白介素-2的部分激动剂的制作方法
相关申请的交叉引用
本申请要求2017年11月21日提交的美国临时专利申请序列号62/589,497的优先权权益。以上引用的申请的公开内容通过引用以其整体(包括任何附图)明确并入本文。
关于联邦政府资助研发的声明
本发明是在由美国国立卫生研究院授予的r37ai051321下在美国政府支持下完成的。政府享有本发明的某些权利。
对序列表的引用
本申请是与电子格式的序列表一起提交的。序列表以2018年11月14日创建的名为“sequence_listing_078430-503001wo.txt”的文件提供,其大小为约32kb。将在电子格式的序列表中的信息通过引用以其整体并入本文。
本文尤其公开了一种展现出对il-2受体信号的部分激动特性的白介素-2突变蛋白,以及使用所述白介素-2突变蛋白治疗自身免疫性疾病的方法。
背景技术:
白介素2(il-2)是主要由激活的cd4+t细胞产生的多能细胞因子,其在产生正常免疫应答中起着至关重要的作用。il-2促进激活的t淋巴细胞的增殖和扩增、增强b细胞生长,并且激活单核细胞和自然杀伤细胞。
il-2除了在正常免疫应答中的生理作用外,还可以促进病理反应,并且治疗目标是在阻断不希望的自身免疫或免疫抑制反应的同时维持该细胞因子的所需作用。两种针对人il-2rα的单克隆抗体(mab)达克珠单抗(daclizumab)和巴利昔单抗(basiliximab)被fda批准并且展现出在肾移植排斥(vincenti等人,nengljmed338:161(1998))、心脏移植(hershberger等人,nengljmed352:2705(2005))、多发性硬化(gold等人,lancet381:2167(2013))和哮喘(bielekova等人,proc.natl.acad.sci.usa101:8705(2004);和busse等人,amjrespircritcaremed178:1002(2008))中的功效,但它们不是通过在nk和记忆cd8+细胞上表达的中等亲和力il-2rγ受体阻断il-2信号传导(tkaczuk等人,amjtransplant2:31(2002))。尽管抗人il-2rβmabmikβ1可以阻断il-2和il-15向表达il-2rγ受体的细胞的反式呈递(morris等人,proc.natl.acad.sci.usa103:401(2006)),但是它在阻断il-2或il-15通过它们的高亲和力异三聚受体复合物进行顺式信号传导方面相对无效(morris等人,proc.natl.acad.sci.usa103:401(2006);以及waldmann等人,blood121:476(2013))。因此,需要新的il-2突变蛋白,其可以促进该细胞因子的治疗有益效应,但是也阻断与不希望的自身免疫或免疫抑制反应相关的一种或多种il-2功能。
技术实现要素:
本公开文本总体上涉及免疫学和医学领域,包括用于调节有需要的受试者中由白介素2(il-2)介导的信号转导途径的组合物和方法。更具体地,在一些实施方案中,本公开文本提供了新型il-2突变蛋白,其对il-2受体(例如白介素2α受体(il-2rα)、白介素2β受体(il-2rβ)、白介素2γ受体(il-2rγ))中的至少一种具有经调节的亲和力。本公开文本的一些实施方案提供了il-2部分激动剂,其不促进引起不期望的不利自身免疫性事件(诸如炎症)的免疫细胞的激活。本公开文本的一些实施方案涉及可用于产生此类il-2突变蛋白的组合物和方法,以及用于治疗与由il-2信号传导途径介导的信号转导的扰动相关的健康病症和障碍的方法。
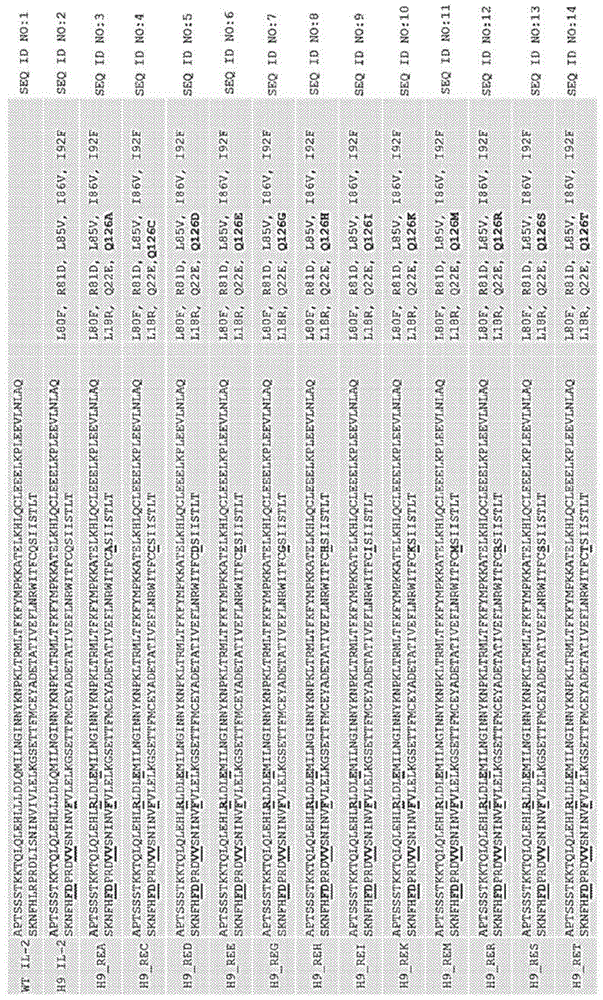
在一个方面,提供了一种白介素2(il-2)突变蛋白,其具有:(a)与由seqidno:2编码的il-2多肽相比降低的对白介素2受体γ(il-2rγ)的结合亲和力;和(b)与由seqidno:2编码的多肽相比15-95%e最大。在一些实施方案中,il-2突变蛋白包含:(i)与由seqidno:1编码的多肽相比增大il-2rβ结合亲和力的一种或多种氨基酸取代,所述一种或多种氨基酸取代选自根据seqidno:1的氨基酸序列编号的l80f、r81d、l85v、i86v和i92f;和/或(ii)与由seqidno:2编码的多肽相比降低il-2rγ受体结合亲和力并导致15-95%的e最大的一种或多种氨基酸取代,所述一种或多种氨基酸取代选自根据seqidno:2的氨基酸序列编号的(a)l18r和q22e;和(b)氨基酸位置126。在一些实施方案中,seqidno:2的位置126处的氨基酸取代选自q126a、q126c、q126d、q126e、q126g、q126h、q126i、q126k、q126m、q126r、q126s、或q126t。在一些实施方案中,il-2突变蛋白包含氨基酸取代q126h、q126k或q126m。在一些实施方案中,il-2突变蛋白包含氨基酸取代q126h。
在一些实施方案中,il-2突变蛋白在结构上经修饰以延长半衰期。在一些实施方案中,修饰包括选自以下的一种或多种修饰:与人fc抗体片段的融合、与白蛋白的融合和peg化。在一些实施方案中,il-2突变蛋白引起treg细胞的扩增,并且不促进潜在炎性t细胞和自然杀伤(nk)细胞或粒细胞的扩增。在一些实施方案中,与由seqidno:1或seqidno:2编码的多肽相比,il-2突变蛋白诱导较少的潜在炎性t细胞增殖。在一些实施方案中,与由seqidno:1或seqidno:2编码的多肽相比,il-2突变蛋白使treg细胞的增殖增加至少3倍和/或诱导更少的ifnγ分泌。在一些实施方案中,潜在炎性t细胞是cd4+ifnγ+t细胞,或者是cd8+ifnγ+t细胞。在本文公开的任何实施方案中的一些实施方案中,突变蛋白不诱导从cd8+t细胞和/或其他炎性免疫细胞子集分泌ifnγ。在一些实施方案中,与由seqidno:1编码的多肽相比,所述突变蛋白具有70-95%e最大。
在一个方面,本文提供了一种白介素2(il-2)突变蛋白,其具有与由seqidno:1编码的多肽相比:(a)降低的对白介素2受体γ(il-2rγ)的结合亲和力;以及(b)15-95%e最大。在一些实施方案中,突变蛋白包含与由seqidno:1编码的多肽相比降低il-2rγ受体结合亲和力并导致15-95%e最大的一种或多种氨基酸取代,所述一种或多种氨基酸取代选自根据seqidno:1的氨基酸序列编号的(a)l18r和q22e;和(b)氨基酸位置126。在一些实施方案中,il-2突变蛋白包含在seqidno:1的位置126处的氨基酸取代,所述氨基酸取代选自q126a、q126c、q126d、q126e、q126g、q126h、q126i、q126k、q126m、q126r、q126s、或q126t。在一些实施方案中,il-2突变蛋白包含氨基酸取代q126h、q126k或q126m。在一些实施方案中,il-2突变蛋白包含氨基酸取代q126h。在一些实施方案中,il-2突变蛋白在结构上经修饰以延长半衰期。在一些实施方案中,修饰包括选自以下的一种或多种修饰:与人fc抗体片段的融合、与白蛋白的融合和peg化。在一些实施方案中,本公开文本的il-2突变蛋白增加调节性t(treg)细胞的增殖和/或诱导潜在炎性t细胞的最小增殖。在一些实施方案中,本公开文本的il-2突变蛋白引起treg细胞的扩增,并且不促进潜在的炎性t细胞和自然杀伤(nk)细胞或粒细胞的扩增。在一些实施方案中,与由seqidno:1或seqidno:2编码的多肽相比,il-2突变蛋白诱导较少的潜在炎性t细胞增殖。在一些实施方案中,潜在炎性t细胞是cd4+ifnγ+t细胞,或者是cd8+ifnγ+t细胞。在一些实施方案中,潜在炎性t细胞是cd4+ifnγ+t细胞,或者是cd8+ifnγ+t细胞。在一些实施方案中,与由seqidno:1或seqidno:2编码的多肽相比,il-2突变蛋白使treg细胞的增殖增加至少3倍和/或诱导更少的ifnγ分泌。在一些实施方案中,突变蛋白不诱导从cd8+t细胞和/或其他炎性免疫细胞子集分泌ifnγ。在一些实施方案中,与由seqidno:1编码的多肽相比,本公开文本的il-2突变蛋白具有70-95%e最大。
在其他方面,本文提供了(i)编码本文公开的任何一种il-2突变蛋白的核酸;(ii)包含所述核酸的载体;(iii)包含所述载体或核酸的宿主细胞;以及(iv)无菌药物组合物,其包含本文公开的任何一种il-2突变蛋白和/或本文公开的任何核酸或载体以及药学上可接受的赋形剂。
在其他方面,本文还提供了注射器和包括注射器的导管,所述注射器和导管包含本文公开的任何一种il-2突变蛋白、本文公开的任何核酸或载体,和/或本文公开的任何药物组合物。
在又其他方面,本文提供了试剂盒,其包括:(i)本文公开的任何一种il-2突变蛋白;(ii)本文公开的任何一种核酸或载体;(iii)本文公开的任何一种注射器或导管;和/或本文公开的任何一种药物组合物,以及使用它们的书面说明书。
在另一方面,本文提供了治疗有需要的个体的自身免疫性疾病的方法,所述方法包括向所述个体给予治疗有效量的(i)本文公开的任何一种il-2突变蛋白;(ii)本文公开的任何一种核酸或载体;和/或(iii)本文公开的任何一种药物组合物。在一些实施方案中,自身免疫性疾病选自类风湿性关节炎、胰岛素依赖性糖尿病、溶血性贫血、风湿热、甲状腺炎、克罗恩病、重症肌无力、肾小球肾炎、自身免疫性肝炎、多发性硬化、斑秃、牛皮癣、白癜风、营养不良性大疱性表皮松解症、系统性红斑狼疮和移植物抗宿主病。在一些实施方案中,自身免疫性疾病是移植物抗宿主病。在本文公开的方法的一些实施方案中,所述方法还包括给予本文公开的任何il-2突变蛋白或药物组合物与将所述突变蛋白靶向特定细胞类型的抗体的组合。在一些实施方案中,细胞类型是调节性t(treg)细胞。在一些实施方案中,抗体与il-2突变蛋白共价或非共价连接。
在其他方面,本文提供了产生本文公开的任何一种突变蛋白的方法,所述方法包括在用于产生所述突变蛋白的合适条件下培养本文公开的任何宿主细胞。在其他实施方案中,所述方法还包括分离和/或纯化所述突变蛋白。在一些实施方案中,所述方法还包括在结构上修饰所述突变蛋白以延长半衰期。在其他实施方案中,所述方法还包括,所述修饰包括选自以下的一种或多种修饰:与人fc抗体片段的融合、与白蛋白的融合和peg化。
在一些方面,本文还提供了用于防止潜在炎性t细胞增殖和/或防止从cd8+t细胞或其他炎性免疫细胞子集分泌ifnγ的方法,所述方法包括使表达白介素2受体γ(il-2rγ)的细胞与本文公开的任何il-2突变蛋白接触。在其他实施方案中,潜在炎性t细胞是cd4+cd44+ifnγ+t细胞,或者是cd8+cd44+ifnγ+t细胞。在一些实施方案中,所述方法是体外、体内或离体进行的。在本文公开的任何实施方案中的一些实施方案中,il-2突变蛋白在结构上经修饰以延长半衰期。在本文公开的任何实施方案中的一些实施方案中,所述修饰是选自以下的一种或多种修饰:与人fc抗体片段的融合、与白蛋白的融合和peg化。
在另一方面,本文提供了用于减少调节性t(treg)细胞增殖的方法,所述方法包括使treg细胞与白介素2(il-2)突变蛋白接触,所述il-2突变蛋白具有:(i)与由seqidno:2编码的多肽相比降低的对白介素2受体γ(il-2rγ)的结合亲和力;和(ii)与由seqidno:2编码的多肽相比0-50%e最大。在一些实施方案中,il-2突变蛋白包含:(i)与由seqidno:1编码的多肽相比增大il-2rβ结合亲和力的一种或多种氨基酸取代,所述一种或多种氨基酸取代选自根据seqidno:1的氨基酸序列编号的l80f、r81d、l85v、i86v和i92f;和(ii)与由seqidno:2编码的多肽相比降低il-2rγ受体结合亲和力并导致0-50%的e最大的一种或多种氨基酸取代,所述一种或多种氨基酸取代选自根据seqidno:2的氨基酸序列编号的(a)l18r和q22e;和(b)氨基酸位置126。
在另一方面,本文提供了用于减少调节性t(treg)细胞增殖的方法,所述方法包括使treg细胞与il-2突变蛋白接触,所述il-2突变蛋白具有:(i)与由seqidno:1编码的多肽相比降低的il-2rγ结合亲和力;和(ii)与由seqidno:1编码的多肽相比0-50%e最大。在一些实施方案中,il-2突变蛋白包括与由seqidno:1编码的多肽相比降低il-2rγ受体结合亲和力并导致0-50%e最大的一种或多种氨基酸取代,所述一种或多种氨基酸取代选自根据seqidno:1的氨基酸序列编号的(a)l18r和q22e;和(b)氨基酸位置126。在一些实施方案中,seqidno:1或seqidno:2的位置126处的氨基酸取代选自q126a、q126c、q126d、q126e、q126g、q126h、q126i、q126k、q126m、q126r、q126s、或q126t。在一些实施方案中,il-2突变蛋白包含氨基酸取代q126h、q126k或q126m。在一些实施方案中,il-2突变蛋白包含氨基酸取代q126h。在本文公开的任何实施方案中的一些实施方案中,所述方法还包括将il2突变蛋白与抗体一起给予,所述抗体将所述突变蛋白靶向treg细胞。在本文公开的任何实施方案中的一些实施方案中,所述抗体与所述突变蛋白共价或非共价连接。在本文公开的任何实施方案中的一些实施方案中,所述方法是体外、体内或离体进行的。
除非从实施方案或方面的上下文中明确或清楚地排除,否则本文所述的每个方面和实施方案都能够一起使用。
前述概述仅是说明性的,而无意以任何方式进行限制。除了本文描述的说明性实施方案和特征之外,根据附图、详细描述和权利要求书,本公开文本的其他方面、实施方案、目的和特征也将变得完全显而易见。
附图说明
图1描绘了野生型il-2和使用h9背景构建的突变蛋白的氨基酸序列。
图2描绘了野生型il-2和使用野生型背景构建的突变蛋白的氨基酸序列。
图3描绘了具有il-2变体的剂量反应的磷酸-stat5信号传导测定的结果。用如图所示不同浓度(从1μm至0μm,10倍稀释)的与小鼠血清白蛋白(msa)融合的各种人il-2变体将人nk样yt细胞刺激15min。y轴显示在每种浓度下归一化的来自每种il-2变体的p-stat5信号与来自wtil-2的p-stat5信号的比率。
图4a至图4b描绘了磷酸-stat5信号传导测定的时间过程。图4a描绘了对用指示浓度的il-2部分激动剂刺激15min的yt细胞进行的p-stat5信号传导测定的结果。y轴显示在每种浓度下归一化的来自每种il-2变体的p-stat5信号与来自wtil-2的p-stat5信号的比率。图4a描绘了如图所示在各个时间点用1μm的不同il-2变体刺激yt细胞。y轴表示p-stat5信号的中位荧光强度(mfi)。
图5a描绘了用于在小鼠中给予il2激动剂的实验方案。图5b、图5c和图5d描绘了每种条件下所示免疫细胞子集的频率,所述频率被归一化为经pbs处理的小鼠中的相应频率。b细胞由在cd3-cd19+上门控的细胞定义。nk细胞在cd3-nk1.1+上门控。在ly6g(gr1)+cd3+cd11b+上门控的细胞被定义为粒细胞。
图6a至图6b描绘了具有il-2变体的剂量反应的磷酸-stat5信号传导测定的结果。图6a描绘了人nk样yt细胞,而图6b描绘了小鼠饥饿t原始细胞(b),所述人nk样yt细胞和小鼠饥饿t原始细胞用如图所示不同浓度(从5μm至0μm)的与小鼠血清白蛋白(msa)融合的各种人il-2变体刺激15min。y轴显示在每种浓度下归一化的来自每种il-2变体的p-stat5信号与来自wtil-2的p-stat5信号的比率。
图7a至图7e描绘了在b16黑色素瘤小鼠模型上进行的il-2变体给予的结果。图7a描绘了肿瘤体积的变化。图7b、图7c、图7d和图7e描绘了每种条件下所示免疫细胞子集的频率,所述频率被归一化为wt小鼠中的相应频率。
图8a至图8g以图形方式汇总了为证明几种示例性il-2r部分激动剂可以在体内引起细胞类型特异性应答而进行的实验的结果。
图9a至图9c以图形方式汇总了为说明il-2r部分激动剂reh增加foxp3+调节性t细胞的频率而进行的实验的结果。
图10是为表明来自用il-2r部分激动剂reh处理过的小鼠的treg抑制cd4+常规t细胞的增殖而进行的实验的结果的图形汇总。
图11是为说明il-2r部分激动剂reh支持cd8+t细胞增殖,但不支持ifnγ产生而进行的实验的结果的图形汇总。
图12是为表明用il-2r部分激动剂reh进行预处理和共处理在啮齿动物eae模型(脑部炎症的动物模型)中防止各种自身免疫性症状而进行的实验的结果的图形汇总。疾病得分如下:0-健康;1-尾巴无力;2-部分后肢瘫痪;3-完全后肢瘫痪;4-全身瘫痪;5-死亡。
具体实施方式
本公开文本总体上涉及免疫学和医学领域,包括用于调节有需要的受试者中由白介素2(il-2)介导的信号转导途径的组合物和方法。更具体地,在一些实施方案中,本公开文本提供了新型il-2突变蛋白,其对il-2受体(例如白介素2α受体(il-2rα)、白介素2β受体(il-2rβ)、白介素2γ受体(il-2rγ))中的至少一种具有经调节的亲和力,借此完全或部分拮抗通过相应的il-2rα、il-2rβ和/或il-2rγ受体介导的下游信号转导。本公开文本的一些实施方案提供了il-2部分激动剂,其不促进引起不期望的不利自身免疫性事件(诸如炎症)的免疫细胞的激活。本公开文本的一些实施方案涉及可用于产生此类il-2突变蛋白的组合物和方法,以及用于治疗与由il-2信号传导途径介导的信号转导的扰动相关的健康病症和障碍的方法。
il-2对免疫系统具有广谱效应,并且其在调节免疫激活和体内平衡方面起着至关重要的作用。作为免疫系统刺激物,il-2已用于治疗癌症和慢性病毒感染。然而,il-2的刺激效应也可引起破坏,从而介导自身免疫和移植排斥。由于il-2分子在免疫调节和疾病中的有用作用,新的il-2分子(诸如il-2部分激动剂)的鉴定仍然是活跃的研究领域。
在大多数情况下,il-2通过以下三种不同的受体起作用:il-2rα、il-2rβ和il-2rγ。大多数细胞(例如静息t细胞)对il-2无反应,因为它们仅表达对il-2具有低亲和力的il-2rβ和il-2rγ。在刺激时,静息t细胞表达相对较高亲和力的il-2rα。il-2与il-2rα的结合导致该受体依次与il-2rβ和il-2rγ接合,从而引发t细胞激活。
本文所述的公开内容尤其提供了新型il-2组合物,所述新型il-2组合物基于对il-2如何与其同源受体,特别是il-2rγ相互作用的新见解。发明人已经惊奇地发现,il-2rγ的il-2结合位点的突变产生了能够激活调节性t细胞(treg)的部分激动剂。此外,这些il-2部分激动剂不会激活cd8+t细胞和其他潜在炎性免疫细胞子集,并且不会诱导炎性细胞分泌干扰素-γ(ifnγ)。如此,这些分子可用于治疗自身免疫性障碍和病症。
i.一般技术
除非另有说明,否则本公开文本的实践将采用本领域技术人员众所周知的分子生物学、微生物学、细胞生物学、生物化学、核酸化学和免疫学的常规技术。此类技术在文献中进行了充分的解释,所述文献如molecularcloning:alaboratorymanual,第四版(sambrook等人,2012)和molecularcloning:alaboratorymanual,第三版(sambrook和russel,2001),(在本文中统称为“sambrook”);currentprotocolsinmolecularbiology(f.m.ausubel等人编辑,1987,包括到2014年的增补);pcr:thepolymerasechainreaction,(mullis等人编辑,1994);beaucage等人编辑,currentprotocolsinnucleicacidchemistry,johnwiley&sons,inc.,纽约,2000(包括至2014年的增补),genetransferand.expressioninmammaliancells(makrides编辑,elseviersciencesb.v.,阿姆斯特丹,2003),以及currentprotocolsinimmunology(horgank和s.shaw(1994))(包括至2014年的增补)。视情况而定,除非另有说明,否则通常根据制造商定义的方案和/或参数进行涉及市售试剂盒和试剂的使用的程序。
ii.定义
除非另有定义,否则本文中使用的所有领域术语、符号和其他科学术语或用辞旨在具有本公开文本所属领域的技术人员通常理解的含义。在一些情况下,为了清楚和/或为了便于引用而在本文中定义具有通常理解的含义的术语,并且本文中包含此类定义不一定应当被解释为表示与本领域通常所理解的含义有实质性差异。本领域技术人员很好地理解并通常使用常规方法采用本文描述或提及的许多技术和程序。
除非上下文另外明确指出,否则单数形式“一个/一种(a)”、“一个/一种(an)”和“所述”包括复数指示物。例如,术语“一种细胞”包括一种或多种细胞,包括其混合物。本文使用“a和/或b”来包括所有以下替代方案:“a”、“b”、“a或b”以及“a和b”。
如本文所使用的,术语“约”具有其近似的普通含义。如果上下文没有另外明确近似程度,在包括所提供的值的所有情况下,“约”表示在所提供的值的加或减10%内,或四舍五入到最接近的有效数字。在提供范围的情况下,所述范围包括边界值。
如本文中所使用的,术语“il-2”是指野生型il-2,无论是天然的还是重组的。因此,il-2多肽是指任何il-2多肽,包括但不限于重组产生的il-2多肽、合成产生的il-2多肽、从细胞或组织提取的il-2。成熟的人il-2作为133个氨基酸的序列出现(减去信号肽,所述信号肽由另外的20个n末端氨基酸组成),如在fujita等人,proc.natl.acad.sci.usa,80,7437-7441(1983)中所述。在genbank中的登录定位器(accessionlocator)np_000577.2下找到了人il-2的氨基酸序列(seqidno:17)。成熟的人il-2的氨基酸序列描绘于seqidno:1中。在genbank中的登录号下发现了鼠(小家鼠(musmusculus))il-2氨基酸序列(seqidno:18)。成熟鼠il-2的氨基酸序列描绘于seqidno:19中。
如本文所用,“il-2突变蛋白”是指其中已经对白介素-2蛋白进行了特定取代的il-2多肽。il-2突变蛋白的特征在于在天然il-2多肽链中的一个或多个位点处或在天然il-2多肽链的其他残基处的氨基酸插入、缺失、取代和修饰。根据本公开文本,任何此类插入、缺失、取代和修饰导致保留了il-2r结合活性的il-2突变蛋白。例如,本文公开的突变蛋白可以具有对il-2rα和/或il-2rβ的高或低亲和力,或者对这些受体的亲和力可以与野生型il-2相同或相似。示例性突变蛋白可包括对1个、2个、3个、4个、5个、6个、7个、8个、9个、10个或更多个氨基酸的取代。突变蛋白还可以在il-2的其他位置处包含保守的修饰和取代(即,对突变蛋白的二级或三级结构具有最小影响的那些修饰和取代)。此类保守取代包括以下文献中所述的那些:dayhoff,theatlasofproteinsequenceandstructure5(1978)和argos,emboj,8:779-785(1989)。例如,属于以下组之一的氨基酸表示保守变化:组i:ala、pro、gly、gln、asn、ser、thr;组ii:cys、ser、tyr、thr;组iii:val、ile、leu、met、ala、phe;组iv:lys、arg、his;组v:phe、tyr、trp、his;以及组vi:asp、glu。
短语“根据……编号”是指参照所选氨基酸在给定氨基酸序列(诸如,野生型il-2的氨基酸序列)中通常存在的位置来鉴定该氨基酸。例如,r81是指在seqidno:1中存在的第八十一个氨基酸精氨酸。
如本文所用的关于多肽或dna序列的术语“同一性”是指两个分子之间的亚基序列同一性。当两个分子中的某一亚基位置都被相同的单体亚基(即,相同的氨基酸残基或核苷酸)占据时,则所述分子在该位置是相同的。两个氨基酸或两个核苷酸序列之间的相似性是相同位置数的直接函数。通常,将序列进行比对,以便获得最高阶匹配。如有必要,可以使用已公布的技术和广泛可用的计算机程序来计算同一性,所述计算机程序如gcs程序包(devereux等人,nucleicacidsres.12:387,1984)、blastp、blastn、fasta(atschul等人,j.molecularbiol.215:403,1990)。可以使用序列分析软件以所述软件的默认参数来测量序列同一性,所述序列分析软件如universityofwisconsinbiotechnologycenter(威斯康星州麦迪逊,大学大道1710号,53705)的geneticscomputergroup的序列分析软件包。
如本文所用,术语“蛋白质”和“多肽”是指氨基酸残基的聚合物,并且不限于最小长度的产物。因此,所述定义内涵盖了多肽、肽、多肽片段、融合多肽、寡肽等。所述定义涵盖了全长蛋白质及其片段两者。所述术语还包括多肽的表达后修饰,例如糖基化、乙酰化、磷酸化等。此外,出于本公开文本的目的,“多肽”是指包含对天然序列的修饰如缺失、添加和取代(通常是保守性)的蛋白质,只要所述蛋白质保持所需活性即可。这些修饰可能是故意的(如通过定点诱变),或者可能是偶然的(诸如通过产生蛋白质的宿主的突变或由于pcr扩增引起的错误)。因此,除非从实施方案或方面的上下文中明确或清楚地排除,否则术语“il-2多肽”是指天然il-2序列,以及il-2类似物、il-2突变蛋白和片段。
如本文所用,“潜在炎性t细胞”或“炎性免疫细胞子集”是指产生、分泌或能够产生或分泌干扰素γ的一种或多种t细胞。在一些实施方案中,潜在炎性t细胞在其细胞表面上表达cd44,并产生或分泌干扰素γ。在其他实施方案中,潜在炎性t细胞在其细胞表面上表达cd44和cd4,并产生或分泌干扰素γ。在其他实施方案中,潜在炎性t细胞在其细胞表面上表达cd44和cd8,并产生或分泌干扰素γ。在又其他实施方案中,潜在炎性t细胞在其细胞表面上表达cd4和cd8,并产生或分泌干扰素γ。
如本文所使用的,“调节性t细胞”或“treg细胞”是指调节其他一种或多种t细胞和/或其他免疫细胞的活性的t细胞(t淋巴细胞),通常通过抑制其活性来调节。在一些实施方案中,treg细胞是cd4+和foxp3+细胞(但本领域技术人员应理解,treg细胞并不完全局限于该表型)。
如本文所提及的“e最大”是在最高浓度下测量的可以由il-2突变蛋白产生的最大p-stat5信号。将来自wtil-2的e最大用作参考,以计算在最高浓度下归一化的来自il-2突变蛋白的p-stat5信号与来自wtil-2的p-stat5信号的比率。
“激动剂”是与靶标相互作用以引起或促进靶标激活增加的化合物。
“部分激动剂”是化合物,与其相互作用的靶标与激动剂的靶标相同,但是即使通过增加所述部分激动剂的剂量,它也不会产生与激动剂一样大的生化和/或生理作用的量值。
“超级激动剂”是一类激动剂,其能够产生比内源激动剂更大的对靶受体的最大反应,并且因此具有大于100%的功效。
“拮抗剂”是例如通过预防、减少、抑制或中和激动剂的活性来对抗激动剂的作用的化合物。即使没有确定的激动剂,“拮抗剂”也可以预防、抑制或降低靶标(例如,靶受体)的组成性活性。
“可操作地连接”意图意指,目的核苷酸序列(即,编码il-2突变蛋白的序列)以允许表达核苷酸序列的方式与一种或多种调节序列连接(例如,在体外转录/翻译系统中,或在将载体引入宿主细胞时在所述宿主细胞中)。“调节序列”包括启动子、增强子和其他表达控制元件(例如,聚腺苷酸化信号)。参见例如goeddel(1990)的geneexpressiontechnology:methodsinenzymology第185卷(加利福尼亚州圣地亚哥的academicpress)。调节序列包括在许多类型的宿主细胞中引导核苷酸序列的组成型表达的那些,和仅在某些宿主细胞中引导核苷酸序列的表达的那些(例如组织特异性调节序列)。本领域的技术人员应理解,表达载体的设计可以取决于诸如以下的因素:待转化的宿主细胞的选择、所需蛋白质的表达水平等。可将本公开文本的表达构建体引入宿主细胞中,从而产生本文公开的人il-2突变蛋白或产生其生物学活性变体。
术语“宿主细胞”和“重组宿主细胞”在本文中可互换使用。应当理解,此类术语不仅指特定的受试者细胞,而且还指这种细胞的子代或潜在子代。因为在后代中可能由于突变或环境影响而发生某些修饰,这种子代可能实际上与亲代细胞不同,但是仍然包括在如本文使用的所述术语的范围内。
术语“载体”在本文中用于指能够转移或转运另一种核酸分子的核酸分子或序列。转移的核酸通常与载体核酸分子连接,例如插入所述载体核酸分子中。载体可以包含引导细胞中自主复制的序列,或者可以包括足以允许整合到宿主细胞dna中的序列。有用的载体包括例如质粒(例如,dna质粒或rna质粒)、转座子、粘粒、细菌人工染色体和病毒载体。有用的病毒载体包括例如复制缺陷型逆转录病毒和慢病毒。在一个方面,载体是基因递送载体。在一个方面,将载体用作基因递送媒介物以将基因转移到细胞中。
如本文所用,术语“转化”和“转染”是指用于将外来核酸(例如,dna)引入宿主细胞中的多种本领域公认的技术,包括磷酸钙或氯化钙共沉淀、deae-葡聚糖介导的转染、脂转染、粒子枪或电穿孔。
如本文所用,“受试者”或“个体”或“患者”包括动物,诸如人(例如,人受试者)和非人动物。术语“非人动物”包括所有脊椎动物,例如哺乳动物(例如,啮齿动物,例如小鼠)和非哺乳动物(例如非人灵长类动物),例如绵羊、狗、牛、鸡、两栖动物、爬行动物等。
如本文所用,术语“药学上可接受的载体”包括但不限于与药物给予相容的盐水、溶剂、分散介质、包衣、抗细菌和抗真菌剂,等渗和吸收延迟剂等。也可以将补充性活性化合物(例如,抗生素)掺入组合物中。
应理解,本文描述的本公开文本的方面和实施方案包括“包含(comprising)方面和实施方案”,“由方面和实施方案组成(consisting)”,以及“基本上由方面和实施方案组成(consistingessentiallyof)”。
如本领域的普通技术人员将理解的,出于任何和所有目的,诸如就提供书面描述方面而言,本文公开的所有范围还涵盖任何和所有可能的子范围以及其子范围的组合。任何所列范围都可容易地识别为充分描述相同范围并且使得相同范围能分解为至少相等的二分之一、三分之一、四分之一、五分之一、十分之一等。作为非限制性例子,本文中论述的每个范围可容易地分解为下三分之一、中三分之一和上三分之一等。同样如本领域技术人员可理解,所有诸如“至多”、“至少”、“大于”、“小于”等言辞都包括所述数字并且涉及随后可如上所述分解为子范围的范围。最后,本领域技术人员将理解,范围包括每个单独的成员。因此,例如,具有1-3个制品的组是指具有1、2或3个制品的组。类似地,具有1-5个制品的组是指具有1、2、3、4或5个制品的组等等。
提供标题(例如,(a)、(b)、(i)等)只是为了便于阅读说明书和权利要求书。说明书或权利要求书中的标题的使用不要求按字母或数字顺序或呈现顺序进行步骤或元件。
ⅲ.本公开文本的组合物
本公开文本的一些方面涉及与野生型il-2多肽相比对一种或多种il-2受体(例如,il-2rα、il-2rβ和/或il-2rγ)具有经调节的亲和力,例如升高或降低的亲和力的新型il-2突变蛋白。具体地,本公开文本的一些实施方案涉及l-2突变蛋白,其中一种或多种分子改变赋予了对il-2rα、il-2rβ和/或il-2rγ的降低的结合亲和力。换句话说,本文公开的新型il-2突变蛋白中的一些是il-2介导的信号传导途径的部分激动剂。在一些实施方案中,本文公开的il-2部分激动剂不促进引起不期望的不利自身免疫性事件(诸如炎症)的免疫细胞的激活。
a.il-2部分激动剂
在一个方面,本公开文本的一些实施方案提供作为部分激动剂的il-2突变蛋白。在一些实施方案中,本文提供了il-2突变蛋白,所述il-2突变蛋白含有与野生型il-2(例如,人il-2,seqidno:2)相比降低il-2突变蛋白对il-2rγc受体的结合亲和力的一种或多种突变。如本文所用,术语“共有γ链”、“γc”、“il-2rγc”、“yc”、“il-2rγ”、“il-2受体亚基γ”和“il-2rg”(genbank登录号:nm_000206和np_000197(人)以及nm_013563和np_038591(小鼠))都是指i型细胞因子受体家族的成员,所述成员是至少六种不同白介素受体(包括但不限于il-2、il-4、il-7、il-9、il-15和il-21受体)的受体复合物的细胞因子受体亚基。il-2rγc与il-2rβ相互作用以形成主要在记忆t细胞和自然杀伤(nk)细胞上的中等亲和力il-2受体,并且与il-2rα和il-2rβ相互作用以形成在激活t细胞和treg上的高亲和力il-2受体。
在一些实施方案中,如本文所公开的il-2突变蛋白是人工重组il-2突变蛋白,并且可以是例如任何重组il-2多肽、经工程化的il-2多肽或天然存在的il-2多肽,所述il-2突变蛋白具有经调节的对il-2受体(例如,il-2rα、il-2rβ和/或il-2rγ)的结合亲和力。
示例性的主题il-2突变蛋白与对应的野生型il-2(wtil-2)至少约50%、至少约65%、至少约70%、至少约80%、至少约85%、至少约87%、至少约90%、至少约91%、至少约92%、至少约93%、至少约94%、至少约95%、至少约96%、至少约97%、至少约98%或至少约99%相同。突变可以由氨基酸残基的数量或含量的变化组成。例如,突变型il-2可以具有比对应的野生型il-2更大或更小的氨基酸残基数量。可替代地或另外地,示例性突变型多肽可含有在野生型il-2中存在的一个或多个氨基酸残基的取代。在各种实施方案中,突变型il-2多肽与野生型il-2的不同之处可以在于单一氨基酸残基的添加、缺失或取代。
通过非限制性说明,包含与参考氨基酸序列seqidno:1至少95%相同的氨基酸序列的il-2突变蛋白是如下多肽:包含与参考序列相同的序列,但包含seqidno:2的参考氨基酸序列的至多五个改变。因此,在一些实施方案中,本公开文本的il-2突变蛋白包含与参考氨基酸序列seqidno:1至少95%相同的氨基酸序列,是如下多肽:包含与参考序列相同的序列,但是包含参考氨基酸序列的1个、2个、3个、4个或5个改变。例如,参考序列中至多5%的氨基酸残基可以被缺失或用另一个氨基酸取代,或者可以将参考序列中总氨基酸残基的至多5%数量的氨基酸插入所述参考序列中。参考序列的这些改变可以发生在参考氨基酸序列的氨基(n-)或羧基(c-)末端位置或那些末端位置之间的任何位置,单独地散布在参考序列中的残基之间,或者散布在参考序列内的一个或多个连续组中。
在一些实施方案中,本公开文本的il-2突变蛋白以如下亲和力结合il-2rγ:比野生型il-2低至少1%、2%、3%、4%、5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%,包含落在这些百分比之间的任何值。il-2突变蛋白的结合亲和力还可以表示为比野生型il-2低1.2倍、1.4倍、1.5倍、2倍、5倍、10倍、15倍、20倍、25倍、50倍、100倍、200倍、250倍或更多倍的对il-2γ的亲和力。可以使用本领域已知的任何合适的方法来测量主题il-2突变蛋白对il-2rγ的结合亲和力。用于测量il-2rγ结合的合适方法包括但不限于放射性配体结合测定(例如,饱和结合、scatchard作图、非线性曲线拟合程序和竞争结合测定);非放射性配体结合测定(例如,荧光偏振(fp)、荧光共振能量转移(fret)和表面等离振子共振测定(参见例如,drescher等人,methodsmolbiol493:323-343(2009));液相配体结合测定(例如,实时聚合酶链反应(rt-qpcr)和免疫沉淀);以及固相配体结合测定(例如,多孔板测定、珠上配体结合测定、柱上配体结合测定和过滤器测定)。不受理论的束缚,据信通过il-2rγ受体结合位点中的突变构建的部分激动剂与il-2的野生型或其他变体相比会具有更低剂量依赖性。
在一些实施方案中,il-2突变蛋白破坏il-2rβ与il-2rγ的缔合,使得该il-2rβ/il-2rγ相互作用相对于野生型il-2减少约2%、约5%、约10%、约15%、约20%、约50%、约75%、约90%、约95%或更多(包含落入这些百分比之间的任何值)。
在一些实施方案中,降低il-2突变蛋白对il-2rγ受体的结合亲和力的一种或多种突变是氨基酸取代。在一些实施方案中,与野生型il-2(seqidno:1)相比,主题il-2突变蛋白由1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个、12个、13个、14个或15个氨基酸取代组成。一个或多个经取代的氨基酸残基可以是但不一定是保守取代,所述取代通常包括以下组内的取代:甘氨酸、丙氨酸;缬氨酸、异亮氨酸、亮氨酸;天冬氨酸、谷氨酸;天冬酰胺、谷氨酰胺;丝氨酸、苏氨酸;赖氨酸、精氨酸;以及苯丙氨酸、酪氨酸。在特定的实施方案中,取代位于il-2中与il-2rγ结合界面接触的氨基酸残基处。
在一些实施方案中,氨基酸取代是在野生型il-2中选自位置18、22和126的一个或多个氨基酸位置处的取代,根据野生型hil-2(例如,seqidno:1)编号。在一些实施方案中,降低il-2rγ受体结合亲和力的氨基酸取代包括氨基酸取代leu至arg(l18r)、gln至glu(q22e),和/或gln至his(q126h)、gln至met(q126m)或gln至lys(q126k)中的一者,或其组合。
在其他实施方案中,降低il-2rγ受体结合亲和力的氨基酸取代包括l18r和q22e,以及q126a、q126c、q126d、q126e、q126g、q126h、q126i、q126k、q126m、q126r、q126s或q126t中的任一者。在又其他实施方案中,il-2突变蛋白可以具有在野生型il-2中选自位置80、81、85、86和192的一个或多个氨基酸位置处的另外的氨基酸取代,根据野生型hil-2(例如,seqidno:1)编号。在另一个实施方案中,il-2突变蛋白可以具有选自l80f、r81d、l85v、i86v和/或i92f中的一个或多个的另外的氨基酸取代。
在一些实施方案中,il-2突变蛋白还可以具有增大的对il-2rβ受体的结合亲和力,并且还可包含增大il-2rβ结合亲和力的1个、2个、3个、4个、5个、6个、7个、8个、9个或10个或更多个突变。如本文所用,术语“il-2rβ”和“cd122”(genbank登录号nm_000878和np_000869(人))均指i型细胞因子受体家族的成员,所述成员与il-2rγ相互作用以形成主要在记忆t细胞和自然杀伤(nk)细胞上的中等亲和力il-2受体,并且与il-2rα和il-2rγ相互作用以形成在激活t细胞和调节性t细胞(treg)上的高亲和力il-2受体。在一些实施方案中,主题il-2突变蛋白包含相对于野生型il-2(例如,seqidno:1)的至少一种突变(例如,1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个、12个、13个、14个、15个、16个、17个、18个、19个、20个或更多个氨基酸残基的缺失、添加或取代),并以高于野生型il-2的亲和力结合il-2rβ。在一些实施方案中,il-2突变蛋白以如下亲和力结合il-2rβ:比野生型il-2高至少1%、2%、3%、4%、5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%(包含落在这些百分比之间的任何值)。il-2突变蛋白的结合亲和力还可以表示为比野生型il-2高1.2倍、1.4倍、1.5倍、2倍、5倍、10倍、15倍、20倍、25倍、50倍、100倍、200倍、250倍或更多倍的对il-2rβ的亲和力。主题il-2突变蛋白与il-2rβ的结合可以通过本领域技术人员已知的任何合适的方法(包括但不限于上述方法)来评估。在一些实施方案中,增大il-2rβ结合亲和力的至少一种突变是氨基酸取代。在一些实施方案中,增大il-2rβ结合亲和力的氨基酸取代包括在根据野生型hil-2(seqidno:1)编号的氨基酸位置i24、p65、q74、l80、r81、l85、i86、i89、i92和/或v93处的取代。在一些实施方案中,所述取代包括i24v、p65h、q74r、q74h、q74n、q74s、l80f、l80v、r81i、r81t、r81d、l85v、i86v、i89v、i92f和/或v93i,或其组合。在一些实施方案中,取代包括q74n、q74h、q74s、l80f、l80v、r81d、r81t、l85v、i86v、i89v和/或i93v,或其组合。
在一些实施方案中,il-2突变蛋白还可具有降低的对il-2rβ受体的结合亲和力,并且还可包含降低il-2rβ结合亲和力的1个、2个、3个、4个、5个、6个、7个、8个、9个或10个或更多个突变。在一些实施方案中,主题il-2突变蛋白包含相对于野生型il-2(例如,seqidno:1)的至少一种突变(例如,1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个、12个、13个、14个、15个、16个、17个、18个、19个、20个或更多个氨基酸残基的缺失、添加或取代),并以与野生型il-2相比降低的亲和力结合il-2rβ。在一些实施方案中,il-2突变蛋白以如下亲和力结合il-2rβ:比野生型il-2低至少1%、2%、3%、4%、5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%(包含落在这些百分比之间的任何值)。il-2突变蛋白的结合亲和力还可以表示为比野生型il-2低1.2倍、1.4倍、1.5倍、2倍、5倍、10倍、15倍、20倍、25倍、50倍、100倍、200倍、250倍或更多倍的对il-2rβ的亲和力。
在其他实施方案中,本文公开的任何il-2突变蛋白都可以以与野生型il-2(例如,seqidno:1)相似(例如,相差小于百分之一)或相同的亲和力结合至il-2rβ受体。
在另外的实施方案中,il-2突变蛋白还可具有增大的对il-2rα受体的结合亲和力,并且还可包含增大il-2rα结合亲和力的1个、2个、3个、4个、5个、6个、7个、8个、9个或10个或更多个突变。如本文所用,术语“il-2rα”和“cd25”(genbank登录号nm_000417和np_000408(人))均指i型细胞因子受体家族的成员,所述成员与il-2rβ和il-2rγ相互作用以形成在激活的t细胞和调节性t细胞(treg)上的高亲和力il-2受体。在一些实施方案中,主题il-2突变蛋白包含相对于野生型il-2(例如,seqidno:1)的至少一种突变(例如,1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个、12个、13个、14个、15个、16个、17个、18个、19个、20个或更多个氨基酸残基的缺失、添加或取代),并以高于野生型il-2的亲和力结合il-2rα。在一些实施方案中,il-2突变蛋白以如下亲和力结合il-2rα:比野生型il-2高至少1%、2%、3%、4%、5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%(包含落在这些百分比之间的任何值)。il-2突变蛋白的结合亲和力还可以表示为比野生型il-2高1.2倍、1.4倍、1.5倍、2倍、5倍、10倍、15倍、20倍、25倍、50倍、100倍、200倍、250倍或更多倍的对il-2rβ的亲和力。主题il-2突变蛋白与il-2rα的结合可以通过本领域技术人员已知的任何合适的方法(包括但不限于上述方法)来评估。在一些实施方案中,增大il-2rβ结合亲和力的至少一种突变是氨基酸取代。
在又其他实施方案中,il-2突变蛋白还可具有降低的对il-2rα受体的结合亲和力,并且还可包含降低il-2rα结合亲和力的1个、2个、3个、4个、5个、6个、7个、8个、9个或10个或更多个突变。在一些实施方案中,主题il-2突变蛋白包含相对于野生型il-2(例如,seqidno:1)的至少一种突变(例如,1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个、12个、13个、14个、15个、16个、17个、18个、19个、20个或更多个氨基酸残基的缺失、添加或取代),并以低于野生型il-2的亲和力结合il-2rα。在一些实施方案中,il-2突变蛋白以如下亲和力结合il-2rα:比野生型il-2低至少1%、2%、3%、4%、5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%、96%、97%、98%或99%(包含落在这些百分比之间的任何值)。il-2突变蛋白的结合亲和力还可以表示为比野生型il-2低1.2倍、1.4倍、1.5倍、2倍、5倍、10倍、15倍、20倍、25倍、50倍、100倍、200倍、250倍或更多倍的对il-2rα的亲和力。
在其他实施方案中,本文公开的任何il-2突变蛋白都可以以与野生型il-2(例如,seqidno:1)相似(例如,相差小于百分之一)或相同的亲和力结合至il-2rα受体。
在一些实施方案中,il-2突变蛋白包括氨基酸取代l80f、r81d、l85v、i86v、i92f、l18r、q22e和q126h。在一些实施方案中,il-2突变蛋白具有以下氨基酸序列:
aptssstkktqlqlehlrldlemilnginnyknpkltrmltfkfympkkatelkhlqcleeelkpleevlnlaqsknfhfdprdvvsninvfvlelkgsettfmceyadetativeflnrwitfchsiistlt(seqidno:8)
在其他实施方案中,il-2突变蛋白包括氨基酸取代l80f、r81d、l85v、i86v、i92f、l18r、q22e和q126k。在一些实施方案中,il-2突变蛋白具有以下氨基酸序列:
aptssstkktqlqlehlrldlemilnginnyknpkltrmltfkfympkkatelkhlqcleeelkpleevlnlaqsknfhfdprdvvsninvfvlelkgsettfmceyadetativeflnrwitfcksiistlt(seqidno:10)
在其他实施方案中,il-2突变蛋白包括氨基酸取代l80f、r81d、l85v、i86v、i92f、l18r、q22e和q126m。在一些实施方案中,il-2突变蛋白具有以下氨基酸序列:
aptssstkktqlqlehlrldlemilnginnyknpkltrmltfkfympkkatelkhlqcleeelkpleevlnlaqsknfhfdprdvvsninvfvlelkgsettfmceyadetativeflnrwitfcmsiistlt(seqidno:11)。
在另一个实施方案中,il-2突变蛋白包含氨基酸取代l18r、q22e和q126h。在一些实施方案中,il-2突变蛋白具有以下氨基酸序列:
aptssstkktqlqlehlrldlemilnginnyknpkltrmltfkfympkkatelkhlqcleeelkpleevlnlaqsknfhlrprdlisninvivlelkgsettfmceyadetativeflnrwitfchsiistlt(seqidno:15)
在另一个实施方案中,il-2突变蛋白包含氨基酸取代l18r、q22e和q126m。在一些实施方案中,il-2突变蛋白具有以下氨基酸序列:
aptssstkktqlqlehlrldlemilnginnyknpkltrmltfkfympkkatelkhlqcleeelkpleevlnlaqsknfhlrprdlisninvivlelkgsettfmceyadetativeflnrwitfcmsiistlt(seqidno:16)。
在各种实施方案中,主题il-2突变蛋白具有根据下式的氨基酸序列:
a-p-t-s-s-s-t-k-k-t-q-l-q-l-e-h-l-(x1)n-l-d-l-(x2)n-m-i-l-n-g-i-n-n-y-k-n-p-k-l-t-r-m-l-t-f-k-f-y-m-p-k-k-a-t-e-l-k-h-l-q-c-l-e-e-e-l-k-p-l-e-e-v-l-n-l-a-q-s-k-n-f-h-(x3)n-(x4)n-p-r-d-(x5)n-(x6)n-s-n-i-n-v-(x7)n-v-l-e-l-k-g-s-e-t-t-f-m-c-e-y-a-d-e-t-a-t-i-v-e-f-l-n-rw-i-t-f-c-(x13)n-s-i-i-s-t-l-t,其中:
每个n单独选自0或1;
x1是l(野生型)或r;
x2是q(野生型)或e;
x3是l(野生型)、f或v;
x4是r(野生型)、i、t或d;
x5是l(野生型)或v;
x6是i(野生型)或v;
x7是i(野生型)或f;
x13是q(野生型)或h、m、k、c、d、e、g、i、r、s或t(seqidno:20)。
在根据seqidno:20的il-2突变蛋白的一些实施方案中,至少在x1、x2、x3、x4、x5、x6、x7、x8、x9、x10、x11、x12或x13中的一处的氨基酸不是野生型氨基酸。在一些实施方案中,至少在x1、x2、x3、x4、x5、x6、x7、x8、x9、x10、x11、x12或x13中的两处、三处、四处、五处、六处、七处、八处、九处、十处、十一处、十二处、十三处或十四处的氨基酸不是野生型氨基酸。在一些实施方案中,il-2突变蛋白具有与seqidno:20的il-2突变蛋白的至少约95%、至少约96%、至少约97%、至少约98%、至少约99%或约100%的同源性。
在一些实施方案中,与野生型il-2相比,作为部分激动剂的主题il-2突变蛋白具有一种或多种降低的功能,例如(i)降低的e最大,(ii)降低的刺激依赖于il-2rβ/il-2rγ异二聚化的信号传导途径的能力,(iii)减少的来自潜在炎性免疫细胞的干扰素-γ(ifnγ)的产生和/或分泌,(iv)减少的潜在炎性t细胞的增殖。
在一些实施方案中,il-2突变蛋白具有降低的刺激一种或多种依赖于il-2rβ/il-2rγ异二聚化的信号传导途径的能力。在一些实施方案中,与野生型hil-2相比,主题il-2突变蛋白具有降低的刺激t细胞中stat5磷酸化的能力(参见例如,实施例1和图4a至图4b)。在一些实施方案中,il-2突变蛋白以如下水平刺激t细胞中的stat5磷酸化:是野生型il-2刺激相同细胞中的stat5磷酸化的水平的1%、5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%(包含这些百分比之间的值)。在一些实施方案中,t细胞是自然杀伤(nk)细胞。stat5信号传导可以例如通过使用本领域已知的任何合适方法依据stat5的磷酸化来测量。例如,可以使用对该分子的磷酸化形式具有特异性的抗体与流式细胞术分析组合来测量stat5磷酸化。
在其他实施方案中,与野生型il-2的e最大相比,本文公开的il-2突变蛋白具有降低的e最大。在一些实施方案中,与野生型il-2(诸如,由seqidno:1编码的多肽)相比,突变蛋白具有约15-95%中任一值的e最大,诸如约20-95%、约30-95%、约40-95%、约50-95%、约60-95%、约70-95%、约80-95%或约90-95%e最大。在一些实施方案中,与野生型il-2的e最大相比,本文公开的il-2突变蛋白具有15-95%e最大。在一些实施方案中,与野生型il-2的e最大相比,本文公开的il-2突变蛋白具有70-95%e最大。在另一个实施方案中,与野生型il-2(诸如,由seqidno:1编码的多肽)相比,本文公开的il-2突变蛋白具有以下中的任一值的e最大:约15%、16%、17%、18%、19%、20%、21%、22%、23%、24%、25%、26%、27%、28%、29%、30%、31%、32%、33%、34%、35%、36%、37%、38%、39%、40%、41%、42%、43%、44%、45%、46%、47%、48%、49%、50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%或95%。在一个非限制性实施方案中,基于由il-2突变蛋白在免疫细胞中诱导的stat5磷酸化(pstat5)的最大水平相对于由野生型il-2产生的最大p-stat5信号的比率来计算e最大。
在又其他实施方案中,与由野生型il-2(诸如,由seqidno:1编码的多肽)诱导的ifnγ产生和/或分泌的量相比,本文公开的il-2突变蛋白可导致更少的来自潜在炎性免疫细胞的干扰素-γ(ifnγ)产生和/或分泌(参见例如,实施例2和图5a至图5d)。ifnγ产生和/或分泌的减少可以处于如下水平:是野生型il-2在可比浓度和类似条件下在潜在炎性免疫细胞中刺激的水平的约1%、5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%或100%(包含这些百分比之间的值)。在一些实施方案中,本文公开的il-2突变蛋白不诱导从潜在炎性免疫细胞分泌ifnγ。在非限制性实施方案中,潜在炎性免疫细胞是cd8+t细胞和/或其他炎性免疫细胞子集。在另一个非限制性实施方案中,潜在炎性免疫细胞是cd4+ifnγ+t细胞或cd8+ifnγ+t细胞。
在其他实施方案中,与由野生型il-2(例如,由seqidno:1编码的多肽)诱导的潜在炎性t细胞的增殖相比,本文公开的il-2突变蛋白可导致更少的潜在炎性t细胞增殖。潜在炎性t细胞的非限制性示例包括cd4+ifnγ+t细胞或cd8+ifnγ+t细胞。在一些实施方案中,潜在炎性t细胞增殖的减少可以处于如下水平:是野生型il-2在可比浓度和类似条件下刺激的水平的1%、5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%或100%(包含这些百分比之间的值)或更低。在一些实施方案中,本文公开的il-2突变蛋白不诱导潜在炎性免疫细胞的任何增殖。在非限制性实施方案中,潜在炎性免疫细胞是cd8+t细胞和/或其他炎性免疫细胞子集。在另一个非限制性实施方案中,潜在炎性免疫细胞是cd4+ifnγ+t细胞或cd8+ifnγ+t细胞。
在仍其他实施方案中,本文公开的il-2突变蛋白可具有增加的功能,诸如增加的treg细胞的增殖。在一些实施方案中,与由野生型il-2(例如,由seqidno:1编码的多肽)诱导的treg细胞增殖的量相比,本文公开的il-2突变蛋白可以使treg细胞的增殖增加1倍、2倍、3倍、4倍、5倍、6倍、7倍、8倍、9倍或10倍中的任一值。在一些实施方案中,与由野生型il-2(诸如,由seqidno:1编码的多肽)诱导的treg细胞的增殖量相比,本文公开的il-2突变蛋白可以使treg细胞的增殖增加以下中的任一值:约5%、6%、7%、8%、9%、10%、11%、12%、13%、14%、15%、16%、17%、18%、19%、20%、21%、22%、23%、24%、25%、26%、27%、28%、29%、30%、31%、32%、33%、34%、35%、36%、37%、38%、39%、40%、41%、42%、43%、44%、45%、46%、47%、48%、49%、50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%或95%。在一些实施方案中,treg细胞是cd4+foxp3+细胞。
如下文更详细地描述,在本公开文本的一些实施方案中,可以将本文公开的il-2突变蛋白和编码此类il-2突变蛋白的核酸掺入组合物,包括药物组合物中。此类组合物通常包含所述多肽或核酸分子和药学上可接受的载体。
b.核酸
在一个方面,本文公开的一些实施方案涉及包括表达盒的编码本公开文本的il-2突变蛋白的核酸分子,以及含有与异源核酸序列可操作连接的这些核酸分子的表达载体,所述异源核酸序列为诸如调节序列,所述调节序列允许il-2突变蛋白在宿主细胞中的体内表达或在离体无细胞表达系统中的表达。
在各种实施方案中,在本公开文本的实践中使用的多肽是合成的,或是通过表达重组核酸分子产生的。在多肽是嵌合体(例如,至少包含突变型il-2多肽和异源多肽的融合蛋白)的情况下,所述多肽可以由杂合核酸分子编码,所述杂合核酸分子含有编码突变型il-2的全部或部分的一个序列,以及编码异源多肽的全部或部分的第二序列。例如,本文所述的主题il-2突变蛋白可以与六组氨酸标签融合以促进细菌表达的蛋白质的纯化,或者与血凝素标签融合以促进在真核细胞中表达的蛋白质的纯化。
术语“核酸分子”和“多核苷酸”在本文中可互换使用,是指rna分子和dna分子两者,包括包含以下的核酸分子:cdna、基因组dna、合成dna,以及包含核酸类似物的dna或rna分子。核酸分子可以是双链或单链的(例如,有义链或反义链)。核酸分子可包含非常规或经修饰的核苷酸。如本文所用的术语“多核苷酸序列”和“核酸序列”可互换地指多核苷酸分子的序列。本文使用37cfr§1.822中所示的核苷酸碱基命名法。
本公开文本的核酸分子可以是任何长度的核酸分子,包含优选在约0.5kb与约50kb之间,或例如在约0.5kb与约10kb之间、在约1kb与约8kb之间、在约2kb与约7kb之间,或在约2kb与约20kb之间,例如在约2kb与10kb之间、约3kb与约15kb之间、约4kb与约10kb之间、约5kb与约15kb之间,或约3kb与约9kb之间的核酸分子。在一些实施方案中,本公开文本的核酸分子可以在5kb至50kb之间,例如在约5kb与约40kb之间、在约5kb与约30kb之间、在约5kb与约20kb之间,或在约10kb与约50kb之间,例如在约15kb与30kb之间、约20kb与约50kb之间、约20kb与约40kb之间、约5kb与约25kb之间,或约30kb与约50kb之间。
用于构建编码il-2突变蛋白的dna序列并在适当转化的宿主中表达那些序列的方法包括但不限于使用pcr辅助的诱变技术。由il-2多肽中氨基酸残基的缺失或添加组成的突变也可以用标准重组技术进行。在缺失或添加的情况下,任选地用适当的限制性核酸内切酶消化编码il-2的核酸分子。所得片段可以直接表达,或通过例如将所得片段连接至第二片段而进一步操纵。如果核酸分子的两个末端含有彼此重叠的互补核苷酸,则可以促进连接,但是也可以连接平末端片段。pcr产生的核酸也可以用于产生各种突变型序列。
完整的氨基酸序列可用于构建反向翻译的基因。可以合成含有编码il-2突变蛋白的核苷酸序列的dna低聚物。例如,可以合成编码所需多肽的各部分的几个小寡核苷酸,然后进行连接。单独寡核苷酸通常包含5'或3'突出端以进行互补装配。
除了通过表达已经通过重组分子生物学技术改变的核酸分子来产生突变型多肽之外,还可以化学合成主题il-2突变蛋白。化学合成的多肽由本领域技术人员常规地产生。
一旦(通过合成、定点诱变或另一种方法)装配后,将编码il-2突变蛋白的dna序列插入表达载体中,并与适合于在所需的转化宿主中表达il-2突变蛋白的表达控制序列可操作地连接。正确的装配可以通过核苷酸测序、限制性酶切作图和生物活性多肽在合适宿主中的表达来确认。如本领域中众所周知的,为了获得转染基因在宿主中的高表达水平,必须将所述基因可操作地连接至在所选表达宿主中起作用的转录和翻译表达控制序列。
不管是通过定点诱变、化学合成还是其他方法制备,编码il-2突变蛋白的dna序列都还可以包括编码信号序列的dna序列。这种信号序列如果存在,应该是由被选择用于表达il-2突变蛋白的细胞所识别的信号序列。它可以是原核的、真核的或这两者的组合。它也可以是天然il-2的信号序列。信号序列的包含取决于是否需要从制备il-2突变蛋白的重组细胞中分泌il-2突变蛋白。如果选择的细胞是原核细胞,则通常优选的是dna序列不编码信号序列。如果选择的细胞是真核细胞,则通常优选的是编码信号序列,并且最优选地使用野生型il-2信号序列。
在一些实施方案中,可以通过表达核酸分子获得单独或作为嵌合多肽(诸如上述那些)的一部分的主题il-2突变蛋白。正如il-2突变蛋白可以用它们与野生型il-2多肽的同一性来描述一样,编码它们的核酸分子也需要与编码野生型il-2的那些核酸分子具有一定的同一性。例如,编码主题il-2突变蛋白的核酸分子可以与编码野生型il-2的核酸至少50%、至少65%、优选至少75%、更优选至少85%、最优选至少95%(例如,99%)相同。因此,在一些实施方案中,编码本文公开的主题il-2突变蛋白的核酸分子与编码具有seqidno:1所示的氨基酸的野生型il-2的核酸至少50%、至少65%、优选至少75%、更优选至少85%、最优选至少95%(例如,96%、97%、98%或99%)相同。在一些实施方案中,编码本文公开的主题il-2突变蛋白的核酸分子与编码具有seqidno:2所示的氨基酸的野生型il-2的核酸至少50%、至少65%、优选至少75%、更优选至少85%、最优选至少95%(例如,96%、97%、98%或99%)相同。
所提供的核酸分子可以含有天然存在的序列,或与天然存在的那些序列不同,但是由于遗传密码的简并性而编码相同多肽的序列。这些核酸分子可以由rna或dna(例如,基因组dna、cdna,或合成dna(例如通过基于亚磷酰胺的合成产生的dna))或这些类型的核酸内的核苷酸的组合或修饰组成。此外,核酸分子可以是双链或单链的(即,有义或反义链)。
核酸分子不限于编码多肽的序列;也可以包括位于编码序列(例如,il-2的编码序列)上游或下游的一些或全部非编码序列。分子生物学领域的普通技术人员熟悉分离核酸分子的常规程序。它们可以例如通过用限制性核酸内切酶处理基因组dna或通过进行聚合酶链式反应(pcr)来产生。在核酸分子是核糖核酸(rna)的情况下,分子可以例如通过体外转录产生。
本公开文本的示例性分离的核酸分子可包括并非以天然状态下的原样被发现的片段。因此,本公开文本涵盖重组分子,例如其中将核酸序列(例如,编码突变型il-2的序列)掺入载体(例如,质粒或病毒载体)中或掺入异源细胞的基因组(或同源细胞的基因组,在天然染色体部位以外的位置掺入)中的重组分子。
如上所述,主题il-2突变蛋白可以作为嵌合多肽的一部分存在。除了上述异源多肽或代替上述异源多肽,主题核酸分子可含有编码“标记”或“报告物”的序列。标记或报告基因的例子包括β-内酰胺酶、氯霉素乙酰转移酶(cat)、腺苷脱氨酶(ada)、氨基糖苷磷酸转移酶(neor,g418)、二氢叶酸还原酶(dhfr)、潮霉素-b-磷酸转移酶(hph)、胸苷激酶(tk)、lacz(编码β-半乳糖苷酶)和黄嘌呤鸟嘌呤磷酸核糖基转移酶(xgprt)。本领域技术人员将知道另外的有用试剂,例如可以起到标记或报告物功能的另外的序列。
主题核酸分子可以通过将突变引入从任何生物细胞(诸如哺乳动物细胞)获得的编码il-2的dna中而获得。因此,主题核酸(以及它们编码的多肽)可以是小鼠、大鼠、豚鼠、牛、绵羊、马、猪、兔子、猴子、狒狒、狗或猫的那些核酸。在一些实施方案中,核酸分子将是人的那些核酸分子。
c.载体和宿主细胞
本文还提供了含有编码本文公开的任何il-2突变蛋白多肽的核酸分子中的一个或多个核酸分子的载体、质粒或病毒。上述核酸分子可以包含在载体中,所述载体能够引导所述核酸分子在例如已用所述载体转导的细胞中的表达。因此,除了主题il-2突变蛋白外,包含编码主题il-2突变蛋白的核酸分子的表达载体和用这些载体转染的细胞在优选的实施方案中。用于真核和原核细胞的合适载体是本领域中已知的并且可以商购获得或由技术人员容易地制备。还可以在例如以下文献中发现另外的载体:ausubel,f.m.等人,currentprotocolsinmolecularbiology,(currentprotocol,1994)和sambrook等人“molecularcloning:alaboratorymanual,”第二版(1989)。
当然应当理解的是,并非所有的载体和表达控制序列都将同样好地作用于表达本文所述的dna序列。也并非所有宿主都将同样好地以相同表达系统发挥作用。然而,本领域技术人员可以在没有进行过多实验的情况下在这些载体、表达控制序列和宿主之中进行选择。例如,在选择载体时,必须考虑宿主,因为载体必须在宿主中复制。还应考虑载体的拷贝数、控制该拷贝数的能力以及由所述载体编码的任何其他蛋白质诸如抗生素标记的表达。例如,可以使用的载体包括允许编码il-2突变蛋白的dna以拷贝数扩增的载体。此类可扩增载体是本领域众所周知的。它们包括例如能够通过dhfr扩增(参见例如,kaufman,美国专利号4,470,461,kaufman和sharp,“constructionofamodulardihydrafolatereductasecdnagene:analysisofsignalsutilizedforefficientexpression”,mol.cell.biol.,2,第1304-19页(1982))或谷氨酰胺合成酶(“gs”)扩增(参见例如,美国专利号5,122,464和欧洲公开申请338,841)而扩增的载体。
在一些实施方案中,本公开文本的人il-2突变蛋白将从载体,优选地表达载体表达。载体可用于在宿主细胞中自主复制,或可在引入宿主细胞后整合到宿主细胞的基因组中,从而与宿主基因组一起复制(例如,非附加型哺乳动物载体)。表达载体能够引导与其可操作地连接的编码序列的表达。通常,在重组dna技术中有用的表达载体往往是质粒(载体)的形式。然而,还包括其他形式的表达载体,诸如病毒载体(例如,复制缺陷型逆转录病毒、腺病毒和腺相关病毒)。
示例性重组表达载体可包含一个或多个调节序列,所述一个或多个调节序列是基于要用于表达的宿主细胞选择的,可操作地连接至待表达的核酸序列。
可以设计表达构建体或载体以在原核或真核宿主细胞中表达il-2突变蛋白或其变体。
可经由常规转化或转染技术将载体dna引入原核或真核细胞中。用于转化或转染宿主细胞的合适方法可以发现于以下文献中:sambrook等人(1989)molecularcloning:alaboratorymanual(第二版,纽约州普莱恩维尤的springharborlaboratorypress)和其他标准分子生物学实验室手册。
蛋白质在原核生物中的表达最常在大肠杆菌(escherichiacoli)中使用含有组成型或诱导型启动子的载体进行。最大化大肠杆菌中的重组蛋白表达的策略可以发现于例如以下文献中:gottesman(1990)的geneexpressiontechnology:methodsinenzymology,第185卷(加利福尼亚州圣迭戈的academicpress),第119-128页和wada等人(1992)nucleicacidsres.20:2111-2118。用于细胞生长、收获、破坏或从细胞提取il-2突变蛋白或其变体的方法基本上描述于例如以下文献中:美国专利号4,604,377;4,738,927;4,656,132;4,569,790;4,748,234;4,530,787;4,572,798;4,748,234;和4,931,543,这些专利通过引用一起整体并入本文。
在一些实施方案中,重组il-2突变蛋白或其生物学活性变体也可以在真核生物(诸如酵母或人细胞)中制备。合适的真核宿主细胞包括昆虫细胞(可用于在培养的昆虫细胞(例如,sf9细胞)中表达蛋白质的杆状病毒载体的例子包括pac系列(smith等人(1983)mol.cellbiol.3:2156-2165)和pvl系列(lucklow和summers(1989)virology170:31-39));酵母细胞(用于在酿酒酵母(s.cerevisiae)中表达的载体的例子包括pyepsec1(baldari等人(1987)emboj.6:229-234)、pmfa(kurjan和herskowitz(1982)cell30:933-943)、pjry88(schultz等人(1987)gene54:113-123)、pyes2(加利福尼亚州圣地亚哥的invitrogencorporation)和ppicz(加利福尼亚州圣地亚哥的invitrogencorporation));或哺乳动物细胞(哺乳动物表达载体包括pcdm8(seed(1987)nature329:840)和pmt2pc(kaufman等人(1987)emboj.6:187:195))。合适的哺乳动物细胞包括中国仓鼠卵巢细胞(cho)或cos细胞。在哺乳动物细胞中,表达载体的控制功能通常由病毒调节元件提供。例如,常用的启动子源自多瘤病毒、腺病毒2、巨细胞病毒和猿猴病毒40。用于原核和真核细胞两者的其他合适的表达系统参见sambrook等人(1989)molecularcloning:alaboratorymanual(第二版,纽约州普莱恩维尤的springharborlaboratorypress)的第16章和第17章。参见goeddel(1990,出处同上)。
可以优化编码本公开文本的人il-2突变蛋白的序列以在目的宿主细胞中表达。可以将序列的g-c含量调节至给定细胞宿主的平均水平,如参照宿主细胞中表达的已知基因所计算的。用于密码子优化的方法是本领域中众所周知的。可以优化il-2突变蛋白编码序列内的密码子以增强在宿主细胞中的表达,使得已针对在特定宿主细胞中的表达优化了编码序列内的密码子的约1%、约5%、约10%、约25%、约50%、约75%或至多100%。
适合使用的载体包括用于细菌中的基于t7的载体(参见例如,rosenberg等人,gene56:125,1987)、用于哺乳动物细胞中的pmsxnd表达载体(lee和nathans,j.biol.chem.263:3521,1988)和用于昆虫细胞中的源自杆状病毒的载体(例如,来自加利福尼亚州帕洛阿尔托市的clontech的表达载体pbacpak9)。
在一些实施方案中,可以将在此类载体中编码主题il-2突变蛋白的核酸插入物可操作地连接至启动子,所述启动子基于例如寻求在其中表达的细胞的类型进行选择。
在选择表达控制序列时,还应考虑多种因素。这些因素包括例如序列的相对强度、序列的可控制性,以及序列与编码主题il-2突变蛋白的实际dna序列的相容性,特别是关于潜在的二级结构。宿主的选择应考虑它们与所选载体的相容性、由本公开文本的dna序列编码的产物的毒性、所述宿主的分泌特征、所述宿主正确折叠多肽的能力、所述宿主的发酵或培养要求,以及由所述dna序列编码的产物的易纯化性。
在这些参数内,本领域技术人员可以选择各种载体/表达控制序列/宿主组合,所述各种载体/表达控制序列/宿主组合将在进行发酵时或大规模动物培养(例如使用cho细胞或cos7细胞)中表达期望的dna序列。
在一些实施方案中,表达控制序列和表达载体的选择将取决于对宿主的选择。可以使用多种表达宿主/载体组合。用于真核宿主的有用的表达载体包括例如具有来自sv40、牛乳头瘤病毒、腺病毒和巨细胞病毒的表达控制序列的载体。用于细菌宿主的有用表达载体包括已知的细菌质粒,例如来自大肠杆菌的质粒,包括colel、pcri、per32z、pmb9以及它们的衍生物;更广泛的宿主范围质粒,诸如rp4;噬菌体dna,例如噬菌体λ的众多衍生物,例如nm989;以及其他dna噬菌体,诸如m13和丝状单链dna噬菌体。用于酵母细胞的有用表达载体包含2μ质粒及其衍生物。用于昆虫细胞的有用载体包括pvl941和pfastbactm1(马里兰州盖瑟斯堡的gibcobrl)。cate等人,cell,45,第685-98页(1986)。
另外,在这些载体中可以使用多种表达控制序列中的任何一种。此类有用的表达控制序列包括与前述表达载体的结构基因相关的表达控制序列。有用的表达控制序列的例子包括例如sv40或腺病毒的早期和晚期启动子、lac系统、trp系统、tac或trc系统、噬菌体λ的主要操纵子和启动子区域(例如pl)、fd外壳蛋白的控制区域、3-磷酸甘油酸激酶或其他糖酵解酶的启动子、酸性磷酸酶的启动子(例如,phoa)、酵母α-交配系统的启动子、杆状病毒的多面体启动子,以及已知控制原核或真核细胞或其病毒的基因表达的其他序列,以及它们的各种组合。
t7启动子可用于细菌中,多角体蛋白启动子可用于昆虫细胞中,并且巨细胞病毒或金属硫蛋白启动子可用于哺乳动物细胞中。同样,在高级真核生物的情况下,组织特异性和细胞类型特异性启动子是广泛可用的。这些启动子由于其引导核酸分子在体内给定组织或细胞类型中表达的能力而得名。技术人员熟知许多可用于引导核酸表达的启动子和其他调节元件。
除了促进插入的核酸分子转录的序列外,载体还可包含复制起点和编码选择性标记的其他基因。例如,新霉素抗性(neor)基因赋予表达它的细胞g418抗性,并且因此允许对经转染的细胞进行表型选择。本领域技术人员可以容易地确定给定的调节元件或选择性标记是否适合用于特定的实验环境。
可以在本公开文本中使用的病毒载体包括例如逆转录病毒、腺病毒和腺相关载体、疱疹病毒、猿猴病毒40(sv40)和牛乳头瘤病毒载体(参见例如,gluzman(编辑),eukaryoticviralvectors,纽约州冷泉港的cshlaboratorypress)。
包含并表达编码本文公开的主题il-2突变蛋白的核酸分子的原核或真核细胞也是本公开文本的特征。本公开文本的细胞是经转染的细胞,即已经借助于重组dna技术将核酸分子(例如编码突变型il-2多肽的核酸分子)引入到其中的细胞。这种细胞的子代也被认为在本公开文本的范围内。
表达系统的精确组分并不重要。例如,il-2突变蛋白可以在原核宿主(例如细菌大肠杆菌)中或在真核宿主(例如昆虫细胞(例如,sf21细胞)或哺乳动物细胞(例如,cos细胞、nih3t3细胞或hela细胞))中产生。这些细胞可从许多来源获得,包括美国典型培养物保藏中心(弗吉尼亚州的马纳萨斯)。在选择表达系统时,重要的仅是组分彼此相容。技术人员或普通技术人员能够做出这样的决定。此外,如果在选择表达系统时需要指导,则技术人员可以咨询ausubel等人,(currentprotocolsinmolecularbiology,johnwileyandsons,纽约州纽约,1993)和pouwels等人(cloningvectors:alaboratorymanual,1985年增刊,1987)。
如本文所述,可以使用常规的生化程序从表达系统纯化表达的多肽,并且可以将其用作例如治疗剂。
在一些实施方案中,获得的il-2突变蛋白将是糖基化或非糖基化的,取决于用于产生突变蛋白的宿主生物。如果选择细菌作为宿主,则产生的il-2突变蛋白将是非糖基化的。另一方面,真核细胞将使il-2突变蛋白糖基化,但可能与天然il-2糖基化的方式不同。可以根据任何合适的方法纯化由转化的宿主产生的il-2突变蛋白。已知有多种纯化il-2的方法。参见例如currentprotocolsinproteinscience,第2卷.编辑:johne.coligan,benm.dunn,hiddel.ploehg,davidw.speicher,pault.wingfield,6.5单元(copyright1997,johnwileyandsons,inc.)。il-2突变蛋白可以使用阳离子交换、凝胶过滤和/或反相液相色谱从大肠杆菌中产生的包涵体,或者从产生给定突变蛋白的哺乳动物或酵母培养的条件培养基分离。
构建编码il-2突变蛋白的dna序列的另一种示例性方法是通过化学合成。这包括通过蛋白质序列的化学手段直接合成肽,所述蛋白质序列编码展现出所述特性的il-2突变蛋白。该方法可以在影响il-2与il-2rα、il-2rβ和/或il-2rγ的相互作用的位置处掺入天然和非天然氨基酸。可替代地,可以使用寡核苷酸合成仪通过化学手段合成编码所需il-2突变蛋白的基因。基于所需il-2突变蛋白的氨基酸序列设计此类寡核苷酸,并且优选地选择将在其中产生重组突变蛋白的宿主细胞中有利的那些密码子。在这方面,众所周知遗传密码是简并的,因此一个氨基酸可以被多于一种密码子编码。例如,phe(f)由两种密码子tic或ttt编码,tyr(y)由tac或tat编码,而his(h)由cac或cat编码。trp(w)由单一密码子tgg编码。因此,应认识到,对于编码特定il-2突变蛋白的给定dna序列,将存在许多将编码该il-2突变蛋白的dna简并序列。例如,应认识到,除了图2所示的突变蛋白5-2的优选dna序列外,还有许多简并dna序列编码所示的il-2突变蛋白。这些简并dna序列被认为在本公开文本的范围内。因此,在本公开文本的情况下,“其简并变体”是指编码特定突变蛋白并由此使得能表达所述特定突变蛋白的所有dna序列。
il-2突变蛋白的生物学活性可以通过本领域中已知的任何合适方法来测定。此类测定包括pha-原始细胞增殖和nk细胞增殖。
d.融合蛋白
本文公开的任何il-2突变蛋白可以作为融合或嵌合多肽来制备,所述融合或嵌合多肽包含主题il-2突变蛋白和异源多肽(即,不是il-2或其突变体的多肽)(参见例如,美国专利号6,451,308)。示例性异源多肽可以延长嵌合多肽在体内的循环半衰期,并且因此可以进一步增强突变型il-2多肽的特性。在各种实施方案中,延长循环半衰期的多肽可以是血清白蛋白,诸如人血清白蛋白,或者是缺少igg重链可变区的抗体的igg亚类的fc区。示例性fc区可包含抑制补体固定和fc受体结合的突变,或者其可以是溶解性的,即能够结合补体或通过另一种机制(例如抗体依赖性补体溶解)溶解细胞(adcc;1994年12月12日提交的美国专利号08/355,502)。
“fc区”可以是与通过用木瓜蛋白酶消化igg而产生的iggc末端结构域同源的天然存在的或合成的多肽。iggfc的分子量为约50kda。突变型il-2多肽可包含完整的fc区,或较小部分,所述较小部分保留了延长嵌合多肽的循环半衰期的能力,所述较小部分是所述嵌合多肽的一部分。另外,全长或片段化的fc区可以是野生型分子的变体。也就是说,所述fc区可以包含可能影响或可能不影响多肽功能的突变;如下文进一步描述的,并非在所有情况下都需要或期望天然活性。在一些实施方案中,il-2突变蛋白融合蛋白(例如,如本文所述的il-2部分激动剂或拮抗剂)包含igg1、igg2、igg3或igg4fc区。
fc区可以是“溶解性的”或“非溶解性的”,但是通常是非溶解性的。非溶解性的fc区通常缺乏高亲和力fc受体结合位点和c'1q结合位点。鼠iggfc的高亲和力fc受体结合位点包括在iggfc的位置235处的leu残基。因此,fc受体结合位点可通过使leu235突变或缺失而被破坏。例如,用glu取代leu235抑制fc区结合高亲和力fc受体的能力。可以通过使igg的glu318、lys320和lys322残基突变或缺失在功能上破坏鼠c'1q结合位点。例如,用ala残基取代glu318、lys320和lys322使igg1fc无法引导抗体依赖性补体溶解。相比之下,溶解性的iggfc区具有高亲和力fe受体结合位点和c'1q结合位点。高亲和力fc受体结合位点包含iggfc的位置235处的leu残基,并且c'1q结合位点包含igg1的glu318、lys320和lys322残基。溶解性iggfc在这些位点处具有野生型残基或保守氨基酸取代。溶解性iggfc可以针对抗体依赖性细胞毒性或补体定向的细胞溶解(cdc)靶向细胞。人igg的适当突变也是已知的(参见例如,morrison等人,theimmunologist2:119-124,1994;和brekke等人,theimmunologist2:125,1994)。
在其他实施方案中,嵌合多肽可包含主题il-2突变蛋白和起抗原标签作用的多肽(诸如flag序列)。如本文所述,flag序列被生物素化的高度特异性抗flag抗体识别(也参见blanar等人,science256:1014,1992;leclair等人,proc.natl.acad.sci.usa89:8145,1992)。在一些实施方案中,嵌合多肽还包含c末端c-myc表位标签。
在其他实施方案中,嵌合多肽包含突变型il-2多肽和起增强突变型il-2多肽的表达或引导突变型il-2多肽的细胞定位的作用的异源多肽,例如aga2p凝集素亚基(参见例如,boder和wittrup,naturebiotechnol.15:553-7,1997)。
在其他实施方案中,可以产生包含突变型il-2和抗体或其抗原结合部分的嵌合多肽。嵌合蛋白的抗体或抗原结合组分可以用作靶向部分。例如,它可以用于将嵌合蛋白定位于特定的细胞子集或靶分子。产生细胞因子-抗体嵌合多肽的方法描述于例如美国专利号6,617,135中。
在一些实施方案中,突变体il-2可用一种或多种聚乙二醇(peg)分子修饰以延长其半衰期。如本文所用的术语“peg”是指聚乙二醇分子。peg在其典型形式下是具有末端羟基的线性聚合物,并且具有化学式ho-ch2ch2-(ch2ch2o)n-ch2ch2-oh,其中n是约8至约4000。
通常,n不是离散值,而是构成一个范围,所述范围在平均值附近具有近似高斯分布。末端氢可以被诸如烷基或链烷醇基的封端基团取代。peg可具有至少一个羟基,更优选地其是末端羟基。该羟基可附接至接头部分,所述接头部分可与肽反应形成共价键。本领域中存在peg的许多衍生物。(参见例如,美国专利号:5,445,090;5,900,461;5,932,462;6,436,386;6,448,369;6,437,025;6,448,369;6,495,659;6,515,100和6,514,491;以及zalipsky,s.bioconjugatechem.6:150-165,1995)。共价附接至本公开文本的il-2突变蛋白的peg分子的平均分子量可为约10,000、20,000、30,000或40,000道尔顿。peg化试剂可以是直链或支链的分子,并且可以单独或串联存在。本公开文本的peg化的il-2突变蛋白肽可具有附接至所述肽的c末端和/或n末端的串联peg分子。如本文所用的术语“peg化”是指如上所述的一个或多个peg分子与分子如本公开文本的il-2突变蛋白的共价附接。
e.药物组合物
适合于注射使用的药物组合物包括无菌水溶液(在水溶性的情况下)或分散液,以及用于临时制备无菌可注射溶液或分散液的无菌粉末。对于静脉内给予,合适的载体包括生理盐水、抑菌水、cremophoreltm.(新泽西州帕西帕尼的basf)或磷酸盐缓冲盐水(pbs)。在所有情况下,组合物应当是无菌的并且应当是易于注射的程度的流体。其在制造和储存条件下应当稳定并且必须抵抗微生物(如细菌和真菌)的污染作用而保存。载体可以是溶剂或分散介质,所述溶剂或分散介质含有例如水、乙醇、多元醇(例如,甘油、丙二醇和液体聚乙二醇等)及其合适的混合物。例如,可以通过使用包衣如卵磷脂、通过在分散液的情况下维持所需的粒度、以及通过使用表面活性剂(例如,十二烷基硫酸钠),维持适当的流动性。防止微生物的作用可以通过各种抗细菌以及抗真菌剂,例如对羟基苯甲酸酯、氯丁醇、苯酚、抗坏血酸、硫柳汞等来实现。在许多情况下,优选在组合物中包含等渗剂,例如糖、多元醇(如甘露糖醇、山梨糖醇)、氯化钠。可以通过在所述组合物中包含延迟吸收的药剂(例如单硬脂酸铝和明胶)来实现可注射组合物的延长吸收。
无菌可注射溶液可以通过以下方式制备:视需要将活性化合物以所需量掺入具有上文所列举成分中的一种或组合的适当溶剂中,之后过滤灭菌。通常,分散液是通过将活性化合物掺入无菌媒介物中来制备,所述无菌媒介物含有基础分散介质和来自以上列举的那些的所需其他成分。在用于制备无菌可注射溶液的无菌粉末的情况下,优选制备方法为真空干燥和冷冻干燥,这些方法由先前无菌过滤的溶液产生活性成分加上任何另外的所需成分的粉末。
口服组合物(如果使用的话)通常包含惰性稀释剂或可食用的载体。为了口服治疗给予的目的,可以将活性化合物与赋形剂一起掺入,并以片剂、锭剂或胶囊(例如,明胶胶囊)的形式使用。口服组合物也可以使用流体载体制备,用作漱口水。可以包括药学上相容的结合剂和/或辅助材料作为组合物的一部分。片剂、丸剂、胶囊剂、锭剂等可含有任何以下成分或具有类似性质的化合物:粘合剂,如微晶纤维素、黄芪胶或明胶;赋形剂,如淀粉或乳糖;崩解剂,如藻酸、primogeltm或玉米淀粉;润滑剂,如硬脂酸镁或sterotestm;助流剂,如二氧化硅胶体;甜味剂,如蔗糖或糖精;或调味剂,如薄荷、水杨酸甲酯或橙味剂。
在通过吸入给予的情况下,将主题il-2突变蛋白或编码它们的核酸从加压容器或分配器(其含有合适的推进剂,例如,气体(诸如二氧化碳))或喷雾器以气溶胶喷雾剂形式递送。此类方法包括在美国专利号6,468,798中描述的那些。
主题il-2突变蛋白或核酸的全身给予也可以通过经粘膜或透皮手段进行。对于经粘膜或透皮给予,在配制品中使用适合于待渗透屏障的渗透剂。此类渗透剂通常是本领域已知的,并且例如对于经粘膜给予包括洗涤剂、胆盐、以及梭链孢酸衍生物。经粘膜给予可以通过使用鼻喷雾剂或栓剂来完成。对于透皮给予,将活性化合物配制成如本领域通常已知的软膏、药膏、凝胶或乳膏。
在一些实施方案中,化合物(突变型il-2多肽或核酸)也可以以栓剂(例如,使用常规栓剂基质,如可可脂和其他甘油酯)或保留灌肠剂的形式来制备以供直肠递送。
在一些实施方案中,化合物(主题il-2突变蛋白或核酸)还可以通过转染或感染使用本领域已知的方法来给予,所述方法包括但不限于以下文献中描述的方法:mccaffrey等人(nature418:6893,2002);xia等人(naturebiotechnol.20:1006-1010,2002);或putnam(am.j.healthsyst.pharm.53:151-160,1996,勘误表,am.j.healthsyst.pharm.53:325,1996)。
在一些实施方案中,主题il-2突变蛋白或核酸是用将保护突变型il-2多肽免于从体内快速消除的载体来制备,此类载体如控释配制品,包括植入物和微囊化递送系统。可以使用可生物降解的生物相容性聚合物,如乙烯乙酸乙烯酯、聚酸酐、聚乙醇酸、胶原、聚原酸酯和聚乳酸。此类配制品可以使用标准技术来制备。所述材料也可以从alzacorporation和novapharmaceuticals,inc.商购获得。脂质体悬浮液(包括用针对病毒抗原的单克隆抗体靶向感染细胞的脂质体)也可用作药学上可接受的载体。这些可以根据本领域技术人员已知的方法来制备,例如如在美国专利号4,522,811中所述。
iv.本公开文本的方法
本公开文本的一些方面涉及可用于产生il-2突变蛋白(例如,本文所述的il-2部分激动剂)的方法和相关材料,以及用于治疗与由il-2信号传导途径介导的信号转导的扰动相关的健康病症和障碍的方法。更具体地,本公开文本的一些实施方案涉及在有需要的受试者中调节由白介素2(il-2)介导的信号转导途径。在本公开文本的一些实施方案中,il-2介导的信号传导通过选择性降低il-2与其受体中的一种、两种或三种(例如,白介素2α受体(il-2rα)、白介素2β受体(il-2rβ)、白介素2γ受体(il-2rγ))的结合来调节。
在一个方面,本公开文本的一些实施方案涉及用于产生本文公开的il-2突变蛋白的组合物和方法,所述方法包括在用于产生il-2突变蛋白的合适条件下培养如本文所述的宿主细胞。
在一些实施方案中,所述方法还包括分离产生的突变蛋白。在一些实施方案中,所述方法还包括纯化产生的突变蛋白。适用于分离和纯化在原核和真核宿主细胞中重组产生的蛋白质的技术、系统和相关材料是本领域中已知的。
在一些实施方案中,可以进一步修饰所产生的il-2突变蛋白以延长其在体内和/或离体的半衰期。适用于修饰本公开文本的il-2突变蛋白的已知策略和方法的非限制性例子包括(1)用高度可溶性的大分子如聚乙二醇(“peg”)对本文所述的il-2突变蛋白多肽进行化学修饰,从而防止所述多肽与蛋白酶接触;(2)将本文描述的il-2突变蛋白与抗体或抗体片段(例如人fc抗体片段)共价连接或缀合;以及(3)将本文所述的il-2突变蛋白与稳定蛋白质(诸如白蛋白)共价连接或缀合。因此,在一些实施方案中,本公开文本的il-2突变蛋白可以与稳定的蛋白质(诸如,白蛋白)融合。例如,已知人白蛋白是增强与之融合的多肽的稳定性的最有效蛋白质之一,并且已经报道了许多此类融合蛋白。
在一些实施方案中,将本公开文本产生的il-2突变蛋白用一个或多个聚乙二醇部分进行化学修饰,例如peg化;或进行类似的修饰,例如pas化。在一些实施方案中,peg分子或pas分子与il-2突变蛋白的一个或多个氨基酸缀合。在一些实施方案中,peg化或pas化的il-2突变蛋白仅在一个氨基酸上包含peg或pas部分。在其他实施方案中,peg化或pas化的il-2突变蛋白含有两个或更多个氨基酸上的peg或pas部分,例如附接至两个或更多个、五个或更多个、十个或更多个、十五个或更多个、或二十个或更多个不同的氨基酸残基的peg或pas部分。在一些实施方案中,peg或pas链为2000da、大于2000da、5000da、大于5,000da、10,000da、大于10,000da、大于10,000da、20,000da、大于20,000da和30,000da。产生的il-2突变蛋白可通过氨基、巯基、羟基或羧基直接偶联至peg或pas(例如,无连接基团)。
在一个方面,本公开文本的一些实施方案涉及可用于治疗与由il-2信号传导途径介导的信号转导的扰动相关的健康病症和障碍的组合物和方法。更具体地,本公开文本的一些实施方案涉及在有需要的受试者中调节由白介素2(il-2)介导的信号转导途径。
在一些实施方案中,可以将本公开文本的il-2突变蛋白和/或表达它们的核酸给予受试者以治疗与不希望的自身免疫或免疫抑制反应有关的障碍。在此类疾病的治疗中,本文公开的il-2突变蛋白可具有有利的特性,例如刺激调节性t细胞,同时最小程度地激活或不激活潜在炎性免疫细胞。
在一些实施方案中,本公开文本的il-2突变蛋白可用于治疗患有自身免疫性疾病、怀疑患有自身免疫性疾病或可能具有患上自身免疫性疾病的高风险的患者。所述方法包括向所述个体给予治疗有效量的(i)本文公开的任何一种il-2突变蛋白;(ii)本文公开的任何一种核酸或载体;和/或(iii)本文公开的任何一种药物组合物。在一些实施方案中,自身免疫性疾病选自类风湿性关节炎、胰岛素依赖性糖尿病、溶血性贫血、风湿热、甲状腺炎、克罗恩病、重症肌无力、肾小球肾炎、自身免疫性肝炎、多发性硬化、斑秃、牛皮癣、白癜风、营养不良性大疱性表皮松解症、系统性红斑狼疮和移植物抗宿主病。在一些实施方案中,自身免疫性疾病是移植物抗宿主病。
在一些实施方案中,将il-2突变蛋白、核酸或载体和/或药物组合物作为单一治疗剂或与一种或多种另外的治疗剂的组合给予受试者。在一些实施方案中,一种或多种另外的治疗剂包括将突变蛋白靶向特定细胞类型的抗体。在一些实施方案中,细胞类型是调节性t(treg)细胞。在一些实施方案中,抗体与il-2突变蛋白共价或非共价连接。
在一个方面,本文还提供了用于防止潜在炎性t细胞增殖和/或防止从cd8+t细胞或其他炎性免疫细胞子集分泌ifnγ的方法,所述方法包括使表达白介素2受体γ(il-2rγ)的细胞与本文公开的任何il-2突变蛋白接触。在其他实施方案中,潜在炎性t细胞是cd4+cd44+ifnγ+t细胞,或者是cd8+cd44+ifnγ+t细胞。在一些实施方案中,所述方法是体外、体内或离体进行的。
如上所述,在一些实施方案中,il-2突变蛋白在结构上经修饰以延长半衰期。在本文公开的任何实施方案中的一些实施方案中,所述修饰是选自以下的一种或多种修饰:与人fc抗体片段的融合、与白蛋白的融合和peg化。
在另一方面,本文提供了用于减少调节性t(treg)细胞增殖的方法,所述方法包括使treg细胞与白介素2(il-2)突变蛋白接触,所述il-2突变蛋白具有:(i)与由seqidno:2编码的多肽相比降低的对白介素2受体γ(il-2rγ)的结合亲和力;和(ii)与由seqidno:2编码的多肽相比0-50%e最大。在一些实施方案中,il-2突变蛋白包含:(i)与由seqidno:1编码的多肽相比增大il-2rβ结合亲和力的一种或多种氨基酸取代,所述一种或多种氨基酸取代选自根据seqidno:1的氨基酸序列编号的l80f、r81d、l85v、i86v和i92f;和(ii)与由seqidno:2编码的多肽相比降低il-2rγ受体结合亲和力并导致0-50%的e最大的一种或多种氨基酸取代,所述一种或多种氨基酸取代选自根据seqidno:2的氨基酸序列编号的(a)l18r和q22e;和(b)氨基酸位置126。
在另一方面,本文提供了用于减少调节性t(treg)细胞增殖的方法,所述方法包括使treg细胞与il-2突变蛋白接触,所述il-2突变蛋白具有:(i)与由seqidno:1编码的多肽相比降低的il-2rγ结合亲和力;和(ii)与由seqidno:1编码的多肽相比0-50%e最大。在一些实施方案中,il-2突变蛋白包括与由seqidno:1编码的多肽相比降低il-2rγ受体结合亲和力并导致0-50%e最大的一种或多种氨基酸取代,所述一种或多种氨基酸取代选自根据seqidno:1的氨基酸序列编号的(a)l18r和q22e;和(b)氨基酸位置126。在一些实施方案中,seqidno:1或seqidno:2的位置126处的氨基酸取代选自q126a、q126c、q126d、q126e、q126g、q126h、q126i、q126k、q126m、q126r、q126s、或q126t。在一些实施方案中,il-2突变蛋白包含氨基酸取代q126h、q126k或q126m。在一些实施方案中,il-2突变蛋白包含氨基酸取代q126h。在本文公开的任何实施方案中的一些实施方案中,所述方法还包括将il2突变蛋白与抗体一起给予,所述抗体将所述突变蛋白靶向treg细胞。在本文公开的任何实施方案中的一些实施方案中,所述抗体与所述突变蛋白共价或非共价连接。在本文公开的任何实施方案中的一些实施方案中,所述方法是体外、体内或离体进行的。
将药物组合物配制为与其预期的给予途径相容。如本文所用,术语“给予(administration和administering)”是指通过如下给予途径递送生物活性组合物或配制品:包括但不限于口服、静脉内、动脉内、肌内、腹膜内、皮下、肌内和局部给予或其组合。该术语包括但不限于由医学专业人员给予和自我给予。在一些实施方案中,本公开文本的突变型il-2多肽可以口服或通过吸入给予,但是它们更可能将通过肠胃外途径给予。肠胃外给予途径的例子包括例如静脉内、皮内、皮下、透皮(局部)、经粘膜和直肠给予。用于肠胃外施用的溶液或悬浮液可包括以下组分:无菌稀释剂,如注射用水、盐水溶液、不挥发油、聚乙二醇、甘油、丙二醇或其他合成溶剂;抗细菌剂,如苯甲醇或对羟基苯甲酸甲酯;抗氧化剂,如抗坏血酸或亚硫酸氢钠;螯合剂,如乙二胺四乙酸;缓冲液,如醋酸盐、柠檬酸盐或磷酸盐;以及用于调节张力的试剂,如氯化钠或右旋糖。ph可用酸或碱来调节,如磷酸二氢钠和/或磷酸氢二钠、盐酸或氢氧化钠(例如,调节至约7.2-7.8,例如7.5的ph)。可以将肠胃外制剂封装在由玻璃或塑料制成的安瓿、一次性注射器或多剂量小瓶中。
此类主题il-2突变蛋白或核酸化合物的剂量、毒性和治疗功效可通过标准药学程序在细胞培养或实验动物中确定,例如用于确定ld50(对50%群体致死的剂量)和ed50(在50%的群体中治疗有效的剂量)。在毒性与治疗效果之间的剂量比是治疗指数,并且其可表示为比率ld50/ed50。展现出高治疗指数的化合物是优选的。尽管可以使用展现出毒副作用的化合物,但应谨慎设计将此类化合物靶向至患病组织部位的递送系统,以最小化对未感染细胞的潜在损伤,从而减少副作用。
从细胞培养测定和动物研究获得的数据可用于配制用于人的剂量范围。此类化合物的剂量优选在包括具有很小或没有毒性的ed50的循环浓度的范围内。所述剂量可在这个范围内变化,根据所采用的剂型和所利用的给予途径而定。对于在本公开文本的方法中使用的任何化合物,治疗有效剂量可以首先根据细胞培养测定估计。可以在动物模型中配制剂量,以达到包括如在细胞培养中确定的ic50(即,测试化合物达到症状的半最大抑制的浓度)在内的循环血浆浓度范围。这个信息可以用于更准确地确定人中的有用剂量。血浆中的水平可以例如通过高效液相色谱法测量。
如本文所定义,主题il-2突变蛋白的“治疗有效量”(即,有效剂量)取决于所选择的多肽。例如,可以给予在约0.001mg/kg患者体重至0.1mg/kg患者体重的范围内的单一剂量;在一些实施方案中,可以给予约0.005mg/kg、0.01mg/kg、0.05mg/kg。在一些实施方案中,给予600,000iu/kg(iu可以通过淋巴细胞增殖生物测定法确定,并以如世界卫生组织的第一白介素-2(人)国际标准(1stinternationalstandardforinterleukin-2(human))建立的国际单位(iu)表示)。所述剂量可以与针对
本文还提供了用于预防、减少、或不促进潜在炎性t细胞的增殖和/或防止从cd8+t细胞或其他炎性免疫细胞子集分泌ifnγ的方法。所述方法是通过使表达白介素2受体γ(il-2rγ)的细胞与本文公开的任何il-2突变蛋白接触来进行。与野生型il-2在可比浓度和相似条件下刺激的增殖和/或ifnγ分泌相比,可以使潜在炎性t细胞的增殖和/或防止从cd8+t细胞或其他炎性免疫细胞子集分泌ifnγ减少以下中的任一值:约1%、5%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、95%或100%(包含这些百分比之间的值)。在一些实施方案中,潜在炎性t细胞是cd4+cd44+ifnγ+t细胞,或者是cd8+cd44+ifnγ+t细胞。所述方法可以另外在体外、体内或离体地进行。
v.系统和试剂盒
本公开文本的系统和/或试剂盒包括本文公开的任何il-2突变蛋白、核酸、载体或药物组合物中的一种或多种;以及注射器(包括预填充的注射器)和/或导管(包括预填充的注射器),所述注射器和/或导管用于向个体给予任何il-2突变蛋白、核酸、载体或药物组合物。所述试剂盒还包括使用本文公开的任何il-2突变蛋白、核酸、载体或药物组合物以及用于它们的给予的注射器和/或导管的书面说明。
在整个本说明书给出的每个最大数值极限意图包括每个数值下限,如同此类数值下限在本文中明确写出一样。在整个本说明书给出的每个最小数值极限将包括每个数值上限,如同此类数值上限在本文中明确地写出一样。在整个本说明书给出的每个数值范围将包括落入这个较宽数值范围内的每个较窄数值范围,如同此类较窄数值范围均在本文中明确地写出一样。
本公开文本中所提到的所有出版物和专利申请都通过引用并入本文中,并入程度如同明确且单独地指示每个单独出版物或专利申请通过引用并入一般。
不承认本文引用的任何参考文献构成现有技术。参考文献的讨论陈述其著者的主张,并且发明人保留质疑所引用文献的准确性和相关性的权利。应当清楚地理解,尽管本文中提及许多信息源,包括科学期刊文章、专利文件和教科书;但是该提及并不意味着承认这些文件中的任何文件构成了本领域中公知常识的一部分。
本文给出的一般方法的讨论意图仅用于说明目的。其他替代方法和替代方案对于本领域技术人员而言在阅读了本公开文本之后将是显而易见的,并且意图包括在本申请的精神和范围内。
实施例
实施例1
该实施例显示了各种人il-2变体刺激stat5磷酸化的能力。用如图3所描绘的图上所示的不同浓度(从1μm至0μm,10倍稀释)的与小鼠血清白蛋白(msa)融合的各种人il-2变体将人nk样yt细胞刺激15min。除了从昆虫细胞中纯化的il-2rek(seqidno:10)以外,所有il-2变体均从转导的hek293细胞中纯化。图3中描绘的图上的y轴示出了在每种浓度下归一化的来自每种il-2变体的p-stat5信号与来自wtil-2的p-stat5信号的比率。如图所示,用il-2h9(seqidno:2)背景中的特定氨基酸取代残基q126导致宽范围的il-2功效。
接下来,通过用图4a所描绘的图上所示浓度的il-2部分激动剂将yt细胞刺激15min来进行时程磷酸-stat5信号传导测定。图4a中的图的y轴示出了在每种浓度下归一化的来自每种il-2变体的p-stat5信号与来自wtil-2的p-stat5信号的比率。如图4b所示,在如图所示的各个时间点用1μm的不同il-2变体刺激yt细胞。在该图中,y轴表示p-stat5信号的中位荧光强度(mfi)。
在图4b中的该数据证明,wt(seqidno:1)、h9(seqidno:2)和新的部分激动剂ree(seqidno:6)、reh(seqidno:8)和rek(seqidno:10)(均在h9背景上)的p-stat5信号的量值是随时间恒定的,其中ree、rek和reh信号传导分别为25%、50%和75%。
实施例2
该实施例显示了il-2变体在小鼠中的体内给予。在第0天,雌性wtc57bl/6c小鼠通过腹膜内(ip)注射接受30μg的每种msa融合的il-2变体。在第6天,收获脾脏并通过流式细胞术分析细胞表面标记表达和细胞内细胞因子(n=3只/组)(参见图5a)。在图5b、图5c和图5d中描绘的图表示每种条件下所示细胞子集的频率,所述频率被归一化为经pbs处理的小鼠中的相应频率。b细胞由在cd3-cd19+上门控的细胞定义。nk细胞在cd3-nk1.1+上门控。在ly6g(gr1)+cd3+cd11b+上门控的细胞被定义为粒细胞。
如图5b至图5d中所示,il-2变体的给予导致其他免疫细胞类型中的轻度改变,il-2h9_reh(reh;seqidno:8)引发了cd4效应记忆t(tem)细胞的优先扩增。值得注意的是,虽然wt(seqidno:1)和h9il-2(seqidno:2)处理导致分泌ifnγ的cd8t细胞频率显著增加,但是在用ree(seqidno:6)、reh(seqidno:8)和rek(seqidno:10)中的任一者处理小鼠时并未观察到这种增加。
实施例3
该实施例公开了在野生型背景上il-2突变蛋白的产生和表征。使用磷酸-stat5信号传导测定法测量这些变体刺激stat5磷酸化的能力。将人nk样yt细胞(图6a)或小鼠饥饿t原始细胞(图6b)用如图所示不同浓度(从5μm至0μm)的与小鼠血清白蛋白(msa)融合的各种人il-2变体刺激15min。从昆虫细胞中纯化出所有il-2变体。图6a和图6b中描绘的图上的y轴示出了在每种浓度下归一化的来自每种il-2变体的p-stat5信号与来自wtil-2的p-stat5信号的比率。如图所示,与h9_reh(seqidno:8)相比,新的部分激动剂wt_reh(seqidno:15)似乎展现出较低的ec50和较高的e最大。两种新的部分激动剂wt_reh(seqidno:1)和h9_reh(seqidno:8)显示出比wt或h9il-2更低的功效(e最大)。
然后将野生型背景il-2突变蛋白给予b16黑色素瘤小鼠模型。在d0,在c57bl/6雌性小鼠中皮下注射106个b16f10细胞。在d5、d9和d14,通过ip注射用30μg的每种msa-il2变体处理小鼠。从d5开始每两天用卡尺测量肿瘤大小。以mm3计的肿瘤体积计算如下:(长度*(宽度)2)/2。在d19,收获脾脏并通过流式细胞术分析细胞表面和细胞内标记的表达。(n=5只/组)。
如图7a所描绘,与用pbs处理的小鼠相比,用wt(seqidno:1)或h9(seqidno:2)处理的小鼠的肿瘤大小明显较小。h9_reh(seqidno:8)和h9_rem(rem)(seqidno:11)给予导致与用pbs处理的小鼠可比的肿瘤大小增加。与用wt(seqidno:1)或h9(seqidno:2)处理的小鼠相比,用wt_reh(seqidno:15)处理的小鼠展现出更大的肿瘤大小。在用rem(seqidno:11)处理的小鼠中观察到了foxp3+调节性t(treg)细胞的频率增加,并且与pbs小鼠相比,当用wt_reh(seqidno:15)处理小鼠时,观察到foxp3+treg细胞数的3倍增加(图7b)。用wt和h9处理的小鼠展现出分泌ifnγ的cd44+cd8记忆细胞的扩增,但用新的部分激动剂wt_reh(seqidno:15)、h9_reh(seqidno:8)或rem(seqidno:11)处理时没有这种扩增。图7c描绘了对nk细胞和粒细胞的作用。使用分泌ifnγ的cd4cd44+记忆t细胞时观察到了相同的趋势(图7d)。最后,虽然wtil-2(seqidno:1)触发了cd4+效应t细胞(teffki67+细胞)和treg细胞的增殖,但wt_reh(seqidno:1)诱导treg细胞而不是teff细胞的特异性扩增(图7e)。
实施例4
该实施例描述了为了证明野生型背景中的il-2r部分激动剂可以在体内引发细胞类型特异性应答的实验。
在这些实验中,向雌性c57/bl6小鼠给予3×30μg腹膜内剂量(第0天、第3天和第6天)的野生型(wt)il-2(seqidno:1)、部分激动剂wt_reh(seqidno:15)、部分激动剂wt_retr(seqidno:21)、部分激动剂wt_rek(seqidno:22)或pbs对照。将所有细胞因子格式化为与小鼠血清白蛋白(msa)的n末端融合物。在第7天,收获脾脏和外周淋巴结并通过流式细胞术进行分析,并表示为昆虫细胞中的分泌蛋白。
这些实验中使用的il-2突变蛋白wt_retr和wt_rek包含以下氨基酸序列:
aptssstkktqlqlehlrldlemilnginnyknpkltrmltfkfympkkatelkhlqcleeelkpleevlnlaqsknfhlrprdlisninvivlelkgsettfmceyadetativeflnrwitfctsiirtlt(wt_retr;seqidno:21)。
aptssstkktqlqlehlrldlemilnginnyknpkltrmltfkfympkkatelkhlqcleeelkpleevlnlaqsknfhlrprdlisninvivlelkgsettfmceyadetativeflnrwitfcksiistlt(wt_rek;seqidno:22)。
观察到il-2r的部分激动剂诱导淋巴器官的增大与它们的激动剂活性相符(拍摄了脾脏和腹股沟淋巴结的照片)。如图8a至图8b所示,发现部分激动剂的给予诱发淋巴器官的增大。il-2和部分激动剂显示对nk细胞(cd3-nk1.1+)的频率几乎没有作用,同时il-2而不是部分激动剂显示b细胞(cd3-cd19+)的频率降低。还观察到il-2和部分激动剂不会改变t细胞(cd3+)的频率,然而il-2导致cd4t细胞与cd8t细胞的比率降低,而一种部分激动剂reh引起cd4t细胞与cd8t细胞的比率增大。
实施例5
该实施例描述了为证明用示例性il-2r部分激动剂reh处理增加了foxp3+调节性t细胞的频率并且降低了常规cd4+t细胞的激活而进行的实验。
在这些实验中,向雌性c57/bl6小鼠给予3×30μg腹膜内剂量(第0天、第3天和第6天)的野生型wtil-2(seqidno:1)、部分激动剂wt_reh(seqidno:15)、部分激动剂wt_retr(seqidno:21)、部分激动剂wt_rek(seqidno:22)或pbs对照。将所有细胞因子格式化为与小鼠血清白蛋白(msa)的n末端融合物,并表达为昆虫细胞中的分泌蛋白。在第7天,收获脾脏和外周淋巴结并通过流式细胞术进行分析,以评估treg(cd25+foxp3+)的频率和常规cd4+t细胞(cd25+foxp3-)的激活。
观察到il-2r部分激动剂reh增加了cd25+foxp3+调节性t细胞的频率,而没有诱导cd25+常规细胞。
实施例6
该实施例描述了为了证明来自用il-2r部分激动剂reh处理过的小鼠的treg抑制cd4+常规t细胞的增殖而进行的实验。
在这些实验中,向雌性b6.foxp3gfp小鼠给予3×30μg腹膜内剂量(第0天、第3天和第6天)的野生型wt(seqidno:1)、部分激动剂wt_reh(seqidno:15)或pbs对照。在第7天,收获脾和淋巴结,并通过磁性分选(macs)分离cd4t细胞。通过荧光激活细胞分选术(facs)基于gfp表达对纯化的cd4+t细胞进行分选,以分离treg(gfp+)和tcon(gfp-)。将来自细胞因子处理的小鼠的foxp3gfp+细胞与来自pbs处理的小鼠的celltraceviolet标记的tcon和acd3acd28包被的珠(珠与tcon的比率为1:1)共培养。72小时后评估载有ctv的tcon的增殖。
观察到来自reh处理的小鼠的foxp3+细胞能够抑制foxp3-效应细胞的增殖。
实施例7
本实施例描述了为了说明il-2r部分激动剂reh支持cd8+t细胞增殖但不支持ifnγ产生而进行的实验。
在这些实验中,通过磁性分离(macs)从c57/bl6小鼠的脾脏和淋巴结分离cd8+t细胞,加载celltraceviolet,并在存在或不存在20nmwt(seqidno:1)或wt_reh(seqidno:15)的情况下与acd3acd28包被的珠共培养三天。在收获前四个小时,将golgistop加入细胞中以防止进一步的细胞因子分泌。在72小时时,将细胞固定,透化处理并用针对干扰素γ(ifnγ)的抗体染色。
观察到il-2r部分激动剂reh和il-2都可以支持增殖,但是只有il-2可以通过使细胞增殖来诱导稳健的ifnγ产生。
实施例8
该实施例描述了为了证明il-2r部分激动剂reh的处理和共处理在啮齿动物eae模型(脑部炎症的动物模型)中防止各种自身免疫性症状而进行的实验。
在这些实验中,将b6小鼠用10μgmsa融合的wt_reh(seqidno:15)进行预处理(第-7天、第-4天、第-1天)或共处理(第0天、第3天和第7天)。在第0天将小鼠用在完全弗氏佐剂(cfa)和百日咳毒素(ptx)中的髓磷脂少突胶质细胞糖蛋白(mog35-55)进行免疫,然后在第2天进行ptx加强。依据体重减轻和疾病得分来跟踪疾病得分的进展。疾病得分如下:0-健康;1-尾巴无力;2-部分后肢瘫痪;3-完全后肢瘫痪;4-全身瘫痪;5-死亡。
观察到il-2r部分激动剂reh预处理是保护性的,而reh共处理以与检查treg在reh处理的小鼠中的动力学的先前实验一致的方式延迟疾病发作。
尽管已经公开了本公开文本的特定替代方案,但是应当理解,各种修改和组合是可能的并且涵盖于所附权利要求的真实精神和范围内。因此,无意限制于本文呈现的确切摘要和公开文本。
序列表
<110>小利兰·斯坦福大学董事会
k·c·加西亚
s·s·玛吉
c·r·格拉斯曼
l·l-r·苏
<120>白介素-2的部分激动剂
<130>078430-503001wo
<160>22
<170>patentinversion3.5
<210>1
<211>133
<212>prt
<213>智人
<220>
<223>wtil-2
<400>1
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysglnserile
115120125
ileserthrleuthr
130
<210>2
<211>133
<212>prt
<213>智人
<220>
<223>h9il-2
<400>2
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuleuleuaspleuglnmetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysglnserile
115120125
ileserthrleuthr
130
<210>3
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白h9_rea
<400>3
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysalaserile
115120125
ileserthrleuthr
130
<210>4
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白h9_rec
<400>4
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecyscysserile
115120125
ileserthrleuthr
130
<210>5
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白h9_red
<400>5
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysaspserile
115120125
ileserthrleuthr
130
<210>6
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白h9_ree
<400>6
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysgluserile
115120125
ileserthrleuthr
130
<210>7
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白h9_reg
<400>7
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysglyserile
115120125
ileserthrleuthr
130
<210>8
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白h9_reh
<400>8
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecyshisserile
115120125
ileserthrleuthr
130
<210>9
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白h9_rei
<400>9
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysileserile
115120125
ileserthrleuthr
130
<210>10
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白h9_rek
<400>10
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecyslysserile
115120125
ileserthrleuthr
130
<210>11
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白h9_rem
<400>11
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysmetserile
115120125
ileserthrleuthr
130
<210>12
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白h9_rer
<400>12
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysmetserile
115120125
ileserthrleuthr
130
<210>13
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白h9_rer
<400>13
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysargserile
115120125
ileserthrleuthr
130
<210>14
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白h9_res
<400>14
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisphe
65707580
aspproargaspvalvalserasnileasnvalphevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysserserile
115120125
ileserthrleuthr
130
<210>15
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白wt_reh
<400>15
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecyshisserile
115120125
ileserthrleuthr
130
<210>16
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白wt_rem
<400>16
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysmetserile
115120125
ileserthrleuthr
130
<210>17
<211>149
<212>prt
<213>智人
<220>
<223>人il-2,genbanknp_000577.2
<400>17
mettyrargmetglnleuleusercysilealaleuserleualaleu
151015
valthrasnseralaprothrserserserthrlyslysthrglnleu
202530
glnleugluhisleuleuleuaspleuglnmetileleuasnglyile
354045
asnasntyrlysasnprolysleuthrargmetleuthrphephetyr
505560
metprolysalathrgluleuhisleuglncysleugluglugluleu
65707580
lysproleuglugluvalleuasnleualaglnserlysasnphehis
859095
leuargproargaspleuileserasnilevalilevalleugluleu
100105110
lysglysergluthrthrphemetcysglutyralaaspgluthrala
115120125
thrilevalglupheleuasnargtrpilethrphecysglnserile
130135140
ileserthrleuthr
145
<210>18
<211>167
<212>prt
<213>小家鼠(musmusculus)
<220>
<223>鼠il-2
<400>18
mettyrsermetglnleualasercysvalthrleuthrleuvalleu
151015
leuvalasnseralaprothrserserserthrserserserthrala
202530
glualaglnglnglnglnglnglnglnglnglnglnglnglnhisleu
354045
gluglnleuleumetaspleuglngluleuleuserargmetgluasn
505560
tyrargasnleulysleuproargmetleuthrphelysphetyrleu
65707580
proglnalathrgluleulysaspleuglncysleugluaspgluleu
859095
glyproleuarghisvalleuaspleuthrglnserlysserphegln
100105110
leugluaspalagluasnpheileserasnileargvalthrvalval
115120125
leulysglyseraspasnthrpheglucysglnpheaspaspgluser
130135140
alathrvalvalasppheleuargargtrpilealaphecysglnser
145150155160
ileileserthrserprogln
165
<210>19
<211>149
<212>prt
<213>小家鼠(musmusculus)
<220>
<223>成熟鼠il-2
<400>19
alaprothrserserserthrserserserthralaglualaglngln
151015
glnglnglnglnglnglnglnglnglnglnhisleugluglnleuleu
202530
metaspleuglngluleuleuserargmetgluasntyrargasnleu
354045
lysleuproargmetleuthrphelysphetyrleuprolysglnala
505560
thrgluleulysaspleuglncysleugluaspgluleuglyproleu
65707580
arghisvalleuaspleuthrglnserlysserpheglnleugluasp
859095
alagluasnpheileserasnileargvalthrvalvallysleulys
100105110
glyseraspasnthrpheglucysglnpheaspaspgluseralathr
115120125
valvalasppheleuargargtrpilealaphecysglnserileile
130135140
serthrserprogln
145
<210>20
<211>133
<212>prt
<213>人工序列
<220>
<223>突变蛋白il-2
<220>
<221>变体
<222>(18)..(18)
<223>xaa=leu或arg
<220>
<221>尚未归类的特征
<222>(18)..(18)
<223>残基18可存在或不存在
<220>
<221>变体
<222>(22)..(22)
<223>xaa=gln或glu
<220>
<221>尚未归类的特征
<222>(22)..(22)
<223>残基22可存在或不存在
<220>
<221>变体
<222>(80)..(80)
<223>xaa=leu或phe或val
<220>
<221>尚未归类的特征
<222>(80)..(80)
<223>残基80可存在或不存在
<220>
<221>变体
<222>(81)..(81)
<223>xaa=arg或ile或thr或asp
<220>
<221>尚未归类的特征
<222>(81)..(81)
<223>残基81可存在或不存在
<220>
<221>变体
<222>(85)..(85)
<223>xaa=leu或val
<220>
<221>尚未归类的特征
<222>(85)..(85)
<223>残基85可存在或不存在
<220>
<221>变体
<222>(86)..(86)
<223>xaa=ile或val
<220>
<221>尚未归类的特征
<222>(86)..(86)
<223>残基86可存在或不存在
<220>
<221>变体
<222>(92)..(92)
<223>xaa=ile或phe
<220>
<221>尚未归类的特征
<222>(92)..(92)
<223>残基92可存在或不存在
<220>
<221>变体
<222>(126)..(126)
<223>xaa=gln或his或met或lys或cys或asp或glu或
gly或ile或arg或ser或thr
<220>
<221>尚未归类的特征
<222>(126)..(126)
<223>残基126可存在或不存在
<400>20
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuxaaleuaspleuxaametileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisxaa
65707580
xaaproargaspxaaxaaserasnileasnvalxaavalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysxaaserile
115120125
ileserthrleuthr
130
<210>21
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白wt_retr
<400>21
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecysthrserile
115120125
ileargthrleuthr
130
<210>22
<211>133
<212>prt
<213>人工序列
<220>
<223>il-2突变蛋白wt_rek
<400>22
alaprothrserserserthrlyslysthrglnleuglnleugluhis
151015
leuargleuaspleuglumetileleuasnglyileasnasntyrlys
202530
asnprolysleuthrargmetleuthrphelysphetyrmetprolys
354045
lysalathrgluleulyshisleuglncysleugluglugluleulys
505560
proleuglugluvalleuasnleualaglnserlysasnphehisleu
65707580
argproargaspleuileserasnileasnvalilevalleugluleu
859095
lysglysergluthrthrphemetcysglutyralaaspgluthrala
100105110
thrilevalglupheleuasnargtrpilethrphecyslysserile
115120125
ileserthrleuthr
130
- 还没有人留言评论。精彩留言会获得点赞!