一种恒温条件下合成核酸的方法与流程
本发明涉及基因工程
技术领域:
,具体地,本发明涉及一种恒温条件下合成核酸的方法,特别涉及由特异性核苷酸序列构成的可形成特殊结构核酸的合成方法,及基于该特异性核酸序列的一种有用的扩增核酸的方法。
背景技术:
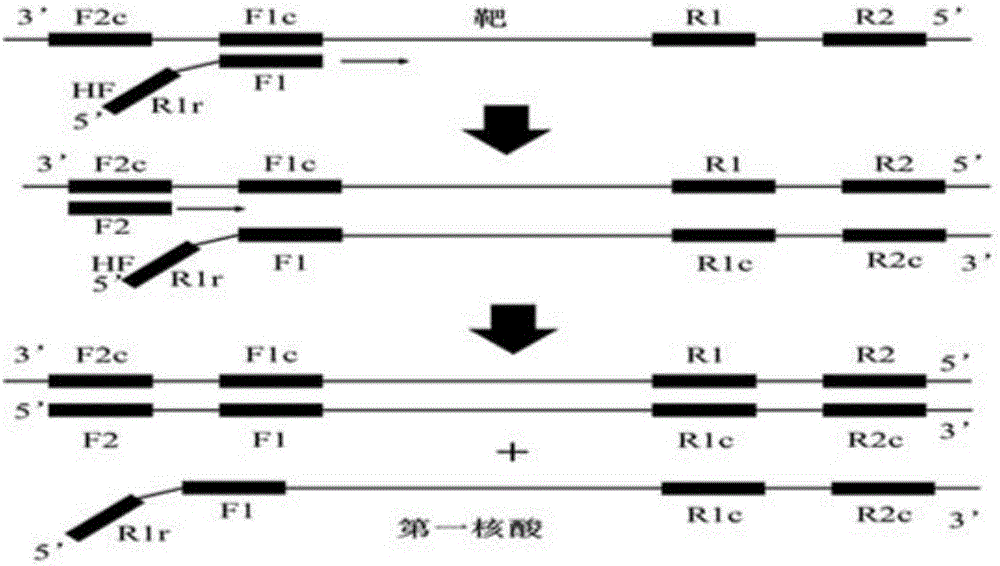
:生物体所携带基因信息的生物体的最根本差异,基于核苷酸序列互补性的分析方法可直接分析基因所携带的遗传特征。该分析是一种鉴定遗传疾病、癌变、微生物等非常强有力的方法。当样品中靶基因含量非常少时一般不易检测,因此必须对靶基因进行扩增或使其检测信号放大。作为扩增靶基因的方法,PCR方法被认为是最经典方法(Saiki,Gelfandetal.1988),亦是体外核酸序列扩增应用最为普遍的技术。该方法指数式扩增结果使其具有高灵敏度,确立了其在分子生物学方法检测领域的地位,经几十年发展已有系列成熟产品。此外,扩增产物可回收使用,因此作为一种支持遗传工程技术如基因克隆和结构决定的重要工具,亦得到广泛应用。然而,PCR方法明显有下述的问题:实际操作中必须要用专门的程序温度控制系统;扩增反应的指数式上升导致难于定量;样品和反应溶液易受到外部污染,假阳性问题较为突出。当前人类基因组计划已经完成,单核苷酸多态性(SNPs)显示出数量多、分布广泛的特点,故其分析逐步受到重视。通过设计引物使其核苷酸序列包含SNPs,依靠PCR扩增来检测SNPs是可行的,即通过是否存在与引物互补的核苷酸序列可通过是否存在反应产物的决定得出推断。然而,一旦PCR中偶然误合成互补链,在接下去的反应中该产物以模板运行,就会造成错误的结果。实际应用中,若引物末端仅一个碱基不同则将很难严格控制PCR,故必须改进特异性使PCR更好应用于SNPs的检测上。另一方面,相较于繁琐的程序温控过程合成核酸,科学家亦开发出在恒温条件下合成核酸的技术(Zhao,Chenetal.2015),主要包括以下几种:依赖核酸序列的扩增(NASBA)、滚环扩增(RCA),链置换扩增(SDA)、环介导等温扩增(LAMP)、依赖解旋酶的扩增(HDA)、重组聚合酶扩增(RPA)。NASBA,也称TMA(转录介导扩增方法)不需要复杂的温度控制。该方法通过DNA聚合酶以靶RNA为模板,加入连接有T7启动子的探针而得以合成,第二探针进入双链使产物得以生成,接着以生成的双链DNA为模板通过T7RNA聚合酶转录而扩增得到大量的RNA产物。NASBA需要热变性步骤直到双链DNA形成,但是接下去的转录反应在等温条件下通过T7RNA聚合酶得以进行。必须要用多种酶组合例如反转录酶,RNaseH,DNA聚合酶和T7RNA聚合酶,然而多种酶的组合对于费用是不利的。同时由于复杂的多种酶反应条件设定,该方法很难作为一般的分析方法得以推广。RCA(滚环扩增,RollingCircleAmplification)旨在模仿微生物中环状DNA的滚环复制过程,对于环状单链DNA模板,与该模板相结合的引物在原方法仅能实现对于环状核酸的扩增。为使得该方法可通用于线性DNA的扩增,通过挂锁探针(padlockprobe)或环形探针互补的单链DNA显示具有一系列核苷酸序列与挂锁探针(padlockprobe)或环形探针互补的单链DNA在靶核苷酸存在下可被连续的合成(Lizardi,Huangetal.1998)。该方法亦存在需多种酶的问题。而且,互补链合成的启动取决于连接两邻近区的反应,并且其特异性基本上与LCR中的相同。链替代扩增(StrandDisplacementAmplification,SDA)方法也是人们所知的扩增具有序列与靶序列互补的模板DNA的方法(Zhang,Cuietal.1992)。SDA方法应用特定的DNA聚合酶从目标核苷酸序列3’-侧互补的引物开始合成互补链以替换双链5’-侧的序列。由于新合成的互补链替换了5’-侧的双链,称该项技术为SDA方法。SDA方法中限制酶识别序列作为引物预先插入到退火序列中就可去除PCR方法中必须的温度变化步骤。即通过限制酶生成的切口供给3’-OH基作为互补链的合成起点,并且先合成的互补链通过链置换合成得以释放单链,接着再次作为模板用于下面的互补链合成。但SDA扩增产物与天然核酸结构不同,并且对用限制酶来断裂或将扩增产物应用到基因克隆上存在限制。这方面也是导致费用较高的主要原因。另外,SDA方法应用在未知序列时,用于引入缺口的限制酶识别序列相同的核苷酸序列可能存在于要被合成的区中,如此可能阻止完全互补链的合成。依赖解旋酶DNA恒温扩增技术(Helicase-dependentIsothermalDNAAmplification,HAD)是由美国NEB公司研究人员于2004年发明的一种新型核酸恒温扩增技术(Vincent,Xuetal.2004)。该技术模拟自然界生物体内DNA复制的自然过程,在恒温条件下利用解旋酶解开DNA双链,同时DNA单链结合蛋白(single-strandedDNA-bindingprotein,SSB)用以稳定解开的单链,并为引物提供结合模板,然后由DNA聚合酶催化合成互补链。新合成的双链在解旋酶的作用下又解成单链,并作为下一轮合成的模板进入上述的循环扩增反应,最终实现靶序列的指数式增长。LAMP技术(Notomi,Okayamaetal.2000)的核心是利用针对靶基因上的六个区域设计四条特异性引物依靠一种高活性链置换DNA聚合酶,使得链置换DNA合成在不停地自我循环。该技术可在15-60分钟内实现109-1010倍的扩增,反应能产生大量的扩增产物即焦磷酸镁白色沉淀,可以通过肉眼观察白色沉淀的有无来判断靶基因是否存在,日本荣研公司亦针对性开发出浊度仪以实现扩增反应实时监测。LAMP方法的优势除了高特异性和高灵敏度外,操作还十分的简单,在应用阶段对仪器的要求低,一个简单的恒温装置就能实现反应,结果检测也非常简单,直接肉眼观察白色沉淀或者绿色荧光即可,是一种适合现场、基层快速检测的方法。其中一个局限,因该方法高特异性和灵敏度等特点依赖于4条引物的性质,最佳引物的获得通常需要进行序列比对、在线引物设计、引物筛选及特异性试验,这一过程十分繁琐。重组酶聚合酶扩增(RecombinasePolymeraseAmplification,RPA),其要点在于:重组酶与引物结合形成的蛋白-DNA复合物,能在双链DNA中寻找同源序列。一旦引物定位了同源序列,随后链置换DNA聚合酶即会介导链交换反应形成并启动DNA合成,对模板上的目标区域进行指数式扩增。被替换的DNA链与单链结合蛋白(SSB)结合,以防止进一步替换。在这个体系中,由两个相对的引物起始一个合成事件。整个过程进行得非常快,一般可在十分钟之内获得可检出水平的扩增产物。但整个过程中需要筛选能与重组酶的结合并且特异性良好的引物,同时需要使用三种酶将极大增加其成本。解放军军事医学科学院的刘威等(Liu,Dongetal.2015)提出了一种称为聚合酶螺旋反应(PSR)的新型核酸等温扩增技术,该反应使用与LAMP反应一样的酶,通过在设计的PCR引物的5’端引入特定序列进行扩增形成5’和3’端可以自组装形成闭合螺旋状的结构以提供3’-OH作为合成的起点,并以之为扩增起始结构,而后大量合成相同序列的DNA单链,最终形成长链的螺旋结构产物。但此方法的引物序列中引入了外源基因,这将极大增加错配的可能性而导致错误的结果。此外,起始结构中的闭合环状结构中重叠成环部分仅为20碱基对左右,此将分子间氢键较弱进而使得螺旋环不稳定,同时若外源基因的长度过长则将影响螺旋环形成过程。技术实现要素:本发明的目的是提供一种合成核酸的方法,受到DNA常见双螺旋结构及LAMP和PSR方法的启发的新原理,以PCR引物为基础重新设计并规划了引物工作模式,螺旋环扩增起始结构,设计该结构环的形成过程,并且具有重叠成环部分碱基对较多、不引入外源基因片段、扩增产物可重新利用的新特点。更具体的是提供一种新型的依靠序列高效合成核酸的低成本方法。亦即,本发明的目的是提供通过在一种单酶及等温条件下完成核酸合成和扩增的方法。本发明的另一目的是提供一种核酸合成的方法,该方法通过改造已有的核酸合成反应原理很难达到的高特异性的核酸快速合成,还提供一种用该合成方法扩增核酸方法。本发明利用聚合酶催化链置换型的互补链合成而不需复杂的温度控制,有益于核酸的合成。该DNA聚合酶是SDA、RCA、LAMP和PSR等方法中用到的酶。本发明人改进了已知方法3’-OH的供给,结果发现通过利用具有特定结构的寡核苷酸,不需任何额外酶反应3’-OH结构就可被提供,由此得出本发明。即本发明涉及合成核酸的方法,通过用所述核酸合成方法扩增核酸的方法和应用所述方法的试剂盒。本发明的具有技术方案如下:本发明的恒温条件下合成核酸的方法,包括以下步骤:1)提供一种核酸,所述核酸的3’末端具有可与同一条链上的F1c区退火的F1r区,并且通过所述F1r区与Flc区的同时退火可形成螺旋环;2)以步骤1)所述核酸为模板合成其自身的互补链,所述互补链的R1rc区与R1区退火,而Flc区退火的F1r区的3'末端组成螺旋环,作为合成起点;3)通过聚合酶催化链置换型互补链合成反应而进行互补链合成,以置换步骤2)中所合成的互补链,其中所述多核苷酸在其3’末端包含一种与步骤2)中合成的互补链的任意区域互补的序列。根据本发明所述的合成核酸的方法,其中,步骤1)所述核酸的制备方法,包括以下步骤:i)退火步骤,使第一寡核苷酸I与模板的F1c区退火,其中该模板的3’末端包括F1c区和位于F1c区3’侧的F2c区,该模板的5’末端包括R1区和位于R1区5’侧的R2区,其中所述第一寡核苷酸I,包括R1r区与Fl区,所述Rlr区与F1区的5’侧相连,其中,F1区:具有与模板F1c区互补的核苷酸序列的区,R1r区:与模板R1区反向的区;ii)合成第一核酸的步骤,所述第一核酸具有与所述模板互补的核苷酸序列,此步骤以第一寡核苷酸I的F2区作为合成的起点合成第一核酸,所述第一核酸的3'末端具有可与同一条链上的部分F1c区退火的F1区,并且通过所述F1区与Flc区的同时退火可形成的螺旋环;iii)利用聚合酶催化链置换反应置换步骤ii)中合成的第一核酸,其中与模板中的F1c的3’侧的F2c区退火的第一外引物I用作合成的起点,和iv)退火步骤,使第二寡核苷酸II与步骤iii)所得第一核酸的R1c区退火,其中所述第二寡核苷酸II包括R1区和F1r区,并且Flr区与R1区的5’侧相连,第二寡核苷酸II为第一寡核苷酸I的反向序列;其中,R1区:具有与第一核酸的R1c区互补的核苷酸序列的区,Flr区:与第一核酸的F1区反向的区;v)以所述第二寡核苷酸II作为合成的起点,合成第二核酸,并利用聚合酶催化链置换反应置换该第二核酸获得步骤1)所述的核酸;其中,置换第二核酸时,与第一核酸中的R1c的3’侧的R2c区退火的第二外引物II用作合成的起点。进一步地,步骤i)所述模板为RNA,步骤ii)中的第一核酸通过具有反转录酶活性的酶来合成。本发明适用于各种RNA,例如各种病毒的RNA等等。比如,用于MERS-CoV病毒的RNA检测,如:该RNA的orf1a,orf1b区段等。本发明在形成起始螺旋结构的过程中,所使用的寡核苷酸中连接F1区与R1r区,或者连接R1区与F1r区的序列可以为无,但如若引入连接序列(linker,L),该序列必须为反向序列,即两寡核苷酸为5’-R1r-L-F1-3’、5’-F1r-Lr-R1-3’中L和Lr须为反向互补序列(R1r为R1的反向序列,F1r为F1的反向序列),linker一般为限制性内切酶位点序列。根据本发明所述的合成核酸的方法,其中,所述合成的核酸是指在其一条链上具有首尾互补核苷酸序列的核酸。根据本发明所述的合成核酸的方法,其中,所述恒温是指整个反应过程在60-65℃的恒定温度下进行合成。根据本发明所述的合成核酸的方法,其中,每种寡核苷酸与其在模板中的互补区之间的解链温度在相同严谨条件下存在以下的关系:(外引物或模板3'侧区)≤(F2c或F2,以及,R2c或R2)≤(F1c或F1,以及,Rlc或Rl)。根据本发明所述的合成核酸的方法,作为优选,所得到的第二核酸可以通过引入加速探针Xin的方法使得核酸扩增加速进行,其中Xin是位于F1区到R1区的中间区段。本发明可以以步骤3)置换后的互补链为模板重复步骤1)-3)合成长链核酸。本发明所述聚合酶催化链置换反应中使用的聚合酶为BstDNA聚合酶、Bca(exo-)DNA聚合酶、DNA聚合酶I克列诺(Klenow)片段、VentDNA聚合酶、Vent(Exo-)DNA聚合酶(缺少核酸外切酶活性的VentDNA聚合酶)、DeepVentDNA聚合酶、DeepVent(Exo-)DNA聚合酶(缺少核酸外切酶活性的DeepVentDNA聚合酶)、Φ29phageDNA聚合酶以及MS-2phageDNA聚合酶等中的一种或几种。其中优选使用BstDNA聚合酶或Bca(exo-)DNA聚合酶。根据本发明所述的合成核酸的方法,其中,所述聚合酶催化链置换反应中可以加入解链温度调节剂。其中,所述解链温度调节剂优选为甜菜碱,进一步优选地,允许反应溶液中甜菜碱的浓度为0.2-3.0M。本发明的用于恒温条件下合成核酸的试剂盒,其特征在于,所述试剂盒包括:一寡核苷酸I,其包括F1区和R1r区,所述R1r区与F1区的5’侧相连,其中,F1区:具有与模板的F1c区互补的核苷酸序列的区,和R1r区:与模板的R1区反向的区;一寡核苷酸II,其包括R1区和F1r区,所述F1r区与R1区的5’侧相连,其中,R1区:具有与模板的R1c区互补的核苷酸序列的区,和F1r:与模板的F1区反向的区;第一引物I,其具有与作为模板的核酸中F1c区3'侧的F2c区互补的核苷酸序列;第二引物II,其具有与作为模板的核酸中R1c区3'侧的R2c区互补的核苷酸序列;催化链置换型互补链合成反应的DNA聚合酶,和,核苷酸,其作为所述DNA聚合酶的底物。根据本发明所述的试剂盒,其中,所述试剂盒还包含用于检测核酸合成反应产物的检测试剂。根据本发明所述的试剂盒,其中,所述试剂盒还包括加速探针Xin,其中所述Xin是位于模板的F1区到R1区的中间区段的核苷酸片段,具体地,即位于F1至R1之间的Fin及Rin区域。根据本发明所述的试剂盒,其中,所述DNA聚合酶为BstDNA聚合酶、Bca(exo-)DNA聚合酶、DNA聚合酶I克列诺片段、VentDNA聚合酶、Vent(Exo-)DNA聚合酶、DeepVentDNA聚合酶、DeepVent(Exo-)DNA聚合酶、Φ29phageDNA聚合酶以及MS-2phageDNA聚合酶等中的一种或几种。其中优选使用BstDNA聚合酶或Bca(exo-)DNA聚合酶。本发明提供了上述试剂盒在合成核酸或检测样品中靶核苷酸序列中的应用。基于本发明的合成核酸的方法,提供一种检测样品中靶核苷酸序列的方法,包括以靶核苷酸为模板,通过本发明的合成核酸的方法进行扩增,并观察是否生成扩增产物。在上述扩增产物中加入包含与形成的螺旋环互补的核苷酸序列的探针,进而观察两者之间的杂交。还可以将所述探针标记在颗粒上,并观察通过杂交而发生的聚集反应。所述的扩增方法可以在核酸检测试剂的存在下实施,并根据该检测试剂的信号变化来观察是否生成扩增产物。基于本发明的合成核酸的方法,还可以提供一种检测样品中靶核苷酸序列突变的方法,包括以靶核苷酸为模板,通过本发明所述的合成核酸的方法进行扩增。其中,核苷酸序列中作为扩增对象的突变阻碍了组成该扩增方法的任一互补链的合成,从而检测出突变。具有首尾为互补核苷酸序列的单链核酸是本发明合成的目的,该核酸指的以靶核酸序列为具有互相互补核苷酸序列首位并排连接在单链里的核酸。此外,本发明中应包含用于在互补链间成螺旋的核苷酸序列。本发明中该序列称为成螺旋环序列。本发明合成的核酸基本上由通过成螺旋环序列连接的互相互补的链组成。一般而言,不管是否部分涉及碱基配对,一个在配对碱基分离时不能被分离成两个或更多分子的链称为单链。同一链中互补核苷酸序列可形成碱基配对,本发明通过容许具有核苷酸序列首尾连接在单链里的核酸在同一链内碱基配对,可获得分子内碱基配对的产物,该产物包含组成明显双链的区和不涉及碱基配对的环。也就是,本发明具有互补核苷酸序列首尾连接在单链里的核酸可被定义为单链核酸,其中包含能在同一链中退火的互补核苷酸序列,并且其退火产物,在弯曲部分组成不涉及碱基配对的环。具有互补核苷酸序列的核苷酸可退火成不涉及碱基配对的环。成环序列可以是任意的核苷酸序列。成环序列能碱基配对以启动用于置换的互补链的合成。并优先地被提供与位于其它区的核苷酸序列不同的序列,以获得特异性退火。本发明中基本相同的核苷酸序列定义如下:当以某序列作为模板合成的互补链与靶核苷酸序列退火以供给合成互补链的起点时,该某序列基本上与靶核苷酸序列相同。例如,基本上与F1相同的序列不但完全包括与F1相同的序列,还包括能作为模板的核苷酸序列,所述模板能给出与F1退火的核苷酸序列并能作为合成互补链的起点。本发明术语“退火”指的是通过根据沃森-克里克定律的碱基配对,形成互补结构的核酸。故而,即使组成碱基配对的核酸链为单链,如果分子内互补核苷酸序列碱基配对,退火亦会发生。通过碱基配对核酸组成双链结构,故本发明退火和杂交所表达的含义有重合部分。本发明组成核酸的核苷酸序列对数至少为1。本发明所期望的模型中,核苷酸序列对数可为1的整倍数。该情况中,本发明组成核苷酸的互补核苷酸序列对数理论上没有上限,在由多组互补核苷酸序列构成的本发明合成产物核酸时,该核酸由重复相同的核苷酸序列组成。本发明合成的具有首尾可互补成环序列的单链核苷酸与天然存在的核酸不可能有相同的结构,并且一般已知当通过核酸聚合酶作用合成核酸时如果用核苷酸衍生物作为底物,就可合成核酸衍生物。所用核苷酸衍生物包括放射性同位素标记的核苷酸或结合配体标记的核苷酸衍生物例如生物素或地高辛。这些核苷酸衍生物可用于标记产物核酸。或者,如果底物是荧光核苷酸,则产物核酸可能为荧光衍生物。而且产物可为DNA亦可为RNA。生成的产物通过结合实现核酸聚合反应的引物结构,聚合反应底物类型,聚合反应的试剂而定。利用DNA聚合酶能启动有上述结构的核酸的合成,该DNA聚合酶具有链置换活性以及F1r区具备在3’-末端与同一链上的部分Flc区退火形成合成互补链。有许多关于互补链合成反应的报道,其中形成发夹环,以发夹环序列自身为模板,亦有形成螺旋环,以螺旋环序列自身为模板,而本发明中提供给螺旋环其首尾部分能碱基配对的区,并且具有在合成互补链时利用该区的新特点。通过将该区用作合成的起点,先前以螺旋环序列自身为模板合成的互补链被替换。本发明用到术语“核酸”,本发明核酸通常既包括DNA又包括RNA。然而,功能为合成互补链的模板的,来自天然DNA或RNA的其核苷酸被人工衍生物所替换的核酸或修饰核苷酸亦包括在本发明的核酸范围中。通常本发明的核酸被包含于生物样品中,生物样品包括动物,植物或微生物的组织,细胞,培养物和分泌物,以及它们的提取物。本发明的生物样品包括细胞内寄生物基因组DNA或RNA,例如病毒或支原体。本发明的核酸一般由包含在所述生物样品的核酸衍生而来。例如由mRNA合成cDNA,基于生物样品衍生来的核酸而扩增的核酸,是本发明的核酸的典型实例。本发明核酸的特征是在3’-末端被提供F1r区,可与同一链上的部分Flc退火,通过该F1r区与同一链上的Flc退火,可形成包含可碱基配对的R1区在内的环,可在各种方法中得到该核酸。组成基于本发明所述寡核苷酸的核苷酸序列特征所用术语“相同的”和“互补的”并不意味着绝对相同和绝对互补。也就是,与某序列相同的序列包括与某序列退火的核苷酸序列互补的序列。另一方面,互补序列是严格条件下能退火的序列,提供作为互补链合成的起点3’-末端。本发明中,寡核苷酸是满足两个要求的核苷酸,即必须能形成互补碱基配对,并且在3’-末端供给-OH基为互补链合成的起点。因此,其主链并不必限于磷酸二酯键一种连接。例如,它可由硫代磷酸衍生物组成主链或者是基于肽连接的肽核酸,所述硫代磷酸衍生物为S取代O。碱基是指那些可互补配对的碱基。天然存在五种碱基,即A,C,T,G和U,碱基也可为类似物例如溴脱氧尿苷。优选的是,本发明寡核苷酸不仅可用做合成的起点还可为互补链合成的模板。本发明术语多核苷酸包括寡核苷酸。本发明所用术语“多核苷酸”的链长无限制,而所用术语“寡核苷酸”指的是有相对较短的链长的核苷酸聚合物。下述各种核酸合成反应中在给定的条件下,本发明寡核苷酸链有能与互补链碱基配对并保持一定的特异性这样一种长度。具体地,它由5-200个碱基组成,更优选10-50个碱基对。识别已知聚合酶的链长至少为5个碱基。该聚合酶催化依靠序列的核酸合成反应。故而退火部分的链长应长于该长度。另外,统计学上所期望10个碱基的长度或更长以获得目标核苷酸特异性。另一方面,由于化学合成制备太长核苷酸序列比较困难。因此上述链长是所期望范围的实例。例证的链长指的是部分与互补链退火的链长。正如下面所描述的,本发明寡核苷酸可最终至少分别与两区退火。因此,这里例证的链长应理解为组成寡核苷酸的每个区的链长。此外,本发明的寡核苷酸可用已知的标记物标记。标记物包括结合配体例如地高辛和生物素,酶,荧光物,发光物,放射性同位素。众所周知通过荧光类似物替换组成寡核苷酸的碱基的技术(W095/05391,Proc.Natl.Acad.Sci.USA,91,6644-6648,1994)。本发明其它寡核苷酸还可被结合到固相。或者,寡核苷酸的任意部分可用结合配体标记,例如生物素,间接地由结合配体例如固定抗生物素蛋白所固定。固定寡核苷酸为合成的起点时,合成反应产物核酸为固相所捕获,这将利于其分离。通过核酸特异性指示物或与标记探针的杂交可对分离部分进行检测。通过本方法所获得的核酸产物,针对其中靶核酸片段可通过限制酶消化产物而得以回收。本发明所用术语“模板”是指用于合成互补链时作为模板的核酸。具有核苷酸序列与模板互补的互补链意思是指对应于模板的链。但是二者的关系只是相对的。即合成的互补链可以再次起到模板的功能。也就是,互补链亦可作为模板。在本发明中,如果目标为RNA,可仅通过添加反转录酶构成,即用RNA为模板,通过反转录酶在模板中F1与F1c的退火有可能合成互补链和从退火为F2c外引物F2为合成起点合成互补链以及同时替换先前合成的互补链,外引物F2位于F1c的3’-侧。当反转录酶以DNA为模板进行合成互补链的反应时,所有通过反转录酶进行合成互补链的反应包括以与Rlc退火的R1作为合成起点的互补链的合成,该互补链在链置换反应中作为模板;以与R2c退火的R2作为合成起点的互补链的合成及同时的链置换发应,R2位于R2c的3’-侧。在所给反应条件下不能预计反转录酶显示DNA/RNA链置换活性时,可以结合具有如上述链置换活性的DNA聚合酶。如上述以RNA为模板获得第一条单链核酸的模式为本发明的优选模式。另一方面,如果使用既具有链置换活性又有反转录酶活性的DNA聚合酶如BcaDNA聚合酶,通过相同的酶不但从RNA的第一条单链核酸的合成,而且接下去以DNA为模板的反应可得以类似地进行。反应在下面成分存在下进行,能使酶反应处于合适pH值的缓冲液,退火或维持酶催化活性的必需盐,保护酶的介质以及调控解链温度(Tm)所必须的调控物。对于缓冲液,例如所用的在中性或弱碱性范围有缓冲作用的Tris-HCl。根据所用的DNA聚合酶调节pH值,对于盐,KCl,NaCl,(NH4)2SO4等适量加入以保持酶的活性并调控核酸的解链温度(Tm),保护酶的介质使用牛血清白蛋白或糖类。此外,一般用二甲基亚砜(DMSO)或甲酰胺作为解链温度(Tm)的调控物。通过利用解链温度(Tm)的调控物在限定的温度条件下寡核苷酸的退火得到了调控。而且,甜菜碱(N,N,N-三甲基甘氨酸)或四烷基铵盐(tetraalkyl)通过其等稳定作用(isostabilization)对于改善链置换的效率也是有效的。通过向反应溶液中加入0.2-3.0M甜菜碱,优选0.5-1.5M,可得到所希望的本发明对核酸扩增的促进作用。因为这些解链温度的调控物有降低解链温度的作用,那些合适的严谨性和反应性条件要结合盐的浓度,反应温度等凭经验而定。本发明重要的特征是除非许多区的位置关系得以保持,否则一系列反应不能进行。由于这个特征,伴随互补链非特异合成的非特异合成反应得到了有效阻止。也就是即使发生某非特异反应,在合成接下去的扩增步骤中产物作为起始物质的可能性也得到了降低,而且,通过许多区调控反应的进展,有可能导致在类似的核苷酸序列中能精确的鉴定出所需产物的检测系统可被随意地组成。本发明合成的核酸是单链,就单链而言,由互补核苷酸序列构成,其大部分均为碱基配对的。通过利用这个特征,对合成的产物可进行检测。通过实施本发明合成核酸的方法,在有荧光色素作为双链特异性嵌入剂(double-specificintercalater)例如溴化乙锭、SYBRGreenI、PicoGreen或EvaGreen,随着产物的增加可观察到荧光的强度增加。通过监测荧光强度,就可能在封闭系统中跟踪实时(real-time)合成反应进行情况。也可考虑在PCR方法中应用该类型的检测系统,但有许多问题,因为不能区分产物信号和引物二聚物的信号等。然而,当本发明应用该系统时,增加非特异碱基配对的能力非常低,因此,预计高灵敏度和低干扰可能同时能获得,与应用双链特异性嵌入剂(double-specificintercalater)相似,在同一系统中可利用荧光能量的转移用于实现检测系统的方法。本发明合成核酸的方法通过DNA聚合酶催化合成链置换型的互补链反应而得到支持。上述反应期间,也包含不必需链置换型聚合酶的反应步骤。然而,为了组成试剂的简单化及经济的观点,使用一种DNA聚合酶有利,该种DNA聚合酶,下列的酶是已知的。此外,本发明范围中可利用这些酶的各种突变体,它们都具有用于互补链合成的序列-依赖活性和链置换活性。其中突变体指的是包括那些仅具有导致酶所需的催化活性的结构或那些通过例如氨基酸中突变对催化活性,稳定性或热稳定性所进行的修饰的突变体。BstDNA聚合酶Bca(exo-)DNA聚合酶DNA聚合酶I克列诺(Klenow)片段VentDNA聚合酶Vent(Exo-)DNA聚合酶(缺少核酸外切酶活性的VentDNA聚合酶)DeepVentDNA聚合酶DeepVent(Exo-)DNA聚合酶(缺少核酸外切酶活性的DeepVentDNA聚合酶)Φ29phageDNA聚合酶MS-2phageDNA聚合酶这些酶中,BstDNA聚合酶或Bca(exo-)DNA聚合酶是特别所需的酶,因为它们具有某种程度的热稳定性和高催化活性。在优选的实施方案中,本发明的反应可等温的实现,但由于解链温度的调节(Tm)等,不可能总是能利用所需温度条件来维持酶的稳定。因此,它是酶的热稳定所需的条件之一。尽管等温反应是可行的,热变性可提供核酸作为最初的模板,在这方面,热稳定酶的使用拓宽了试验方案的选择。本发明合成或扩增核酸所必需的各种试剂可被预先包装,并以试剂盒的形式提供,具体地,本发明所提供的试剂盒包含作为合成互补链合成的引物和用于置换反应的外引物所必需的各种寡核苷酸,用于互补链合成的底物dNTP,用于实现链置换型互补链合成的DNA聚合酶,为酶反应提供合适条件的缓冲液,和用于检测合成反应产物所必需的介质。具体地,本发明优选的模式中,反应期间无需加入的试剂,并由此对于移入反应容器后一个反应所必需供给的试剂,其中仅通过加入样品就可启动该反应。通过利用可见光信号或荧光信号可在容器内检测反应产物的系统。反应后不必打开和关闭容器。这对于预防污染是非常有利的。本发明合成具有首尾序列可退火成环的核苷酸序列的单链核酸。该核酸具有例如下面的用途:第一特征是利用一分子中具有互补序列的特定结构带来的优势,该特征可能利于检测,即有已知用于检测核酸的系统,其中其变化的信号取决于与互补核苷酸序列碱基配对。例如,通过结合使用双链特异性嵌入剂作为如上所述的检测试剂的方法,充分利用本发明合成产物特征的检测系统可得以实现。如果本发明合成反应的产物在所述检测系统发生一次热变性,并且返回到原始温度,分子内退火优先发生,并因此容许互补序列之间快速碱基配对。如果存在非特异性产物,分子中它们没有互补序列从而使通过热变性分离成2个或更多的分子后,它们不能立刻就返回到原始双链。通过在检测前提供的热变性步骤,伴随非特异反应的干扰得以降低。如果所用的DNA聚合酶不抗热,热变性步骤有反应终止的意思,并因此有利于控制反应温度。第二特征是常常形成能碱基配对的首尾连接的闭合环。能碱基配对的环的结构在图3中显示。如图3中看到的该环由核苷酸序列F1,R1,F1c,R1c构成,可进行分子内退火形成闭合环。根据本发明优选的模式,在单链核酸中供给大量能碱基配对的环。这意味着大量的探针可与一分子核酸杂交以容许高灵敏度的检测。因此不仅可能实现改进灵敏度还可能实现基于特殊反应原理例如聚集作用来检测核酸的方法。例如,将固定在精细颗粒例如聚苯乙烯乳胶上的探针加入到本发明反应产物中,观察乳胶颗粒的聚集作用为产物与探针杂交。聚集作用的强度通过光学测定就可进行高灵敏度和定量观察。或者还可通过裸眼观察聚集作用,故还可建立不用光学的测定装置的反应系统。此外,本发明反应产物允许一些可结合的标记,其中每核酸分子可进行层析检测。在免疫测定领域里,实际所应用是利用可见的检测标记使用层析介质的分析方法(免疫层析)。该方法基于分析物夹在固定于层析介质上的抗体和标记抗体间的原理,未反应的标记成分被洗脱。本发明的反应产物使该原理应用到核酸分析上。也就是,制备针对环部分的标记探针并固定在层析介质上为捕获准备捕获探针,以允许在层析介质里进行分析。序列与环部分互补的捕获探针得以利用,由于本发明的反应产物具有大量的环,产物与大量标记的探针结合以给出肉眼可识别信号。本发明反应产物常常能供给碱基配对的环区,能拓宽其它各种检测系统。例如,利用表面胞质基因组使用固定探针检测该环部分的系统是可行的。此外,如果用双链特异嵌入物标记该环部分的探针,就可进行更灵敏的荧光分析。或积极利用本发明合成核酸的能力在3’-和5’-侧以形成能碱基配对的螺旋环。例如,设计一个环使其在正常型和不正常型间有共同的核苷酸序列,而设计其它环使其在其中产生差异。通过探针证实共同部分存在基因,而在其它区证实有不正常存在时,有可能组成特征分析系统。因为本发明合成核酸的反应也能等温的进行,值得一提的优点是,通过一般荧光光度计可进行实时(real-time)分析。直到此时,同一链中要退火的核酸的结构是已知的。然而,通过本发明获得的具有首尾退火成环的序列单链里的核酸是新的,它包含大量的能与其它碱基配对的环。另一方面,通过本发明反应产物所给的大量的环自身能被用作探针,例如,在DNA芯片里,探针在有限的区域内高密度堆积,而该技术中可固定在某区域寡核苷酸数量有限,因此通过利用本发明产物大量能退火的探针可被高密度固定,即本发明的反应产物在DNA芯片上可用作固定的探针,扩增后反应产物可通过本领域已知的任何技术得以固定,或用固定的寡核苷酸作为本发明扩增反应的寡核苷酸,导致生成固定反应产物。因此通过使用固定的探针,大量样品DNA在有限的区域内得以杂交,结果预计可得到高信号值。附图说明图1是本发明合成第一核酸的步骤图解。图2是本发明合成第二核酸的步骤图解。图3是通过本发明单链核酸所形成的环结构的图解。图4是通过本发明单链核酸所形成的环结构通过添加额外引物加速图解。图5是显示MERS-orf1b靶核苷酸序列中组成寡核苷酸的每个核苷酸序列的位置关系。图6是是显示通过以MERS-orf1b为模板合成本发明单链核酸的方法获得的产物琼脂糖电泳结果的照片。泳道1:碧云天O0107DNALadder泳道2:1fmolMERS-orf1bdsDNA图7是显示MERS-orf1b靶核苷酸序列中组成寡核苷酸的每个核苷酸序列的位置关系。图8是显示限制酶消化产物的琼脂糖凝胶电泳结果的照片,其中所述产物是通过本发明核酸合成反应在实施例1中得到的。其中,泳道1:碧云天O0107DNALadder;泳道2:1fmolMERS-orf1bdsDNA;泳道3:纯化产物的HindIII消化。图9是显示在引物的作用下,含MERS-orf1b靶核苷酸序列DNA增加的实时荧光曲线。图10是显示在引物的作用下,含MERS-orf1b靶核苷酸序列DNA增加的实时荧光曲线。图11是显示在引物的作用下,通过添加加速引物条件下,不同DNA靶浓度扩增体系的荧光强度随反应时间变化的曲线。图12是本发明的合成核酸的方法的流程示意图。图13是本发明合成的核酸螺旋示意图。具体实施方式下述实施例中所使用的实验方法如无特殊说明,均为常规方法,具体可参照《分子克隆实验指南》(第三版)J.萨姆布鲁克一书中所列的具体方法进行,或者按照试剂盒和产品说明书进行;下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。实施例1对于MERS-orf1b中片段的扩增本发明具有互补链以螺旋环的形式连接到单链里的核酸是利用MERS-orf1b(来自于GenBank:NM_001012270.1)为模板尝试的。实验中用到四种引物即Mo1bHF,Mo1bHR,Mo1bF2和Mo1bR2。Mo1bF2和Mo1bR2是置换分别以Mo1bHF和Mo1bHR为合成起点获得的第一条核酸的外引物。因为以Mo1bHF(或Mo1bHR)合成后外引物为互补链合成起点的引物。通过利用临近堆积现象将这些设计成退火成环的区。此外,将这些引物设置为高浓度使Mo1bHF(或Mo1bHR)的退火优先发生。组成每个引物的核苷酸序列如序列表所示,引物的结构特征在下面概括。此外,针对靶核苷酸序列每个区的位置关系在图5中显示。通过这些引物合成本发明核酸的方法的反应溶液的组合如下表1所示。表1引物与寡核苷酸信息引物5’-侧的区/3’-侧的区Mo1bHF与通过Mo1bHF合成的互补链中Flc区相同/与MERS-orf1b中R1区反向Mo1bHR与通过Mo1bHB合成的互补链中Rlc区相同/与MERS-orf1b中F1区反向Mo1bF2与MERS-orf1b中F1c区3’-侧临近的F2c互补Mo1bR2与MERS-orf1b中R1区5’-侧临近的R2c互补通过所述引物,合成R1与R1rc、F1c与F1r互补成螺旋环的核酸。通过这些引物合成本发明核酸的方法的反应溶液的组合在下面所示。反应溶液组合(25μL)20mMTris-HClpH8.810mMKCl10mM(NH4)2SO414mMMgSO40.1%TritonX-1001M甜菜碱1.25mMdNTP8UBstDNA聚合酶(NEWENGLANDBiolabs)引物:1600nMMo1bHF/SEQIDNO.l1600nMMo1bHR/SEQIDNO.2200nMMo1bF2/SEQIDNO.3200nMMo1bR2/SEQIDNO.4靶:MERS-orf1bdsDNA/SEQIDNO.5具体的各核苷酸序列如下:SEQIDNO.l:GTACGAAGGGCATTACGCTCTCGTGTTATTTCCAGG;SEQIDNO.2:GGACCTTTATTGTGCTCTCGCATTACGGGAAGCATG;SEQIDNO.3:TACCCGCAAATGTCCCATA;SEQIDNO.4:TGTAGAGGCACATTGGTG;SEQIDNO.5:AAGACGAGTGATGAGCTTTGCGTGAATCTTAATTTACCCGCAAATGTCCCATACTCTCGTGTTATTTCCAGGATGGGCTTTAAACTCGATGCAACAGTTCCTGGATATCCTAAGCTTTTCATTACTCGTGAAGAGGCTGTAAGGCAAGTTCGAAGCTGGATAGGCTTCGATGTTGAGGGTGCTCATGCTTCCCGTAATGCATGTGGCACCAATGTGCCTCTACAA。混合物于63℃反应1小时,反应后,于80℃10分钟终止该反应,然后重新转到用冰预冷的水中。反应的证实:将1μL上样缓冲液力加到5μL上面的反应溶液中,样品于90mV在GelRed预染的(Biotum)1%琼脂糖凝胶(TAE溶解)电泳1小时。用碧云天O0107DNALadder作为分子量标记。电泳后的凝胶以验证核酸。结果在图6中显示,各泳道分别相对于下面的样品。1.分子量标记DNAladder2.1fmolM13mpl8dsDNA。实施例2通过限制酶的消化证实反应产物为了阐清具有互补核苷酸序列以环状结构连接在单链内的本发明实施例1获得的核酸结构,用限制酶消化产物。如果通过消化能生成理论上的片段,同时不存在(disappear)如实施例1中观察到的高分子量处产生不清晰成片条带模式和未被电泳的带,就可预计任何这些产物为本发明具有互补序列交替地连接在单链内的核酸。实施例1中反应溶液通过用酚处理及乙醇的沉淀作用得以沉积和纯化,回收产生的沉淀并重新溶于超纯水中,用限制酶HindIII于37℃消化2小时,样品于90mV在GelRed预染的(Biotum)1%琼脂糖凝胶(TAE溶解)电泳1小时。用碧云天O0107DNALadder作为分子量标记。电泳后的凝胶以验证核酸。结果在图7中显示,各泳道分别相对于下面的样品。1.分子量标记DNAladder2.1fmolM13mpl8dsDNA3纯化产物的HindIII消化实施例3应用EvaGreen验证反应产物EvaGreen同SYBRGreenI类似,是一种结合于所有dsDNA双螺旋小沟区域的具有绿色激发波长的染料,其对PCR等核酸扩增反应的抑制远小于后者。在游离状态下,EvaGreen发出微弱的荧光,但一旦与双链DNA结合后,荧光大大增强。因此,EvaGreen的荧光信号强度与双链DNA的数量相关,可以根据荧光信号检测出核酸扩增体系存在的双链DNA数量。通过这些引物合成本发明核酸的方法的反应溶液的组合在下面所示。反应溶液组合(25μL)20mMTris-HClpH8.810mMKCl10mM(NH4)2SO414mMMgSO40.1%TritonX-1001M甜菜碱1.25mMdNTP8UBstDNA聚合酶(NEWENGLANDBiolabs)1XEvaGreen(Biotum)120μM引物:1600nMMo1bHF/SEQIDNO.l1600nMMo1bHR/SEQIDNO.2200nMMo1bF2/SEQIDNO.3200nMMo1bR2/SEQIDNO.4靶:MERS-orf1bdsDNA/SEQIDNO.5设置ABIStepOnerealtimePCR反应温度恒定为63℃,反应时间为75min。荧光强度随反应时间变化的曲线如图8所示。将荧光检测应用于其中可实现实时监测的目的,通过实时扩增曲线可提前判断结果。实施例4应用基于EvaGreen的实时荧光比较不同浓度靶基因扩增反应溶液组合(25μL)八管20mMTris-HClpH8.810mMKCl10mM(NH4)2SO414mMMgSO40.1%TritonX-1001M甜菜碱1.25mMdNTP8UBstDNA聚合酶(NEWENGLANDBiolabs)1XEvaGreen(Biotum)引物:1600nMMo1bHF/SEQIDNO.l1600nMMo1bHR/SEQIDNO.2200nMMo1bF2/SEQIDNO.3200nMMo1bR2/SEQIDNO.4靶:MERS-orf1bdsDNA/SEQIDNO.5相同的反应液设置8管,仅靶的添加有区别分别为108Copies,107Copies,106Copies,105Copies,104Copies,103Copies,102Copies,纯水(0Copies)。设置ABIStepOnerealtimePCR反应温度恒定为63℃,反应时间为90min。不同靶浓度扩增体系的荧光强度随反应时间变化的曲线如图9所示。此结果说明本发明方法所获得的结果线性关系较好,可应用于目标核酸定量检测。实施例5应用基于EvaGreen的实时荧光比较不同浓度RNA靶基因扩增AMV反转录酶可以RNA为模板合成cDNA,配合BstDNA聚合酶可以实现RNA的检测。反应溶液组合(25μL)20mMTris-HClpH8.810mMKCl10mM(NH4)2SO414mMMgSO40.1%TritonX-1001M甜菜碱1.25mMdNTP8UBstDNA聚合酶(NEWENGLANDBiolabs)10UAMV反转录酶1XEvaGreen(Biotum)引物:1600nMMo1bHF/SEQIDNO.l1600nMMo1bHR/SEQIDNO.2200nMMo1bF2/SEQIDNO.3200nMMo1bR2/SEQIDNO.4靶:MERS-orf1bssRNA/SEQIDNO.5相同的反应液设置8管,仅靶的添加有区别,分别为108Copies,107Copies,106Copies,105Copies,104Copies,103Copies,102Copies,纯水(0Copies)。设置ABIStepOnerealtimePCR反应温度恒定为63℃,反应时间为90min。不同靶浓度扩增体系的荧光强度随反应时间变化的曲线如图10所示。此结果说明本方法应用于RNA检测同样可行。实施例6应用Fin对于不同浓度靶基因扩增反应溶液组合(25μL)八管20mMTris-HClpH8.810mMKCl10mM(NH4)2SO414mMMgSO40.1%TritonX-1001M甜菜碱1.25mMdNTP8UBstDNA聚合酶(NEWENGLANDBiolabs)1XEvaGreen(Biotum)引物:1600nMMo1bHF/SEQIDNO.l1600nMMo1bHR/SEQIDNO.2200nMMo1bF2/SEQIDNO.3200nMMo1bR2/SEQIDNO.4800nMMo1bFin/SEQIDNO.6800nMMo1bFin/SEQIDNO.7靶:MERS-orf1bdsDNA/SEQIDNO.5Fin及Rin引物的具体核苷酸序列如下:SEQIDNO.6:ACAGTTCCTGGATATCCTAAGCT;SEQIDNO.7:ACAGCCTCTTCACGAGTAATG。相同的反应液设置8管,仅靶的添加有区别,分别为106Copies,105Copies,104Copies,103Copies,102Copies,101Copies,100Copies,纯水(0Copies)。设置ABIStepOnerealtimePCR反应温度恒定为63℃,反应时间为75min。不同靶浓度扩增体系的荧光强度随反应时间变化的曲线如图11所示。此结果同实施例6中的结果对比说明加入Fin寡核苷酸可明显减少扩增时间。当然,本发明还可以有多种实施例,在不背离本发明精神及其实质的情况下,熟悉本领域的技术人员可根据本发明的公开做出各种相应的改变和变形,但这些相应的改变和变形都应属于本发明所附的权利要求的保护范围。<110>中国科学院过程工程研究所<120>一种恒温条件下合成核酸的方法<160>7<210>1<211>36<212>DNA<213>人工序列<220><223>引物<400>1gtacgaagggcattacgctctcgtgttatttccagg36<210>2<211>36<212>DNA<213>人工序列<220><223>引物<400>2ggacctttattgtgctctcgcattacgggaagcatg36<210>3<211>19<212>DNA<213>人工序列<220><223>引物<400>3tacccgcaaatgtcccata19<210>4<211>18<212>DNA<213>人工序列<220><223>引物<400>4tgtagaggcacattggtg18<210>5<211>225<212>DNA<213>人工序列<220><223>MERS-orf1b<400>5aagacgagtgatgagctttgcgtgaatcttaatttacccgcaaatgtcccatactctcgt60gttatttccaggatgggctttaaactcgatgcaacagttcctggatatcctaagcttttc120attactcgtgaagaggctgtaaggcaagttcgaagctggataggcttcgatgttgagggt180gctcatgcttcccgtaatgcatgtggcaccaatgtgcctctacaa225<210>6<211>23<212>DNA<213>人工序列<220><223>引物<400>6acagttcctggatatcctaagct23<210>7<211>21<212>DNA<213>人工序列<220><223>引物<400>7acagcctcttcacgagtaatg21当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1