一种褐藻胶裂解酶及其应用的制作方法
本发明属于基因工程技术领域,具体涉及一株褐藻胶裂解酶基因及其应用。
背景技术:
我国拥有广阔的海域面积,其中蕴含着丰富的海洋生物资源,尤其是海藻资源。褐藻主要包括海带、裙带等,其细胞壁中富含的褐藻胶是一种直链酸性多糖,在天然状态下,褐藻胶的主要存在状态为水溶性的褐藻酸钠(sodiumalginate)、褐藻酸钾等褐藻酸盐类和水不溶性的褐藻酸(alginicacid)。目前市场上的褐藻酸钠(商品名为海藻酸钠)或其他褐藻酸盐主要是从褐藻中获得。研究发现降解褐藻酸钠所得到的寡糖具有多种生物活性,比如免疫调节、促生长、诱导植物抗性和提高蛋白质稳定性等,因而可广泛应用于化工、农业、食品、饲料添加和医药等领域(刘航,天然产物研究与开发,2012,24:201-204)。褐藻酸钠可用多种方法降解,包括化学降解法、物理降解法和酶降解法。化学降解法以酸降解为主,但该方法降解条件难以控制,操作较复杂,耗时长。物理降解法包括辐射法和超声法等,一般与其他降解法一起使用,降解产物的极限分子质量为50kda左右,不易制得寡糖。而用褐藻胶裂解酶降解褐藻酸钠具有降解条件温和,得率高等优点,且因为酶的底物专一性,能为后续研究寡糖化学结构提供信息,故而褐藻胶裂解酶逐步成为优先降解褐藻酸钠的方法。另外,褐藻胶裂解酶还能应用于肺囊肿性纤维化症的治疗、海藻饲料加工及海藻遗传工程等领域的研究(wongtyetal.annualreviewofmicrobiology,2000,54:289-340)。
褐藻酸裂解酶来源于海洋动植物和多种微生物(包括海洋细菌、土壤细菌和真菌)。褐藻酸钠裂解酶按其底物专一性可分为两大类:1,4-d-甘露糖醛酸片段裂解酶(ec4.2.2.3)和1,4-l-古罗糖醛酸片段裂解酶(ec4.2.2.11)。至今,褐藻胶裂解酶的生产大多依赖原始产酶动植物或微生物来获得酶蛋白,此种方法虽然能有效的获得一定量的酶蛋白,但产量有限,成本较高,较难满足实际应用需求。随着生物技术的发展,提供了利用基因工程高效生产褐藻胶裂解酶的技术方法。dongeunkim等根据已知的相关功能基因的相似序列设计pcr引物,从streptomycessp.alg-5菌株中克隆到了一个褐藻胶裂解酶基因,并在escherichiacolibl21(de3)中成功表达(kimetal.marinebiotechnology.2009.11:10-16)。采用这一策略必须对相关基因序列有一定的了解才能设计pcr引物,并且找到的是某一类结构或功能相似蛋白质中的新分子,较难发现全新的基因。
随着高通量测序技术的不断发展,越来越多的产褐藻胶裂解酶的微生物基因组序列得以测定,这使得通过基因组挖掘技术来分析序列信息,对编码褐藻胶裂解酶的基因进行钓取变得简洁快速,badurah等对来源于海洋的一株细菌vibriosplendidus的基因组进行序列测定,通过分析发现该菌株中含有4个编码褐藻胶裂解酶的基因,对其进行了基因克隆和在大肠杆菌escherichiacolibl21(de3)中的重组表达(baduretalappl.environ.microbiol.2015.81:1865-1873);目前获得褐藻胶裂解酶大多数是专一性降解均聚甘露糖醛酸的褐藻胶裂解酶,少数是具有具有降解古洛糖醛酸活性的褐藻胶裂解酶,而具有广泛底物特异性的褐藻胶裂解酶则非常稀少,只有来源于pseudoalteromonassp.的褐藻胶裂解酶aly-sj02(李建伟,marinedrugs,2011,21:1374-80),来源于白蚁肠道的isoptericolahalotolerans的褐藻胶裂解酶alyih(窦文芳等,carbohydratepolymers,2013,98:1476-82)等具有广泛底物特异性的酶活力较低,而且稳定性差,其中alyih只是获得了纯化的酶,尚未获得其编码基因,不能进行重组表达和分子改造;而且大多数褐藻胶裂解酶活性较低。本发明涉及的重组褐藻胶裂解酶具有高活性和高稳定性的特点,其降解产物为低聚合度(主要是二糖、三糖和四糖)的褐藻胶寡糖,具有重大的工业化应用前景。
技术实现要素:
本发明要解决的技术问题是,提供一种新型褐藻胶裂解酶。
本发明还要解决的技术问题是,提供包含上述褐藻胶裂解酶的基因工程菌及其构建方法。
本发明最后要解决的技术问题是,提供上述褐藻胶裂解酶的应用。
为解决上述技术问题,本发明采用如下技术方案:
一种褐藻胶裂解酶基因,其核苷酸序列如seqidno.1所示。
一种褐藻胶裂解酶,其氨基酸序列如seqidno.2所示。
包含上述褐藻胶裂解酶的基因工程菌,该菌株中导入了褐藻胶裂解酶基因,所述的褐藻胶裂解酶基因的核苷酸序列如seqidno.1所示。
上述产褐藻胶裂解酶的基因工程菌的构建方法,其特征在于,包括如下步骤:
(1)将褐藻胶裂解酶基因克隆到质粒中,得到重组载体;
(2)将重组载体转化宿主菌,得到产褐藻胶裂解酶的基因工程菌。
步骤(1)中,所述的质粒为pet21a(+)。
步骤(2)中,所述的宿主菌为大肠杆菌dh5α。
上述褐藻胶裂解酶在裂解褐藻酸盐和褐藻多糖中的应用在本发明的保护范围之内。
上述褐藻胶裂解酶的基因工程菌在发酵产褐藻胶裂解酶中的应用在本发明的保护范围之内。
有益效果:
本发明的algl来源于海洋细菌vibriosp.nj-04,通过设计同源引物,获得编码褐藻胶裂解酶algl的dna序列,该基因编码区长1359bp,编码452个氨基酸,其中1-21个氨基酸为信号肽,理论分子量为50.19kda,属于多糖裂解酶7家族。大肠杆菌重组表达获得的algl,对于褐藻胶、polym和polyg均具有较高活性,属于双功能褐藻胶裂解酶。本发明的内切型褐藻胶裂解酶可广泛用于化工、农业、食品和饲料添加、医药及海藻遗传工程等领域。
附图说明
图1:褐藻胶裂解酶algl的蛋白质三维结构模型。
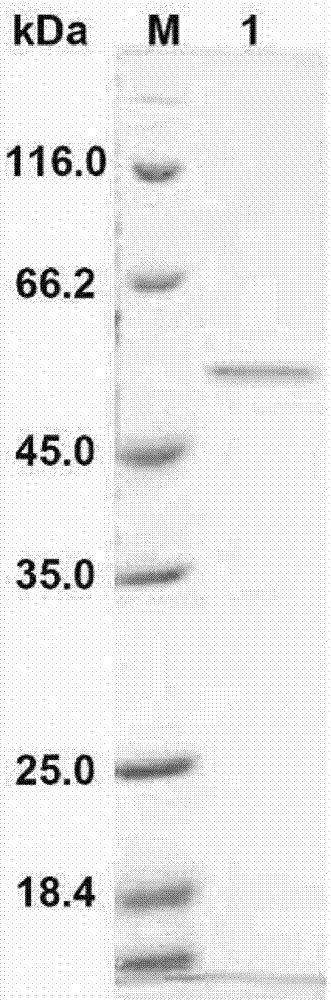
图2:重组褐藻胶裂解酶algl表达纯化的聚丙烯酰氨凝胶电泳图(sds-page)。
图3:温度、ph对于褐藻胶裂解酶algl的活性及稳定性影响曲线。
图4:褐藻胶裂解酶降解褐藻胶、polym和polyg所得产物的电喷雾质谱(esi-ms)分析图。
具体实施方式
根据下述实施例,可以更好地理解本发明。然而,本领域的技术人员容易理解,实施例所描述的内容仅用于说明本发明,而不应当也不会限制权利要求书中所详细描述的本发明。
实施例1:菌株vibriosp.nj-04的培养及鉴定
所采用的菌株来源于采集自黄海海域的海藻样品,经分离得到一株产褐藻胶裂解酶的菌株nj-04,使用的选择性培养基的配方为:
牛肉膏5g、葡萄糖15g、酵母浸粉1.0g、nacl5.0g、mgso4·7h2o0.5g、cacl20.2g、kh2po41.0g、feso4·7h2o0.02g,ph值为7.0。固体培养基添加1.5%的琼脂。
将菌落接入100ml液体培养基中进行富集培养,30℃,150rpm培养36h。将获得的菌液进行系列稀释后涂布到固体选择培养上取4ml已经培养好的菌液,置于2mltube管中,5000g,10min充分离心。弃去上清,用180μl消化液(digestionsolution)将菌体重悬,再加入20μl蛋白酶k,充分混匀后,56℃温育30min。加入20μlrnasea,充分混匀后,室温放置10min。加入200μl裂解液(lysissolution),快速混匀,过程不要超过15s。加入400μl50%(v/v)乙醇溶液,充分混匀。将上述溶液转移至吸附柱中,静置1min,离心6000g,1min,弃废液,将吸附柱转移至一个新的2mltube管中。加入500μlwashbufferi,8000g,1min离心,弃废液。加入500μlwashbufferii,8000g,1min离心,弃废液。重复一次。将空吸附柱12000g,3min离心,将柱内残留液体充分甩干。在吸附柱基质膜中间位置加入200μl洗脱液(elutionbuffer)静置1min,8000g,1min离心,即得基因组。
以提取的菌株基因组为模板,以细菌鉴定通用引物27f和1492r为引物,进行pcr扩增得到16srdna基因全长序列。pcr条件为:94℃预变性3min,随后以94℃30s、55℃30s、72℃2min进行30个循环,最后在72℃延伸10min。琼脂糖凝胶电泳显示在1.0kb和2.0kb之间有一条特异性条带,将其从琼脂糖凝胶上切下,采用dna凝胶回收试剂盒进行纯化后,送南京金斯瑞生物技术有限公司进行序列测定,序列见附录。
实施例2:褐藻胶裂解酶algl编码基因的克隆与鉴定
根据pl7家族褐藻胶裂解酶的保守区序列设计如下扩增引物:上游引物(5’-ggntcnaaywanaaraayta-3’)和下游引物(5’-ggnagyaaywanaaraayta’)。以提取的菌株基因组为模板,进行pcr扩增得到褐藻胶裂解酶algl基因部分序列。
pcr条件为:94℃预变性3min,随后以94℃30s、55℃30s、72℃2min进行30个循环,最后在72℃延伸10min。琼脂糖凝胶电泳显示在1.0kb左右一条特异性条带,将其从琼脂糖凝胶上切下,采用dna凝胶回收试剂盒进行纯化。
将纯化的dna片段连接到克隆载体peasy-bluntzerocloningvector上,转化大肠杆菌dh5α感受态细胞中,在lb固体培养基(含有氨苄青霉素)中培养后,挑取白色菌落使用扩增引物进行pcr验证。pcr条件为94℃预变性3min,随后以94℃30s、55℃30s、72℃2min进行30个循环,最后在72℃延伸10min。测序所得基因序列设计巢式pcr引物来按照上述条件来克隆褐藻胶裂解酶基因的全长。将出现特异性条带所对应的重组菌株大量培养,使用质粒提取试剂盒进行质粒抽提,进行测序分析。结果表明褐藻胶裂解酶基因的全核苷酸序列全长1359bp,核苷酸序列如seqidno.1所示;编码452个氨基酸,氨基酸序列如seqidno.2所示,蛋白理论分子量为50.19kda。用phyre2同源建模服务器对褐藻胶裂解酶algl的蛋白质三维结构进行同源建模。最终得到的algl的蛋白质三维结构模型如图1所示。
seqidno.1:algl编码基因开放阅读框(orf)全长1359bp,编码褐藻胶裂解酶的成熟酶序列,不含有信号肽序列。
seqidno.2:褐藻胶裂解酶algl结构中包含一个褐藻胶裂解酶家族2(alginatelyase2family)结构域。
实施例3:褐藻胶裂解酶algl重组表达载体构建。
根据褐藻胶裂解酶基因全序列设计上游引物(5’-gccatatgatgttattgaacaaaattattac-3’)和下游引物(5’-ggcctcgaggtaagaataatttgaatgtt-3’),进行pcr扩增得到褐藻胶裂解酶algl基因全长序列。pcr条件为:94℃预变性3min,随后以94℃30s、55℃30s、72℃2min进行30个循环,最后在72℃延伸10min。pcr产物用ndei和xhoi进行酶切后回收;将大肠杆菌表达载体pet21a(+)同样用ndei和xhoi进行酶切后回收,将其与上述所得目的基因连接后转化大肠杆菌dh5α中,筛选具有氨苄青霉素抗性的转化子。用质粒提取试剂盒抽提阳性转化子质粒,使用ndei和xhoi进行酶切验证,得到长度分别为5.5kb和1.5kb左右两个片段,证明褐藻胶裂解酶基因已经克隆到表达载体pet21a(+)上,将此重组质粒命名为pet21a-algl。
实施例4:褐藻胶裂解酶algl基因在大肠杆菌中的重组表达与纯化制备。
将重组表达质粒pet21a-algl转化大肠杆菌菌株bl21(de3)(购自美国novagen公司),然后按照该公司提供的操作步骤进行褐藻胶裂解酶algl诱导表达及纯化。用聚丙烯酰胺凝胶电泳检测褐藻胶裂解酶algl的纯化情况,结果如图2所示,纯化后的algl在电泳胶上呈现单一条带,且位置与预测的分子量相吻合。
实施例5:褐藻胶裂解酶algl的酶学性质分析。
1、ph和温度对酶活性的影响
将质量浓度为1%褐藻酸钠底物、纯化的algl酶液以及不同ph值的20mmtris-hcl、磷酸盐、醋酸盐缓冲液(ph范围为2.0-11.0)按9:1:10(体积比)的比例混合后,在30℃反应10分钟,紫外吸收法测酶活力。结果显示algl在ph8.0时达到最大活力,表明algl的最适反应ph为4.0。在最适ph下,将质量浓度为1%褐藻酸钠底物、纯化的algl酶液以及20mmtris-hcl缓冲液(ph7.0)按9:1:10(体积比)的比例混合,分别在不同温度(20℃-70℃)反应10分钟,按紫外吸收法测酶活力。结果显示algl在30℃时达到最大活力,表明algl的最适反应温度为30℃(如图3-a,c)。
2、ph和温度对酶稳定性的影响
将在不同温度(20℃-80℃)下热处理30min后的algl酶液与质量浓度为1%褐藻酸钠底物溶液按1:9(体积比)的比例混合,然后在最适温度和最适ph下测定剩余酶活,以不经过热处理的酶液酶活定义为100%相对活力(relativieactivity),结果表明algl在低于40℃的温度下具有较好的热稳定性;将在30℃,不同的ph(ph4-10)预孵育24h后的algl酶液与质量浓度为1%褐藻酸钠底物溶液按1:9(体积比)的比例混合,然后在最适温度和最适ph下测定剩余酶活,以不经过ph处理的酶液酶活定义为100%相对活力(relativieactivity),结果显示在ph6~10的范围内,algl酶活仍保持70%以上,表明algl对ph值耐受范围较广(如图3-b,d)。
3、algl的酶活性及动力学
将纯化的algl分别与质量浓度为0.1%的褐藻酸钠、聚甘露糖醛酸片段(polym)和聚古洛糖醛酸片段(polyg)三种不同底物按1:9(体积比)的比例混合,然后在30℃,ph7.0条件下反应。取不同反应时间点的产物测其235nm紫外吸收值,发现随着酶解时间延长,235nm吸收值逐渐增加,根据紫外法测褐藻胶裂解酶的原理(qing-daanetal.processbiochemistry,2008,43:842–847),证实algl是裂解酶。另外,如图4所示,algl对聚古洛糖醛酸片段、褐藻酸钠和聚甘露糖醛酸片段的降解均具有较好的降解活性,表明algl是一个双功能褐藻胶裂解酶。同时,该酶对于三种不同底物均具有较好的底物亲和力。
表1.褐藻胶裂解酶algl的底物特异性及酶动力学
实施例6:金属离子对algl活性的影响
将质量浓度为1%褐藻酸钠底物、纯化的algl酶液以及50mmtris-hcl缓冲液(ph7.0)按8:2:10(体积比)的比例混合,接着向反应体系中添加不同的金属离子,添加的离子终浓度为1mm和5mm,然后在30℃反应10分钟,按前述的紫外吸收法测酶活力。对照组为不加任何金属离子时algl的活性(设定为100%),结果如下表所示。实验结果显示,mg2+、co2+能增加algl活性,cu2+、zn2+等其他离子对酶活呈现抑制作用。
表2.金属离子对于algl酶活性的影响
实施例7:褐藻胶裂解酶algl对三种底物的降解产物的esi-ms分析。
将质量浓度为0.1%底物(褐藻酸钠、polym和polyg)、纯化的algl酶液以及50mmtris-hcl缓冲液按9:1:10(体积比)的比例混合后,在ph7.0,30℃条件下反应,对酶解72小时后的产物进行电喷雾质谱分析。电喷雾质谱条件为采用正离子模式,离子源电压:4.5kv;鞘气流速:30arb;毛细管温度:275~300℃;管镜电压:250v;扫描范围:150-2000。如图4所示。
序列表
<110>五洲丰农业科技有限公司
<120>一种褐藻胶裂解酶及其应用
<130>sg20160919001
<160>2
<170>patentinversion3.5
<210>1
<211>903
<212>dna
<213>弧菌(vibriosp.nj-04)褐藻胶裂解酶algl核苷酸序列
<400>1
1atgttattgaacaaaattattactacatctgccatcgccatttttagttcattttcaatggccgatacactgccgatcctagaagcaacggattggggga
tacaataacttgttttaataatgatgtagacggtagcggtaaaaatcaagtaaaagttaccggctatgtgacggctaggatcttcgttgcctaaccccct
101gttcgcatacaagttatcctgcctcaaacgcgattgatggtgatactgcttggagttctcgttgggccgctcatgattcaggttcagaagtgaacttagt
caagcgtatgttcaataggacggagtttgcgctaactaccactatgacgaacctcaagagcaacccggcgagtactaagtccaagtcttcacttgaatca
201gttggatctaggatcgatacaaaccgtatctgcagtaaatattgcttggggacgtggcaatgagagcacgaataaatttgaaattagagcccgttcagat
caacctagatcctagctatgtttggcatagacgtcatttataacgaacccctgcaccgttactctcgtgcttatttaaactttaatctcgggcaagtcta
301gccagtaatactacttgggataaaatttactctggtgatagtcaaggtaacacgacggatttcgaaagttatgatgttcaagatattaaagctcgttgga
cggtcattatgatgaaccctattttaaatgagaccactatcagttccattgtgctgcctaaagctttcaatactacaagttctataatttcgagcaacct
401ttcgaattaaagtattcagtaacagctcaggaagtgactggaccaatattaccgaagtcaatattcttggtagtaacaagaacaattatgggctagatgc
aagcttaatttcataagtcattgtcgagtccttcactgacctggttataatggcttcagttataagaaccatcattgttcttgttaatacccgatctacg
501gtcaaagcctccttcctataacttcgatttacttgactggtacctaagtattccagtcgacgagggtgatgggtatgccagctcgatcaaagaaaatgct
cagtttcggaggaaggatattgaagctaaatgaactgaccatggattcataaggtcagctgctcccactacccatacggtcgagctagtttcttttacga
601ttatctgcgagttatgaaagcgaatttttttatactggccaggatgggggaatggttttttatacaccagtaaaaggcgttaagacatcggaaaatacca
aatagacgctcaatactttcgcttaaaaaaatatgaccggtcctacccccttaccaaaaaatatgtggtcattttccgcaattctgtagccttttatggt
701agtatgtaagaaccgaactgcgagagatgttaagaagaggcgacacatcaaagtcaaccagtggtaaagaaaataactgggcattttcgagtatacccaa
tcatacattcttggcttgacgctctctacaattcttctccgctgtgtagtttcagttggtcaccatttcttttattgacccgtaaaagctcatatgggtt
801tgagacacaatctgattttggcggtattgatggtcatttaaaagcgactttagcggttaaccatgtcacaacaacgacgagtgattcgaaacaagttgga
actctgtgttagactaaaaccgccataactaccagtaaattttcgctgaaatcgccaattggtacagtgttgttgctgctcactaagctttgttcaacct
901agaatcgtgatcggtcagattcatgcaaaaaacaatgaacctattcgtttgtattatcacaaattacctgagaatgaaaaaggtgctatctattttgcac
tcttagcactagccagtctaagtacgttttttgttacttggataagcaaacataatagtgtttaatggactcttactttttccacgatagataaaacgtg
1001atgaatcatcgaaagcctctggcggtaatgaatcttggtataacttgcttggtgaaatggtcacttcggaaggtgagcttaatagtacgtctaatccgtc
tacttagtagctttcggagaccgccattacttagaaccatattgaacgaaccactttaccagtgaagccttccactcgaattatcatgcagattaggcag
1101aaaaggggtggcattaaatgagcatttcacgtatgaaattgatgtagttgctgattcacttaccgtaactcttcgccaaaatgatattgaattggcgaag
ttttccccaccgtaatttactcgtaaagtgcatactttaactacatcaacgactaagtgaatggcattgagaagcggttttactataacttaaccgcttc
1201aaaaccgtcgatatgtcagacagtggctatgatgatggtgacaattatatgtatttcaaggctggaatttatttacaagacaatacaagtgaagaggatg
ttttggcagctatacagtctgtcaccgatactactaccactgttaatatacataaagttccgaccttaaataaatgttctgttatgttcacttctcctac
1301attatgcacaagtcactttttatgagttggtaaacgaacattcaaattattcttactaa
taatacgtgttcagtgaaaaatactcaaccatttgcttgtaagtttaataagaatgatt
<210>2
<211>300
<212>prt
<213>褐藻胶裂解酶氨基酸序列
<400>2
1metleuleuasnlysileilethrthrseralailealailepheserserphesermet
21alaaspthrleuproileleuglualathrasptrpglyserserhisthrsertyrpro
41alaserasnalaileaspglyaspthralatrpserserargtrpalaalahisaspser
61glysergluvalasnleuvalleuaspleuglyserileglnthrvalseralavalasn
81ilealatrpglyargglyasngluserthrasnlysphegluileargalaargserasp
101alaserasnthrthrtrpasplysiletyrserglyaspserglnglyasnthrthrasp
121pheglusertyraspvalglnaspilelysalaargtrpileargilelysvalpheser
141asnserserglyserasptrpthrasnilethrgluvalasnileleuglyserasnlys
161asnasntyrglyleuaspalaserlysproprosertyrasnpheaspleuleuasptrp
181tyrleuserileprovalaspgluglyaspglytyralaserserilelysgluasnala
201leuseralasertyrglusergluphephetyrthrglyglnaspglyglymetvalphe
221tyrthrprovallysglyvallysthrsergluasnthrlystyrvalargthrgluleu
241argglumetleuargargglyaspthrserlysserthrserglylysgluasnasntrp
261alapheserserileproasngluthrglnserasppheglyglyileaspglyhisleu
281lysalathrleualavalasnhisvalthrthrthrthrseraspserlysglnvalgly
301argilevalileglyglnilehisalalysasnasngluproileargleutyrtyrhis
321lysleuprogluasnglulysglyalailetyrphealahisgluserserlysalaser
341glyglyasnglusertrptyrasnleuleuglyglumetvalthrsergluglygluleu
361asnserthrserasnproserlysglyvalalaleuasngluhisphethrtyrgluile
381aspvalvalalaaspserleuthrvalthrleuargglnasnaspilegluleualalys
401lysthrvalaspmetseraspserglytyraspaspglyaspasntyrmettyrphelys
421alaglyiletyrleuglnaspasnthrserglugluaspasptyralaglnvalthrphe
441tyrgluleuvalasngluhisserasntyrsertyr***
- 还没有人留言评论。精彩留言会获得点赞!