木质纤维素材料的降解的制作方法
本申请是申请号为200980131289.8的中国专利申请的分案申请,原申请是申请日为2009年8月4日的国际申请pct/ep2009/060098的中国国家阶段申请。本发明涉及用于降解木质纤维素材料的方法,和由此类材料生产一种或多种糖的方法。本发明还涉及用于生产发酵产物的方法。
背景技术:
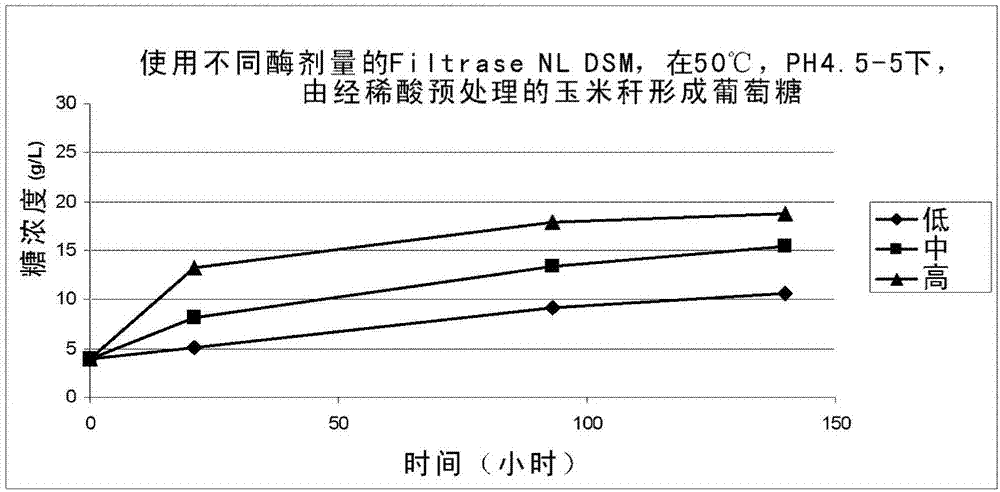
:碳水化合物组成地球上最丰量的有机化合物。然而,许多这类碳水化合物隐蔽在复杂聚合物中,包括淀粉(种子和谷物中的主要储存碳水化合物)和已知为木质纤维素的碳水化合物与木质素的集合。木质纤维素的主要碳水化合物组分是纤维素、半纤维素和果胶。这些复杂的聚合物通常被统称为木质纤维素。从由于opec减少石油输出而导致石油危机爆发的70年代开始,可再生的木质纤维素生物质成为可发酵的糖的生物转化吸引了研究人员强烈的注意力,所述可发酵的糖随后被发酵产生醇(例如乙醇),作为液体燃料的替代品。在过去二十年间,乙醇已被广泛使用,在美国作为与汽油的10%的掺合物,或在巴西作为车辆的纯燃料。更最近实现了e85(85%乙醇掺合物)的使用,特别是用于清洁城市应用。燃料生物乙醇的重要性将会随着油价的提高及其来源的逐渐耗尽而提高。另外,可发酵的糖被用于生产塑料、聚合物和其他基于生物的制品,并且预期这一工业将大幅增长,从而提高了对丰量的低成本可发酵糖的需求,所述可发酵糖可以被用作原料来代替基于石油的原料。这类大量碳水化合物在植物生物质中的隐蔽提供了糖(五碳糖以及六碳糖)形式的大量潜在的能量来源,所述糖能够被用于大量工业和农业过程。然而,这些碳水化合物的大量能源潜力目前利用不足,因为糖封闭在复杂聚合物中并因此不容易进行发酵。由植物生物质生产糖的方法会提供大量的、经济上有竞争性的原料,用于发酵成化学品、塑料和燃料。与纤维素原料的种类无关,酶的成本和水解效率是限制生物质生物转化方法商业化的主要因素。微生物生产的酶的生产成本与产酶菌株的生产力和发酵液中最终的活性产率密切相关。尽管过去几十年为了理解酶促木质纤维素生物质降解和纤维素酶生产而进行了持续的研究,但是仍然期望发现或者改造新的有高度活性的纤维素酶和半纤维素酶。还高度期望构建能够进行迅速和高效的木质纤维素材料生物降解的高效的酶组合物。另外,目前可获得的具有纤维素酶活性的酶(典型地源自trichoderma)在嗜温温度(mesophilictemperatures)如45℃到50℃下和ph5.0下发挥功能。然而,这可导致降低产率的细菌感染,因此期望在65℃或更高的温度下进行糖化。另外,嗜温温度的使用提高了使用的生物质的粘度,从而限制了使用的干物质含量。另外,使用经酸预处理的生物质时,必须提高ph,使得酶能够糖化生物质中的糖。在商业可行的燃料乙醇工业的情形中,这意味着需要例如氢氧化钠或硫酸钙和生产大量相应的盐,例如在氢氧化钠情况下生产石膏。因此,期望使用能够在ph4.0或更低的ph下工作的酶进行糖化。技术实现要素:我们已展示了源自talaromycesemersonii的酶制剂能够极度有效地将木质纤维素材料,例如玉米秆或小麦秸,水解成单体糖,所述单体糖随后可以被转化成有用的产物,如乙醇。酶制剂包含纤维素酶和半纤维素酶活性。令人惊讶的是,本发明现在表明所述酶制剂可以被用于进行高度有效的木质纤维素底物水解(实现超过90%的纤维素转化)。制剂比市面上能够获得的其他产品具有更高的比活性。这在从木质纤维素生物质生产商业可行的燃料乙醇的情况下意义高度重大,因为会需要(与目前可获得的产品相比)更少量的酶。另外,所述水解可以在高温下进行,这(i)降低了细菌感染的风险,并且(ii)导致更不粘稠的生物质浆体。后者的效果是显著的,因为其使得能够更好地掺合酶,导致植物中更高的可加工的干物质,并允许因此实现更高的乙醇浓度。因此,需要使用更少的能量来提高可持续性(sustainability),并且需要更少的发酵过程,从而需要更低的投资。另外,所述水解可以在低ph下进行。这是期望的,因为通常用酸预处理生物质。如果随后用于糖化的酶能够在低ph下作用,则用这种方式预处理的生物质不需要进行ph调节。这意味着对例如氢氧化钠或硫酸钙的更低需要,和其中没有废弃盐的方法。这在例如要生产乙醇的方法中是意义重大的,因为在此类方法中要消耗巨大量的材料。这允许进行其中不需要ph调节的方法,即不需要添加酸或碱。因此,所述方法可以作为零废弃物方法进行和/或作为其中不需要无机化学品输入的方法来进行。另外,还表明了使用高干物质含量时,酶组合物能够有效水解生物质。高度期望在例如燃料乙醇生产中使用的酶能够作用于具有高粘度的底物(即高干重组合物),因为这允许实现更高量的终产物,例如燃料乙醇。因此根据本发明提供了用于处理木质纤维素材料的方法,所述方法包括将所述木质纤维素材料与包含两种或更多酶活性的组合物接触,所述酶活性是纤维素酶和/或半纤维素酶活性,其中处理期间的ph约为4.5或更低,并且所述处理在15%或更高的干物质含量下进行。本发明还提供了:-由木质纤维素材料生产一种或多种糖的方法,所述方法包括使所述木质纤维素材料与上文定义的组合物接触;-用于生产发酵产物的方法,所述方法包括:使用上文公开的方法生产可发酵的糖;和发酵所得到的可发酵的糖,从而生产发酵产物;-上文定义的组合物在木质纤维素材料的处理中的用途;和-上文定义的组合物在由木质纤维素材料生产一种或多种糖中的用途。附图说明图1示出了在50℃,ph4.5-5下,使用不同酶剂量由经稀酸预处理的玉米秆形成葡萄糖:图1a=gc220;图nl;和fig.1c=laminexbg。图2和3示出了由经稀酸预处理的玉米秆形成糖的比活性数据:图2a和3a=21小时处的比活性;图2b和3b=93小时处的比活性;且图2c和3c=140小时处的比活性。图4示出了同时进行的糖化和发酵和蒸馏实验的流程图。图5示出了使用nl在同时进行的糖化和发酵和蒸馏实验中由经稀酸预处理的玉米秆生产乙醇。图6示出了使用nl在同时进行的糖化和发酵和蒸馏实验中由经稀酸预处理的玉米秆的水解产率。图7示出了在60℃,ph3.8下使用nl由用蒸汽预处理的小麦秸形成糖。图8示出了使用talaromyces纤维素酶生产葡萄糖。具体实施方式发明详述在本说明书和所附权利要求书中,词语“包含”和“包括”及其变体如“包含”(″comprises″,″comprising″)、“包括”(″includes″and″including″)应被解释为开放性的。也就是说,在上下文允许时,这些词语旨在表达可能包括未明确指出的其他要素或成分。冠词“一个”(“a”和“an”)在本文中被用于表示一个或多于一个(即一个或至少一个)所述冠词的语法客体。例如,“一个要素”可表示一个要素或多于一个要素。本发明涉及下述组合物,所述组合物包含纤维素水解和/或半纤维素水解酶活性并且具有改性(例如降解)非淀粉的碳水化合物材料的能力。非淀粉的碳水化合物材料是包含一种或多种非淀粉的碳水化合物的材料、由一种或多种非淀粉的碳水化合物组成的材料或基本由一种或多种非淀粉的碳水化合物组成的材料。碳水化合物在本文上下文中包括所有糖,例如多糖、寡糖、二糖或单糖。本文所述的组合物典型地通过此类材料的化学改性来改性非淀粉的碳水化合物材料。碳水化合物材料的化学改性可例如通过水解、氧化或其他化学改性(例如通过裂合酶的作用)导致此类材料的降解。适用于被本文所述的组合物改性的非淀粉的碳水化合物是木质纤维素。可以被认为是潜在的可再生原料的、包含不同木质纤维素残基的主要的多糖是纤维素(葡聚糖)、半纤维素(木聚糖、杂木聚糖(heteroxylans)和木葡聚糖(xyloglucans))。另外,一些半纤维素可以作为葡甘露聚糖(glucomannans)存在于例如木材衍生的原料中。这些多糖成为可溶糖(包括单体和多体,例如葡萄糖、纤维二糖、木糖、阿拉伯糖、半乳糖、果糖、甘露糖、鼠李糖、核糖、半乳糖醛酸、葡糖醛酸和其他己糖和戊糖)的酶促水解发生于共同作用(actinginconcert)的不同酶的作用下。另外,果胶和其他果胶物质如阿拉伯聚糖(arabinans)可占来自非木本植物组织典型的细胞壁干物质的可观的比例(约四分之一到一半的干物质可以是果胶)。纤维素是由通过β-1,4键连接的葡萄糖残基组成的线性多糖。纤维素纤维的线性本质以及化学计量的β-连接的葡萄糖(相对于α)产生比淀粉的高度枝化的α-连接的结构更倾向于链间(interstrand)氢键合的结构。因此,纤维素聚合物与淀粉中存在的纤维相比通常溶解度更小,并且形成更紧密结合的纤维。内切葡聚糖酶(eg)和外切纤维二糖水解酶(cbh)催化不溶性纤维素水解成为纤维寡糖(纤维二糖作为主要产物),而β-葡糖苷酶(bg)将寡糖(主要是纤维二糖和纤维三糖)转化成葡萄糖。半纤维素是复合聚合物,并且其组成常常在生物之间、组织类型之间大幅变化。通常,半纤维素的主要成分是β-1,4-连接的木糖(一种五碳糖)。然而,所述木糖通常在木糖的0-3和/或0-2原子处被枝化,并且可以被与阿拉伯糖、半乳糖、甘露糖、葡糖醛酸、半乳糖醛酸形成的键取代,或者被对乙酸的酯化(和阿魏酸对阿拉伯糖的酯化)取代。半纤维素也可含有葡聚糖,所述葡聚糖是β-连接的六碳糖(如先前提到的β-(1,3)(1,4)葡聚糖和杂葡聚糖)和额外的葡甘露聚糖(其中葡萄糖和甘露糖均存在于线性主链中,彼此通过β-键连接)的总称。木聚糖酶与其他附属酶例如α-l-阿拉伯呋喃糖酶、阿魏酸酯酶和乙酰木聚糖酯酶、葡糖醛酸糖苷酶和β-木糖苷酶一起,催化半纤维素的水解。果胶物质包括果胶、阿拉伯聚糖、半乳聚糖和阿拉伯半乳聚糖。果胶是植物细胞壁中最复杂的多糖。它们沿着一定程度上散布着l-鼠李糖的α(1,4)-连接的d-半乳糖醛酸单元核心链周围构建。在任何细胞壁中都存在符合所述描述的大量结构单元,并且通常认为在单个果胶分子中,不同结构单元的核心链彼此连续。结构单元的主要类型是:聚半乳糖醛酸(同聚半乳糖醛酸),其可被甲醇在羧基上取代,被乙酸酯在o-2和o-3上取代;鼠李半乳糖醛酸聚糖i(rgi),其中半乳糖醛酸单元与带有(1,4)-连接的半乳聚糖和(1,5)-连接的阿拉伯聚糖侧链的鼠李糖交替存在。阿拉伯聚糖侧链可直接与鼠李糖结合,或者通过半乳聚糖链间接结合;木半乳糖醛酸聚糖,在半乳糖醛酸的o-3上具有单个木糖基单元(与rgi紧密结合);和鼠李半乳糖醛酸聚糖ii(rgii),含有罕见糖例如芹糖的特别复杂的小型单元。rgii单元可含有两个芹菜糖残基,所述芹菜糖残基在合适的离子条件下能够与硼酸盐可逆地形成酯。在本发明方法中使用的组合物应包含下述酶活性,所述酶活性典型地源自penicillium纲和talaromyces属,例如talaromycesemersonii的腐生生物真菌微生物。talaromycesemersonii也可以被称作geosmithiaemersonii或penicilliumemersonii。talaromycesemersonii也曾被称作talaromycesduponti和penicilliumduponti。在本发明方法中使用的组合物包含至少两种活性,但是典型地组合物将包含多于两种活性,例如三种、四种、五种、六种、七种、八种、九种或更多。典型地,本发明的组合物可包含至少一种纤维素酶和至少一种半纤维素酶。然而,本发明的组合物可包含纤维素酶,但是不含木聚糖酶。另外,本发明的组合物可包含辅助酶活性,即直接或间接导致木质纤维素降解的额外的活性。此类辅助活性的例子在本文中有提到。因此,在本发明中使用的组合物可含有内切葡聚糖酶活性和/或纤维二糖水解酶活性和/或β-葡糖苷酶活性。在本发明中使用的组合物可包含一个或多个这些种类中的多于一种的酶活性。例如,在本发明中使用的组合物可包含两种内切葡聚糖酶活性,例如内切-1,3(1,4)-β葡聚糖酶活性和内切-β-1,4-葡聚糖酶活性。此类组合物也可包含一种或多种木聚糖酶活性。此类组合物可包含辅助酶活性。在本发明中使用的组合物可源自talaromycesemersonii。在本发明中,预期核心的一组(降解木质纤维素的)酶活性可源自talaromycesemersonii。talaromycesemersonii能够提供本文中证明对木质纤维素生物质的水解而言高度有效的活性组。随后可对所述活性补充来自其他来源的额外的酶活性。此类额外的活性可源自经典来源和/或由经遗传修饰的生物生产。本发明中使用的组合物中的活性可以是热稳定的。本文中这表示该活性具有40℃或更高,例如约50℃或更高,例如约60℃或更高,例如约70℃或更高,例如约75℃或更高,例如约80℃或更高,例如85℃或更高的温度最适度。本发明中使用的组合物中的活性一般不具有相同的温度最适度,但是优选地应是热稳定的。另外,本发明中使用的组合物中的酶活性可以能够在低ph下工作。就本发明的目的而言,低ph表示约5.5或更低,约5或更低,约4.9或更低,约4.8或更低,约4.7或更低,约4.6或更低,约4.5或更低,约4.4或更低,约4.3或更低,约4.2或更低,约4.1或更低,约4.0或更低,约3.9或更低,或约3.8或更低,约3.7或更低,约3.6或更低,或约3.5或更低的ph。本发明中使用的组合物中的活性可以通过任何上述温度最适度和ph值的组合来限定。除了源自talaromyces的活性以外,本发明方法中使用的组合物还可包含纤维素酶(例如源自除talaromyces之外其他来源的纤维素酶)和/或半纤维素酶(例如源自除talaromyces之外其他来源的半纤维素酶)和/或果胶酶。本发明中使用的组合物可包含一类、两类或三类纤维素酶,例如内切葡聚糖酶(eg)、外切纤维二糖水解酶(cbh)和β-葡糖苷酶(bg)中的一种、两种或所有。本发明中使用的组合物可包含两种或更多种任何这些种类的纤维素酶。已知talaromyces原产的β-葡糖苷酶非常有活性,talaromycesβ-葡糖苷酶cel3a的vmax值是512iu/mg,可观地高于针对来自其他真菌来源的β-葡糖苷酶所报道的数值(p.murrayetal./proteinexpressionandpuriwcation38(2004)248-257)。尽管根据本发明的组合物中β-葡糖苷酶有高活性并且达到了高葡萄糖水平,但是不发生葡萄糖抑制。这是有利的,因为使用根据本发明的组合物可以将高活性和高葡萄糖水平组合在一起。本发明的组合物可包含下述活性,所述活性与本发明方法中使用的组合物所提供的活性具有不同种类的纤维素酶活性和/或半纤维素酶活性和/或果胶酶活性。例如,本发明的组合物可包含一类由本文所述的组合物提供的纤维素酶和/或半纤维素酶活性和/或果胶酶活性,和由额外的纤维素酶/半纤维素酶/果胶酶提供的第二类纤维素酶和/或半纤维素酶活性和/或果胶酶活性。在本文中,纤维素酶是能够降解或改性纤维素的任何多肽。能够降解纤维素的多肽是能够催化下述过程的多肽:将纤维素降解成更小的单元,例如部分地降解成纤维素糊精,或者完全降解成葡萄糖单体。与纤维素酶接触时,根据本发明的纤维素酶可得到纤维素糊精与葡萄糖单体的混合种群。此类降解将典型地通过水解反应而发生。本文中,半纤维素是能够降解或改性半纤维素的任何多肽。也就是说,半纤维素酶可以能够降解或改性木聚糖、葡糖醛酸木聚糖、阿拉伯木聚糖、葡甘露聚糖和木葡聚糖之一种或多种。能够降解半纤维素的多肽是能够催化下述过程的多肽:将半纤维素降解成更小的多糖,例如部分地降解成寡糖,或者完全降解成糖单体,例如己糖或戊糖单体。与半纤维素酶接触时,根据本发明的半纤维素酶可得到寡糖与糖单体的混合种群。此类降解将典型地通过水解反应而发生。本文中,果胶酶是能够降解或改性果胶的任何多肽。能够降解果胶的多肽是能够催化下述过程的多肽:将果胶降解成更小的单元,例如部分地降解成寡糖,或者完全降解成糖单体。与果胶酶接触时,根据本发明的果胶酶可得到寡糖与糖单体的混合种群。此类降解将典型地通过水解反应而发生。因此,本发明的组合物可包含任何纤维素酶,例如纤维二糖水解酶,内切-β-1,4-葡聚糖酶,β-葡糖苷酶或β-(1,3)(1,4)-葡聚糖酶。本文中,纤维二糖水解酶(ec3.2.1.91)是能够催化纤维素或纤维四糖中1,4-β-d-葡糖苷键水解、从链末端释放纤维二糖的任何多肽。所述酶也可以被称作纤维素酶1,4-β-纤维二糖苷酶(1,4-β-cellobiosidase),1,4-β-纤维二糖水解酶,1,4-β-d-葡聚糖纤维二糖水解酶,微晶纤维素酶(avicelase),外切-1,4-β-d-葡聚糖酶,外切纤维二糖水解酶或外切葡聚糖酶。本文中,内切-β-1,4-葡聚糖酶(ec3.2.1.4)是能够催化纤维素、地衣淀粉或谷类β-d-葡聚糖中11,4-β-d-葡糖苷键内切水解的任何多肽。此类多肽也可以能够水解还含有1,3-键的β-d-葡聚糖中的1,4-键。所述酶也可以被称作纤维素酶、微晶纤维素酶、β-1,4-内切葡聚糖水解酶、β-1,4-葡聚糖酶、羧甲基纤维素酶、纤维糊精酶、内切-1,4-β-d-葡聚糖酶、内切-1,4-β-d-葡聚糖水解酶、内切-1,4-β-葡聚糖酶或内切葡聚糖酶。本文中,β-葡糖苷酶(ec3.2.1.21)是能够催化末端、非还原性β-d-葡萄糖残基水解并释放β-d-葡萄糖的任何多肽。这样的多肽可具有针对β-d-葡糖苷的广泛特异性,并且也可以水解以下一种或多种:β-d-半乳糖苷、α-l-阿拉伯糖苷、β-d-木糖苷或β-d-岩藻糖苷。所述酶也可以被称作苦杏仁苷酶、β-d-葡糖苷葡糖水解酶、纤维二糖酶或龙胆二糖酶。本文中,β-(1,3)(1,4)-葡聚糖酶(ec3.2.1.73)是能够催化含有1,3-和1,4-键的β-d-葡聚糖中1,4-β-d-葡糖苷键水解的任何多肽。此类多肽可作用于地衣淀粉和谷类β-d-葡聚糖,但不作用于仅含1,3-或1,4-键的β-d-葡聚糖。所述酶也可以被称作地衣淀粉酶(licheninase),1,3-1,4-β-d-葡聚糖4-葡聚糖水解酶,β-葡聚糖酶,内切-β-1,3-1,4葡聚糖酶,地衣多糖酶(lichenase)或混合键β-葡聚糖酶。这类酶的替代方案是被描述为内切-1,3(4)-β-葡聚糖酶的ec3.2.1.6。当下述葡萄糖残基在c-3处被自身取代时,这类酶水解β-d-葡聚糖酶中的1,3-键或1,4-键,所述葡萄糖残基的还原基团涉及要水解的键。替代性的名称包括内切-1,3-β-葡聚糖酶,昆布多糖酶,1,3-(1,3;1,4)-β-d-葡聚糖3(4)葡聚糖水解酶,底物包括昆布多糖,地衣淀粉和谷类β-d-葡聚糖。本发明的组合物可以包含任何半纤维素酶,例如内切木聚糖酶、β-木糖苷酶、α-l-阿拉伯呋喃糖苷酶、α-d-葡糖醛酸糖苷酶、乙酰木聚糖酯酶、阿魏酸酯酶、香豆酸酯酶、α-半乳糖苷酶、β-半乳糖苷酶、β-甘露聚糖酶或β-甘露糖苷酶。本文中,内切木聚糖酶(ec3.2.1.8)是能够催化木聚糖中1,4-β-d-木糖苷键内切水解的任何多肽。所述酶也可以被称作内切-1,4-β-木聚糖酶或1,4-β-d-木聚糖木聚糖水解酶。一种替代方案是ec3.2.1.136,葡糖醛酸阿拉伯木聚糖内切木聚糖酶,一种能够水解葡糖醛酸阿拉伯木聚糖中1,4木糖苷键的酶。本文中,β-木糖苷酶(ec3.2.1.37)是能够催化1,4-β-d-木聚糖的水解、从非还原性末端去除相继的d-木糖残基的任何多肽。这类酶也可以水解木二糖。所述酶也可以被称作木聚糖1,4-β-木糖苷酶、1,4-β-d-木聚糖木糖水解酶、外切-1,4-β-木糖苷酶或木二糖酶。本文中,α-l-阿拉伯呋喃糖苷酶(ec3.2.1.55)是能够作用于阿拉伯呋喃糖苷、含(1,2)和/或(1,3)-和/或(1,5)-键的α-l-阿拉伯聚糖、阿拉伯木聚糖和阿拉伯半乳聚糖的任何多肽。所述酶也可以被称作α-n-阿拉伯呋喃糖苷酶、阿拉伯呋喃糖苷酶或阿拉伯糖苷酶。本文中,α-d-葡糖醛酸糖苷酶(ec3.2.1.139)是能够催化以下形式反应的任何多肽:α-d-葡糖醛酸苷+h(2)o=醇+d-葡糖醛酸酯。所述酶也被称作α-葡糖醛酸糖苷酶或α-葡糖苷酸酶。这些酶也可水解可作为木聚糖中取代基存在的4-o-甲基化的葡糖醛酸。替代方式是ec3.2.1.131:木聚糖α-1,2-葡糖醛酸糖苷酶,其催化α-1,2-(4-o-甲基)葡糖醛酰基连接的水解。本文中,乙酰基木聚糖酯酶(ec3.1.1.72)是能够催化木聚糖和木寡糖脱乙酰化的任何多肽。此类多肽可催化来自多聚木聚糖、乙酰化木糖、乙酰化葡萄糖、α-萘基乙酸酯或对硝基苯基乙酸酯的乙酰基水解,但是一般不催化来自于三乙酰基丙三醇的乙酰基水解。此类多肽一般不作用于乙酰化的甘露聚糖或果胶。本文中,阿魏酸酯酶(ec3.1.1.73)是能够催化以下形式反应的任何多肽:阿魏酸-糖+h(2)o=阿魏酸酯+糖。糖可以是例如寡糖或多糖。其典型地可以催化来自酯化的糖的4-羟基-3-甲氧基肉桂酰(阿魏酰)基的水解,所述糖通常是“天然”底物中的阿拉伯糖。对硝基苯酚乙酸酯和阿魏酸甲基酯是典型地更弱的底物。所述酶也可以被称作肉桂酰基酯水解酶,阿魏酸酯酶或羟基肉桂酰基酯酶。也可以被称作半纤维素附属酶,因为其可帮助木聚糖酶和果胶酶分解植物细胞壁半纤维素和果胶。本文中,香豆酰基酯酶(ec3.1.1.73)是能够催化以下形式反应的任何多肽:香豆酰基-糖+h(2)o=香豆酸酯+糖。糖可以是例如寡糖或多糖。所述酶也可以被称作反式-4-香豆酰基酯酶、反式-对香豆酰基酯酶、对香豆酰基酯酶或对香豆酸酯酶。所述酶也落入ec3.1.1.73中,从而也可以被称作阿魏酸酯酶。本文中,α-半乳糖苷酶(ec3.2.1.22)是能够催化α-d-半乳糖苷(包括半乳糖寡糖、半乳甘露聚糖、半乳聚糖和阿拉伯半乳聚糖)中末端、非还原性α-d-半乳糖残基水解的任何多肽。此类多肽也可以能够水解α-d-岩藻糖苷。所述酶也可以被称作蜜二糖酶。本文中,β-半乳糖苷酶(ec3.2.1.23)是能够催化β-d-半乳糖苷中末端非还原性β-d-半乳糖残基水解的任何多肽。这样的多肽也可以能够水解α-l-阿拉伯糖苷。所述酶也可以被称作外切-(1->4)-β-d-半乳聚糖酶或乳糖酶。本文中,β-甘露聚糖酶(ec3.2.1.78)是能够催化甘露聚糖、半乳甘露聚糖和葡甘露聚糖中1,4-β-d-甘露糖苷键随机水解的任何多肽。所述酶也可以被称作甘露聚糖内切-1,4-β-甘露糖苷酶或内切-1,4-甘露聚糖酶。本文中,β-甘露糖苷酶(ec3.2.1.25)是能够催化β-d-甘露糖苷中末端非还原性β-d-甘露糖残基水解的任何多肽。所述酶也可以被称作甘露聚糖酶或甘露糖酶。本发明的组合物可包含任何果胶酶,例如内切多聚半乳糖醛酸酶,果胶甲基酯酶,内切-半乳聚糖酶,β-半乳糖苷酶,果胶乙酰基酯酶,内切-果胶裂合酶,果胶酸裂合酶,α-鼠李糖苷酶,外切-半乳糖醛酸酶,外切多聚半乳糖醛酸酯裂合酶,鼠李半乳糖醛酸聚糖水解酶,鼠李半乳糖醛酸聚糖裂合酶,鼠李半乳糖醛酸聚糖乙酰基酯酶,鼠李半乳糖醛酸聚糖半乳糖醛酸水解酶,木半乳糖醛酸酶。本文中,内切-多聚半乳糖醛酸酶(ec3.2.1.15)是能够催化果胶酸酯和其他半乳糖醛酸聚糖中1,4-α-d-半乳糖醛酸键的随机水解的任何多肽。所述酶也可以被称作多聚半乳糖醛酸酶果胶解聚酶,果胶酶(pectinase),内切多聚半乳糖醛酸酶,果胶酶(pectolase),果胶水解酶,果胶多聚半乳糖醛酸酶,多聚-α-1,4-半乳糖醛酸苷聚糖水解酶,内切半乳糖醛酸酶;内切-d-半乳糖醛酸酶或多聚(1,4-α-d-半乳糖醛酸苷)聚糖水解酶。在本文中,果胶甲基酯酶(ec3.1.1.11)是能够催化下述反应的任何酶:果胶+nh2o=n甲醇+果胶酸。所述酶也已知为果胶酯酶(pectinesterase),果胶脱甲氧基酶,果胶甲氧基酶,果胶甲基酯酶,果胶酶,果胶酯酶(pectinoesterase)或果胶果酰基水解酶(pectinpectylhydrolase)。在本文中,内切-半乳聚糖酶(ec3.2.1.89)是能够催化阿拉伯半乳聚糖中1,4-β-d-半乳糖苷键的内切水解的任何酶。所述酶也已知为阿拉伯半乳聚糖内切-1,4-β-半乳糖苷键,内切-1,4-β-半乳聚糖酶,半乳聚糖酶,阿拉伯半乳聚糖酶或阿拉伯半乳聚糖4-β-d-半乳聚糖水解酶。本文中,果胶乙酰基酯酶在本文中被定义为下述任何酶,所述酶具有催化果胶gaiua残基羟基处乙酰基的脱乙酰基作用的乙酰基酯酶活性。本文中,内切-果胶裂合酶(ec4.2.2.10)是能够催化(1→4、)-α-d-半乳糖醛酸聚糖甲基酯的清除性切割,得到在其非还原末端具有4-脱氧-6-o-甲基-α-d-半乳-4-糖醛酰基的寡糖。酶也可已知为果胶裂合酶,果胶反式消除酶(trans-eliminase),内切果胶裂合酶,多聚甲基半乳糖醛酸反式消除酶,果胶甲基反式消除酶,果胶酶(pectolyase),pl,pnl或pmgl或(1→4)-6-o-甲基-α-d-半乳糖醛酸聚糖裂合酶。本文中,果胶酸裂合酶(ec4.2.2.2)是能够催化(1→4)-α-d-半乳糖醛酸聚糖的消除性切割,得到在其非还原性末端具有4-脱氧-α-d-半乳-4-糖醛酰基的任何酶。酶也可以已知为多聚半乳糖醛酸反式消除酶,果胶酸反式消除酶,多聚半乳糖醛酸酯裂合酶,内切果胶甲基反式消除酶,果胶酸反式消除酶,内切半乳糖醛酸酯反式消除酶,果胶酸裂合酶,果胶裂合酶,α-1,4-d-内切多聚半乳糖醛酸裂合酶,pga裂合酶,ppase-n,内切-α-1,4-多聚半乳糖醛酸裂合酶,多聚半乳糖醛酸裂合酶,果胶反式消除酶,多聚半乳糖醛酸反式消除酶或(1→4)-α-d-半乳糖醛酸聚糖裂合酶。本文中,α-鼠李糖苷酶(ec3.2.1.40)是能够催化α-l-鼠李糖苷或鼠李半乳糖醛酸聚糖中末端非还原性α-l-鼠李糖残基水解的任何多肽。所述酶也可以已知为α-l-鼠李糖苷酶t,α-l-鼠李糖苷酶n或α-l-鼠李糖苷鼠李糖水解酶。本文中,外切-半乳糖醛酸酶(ec3.2.1.82)是能够从非还原性末端水解果胶酸、释放二半乳糖醛酸的任何多肽。酶也可以已知为外切-多聚-α-半乳糖醛酸苷酶(exo-poly-α-galacturonosidase)、外切多聚半乳糖醛酸苷酶(exopolygalacturonosidase)或外切多聚聚半乳糖醛酸苷酶(exopolygalacturanosidase)。本文中,外切-半乳糖醛酸酶(ec3.2.1.67)是能够催化以下的任何多肽:(1,4-α-d-半乳糖醛酸苷)n+h2o=(1,4-α-d-半乳糖醛酸苷)n-1+d-半乳糖醛酸酯。所述酶也可以已知为半乳糖醛1,4-α-半乳糖醛酸苷酶(galacturan1,4-α-galacturonidase),外切多聚半乳糖醛酸酶,多聚(半乳糖醛酸酯)水解酶,外切-d-半乳糖醛酸酶,外切-d-半乳糖醛酸聚糖酶,外切多聚-d-半乳糖醛酸酶或多聚(1,4-α-d-半乳糖醛酸苷)半乳糖醛酸水解酶。本文中,外切多聚半乳糖醛酸酯裂合酶(ec4.2.2.9)是能够催化从果胶酸(即脱脂化的果胶)的还原末端消除性切割4-(4-脱氧-α-d-半乳-4-糖醛酰基)-d-半乳糖醛酸酯的任何多肽。所述酶可已知为果胶酸二糖裂合酶,果胶酸外切裂合酶,外切果胶酸反式消除酶,外切果胶酸裂合酶,外切多聚半乳糖醛酸-反式-消除酶,pate,外切-pate,外切-pgl或(1→4)-a-d-半乳糖醛酸聚糖还原末端二糖裂合酶。本文中,鼠李半乳糖醛酸聚糖水解酶是能够在严格交替出现的鼠李半乳糖醛酸聚糖结构中以内切方式水解半乳基糖醛酸酸和吡喃鼠李糖基之间键的任何多肽,所述鼠李半乳糖醛酸聚糖结构由二糖[(1,2-α-l-鼠李糖基-(1,4)-α-半乳糖醛酸]组成。本文中,鼠李半乳糖醛酸聚糖裂合酶是能够以内切方式在鼠李半乳糖醛酸聚糖中通过β-消除切割α-l-rhap-(1→4)-α-d-galpa键的任何多肽。本文中,鼠李半乳糖醛酸聚糖乙酰基酯酶是能够催化鼠李半乳糖醛酸聚糖中交替出现的鼠李糖和半乳糖醛酸残基主链的脱乙酰化的任何多肽。本文中,鼠李半乳糖醛酸聚糖半乳糖醛酸水解酶是能够以外切方式、从严格交替出现的鼠李半乳糖醛酸聚糖结构的非还原性末端水解半乳糖醛酸的任何多肽。本文中,木半乳糖醛酸酶是通过以内切方式切割β-木糖取代的半乳糖醛酸主链而作用于木半乳糖醛酸聚糖的任何多肽。所述酶也可以已知为木半乳糖醛酸聚糖水解酶。本文中,α-l-阿拉伯糖呋喃糖苷酶(ec3.2.1.55)是能够作用于α-l-阿拉伯呋喃糖苷、含有(1,2)和/或(1,3)-和/或(1,5)-键的α-l-阿拉伯聚糖、阿拉伯木聚糖和阿拉伯半乳聚糖的任何多肽。所述酶也可以被称作α-n-阿拉伯糖呋喃糖苷酶,阿拉伯糖呋喃糖苷酶或阿拉伯糖苷酶。本文中,内切阿拉伯聚糖酶(ec3.2.1.99)是能够催化1,5-阿拉伯聚糖中1,5-α-阿拉伯呋喃糖苷键内切水解的任何多肽。所述酶也已知为内切阿拉伯糖酶,阿拉伯聚糖内切-1,5-α-l-阿拉伯糖苷酶,内切-1,5-α-l-阿拉伯聚糖酶,内切-α-1,5-阿拉伯聚糖酶;内切-阿拉伯聚糖酶或1,5-α-l-阿拉伯聚糖1,5-α-l-阿拉伯聚糖水解酶。本发明的组合物将一般包含至少一种纤维素酶和/或至少一种半纤维素酶和/或至少一种果胶酶(其中之一是根据本发明的多肽)。本发明的组合物可包含纤维二糖水解酶,内切葡聚糖酶和/或β-葡糖苷酶。此类组合物也可以包含一种或多种半纤维素酶和/或一种或多种果胶酶。另外,本发明的组合物中可以存在淀粉酶、蛋白酶、脂肪酶、木质酶、己糖基转移酶葡糖醛酸糖苷酶或扩展蛋白或纤维素诱导蛋白或纤维素整合蛋白或类似蛋白质中的一种或多种(例如两种、三种、四种或所有)(这些也被称作上文的辅助活性)。“蛋白酶”包括水解肽键的酶(肽酶),以及水解肽和其它部分如糖之间键的酶(糖肽酶)。许多肽酶根据ec3.4表征,适合在本发明中使用并且通过引用并入本文。一些特定类型的蛋白酶包括半胱氨酸蛋白酶(包括胃蛋白酶,木瓜蛋白酶)和丝氨酸蛋白酶(包括糜蛋白酶,羧肽酶和金属内切蛋白酶)。“脂肪酶”包括水解脂质、脂肪酸和酰基甘油酯(包括磷酸甘油酯)、脂蛋白、二酰基甘油等等的酶。在植物中,脂质被用作结构组分,来限制水损失和病原体感染。这些脂质包括来自脂肪酸的蜡质,以及角质和木栓素(suberin)。“木质酶”包括能够水解或分解木质素聚合物结构的酶。能够分解木质素的酶包括木质素过氧化物酶、锰过氧化物酶、漆酶和阿魏酸酯酶,和已知解聚或以其他方式分解木质素聚合物的其他酶。还包括在内的是能够水解在半纤维素糖(主要是阿拉伯糖)和木质素之间形成的键的酶。木质酶包括但不限于以下组的酶:木质素过氧化物酶(ec1.11.1.14)、锰过氧化物酶(ec1.11.1.13)、漆酶(ec1.10.3.2)和阿魏酸酯酶(ec3.1.1.73)。“己糖基转移酶”(2.4.1-)包括能够催化转移酶反应,但是也能够催化例如纤维素和/或纤维素降解产物的水解反应的酶。可以在本发明中使用的己糖基转移酶的例子是β-葡聚糖基转移酶。此类酶可以能够催化(1,3)(1,4)葡聚糖和/或纤维素和/或纤维素降解产物的降解。“葡糖醛酸糖苷酶”包括催化葡糖醛酸苷(例如β-葡糖醛酸苷)水解得到醇的酶。许多葡糖醛酸糖苷酶已被表征,并可适合在本发明中使用,例如β-葡糖醛酸糖苷酶(ec3.2.1.31),透明质酸酶葡糖醛酸糖苷酶(ec3.2.1.36),葡糖醛酰基-二硫葡糖胺葡糖醛酸糖苷酶(3.2.1.56),甘草酸β-葡糖醛酸糖苷酶(3.2.1.128)或α-d-葡糖醛酸糖苷酶(ec3.2.1.139)。本发明中使用的组合物可包含扩展蛋白或扩展蛋白样蛋白质,如膨胀因子(swollenin)(见salheimoetal.,eur.j.biohem.269,4202-4211,2002)或膨胀因子样蛋白质。扩展蛋白涉及植物细胞生长期间细胞壁的松弛。已经提出扩展蛋白破坏纤维素和其他细胞壁多糖之间的氢键,但不具有水解活性。认为它们通过这种方式允许纤维素纤维的滑动和细胞壁的扩大。一种扩展蛋白样蛋白质——膨胀因子,含有n端碳水化合物结合模块家族1结构域(cbd)和c端扩展蛋白样结构域。就本发明的目的而言,扩展蛋白样蛋白质或膨胀因子样蛋白质可包含此类结构域中的一种或两种和/或可破坏细胞壁的结构(如破坏纤维素结构),任选地不生产可检测量的还原糖。本发明中使用的组合物可以是纤维素诱导的蛋白质,例如cip1或cip2基因的多肽产物(见foremanetal.,j.biol.chem.278(34),31988-31997,2003),纤维素/纤维体(cellulosome)整合蛋白质,例如cipa或cipc基因的多肽产物,或支架蛋白或支架蛋白样蛋白质。支架蛋白和纤维素整合蛋白是多功能整合亚基,其可将分解纤维素的亚基组合进多酶复合物中。这通过两类互补的结构域(即支架蛋白上的内聚结构域(cohesiondomain)和每个酶单元上的锚定结构域(dockerindomain))的相互作用完成。支架蛋白亚基还带有介导纤维体与其底物结合的纤维素结合模块(cbm)。就本发明的目的而言,支架蛋白或纤维素整合蛋白可包含此类结构域中的一种或两种。本发明中使用的组合物可由大量上文提到的各种酶、一个酶种类的若干成员、或这些酶种类或辅助蛋白(即本文提到的本身不具有酶活性,但是帮助木质纤维素降解的这些蛋白质)的任何组合组成。在本发明的方法中使用的组合物可以由来自以下的酶组成:(1)供应商;(2)经克隆的表达酶的基因;(3)复合培养液(例如由培养基中微生物菌株的生长得到的,其中所述菌株向培养基中分泌蛋白质和酶);(4)如(3)中培养的菌株的细胞裂解物;和/或(5)表达酶的植物材料。本发明组合物中不同的酶可得自不同的来源。酶可以在微生物、酵母、真菌、细菌或植物中外源产生,然后分离并加入例如木质纤维素原料中。或者,酶可以被生产但不分离,并可将粗制细胞群发酵液或植物材料(如玉米秆或小麦秸)等等加入例如原料中。或者,可以处理粗制细胞群或酶生产培养基或植物材料,以预防进一步的微生物生长(例如通过加热或添加抗微生物剂),然后添加至原料。这些粗制酶混合物可包含生产酶的生物。或者,酶可以在下述发酵中生产,所述发酵施用原料(例如玉米秆或小麦秸)对生产酶的生物提供营养。通过这种方式,生产酶的植物自身可以发挥木质纤维素原料的作用,并且被添加进木质纤维素原料中。在本文所述的用途和方法中,上述组合物的组分可以共同(即本身作为单一组合物)或单独或先后提供。本发明因此涉及其中使用上述组合物的方法,和所述组合物在工业方法中的用途。原则上,本发明的组合物可以在任何下述方法中使用,所述方法需要处理包含非淀粉的多糖的材料。因此,本发明的多肽或组合物可以在非淀粉的多糖材料的处理中使用。本文中,非淀粉的多糖材料是下述材料,所述材料包含一种或更典型地包含多于一种非淀粉的多糖或者基本由所述非淀粉的多糖组成。典型地,植物和真菌和源自它们的材料包含大量非淀粉的多糖材料。因此,本发明的多肽可以用于植物或真菌材料或源自它们的材料的处理。植物非淀粉多糖材料的一个重要的组分是木质纤维素(在本文中也称作木质纤维素生物质)。木质纤维素是由纤维素和半纤维素和木质素组成的植物材料。碳水化合物聚合物(纤维素和半纤维素)与木质素通过氢键和共价键紧密结合。因此,本发明的多肽可以用于木质纤维素材料的处理。本文中,木质纤维素材料是包含木质纤维素或基本由木质纤维素组成的材料。因此,在用于处理非淀粉的多糖的本发明方法中,非淀粉的多糖可以是木质纤维素材料/生物质。因此,本发明提供了用于处理非淀粉的多糖的方法,其中所述处理包括纤维素和/或半纤维素的降解和/或改性。降解在本文上下文中表示导致产生纤维素和/或半纤维素和/或果胶物质的水解产物(即与类似的未经处理的非淀粉多糖中存在的相比,由于处理而存在更短链的糖)的处理。因此,降解在本文上下文中可以导致寡糖和/或糖单体的释放。所有植物和真菌都含有非淀粉的多糖,同样,基本上所有来自植物和真菌的多糖材料也含有非淀粉的多糖。因此,在本发明的用于处理非淀粉的多糖的方法中,所述非淀粉的多糖可以以植物或源自植物的材料或包含植物或源自植物的材料的形式提供,例如以植物浆、植物提取物、食品或其成分、织物、纺织品或衣物。本发明提供了由木质纤维素材料生产糖的方法,所述方法包括将本文所述的组合物与木质纤维素材料接触。此类方法允许由木质纤维素生物质生产游离的糖(单体)和/或寡糖。这些方法涉及用本发明的多肽或组合物将木质纤维素生物质转化成游离糖和小寡糖。将复合碳水化合物如木质纤维素转化为糖的方法优选地允许转化成可发酵的糖。此类方法可以被称作“糖化”。因此,本发明的方法可导致一种或多种己糖和/或戊糖(如葡萄糖、纤维二糖、木糖、阿拉伯糖、半乳糖、半乳糖醛酸、葡糖醛酸、甘露糖、鼠李糖、蔗糖和果糖中的一种或多种)的释放。适合在本发明的方法中使用的木质纤维素生物质包括下述生物质,所述生物质可包括原生生物质(virginbiomass)和/或非原生生物质如农业生物质,商业有机物,建筑和拆除碎片,市政固体废弃物,废纸和庭院废弃物。常见的生物质形式包括树木,灌木丛(shrubs)和草,小麦,小麦秸,甘蔗渣,柳枝稷,芒草(miscanthus),玉米,玉米秆(cornstover),玉米壳,玉米芯(corncobs),芸苔茎,大豆茎,高粱,玉米粒(包括来自玉米粒的纤维),来自谷物如玉米、小麦和大麦研磨(包括湿磨和干磨)的通常称作麸或纤维的产物和副产物,以及市政固体废弃物,废纸和庭院废弃物。生物质也可以是,但不限于草本材料,农业残余物,林业残余物,市政固体废弃物,废纸,和纸浆和造纸厂残余物。“农业生物质”包括树枝,灌木(bushes),藤蔓(canes),玉米和玉米壳,能量作物,森林,水果,鲜花,谷物,草,草本作物,树叶,树皮,针叶,原木,根,树苗,短期轮种木本作物(short-rotationwoodycrops),灌木丛,柳枝稷,树木,蔬菜,水果皮,蔓藤(vines),甜菜浆,小麦麸皮(wheatmidlings),燕麦壳,和硬木材或软木材(不包括带有有害材料的木材)。另外,农业生物质包括由农业加工产生的有机废弃物材料,所述农业加工包括农业和林业活动,特定地包括林业木材废弃物。农业生物质可以是前述任一或其任何组合或混合物。除了已经在食物和饲料或造纸和制浆工业中加工的原生生物质或原料以外,生物质/原料可额外地用热、机械和/或化学改性或此类方法的任何组合预处理,从而增强酶降解。可发酵的糖可以被转化成有用的增值的(value-added)发酵产物,其非限制性例子包括氨基酸、维生素、药物、动物饲料补充剂、特种化学品(specialtychemicals)、化学原料、塑料、溶剂、燃料或其他有机聚合物、乳酸和乙醇,包括燃料乙醇。可以通过本发明的方法生产的特定的增值产物包括但不限于生物燃料(包括乙醇和甲醇和沼气(biogas));乳酸;塑料;特种化学品;有机酸,包括柠檬酸、琥珀酸、反丁烯二酸、衣康酸和顺丁烯二酸;3-羟基-丙酸,丙烯酸;乙酸;1,3-丙烷-二醇;乙烯,丙三醇;溶剂;动物饲料补充剂;药物,如β-内酰胺抗生素或头孢菌素;维生素;氨基酸,如赖氨酸,甲硫氨酸,色氨酸,苏氨酸和天冬氨酸;工业酶,如蛋白酶,纤维素酶,淀粉酶,葡聚糖酶,乳糖酶,脂肪酶,裂合酶,氧化还原酶,转移酶或木聚糖酶;和化学原料。与单子叶植物(monocots,即具有单个子叶或种子叶的植物,如玉米、小麦、水稻、草、大麦)相比,双子叶植物(dicot,即其种子具有两片子叶或种子叶的植物,如利马豆(limabeans)、花生、杏仁、豌豆、四季豆)中半纤维素的组成、取代性质和枝化程度差异很大。在双子叶植物中,半纤维素主要由下述木葡聚糖组成,所述木葡聚糖是具有1,6-β-连接的木糖基侧链的1,4-β-连接的葡萄糖链。在单子叶植物(包括大部分谷类作物)中,半纤维素的主要组分是异木聚糖。这些主要由下述1,4-β-连接的木糖主链聚合物组成,所述木糖主链聚合物与阿拉伯糖、半乳糖、甘露糖和葡糖醛酸或4-o-甲基-葡糖醛酸以及由与酯连接的乙酸改性的木糖具有1,3-α连接。还存在由1,3-和1,4-β-连接的葡糖基链组成的β葡聚糖。在单子叶植物中,纤维素、杂木聚糖和β-葡聚糖可以大致等量地存在,各自包含约15-25%的细胞壁干物质。另外,不同的植物可包含不同量和不同组成的果胶物质。例如,甜菜含有以干重为基础约19%的果胶和约21%的阿拉伯聚糖。因此,本发明的组合物可根据要使用的具体原料来定制。也就是说,本发明组合物中的活性谱可根据考虑的原料来变化。酶组合或物理处理可以并发或先后地施用。酶可以在微生物、酵母、真菌、细菌或植物中外源产生,然后分离并加入木质纤维素原料中。或者,酶可以被生产但不分离,并且将粗制细胞群发酵液或植物材料(如玉米秆)等等加入原料中。或者,可以处理粗制细胞群或酶生产培养基或植物材料,以预防进一步的微生物生长(例如通过加热或添加抗微生物剂),然后添加至原料。这些粗制酶混合物可包含生产酶的生物。或者,酶可以在下述发酵中生产,所述发酵施用原料(例如玉米秆)对生产酶的生物提供营养。通过这种方式,生产酶的植物可以发挥木质纤维素原料的作用,并且被添加进木质纤维素原料中。在本发明的方法中,酶或酶的组合作用于发挥原料作用的木质纤维素底物或植物生物质上,从而将所述复合底物转化成简单的糖和寡糖,用于生产乙醇或其它有用的发酵产物。因此,本发明的另一方面包括下述方法,所述方法与其它酶或物理处理(例如温度和ph)一起利用上述组合物,将木质纤维素植物生物质转化成糖和寡糖。尽管组合物已作为单一混合物被讨论,但是认为酶可以先后添加,其中温度、ph和其他条件可以被改变,以提高每种个体酶的活性。或者,可以针对酶混合物测定最适ph和温度。组合物在任何适当的条件下与底物反应。例如,酶可以在约25℃,约30℃,约35℃,约37℃,约40℃,约45℃,约50℃,约55℃,约60℃,约65℃,约70℃,约75℃,约80℃,约85℃,约90℃或更高下孵育。也就是说,它们可以在从约20℃到约95℃下,例如在低到中等离子强度的缓冲液中和/或从低到中性ph下孵育。“中等离子强度”表示对任何单个离子组分而言,缓冲液具有约200毫摩尔(mm)或更少的离子浓度。ph可在从约ph2.5,约ph3.0,约ph3.5,约ph4.0,约ph4.5,约ph5,约ph5.5,约ph6,约ph6.5,约ph7,约ph7.5,约ph8.0,到约ph8.5的范围内。通常,ph范围会从约ph3.0到约ph9。典型地,反应可以在上文定义的低ph条件下进行。因此,可以进行本发明的方法从而不需要调节ph(即至更中性的ph)。也就是说,可以使用经酸预处理的原料,因为在添加本发明的组合物之前不需要添加例如过氧化钠。可以在液化/水解之前洗涤原料。所述洗涤可以例如用水进行。在这些条件下孵育组合物导致大量糖从木质纤维素材料释放或解离。大量表示至少约20%、至少约30%、至少约40%、至少约50%、至少约60%、至少约70%、至少约80%、至少约90%、至少约95%或更多的可利用的糖。可以使用涉及用酶或酶混合物孵育的液化/水解或预糖化步骤。所述步骤可以在许多不同的温度下进行,但是优选预处理在最适合要测试的酶混合物的温度下,或针对要测试的酶预测的酶最适度下进行。预处理温度可以在从约10℃到约80℃,约20℃到约80℃,约30℃到约70℃,约40℃到约60℃,约37℃到约50℃,优选地约37℃到约80℃,更优选地约50℃的范围内。缺失温度最适度数据时,优选首先在37℃下,然后在更高温度如50℃下进行预处理反应。预处理混合物的ph可以在从约2.0到约10.0,但是优选地约3.0到约5.0。另外,在糖化之前可能不必须调节ph,因为本发明中使用的组合物典型地适合在本文定义的低ph下使用。液化/水解或预糖化步骤反应可以进行数分钟到数小时,例如从约1小时到约120小时,优选地从约2小时到约48小时,更优选地从约2小时到约24小时,最优选地从约2小时到约6小时。纤维素酶处理可进行数分钟到数小时,例如从约6小时到约168小时,优选地约12小时到约96小时,更优选地约24小时到约72小时,进一步更优选地从约24小时到约48小时。在以单独水解和发酵(shf)模式进行液化/水解或预糖化步骤时,这些条件特别适合。ssf模式对同时糖化和发酵(ssf)的模式而言,液化/水解或预糖化步骤的反应时间取决于实现期望产率(即纤维素成为葡萄糖的转化产率)的时间。这类产率优选地尽可能高,优选地为60%或更多,65%或更多,70%或更多,75%或更多,80%或更多,85%或更多,90%或更多,95%或更多,96%或更多,97%或更多,98%或更多,99%或更多,甚至99.5%或更多或99.9%或更多。根据本发明,在shf模式下实现了非常高的糖浓度,在ssf模式下实现了非常高的产物浓度(例如乙醇)。在shf操作中,葡萄糖浓度是25g/l或更高,30g/l或更高,35g/l或更高,40g/l或更高,45g/l或更高,50g/l或更高,55g/l或更高,60g/l或更高,65g/l或更高,70g/l或更高,75g/l或更高,80g/l或更高,85g/l或更高,90g/l或更高,95g/l或更高,100g/l或更高,110g/l或更高,120g/l或更高,或者可以例如是25g/l-250g/l,30gl/l-200g/l,40g/l-200g/l,50g/l-200g/l,60g/l-200g/l,70g/l-200g/l,80g/l-200g/l,90g/l,80g/l-200g/l。ssf模式中的产物浓度在ssf操作中,产物浓度(g/l)取决于生产的葡萄糖量,但因为在ssf中糖被转化成产物,所以这是不可见的,并且糖浓度可以通过与理论最大产率(以gr产物/克葡萄糖计的ypsmax)相乘而与潜在的葡萄糖浓度相关。发酵产物的理论最大产率(以gr产物/克葡萄糖计的ypsmax)可源自生物化学教科书。对乙醇而言,1摩尔葡萄糖(180gr)根据酵母中正常的糖酵解发酵通路产生2摩尔乙醇(=2x46=92gr乙醇)。因此,基于葡萄糖的乙醇的理论最大产率为92/180=0.511gr乙醇/gr葡萄糖。对丁醇(mw74gr/摩尔)或异丁醇而言,理论最大产率是每摩尔葡萄糖1摩尔丁醇。因此,(异)丁醇的ypsmax=74/180=0.411gr(异)丁醇/gr葡萄糖。对乳酸而言,纯乳酸发酵(homolacticfermentatio)的发酵产率是每摩尔葡萄糖2摩尔乳酸(mw=90gr/摩尔)。根据所述化学计量法,ypsmax=1gr乳酸/gr葡萄糖。对其他发酵产物而言,可以进行类似的计算。ssf模式在ssf操作中,产物浓度为25g*ypsg/l/l或更高,30*ypsg/l或更高,35g*yps/l或更高,40*ypsg/l或更高,45*ypsg/l或更高,50*ypsg/l或更高,55*ypsg/l或更高,60*ypsg/l或更高,65*ypsg/l或更高,70*ypsg/l或更高,75*ypsg/l或更高,80*ypsg/l或更高,85*ypsg/l或更高,90*ypsg/l或更高,95*ypsg/l或更高,100*ypsg/l或更高,110*ypsg/l或更高,120g/l*yps或更高,或者可以例如是25*ypsg/l-250*ypsg/l,30*ypsgl/l-200*ypsg/l,40*ypsg/l-200*ypsg/l,50*ypsg/l-200*ypsg/l,60*ypsg/l-200*ypsg/l,70*ypsg/l-200*ypsg/l,80*ypsg/l-200*ypsg/l,90*ypsg/l,80*ypsg/l-200*ypsg/l。从生物质释放的糖可以被转化成有用的发酵产物,例如包括但不限于以下之一:氨基酸、维生素、药物、动物饲料补充剂、特殊化学品、化学原料、塑料和乙醇(包括燃料乙醇)。值得注意的是,本发明的方法可以在水解反应中使用高水平的(木质纤维素材料的)干物质。因此,本发明可以用约5%或更高,约8%或更高,约10%或更高,约11%或更高,约12%或更高,约13%或更高,约14%或更高,约15%或更高,约20%或更高,约25%或更高,约30%或更高,约35%或更高或约40%或更高的干物质含量进行。因此,本发明提供了制备发酵产物的方法,所述方法包括:a.使用本文所述的方法降解木质纤维素;和b.发酵所得到的材料,从而制备发酵产物。可以进行此类方法而不需要在所述方法期间调节ph。也就是说,所述方法是可以不添加任何酸或碱而进行的方法。然而,这排除了其中可以添加酸的预处理步骤。关键是本发明的组合物能够在低ph下发挥作用,因此不需要调节经酸预处理的原料的ph,从而可以发生糖化。因此,本发明的方法可以是仅使用有机产物而不需要输入无机化学品的零废弃物方法。可以根据本发明生产的发酵产物包括氨基酸、维生素、药物、动物饲料补充剂、特种化学品、化学原料、塑料、溶剂、燃料或其他有机聚合物、乳酸和乙醇,包括燃料乙醇(术语“乙醇”被理解为包括乙醇或乙醇和水的混合物)。可以通过本发明的方法生产的特定的增值产物包括但不限于生物燃料(包括乙醇和丁醇);乳酸;3-羟基-丙酸,丙烯酸;乙酸;1,3-丙烷-二醇;乙烯;丙三醇;塑料;特种化学品;有机酸,包括柠檬酸、琥珀酸和顺丁烯二酸;溶剂;动物饲料补充剂;药物,如β-内酰胺抗生素或头孢菌素;维生素;氨基酸,如赖氨酸,甲硫氨酸,色氨酸,苏氨酸和天冬氨酸;酶,如蛋白酶,纤维素酶,淀粉酶,葡聚糖酶,乳糖酶,脂肪酶,裂合酶,氧化还原酶,转移酶或木聚糖酶;化学原料;或动物饲料补充剂。用于制备发酵产物的方法可任选的包含发酵产物的回收。此类方法可在需氧或厌氧条件下进行。优选地,所述方法在微需氧或氧受限的条件下进行。厌氧发酵方法在本文中被定义为在不存在氧时运行的,或者其中基本不消耗氧,优选地消耗约5或更少、约2.5或更少或约1mmol/l/h或更少,并且其中有机分子既作为电子供体又作为电子受体的发酵方法。氧受限的发酵方法是其中氧消耗受到从气体到液体的氧转移限制的方法。氧限制程度由进入气流的量和组成以及使用的发酵设备的实际混合/物料转移性质决定。优选地,在氧受限条件下进行的方法中,氧消耗速率至少约为5.5,更优选地至少约为6,进一步更优选地至少约为7mmol/l/h。以下实施例阐述本发明:实施例1使用多种纤维素酶糖化玉米秆水解产物材料和方法评价三种不同的纤维素酶制剂糖化玉米秆水解产物的能力。将名为nl(dsmfoodspecialties,delft,荷兰)的talaromycesemersonii酶制品与bg和gc220(genencor-danisco,rochester,usa)比较。bg和gc220被认为是市场上之前可获得的基准酶。使用通过nrel制备的经稀酸预处理的玉米秆作为底物,用纤维素酶制剂进行糖化。将经预处理的玉米秆储存于4℃。浆体约为34%总固体,含约17%的不溶固体。用于预处理的玉米芯的组成在表1中公开。表1:秸秆原料的组成如下:a4个样品的平均值±标准差b计算值验证干物质浓度并证实为32.8%干物质浆(105℃,干燥48小时)。将60gr纤维与120gr水混合,使用4nnaoh将ph(1.9)调节至5.0,之后用水加至200gr,以获得9.45%的干物质软泥(sludge)。将10gr的软泥部分分装进50mlschott烧瓶中,并以三种不同的剂量(分别为20、61和204μl)添加每种酶制剂。随后封闭烧瓶,在50℃和280rpm下孵育140小时,在0,21,93和140小时时取样(3ml),使用eppendorf离心机离心,将上清液倾析进小瓶中,并使用nmr针对葡萄糖、阿拉伯糖、木糖和半乳糖进行分析。结果和讨论孵育开始时,显然稀酸预处理已经完成了对半纤维素级分的作用,因为与30g/l的理论酯相比已经存在24g/l的木糖。糖化开始时的糖组成在表2中公开;零时间时的游离葡萄糖浓度在4g/l左右。表2.糖化开始时的糖组成原材料时间(hrs)glug/lgalg/lxylg/larag/laceticg/llacticg/l总糖样品103.90.524.41.62.30.130.4样品203.90.523.91.52.30.129.8平均03.90.524.21.62.30.130.1在t=0时将ph设定为5.0,在93小时和140小时时测量并且在这两种情况下均为ph4.5,但是有机酸浓度未显著提高。ph降低可能影响了酶性能,但是对应用而言ph4.5比ph5.0更理想(归因于50℃下更低的细菌污染)。表3:对低、中和高酶剂量而言,以每克玉米秆干物质的mg酶蛋白质(bradford)为单位(mgep/gcsdm)的实施例1的酶剂量。图1a到c公开了这些实验中使用的三种酶制剂的糖化结果。图1a到1c中展示的数据表明,bg与gc220非常相似。然而,talaromycesnl制剂似乎非常有活性,因为其与gc220或bg相比每单位量酶蛋白质释放更多糖。通过bradford蛋白质测定法测定酶混合物的蛋白质含量,并且测定出对talaromycesnl制剂而言,实验中使用的蛋白质量比gc220或bg更少。见表3。这证明talaromycesnl制剂的比活性比所述比较中使用的其他纤维素酶制剂的比活性更高。比活性根据以下等式计算蛋白质的比活性:生产力={时间x时的[葡萄糖+阿拉伯糖+半乳糖+木糖](克单体糖)-时间0时的[葡萄糖+阿拉伯糖+半乳糖+木糖](克单体糖)]}/[总孵育时间(hrs)]/[孵育的蛋白质量(gr蛋白质)]以gr可发酵糖/gr酶蛋白质/hr为单位。仅针对与时间0时初始可获得的30g/l糖相比生产了多于34g/l总糖的实验计算比活性数据,因为低净生产水平下小的测量误差对比活性影响较大。在不同的时间点检验3种酶制剂的比活性,获得的结果公开于图2a到c和图3a到3c中。这些实验中获得的结果证明,对每份蛋白质量的糖或葡萄糖生产而口,nl酶制剂胜过其他基准市售酶。gc220与bg相当。实施例2使用经预处理的玉米秆用nl和saccharomycescerevisiae237ng进行ssf实验作为实施例1中所述糖化实验的跟进,使用nl和酵母(saccharomycescerevisiae237ng),以100ml的规模用经稀酸预处理的玉米秆针对纤维素乙醇进行同时糖化和发酵和蒸馏实验。使用这一技术去除抑制性葡萄糖,然后设想与原样水解相比获得更高的水解产率。这些实验的流程图公开于图4中。材料和方法如下计算每份生物质干重输入的乙醇产率和水解产率:1.干物质输入:干重测定48hrs,105c2.乙醇输出:在dma上测量的ml乙醇@100%3.葡聚糖含量:使用nrel样品,数据由nrel提供4.通过nmr测定洗涤液体中的糖(葡萄糖、半乳糖、木糖和阿拉伯糖的总量),从洗涤步骤的质量平衡计算水解产物中存在的总糖(糖浓度乘以测量的洗涤液重量)5.通过以下计算来自葡聚糖的理论乙醇产率:a)250ml烧瓶中的纤维干重量;纤维浆体的精确(100,0+/-0.1gr)重量乘以起始材料的干重含量(gr纤维dm)*100/330(我们制备了3烧瓶的100gr醪液,各来自调节ph后的330gr纤维-浆体制剂)数值100gr浆体中10gr左右纤维=10%dmb)通过由原料供应商获得的葡聚糖含量乘以我们自己测定的干物质含量(未洗涤的情况下3-4gr葡聚糖/100gr纤维浆体和经洗涤的纤维制剂中5-6gr葡聚糖)计算葡聚糖量c)通过用葡聚糖乘以180/162(由葡聚糖水解造成的化学增益因子(葡聚糖=纤维素=葡萄糖的聚合物))来计算潜在的葡萄糖量d)通过用潜在的葡萄糖的量乘以0.511gr/gr葡萄糖的基于葡萄糖的乙醇理论最大产率来计算潜在的葡萄糖量(预计为0.79gr乙醇/ml乙醇@100%),并预计91.5%的理论最大发酵产率(工业均值预计为91.5%+/-1.5%,这表示在90%和93%之间)e)(纤维素或葡聚糖)水解%=100*生产的乙醇量(gr)/理论最大乙醇(gr)结果和讨论使用不同的酶制剂时,低、中和高酶(分别为1、2和3)剂量获得了递增量的乙醇。10.2%干物质和34%葡聚糖含量(nrel-分析)下能够生产的理论最大乙醇量应当是102*0.34*180/162*0.91*0.511=17.9g乙醇/l培养基。如图5和6中所示,能够看出在最高酶剂量下达到所述乙醇量,证明可能用nl完全糖化。令人惊讶的是,所述嗜热酶在33℃的嗜温温度下也是非常有效的。实施例3使用nl糖化小麦秸如janlarsenetal.chem.eng.technol.2008,31,no.5,1-9所述,将小麦秸在195℃下用蒸汽处理12分钟。在摇床孵育箱中,在175rpm摇动下于ph3.8和60℃下,在8%的干物质下使用nl水解纤维,不进行任何酸或碱的添加。在纤维中50%葡聚糖含量下,应当预期所述实验中最多生产40g/l的葡萄糖,水解产率>85%,并且相对于40g/l总糖在92%发酵产率下所述水解产物的总乙醇潜能应为19g/l。图7公开了所述实验的结果,证明实现了所述葡萄糖水平。因此,酶制剂的酸性特性使得能够进行乙醇生产而不进行任何酸或碱添加,因为ph3.8对于酵母saccharomycescerevisiae而言也是最适的。实施例4使用nl糖化小麦秸以10l规模将实施例3中使用的含33-34%干物质的经预处理的小麦秸原料与水和下述酶溶液混合,所述酶溶液通过在10kd渗滤uf-单元上用水从市售制剂中透析出甘油(作为配制剂存在于商业产品中)并将所述酶浓缩10倍而获得。6小时后制剂中总干物质浓度为28%的小麦秸干物质。见表4。表4:进料分批操作中原料经预处理的小麦秸的糖化,在高酶剂量(高)和中酶剂量(中)下,t=1h、t=3h和t=6h时剂量的总的观察结果。-使用8nkoh和4nh2so4将ph控制在5.0+/-0.2-将温度控制在55和60℃之间。-使用1标准rushton涡轮在700rpm进行搅拌一天后已经能够测量到>110g/l的非常高的葡萄糖浓度,孵育两天后,通过离心去除剩余的木质素固体后在上清液中使用nmr糖测量法测量到128g/l的葡萄糖浓度,显示与根据文献预期的相比,所述酶被葡萄糖抑制的严重性较低。见图8。所述葡萄糖浓度是迄今为止用talaromyces纤维素酶制剂观察到的最高葡萄糖浓度,这使得能够使用所述酶进行商业shf方法同时实现65g/l乙醇的理论最大值,此时所有葡萄糖会被转化成乙醇(=0.511*128g/l)。理论最大产率如下计算发酵的理论最大产率(以每克葡萄糖gr产物为单位的ypsmax)。对乙醇而言,1摩尔葡萄糖(180gr)根据酵母中正常的糖酵解发酵通路产生2摩尔乙醇(=2×46=92gr乙醇)。乙醇相对于葡萄糖的理论最大产率因此是92/180=0.511gr乙醇/gr葡萄糖。对丁醇(mw74gr/摩尔)或异丁醇而言,理论最大产率是每摩尔葡萄糖1摩尔丁醇。因此(异)丁醇的ypsmax=74/180=0.411gr(异)丁醇/gr葡萄糖。对乳酸而言,纯乳酸发酵的发酵产率是每摩尔葡萄糖2摩尔乳酸(mw=90gr/摩尔)。根据这一化学计量,ypsmax=1gr乳酸/gr葡萄糖。表5:shf中可实现的葡萄糖浓度,和对具有不同ypsmax(g/g)的产物而言ssf中可实现的产物浓度。无葡萄糖抑制:表6示出了β葡糖苷酶的动力学比较。从表6中可以看出,talaromycesβ葡糖苷酶的ki(葡萄糖)非常低(0.045),这显示根据本发明的组合物不被葡萄糖阻抑。表6:来自不同来源的β葡糖苷酶的动力学比较:talaromycestrichodermaa.nigera.oryzae单位vmax(纤维二糖)108047040010131gr/gr蛋白质/hrkm(纤维二糖)0,020,680,052,39g/lki(葡萄糖)0,0450,1083245g/ls(纤维二糖)2,52,52,52,5g/l当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1