤候选药物,测试其在无所述病毒蛋白存在情况下,对肿瘤细胞的抑制或杀伤能力。在另一优选例中,所述步骤(iii)中的测试包括体外细胞测试、动物测试等。在另一优选例中,所述的方法是非诊断性和非治疗性的。在另一优选例中,所述的筛选方法包括:(i)在实验组中,在所述测试物存在下,并且在病毒蛋白和CPSF30存在下,并测试实验组中所述病毒蛋白与CPSF30结合形成复合物的情况;并且在对照组中,在测试条件相同但无所述测试物存在下,测试实验组中所述病毒蛋白与CPSF30结合形成复合物情况,其中,所述的病毒蛋白选自下组:NP蛋白、NS1蛋白、或其组合;(ii)将实验组中所述病毒蛋白与CPSF30结合形成复合物的数量Vs与对照组中所述病毒蛋白与CPSF30结合形成复合物的数量Vc进行比较,如果Vs显著高于Vc,则表明该测试物为潜在的抗肿瘤候选药物(即与所述病毒蛋白存在相互作用关系的潜在的抗肿瘤候选药物),并可进行步骤(iii);如果Vs不显著高于Vc,则不将该测试物视为潜在的抗肿瘤候选药物;(iii)任选地,对于上一步骤中选出的潜在的抗肿瘤候选药物,测试其在无所述病毒蛋白存在情况下,对肿瘤细胞的抑制或杀伤能力。在另一优选例中,所述的“显著高于”指Vs/Vc之比≥1.20,较佳地≥1.50,更佳地≥2.0。在另一优选例中,所述的复合物为CPSF30/NP二元复合物、CPSF30/NS1二元 复合物、或CPSF30/NP/NS1三元复合物。本发明的主要优点包括:(a)首次揭示了流感病毒的蛋白NP和NS1能够使得人细胞(尤其是癌细胞)停滞于G0/G1期,从而可用于抑制癌细胞的生长。(b)首次揭示了流感病毒的蛋白NS1和NP可有效地通过结合于人CPSF30而使得抑制CPSF30的活性,进而抑制癌细胞的生长。(c)基于本发明的意外发现,可以开发利用NS1与CPSF30的结合和/NP与CPSF30的结合,从而抑制癌细胞生长的药物。下面结合具体实施例,进一步阐述本发明。应理解,这些实施例仅用于说明本发明而不用于限制本发明的范围。下列实施例中未注明具体条件的实验方法,通常按照常规条件,例如Sambrook等人,分子克隆:实验室手册(NewYork:ColdSpringHarborLaboratoryPress,1989)中所述的条件,或按照制造厂商所建议的条件。除非另外说明,否则百分比和份数为重量百分比和重量份数。通用方法和材料细胞培养:人肺腺癌细胞系(A549),人宫颈癌细胞(Hela)以及人胚肾细胞HEK293均从ATCC购得,在含10%胎牛血清的DMEM培养基(Hyclone),5%浓度的CO2,37℃的细胞培养箱中培养,培养基中同时添加50unit/ml青霉素(Invitrogen),50ug/ml链霉素(Invitrogen)和0.1mg/ml的卡那霉素(Invitrogen)。病毒基因:流感病毒A/Sichuan/1/2009(H1N1)(2009pH1N1)病毒基因由中国疾病和预防控制中心提供。流感病毒A/Duck/Hubei/L-1/2004(H5N1)病毒基因由武汉病毒所提供。流感病毒A/WSN/33(H1N1)病毒基因获自公共数据库或马尔堡大学。质粒构建:所有病毒基因的克隆都用相应的引物进行PCR扩增后克隆至pTM1载体(购自Promega公司)用于体外翻译病毒蛋白。病毒基因另克隆至慢病毒表达载体pLenti-CMV-NS1-HA-3Flag-P2A-LUC-T2A-Puro中(购自和元生物),用于包装可表达病毒蛋白的慢病毒。或者,将NP和NS1基因克隆在pEGFP-C1质粒(购自Clontech公司)上。突变质粒的构建是根据基于PCR的定点诱变说明书完成的,通过KOD+DNA聚合酶(Toyobo)合成新的突变的质粒,然后经过DpnI限制性酶(NEB)将原有的质粒切碎,然后转染大肠杆菌来完成。突变质粒的序列均通过测序鉴定确认。人的全长CPSF-30cDNA由细胞内抽提的RNA经过RT-PCR得到,通过CPSF30的cDNA序列设计引物,将全长序列并克隆至pGEX-4T1载体(购自GE公司)上,分别获得pGEX-4T1-CPSF-30。纯化的GST-CPSF-30蛋白,是通过将pGEX-4T1-CPSF-30转入大肠杆菌BL21,并通过常规纯化方法获得。CPSF30的siRNA1序列克隆至慢病毒载体。GV115(购自吉凯生物公司),用于包装可表达该siRNA的慢病毒。转染:通过lipofectamine2000(Invitrogen)将带有NP和NS1基因的pEGFP-C1质粒转入6孔板的50-60%融合状态的人胚肾细胞HEK293细胞,24小时后加入400ng/ml市售的Nocodazole(Sigma公司)(一种可以通过使微管解聚而使细胞周期停留在G2/M期的药物),一天后用胰酶消化细胞,收集到流式管中,用1ml冷的70%乙醇固定,在4度存放用于流式细胞仪分析细胞周期。siRNA干扰:两条siRNA序列为:siRNA1:ACGCCAACAGAUGCACCAAA(SEQIDNO.:5),siRNA2:AGAGUCAUCUGUGUGAAUUACCUCG(SEQIDNO.:6)。使用X-tremeGENEsiRNATransfectionReagent(Roche),将这些siRNA转染入HEK293细胞和A549细胞。48小时后,收集细胞做免疫印迹和实时定量PCR(RT-PCR)分析CPSF30的表达情况。CPSF30表达水平降低的HEK293细胞在加入400ng/mlNocodazole处理后,收集下来用于细胞周期的分析。实时定量PCR:从6孔板中收取106个细胞,加入1mlTRIZOL,吹吸至液体澄清,室温放置5分钟,然后加0.2ml氯仿,颠倒混匀,4℃12000g离心15分钟,将上层水相转入另一干净EP管中,然后加入等体积异丙醇,混匀,室温放置10分钟。12000g4℃离心10分钟。弃上清,加入75%冷乙醇1ml,7500g4℃离心5分钟,弃上清,空气干燥,溶于20μlDEPC水中。将提取的RNA(2μg)使用ReverTraqPCRRTKit(Toyobo)反转录为cDNA,使用RealtimePCRMasterMix(Toyobo)来进行特异性的PCR产物扩增检测。CPSF-30定量PCR引物序列:Forward:5’-TTGACTTGGAGATCGCGG-3’(SEQIDNO.:7)Revers:5’-GCAGGCAGCTTTCAAAAAGA-3’(SEQIDNO.:8)。GAPDH定量PCR引物序列:Forward:5’-GAAATCCCATCACCATCTTCCAC-3’(SEQIDNO.:9)Reverse:5’-GAGCCCCAGCCTTCTCCATG-3’(SEQIDNO.:10)。细胞周期分析:核内DNA含量用PI染色然后用流式细胞仪进行分析贴壁生长的细胞用胰酶消化,然后用PBS清洗。细胞在1ml70%的冷乙醇中固定,然后重悬于染色液(50ug/mlPI[Sigma],20ug/mlRNAase)中。在室温下放置20分钟后用流式细胞仪(FACScan;BDLSRII)进行分析。每个样品至少收30,000个细胞进行分析。数据分析软件为ModfitLT2.0(VeritySoftwareHouse)。融合蛋白的原核表达与纯化:为了纯化融合蛋白GST-CPSF-30,将原核表达质粒pGEX4T1-CPSF-30转化至大肠杆菌表达菌株BL21中,挑取单菌落于15ml含有相应抗生素的LB培养基中37℃摇床培养过夜,然后以1:100的比例将过夜培养物接种至1L含有相应抗生素的LB培养基中37℃摇床培养2h左右至OD600=0.4-0.6,取出1ml未诱导的培养物于1.5ml的离心管中,作为阴性对照。剩下的加入终浓度为1mM的IPTG(Isopropyl-1-thio-β-D-galactopyranoside),18℃摇床培养4h左右,收集菌液,7000rpm,离心15min,去上清,用预冷的PBS缓冲液重悬后超声,细胞裂解液于12000rpm,4℃,离心10min,取40μl上清加入6×SDS上样缓冲液重悬,于100℃煮沸5min,取20μl上样于SDS-PAGE电泳检测。上清冻存于-20℃冰箱待纯化。融合蛋白的纯化采取亲和纯化树脂,纯化后的融合蛋白GST-CPSF-30用于GSTpull-down。GST共沉淀分析(PullDownanalysis):将构建在pTM1载体上的NS1和NP基因直接用带有T7及转录终止信号的引物对来PCR目的基因,然后利用TNTtranscription/translationkit(Promega)体外合成HA-taggedNS1和NP。将等量的GST与GST-CPSF-30蛋白加到GlutathioneSepharoseBeads(GE),4℃旋转混匀1h,用预冷的PBS洗珠三次以去除未结合的GST与GST-CPSF-30,然后加入体外转录的目的蛋白,4℃旋转过夜,预冷的PBS洗珠三次,加入2×SDS上样缓冲液,100℃煮5min,上样于SDS-PAGE,Westernblot检测。免疫印迹实验:细胞用PBS洗一遍,然后直接用SDS缓冲液(60mMTris-HCl[pH6.8],2%SDS,10%glycerol,5%2-mercaptoethanol,0.01%bromophenolblue)裂解,之后煮10分钟。全细胞裂解液跑SDS-PAGE电泳。蛋白被转移到硝酸纤维素膜(BioRad)上,用5%脱脂奶粉溶于TBST(20mMTris-HCL,150mMNaCL,0.02Tween),室温条件下封闭1小时,然后用对应的一抗和二抗检测。显影使用enhancedchemiluminescencekit(PIERCE)。在免疫印迹实验中使用了下列抗体:鼠源单克隆抗体:CPSF-30抗体(购自Abgent);辣根过氧化物酶标记的抗鼠IgG购自Sigma;抗NP和抗NS1多克隆抗体购自上海生命科学研究院抗体研究中心。实施例1、流感病毒蛋白NS1和NP将细胞周期抑制在G0/G1期在本实施例中,通过感染试验或转染试验,观察流感病毒以及流感病毒的各蛋白对细胞周期的影响。结果如图1所示,甲型流感病毒(具体为A/WSN/33(H1N1))感染人肺癌细胞A549引起细胞周期停滞在G0/G1期。在图1A中,病毒感染效率超过90%。此时,相比较于对照组,感染组的细胞明显更多的聚集于G0/G1期(图1B),数据统一表明,只有约30%的细胞处于G0/G1期,而感染组细胞则有约60%处于G0/G1期(图1C)。此外,通过大量筛选,意外地发现,通过在HEK293细胞中转染表达甲型流感病毒A/WSN/33(H1N1)的病毒蛋白基因时,该流感病毒的两种蛋白(即NS1蛋白和NP蛋白)的表达可以分别将细胞周期抑制到G0/G1期(图2)。如图2A所示,GFP本身的表达对细胞周期几乎没有影响。加入Noc将细胞周期停止在G2/M期可以更清楚的看到细胞在此前的G0/G1和S期的分布,加入noc后,由于GFP的表达对细胞周期没有影响,因此大多数细胞顺利通过G0/G1和S期进入G2/M期并停止在G2/M期;而表达有NS1-GFP和NP-GFP融合蛋白的细胞则被停滞在G0/G1期,不能其后的S和G2/M期进行(图2A)。数据统计表明,在NP组中,约50%细胞被抑制G0/G1期;在NS1组中,接近60%细胞被抑制G0/G1期,而在GFP对照组中,只有不到20%细胞被抑制G0/G1期(图2B)。实施例2、NS1和NP蛋白抑制细胞周期的能力依赖于其与宿主因子CPSF30的结合。为了找出NS1蛋白和NP蛋白抑制细胞周期的分子机制,在本实施例中分析了诸多甲型流感病毒毒株的NS1蛋白和NP蛋白的功能,并且还构建了NS1蛋白和NP 蛋白的多种突变蛋白并观察这些突变蛋白对细胞周期的影响实验结果表明,(1)NS1可与CPSF30结合形成复合物。能够和宿主因子CPSF30相互作用的H5N1病毒的NS1蛋白具有细胞周期的抑制作用,而该H5N1病毒的NS1的F103S,M106I突变体和CPSF30的结合大大减弱,其抑制细胞周期的能力也随着丧失(图3)。(2)来自于2009pH1N1病毒的NS1蛋白由于自身和CPSF30的结合能力较差,因此,只有引入三个位点(108K/125D/189D)的突变(TripleMut)增强其与CPSF30的结合才能使其获得更强的抑制细胞周期的能力(图3)。(3)NP可与CPSF30结合形成复合物。来自于H5N1和2009pH1N1的NP蛋白都与CSPF4相互作用并具有抑制细胞周期的能力(图4)。但是,当在NP蛋白上引入三个位点的突变(17Gdel/400R/451T)时(TripleMut),NP蛋白和CPSF30的结合大大减弱,其抑制细胞周期的能力也随着丧失。这些实验结果提示,NS1和NP蛋白抑制细胞周期的能力与其与宿主因子CPSF30的结合有关。另外,由于NP和NS1结合于CPSP30,因此三者可形成CPSF30/NP/NS1三元复合物。实施例3、CPSF30是保持细胞周期正常进行的关键性分子实施例1-2的研究表明,CPSF30是细胞周期中的一种重要分子,病毒蛋白结合并劫持CPSF30(降低CPSF30的活性),从而导致细胞周期的停滞。为了验证CPSF30对细胞周期的调控作用,在本实施例中使用siRNA的方法对细胞内源性的CPSF30进行表达抑制。结果表明,两条siRNA序列(siRNA1和siRNA2)都可以有效地抑制CPSF30的mRNA和蛋白表达(图5A和5B)。转染有这两种siRNA的细胞,其细胞周期显著地停滞在G0/G1期(图5C和5D)。这表明,抑制CPSF30的表达或活性可以抑制细胞周期的正常进行。此外,通过抑制CPSF30,可以有效地使得细胞周期停滞于G0/G1期。实施例4、在癌细胞中过表达甲型流感病毒的NS1和NP基因可以抑制癌细胞系的增殖为了在癌细胞中高效表达NS1和NP基因,在本实施例中,将两个病毒基因NP和NS1分别构建于慢病毒表达载体中。以表达有NS1或者NP基因的慢病毒感染癌细胞系A549和Hela,通过MTT方法检测NS1、NP感染细胞组与阴性对照慢病毒感染细胞组的增殖能力,研究NS1、NP基因对癌细胞增殖能力的影响。结果表明,在感染后96和120小时,表达有NS1蛋白的实验组的细胞增殖显 著低于A549阴性对照感染组(**p<0.01,***p<0.001),而表达有NP蛋白的实验组的细胞增殖也略低于A549阴性对照感染组(图6A和B);同样的,在感染后96和120小时,表达有NS1蛋白的实验组的细胞增殖显著低于Hela阴性对照感染组(**p<0.01,***p<0.001),表达有NP蛋白的实验组也显著低于Hela阴性对照感染组(*p<0.05,***p<0.001)(图6C和D)。这些结果提示,NS1和NP的基因表达都可抑制A539细胞和Hela细胞的增殖。实施例5、抑制CPSF30的表达可以抑制癌细胞增殖在本实施例中,将抑制CPSF30表达的siRNA系列(siRNA1)克隆于慢病毒表达载体上,并以此慢病毒感染癌细胞系A549和Hela,通过MTT方法检测抑制CPSF30的表达对A549和Hela细胞增殖能力的影响。结果表明,在A549细胞中,虽然阴性对照感染组与空白组相比,对细胞增殖有抑制作用,但从感染后72小时开始,表达有CPSF30的siRNA的实验组的细胞增殖就低于A549阴性对照感染组,数据分析显示,在感染后96小时,表达有CPSF30的siRNA的实验组的细胞增殖显著低于A549阴性对照感染组(*P<0.05)(图7A和B)。在Hela细胞中,阴性对照感染组与空白组相比,对细胞增殖几乎没有影响,因此,但从感染后24小时开始,表达有CPSF30的siRNA的实验组的细胞增殖就显著低于阴性对照感染组(*P<0.05,***P<0.001)(图7C和D)。这表明,通过RNA干扰技术抑制CPSF30基因表达,可以抑制A549和Hela癌细胞的增殖。讨论病毒感染对正常机体来说是不利于健康的,但是人们对病毒学的研究发现,病毒通过和宿主的相互作用可以影响很多宿主信号通路和生理反应,这一点使得病毒也有被加以利用的可能,利用病毒的特性用来治疗病变的细胞。在癌症治疗中,利用病毒感染导致细胞死亡的原理杀死癌细胞的想法早在1940年左右出现,但直到1991年,才发现第一个溶癌病毒(Oncolyticvirus)疱疹病毒可以裂解脑胶质瘤细胞,诞生了第一个病毒疗法(Virotherapy)。随后,很多不同的病毒都被用来研究其溶癌特性。然而,这种病毒疗法主要还是利用病毒感染最终导致细胞裂解的原理,虽然可以实现癌细胞感染特异性,但需要具有复制能力的活病毒,其安全性导致很大的应用限制。一个完整的细胞周期,即连续分裂的细胞从上一次有丝分裂结束到下一次有丝分裂完成,包含G0期(静息期)、G1期(前期)、S期(中期)、G2期(后期)、M期(有丝分裂期)四个阶段。目前,很多抗癌药物都是抑制癌细胞的有丝分裂。如德国拜尔公司研发的SORAFENIB(索拉非尼),就含有RAF抑制剂,抑制有丝分裂酶的活性。英国阿斯利康公司研发的抗癌新药易瑞沙(吉非替尼)也是通过抑制有丝分裂酶达到抑制癌细胞分裂增殖的目的。然而,有丝分裂(mitosis)发生在整个细胞周期的末期。如果能够将癌细胞抑制在细胞周期的早期,如G0/G1期,从源头抑制细胞周期的进行,将具有更优的抑制效果。本发明完全跳出这种病毒溶癌原理的局限,开发了可直接干扰细胞周期的病毒蛋白,从而更安全和更有针对性地抑制细胞周期。通过将甲型流感病毒的NS1和NP基因在癌细胞系中表达,或者通过在癌细胞周中抑制CPSF30基因表达,实现对癌细胞细胞增殖的抑制,对开发抗癌新方法有重要意义。此外,本发明的NP和NS1蛋白或其激动剂可有效地将细胞(尤其肿瘤细胞)停滞于G0/G1期,因此,结合现有的抑制有丝分裂的药物,将会达到更彻底的抑制癌细胞分裂的功效。在本发明提及的所有文献都在本申请中引用作为参考,就如同每一篇文献被单独引用作为参考那样。此外应理解,在阅读了本发明的上述讲授内容之后,本领域技术人员可以对本发明作各种改动或修改,这些等价形式同样落于本申请所附权利要求书所限定的范围。当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1