原种事件EE-GM5及在生物样品中用于鉴定该事件的方法和试剂盒与流程
本申请要求于2016年12月22日提交的美国临时申请序列号62/437,874和于2017年4月4日提交的美国临时申请序列号62/481,284的优先权,这些文件的内容通过引用以其全部内容结合在此。
本发明涉及新颖的核酸和转基因大豆植物、植物材料、和种子,其特征在于在大豆基因组的特定位置处携带特定的转化事件,特别是赋予线虫抗性和除草剂耐受性的基因的存在。本发明的大豆植物将线虫抗性和除草剂耐受性表型与不同遗传背景中的农艺性状、遗传稳定性和功能性相结合,其中在无(一种或多种)hppd抑制剂型除草剂或线虫侵染情况下所述的遗传背景与相应的非转化大豆遗传背景等同。本发明进一步提供了用于鉴定生物样品中植物材料存在的方法和试剂盒,其中所述植物材料特别地包含转化事件ee-gm5。
背景技术:
植物中转基因的表型表达由一个基因或多个基因本身的结构以及它或它们在植物基因组中的位置决定。同时,转基因或“插入的t-dna”在基因组中不同位置处的存在将以不同方式影响植物的整体表型。通过遗传操作在植物中农业或工业成功地引入商业上的目的性状,取决于不同因素,可以是一个漫长过程。基因转化植物的实际转化和再生只是一系列选择步骤中的第一步,这些步骤包括广泛的遗传表征、基因渗入和田间试验评估,最终导致选择原种事件。
鉴于新食品/饲料上的讨论、gmo和非gmo产品的分离、以及专利材料的鉴定,对原种事件的明确鉴定变得越来越重要。理想地,鉴定方法应既快速又简单,无需广泛的实验室设置。此外,该方法应该提供可以允许在没有专家解释的情况下对原种事件实现明确鉴定的结果,但是如果必要的话,该结果也经得起专家审查。
种植线虫抗性和除草剂耐受性大豆ee-gm5品种为种植者提供了使用hppd抑制剂型除草剂(例如,异噁唑草酮(ift)、苯唑草酮、或硝磺草酮(mst)除草剂)用于线虫和杂草控制的新选择。hppd抑制剂型除草剂为大豆种植者提供了一种可替代的杂草控制选择,以帮助管理问题杂草物种,并作为替代性作用模式工具,有助于减缓除草剂抗性杂草的蔓延。
大豆胞囊线虫(scn,heteroderaglycines(ichinohe)是大豆生产的全球性问题,对生产者持续构成威胁。自1954年在美国北卡罗来纳州的一个县首次发现以来,scn已经蔓延到美国几乎所有的大豆生产州,并估计导致美国的年产量损失超过12亿美元,成为当地最具破坏性的大豆病原体。scn于90年代初首次在巴西被发现,目前已经遍布南美洲,并且在巴西是最重要的病原体之一,导致实际上所有巴西种植区域的损失。类似地,scn持续在中国大豆产区蔓延,其中在15个省份中检测到,产量损失估计超过1.2亿美元。在美国爱荷华州(2001至2015年)进行的一项为期多年的研究,其中几乎所有的scn抗性大豆品种都含有来自pi88788的scn抗性,该研究发现scn群体的毒力逐年增加,导致季末scn群体密度的增加和具有pi88788来源抗性的scn抗性大豆品种产量的降低(mitchum(2016),phytopathology[植物病理学]106(12):1444-1450,allen等人.(2017)planthealthprogr.[植物健康进展]18:19-27,arias等人.(2017)www.researchgate.net/publication/266907703_resistance_to_soybean_cyst_nematode_genetics_and_breeding_in_brazil;mccarville等人.(2017)planthealthprogress[植物健康进展]18:146-155)。
根腐线虫最短尾短体线虫(pratylenchusbrachyurus)已成为越来越重要的大豆病原体。它具有广泛的寄主范围,并且广泛分布于热带和亚热带地区,特别是巴西、非洲和美国南部。最短尾短体线虫已成为巴西塞拉多(cerrado)地区棉花和大豆种植者的顾虑,并且被认为是该地区大豆的主要线虫病原体。在大豆中,这种线虫可以使产量降低30%至50%,其中在砂土上观察到更大的损害。抗性大豆品种的使用将是控制该线虫的最佳方式,然而,迄今尚未鉴定出最短尾短体线虫抗性大豆品种。虽然已经研究了针对最短尾短体线虫抗性的若干大豆基因型,并且鉴定了具有耐受性增加的一些栽培品种,但由于这种线虫是杂食性并缺少与其宿主的密切相互作用,因此育种产生针对最短尾短体线虫的抗性栽培品种很难(machado(2014)currentagriculturalscienceandtechnology[当前农业科学与技术]20:26-35;antonio等人.(2012)soilproductivitylossesinareainfestedbythenematoidoftherootlesionsinvera,mt[在vera,mt中根病变的线虫侵染的区域中土壤生产力的损失].在:braziliancongressofsoy[巴西大豆大会],6,2012,cuiabá.摘要.londrina:embrapasoja,4pp;rios等人.(2016)ciênciarural46:580-584;lima等人.,2017,在书:soybean-thebasisofyield,biomassandproductivity[大豆——产量、生物量和生产力的基础]中第6章;由minobukasai编辑,isbn978-953-51-3118-2,printisbn978-953-51-3117-5,intech;inomoto等人.(2011)
众所周知,保护植物对抗线虫(例如scn)可以帮助植物更好地应对其他胁迫(例如,土壤组分/含量、天气条件、病原菌胁迫、除草剂施用等)。特别是当此类其他胁迫产生易于看到的表型时,例如叶子的萎黄/变黄,scn控制的效果更容易看到,而其他情况下常不“可见”。例如,当大豆植物患有猝死综合征(sds)或缺铁性萎黄病(idc)时,针对scn的保护将导致植物更绿或具有不太严重的sds/idc症状。尽管进行了广泛的研究和品种筛选工作,但在美国中北部的大型大豆生产区域,铁缺乏仍然是一个挑战。由于在易受铁缺乏影响的土壤上扩大的大豆生产以及与种植系统变化的可能的相互作用,这一问题的重要性已经增加。铁缺乏发生在具有高ph和碳酸盐的土壤中,但由于与空间可变土壤特性(例如,水分含量、盐度、铁以及其他微量营养素的可用性、和金属浓度)的相互作用,缺铁的表现在空间上变化很大。此外,缺铁表现与生物因子(例如固氮、害虫、疾病)和管理诱导的胁迫(例如,除草剂施用)相互作用。品种选择是管理铁缺乏的最重要的手段,但品种选择由于与萎黄病耐受性相关的环境相互作用以及大的基因型而复杂化(hansen等人.(2004)soilsci.plantnutr.[土壤科学与植物营养学]50(7):983-987)。
大豆猝死综合征(sds)最早于1971年在阿肯色州被发现,并且从那时起已在美国的大多数大豆种植区域得到证实。sds是真菌疾病,也发生在与大豆胞囊线虫(scn)复合的疾病中。sds是美国最具破坏性的大豆土传疾病。当这种疾病在scn存在的情况下发生时,症状发生得更早并且更严重。sds是由一组茄病镰刀菌(fusariumsolani)物种复合体中的土传真菌引起。在北美,fusariumvirguliforme(先前是,茄病镰刀菌甘氨酸专化型(fusariumsolanif.sp.glycines))是致病因子。在南美,巴西镰刀菌(f.brasiliense)、镰刀菌f.cuneirostrum、镰刀菌f.tucumaniae、和fusariumvirguliforme引起sds症状。尽管已经开发出对sds不太敏感的大豆栽培品种,但是没有高抗性栽培品种。这些真菌可以在种植后很快侵染大豆幼苗的根,但在大豆植物达到生殖阶段之前,很少出现sds的地上症状。真菌在根部产生毒素,转移到叶中。sds的第一个明显症状是上部叶片变黄和脱叶。如果该疾病在季节早期发展,花和幼荚将会中止。当该疾病在晚期发展时,植物每荚将产生较少的种子或较小的种子。严重疾病发展越早,产量减少得越多。因为sds真菌可以在土壤中长期存在,所以每个生长季节,田地的较大区域将显示疾病的症状,直到大部分田地受到影响(westphal等人.(2008).suddendeathsyndromeofsoybean[大豆猝死综合征].theplanthealthinstructor[植物健康指导].doi:10.1094/phi-i-2008-0102-01,www.apsnet.org/edcenter/intropp/lessons/fungi/ascomycetes/pages/suddendeath.aspx)。
目前,没有针对线虫抗性进行基因工程改造的大豆植物被商业化。本领域已经披露了包含一种或多种除草剂耐受性基因的大豆植物。wo2006/130436描述了包含epsps基因的草甘膦耐受性大豆事件,并且wo2011/034704描述了麦草畏耐受性大豆事件。wo2012/082548描述了包含hppd和pat基因的大豆植物。wo2011/063411描述了对hppd抑制剂和草甘膦具有耐受性的大豆事件,而wo2011/063413描述了对hppd抑制剂、草铵膦和草甘膦具有耐受性的大豆植物。wo2011/066384描述了对2,4-d和草铵膦具有耐受性的大豆事件,而wo2012/075426描述了对2,4-d、草铵膦和草甘膦具有耐受性的大豆事件,并且wo2017/059795描述了对草甘膦具有耐受性的大豆事件。wo2009/064652描述了对鳞翅目昆虫具有抗性的大豆事件,并且wo2013/016527描述了对鳞翅目昆虫具有抗性和具有草铵膦耐受性的大豆事件。
对hppd抑制剂型除草剂赋予改善的耐受性的hppd基因和蛋白质已经被披露于例如wo2015138394、wo2015135881、wo2014043435中,并且cry蛋白的杀线虫活性已经被披露于例如,wo2010027805、wo2010027809、wo2010027804、wo2010027799、wo2010027808和wo2007147029中。
现有技术没有教导或提示在包含线虫活性cry基因的大豆中的原种事件,并且当然没有教导或提示在包含线虫活性cry基因以及赋予hppd抑制剂耐受性的基因的大豆中的原种事件。
本领域已知在具有可接受的农艺性状的大豆植物中获得商业原种转化事件决非简单之事。
本发明优选实施方案的概述
本发明提供了编码cry14ab-1蛋白的核酸,例如seqidno.7的cry14ab-1.b编码序列、或与seqidno.7具有至少95%、96%、97%、98%、或至少99%序列同一性的编码杀线虫cry14ab蛋白的序列。本文还提供了编码hppd-4蛋白的核酸,例如seqidno.9的hppdpf-4pa编码序列、或与seqidno.9具有至少95%、96%、97%、98%、或至少99%序列同一性的序列,其中当在植物中表达时所述序列编码提供对hppd抑制剂型除草剂的耐受性的hppd蛋白。本文还提供了嵌合cry14ab-1.b基因,该基因包含seqidno.11的核苷酸位置131至核苷酸位置5276的序列或其互补序列;或嵌合cry14ab-1.b基因,该基因包含有效连接至植物可表达型启动子的、seqidno.11的核苷酸位置412至核苷酸位置3969的序列;或编码杀线虫cry14ab蛋白的序列,该序列与seqidno.11的核苷酸位置131至核苷酸位置5276的序列或其互补序列、或与seqidno.11的核苷酸位置412至核苷酸位置3969的序列(当有效连接至植物可表达型启动子时)具有至少95%、96%、97%、98%、或至少99%序列同一性。本文进一步提供了嵌合hppdpf-4pa基因,该基因包含seqidno.11的核苷酸位置5382至核苷酸位置7459的序列或其互补序列;或包含有效连接至植物可表达型启动子的、seqidno.11的核苷酸位置5589至核苷酸位置6665的序列;或与seqidno.11的核苷酸位置5382至核苷酸位置7459的序列或其互补序列、或与seqidno.11的核苷酸位置5589至核苷酸位置6665的序列(当有效连接至植物可表达型启动子时)具有至少95%、96%、97%、98%、或至少99%序列同一性的序列;其中所述序列编码hppd蛋白(当该蛋白在植物中表达时提供对hppd抑制剂型除草剂的耐受性);以及提供包含所述嵌合cry14ab-1.b基因和所述嵌合hppdpf-4pa基因的核酸。这些核酸或基因可用于转化植物(例如大豆、棉花、玉米、稻、油菜和小麦),从而使它们可以控制线虫和/或具有hppd抑制剂型除草剂耐受性。
本文还提供了嵌合dna分子,该嵌合dna分子包含seqidno.11的核苷酸位置131至核苷酸位置7941的核苷酸序列,或与其具有至少95%、至少96%、至少97%、至少98%、或至少99%序列同一性的核苷酸序列。在一个实施例中,该dna分子编码对hppd抑制剂具有耐受性的蛋白质和负面影响植物有害生物线虫(例如,scn、rkn或短体线虫属线虫)的蛋白质。在一个实施例中,该嵌合dna分子编码seqidno.8的蛋白或与其具有至少99%同一性的线虫控制蛋白、和seqidno.10的蛋白或与其具有至少99%同一性的hppd抑制剂耐受性蛋白。还提供了经转化而包含此类dna分子的植物、种子、或细胞(例如,大豆植物、种子、或细胞)以及此类dna分子用于赋予植物或种子(例如,大豆植物或种子)对线虫的抗性和对hppd抑制剂型除草剂的耐受性的用途。
本发明涉及包含稳定整合到其基因组中的表达盒的转基因大豆植物、植物部分、种子、其细胞或组织,该表达盒包含:含有cry14ab-1.b基因的编码序列的线虫抗性基因以及含有hppdpf-4pa基因的编码序列的除草剂耐受性基因(二者均如本文中实例1.1所述并分别如seqidno.7和9中所表示),这两种基因提供对植物寄生线虫(如大豆胞囊线虫)的抗性和对hppd抑制剂型除草剂(例如,异噁唑草酮、苯唑草酮、或硝磺草酮)的耐受性。在不存在hppd抑制剂型除草剂和线虫压力的情况下,这种大豆植物具有基本上等同于非转基因等基因系的农艺性状。当遇到影响田地中植物性能的大豆胞囊线虫(scn)压力时,与非转基因植物相比,本发明的植物将具有优异的农艺表型。此外,在杂草存在下,在施用耐受性所针对的hppd抑制剂型除草剂后,与未用除草剂处理的植物相比,本发明的植物将具有优异的农艺表型。
根据本发明,大豆植物或种子、其细胞或组织包含原种事件ee-gm5。在一个实施方案中,原种事件ee-gm5包含seqidno.1、3、5、或24中任一个的序列,或seqidno.2、4、6、或25中任一个的序列,或与其基本上相似的序列。在一个实施方案中,ee-gm5包含seqidno.1、3、5、或24中任一个的序列和seqidno.2、4、6、或25中任一个的序列,或与其基本上相似的序列;和seqidno.7的cry14ab-1.b编码序列和seqidno.9的hppdpf-4pa编码序列,或与其基本上相似的序列。在一个实施方式中,原种事件ee-gm5是在大豆基因组中在特定位置处插入的t-dna,如包含于以保藏号pta-123625保藏在atcc的参考种子中的原种事件。在一个实施方案中,ee-gm5中的t-dna包含嵌合cry14ab-1-编码基因和hppd-4-编码基因。在另一个实施方案中,所述事件的特征在于seqidno.1或3的5’连接序列,或seqidno.2或4的3’连接序列;或seqidno.1或3的5’连接序列和seqidno.2或4的3’连接序列。在一个实施方案中,当使用分别包含seqidno.12和seqidno.13的核苷酸序列的两种引物、通过聚合酶链式反应(本文中“pcr”)分析时,包含ee-gm5的基因组dna产生85bp的dna片段。在一个实施方案中,当使用分别包含seqidno.18和seqidno.19的核苷酸序列的两种引物、通过pcr分析时,包含ee-gm5的基因组dna产生84bp的dna片段。
在本文的一个实施方案中提供了在其基因组中包含原种事件ee-gm5的大豆植物、细胞、植物部分、种子或其子代,其中包含所述事件的参考种子在atcc以保藏号pta-123625保藏。在一个实施方案中,通过繁殖由在atcc以保藏号pta-123625保藏的种子生长的大豆植物、和/或用其育种,可获得包含ee-gm5的植物或种子。
更具体地,本发明涉及转基因大豆植物、植物部分、花粉、种子、其细胞或组织,其基因组dna的特征在于:当在如本文所述的pcr中分析时,使用至少两个引物,所述引物针对由ee-gm5的5’或3’t-dna侧翼区的部分和插入的t-dna的部分形成的区域,扩增对事件ee-gm5具有特异性的片段。这些引物可以针对在seqidno:6或seqidno.25中的3’t-dna侧翼区、或其下游并与其邻接的大豆植物基因组dna、以及其上游并与其邻接的插入的t-dna。这些引物还可以针对在seqidno:5或seqidno.24中的5’t-dna侧翼区、或其上游并与其邻接的大豆植物基因组dna、和其下游并与其邻接的插入的t-dna。在一个实施方案中,所述引物分别包含以下核苷酸序列或(基本上)由以下核苷酸序列组成:seqidno:12和seqidno:13的核苷酸序列、或seqidno.18和seqidno.19的核苷酸序列、或seqidno.26和seqidno.28的核苷酸序列、或seqidno.27和seqidno.29的核苷酸序列(例如,引物对包含:在其最3’端含有seqidno:12的核苷酸序列的引物;和在其最3’端含有seqidno:13的核苷酸序列的引物;或引物对包含:在其最3’端含有seqidno.18的核苷酸序列的引物和在其最3’端含有seqidno.19的核苷酸序列的引物;或引物对包含:在其最3’端含有seqidno:26的核苷酸序列的引物和在其最3’端含有seqidno:28的核苷酸序列的引物;或引物对包含:在其最3’端含有seqidno:27的核苷酸序列的引物和在其最3’端含有seqidno:29的核苷酸序列的引物),并且产生在50和1000bp之间的dna片段(例如85bp或84bp的片段)。
已经将包含本发明的原种事件的参考种子在atcc以保藏号pta-123625保藏。本发明的一个实施方案是如包含在以保藏号pta-123625保藏的种子中的原种事件ee-gm5,当将该原种事件ee-gm5引入大豆植物时将提供对线虫的抗性和对除草剂的耐受性,特别是对大豆胞囊线虫(heteroderaglycines,本文中“scn”)和/或腐线虫(如本文所用的腐线虫是指短体线虫属物种的大豆有害线虫,包括但不限于最短尾短体线虫)的抗性以及对hppd抑制剂(例如,异噁唑草酮、苯唑草酮、或硝磺草酮)的耐受性。本发明的具有ee-gm5的植物还控制根结线虫(如本文所用的根结线虫是指根结线虫属物种(meloidogynespp.)的大豆有害生物线虫,包括但不限于南方根结线虫(meloidogyneincognita)、花生根结线虫(meloidogynearenaria)、北方根结线虫(meloidogynehapla)、或爪哇根结线虫(meloidogynejavanica)或其任何组合)、肾形线虫(肾状肾形线虫(rotylenchulusreniformis))和矛线虫(lancenematode)(纽带线虫属物种(hoplolaimusspp.),例如哥伦布纽带线虫(h.columbus)、帽状纽带线虫(h.galeatus)、和h.magnistylus)。该事件的轻微变体包括在本发明中,例如:具有hppd抑制剂耐受性和scn线虫抗性且具有如下核苷酸序列的大豆事件,其中所述核苷酸序列与在atcc保藏号pta-123625保藏的种子中所包含的ee-gm5的核苷酸序列具有至少90%、至少95%、至少98%、至少99%、至少99,5%、或至少99,9%序列同一性;或具有hppd抑制剂耐受性和scn线虫抗性且具有如下核苷酸序列的大豆事件,其中所述核苷酸序列与在atcc保藏号pta-123625的保藏种子中包含的ee-gm5的核苷酸序列相差1至200、1至150、1至100、1至75、1至50、1至30、1至20、1至10、或1至5个核苷酸;或与以下连续核苷酸序列(5’至3’)所形成的核苷酸序列相差1至200、1至150、1至100、1至75、1至50、1至30、1至20、1至10、或1至5个核苷酸:seqidno.5或seqidno.24、seqidno.11的核苷酸位置188至核苷酸位置7101、和seqidno.6或seqidno.25。在一个实施方案中,ee-gm5包含核苷酸序列,所述核苷酸序列与以下连续核苷酸序列(5’至3’)所形成的序列具有至少95%、至少96%、至少97%、至少98%、至少99%、至少99.5%、或至少99.9%序列同一性:seqidno.5或24、seqidno.11的核苷酸位置188至核苷酸位置7101、和seqidno.6或25。由于天然的遗传变异,在相同物种的植物中,同源dna序列中的dna单碱基差异和小的插入和缺失(例如,单核苷酸多态性(snp))是常见的(zhu等人.(2003)genetics[遗传学]163:1123-1134)。
atcc保藏号pta-123625的种子是本发明原种事件ee-gm5纯合的转基因种子的纯种子批,该种子将生长为线虫抗性植物,从而使这些植物还对hppd抑制剂(例如,异噁唑草酮、苯唑草酮、或硝磺草酮)具有耐受性。可以播种可从保藏种子获得的种子或子代种子(例如,在与具有不同遗传背景的其他大豆植物杂交后),并且可以用如本文所述的hppd抑制剂(例如,异噁唑草酮、苯唑草酮、或硝磺草酮)处理生长的植物,或可以测试如本文所述的ee-gm5的存在,以获得包含本发明原种事件的植物。本发明进一步涉及来自植物的细胞、种子、组织、子代、和后代,其中所述植物包含本发明原种事件,并从atcc保藏的保藏号pta-123625种子生长而成。本发明进一步涉及,可从包含本发明的原种事件的大豆植物(例如,由在atcc保藏的保藏号pta-123625种子生长的植物;或包含位于seqidno.1、3或5的序列和seqidno.2、4或6的序列之间的seqidno.9的hppd-4编码序列和seqidno.7的cry14ab-1.b编码序列的植物;或包含位于seqidno.1、3、5、或24的任一序列和seqidno.2、4、6、或25的任一序列之间的seqidno.9的hppdpf-4pa编码序列和seqidno.7的cry14ab-1.b编码序列的植物)可获得的(例如通过繁殖和/或育种)植物。本发明还涉及可获得自上述植物或种子的子代植物和种子,并且这些子代植物和种子包含以下序列:seqidno.1的序列和seqidno.2的序列、或seqidno.3的序列和seqidno.4的序列、或seqidno.5的序列和seqidno.6的序列、或seqidno.24的序列和seqidno.25的序列。
本发明进一步涉及用于鉴定包含原种事件ee-gm5的转基因植物或其细胞或组织的方法,该方法基于在转基因植物、细胞或组织中鉴定表征性dna序列或由此类dna序列编码的氨基酸的存在。根据本发明的优选的实施方案,此类表征性dna序列是允许特异性鉴定原种事件的15bp或至少15bp,优选地20bp或至少20bp,最优选30bp或以上的序列,所述序列包含事件的插入位点,即序列包含含有hppd抑制剂和线虫抗性转基因的插入t-dna的部分和与其邻接并延伸到大豆植物基因组中的5’或3’t-dna侧翼区的部分。本发明还涉及如本文所鉴定的包含事件ee-gm5的植物、种子、和细胞。
本发明进一步涉及用于在生物样品中鉴定原种事件ee-gm5的方法,该方法基于特异性识别ee-gm5中的5’和/或3’t-dna侧翼序列和与其邻接的插入t-dna序列的引物对或探针。本文还包括鉴定ee-gm5的任何其他方法,例如鉴定其特异性表征性序列的方法,例如全基因组或部分(定向)基因组测序。
更特别地,本发明涉及用于在生物样品中鉴定原种事件ee-gm5的方法,该方法包括使用具有至少两个引物的聚合酶链式反应或具有至少两个引物和探针的聚合酶链式反应,扩增存在于所述生物样品中的核酸序列,其中这些引物之一识别ee-gm5中的5’或3’t-dna侧翼区,并且另一引物识别包含除草剂耐受性和线虫抗性基因的t-dna内邻接所述5’或3’t-dna侧翼区的序列,优选地以获得50至1000bp大小的dna片段。在一个实施方案中,第一引物识别ee-gm5中的5’t-dna侧翼区,并且第二引物识别在包含除草剂耐受性和线虫抗性基因的t-dna内邻接所述5’t-dna侧翼区并在其下游的序列;或第一引物识别ee-gm5中的3’t-dna侧翼区,并且第二引物识别在包含除草剂耐受性和线虫抗性基因的t-dna内邻接所述3’t-dna侧翼区并在其上游的序列,从而获得原种事件ee-gm5特征性的dna片段。在一个实施方案中,所述聚合酶链式反应方法进一步包括使用探针,所述探针识别由所述引物扩增的dna,例如连接dna,所述连接dna包含ee-gm5中的插入t-dna的部分和位于所述t-dna侧翼的dna的部分(酌情在事件的5’或3’侧,例如包含本文seqidno.14或20的核苷酸序列或其互补序列的探针),以检测由所述引物产生的扩增产物。这些引物可以分别识别在ee-gm5的5’t-dna侧翼区(seqidno.5,核苷酸位置1至核苷酸位置166,或seqidno.24的核苷酸位置1至核苷酸位置1113)内或在ee-gm5的3’t-dna侧翼区(seqidno.6的核苷酸位置359至核苷酸位置691的互补序列,或seqidno.25的核苷酸位置359至核苷酸位置1449)内的序列以及在插入t-dna(seqidno.5的核苷酸位置167至353、或seqidno.6的核苷酸位置1至核苷酸位置358、或seqidno.23的核苷酸位置1114至8572,或其互补序列)内的序列。识别5’或3’t-dna侧翼区的引物可以包含seqidno.13、seqidno.19、seqidno.26或seqidno.27的核苷酸序列,并且识别在插入的t-dna内的序列(包含线虫抗性和除草剂耐受性基因)的引物可以包含本文所述的seqidno.12、seqidno.18、seqidno.28或seqidno.29的核苷酸序列。本发明还涉及任何事件特异性引物对、以及使用该引物对从ee-gm5事件序列扩增的特异性dna,其中所述引物对可以由本领域普通技术人员获得,或可以从商业来源获得,其中所述ee-gm5事件序列是本文中提供的序列,或者是包含在atcc的保藏号pta-123625保藏种子中的序列。
本发明更具体地涉及用于在生物样品中鉴定原种事件ee-gm5的方法,该方法包括使用两个引物和聚合酶链式反应扩增存在于生物样品中的核酸的序列,其中两个引物分别包含seqidno.12和seqidno.13的核苷酸序列或(基本上)由其组成,以扩增获得85bp的dna片段;或其中两个引物分别包含seqidno.18和seqidno.19的核苷酸序列或(基本上)由其组成,以扩增获得84bp的dna片段。包含如此鉴定的原种事件ee-gm5的植物也包括在本发明中。
本发明进一步涉及本文所述的ee-gm5的特异性t-dna侧翼序列,可以将该序列用于开发在生物样品特异性鉴定ee-gm5的方法。此类特异性t-dna侧翼序列还可以在鉴定试验中用作参照对照材料。更具体地,本发明涉及ee-gm5的5’和/或3’t-dna侧翼区,该侧翼区可以被用于开发如本文进一步描述的特异性引物和探针。还适合作为参考材料的是核酸分子,优选约150-850bp,所述核酸包含可以通过如下引物扩增的序列,所述引物包含seqidno.12和seqidno.13的核苷酸序列或seqidno.18和seqidno.19的核苷酸序列,或(基本上)由其组成。
本发明进一步涉及基于使用此类特异性引物或探针,鉴定生物样品中ee-gm5存在的方法。引物可以包含选自以下序列的17至约200个连续核苷酸的核苷酸序列,由其组成或基本上由其组成:seqidno.5的核苷酸1至核苷酸166或seqidno.24的核苷酸1至核苷酸1113的核苷酸序列、或seqid6的核苷酸359至核苷酸691的核苷酸序列的互补序列、或seqidno.25的核苷酸359至核苷酸1449的核苷酸序列的互补序列;并与如下引物组合,所述引物包含选自以下序列的17至约200个连续核苷酸的核苷酸序列,或由其组成或基本上由其组成:seqidno.11的核苷酸1至核苷酸7459或seqidno.23的核苷酸位置1114至核苷酸位置8572的核苷酸序列,例如选自以下序列的17至约200个连续核苷酸的核苷酸序列:seqidno.5的核苷酸167至核苷酸353的核苷酸序列或seqidno.6的核苷酸1至核苷酸358的核苷酸序列,或其互补序列。引物还可以包含位于它们的最3’末端处的这些核苷酸序列,并且进一步包含不相关的序列或衍生自所述核苷酸序列但包含错配的序列。在一个实施方案中,如本文所用的这些引物还可以与靶dna或其互补序列相同,其中所述靶dna是含有来自不同来源的核苷酸序列的杂合体,这些核苷酸序列在自然中不以这种组合形式存在。
本发明进一步涉及用于在生物样品中鉴定原种事件ee-gm5的试剂盒,所述试剂盒包含至少一个引物对或探针,其在ee-gm5中特异性识别5’或3’t-dna侧翼区以及与其邻接的包含除草剂耐受性和线虫抗性基因的插入t-dna。
除了特异性识别ee-gm5的5’或3’t-dna侧翼区的引物,本发明的试剂盒还可以包含特异性识别插入t-dna内的序列的第二引物,用于在pcr鉴定方案中使用,其中所述插入t-dna包含ee-gm5的hppd抑制剂除草剂耐受性和线虫抗性基因。本发明试剂盒可以包含至少两个特异性引物,其中一个引物识别在ee-gm5的5’t-dna侧翼区的序列或在ee-gm5的3’t-dna侧翼区的序列,并且另一个引物识别在包含hppd抑制剂除草剂耐受性和线虫抗性基因的插入t-dna内的序列。识别5’t-dna侧翼区的引物可以包含seqidno.19的核苷酸序列,且识别邻接所述5’t-dna侧翼区的插入t-dna的引物可以包含seqidno.18的核苷酸序列;或识别3’t-dna侧翼区的引物可以包含seqidno.13的核苷酸序列,且识别邻接所述3’侧翼区的插入t-dna的引物可以包含seqidno.12的核苷酸序列;或如本文所述的或可从说明书或种子保藏物中可获得的任何其他引物或引物组合。该试剂盒可以进一步包含探针,该探针识别位于识别5’t-dna侧翼区的引物和识别插入t-dna内的序列的引物之间的序列,或识别位于识别3’t-dna侧翼区的引物和识别插入t-dna内序列的引物之间的序列,例如包含seqidno.14的序列的探针或包含seqidno.20的序列的探针。
本发明进一步涉及用于鉴定生物样品中原种事件ee-gm5的试剂盒,所述试剂盒包含pcr引物,这些引物包含seqidno.12和seqidno.13的核苷酸序列,或seqidno.18和seqidno.19的核苷酸序列,或(基本上)由其组成,所述引物用于在本文所述的ee-gm5pcr方案中使用。包含引物的所述试剂盒,其中所述引物包含seqidno.12和seqidno.13的核苷酸序列或(基本上)由其组成,可以进一步包含探针,所述探针可以包含seqidno.14的核苷酸序列,或(基本上)由其组成;包含引物的所述试剂盒,其中所述引物包含seqidno.18和seqidno.19的核苷酸序列或(基本上)由其组成,可以进一步包含探针,所述探针可以包含seqidno.20的核苷酸序列,或(基本上)由其组成。所述试剂盒可进一步包含缓冲液和试剂,例如以下化合物中的任何一种或每种:dntp、(taq)dna聚合酶、mgcl2、稳定剂和任选地染料。
本发明还涉及用于在生物样品中鉴定原种事件ee-gm5的试剂盒,该试剂盒包含特异性探针,该探针包含以下序列或(基本上)由其组成,所述序列对应于与ee-gm5的特异性区域具有80%至100%序列同一性的序列(或与其互补),其中该特异性区域包含ee-gm5的5’或3’t-dna侧翼区的部分、和与其邻接的插入t-dna的部分。在一个实施方案中,探针的序列对应于特定区域,该区域包含ee-gm5的5’或3’t-dna侧翼区的部分和与其邻接的插入t-dna的部分。最优选地,该特异性探针包含与seqidno.1、3或5中任一个的序列具有80%至100%序列同一性的序列,或与seqidno.2、4或6中任一个的序列具有80%至100%序列同一性的序列,或(基本上)由其组成(或与其互补);或该特异性探针包含与seqidno.5的序列的至少50个连续核苷酸的部分具有80%至100%序列同一性的序列,或与seqidno.6的序列的至少50个连续核苷酸的部分具有80%至100%序列同一性的序列,或(基本上)由所述序列组成(或与其互补),其中seqidno.5或6的所述部分中的每一个包含插入t-dna的序列和大约相等长度的t-dna侧翼序列。
根据本发明的另一个方面,公开了dna分子,该dna分子包含ee-gm5的t-dna侧翼序列和插入t-dna两者的足够长度的多核苷酸,以便用作检测ee-gm5的引物或探针,或用于表征包含事件ee-gm5的植物。此类序列可以包含,在ee-gm5事件的5’和3’连接位点之一或两者处,分别位于连接位点两侧的ee-gm5的t-dna侧翼序列和插入t-dna序列,其中所述t-dna侧翼序列可以为至少9、至少10、至少15、至少20、或至少30个核苷酸、或可以为9、10、15、20、或30个核苷酸,其中所述插入t-dna序列可以为相同的核苷酸数目。最优选地,此dna分子包含seqidno.1、3、或5中任一个的序列或seqidno.2、4、或6中任一个的序列。在一个实施方案中,此dna分子包含seqidno.23、24或25的序列。在本发明的一个方面,提供了包含此特异性dna分子的大豆植物和种子。
本发明的方法和试剂盒可用于不同目的,例如但不限于以下目的:鉴定植物、植物材料或产品中ee-gm5的存在或确定ee-gm5的(较低)阈值,所述产品例如但不限于包含植物材料或衍生自植物材料的食品或饲料产品(新鲜的或加工的);另外地或可替代地,本发明的方法和试剂盒可用于鉴定转基因植物材料,以分离转基因和非转基因材料;另外地或可替代地,本发明的方法和试剂盒可用于确定包含ee-gm5的植物材料的质量(即,纯物质的百分比)。
本发明进一步涉及ee-gm5的5’和/或3’t-dna侧翼区以及从ee-gm5的5’和/或3’t-dna侧翼序列开发的特异性引物和探针。
本发明还涉及从包含原种事件ee-gm5的植物中获得的基因组dna,特别是包含ee-gm5事件-特异性序列的基因组dna,所述特异性序列例如ee-gm5连接序列的一个或两个(包含t-dna侧翼dna的部分和与其邻接的插入t-dna,是ee-gm5特征性的),例如,seqidno.1、3、5、或24的序列中的任一个和/或seqidno.2、4、6、或25的序列中的任一个。这种基因组dna可用作本文所述的鉴定试验中的参考对照材料。
本文还提供了转基因线虫抗性和除草剂耐受性的大豆植物、或其细胞、部分、种子或子代,其分别包含至少一个原种事件,所述原种事件包含插入t-dna,包含:
i)第一嵌合基因,其包含在植物可表达型启动子控制下的编码cry14ab-1蛋白的来自苏云金芽孢杆菌的cry14ab-1.b基因,例如包含植物可表达型启动子和seqidno.7的编码序列的嵌合基因;和
ii)第二嵌合基因,其包含在植物可表达型启动子控制下的编码更耐受的hppd酶的来自假单胞菌属的经修饰的hppdpf-4pa基因,例如包含植物可表达型启动子和seqidno.9的编码序列的嵌合基因。
在一个实施方案中,所述原种事件包含紧邻所述插入t-dna的上游并与其邻接的seqidno.5的核苷酸1至166或seqidno.24的核苷酸1至1113,以及紧邻所述插入t-dna的下游并与其邻接的seqidno.6的核苷酸359至691或seqidno.25的核苷酸359至1449。
在另外的实施方案中,通过用生长自参考种子的大豆植物育种,可获得所述原种事件,该参考种子包含所述事件,在atcc处保藏号pta-123625下保藏。
在另一个实施方案中,当使用两个引物(分别包含seqidno.12和seqidno.13的核苷酸序列)和pcr分析时,由所述大豆植物、或其细胞、部分、种子或子代的基因组dna产生85bp的dna片段,或当使用两个引物(分别包含seqidno.18和seqidno.19的核苷酸序列)和pcr分析时,产生84bp的dna片段。
本文还提供了用于在生物样品中鉴定具有线虫抗性(例如,scn和/或短体线虫属和/或根结和/或肾形线虫抗性)和hppd抑制剂型除草剂(例如,异噁唑草酮、苯唑草酮或硝磺草酮)耐受性的转基因大豆植物、或其细胞、部分、种子或子代的方法,所述方法包括使用至少两个引物和聚合酶链式反应从存在于生物样品中的核酸扩增50和150bp之间的dna片段,所述引物之一识别原种事件ee-gm5的5’t-dna侧翼区(所述5’t-dna侧翼区包含seqidno.5的核苷酸1至核苷酸166的核苷酸序列或seqidno.24的核苷酸1至核苷酸1113的核苷酸序列)或识别所述原种事件的3’t-dna侧翼区(所述3’t-dna侧翼区包含seqidno.6的核苷酸359至核苷酸691的互补序列的核苷酸序列,或seqidno.25的核苷酸359至核苷酸1449的互补序列核苷酸序列);所述引物中的另一引物识别插入t-dna内的序列,该插入t-dna包含seqidno.5的核苷酸167至核苷酸353的互补序列的核苷酸序列,或seqidno.6的核苷酸1至核苷酸358的核苷酸序列,或其中所述插入t-dna包含seqidno.11的核苷酸位置1至核苷酸位置7459的核苷酸序列,或其互补序列。
本文还提供了用于在生物样品中鉴定具有线虫抗性和hppd抑制剂型除草剂耐受性的转基因大豆植物、或其细胞、部分、种子或子代的试剂盒,所述试剂盒包含识别原种事件ee-gm5的5’t-dna侧翼区的一个引物(所述5’t-dna侧翼区包含seqidno.5的核苷酸1至核苷酸166的核苷酸序列,或seqidno.24的核苷酸1至核苷酸1113的核苷酸序列);或识别所述原种事件的3’t-dna侧翼区的一个引物(所述3’t-dna侧翼区包含seqidno.6的核苷酸359至核苷酸691的互补序列的核苷酸序列,或seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列);以及识别插入t-dna内的序列的一个引物,所述插入t-dna包含seqidno.5的核苷酸167至核苷酸353的互补序列的核苷酸序列、或seqidno.6的核苷酸1至核苷酸358的核苷酸序列,或所述插入t-dna包含seqidno.11的核苷酸位置1至核苷酸位置7459的核苷酸序列,或其互补序列。
在本发明的一个实施方案中,如本文所用的原种事件ee-gm5的插入t-dna包含seqidno.5的核苷酸167至核苷酸353的核苷酸序列或其互补序列,和seqidno.6的核苷酸359至核苷酸691的核苷酸序列或其互补序列,或包含与seqidno.11的核苷酸位置7至核苷酸位置7459的核苷酸序列或其互补序列具有至少95%、98%、99%、99.5%、或99.9%序列同一性的序列。
本文还提供了在其基因组中包含核酸分子的大豆植物、植物细胞、组织、或种子,该核酸分子包含seqidno.1、3、5、或24中任一个的核苷酸序列或与其具有80%至100%序列同一性的核苷酸序列,和/或seqidno.2、4、6、或25、或与其具有80%至100%序列同一性的核苷酸序列、和与seqidno.11的核苷酸位置188至核苷酸位置7101的核苷酸序列或其互补序列具有至少80%、85%、90%、95%、97%、98%、99%、99.5%或至少99.9%序列同一性的核苷酸序列。
本发明的一个实施方案提供了大豆植物、植物细胞、组织、或种子,这些大豆植物、植物细胞、组织、或种子在其基因组中包含在标准严格条件下与seqidno.1、3、或5中任一个的核苷酸序列或其互补序列杂交的,或与seqidno.2、4或6中任一个的核苷酸序列或其互补序列杂交的核酸分子。
本文还提供了分离的核酸分子,该核酸分子包含与seqidno.1、3、5、或24中任一个的核苷酸序列或其互补序列具有至少90%、至少95%、至少98%、或至少99%序列同一性的核苷酸序列,或与seqidno.2、4、6、或25中任一个的核苷酸序列或其互补序列具有至少90%、至少95%、至少98%、或至少99%序列同一性的核苷酸序列;或提供了分离的核酸分子,该核酸分子包含在标准严格条件下与seqidno.1、3、5、或24中任一个的核苷酸序列或其互补序列杂交的核苷酸序列,或与seqidno.2、4、6、或25中任一个的核苷酸序列或其互补序列杂交的核苷酸序列。
本文还提供了分离的核酸分子,该核酸分子包含与seqidno.7的核苷酸序列或其互补序列具有至少90%、至少95%、至少98%、或至少99%序列同一性的核苷酸序列,或提供了分离的核酸分子,该核酸分子包含在标准严格条件下与seqidno.7的核苷酸序列或其互补序列杂交的核苷酸序列,其中所述核酸分子编码对胞囊线虫和/或腐线虫(lesionnematode)和/或根结线虫和/或肾形线虫(例如大豆胞囊线虫和/或最短尾短体线虫和/或南方根结线虫和/或肾状肾形线虫)具有活性的杀线虫毒素。在一个实施方案中,将这种核酸分子有效连接至包含(异源)植物可表达型启动子的核酸分子,从而形成嵌合基因。本文还提供了所述核酸分子在转化的植物或种子中用于控制植物致病线虫的用途。本文进一步提供了用于控制根结线虫(例如南方根结线虫、花生根结线虫、北方根结线虫、或爪哇根结线虫,特别是南方根结线虫)的方法,该方法包括使用cry14ab蛋白或编码cry14ab蛋白的dna或包含在植物可表达型启动子控制下的所述dna的植物或种子,其中所述cry14ab蛋白是包含seqidno.8的氨基酸序列的蛋白,或与其具有至少96%、或至少98或至少99%序列同一性的蛋白,或包含seqidno.8的氨基酸位置1至氨基酸位置706的氨基酸序列的蛋白,或与其具有至少96%、或至少98、或至少99%序列同一性的蛋白。本文进一步提供了用于控制肾形线虫(rotylenchulusreniformis)的方法,该方法包括使用cry14ab蛋白或编码cry14ab蛋白的dna或包含在植物可表达型启动子的控制下的所述dna的植物或种子,其中所述cry14ab蛋白是包含seqidno.8的氨基酸序列的蛋白,或与其具有至少96%、或至少98%或至少99%序列同一性的蛋白,或包含seqidno.8的氨基酸位置1至氨基酸位置706的氨基酸序列的蛋白,或与其具有至少96%、或至少98%、或至少99%序列同一性的蛋白。
本文还提供了包含从seqidno.11的核苷酸位置131至核苷酸位置7941的核苷酸序列、或与其具有至少95%、至少96%、至少97%、至少98%、或至少99%序列同一性的核苷酸序列的核酸分子,其中所述核酸分子编码杀线虫cry14ab蛋白和对hppd抑制剂具有耐受性的hppd蛋白。在一个实施方案中,该核酸分子编码seqidno.8的蛋白或与其具有至少99%同一性的蛋白、和seqidno.10的蛋白或与其具有至少99%同一性的蛋白。
本文还提供了在其基因组中包含原种事件ee-gm5的大豆植物细胞,该原种事件ee-gm5是在定义的基因座处的外源dna或插入的t-dna,其中该原种事件ee-gm5如包含于在atcc处保藏号pta-123625下保藏的参考种子中的原种事件ee-gm5,其中所述插入的t-dna包含嵌合cry14ab-1-编码基因和嵌合hppd-4-编码基因,并且其中所述原种事件的特征在于seqidno.1或3的5’连接序列以及seqidno.2或4的3’连接序列;或这种细胞是种子细胞,或这种细胞当使用两个引物(分别包含seqid12和seqid13的核苷酸序列)的pcr分析其基因组dna时产生85bp的dna片段。
本发明提供了核酸分子,该核酸分子包含在atcc处保藏号pta-123625下保藏的参考种子中的原种事件ee-gm5的核苷酸序列,其中所述原种事件包含嵌合cry14ab-1-编码基因和hppd-4-编码基因,并且包含seqidno.1或3的序列和seqidno.2或4的序列。
本发明还提供了核酸分子,该核酸分子按顺序包含以下核苷酸序列:a)从seqidno.5的核苷酸1至166的核苷酸序列或与其具有至少99%同一性的序列,b)从seqidno.11的核苷酸188至核苷酸7101的核苷酸序列或与其具有至少99%同一性的序列,和c)从seqidno.6的核苷酸359至核苷酸691的核苷酸序列或与其具有至少99%同一性的序列,例如这种核酸分子包含与从seqidno.11的核苷酸188至核苷酸7101的核苷酸序列具有至少99,5%或至少99,9%同一性的序列b)。
本发明还提供了核酸分子,该核酸分子按顺序包含以下核苷酸序列:a)从seqidno.24的核苷酸1至1113的核苷酸序列或与其具有至少99%同一性的序列;b)从seqidno.23的核苷酸1114至核苷酸8572的核苷酸序列或与其具有至少99%同一性的序列;和c)从seqidno.25的核苷酸359至核苷酸1449的核苷酸序列或与其具有至少99%同一性的序列,例如这种核酸分子包含与seqidno.23的核苷酸序列具有至少99,5%或至少99,9%同一性的序列b)。根据本发明,还提供了用于生产大豆产品的方法,该方法包括获得包含如上所述的原种事件ee-gm5的大豆种子,并由其生产大豆产品。在一个实施方案中,这种方法中的大豆产品是或包括以下各项:大豆粕、磨碎的种子、粉、或片、或大豆油、大豆蛋白、卵磷脂、豆浆、豆腐、人造黄油、生物柴油、生物复合材料、粘合剂、溶剂、润滑剂、清洁剂、泡沫、涂料、油墨、蜡烛、或含有大豆油或大豆蛋白的食品或饲料产品。在另一个实施方案中,这种大豆产品包含对原种事件ee-gm5具有特异性的核酸。在一个实施方案中,对原种事件ee-gm5具有特异性的所述核酸包含seqidno.1或3的序列,或seqidno.2或4的序列。
本文还提供了由包含如上所述原种事件ee-gm5的种子产生的大豆产品,其中所述大豆产品是或包括大豆粕、磨碎的种子、粉、或片,并且包含对原种事件ee-gm5具有特异性的核酸,其中使用如本文所述的方法可检测所述核酸。在一个实施方案中,对原种事件ee-gm5具有特异性的所述核酸包含seqidno.1或3的序列,或seqidno.2或4的序列。在另一个实施方案中,对原种事件ee-gm5具有特异性的所述核酸包含seqidno.5或24的序列,或seqidno.6或25的序列。在一个实施方案中,对原种事件ee-gm5具有特异性的所述核酸包含seqidno.5或24的序列,和seqidno.6或25的序列。
本文还提供了包含原种事件ee-gm5的大豆种子用于获得大豆产品的用途,其中所述原种事件包含seqidno.1、3、5、或24中任一个的序列和/或seqidno.2、4、6、或25中任一个的序列。在一个实施方案中,在这种用途中,大豆产品是大豆粕、磨碎的大豆种子、大豆粉、或大豆片中的任一种。
此外,本文提供了用于产生包含与另一scn抗性基因座/基因相组合的原种事件ee-gm5的大豆植物或种子的方法,例如可以将原种事件ee-gm5与存在于相同大豆植物/种子中的另一scn抗性基因座/基因进行组合,并且种植包含ee-gm5和所述另一scn抗性基因座/基因的种子。在一个实施方案中,本发明的植物、细胞或种子包含存在于大豆中的一个或多个其他的scn抗性基因座/基因,从而在本发明的大豆植物、细胞、或种子中得到不同scn抗性来源的组合。若干大豆scn抗性基因座或基因是已知的,并且可以将这些基因座或基因中的一个或多个与ee-gm5组合在相同植物、细胞或种子中,大豆scn抗性基因座或基因可以是例如来自抗性来源pi88788、pi548402(peking)、pi437654(hartwig或
表3.
本文还提供了用于保护出苗的大豆植物免受杂草竞争的方法,该方法包括用hppd抑制剂型除草剂在处理其中播种了包含如上所述原种事件ee-gm5的种子的田地,其中这些植物对hppd抑制剂型除草剂具有耐受性。在一个实施方案中,在这种方法中该hppd抑制剂型除草剂是异噁唑草酮、苯唑草酮、或硝磺草酮。
本文还提供了用于保护出苗的大豆植物免受杂草竞争的方法,该方法包括在种植大豆植物或播种种子之前,用hppd抑制剂型除草剂对待种植包含如上所述原种事件ee-gm5的大豆植物的田地进行处理,随后在所述预处理的田地中种植或播种所述大豆植物或种子,其中这些植物对hppd抑制剂型除草剂具有耐受性。
本文还提供了用于在包含如上所述原种事件ee-gm5的大豆植物的田地中控制杂草的方法,该方法包括用有效量的hppd抑制剂型除草剂处理所述田地,其中这些植物对这种除草剂具有耐受性。
本文还另外提供了转基因大豆植物、种子或其子代在大豆田地中控制杂草的用途,其中所述植物、种子或子代分别在其基因组中包含原种事件ee-gm5,其中ee-gm5是在定义的基因座处的t-dna(如包含于在atcc处保藏号pta-123625下保藏的参考种子中),其中所述t-dna包含嵌合cry14ab-1-编码基因和嵌合hppd-4-编码基因,并且其中所述原种事件的特征在于seqidno.1或3的5’连接序列以及seqidno.2或4的3’连接序列。在一个实施方案中,在这种用途中,转基因大豆植物、其种子或子代对线虫具有抗性和/或对hppd抑制剂型除草剂具有耐受性。在一个实施方案中,所述t-dna包含嵌合cry14ab-1-编码基因和嵌合hppd-4-编码基因,并且所述原种事件特征在于seqidno.5或24的5’连接序列和seqidno.6或25的3’连接序列。
本文还提供了应用在其基因组中包含原种事件ee-gm5的大豆植物或种子来生长线虫抗性和/或除草剂耐受性植物的用途,其中所述原种事件ee-gm5是在定义的基因座处的插入t-dna(如包含于在atcc处保藏号pta-123625下保藏的参考种子中的),其中所述插入的t-dna包含嵌合cry14ab-1-编码基因和嵌合hppd-4-编码基因,并且其中所述原种事件特征在于seqidno.1或3的5’连接序列以及seqidno.2或4的3’连接序列。在一个实施方案中,在这种用途中,大豆植物或种子对scn线虫具有抗性和/或对hppd抑制剂型除草剂具有耐受性。在一个实施方案中,所述t-dna包含嵌合cry14ab-1-编码基因和嵌合hppd-4-编码基因,并且所述原种事件特征在于seqidno.5或24的5’连接序列和seqidno.6或25的3’连接序列。
本文还提供了包含原种事件ee-gm5的大豆种子用于获得大豆产品的用途,其中ee-gm5是如上所述的。
本文还提供了用于生产包含原种事件ee-gm5的大豆植物或种子的方法,该方法包括将包含ee-gm5的植物与另一大豆植物杂交,并且种植从所述杂交获得的包含ee-gm5的种子。在一个实施方案中,这种方法包括在所述种子或植物上施用hppd抑制剂型除草剂的步骤。
根据本发明,还提供了应用包含如上所述的原种事件ee-gm5的大豆种子和hppd抑制剂型除草剂来控制大豆田地中杂草的用途,以及包含原种事件ee-gm5的大豆种子在生长hppd抑制剂型除草剂耐受性大豆的方法中的用途,其中所述种子如上所述。
本文进一步提供了如上所述的原种事件ee-gm5用于赋予大豆植物或种子对线虫的抗性和/或对hppd抑制剂型除草剂的耐受性的用途,或包含原种事件ee-gm5的大豆植物或种子与hppd抑制剂型除草剂组合用于生长大豆的用途。
本文还提供了对ee-gm5具有特异性的引物对,以及使用这种引物对的试剂盒或方法,其中将所述引物对的至少一个引物进行标记(例如用可检测或可筛选的结构部分进行标记),或其中所述引物中的至少一个的5’末端包含一个或多个错配或与ee-gm5的5’或3’侧翼序列无关或与ee-gm5的t-dna序列无关的核苷酸序列;或其中所述引物中的至少一个在其3’末端处包含跨t-dna侧翼序列和t-dna序列之间的连接区域的核苷酸序列(其中,所述连接区域是seqidno.5中的核苷酸166-167、seqidno.24中的核苷酸1113-1114、或seqidno.6或25中的核苷酸358-359),条件是在3’末端处的17个连续核苷酸非唯一地来源于seqidno.5或24、或6或25中的t-dna或t-dna侧翼序列;或其中所述引物中的至少一个包含与ee-gm5的5’或3’侧翼区内的序列或与ee-gm5的插入t-dna内的序列具有在80%和100%之间的同一性的序列,并且所述引物序列包含与所述5’或3’侧翼区或所述t-dna的至少一个错配,条件是该至少一个错配仍允许在优化的检测条件(例如,优化的pcr条件)下用这些引物特异性鉴定原种事件ee-gm5;或其中所述引物中至少一个的核苷酸序列包含与来自另一来源的核酸融合的核酸的核苷酸序列,或其互补序列。
根据本发明的其他实施方案总结在以下段落中:
1.一种用于在生物样品中鉴定原种事件ee-gm5的方法,该方法包括用特异性引物对或探针检测ee-gm5特异区域,该引物对或探针特异性识别ee-gm5中的5’或3’t-dna侧翼区(的至少一部分)以及与其邻接的插入t-dna(的至少一部分)。
2.根据段落1所述的方法,所述方法包括使用至少两个引物和聚合酶链式反应从存在于所述生物样品中的核酸扩增50bp和1000bp之间的dna片段,其中所述引物之一识别ee-gm5中的5’t-dna侧翼区或识别ee-gm5中的3’t-dna侧翼区,其中所述5’t-dna侧翼区包含seqidno.5核苷酸1至核苷酸166的核苷酸序列或seqidno.24的核苷酸1至核苷酸1113的核苷酸序列,其中所述3’t-dna侧翼区包含seqidno.6的核苷酸359至核苷酸691的互补序列的核苷酸序列或seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列;其中所述引物的另一引物识别插入t-dna内的序列,其中所述插入t-dna包含seqidno.5的核苷酸167至核苷酸353的核苷酸序列或其互补序列、或seqidno.6的核苷酸1至核苷酸358的核苷酸序列或其互补序列、或seqidno.23的核苷酸1114至核苷酸8572的核苷酸序列或其互补序列。
3.根据段落2所述的方法,其中识别5’t-dna侧翼区的所述引物包含选自以下的17至200个连续核苷酸的核苷酸序列:seqidno.5核苷酸1至核苷酸166的核苷酸序列、或seqidno.24的核苷酸1至核苷酸1113的核苷酸序列,或识别ee-gm5的3’t-dna侧翼区的所述引物包含选自以下的17至200个连续核苷酸的核苷酸序列:seqidno.6的核苷酸359至核苷酸691的互补序列的核苷酸序列、或seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列,并且识别插入t-dna内的序列的所述引物包含选自以下的17至200个连续核苷酸的核苷酸序列:seqidno.5的核苷酸167至核苷酸353的核苷酸序列或其互补序列、或seqidno.6的核苷酸1至核苷酸358的核苷酸序列或其互补序列、或seqidno.11的核苷酸1至核苷酸7459的核苷酸序列或其互补序列、或seqidno.23的核苷酸1114至核苷酸8572的核苷酸序列或其互补序列。
4.根据段落2所述的方法,其中识别5’t-dna侧翼区的所述引物在其最3’末端包含选自以下的至少17个连续核苷酸的核苷酸序列:seqidno.5的核苷酸1至核苷酸166的核苷酸序列或seqidno.24的核苷酸1至核苷酸1113的核苷酸序列,或识别ee-gm5的3’t-dna侧翼区的所述引物在其最3’末端包含选自以下的至少17个连续核苷酸的核苷酸序列:seqidno.6的核苷酸359至核苷酸691的互补序列的核苷酸序列或seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列,并且识别插入t-dna内序列的所述引物在其最3’末端包含选自以下的至少17个连续核苷酸:seqidno.5的核苷酸167至核苷酸353的核苷酸序列的互补序列、或seqidno.6的核苷酸1至核苷酸358的核苷酸序列、或seqidno.11的核苷酸1至核苷酸7459的核苷酸序列或其互补序列、或seqidno.23的核苷酸1114至核苷酸8572的核苷酸序列或其互补序列。
5.根据段落4所述的方法,其中所述引物分别包含seqidno.12和seqidno.13的序列,或分别包含seqidno.18和seqidno.19的序列。
6.根据段落5所述的方法,该方法包括使用pcr扩增85bp或84bp的ee-gm5特异性片段。
7.根据段落2至6中任一项所述的方法,该方法进一步包括探针杂交的步骤,该探针对于用所述至少两个引物扩增的dna片段具有特异性。
8.根据段落7所述的方法,其中所述探针识别所述5’t-dna侧翼区的部分和与其邻接的插入t-dna的部分,或其中所述探针识别所述3’t-dna侧翼区的部分和与其邻接的插入t-dna的部分,或识别所述5’t-dna侧翼区的部分和与其邻接的插入t-dna的部分,例如其中所述探针包含seqidno.1或3或seqidno2或4的核苷酸序列。
9.根据段落8所述的方法,其中所述引物分别包含seqidno.12和seqidno.13的序列,并且其中所述探针包含seqidno.14的序列,或其中所述引物分别包含seqidno.18和seqidno.19的序列,并且其中所述探针包含seqidno.20的序列。
10.一种试剂盒,该试剂盒包含识别ee-gm5的5’t-dna侧翼区的一个引物(所述5’t-dna侧翼区包含seqidno.5的核苷酸1至核苷酸166的或从seqidno.24的核苷酸1至核苷酸1113的核苷酸序列)、或识别ee-gm5的3’t-dna侧翼区的一个引物(所述3’t-dna侧翼区包含seqidno.6的核苷酸359至核苷酸691的互补序列的核苷酸序列或seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列),和识别插入t-dna内的序列的一个引物(所述插入t-dna包含seqidno.5的核苷酸167至核苷酸353的核苷酸序列的互补序列、或seqidno.6的核苷酸1至核苷酸358的核苷酸序列、或seqidno.11的核苷酸1至核苷酸7459的核苷酸序列或其互补序列、或seqidno.23的核苷酸1114至核苷酸8572的核苷酸序列或其互补序列)。
11.根据段落10所述的试剂盒,其中识别5’t-dna侧翼区的所述引物包含选自以下的17至200个连续核苷酸的核苷酸序列:seqidno.5核苷酸1至核苷酸166的或seqidno.24的核苷酸1至核苷酸1113的核苷酸序列,或识别ee-gm5的3’t-dna侧翼区的所述引物包含选自以下的17至200个连续核苷酸的核苷酸序列:seqidno.6的核苷酸359至核苷酸691的互补序列的核苷酸序列或seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列,并且识别插入t-dna内的序列的所述引物包含选自以下的17至200个连续核苷酸的核苷酸序列:seqidno.5的核苷酸167至核苷酸353的核苷酸序列的互补序列、或seqidno.6的核苷酸1至核苷酸358的核苷酸序列、或seqidno.11的核苷酸1至核苷酸7459的核苷酸序列或其互补序列、或seqidno.23的核苷酸1114至核苷酸8572的核苷酸序列或其互补序列。
12.根据段落10所述的试剂盒,其中识别5’t-dna侧翼区的所述引物在其最3’末端包含选自以下的至少17个连续核苷酸的核苷酸序列:seqidno.5的核苷酸1至核苷酸166的核苷酸序列或seqidno.24的核苷酸1至核苷酸1113的核苷酸序列,或识别ee-gm5的3’t-dna侧翼区的所述引物在其最3’末端包含选自以下的至少17个连续核苷酸的核苷酸序列:seqidno.6的核苷酸359至核苷酸691的互补序列的核苷酸序列或seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列,并且识别插入t-dna内序列的所述引物在其3’末端包含选自以下的至少17个连续核苷酸:seqidno.5的核苷酸167至核苷酸353的核苷酸序列的互补序列、或seqidno.6的核苷酸1至核苷酸358的核苷酸序列、或seqidno.11的核苷酸1至核苷酸7459的核苷酸序列或其互补序列、或seqidno.23的核苷酸1114至核苷酸8572的核苷酸序列或其互补序列。
13.根据段落10所述的试剂盒,该试剂盒包含含有seqidno.12的序列的引物、和含有seqidno.13的序列的引物,或包含含有seqidno.18的序列的引物和含有seqidno.19的序列的引物。
14.根据段落10所述的试剂盒,该试剂盒进一步包含探针,所述探针识别在识别5’t-dna侧翼区的引物和识别插入t-dna内序列的引物之间的序列,或识别在识别3’t-dna侧翼区的引物和识别插入的t-dna内序列的引物之间的序列。
15.根据段落14所述的试剂盒,其中所述探针识别所述5’t-dna侧翼区的部分和与其邻接的插入t-dna的部分,或其中所述探针识别所述3’t-dna侧翼区的部分和与其邻接的插入t-dna的部分。
16.根据段落15所述的试剂盒,其中所述引物包含seqidno.12和seqidno.13的序列,并且其中所述探针包含seqidno.14的序列,或其中所述引物包含seqidno.18和seqidno.19的序列,并且其中所述探针包含seqidno.20的序列。
17.适用于ee-gm5特异性检测的引物对,其包含第一引物和第二引物,其中该第一引物包含在优化的检测条件下特异性识别ee-gm5中的5’或3’t-dna侧翼区内的序列的序列,该第二引物包含在优化的检测条件下特异性识别在ee-gm5中邻接所述侧翼5’或3’区域的插入t-dna内序列的序列,所述5’t-dna侧翼区包含seqidno.5的核苷酸1至核苷酸166的核苷酸序列或seqidno.24的核苷酸1至核苷酸1113的核苷酸序列,所述3’t-dna侧翼区包含seqidno.6的核苷酸359至核苷酸691的互补序列的核苷酸序列或seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列,所述插入的t-dna包含seqidno.5的核苷酸167至核苷酸353的核苷酸序列、或seqidno.6的核苷酸1至核苷酸358的核苷酸序列、或seqidno.11的核苷酸1至核苷酸7459的核苷酸序列或其互补序列、或seqidno.23的核苷酸1114至核苷酸8572的核苷酸序列或其互补序列。
18.引物,该引物在其最3’末端包含seqidno.12的序列、或seqidno.13的序列、或seqidno.18的序列、或seqidno.19的序列。
19.引物对,该引物对包含第一引物和第二引物,该第一引物在其最3’末端处包含seqidno.12的序列,该第二引物在其最3’末端处包含seqidno.13的序列;或该引物对包含第一引物和第二引物,该第一引物在其最3’末端处包含seqidno.18的序列,该第二引物在其最3’末端处包含seqidno.19的序列。
20.根据段落1所述的方法,该方法包括将生物样品的核酸与ee-gm5的特异性探针杂交。
21.根据段落20所述的方法,其中所述特异性探针的序列与如下序列具有至少80%序列同一性,所述序列包含ee-gm5的5’t-dna侧翼序列或3’t-dna侧翼序列的部分以及与其邻接的插入t-dna的序列。
22.根据段落21所述的方法,其中所述特异性探针的序列包含与seqidno.1、3、或5中任一个的序列或seqidno.2、4、或6中任一个的序列或所述序列的互补序列具有至少80%序列同一性的序列。
23.根据段落22所述的方法,其中所述探针包含seqidno.1或3中任一个的序列或seqidno.2或4中任一个的序列。
24.一种用于在生物样品中鉴定原种事件ee-gm5的试剂盒,所述试剂盒包含能够与ee-gm5的特异区域特异性地杂交的特异性探针。
25.根据段落24所述的试剂盒,其中所述特异性探针的序列与如下序列具有至少80%序列同一性,其中所述序列包含ee-gm5的5’t-dna侧翼序列的部分或3’t-dna侧翼序列的部分以及与其邻接的插入t-dna序列的部分。
26.根据段落25所述的试剂盒,其中所述特异性探针的序列包含与seqidno.1、3或5中任一个或seqidno.2、4或6中任一个的序列或所述序列的互补序列具有至少80%序列同一性的核苷酸序列。
27.一种特异性探针,该特异性探针用于在生物样品中鉴定原种事件ee-gm5。
28.根据段落27所述的探针,该探针包含核苷酸序列,该核苷酸序列与如下序列或其互补序列具有至少80%序列同一性,其中所述序列包含ee-gm5的5’t-dna侧翼序列的部分或3’t-dna侧翼序列的部分和与其邻接的插入t-dna序列的部分。
29.根据段落28所述的探针,该探针与seqidno.1、3、或5中的任一个的序列、或seqidno.2、4、或6中任一个的序列、或所述序列的互补序列具有至少80%序列同一性。
30.一种包含核苷酸序列的特异性探针,该核苷酸序列与seqidno.1、3、或5中的任一个的序列或seqidno.2、4、或6中任一个的序列或所述序列的互补序列是基本上相似的。
31.一种特异性探针,该探针包含seqidno.1或3的序列、或seqidno.2或4的序列。
32.一种用于确定种子纯度的方法,该方法包括在种子样品中用特异性引物对或探针检测ee-gm5特异区域,该引物对或探针特异性识别ee-gm5中的5’或3’t-dna侧翼区和与其邻接的插入t-dna。
33.根据段落32所述的方法,该方法包括使用至少两个引物和聚合酶链式反应存在于所述生物样品中的核酸扩增50bp和1000bp之间的dna片段,其中所述引物之一识别ee-gm5的5’t-dna侧翼区(所述5’t-dna侧翼区包含seqidno.5的核苷酸1至核苷酸166的或seqidno.24的核苷酸1至核苷酸1113的核苷酸序列)或识别ee-gm5的3’t-dna侧翼区(所述3’t-dna侧翼区包含seqidno.6的核苷酸359至核苷酸691的互补序列的核苷酸序列、或seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列),所述引物的另一引物识别插入t-dna内的序列(所述插入t-dna包含seqidno.5的核苷酸167至核苷酸353的核苷酸序列的互补序列、或seqidno.6的核苷酸1至核苷酸358的核苷酸序列、或seqidno.11的核苷酸1至核苷酸7459的核苷酸序列或其互补序列内的序列、或seqidno.23的核苷酸1114至核苷酸8572的核苷酸序列或其互补序列内的序列);并且与探针杂交,该探针对由所述至少两个引物扩增的dna片段具有特异性。
34.根据段落33所述的方法,该方法包括扩增85bp的dna片段,并且其中所述引物分别包含seqidno.12和seqidno.13的序列,并且其中所述探针包含seqidno.14的序列,或扩增84bp的dna片段,并且其中所述引物分别包含seqidno.18和seqidno.19的序列,并且其中所述探针包含seqidno.20的序列。
35.一种用于筛选存在ee-gm5的种子的方法,该方法包括在种子批的样品中用特异性引物对或探针检测ee-gm5特异区域,该引物对或探针特异性识别ee-gm5中5’或3’t-dna侧翼区和与其邻接的插入t-dna。
36.根据段落35所述的方法,该方法包括使用至少两个引物和聚合酶链式反应从存在于所述生物样品中的核酸扩增50bp和1000bp之间的dna片段,所述引物之一识别ee-gm5中的插入t-dna的5’t-dna侧翼区(所述5’t-dna侧翼区包含seqidno.5的核苷酸1至核苷酸166的或seqidno.24的核苷酸1至核苷酸1113的核苷酸序列)或识别ee-gm5中的插入t-dna的3’t-dna侧翼区(所述3’t-dna侧翼区包含seqidno.6的核苷酸359至核苷酸691的互补序列的核苷酸序列、或seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列),所述引物的另一引物识别插入t-dna内的序列(所述插入t-dna包含seqidno.5的核苷酸167至核苷酸353的核苷酸序列的互补序列、或seqidno.6的核苷酸1至核苷酸358的核苷酸序列、或包含seqidno.11的核苷酸1至核苷酸7459的核苷酸序列或其互补序列、或seqidno.23的核苷酸1114至核苷酸8572的核苷酸序列或其互补序列);并且与探针杂交,该探针对由所述至少两个引物扩增的dna片段具有特异性,例如探针包含seqidno.1或3、或seqidno.2或4的序列、或其互补序列。
37.根据段落36所述的方法,该方法包括扩增85bp的dna片段,并且其中所述引物分别包含seqidno.12和seqidno.13的序列,并且其中所述探针包含seqidno.14的序列。
38.一种用于确定包含原种事件ee-gm5的植物、植物材料、或种子的合子状态(zygositystatus)的方法,所述方法包括使用至少三个引物和聚合酶链式反应从存在于所述生物样品中的核酸扩增50bp和1000bp之间的dna片段,所述引物中的两个特异性识别插入前的植物dna,例如包含seqidno.21的核苷酸序列的引物和包含seqidno.19的核苷酸序列的引物,所述引物中的第三个识别插入t-dna内的序列,例如seqidno.18的核苷酸序列,例如所述方法使用所述引物,其中扩增84bp和72bp的dna片段。
39.一种通过与基本上互补的经标记的核酸探针杂交来检测生物样品中原种事件ee-gm5的存在的方法,其中通过再循环靶核酸序列来放大探针:靶核酸比率,所述方法包括:
a)将所述靶核酸序列与第一核酸寡核苷酸杂交,该第一核酸寡核苷酸包含seqidno.5的核苷酸位置167至核苷酸位置184的核苷酸序列或其互补序列,或所述第一核酸寡核苷酸包含seqidno.6的核苷酸位置341至核苷酸位置358的核苷酸序列或其互补序列;
b)将所述靶核酸序列与第二核酸寡核苷酸杂交,该第二核酸寡核苷酸包含seqidno.5的核苷酸149至核苷酸166的核苷酸序列或其互补序列,或所述核酸寡核苷酸包含seqidno.6的核苷酸359至核苷酸376的核苷酸序列或其互补序列,其中所述第一和第二寡核苷酸重叠至少一个核苷酸,并且其中所述第一或所述第二寡核苷酸被标记以构成所述的经标记的核酸探针;
c)用酶仅切割探针:靶核酸序列双链体内经标记的探针,该酶引起选择性探针切割,导致双链体解离,而留下完整的靶序列;
d)通过重复步骤(a)至(c),再循环该靶核酸序列;以及
e)检测切割的经标记的探针,由此确定所述靶核酸序列的存在,以及检测所述生物样品中原种事件ee-gm5的存在。
40.一种转基因大豆植物、或细胞、部分、种子或其子代,该转基因大豆植物、或细胞、部分、种子或其子代在基因组中分别包含原种事件ee-gm5,包含所述事件的参考种子已经在atcc以保藏号pta-123625保藏。
41.根据段落40所述的转基因大豆植物、种子、细胞、部分或子代,当使用分别包含seqid12和seqid13的核苷酸序列的两个引物和pct分析ee-gm5时,该转基因大豆植物、种子、细胞、部分或子代的基因组dna产生85bp的dna片段。
42.包含原种事件ee-gm5的种子,该原种事件ee-gm5是在大豆基因组中特定位置处的插入的t-dna,如已经保藏于atcc处保藏号pta-123625下的保藏种子或来自该种子的衍生物中的事件。
43.一种包含原种事件ee-gm5的大豆植物、植物部分、细胞、或组织、或种子,其可从根据段落42所述的种子获得。
44.在基因组中包含原种事件ee-gm5的大豆植物、或其种子、细胞或组织,其可以通过繁殖大豆植物和/或用其育种来获得,其中所述大豆植物从以保藏号pta-123625保藏在atcc的种子生长而成。
45.一种包含原种事件ee-gm5的大豆种子,其中包含所述事件的参考种子已经在atcc以保藏号pta-123625保藏。
46.一种包含原种事件ee-gm5的转基因大豆植物、细胞、或组织,其可以从根据段落45所述的种子获得。
47.根据段落40、41、43、44和46中任一项所述的大豆植物细胞,该细胞是非繁殖性植物细胞。
48.一种用于产生包含原种事件ee-gm5的大豆植物或种子的方法,该方法包括将根据段落40、41、43、44和46中任一项所述的植物与另一大豆植物杂交,并种植由所述杂交获得的种子。
49.包含原种事件ee-gm5的大豆基因组dna。
50.一种核酸分子,该核酸分子包含与seqidno.1、3、或5中任一个的序列或seqidno.2、4、或6中任一个的序列或所述序列的互补序列基本上相似的核苷酸序列,例如包含与seqidno.5或24或6或25的核苷酸序列或其互补序列具有至少99%或至少99,5%序列同一性的核苷酸序列的核酸分子,例如在大豆中赋予hppd抑制剂型除草剂耐受性和/或scn抗性的所述核酸。
51.根据段落50所述的核酸分子,其包含seqidno.1或3、或seqidno.2或4中任一个的核苷酸序列或所述序列的互补序列,例如这样的核酸分子或其互补序列,所述核酸分子还包含seqidno.7和9的核苷酸序列或与其具有至少98%序列同一性的核苷酸序列,或这样的核酸分子,其包含seqidno.11的核苷酸位置188至核苷酸位置7101的核苷酸序列或与其具有至少98%序列同一性的核苷酸序列。
52.一种大豆植物、细胞、植物部分、种子或其子代,该大豆植物、细胞、植物部分、种子或其子代包含这些段落中任一项的核酸分子,例如包含以下核酸分子的大豆植物、细胞、植物部分、种子或其子代,该核酸分子包含seqidno.1、3、或5的核苷酸序列或seqidno.2、4、或6的核苷酸序列;或在基因组中包含seqidno.3的核苷酸序列和seqidno.4的核苷酸序列的大豆植物、细胞、植物部分、种子或其子代;或在基因组中包含seqidno.5的核苷酸序列和seqidno.6的核苷酸序列的大豆植物、细胞、植物部分、种子或其子代;或在基因组中包含seqidno.24的核苷酸序列和seqidno.25的核苷酸序列的大豆植物、细胞、植物部分、种子或其子代,例如这样的大豆植物还可以包含cry14ab-1编码嵌合基因和hppd-4编码嵌合基因,特别地此类嵌合基因分别包含seqidno.7和9的核苷酸序列。
53.一种核酸分子,该核酸分子包含seqidno.1、3、或5或seqidno.2、4、或6中任一个的核苷酸序列,例如包含seqidno.5和seqidno.6的核苷酸序列或其互补序列的核酸分子,或例如包含seqidno.24和seqidno.25的核苷酸序列或其互补序列的核酸分子。
54.一种转基因大豆植物、植物细胞、组织、或种子,该转基因大豆植物、植物细胞、组织、或种子在其基因组中包含事件ee-gm5,该事件特征在于以下核酸分子,该核酸分子包含与seqidno.1、3、5、或24中任一个的序列或seqidno.2、4、6、或25中任一个的序列或所述序列的互补序列基本上相似的核苷酸序列,其中所述大豆植物还包含cry14ab-1编码嵌合基因和hppd-4编码嵌合基因。
55.一种大豆植物、细胞、组织、或种子,该大豆植物、细胞、组织、或种子包含ee-gm5,并且在其细胞的基因组中包含与seqidno.1、3、或5中任一个的序列或seqidno.2、4、或6中任一个的序列或所述序列的互补序列具有至少80%、90%、95%或100%序列同一性的核酸序列,例如还包含cry14ab-1编码嵌合基因和hppd-4编码嵌合基因的大豆植物,或在其细胞的基因组中包含与seqidno.24或seqidno.25的序列具有至少80%、90%、95%或100%序列同一性的核酸序列的大豆植物、细胞、组织、或种子。
56.一种大豆植物、植物细胞、组织、或种子,该大豆植物、植物细胞、组织、或种子在其基因组中包含以下核酸分子,该核酸分子包含与seqidno.5或24或seqidno.6或25的核苷酸序列或其互补序列具有至少99%序列同一性的核苷酸序列;或该大豆植物、植物细胞、组织、或种子在其基因组中包含以下核酸分子,该核酸分子包含与seqidno.5或24和seqidno.6或25的核苷酸序列或其互补序列具有至少99%序列同一性的核苷酸序列。
57.一种大豆植物、植物细胞、组织、或种子,该大豆植物、植物细胞、组织、或种子在其基因组中包含在标准严格条件下与seqidno.5或6的核苷酸序列或其互补序列杂交的核酸分子。
58.一种核酸分子,该核酸分子包含与seqidno.5或24或seqidno.6或25的核苷酸序列或其互补序列具有至少99%序列同一性的核苷酸序列,例如包含与seqidno.5或24和seqidno.6或25的核苷酸序列或其互补序列具有至少99%序列同一性的核苷酸序列的核酸分子。
59.一种核酸分子,该核酸分子包含在标准严格条件下与seqidno.5或6的核苷酸序列或其互补序列杂交的核苷酸序列。
60.根据段落50、51、58、和59中任一项所述的核酸分子,该核酸分子还包含seqidno.7和9的核苷酸序列。
61.一种嵌合dna,该嵌合dna包含t-dna5’侧翼区、插入t-dna、和t-dna3’侧翼区,其中所述插入t-dna的序列包含seqidno.11的核苷酸188至核苷酸7101的序列或与其具有至少95%、96%、97%、98%、99%、或至少99,5%同一性的序列,或其中所述插入t-dna的序列包含seqidno.7和9的序列,并且其中所述t-dna5’侧翼区位于所述插入t-dna的紧上游并与其邻接,并且包含seqidno.5的核苷酸1至核苷酸166的序列或与其具有至少95%、96%、97%、98%、99%、或至少99,5%同一性的序列、或包含seqidno.24核苷酸1至核苷酸1113的序列或与其具有至少95%、96%、97%、98%、99%、或至少99,5%同一性的序列,并且其中所述t-dna3’侧翼区位于所述插入的t-dna的紧下游并与其邻接,并且包含seqidno.6的核苷酸359至核苷酸691的序列或与其具有至少95%、96%、97%、98%、99%、或至少99,5%同一性的序列,或包含seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列或与其具有至少95%、96%、97%、98%、99%或至少99,5%同一性的序列。
62.一种核酸分子,该核酸分子包含与seqidno.7的核苷酸序列或其互补序列具有至少98%序列同一性的核苷酸序列,例如seqidno.7的核苷酸序列,例如dna分子,该dna分子包含seqidno.11的核苷酸131至5276的核苷酸序列或其互补序列,或与seqidno.7的序列或seqidno.11的核苷酸位置131至核苷酸位置5276的序列或其互补序列具有至少95%、96%、97%、98%或至少99%序列同一性的编码杀线虫cry14ab蛋白的序列。
63.一种核酸分子,该核酸分子包含与seqidno.9的核苷酸序列或其互补序列具有至少98%序列同一性的核苷酸序列,例如seqidno.9的核苷酸序列,例如dna分子,该dna分子包含seqidno.11的核苷酸5382至7459的核苷酸序列或其互补序列、或与seqidno.9的序列或与seqidno.11的核苷酸位置5382至核苷酸位置7459的序列或其互补序列具有至少95%、96%、97%、98%或至少99%序列同一性的序列,其中当在植物中表达时所述序列编码提供针对hppd抑制剂型除草剂的耐受性的hppd蛋白。
64.一种用于生产大豆产品的方法,该方法包括获得包含如上所述的原种事件ee-gm5的大豆种子,并由其生产大豆产品。
65.根据段落64所述的方法,其中大豆产品是或包含大豆粕(meal)、磨碎的种子、粉、或片。
66.根据段落4或65所述的方法,其中大豆产品包含对原种事件ee-gm5特异性的核酸,例如包含以下核酸的产品,该核酸产生对事件ee-gm5具有诊断性或特异性的扩增子,该核酸是例如seqidno.1或3的序列或seqidno.2或4的序列。
67.一种包含如上所述的原种事件ee-gm5的大豆产品,例如由根据这些段落中任一项所述的大豆植物、细胞、部分、种子或子代生产的大豆产品。
68.根据段落67所述的大豆产品,其中该大豆产品是或包括大豆粕、磨碎的种子、粉、或片。
69.根据段落67或68所述的大豆产品,其中所述大豆产品包含对原种事件ee-gm5特异性的核酸,例如包含以下核酸的产品,该核酸产生对事件ee-gm5具有诊断性或特异性的扩增子,该核酸是例如seqidno.1或3的序列、或seqidno.2或4的序列,或它们的互补序列。
70.一种用于保护出苗的大豆植物免受杂草竞争的方法,该方法包括用hppd抑制剂型除草剂处理播种了包含这些段落中任一项所述的原种事件ee-gm5的种子的田地,其中这些植物对hppd抑制剂型除草剂具有耐受性。
71.一种用于保护出苗的大豆植物免受杂草竞争的方法,该方法包括在种植大豆植物或播种种子之前,用hppd抑制剂型除草剂对待种植包含如上所述原种事件ee-gm5的大豆植物的田地进行处理,随后在所述预处理的田地中种植或播种所述大豆植物或种子,其中这些植物对hppd抑制剂型除草剂具有耐受性。
72.一种用于在包含如上所述原种事件ee-gm5的大豆植物的田地中控制杂草的方法,该方法包括用有效量的hppd抑制剂型除草剂处理所述田地,其中这些植物对hppd抑制剂型除草剂具有耐受性。
73.根据段落70至72中任一项所述的方法,其中该hppd抑制剂型除草剂是异噁唑草酮、苯唑草酮、或硝磺草酮。
74.包含如上所述原种事件ee-gm5的转基因大豆植物、种子或其子代用于产生大豆籽粒或种子的用途。
75.在其基因组中包含如上所述的原种事件ee-gm5的大豆植物或种子用于种植线虫抗性和/或hppd抑制剂型除草剂耐受性的植物的用途。
76.包含原种事件ee-gm5的大豆种子用于获得大豆产品的用途,其中ee-gm5是如上所述的,例如其中这种大豆产品是或包含磨碎的大豆籽粒、大豆粉、大豆粕、或大豆片。
77.包含如上所定义的原种事件ee-gm5的大豆植物或种子与hppd抑制剂型除草剂组合用于种植大豆田地或用于种植大豆作物的用途。
78.一种可从atcc处保藏号pta-123625下保藏的种子中获得的核酸分子,其中所述核酸分子包含seqidno.1、3、或5中任一者的核苷酸序列以及seqidno.2、4、或6中任一者的核苷酸序列。
79.一种大豆植物、细胞、部分或种子,其在基因组中包含原种事件ee-gm5,其中所述原种事件是包含插入的t-dna以及紧邻所述插入的t-dna的5’和3’侧翼序列的遗传基因座,该插入的t-dna含有嵌合hppd-4蛋白编码基因和嵌合cry14ab-1蛋白编码基因,如存在于atcc处保藏号pta-123625下保藏的参考种子中的原种事件ee-gm5。
80.根据段落79所述的植物、细胞、植物部分、或种子的子代植物、细胞、植物部分或种子,其中所述子代植物、细胞、植物部分或种子包含seqidno.3的核苷酸序列和seqidno.4的核苷酸序列。
81.根据段落79所述的大豆植物、细胞、部分、种子或子代,当使用两个引物和pcr分析时,该大豆植物、细胞、部分、或种子的基因组dna产生84bp的dna片段,所述两个引物分别包含seqidno.18和seqidno.19的核苷酸序列。
82.根据以上段落中任一项所述的植物,该植物对异噁唑草酮和/或苯唑草酮和/或硝磺草酮具有耐受性,例如对异噁唑草酮、苯唑草酮和硝磺草酮具有耐受性的植物。
83.一种用于生产对scn具有抗性和对hppd抑制剂型除草剂具有耐受性的大豆植物的方法,该方法包括通过将缺少cry14ab-1-编码基因和缺少hppd-4-编码基因的第一大豆植物与根据以上段落中任一项所述的大豆植物杂交,并且选择对scn具有抗性和对hppd抑制剂型除草剂具有耐受性的子代植物,从而将对scn的抗性和对hppd抑制剂型除草剂的耐受性引入大豆植物的基因组中。
84.包含如上所定义的原种事件ee-gm5的大豆植物或种子用于获得大豆作物的用途,该大豆作物例如是当受线虫或猝死综合征侵染时产量更高的大豆作物。
85.一种生产对线虫或猝死综合征具有改善抗性的大豆作物的方法,该方法包括以下步骤:(a)使用如以上段落中任一项所述的种子种植田地;和(b)收获从所述种子生长的植物上产生的大豆种子,并且任选地(c)在种子出苗之前或之后向种植有所述种子的田地,或在所述大豆植物上,施用一个剂量或多个剂量的hppd抑制剂型除草剂,该剂量足以杀死杂草但对于所述大豆种子或植物是耐受的,例如其中所述线虫是scn或短体线虫属物种或根结线虫或肾形线虫属物种线虫。
86.上述段落中描述的大豆种子用于制备加工食品或饲料商品的用途,其中所述加工食品或饲料商品包含可检测量的核酸,该核酸包含seqidno.1和/或seqidno:2的核苷酸序列或其互补序列。
87.根据段落86所述的用途,其中(i)所述食品或所述饲料商品包括大豆粕、大豆粉、大豆片、或大豆油;(ii)所述核酸包含seqidno:3和/或seqidno:4的核苷酸序列,或其互补序列;或(iii)所述核酸进一步包含在seqidno:7和seqidno.9中包含的核苷酸序列。
88.一种大豆植物、种子或细胞,该大豆植物、种子或细胞在其基因组中包含原种事件ee-gm5,其中原种事件ee-gm5包含与seqidno23中所示序列具有至少90%同一性的核苷酸序列,其中所述原种事件包含嵌合hppd-4-编码基因和嵌合cry14ab-1-编码基因,其中所述植物、种子或细胞对hppd抑制剂型除草剂具有耐受性并具有scn抗性。
89.根据段落88所述的植物,其中原种事件ee-gm5包含核苷酸序列,该核苷酸序列与seqidno.23中列出的序列具有至少95%同一性。
90.根据段落88所述的植物,其中原种事件ee-gm5包含核苷酸序列,该核苷酸序列与seqidno.23中列出的序列具有至少99%、至少99,5%或至少99,9%同一性。
91.一种核酸分子,该核酸分子包含seqidno.23的核苷酸序列或与seqidno.23具有至少99%序列同一性的核苷酸序列,该核酸分子赋予对hppd抑制剂型除草剂的耐受性和/或线虫抗性,例如其中所述线虫是scn或短体线虫属物种或根结线虫或肾形线虫属物种线虫。
92.一种核酸分子,该核酸dna分子包含seqidno.11的核苷酸位置131至核苷酸位置7941的核苷酸序列,或与其具有至少95%、至少96%、至少97%、至少98%、或至少99%序列同一性的核苷酸序列。
93.根据段落92所述的核酸分子,该核酸分子编码对hppd抑制剂具有耐受性的hppd蛋白以及负面影响植物有害生物线虫例如scn、rkn或短体线虫属物种线虫的蛋白质。
94.根据段落93所述的核酸分子,该核酸分子编码seqidno.8的蛋白质或与其具有至少99%同一性的蛋白质,和seqidno.10的蛋白质或与其具有至少99%同一性的蛋白质。
95.一种用于在待种植大豆植物的田地中控制杂草和/或线虫的方法,该方法包括以下步骤:1)用hppd抑制剂型除草剂(例如,异噁唑草酮、苯唑草酮或硝磺草酮)处理所述田地,和2)在所述经处理的田地中种植或播种包含如上所述原种转化事件ee-gm5的大豆植物或种子,其中包含所述原种事件的参考种子保藏在atcc处保藏号pta-123625下。
96.一种控制杂草的方法,其特征在于该方法包括以下步骤:1)在田地中种植对hppd抑制剂型除草剂(例如,异噁唑草酮、苯唑草酮、或硝磺草酮)具有耐受性的大豆植物或种子,和2)在所述田地中在种植所述植物或种子之前施用或在种植后(可以是种子萌发之前或之后)在所述大豆植物或种子上施用hppd抑制剂型除草剂(例如,异噁唑草酮、苯唑草酮、或硝磺草酮),其中所述植物或种子在其基因组中包含大豆原种转化事件ee-gm5,包含所述原种事件的参考种子在atcc处保藏号pta-123625下保藏。
97.一种用于控制杂草的方法,特征在于该方法包括以下步骤:1)在种植大豆植物或播种大豆种子之前,用hppd抑制剂型除草剂(例如,异噁唑草酮、苯唑草酮、或硝磺草酮)处理待种植所述大豆植物的田地或待播种所述大豆种子的田地,和2)在所述预处理的田地中,种植包含大豆原种转化事件ee-gm5的大豆植物或播种包含大豆原种转化事件ee-gm5的大豆种子,其中包含所述大豆原种转化事件ee-gm5的参考种子在atcc处保藏号pta-123625下保藏。
98.一种用于在待种植大豆植物的田地中减少产量损失的方法,所述田地特别是包含或预期包含线虫(如scn、rkn或短体线虫属或肾形线虫或其组合)的田地,该方法包括以下步骤:1)获得包含如上所述原种转化事件ee-gm5的植物或种子,和2)种植或播种大豆植物或种子,其中包含所述原种事件的参考种子在atcc处保藏号pta-123625下保藏。
99.一种用于增加大豆植物的产量的方法,其中所述大豆植物种植在包含线虫例如scn、rkn、或短体线虫属或肾形线虫或其组合的田地中,该方法包括以下步骤:1)获得包含如上所述的原种转化事件ee-gm5的植物或种子,和2)种植或播种大豆植物或种子,其中包含所述原种事件的参考种子在atcc处保藏号pta-123625下保藏。
100.一种方法,该方法用于生产对hppd抑制剂型除草剂例如异噁唑草酮、苯唑草酮或硝磺草酮具有耐受性的大豆植物或种子;或用于生产对线虫例如scn、rkn、或短体线虫属或肾形线虫具有耐受性的大豆植物或种子;或用于生产对hppd抑制剂型除草剂例如异噁唑草酮、苯唑草酮或硝磺草酮具有耐受性且对线虫例如scn、rkn、或短体线虫属或肾形线虫具有耐受性的大豆植物或种子,该方法特征在于以下步骤:将上面所述的原种大豆转化事件ee-gm5引入大豆植物或种子的基因组中,并且任选地用hppd抑制剂型除草剂例如异噁唑草酮、苯唑草酮或硝磺草酮处理所述植物或种子,或任选地用hppd抑制剂型除草剂例如异噁唑草酮、苯唑草酮或硝磺草酮处理将要种植所述植物或种子的田地,并且在所述预处理的田地中种植所述植物或种子。
101.一种特异地表征大豆原种转化事件ee-gm5的核酸分子,该核酸分子的特征在于包含seqidno.1、3或5中任一个的核苷酸序列,该核苷酸序列包含大豆植物基因组dna的部分以及位于其下游并与其邻接的ee-gm5的插入外源dna的部分;和/或特征在于包含seqidno.2、4、或6的核苷酸序列,该核苷酸序列包含ee-gm5的插入外源dna的部分以及位于其下游并与其邻接的大豆植物基因组dna的部分。
102.一种植物或种子,该植物或种子包含如上所述的ee-gm5并且还包含由大豆抗性基因座/基因提供的对scn、rkn或短体线虫属或肾形线虫或其组合的耐受性或抗性。
103.根据段落102所述的植物或种子,其中所述植物或种子包含ee-gm5、和以下的scn抗性等位基因/基因座中的任一者或其组合:pi548316、pi567305、pi437654、pi90763、pi404198b、pi88788、pi468916、pi567516c、pi209332、pi438489b、pi89772、peking、pi548402、pi404198a、pi561389b、pi629013、pi507471、pi633736、pi507354、pi404166、pi437655、pi467312、pi567328、pi22897、或pi494182。
104.一种包含如上所述ee-gm5的植物或种子,其还包含由除草剂耐受性基因(天然的或突变的大豆基因或转基因)提供的对其他除草剂的耐受性,例如对基于草甘膦、草铵膦、磺酰脲、咪唑啉酮、hppd抑制剂、麦草畏、2,4-d、或ppo抑制剂的除草剂、或其任何组合的耐受性。
105.根据段落103所述的植物或种子,其中所述植物或种子包含如上所述的ee-gm5和赋予除草剂耐受性的以下大豆转化事件中的一或多者:mst-
106.一种方法,所述方法用于在scn侵染存在下降低猝死综合征或缺铁性萎黄病对大豆植物的影响的严重性、或用于在猝死综合征侵染的包含scn的田地中或在导致大豆缺铁性萎黄病的包含scn的田地中增加大豆植物产量的方法,该方法包括种植包含原种事件ee-gm5的大豆植物或播种包含原种事件ee-gm5的大豆种子,其中包含所述原种事件的参考种子在atcc处保藏号pta-123625下保藏。
附图说明
以下实例(不旨在将本发明限制于所描述的具体实施方案)可以结合附图来理解,这些附图通过引用并入本文,其中:
图1:所示核苷酸序列和引物之间关系的示意图.
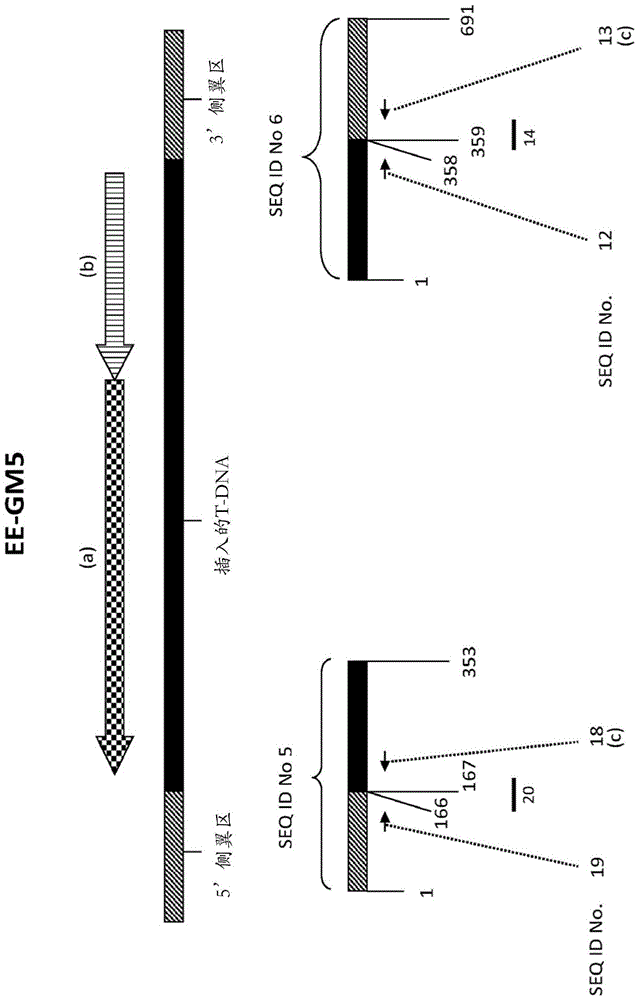
黑条:插入的t-dna;阴影条:位于t-dna侧翼的dna;方格箭头(a):嵌合cry14ab-1.b基因(该嵌合基因的组成参见表1);阴影箭头(b):嵌合hppdpf-4pa基因(该嵌合基因的组成参见表1);黑色箭头:寡核苷酸引物;(c)指所示核苷酸序列的互补序列;黑线:寡核苷酸探针(下面的数字是代表性的seqidno.)。表示seqidno.5和6的条下面的数字是所述序列中不同元件的核苷酸位置。注:该方案未按比例绘制。
图2:用于ee-gm5身份分析的终点方法.
图2显示了实施例2.1中描述的方法用于含有ee-gm5的一系列大豆样品和常规大豆样品的结果的例子。对于每个样品,显示了ee-gm5特异性反应和内源性反应的s/b比率。在该图中,标有“a”的线内的样品是不包含ee-gm5的大豆样品,标有“b”的线内的样品是包含ee-gm5的大豆样品,并且在标有“c”的线形成的框内的样品是不确定的样品。
图3:用于ee-gm5gm5身份和合子型(zygosity)分析的终点方法.
图3显示,对于一系列包含纯合状态的ee-gm5的大豆样品、包含半合子状态的ee-gm5的大豆样品和常规大豆样品,实施例2.2中描述的方法的结果的实例。在该图中,标有“a”的线内的样品是包含纯合状态的ee-gm5的大豆样品,标有“b”的线内的样品是包含半合子状态的ee-gm5的大豆样品,在标有“c”的线内的样品是不包含ee-gm5的大豆样品,并且在标有“d”的线形成的框内的样品是不确定的样品。图4:用于ee-gm5低水平存在分析的实时pcr方法
图4显示,对于在校准样品上进行的低水平存在分析,实施例2.4中描述的rt-pcr方法的结果的实例。“a”、“b”、“c”、“d”、“e”分别表示校准样品“a”、“b”、“c”、“d”、“e”的ct值。校准样品“a”、“b”、“c”、“d”、“e”具有减少量的ee-gm5dna。
图5:除草剂处理的平均最大植物结果
图5显示,与未经转化的/常规的大豆植物(thorne)相比,对于包含事件ee-gm5的大豆植物,在2年的几个田间试验中记录的除草剂处理的最大植物毒性数据的平均值。处理下方在()中的数字给出了条中包含的试验数,每个条顶部的数字给出针对该处理的平均最大植物毒性值。施用的处理是:ift=异噁唑草酮,mst=硝磺草酮,pe=出苗前,po=出苗后(在v2-v3阶段,其中添加助剂作物油浓缩物和硫酸铵以增加除草剂活性)。显示的比率为克活性成分/公顷(出苗前4x剂量,出苗后2x剂量)。
图6.在scn侵染的田地中thorne的ee-gm5的籽粒产量.
在2015和2016年,在爱荷华州,伊利诺伊州,印第安纳州,密苏里州和田纳西州的9个不同位置,在scn侵染的田地(从低到高scn侵染)中测试原始转化体背景(thorne)中的ee-gm5。点是每个试验的纯合子事件的估计产量(与无效分离子(nullsegregant)的差异百分比),水平线代表纯合子事件和无效分离子之间对比的95%置信限(如果该线不与无效分离子的100%产率处的垂直线重叠,则该事件与无效分离子显著不同)。“acrosslocs”是所有9个位置的综合分析的估计产量。图7.在scn侵染的田地中在scn易感原种背景中ee-gm5的籽粒产量.
将ee-gm5渐渗入(bc2f3)易感scn的原种mgi(成熟组i)系中,并且在2016年在明尼苏达州的一个位置和北达科他州的一个位置(每个位置具有高scn侵染水平)处进行测试。点是每个试验的纯合子事件的估计产量(与无效分离子的差异百分比),点周围的水平线代表纯合子事件和无效分离子之间对比的95%置信限(如果该线不与无效分离子的100%产率处的垂直线重叠,则该事件与无效分离子显著不同)。“acrosslocs”是两个位置的综合分析的估计产量。
图8.scn侵染的田地中原种scn抗性背景中ee-gm5的籽粒产量.
将ee-gm5杂交到对scn具有抗性(由于来自pi88788的rhg1基因座)的原种mgiii(成熟组iii)系中,并且在2016年在3个位置进行测试(以“16”,例如16-in1开始的试验)并且在2017年在7个位置(以“17”,例如17-in1开始的试验)进行测试,scn侵染水平从低到高(参见箭头,具有低scn压力的位置在图的底部(例如,17-il2),具有高scn压力的位置在顶部(例如,17-in1))。通过考虑以下若干因素来确定scn压力,所述因素包括已知的田地历史、土壤中的scn群体、抗性和易感对照品种的相对产量、土壤特性(ph和%砂),以及易感条目中根侵染的视觉评估。点是与无效分离子相比,每个试验中纯合子事件的平均产量差异(以吨/公顷表示),点周围的水平线表示纯合子事件和无效分离子之间对比的95%置信限(如果相对于无效分离子的差异为0的垂直线不与该线重叠,那么该事件的产量与无效分离子显著不同)。“avg”是每年所有位置的平均产量。
图9.在美国进行的短体线虫属抗性温室试验
在美国温室试验中具有ee-gm5的原种大豆植物控制最短尾短体线虫。将具有ee-gm5的植物(”ee-gm5”)与其他原种大豆系比较:一个scn易感成熟组(mg)3系(“thorne”)、一个mg3scn易感系、一个mg6.2scn易感系和一个mg9scn易感系(“suscwt”显示这3个系的平均值)、一个mg3scn抗性系(具有来自pi88788的rhg1抗性等位基因,“scnres(pi88788)”)、以及一个mg6.2scn抗性系(具有来自peking的rhg1和rhg4scn抗性)(“scnres(peking)”)。绘图在侵染后30天根中短体线虫属线虫的平均数量(5株植物/条目),还显示了在各品种间观察到的变异(如温室试验中常见的)。结果显示ee-gm5系中短体线虫属线虫的约90%控制。具有天然scn抗性的大豆系(来自peking或pi88788)不控制最短尾短体线虫。图10.在巴西进行的短体线虫属抗性温室试验
ee-gm5纯合的大豆植物(“ee-gm5hh”)显著减少大豆根中的最短尾短体线虫。从巴西的当地田地中分离出最短尾短体线虫。在巴西的温室试验中针对短体线虫属控制评估以下各项:ee-gm5植物(在两个不同的us原种系中(两者均是成熟组6.2,一个scn易感和一个具有pekingscn抗性(“ee-gm5”))和五个巴西大豆系(具有有限的短体线虫属线虫控制(“巴西系”)),标记为短体线虫低rf(繁殖因子)的一个巴西系(“brs7380(低rf)”),scn-易感的一个us原种系(成熟组6.2)(“scnsusc”),和具有pekingscn-抗性的一个us原种系(mg6.2)(“scnres(peking)”)。绘制这些条目的平均值,也显示了各品种间观察到的变异(如通常见于温室试验的)。一个巴西大豆系(brs7380)显示短体线虫属线虫的约89%减少。ee-gm5系给出短体线虫属线虫的约99%控制。携带对scn的peking天然抗性的大豆系不控制最短尾短体线虫。
图11.与无效分离子相比,ee-gm5植物的缺铁性萎黄病(idc)得分
图11显示在一个位置(具有高scn侵染)处具有ee-gm5的大豆植物的idc得分。该试验是裂区(split-plot)设计(4个地块/条目),观察事件在3个不同背景(2个易感性大豆系和1个具有来自pi88788的scn抗性)中的影响。显示了,跨越三个遗传背景(1个scn抗性,1个scn易感性、和scn易感性thorne背景),具有事件ee-gm5的植物(“ee-gm5”)和相应无效分离子(“无效”,缺少ee-gm5)的idc得分的平均值。一个条形代表总共12个地块。垂直线表示标准误(“sem”是平均值的标准误)。
本发明优选实施方案的详细说明
在本发明中,已在线虫抗性大豆(glycinemax)的开发中,自一系列转基因大豆植物中,将ee-gm5鉴定为原种事件,该事件包含编码4-羟基苯基丙酮酸双加氧酶(hppd)抑制剂耐受性的基因以及赋予线虫抗性的基因(每种基因在植物-可表达的启动子的控制下)。本文描述了用于鉴定生物样品中的原种事件ee-gm5的特异工具。
在植物基因组中掺入重组dna分子通常由细胞或组织的转化产生。掺入的具体位点通常由随机整合所致。
通过用重组dna或“转化dna”转化植物细胞或组织而引入植物基因组的并且源自这种转化dna的dna,在下文中被称为包含一个或多个“转基因”的“插入t-dna”(insertedt-dna)。ee-gm5的转基因是线虫抗性和hppd抑制剂除草剂耐受性基因。在本发明的上下文中,“植物dna”将指,源自转化的植物的dna。植物dna通常存在于相应的野生型植物的相同遗传基因座中。该插入的t-dna可以通过在植物基因组中重组dna分子的掺入位点的位置和构型来表征。其中已插入了重组dna的植物基因组中的位点还被称为“插入位点”或“靶位点”。将重组dna插入称为“插入前植物dna”(或“插入前基因座”)的植物基因组区域,可以与植物dna的缺失(称为“靶位点缺失”)相关联。如本文所用的“侧翼区”或“侧翼序列”是指,与引入的t-dna不同的至少10bp、至少20bp、至少50bp、和高达5000bp的dna的序列,优选来自植物基因组的dna,该dna位于插入t-dna的紧上游并与其邻接或位于插入t-dna的紧下游并与其邻接。导致插入t-dna随机整合的转化过程将导致具有不同侧翼区的转化体,这些侧翼区对于每个转化体是特征性的和独特的。当通过传统杂交将该重组dna引入植物中时,该dna在植物基因组中的插入位点或其侧翼区通常不会改变。
如本文所用的“分离的核酸(序列/分子)”或“分离的dna(序列/分子)”是指,该核酸或dna(序列/分子)不再处于其所分离自的天然环境中,例如,该核酸序列在另一(细菌)宿主或植物基因组中,或该核酸或dna(序列/分子)与来自另一来源的dna或核酸(序列/分子)融合,例如包含在嵌合基因中位于(异源)植物-可表达的启动子的控制下。本发明的任何核酸或dna(包括任何引物)还可以是非天然存在的,例如该核酸或dna可以与天然存在的序列具有相同序列,但具有标记(天然存在的对应物中缺失);或其序列与天然存在的核酸或dna或其片段相比具有至少一个核苷酸添加或取代或至少一个内部核苷酸缺失;或其序列与天然存在的核酸或dna或其片段相比具有低于100%的序列同一性(不相同);或该核酸或dna具有由不同来源的核苷酸序列组成的序列,其中这些核苷酸序列在自然界中不在一起出现(嵌合或杂合dna);或具有不同于天然核酸或dna或其片段的序列的人造合成核酸或dna。
事件被定义为(人工)遗传基因座,其作为基因工程的结果而携带包含一个或多个目的基因的至少一个拷贝的插入t-dna或转基因。事件的典型等位基因状态是插入t-dna的存在或不存在。可以通过该一个或多个转基因的表达,在表型上表征事件。在遗传水平上,事件是植物遗传构成的一部分。在分子水平上,事件可以通过限制性图谱(例如,通过dna印迹法确定)、通过转基因的上游和/或下游侧翼序列、分子标记的位置和/或转基因的分子构型来表征。通常,用包含至少一个目的基因的转化dna转化植物,将形成包含多个独立事件的一群转化体,其中每个事件都是独特的。事件可以通过插入的t-dna和至少一个侧翼序列来表征。
如本文所用,原种事件是,从使用相同转化dna转化而获得的一组事件中,基于最佳性状功效和优异表达、一个或多个转基因的稳定性、以及事件与包含其的植物的最佳农艺特征的相容性,而选择的事件。因此,原种事件选择的标准是以下中的一个或多个,优选两个或更多个,有利地是所有:
a)性状功效;
b)插入t-dna的存在不会损害植物的其他所希望的特征,例如与农艺性状或商业价值有关的特征;
c)事件可以通过十分明确的分子构型来表征,所述分子构型可以稳定遗传,并且针对该分子构型可以开发适当的工具用于身份控制;
d)目的基因(一个或多个)在一系列环境条件下以商业上可接受的水平显示正确、适当和稳定的空间和时间表型表达,其中所述环境条件为携带该事件的植物在正常的农艺应用中可能暴露的环境条件。
优选的是,插入的t-dna与植物基因组中的位置相关,所述位置允许插入的t-dna容易地渐渗到期望的商业遗传背景中。
通过在不同相关遗传背景中渐渗原种事件并观察对一个、两个、三个或所有标准(例如以上a)、b)、c)和d))的遵从性,来确认事件作为原种事件的状态。
因此“原种事件”是指,满足上述标准的、包含插入t-dna的遗传基因座。植物、植物材料或子代(例如种子)可以在其基因组中包含一个或多个不同的原种事件。
开发用于鉴定原种事件或包含原种事件的植物或植物材料或包含含有原种事件的植物材料的产品的工具,基于原种事件的特异性基因组特征,例如,包含插入t-dna、分子标记物、或插入t-dna的一个或多个侧翼区序列的基因组区域的特异性限制性图谱。
一旦对插入t-dna的侧翼区中的一个或两个进行了测序,则通过分子生物学技术可以开发引物和/或探针,这些引物和/或探针特异性地识别样品的核酸(dna或rna)中的该(这些)序列。例如,可以开发pcr方法以鉴定生物样品(例如植物样品、植物材料或包含植物材料的产品)中的原种事件。这种pcr基于至少两个特异性“引物”,其中一个引物识别原种事件的5’或3’t-dna侧翼区内的序列并且另一引物识别插入t-dna内的序列。这些引物优选地具有在15和35个核苷酸之间的序列,这些序列在优化的pcr条件下分别“特异性识别”原种事件的5’或3’t-dna侧翼区内的序列和原种事件的插入t-dna内的序列,由此从包含原种事件的核酸样品中扩增特异性片段(“整合片段”或区分性扩增子)。这意味着,在优化的pcr条件下仅扩增靶向的整合片段,而不扩增植物基因组中的其他序列或插入的t-dna。
适用于本发明的pcr引物可以如下:
-长度在17nt至约200nt范围内的寡核苷酸,其在3’末端包含选自5’t-dna侧翼序列(seqidno.5的核苷酸1至核苷酸166或seqidno.24的核苷酸1至核苷酸1113或位于其上游并与其邻接的植物基因组序列)的至少17个连续核苷酸(优选20个连续核苷酸)的核苷酸序列(识别5’t-dna侧翼序列的引物);或者
-长度在17nt至约200nt范围内的寡核苷酸,其在3’末端包含选自3’t-dna侧翼序列(seqidno.6的核苷酸359至核苷酸691的互补序列、或seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列、或位于其下游并与其邻接的植物基因组序列)的至少17个连续核苷酸(优选20个连续核苷酸)的核苷酸序列(识别3’t-dna侧翼序列的引物);或者
-长度在17nt至约200nt范围内的寡核苷酸,其在3’末端包含选自插入t-dna序列(seqidno.5的核苷酸167至核苷酸353的互补序列、或seqidno.6的核苷酸1至核苷酸358的序列、或seqidno.11的核苷酸1至核苷酸7459的序列、或seqidno.23的核苷酸1114至核苷酸8572的序列,或其互补序列)的至少17个连续核苷酸(优选20个连续核苷酸)的核苷酸序列(识别插入t-dna的引物)。
应当理解,可以将识别5’t-dna侧翼序列的引物连同识别插入t-dna的引物(这些引物可以选自seqidno.5的核苷酸167至核苷酸353的互补序列或位于其下游并与其邻接的t-dna序列)一起用于pcr反应,也可以将识别3’t-dna侧翼序列的引物连同识别插入t-dna的引物(这些引物可以选自seqidno.6的核苷酸1至核苷酸358的序列或位于其上游并与其邻接的t-dna)一起用于pcr反应。识别插入t-dna的引物还可以选自seqidno.11的核苷酸1至核苷酸7459的序列,或seqidno.23的核苷酸1114至核苷酸8572的序列,或其互补序列。
这些引物当然可以比所提及的17个连续核苷酸更长,并且可以是例如,20,21、30、35、50、75、100、150、200nt长或甚至更长。这些引物可以完全由核苷酸序列组成,该核苷酸序列选自所提及的侧翼序列的核苷酸序列和插入t-dna的序列。然而,这些引物在其5’末端的核苷酸序列(即,3’末端的17个连续核苷酸之外)不太重要。因此,视情况而定,这些引物的5’序列可以包含选自侧翼序列或插入t-dna的核苷酸序列,或由其组成,但是与t-dna或t-dna侧翼dna相比可以包含几个(例如,1、2、5或10个)错配。这些引物的5’序列甚至可以完全是与侧翼序列或插入t-dna无关的核苷酸序列,例如一个或多个限制酶识别位点的核苷酸序列,或例如能够结合其他寡核苷酸,例如标记的寡核苷酸,例如fret盒(lgc基因组学;参见semagn等人.,2014,molbreeding[分子育种]33:1-14,和us7615620)的核苷酸序列。此类无关的序列或具有错配的侧翼dna序列应优选不长于100,更优选不长于50或甚至25个核苷酸。这些引物还可以用标记(例如荧光标记)修饰。
此外,合适的引物可以在其3’末端处包含跨5’或3’t-dna侧翼区来源序列和插入t-dna序列之间的连接区域(位于seqidno.5中的核苷酸166和167、和seqidno.6中的核苷酸358和359、或seqidno.24中的核苷酸1113和1114、以及seqidno.25中的核苷酸358和359处)的核苷酸序列,或(基本上)由其组成,条件是所提及的3’位置处的17个连续核苷酸非唯一地来源于seqidno.5或6或seqidno.24或25中的插入t-dna或t-dna侧翼序列。
本领域技术人员还将立即清楚,适当选择的pcr引物对也不应包含彼此互补的序列。
出于本发明的目的,“seqidno:x中所示的核苷酸序列的互补序列”是可以根据chargaff的规则
适合的引物的实例是seqidno.13或seqidno.19或seqidno.26或27(识别3’或5’t-dna侧翼序列的引物),或seqidno.12或seqidno.18或seqidno.28或29(识别插入t-dna的引物,与识别3’或5’t-dna侧翼序列的引物一起使用)的寡核苷酸序列。
优选地,扩增的片段具有在50和500个核苷酸之间的长度,例如在50和150个核苷酸之间的长度。这些特异性引物可以具有分别与原种事件的5’或3’t-dna侧翼区内和原种事件的插入t-dna内的序列具有80%和100%之间同一性的序列,条件是错配仍然允许在优化的pcr条件下用这些引物特异性鉴定原种事件。然而,可允许的错配的范围可以容易地通过实验确定并且是本领域技术人员已知的。
整合片段的检测可以按不同方式进行,例如,通过凝胶分析后的大小估计。还可以对整合片段直接测序。用于检测扩增的dna片段的其他序列特异性方法在本领域也是已知的。还可以使用经标记的序列和对标记进行检测来检测扩增的dna片段。例如,经标记的探针可以包含在反应混合物中,该探针特异性结合扩增的片段。在一个实施方案中,经标记的探针(fret杂交探针)可以包含荧光标记和猝灭剂,由此fret盒当与pcr产物结合时不再被猝灭并且发射荧光。可替代地,经标记的fret盒,即用荧光标记和猝灭剂标记的寡核苷酸,可以包括在反应混合物中,特异性结合反应混合物中的引物之一,例如针对反应混合物中使用的引物的5'延伸的fret盒(参见,例如,semagn等人.,2014,molbreeding[分子育种]33:1-14,和us7615620)。使用本领域已知的方法可以测量荧光。可以实时(即在pcr反应的每个循环期间)测量荧光。也可以在pcr反应结束时测量荧光。
由于引物的序列及其在基因组中的相对位置对于原种事件是独特的,因此整合片段的扩增将仅在包含原种事件(的核酸)的生物样品中发生。优选地,当进行pcr以鉴定未知样品中ee-gm5的存在时,包括一组引物作为对照,利用该对照可以扩增该事件的植物物种的“持家基因”内的片段。持家基因是在大多数细胞类型中表达的基因,并且涉及所有细胞共有的基本代谢活动。优选地,从持家基因扩增的片段是比扩增的整合片段更大的片段。根据待分析的样品,可以包括其他对照。
在本领域中描述了标准pcr方案,例如在“pcrapplicationsmanual[pcr应用手册]”(rochemolecularbiochemicals[罗氏分子生物化学],第二版,1999,或第3版,2006)和其他参考文献。pcr的最佳条件(包括特异性引物的序列)在每个原种事件的“pcr(或聚合酶链式反应)鉴定方案”中可以被规定。然而,应理解,pcr鉴定方案中的许多参数可能需要针对特定的实验室条件调整,并且可以稍做修改以获得类似的结果。例如,使用不同的制备dna的方法时,可能需要调整,例如,引物的量、聚合酶和使用的退火条件。类似地,其他引物的选择可能决定pcr鉴定方案的其他最佳条件。然而,这些调整对于本领域技术人员而言是显而易见的,并且在当前的pcr应用手册中进一步详述,例如上面引用的那些文献。
可替代地,可以将特异性引物用于扩增整合片段,该整合片段可以被用作“特异性探针”用于鉴定生物样品中的ee-gm5。在允许探针与核酸中的相应片段杂交的条件下,使生物样品的核酸与探针接触,可以导致核酸/探针杂交体的形成。可以检测该杂交体的形成(例如,通过标记核酸或探针),由此该杂交体的形成表明ee-gm5的存在。基于与特异性探针杂交的这种鉴定方法(在固相载体上或在溶液中)已在本领域中进行了描述。该特异性探针优选地是序列,该序列在优化的条件下与包含原种事件的5’或3’t-dna侧翼区的部分和与其邻接的插入t-dna的部分的区域(下文被称为“特异区域”)特异性地杂交。优选地,特异性探针包含在50和500bp之间的序列,或100至350bp的序列,该序列与ee-gm5的特异区域的核苷酸序列至少80%、或在80%和85%之间、或在85%和90%之间、或在90%和95%之间、或在95%和100%之间相同(或互补),或与该特异区域的核苷酸序列相同(或互补)。优选地,特异性探针包含与原种事件的特异区域相同(或互补)的约15至约100个连续核苷酸的序列。
适合作为检测原种事件ee-gm5的pcr引物的寡核苷酸也可以用于开发基于pcr的方案以确定包含原种事件的植物的合子状态。为此,识别整合之前的野生型基因座的两个引物以这样的方式设计,即它们彼此相向并且插入位点位于这两个引物之间。这些引物可以包含特异性识别ee-gm5的5’和/或3’t-dna侧翼序列的引物。识别整合前的野生型基因座的这组引物,连同与转化dna序列(插入t-dna)互补的第三引物,可以允许ee-gm5特异性基因座和野生型基因座的同时诊断性pcr扩增。如果植物在转基因基因座或相应的野生型基因座上是纯合的,则该诊断性pcr将产生对于转基因或野生型基因座而言典型的单个pcr产物(优选,长度典型的)。如果植物在转基因基因座上是半合子的,将出现两个基因座-特异的pcr产物,反映了转基因和野生型基因座的扩增。
可替代地,为确定包含原种事件的植物的合子状态,识别整合前的野生型基因座的两个引物以这样的方式设计,即这两个引物彼此相对,并且其中一个引物特异性识别包含在seqidno.5或6或包含在seqidno.24或25中的5’或3’t-dna侧翼序列,并且一个引物特异性识别包含在seqidno.6或5或seqidno.24或25中的3’或5’t-dna侧翼序列,或特异性识别插入前基因座。对于本发明,识别整合之前的野生型基因座的合适引物对是包含以下引物的引物对:包含seqidno.21的核苷酸序列或(基本上)由其组成的一个引物,以及包含seqidno.19的核苷酸序列或(基本上)由其组成的一个引物。这组引物,连同与转化dna序列(插入t-dna)互补的第三引物,或与转化dna序列和与其邻接的5’或3’t-dna侧翼序列互补的第三引物,其中第三引物的朝向与特异性识别5’或3’t-dna侧翼序列的引物的朝向相对(例如,包含seqidno.18的核苷酸序列或(基本上)由其组成的引物,与包含seqidno.19的核苷酸序列或(基本上)由其组成的引物,方向相对),由此允许ee-gm5特异性基因座和野生型基因座的同时诊断性pcr扩增。如果植物对于转基因基因座或相应的野生型基因座是纯合的,则诊断性pcr将产生对于转基因或野生型基因座而言典型的单个pcr产物。如果植物对于转基因基因座是半合子的,将出现两个基因座-特异的pcr产物,反映了转基因和野生型基因座两者的扩增。
对于野生型和转基因基因座典型的pcr产物的检测,可以基于pcr产物的长度来确定,所述长度对于野生型和转基因基因座可以是典型的。可替代地,可以通过修饰对插入前基因座具有特异性的引物并通过修饰对插入t-dna具有特异性的引物,以及检查经修饰的引物向pcr产物中的掺入,来检测野生型基因座所典型的以及转基因基因座所典型的pcr产物。例如,可以使用荧光标记对插入前基因座的特异性引物和插入t-dna的特异性引物进行标记,其中两个引物的标记是不同的。当引物掺入pcr产物时,可以检测到荧光。如果植物对于转基因基因座或相应的野生型基因座是纯合的,则可以仅检测到对插入t-dna具有特异性的引物的标记的荧光、或仅检测到对插入前基因座具有特异性的引物的标记的荧光。如果植物对于转基因基因座是半合子的,则可以检测到对插入t-dna具有特异性的引物的标记的荧光和对插入前基因座具有特异性的引物的标记的荧光二者,反映了转基因和野生型基因座二者的扩增。
可替代地,对插入前基因座具有特异性的引物和对插入t-dna具有特异性的引物可以具有5’延伸,该延伸特异性地结合经标记的fret盒,即用荧光标记和猝灭剂标记的寡核苷酸,其中针对两个引物的该5’延伸和相应的fret盒是不同的(参见,例如,semagn等人.,2014,molbreeding[分子育种]33:1-14,和us7615620)。当引物掺入到pcr产物中并随后将fret盒掺入pcr产物中后,可以检测荧光。如果植物对于转基因基因座或相应的野生型基因座是纯合的,则可以仅检测到与插入t-dna特异性引物特异性结合的fret盒的荧光、或仅检测到与插入前基因座特异性引物特异性结合的fret盒的荧光。如果植物对于转基因基因座是半合子的,则可以检测到与插入t-dna特异性引物特异性结合的fret盒的荧光和与对插入前基因座特异性引物特异性地结合的fret盒的荧光两者,反映了转基因和野生型基因座二者的扩增。
如果植物对于转基因基因座或相应的野生型基因座是纯合的,则诊断性pcr将产生对于转基因或野生型基因座而言典型的单个pcr产物(优选,长度典型的)。如果植物对于转基因基因座是半合子的,将出现两个基因座-特异的pcr产物,反映了转基因和野生型基因座的扩增。
可替代地,为确定包含原种事件的植物的合子状态,可以如实施例中所述以定量的方式在pcr反应中确定事件的存在。为此,以这样的方式设计识别事件ee-gm5的两个引物,使得这两个引物彼此相对,其中一个引物特异性识别包含在seqidno.5或6中或seqidno.24或25中的5’或3’t-dna侧翼序列,并且其中一个引物特异性识别seqidno.5或6中或seqidno.24或25中或seqidno.11或23中的插入t-dna。这组引物允许ee-gm5特异性基因座的pcr扩增。可以使用经标记的探针定量检测扩增的dna片段,该经标记的探针包含在反应混合物中,特异性结合扩增片段。经标记的探针可以包含荧光标记和猝灭剂,由此当与pcr产物结合时标记不再被猝灭并且发出荧光。可以使用本领域已知的方法实时测量荧光,即在pcr反应的每个循环期间测量。荧光超过某一阈值水平时的pcr循环数是分析的生物样品中ee-gm5特异性基因座的量的量度,并且可以基于纯合和杂合参考样品计算合子状态。
可替代地,还可以使用taqman化学和实时pcr原理,基于拷贝数分析,确定包含ee-gm5的植物的合子状态。可替代的方法通常包括ee-gm5特异性反应以量化ee-gm5拷贝数,以及内源基因特异性反应以标准化ee-gm5拷贝数。包含纯合状态的ee-gm5事件的样品具有比半合子样品高两倍的相对拷贝数。在这种方法中,无合子(azygous)的样品不会扩增ee-gm5序列。
此外,还可以使用本文提供的原种事件特异性序列信息,开发不同于基于pcr的扩增方法的原种事件ee-gm5特异性检测方法。此类可替代的检测方法包括基于特定核酸结构的侵入性切割的线性信号放大检测方法,也称为invadertm技术(如描述于例如美国专利5,985,557“invasivecleavageofnucleicacids[核酸的侵入性切割]”,6,001,567“detectionofnucleicacidsequencesbyinvaderdirectedcleavage[通过侵入者引导的切割检测核酸]”中,通过引用并于本文中)。为此,将靶序列与经标记的第一核酸寡核苷酸杂交,该第一核酸寡核苷酸包含从seqidno.5的核苷酸位置167至核苷酸位置184的核苷酸序列或其互补序列,或包含从seqidno.6的核苷酸位置341至核苷酸位置358的核苷酸序列或其互补序列;并且所述靶序列与第二核酸寡核苷酸进一步杂交,该第二核酸寡核苷酸包含从seqidno.5的核苷酸149至核苷酸166的核苷酸序列或其互补序列,或所述核酸寡核苷酸包含从seqidno.6的核苷酸359至核苷酸376的核苷酸序列或其互补序列,其中所述第一和第二寡核苷酸重叠至少一个核苷酸。
通过该杂交产生的双链体或三链体结构允许用酶
在一个实施方案中,提供了通过与基本上互补的经标记的核酸探针杂交来检测生物样品中原种事件ee-gm5的存在的方法,其中通过再循环靶核酸序列来扩大探针:靶核酸比率,所述方法包括:
a)将所述靶核酸序列与第一核酸寡核苷酸杂交,该第一核酸寡核苷酸包含从seqidno.5的核苷酸位置167至核苷酸位置184的核苷酸序列或其互补序列,或所述第一核酸寡核苷酸包含从seqidno.6的核苷酸位置341至核苷酸位置358的核苷酸序列或其互补序列;
b)将所述靶核酸序列与第二核酸寡核苷酸杂交,该第二核酸寡核苷酸包含从seqidno.5的核苷酸149至核苷酸166的核苷酸序列或其互补序列,或所述第二核酸寡核苷酸包含从seqidno.6的核苷酸359至核苷酸376的核苷酸序列或其互补序列,其中所述第一和第二寡核苷酸重叠至少一个核苷酸,并且其中所述第一或所述第二寡核苷酸被标记形成所述经标记的核酸探针;
c)用酶仅切割探针:靶核酸序列双链体内经标记的探针,该酶引起选择性探针切割,导致双链体解离,留下该靶序列保持完整;
d)通过重复步骤(a)至(c),再循环该靶核酸序列;以及
e)检测切割的经标记的探针,从而确定所述靶核酸序列的存在,并检测所述生物样品中原种事件ee-gm5的存在。
当两个核酸不是彼此的完整互补序列(如本文所定义)时,例如当它们的序列为至少80%、至少85%、至少90%、至少95%、或至少99%彼此互补时,这两个核酸是如本文所用的“基本上互补的”。
如本文所用的“试剂盒”是指用于实施本发明方法(更具体地,生物样品中原种事件ee-gm5的鉴定,或包含ee-gm5的植物材料的合子状态的确定)的一组试剂。更具体地,本发明的试剂盒的优选实施方案包含至少一个或两个特异性引物,如上所述用于鉴定原种事件,或包含三个特异性引物,或两个特异性引物和一个特异性探针,如上所述用于确定合子状态。任选地,该试剂盒可以进一步包含本文在pcr鉴定方案或用于ee-gm5检测的任何其他方案中描述的任何其他试剂。可替代地,根据本发明的另一个实施方案,试剂盒可以包含如上所述的特异性探针,该探针与生物样品的核酸特异性杂交以鉴定其中ee-gm5的存在。任选地,该试剂盒可以进一步包含任何其他试剂(例如但不限于杂交缓冲液,标记物),用于使用特异性探针鉴定生物样品中的ee-gm5。
可以使用本发明的试剂盒,并且可以出于以下目的特定地调整其组分:质量控制(例如,种子批的纯度),在植物材料中或在包含植物材料或由其衍生的材料(例如但不限于食品或饲料或工业产品)中检测原种事件的存在或缺失。
如本文所用的,关于核苷酸序列(dna或rna)的“序列同一性”是指,具有相同核苷酸的位置数除以两个序列中较短序列的核苷酸数。可以使用窗口大小为20个核苷酸、字长为4个核苷酸、和缺口罚分为4,通过wilbur和lipmann算法进行两个核苷酸序列的比对(wilbur和lipmann,1983,proc.nat.acad.sci.usa[美国国家科学院院刊]80:726)。计算机辅助分析和序列数据的解释,包括如上所述的序列比对,可以例如使用遗传学计算机集团(geneticscomputergroup)(gcg,威斯康星大学生物技术中心(universityofwisconsinbiotechnologycenter))的序列分析软件包,方便地进行。当此类序列具有至少约75%、至少约80%、至少约85%、至少约90%、至少约95%、至少约98%、或至少约99%、或至少95%、至少96%、至少97%、至少98%、至少99%、至少99,5%或至少99,9%的序列同一性时,序列被表示为“基本上相似”。很明显的是,当rna序列被认为与dna序列基本相似或具有一定程度的序列同一性时,dna序列中的胸苷(t)被认为与rna序列中的尿苷(u)等同。而且,明显的是,随着时间在dna序列中可以出现小的差异或突变,并且对于本发明的事件-特异性引物或探针可以允许一些错配,因此在本发明的任何实施方案中,针对任何3’或5’t-dna侧翼dna或针对任何插入物或插入t-dna或本发明的任何引物或探针,在本文中给出的任何dna序列也包括与本文所提供的序列基本上相似的序列,例如与针对本发明的任何3’或5’t-dna侧翼dna、针对任何引物或探针、或针对任何插入物或插入t-dna所给出的序列杂交的序列、或与其具有至少90%、95%、96%、97%、98%、或至少99%序列同一性的序列,例如与任何给定的序列相差1至200、1至150、1至100、1至75、1至50、1至30、1至20、1至10、1至5、或1至3个核苷酸的核苷酸序列。
如本文所用的术语“引物”包括能够在模板依赖性过程(例如pcr)中引发新生核酸合成的任何核酸。通常,引物是从10至30个核苷酸的寡核苷酸,但可以使用更长的序列。引物可以按双链形式提供,但优选单链形式。探针可被用作引物,但被设计用于结合靶dna或rna,并且在扩增过程中不需要使用。
当提及特异性引物或探针时,如本文所用的术语“识别”是指特异性引物或探针在该方法中列出的条件(例如pcr鉴定方案的条件)下与原种事件中的核酸序列特异性杂交的事实,其中通过阳性和阴性对照的存在来确定特异性。
当提及特异性探针时,如本文所用的术语“杂交”是指在标准严格条件下该探针与原种事件核酸序列中的特异区域结合的事实。如本文所用的标准严格条件是指本文所述用于杂交的条件或如sambrook等人.,1989(molecularcloning:alaboratorymanual[分子克隆:实验室手册],第二版,冷泉港实验室出版社(coldspringharborlaboratorypress),纽约)所述的常规杂交条件,这些条件例如可以包括以下步骤:1)将植物基因组dna片段固定在滤膜上;2)在42℃下在50%甲酰胺、5xsspe、2x登哈特试剂(denhardt’sreagent)和0.1%sds中将滤膜预杂交1至2小时,或在68℃下在6xssc、2x登哈特试剂和0.1%sds中将滤膜杂交1至2小时;3)添加已标记的杂交探针;4)孵育16至24小时;5)在室温下在1xssc,0.1%sds中将滤膜洗涤20分钟;6)将滤膜洗涤三次20分钟,每次在68℃下在0.2xssc,0.1%sds中进行;和7)在-70℃下用增感屏将滤膜暴露于x射线胶片24至48小时。
如本文所用,生物样品是植物、植物材料或包含植物材料的产品的样品。术语“植物”旨在包括在任何成熟阶段的大豆(glycinemax)植物组织,以及取自或衍生自任何此类植物的任何细胞、组织或器官,包括但不限于任何种子、叶、茎、花、根、单细胞、配子、细胞培养物、组织培养物或原生质体。如本文所用,“植物材料”是指获得自或衍生自植物的材料。包含植物材料的产品涉及食品,饲料,或使用植物材料生产或可被植物材料污染的其他产品。应当理解,在本发明的上下文中,测试这些生物样品中是否存在ee-gm5特异性的核酸,这意味着样品中核酸的存在。因此,本文提及的用于鉴定生物样品中原种事件ee-gm5的方法,涉及在生物样品中鉴定包含原种事件的核酸。
如本文使用的,“包含”应被解释为指,存在所提及的特征、整数、步骤、试剂或组分,但不排除存在或添加一个或多个特征、整数、步骤或组分或其组。因此,例如,包含核苷酸或氨基酸序列的核酸或蛋白质可以包含比实际所示的核苷酸或氨基酸更多的核苷酸或氨基酸,即嵌入在更大的核酸或蛋白质中。包含功能上或结构上确定的dna序列的嵌合基因可以包含另外的dna序列,例如启动子、前导序列、尾随序列(trailer)和/或转录终止序列(可以也包括编码靶向肽或转运肽的dna)。
本发明还涉及在包含原种事件ee-gm5的大豆植物、获得自这些植物的包含原种事件ee-gm5的子代植物和种子中开发原种事件ee-gm5,并且涉及自包含该事件的植物衍生的植物细胞或植物材料。包含原种事件ee-gm5的植物可如实施例所述获得。本发明还涉及在atcc以保藏号pta-123625保藏的包含原种事件ee-gm5的种子、或包含原种事件ee-gm5的来自该种子的衍生物。如本文所用的“(种子的)衍生物”是指可以从该种子生长的植物;由自交、杂交或回交产生的子代;以及其植物细胞、器官、部分、组织、细胞培养物、原生质体和植物材料。
根据如实施例所述的ee-gm5鉴定方案中的任一种,可以鉴定包含ee-gm5的大豆植物或植物材料,这些鉴定方案包括实施例2.1中用于ee-gm5身份分析的终点方法;如实施例2.2所述用于ee-gm5身份和合子型分析的终点方法;如实施例2.3所述用于ee-gm5低水平存在分析的实时pcr方法;或如实施例2.4所述用于ee-gm5低水平存在分析的实时pcr。简言之,通过pcr使用以下引物扩增存在于生物样品中的大豆基因组dna:特异性识别ee-gm5的5’或3’t-dna侧翼序列中序列的引物,例如具有seqidno:13或seqidno.19的序列的引物,和识别插入t-dna中序列的引物,例如具有seqidno.12或seqidno.18的序列的引物,或识别ee-gm5的5’或3’t-dna侧翼序列和与其邻接的插入t-dna的引物。可以将扩增内源性大豆序列的一部分的dna引物用作pcr扩增的阳性对照。如果在pcr扩增后,该材料产生预期大小的片段或产生预期荧光标记的荧光,则该材料包含来自携带原种事件ee-gm5的大豆植物的植物材料。
携带ee-gm5的植物的特征在于它们的线虫抗性(特别是scn、腐线虫和/或根结线虫(“rkn”)和/或肾形线虫抗性)以及它们对hppd抑制剂(异噁唑草酮、苯唑草酮、或硝磺草酮)的耐受性。携带ee-gm5的不同可商购品种的大豆植物的特征还在于,在不存在hppd抑制剂除草剂施用和scn侵染的情况下,具有与相应的非转基因等基因可商购品种相当的农艺特征。已经观察到,在本文所述的大豆植物基因组的插入区域中存在插入t-dna,可以对包含该事件的植物赋予特别令人感兴趣的表型特征和分子特征。
本文还提供了用于生产对scn具有抗性和对hppd抑制剂型除草剂具有耐受性的大豆植物的方法,该方法包括,通过将缺少cry14ab-1-编码基因和缺少hppd-4-编码基因的第一大豆植物与包含ee-gm5的大豆植物杂交,并且选择对scn具有抗性和/或对hppd抑制剂除草剂具有耐受性的子代植物,从而将对scn的抗性和对hppd抑制剂除草剂的耐受性引入大豆植物的基因组中。可以使用标准scn温室测定法测量对scn的抗性,例如
www.plantpath.iastate.edu/tylkalab/greenhouse-resistance-screening和
www.plantmanagementnetwork.org/pub/php/review/2009/sce08/。
本发明的一个实施方案提供了大豆植物中的原种事件,该原种事件可通过在大豆基因组的特定位置处插入2个转基因而获得,该原种事件在此类大豆植物上赋予对线虫的抗性和对hppd抑制剂型除草剂(异噁唑草酮、苯唑草酮、或硝磺草酮)的耐受性,并且其中该原种事件与等基因系(如本文所用的,“等基因系”或“近等基因系”是具有相同遗传背景但缺乏转基因的大豆系,例如与用于转化的植物具有相同遗传背景的植物,或失去转基因的分离姐妹系(“无效分离子”)具有基本上相似的农艺性能。特别地,本发明在大豆植物中提供了原种事件,其中,与等基因系或商业大豆栽培种相比,在此类大豆植物的基因组中所述原种事件的插入或存在表现出不引起对疾病的易感性增加,不导致产量损失,或不导致倒伏增加。因此,本发明提供了大豆植物中的原种事件(称为ee-gm5),与等基因系相比,该原种事件导致大豆植物对线虫具有改善的抗性并且可以耐受hppd抑制剂型除草剂(异噁唑草酮、苯唑草酮、或硝磺草酮)的施用,而不负面影响所述大豆植物的产量,该大豆植物与等基因大豆植物或商业大豆栽培品种在其疾病易感性或倒伏方面无统计学显著差异。这些特征使得该原种事件成为在控制线虫和杂草抗性管理程序中有用的工具。在一个实施方案中,将事件ee-gm5与一个或多个大豆gm事件组合,所述大豆gm事件对基于草甘膦、基于草铵膦、基于hppd抑制剂、基于磺酰脲或咪唑啉酮的除草剂、以及ahas或als抑制型和/或生长素型(例如,麦草畏,2,4-d)的除草剂中的任一种或其组合提供耐受性,例如事件ee-gm3(akafg-072,mst-
本文还提供了包含事件ee-gm5的大豆植物或其部分,其中包含事件ee-gm5的代表性大豆种子已经在atcc保藏号pta-123625下保藏。本文进一步提供了包含这种事件的此类植物的种子,以及由此类种子产生的大豆产品,其中所述大豆产品包含事件ee-gm5。大豆产品可以是或可以包括大豆粕、磨碎的大豆籽粒、大豆片、或包含任何该加工的大豆产品的产品。特别地,大豆产品包含核酸,该核酸可以产生对事件ee-gm5具有诊断性或特异性的扩增子,所述扩增子包含seqidno.1或3或seqidno.2或4中任一个的序列。本文还提供了用于生产大豆产品的方法,该方法包括获得包含事件ee-gm5的大豆种子或籽粒,并由其生产这样的大豆产品。本文还提供了自大豆籽粒获得加工食品、饲料或工业产品的方法,例如大豆油、大豆蛋白、卵磷脂、豆浆、豆腐、人造黄油、生物柴油、生物复合材料、粘合剂、溶剂、润滑剂、清洁剂、泡沫、涂料、油墨、蜡烛、大豆油、或含大豆蛋白的食品或(动物)饲料产品,所述方法包括获得包含ee-gm5的籽粒并从所述籽粒生产所述加工食品、饲料或工业产品。在一个实施方案中,该方法还可以包括以下步骤:获得包含事件ee-gm5的大豆种子或植物,使所述种子或植物在田地中生长,以及收获大豆籽粒。任选地,该方法包括在种植前、出苗前、出苗后、或在包含ee-gm5的植物顶部,施用hppd抑制剂型除草剂(例如,ift、苯唑草酮、或硝磺草酮)。在一个实施方案中,上述大豆衍生的加工食品、饲料或工业产品包括在本发明中,例如使用本文所述的方法产生ee-gm5事件-特异性扩增子的加工产品,或包含seqidno.1、3或5,或seqidno.2、4、或6中任一个的核苷酸序列的加工产品。
本文还提供了大豆植物,该大豆植物是以上任何大豆植物的子代,并且包含事件ee-gm5,例如包含seqidno.1或3的序列或seqidno.2或4的序列的以上任何大豆植物的子代植物或种子;或包含seqidno.1或3的序列和seqidno.2或4的序列的以上任何大豆植物的子代植物或种子;或包含seqidno.5或seqidno.24的序列或seqidno.6或seqidno.25的序列的以上任何大豆植物的子代植物或种子;或包含seqidno.5或seqidno.24的序列和seqidno.6或seqidno.25的序列的以上任何大豆植物的子代植物或种子。
本文进一步提供了用于生产对线虫具有抗性和对异噁唑草酮和/或苯唑草酮和/或硝磺草酮除草剂具有耐受性的大豆植物的方法,该方法包括,特别地通过将缺少事件ee-gm5的第一大豆植物与包含ee-gm5的大豆植物杂交并且选择对线虫具有抗性和对异噁唑草酮和/或苯唑草酮和/或硝磺草酮除草剂具有耐受性的子代植物,将事件ee-gm5引入到所述植物的基因组中。
本文还提供了对线虫具有抗性和对异噁唑草酮、苯唑草酮、或硝磺草酮除草剂具有耐受性的、具有可接受的农艺特征的大豆植物,该大豆植物包含cry14ab-1-编码基因和hppd-4-编码基因,并且能够产生对事件ee-gm5具有诊断性的扩增子。本文还提供了该特异的分离扩增子(dna序列片段)本身,其可以使用本文所述的特异性检测工具获得,所述扩增子特别是在其序列中包括如下dna片段的扩增子,所述dna片段源自5’至3’t-dna侧翼dna和通过转化在植物基因组中插入的dna,如本文所定义的。
本文进一步提供了用于在包含事件ee-gm5的大豆植物的田地中、或待种植该大豆植物(其中在处理后将所述植物种植在所述田地中)的田地中控制杂草的方法,该方法包括用有效量的hppd抑制剂型除草剂(例如,基于异噁唑草酮、或基于苯唑草酮、或基于硝磺草酮的除草剂)处理田地,其中所述植物对该除草剂具有耐受性。
本文进一步提供了包含seqidno.5或6的序列或与其基本上相似的序列的dna,以及包含该dna序列的任何植物、细胞、组织、或种子(特别是大豆的任何植物、细胞、组织、或种子),例如包含ee-gm5的植物、细胞、组织、或种子。本文还包括任何大豆植物、细胞、组织、或种子,该大豆植物、细胞、组织、或种子包含seqidno.5或6的dna序列(所述dna序列对于常规大豆植物、种子、组织、或细胞是异源或外来的),或包含与seqidno.5或24或seqidno.6或25的序列具有至少99%或99.5%序列同一性的dna序列。
本发明还描述了嵌合dna,其包含:插入t-dna,其中所述插入t-dna的序列包含从seqidno.11的核苷酸1至核苷酸7459的序列、或从seqidno.23的核苷酸1114至核苷酸8572的序列、或与其具有至少97%、98%、99%、99,5%、或至少99,9%序列同一性的序列,并且侧翼为5’和3’t-dna侧翼区,其中位于所述插入t-dna紧上游并与其邻接的该5’t-dna侧翼区的特征在于以下序列,所述序列包含从seqidno.5的核苷酸1至核苷酸166的序列或从seqidno.24的核苷酸1至核苷酸1113的序列,并且其中位于所述插入t-dna紧下游并与其邻接的3’t-dna侧翼区的特征在于以下序列,所述序列包含从seqidno.6的核苷酸359至核苷酸691的序列,或从seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列。在一个实施方案中,所述插入t-dna的序列由seqidno.11的核苷酸1至核苷酸7459的序列、或seqidno.23的核苷酸1114至核苷酸8572的序列、或与其具有至少97%、98%、99%、99,5%、或至少99,9%序列同一性的序列组成,侧翼为5’和3’t-dna侧翼区的部分,其中位于所述插入t-dna紧上游并与其邻接的所述5’t-dna侧翼区的部分的特征在于以下序列,所述序列由seqidno.5的核苷酸1至核苷酸166的序列、或seqidno.24的核苷酸1至核苷酸1113的序列、或与其具有至少97%、98%、99%、99,5%、或至少99,9%序列同一性的序列组成,并且其中位于所述插入t-dna紧下游并与其邻接的3’t-dna侧翼区的部分的特征在于以下序列,所述序列由seqidno.6的核苷酸359至核苷酸691的序列、或seqidno.25的核苷酸359至核苷酸1449的互补序列的核苷酸序列、或与其具有至少97%、98%、99%、99,5%、或至少99,9%序列同一性的序列组成。
嵌合dna是指dna序列,包括在自然界中不在一起的调节序列和编码序列。因此,嵌合dna可以包含彼此相邻的dna区域,这些dna区域衍生自不同来源,或以不同于天然存在的方式排列。嵌合dna的实例是seqidno.5或6的序列。
本文还提供了在其基因组中包含事件ee-gm5的转基因大豆植物、植物细胞、组织、或种子,其特征在于包含与seqidno.1或3基本上相似的核苷酸序列的核酸分子以及包含与seqidno.2或4基本上相似的核苷酸序列的核酸分子,或所述这些序列的互补序列;以及在其基因组中包含事件ee-gm5的大豆植物、植物细胞、组织、或种子,其特征在于包含与seqidno.5或24和seqidno.6或25或所述序列的互补序列基本上相似的核苷酸序列的核酸分子。
本文甚至进一步提供了包含ee-gm5的大豆植物、细胞、组织、或种子,其特征在于在其细胞的基因组中包含与seqidno.1、3、5、或24中任一个具有至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%或100%序列同一性的核酸序列以及与seqidno.2、4、6、或25中任一个具有至少80%、至少85%、至少90%、至少95%、至少96%、至少97%、至少98%、至少99%或100%序列同一性的核酸序列,或所述序列的互补序列。
如本文所用的术语“异噁唑草酮”是指,除草剂异噁唑草酮[即(5-环丙基-4-异噁唑基)[2-(甲基磺酰基)-4-(三氟甲基)苯基]甲酮],其活性代谢物二酮腈,以及包含所述化合物的任何混合物或溶液。可施用于本发明事件上的hppd抑制性除草剂可以是二酮腈类,例如,2-氰基-3-环丙基-1-(2-甲基磺酰基-4-三氟甲基苯基)-丙烷-1,3-二酮和2-氰基-1-[4-(甲基磺酰基)-2-三氟甲基苯基]-3-(1-甲基环丙基)丙烷-1,3-二酮;其他异噁唑类;和吡唑特类(pyrazolinates),例如苯唑草酮[即[3-(4,5-二氢-3-异噁唑基)-2-甲基-4-(甲基磺酰基)苯基](5-羟基-1-甲基-1h-吡唑-4-基)甲酮],和磺酰草吡唑(pyrasulfotole)[(5-羟基-1,3-二甲基吡唑-4-基(2-甲磺酰基-4-三氟甲基苯基)甲酮];或硝磺草酮[2-[4-(甲基磺酰基)-2-硝基苯甲酰基]环己烷-1,3-二酮];或2-氯-3-(甲基硫烷基)-n-(1-甲基-1h-四唑-5-基)-4-(三氟甲基)苯甲酰胺];或2-甲基-n-(5-甲基-1,3,4-噁二唑-2-基)-3-(甲基磺酰基)-4-(三氟甲基)苯甲酰胺;或吡唑芬(pyrazofen)[2-[4-(2,4-二氯苯甲酰基)-1,3-二甲基吡唑-5-基氧基]乙酰苯酮]。
在本发明的一个实施方案中,在播种大豆之前,可以用hppd抑制剂型除草剂(例如,异噁唑草酮('ift')、苯唑草酮或硝磺草酮)或用hppd抑制剂型除草剂和草甘膦处理待种植包含ee-gm5事件的大豆植物的田地,清理可以被hppd抑制剂和/或草甘膦杀死的杂草田,允许免耕措施;然后在相同的预处理田地上种植或播种大豆(使用hppd抑制剂除草剂的快速杀灭(burndown)施用)。ift的残余活性还将保护出苗的大豆植物和正在生长的大豆植物在早期生长阶段免受杂草竞争。一旦大豆植物具有一定的大小,并且杂草倾向于重新出现,可以在植物的顶部作为出苗后除草剂施用hppd抑制剂、或hppd抑制剂与选择性的(常规的)大豆除草剂的混合物、或hppd抑制剂与在大豆中非选择性的除草剂(例如草甘膦或草铵膦)的混合物,其中所述植物包含针对该非选择性除草剂的耐受性基因/基因座,因此所述植物对所述除草剂具有耐受性。
在本发明的另一个实施方案中,在大豆植物出苗前但在播种种子后,可以用hppd抑制剂型除草剂(例如,ift、苯唑草酮、或硝磺草酮)处理在其中播种了包含ee-gm5事件的种子的田地(在播种之前,可以使用其他方法使该田地不含杂草,包括常规耕作措施,如犁耕(ploughing)、凿耕(chiselploughing)或种子床准备),其中残余活性将使该田地保持无可以被该除草剂杀死的杂草,以便使出苗和生长的大豆植物没有杂草竞争(hppd抑制剂除草剂的出苗前施用)。一旦大豆植物具有一定的大小,并且杂草倾向于重新出现,可以在植物的顶部作为出苗后除草剂施用hppd抑制剂、或hppd抑制剂-大豆选择性(常规的)除草剂混合物、或hppd抑制剂与大豆中非选择性的除草剂(例如草甘膦或草铵膦)的混合物,其中所述植物包含针对该非选择性除草剂的耐受性基因/基因座,因此所述植物对所述除草剂具有耐受性。在本发明的一个实施方案中提供了用于控制杂草的方法,该方法包括在田地中播种含有ee-gm5的大豆种子,并且在植物从所述种子出苗前但在播种种子后,用hppd抑制剂型除草剂处理所述田地。
在本发明的另一个实施方案中,可以用hppd抑制剂型除草剂(例如,ift、苯唑草酮、或硝磺草酮)在已经从播种的种子出苗的大豆植物的顶部处理包含ee-gm5事件的植物,这清除被hppd抑制剂杀死的田地杂草,其中该除草剂可以在植物的顶部使用,并与以下处理一起施用(例如,在喷雾罐混物中)、在以下处理之前施用或在以下处理之后施用:用选择性的大豆出苗后除草剂或在大豆中非选择性的除草剂(例如,草甘膦或草铵膦)处理,其中所述植物包含针对所述非选择性除草剂的耐受性基因/基因座,因此所述植物对所述除草剂具有耐受性(hppd抑制剂型除草剂的出苗后施用(组合或不组合所述大豆选择性或非选择性除草剂))。
此外,根据本发明,可以用任何大豆杀昆虫剂、除草剂、或杀真菌剂处理携带ee-gm5的大豆植物(这些植物还可以包含如本文所述的其它除草剂耐受性大豆事件/性状),或可以用任何大豆杀昆虫剂、除草剂、或杀真菌剂包衣携带ee-gm5的大豆种子。
以下实施例描述了原种事件ee-gm5的开发和鉴定,包含该事件的不同大豆系的开发、以及用于在生物样品中特异性鉴定原种事件ee-gm5的工具的开发。
除非在实施例中另有说明,否则所有重组技术均根据如在“sambrookj和russelldw(编辑)(2001)molecularcloning:alaboratorymanual[分子克隆:实验室手册],第3版,冷泉港实验室出版社(coldspringharborlaboratorypress),纽约”和在“ausubelfa,brentr,kingstonre,mooredd,seidmanjg,smithja和struhlk(编辑)(2006)currentprotocolsinmolecularbiology.[分子生物学现代方法]约翰威利父子出版公司(johnwiley&sons),纽约”中所述的标准方案进行。
标准材料和参考文献描述于“croyrdd(编辑)(1993)plantmolecularbiologylabfax[植物分子生物学labfax系列],bios科技出版社公司(biosscientificpublishersltd.),牛津和布莱克威尔科技出版公司(blackwellscientificpublications),牛津”和“brownta,(1998)molecularbiologylabfax[植物分子生物学labfax系列],第2版,学术出版社(academicpress),圣地亚哥”中。聚合酶链式反应(pcr)的标准材料和方法可见于“mcphersonmj和
应当理解,以下实施例中的任何实验室方案(例如pcr方案)中的许多参数可能需要针对特定的实验室条件调整,并且可以稍做修改以获得类似的结果。例如,在pcr方法中使用不同的dna制备方法或选择其他引物,可以确定pcr方案的其他最佳条件。然而,这些调整对于本领域技术人员而言是显而易见的,并且在当前的pcr应用手册中有进一步详述。
在说明书和实施例中,提及在所附序列表中的以下序列:
seqidno.1:5’连接ee-gm5
seqidno.2:3’连接ee-gm5
seqidno.3:ee-gm55’连接
seqidno.4:ee-gm53’连接
seqidno.5:ee-gm55’区域
seqidno.6:ee-gm53’区域
seqidno.7:cry14ab-1.b编码序列
seqidno.8:cry14ab-1蛋白质氨基酸序列
seqidno.9:hppdpf-4pa编码序列
seqidno.10:hppd-4蛋白质氨基酸序列
seqidno.11:转化质粒psz8832——t-dna边界之间的序列
seqidno.12:引物prim1038
seqidno.13:引物prim1039
seqidno.14:探针tm1788
seqidno.15:引物kvm164
seqidno.16:引物kvm165
seqidno.17:探针tm1242
seqidno.18:引物prim1041
seqidno.19:引物prim1040
seqidno.20:探针tm1789
seqidno.21:引物prim1629
seqidno.22:探针tm2083
seqidno.23:大豆事件ee-gm5
seqidno.24:ee-gm55’连接序列
seqidno.25:ee-gm53’连接序列
seqidno.26:引物glpa210
seqidno.27:引物glpa212
seqidno.28:引物glpb167
seqidno.29:引物glpb170
seqidno.30:引物prim2123
seqidno.31:引物prim2122
seqidno.32:探针tm2327
seqidno.33:插入前基因座序列
实施例
1.用线虫抗性和除草剂耐受性基因转化大豆
1.1.对包含cry14ab-1.b和hppdpf-4pa嵌合基因的插入t-dna的描述
使用含有hppdpf-4pa和cry14ab-1.b表达盒的载体psz8832,通过农杆菌(agrobacterium)介导的转化,开发ee-gm5大豆:
(i)编码hppd-4蛋白的突变体hppdpf-4pa基因(该蛋白的氨基酸序列示出在seqidno.10中)。在编码来自荧光假单胞菌(pseudomonasfluorescens)菌株a32的hppd蛋白的dna中,通过在位置335(由pro取代glu)处、位置336(由trp取代gly)处、位置339(由ala取代lys)处、和在位置340(由gln取代ala)处引入点突变,开发了hppdpf-4pa编码序列。hppd-4蛋白的表达赋予对hppd抑制剂型除草剂(例如,异噁唑草酮、苯唑草酮、或硝磺草酮)的耐受性。
(ii)cry14ab-1.b基因编码cry14ab-1蛋白(该蛋白的氨基酸序列示出在seqidno.8中)。cry14ab-1蛋白的表达赋予对线虫(例如,大豆胞囊线虫heteroderaglycines的抗性。
质粒psz8832是植物转化载体,该载体包含位于右t-dna边界(rb)和左t-dna边界(lb)之间的嵌合cry14ab-1.b基因和嵌合hppdpf-4pa基因。右和左t-dna边界之间的t-dna中包含的遗传元件的描述在下表1中给出。该质粒的t-dna(在t-dna边界之间)的确证性测序产生了seqidno.11的序列。cry14ab-1.b和hppdpf-4pa编码序列的核苷酸序列(显示编码链)分别在seqidno.7和9中给出。
表1:psz8832中t-dna边界之间的遗传元件以及在seqidno.11中的核苷酸位置的描述
1.2.事件ee-gm5
将t-dna载体psz8832引入根癌农杆菌(agrobacteriumtumefaciens)中,并根据本领域已知的方法使用hppd抑制剂耐受性选择转化的大豆植物(品种thorne)。然后将存活的植物自花授粉以产生t1种子。通过自花授粉或通过杂交到其他大豆种质中产生随后的世代。
1.2.1原种事件ee-gm5的鉴定
基于广泛选择方法(基于以下参数,包括但不限于温室和田地中的性状功效、分子特征和农学特征),从使用相同嵌合基因而获得的各种不同转化事件中,选择原种事件ee-gm5。发现包含ee-gm5的大豆植物在大豆植物基因组的单个基因座处具有转基因插入,具有与用于转化的亲本植物相似的整体农学性状,不因插入转化dna而导致产量损失(与不包含事件的相应的等基因系相比,例如从分离丢失了转基因的转化植物获得的“无效”植物系);在标准scn温室试验中导致侵染根的成年雌性的显著减少;并且与不包含ee-gm5的等基因无效系相比,田地中在scn线虫压力下具有提高的产量。此外,在田间试验中测量了对hppd抑制剂型除草剂施用的耐受性,但除草剂耐受性不是原种事件选择的选择标准。
1.2.1.1对事件的分子分析
dna印迹结果显示ee-gm5含有单个转基因基因座,该基因座包含cry14ab-1.b嵌合基因的单拷贝和hppdpf-4pa嵌合基因的单拷贝。ee-gm5丢失hppdpf-4pa嵌合基因的35s启动子的一部分(表明在转化期间不是整个seqidno.11的t-dna插入大豆基因组)。使用靶向载体骨架序列以及aada序列的引物,在pcr分析后未获得pcr片段,其中所述载体骨架序列为位于t-dna的左侧和右侧边界侧翼的序列。此外,在ee-gm5的多代中相同ee-gm5整合片段的存在表明了该事件的结构稳定性。
1.2.1.2事件的遗传
通过pcr分析测试hppdpf-4pa和cry14ab-1.b基因的基因型,插入的t-dna插入物在后续世代中的遗传表明,在ee-gm5插入物中包含的hppdpf-4pa和cry14ab-1.b基因以可预测的方式遗传,如针对单插入所预期的。这些数据与孟德尔原理一致,并支持以下结论,即ee-gm5事件由整合到大豆核基因组内的单个染色体基因座中的单个插入物组成。
此外,在ee-gm5渐渗入5个原种大豆系后,对后续世代中ee-gm5的分离模式的分析,证实了正常的孟德尔分离。表2显示了在不同的分离群体中观察到的ee-gm5的分离。
表2.分离分析ee-gm5
在表2中,“hh”代表纯合子(homozogous)植物;“hemi”代表半合子植物;和“null”代表丢失ee-gm5的无效分离子;和“ns”代表无统计学显著性(相对于来自正常/预期分离的任何变化)。在这些试验中,亲本1是具有rhg1和rhg4天然scn抗性的mgvi系;亲本2是对scn易感的mgvi系;亲本3是对scn易感的mgix系;亲本4是具有rhg1天然scn抗性的mgiii系;并且亲本5是对scn易感的mgi系。
1.2.1.3蛋白质表达的稳定性
通过夹心酶联免疫吸附测定(elisa),在从ee-gm5大豆的不同世代(例如,t4、t6和bc2f3)收集的叶、根和种子样品中,测定温室生长的植物中hppd-4和cry14ab-1蛋白的蛋白质表达水平。在所有测试的世代中,在叶、根和种子中hppd-4和cry14ab-1均表现出相似的平均表达水平。在cry14ab-1和hppd-4浓度方面观察到的任何差异归因于天然的植物与植物之间的变异性。
1.2.1.4农艺性能和对hppd抑制剂型除草剂的耐受性
在农艺等效性试验中,将在原始转化背景(thorne)中包含ee-gm5的植物与无效分离子(缺少ee-gm5)和野生型thorne植物,在scn不存在下生长时,进行比较。试验小区(plot)没有用hppd除草剂处理,而是在必要时通过使用常规除草剂和手工除草来保持无杂草状态。当在不同位置处的可比较试验中生长时,当检查定性植物特征(如花色、豆荚颜色、种子颜色、和短柔毛(pubscence))以及定量特征(如产量、高度、倒伏、直立和到达成熟的天数)时,在包含事件的植物和无效分离子(缺少ee-gm5)之间没有观察到以生物学显著方式影响农艺性能的差异。因此,包含ee-gm5的植物显示出与相应的非转基因植物相当的正常农艺学特征。
2017年进行了在原始thorne转化背景中ee-gm5的另外试验。其中ee-gm5处于原种mg1和mg3遗传背景中的初步试验也于2017年在若干位置建立。当检查定性植物特征(如花色、豆荚颜色、种子颜色、和短柔毛)以及定量特征(如产量、高度、倒伏、直立、测试重量、和达到成熟的天数)时,在三种遗传背景的任何一种中均未检测到在ee-gm5事件和无效分离子(缺少ee-gm5)之间一致且有意义的差异,证实包含ee-gm5的植物显示出了正常的农艺学特征。
经2年,在不同位置的田地中测验了包含ee-gm5的植物对hppd抑制剂型除草剂的耐受性。在这些试验中,发现在出苗前和出苗后施用时具有ee-gm5的植物对异噁唑草酮(ift)具有商业上相关的耐受性,但对于ift出苗前施用,作物损害略高。这些试验还表明,在出苗前或出苗后施用时,包含事件ee-gm5的植物对硝磺草酮(mst)具有商业上相关的耐受性。所有的出苗后处理均在v2-v3阶段,其中添加助剂作物油浓缩物和硫酸铵以增加除草剂活性。
图5显示,与未经转化的/常规的大豆植物相比,对于包含事件ee-gm5的大豆植物,在2年的若干田间试验中记录的除草剂处理的最大植物毒性(植物损害)数据的平均值。在这些相同的试验中,对照未转化的thorne植物显示平均最大植物毒性值为约80%-90%,表明(非gm)大豆不耐受这些hppd抑制剂型除草剂。如本文所用的“最大植物毒性”是在试验持续期间在任何观察中记录的最高植物毒性等级(其中每试验3至4个观测值)。在现有的杂草控制应用中,对于在出苗前或出苗后施用的异噁唑草酮(ift)和对于在出苗后施用的mst,正常(1x)剂量为105gr/ha;对于在出苗前施用的硝磺草酮,正常(1x)剂量为210gr/ha。因此,在图5中记录的这些试验中,图5中在出苗前的施用(对于ift为420gr/ha,对于硝磺草酮为840gr/ha)是正常剂量的4倍;而在出苗后的施用(ift和硝磺草酮各210gr/ha)是正常剂量的2倍。
在第3年,在一个田间试验位置,当在出苗前用异噁唑草酮(ift,按410g/ha)处理时,具有ee-gm5(在thorne背景中)的植物具有9%最大植物毒性,并且在4个位置用异噁唑草酮(ift)在出苗后(v2-v3阶段,按210h/ha)处理时,具有10.9%平均最大植物毒性,证实了之前观察到的耐受性。
此外,在2年的若干田间试验中,当分别在出苗前按400grai/ha或出苗后按200grai/ha施用时,具有事件ee-gm5的大豆植物对实验性hppd抑制剂化合物2-甲基-n-(5-甲基-1,3,4-噁二唑-2-基)-3-(甲基磺酰基)-4-(三氟甲基)苯甲酰胺(美国专利9101141)具有良好的耐受性(针对每个处理,平均最大植物毒性值为低于20%)。在这些试验中,当在出苗后按100-150grai/ha施用时,具有事件ee-gm5的大豆植物还显示对实验性hppd抑制剂化合物2-氯-3-(甲基硫烷基)-n-(1-甲基-1h-四唑-5-基)-4-(三氟甲基)苯甲酰胺(美国专利8481749)的良好耐受性(平均最大植物毒性为20%)。所有的出苗后处理均在v2-v3阶段,其中添加助剂作物油浓缩物和硫酸铵以增加除草剂活性。在第3年,在3个田间试验位置,当用2-氯-3-(甲基硫烷基)-n-(1-甲基-1h-四唑-5-基)-4-(三氟甲基)苯甲酰胺按150g/ha出苗后施用时,具有ee-gm5的植物(在thorne背景下)具有平均最大植物毒性为13.3%。
当添加从第3季的除草剂耐受性田间试验获得的数据时(在ee-gm5之前或之后但在另一个地理位置以相同剂量施用异噁唑草酮除草剂),对于ift,获得了如图5中所述的相同或非常相似的平均最大植物毒性分级。
此外,在美国的2个田间试验中,当在出苗后(v2-v3)用苯唑草酮按36gai/ha(+coc和ams)处理时,具有ee-gm5的植物给出平均最大植物毒性为11%,证实ee-gm5也赋予对这种hppd抑制剂良好的耐受性。
1.2.1.5线虫抗性
温室中测量雌性指数的标准scn测定试验显示,与thorne野生型大豆植物相比,含有ee-gm5的植物在根上具有显著减少的scn胞囊。此外,在温室中测量雌性指数的标准scn测定试验还显示,与不含有ee-gm5的scn抗性原种大豆系相比,包含事件ee-gm5和天然scn抗性的大豆植物显示出根上的scn胞囊显著减少。当ee-gm5渐渗入具有pi88788大豆抗性的原种大豆系(成熟组3)或渗入具有peking大豆抗性的原种大豆系(成熟组6.2)时,与仅具有天然抗性的根相比,在根上观察到一致性地减少的scn胞囊数量。
在若干位置的2年田间试验中,在scn侵染的田地中,与等基因无效分离子相比,含有ee-gm5的大豆植物产生显著的产量增加。图6显示在2015和2016年,在爱荷华州,伊利诺伊州,印第安纳州,密苏里州和田纳西州的9个不同位置,在scn侵染的田地中(从低到高的scn侵染),测试的原始转化体背景(thorne)中的ee-gm5的籽粒产量。在2017年,在具有不同scn压力的总共12个位置,用原始转化体背景(thorne)中的ee-gm5进行另外的试验。在所有这12项试验中,与缺少ee-gm5的无效分离子相比,含有ee-gm5的植物产生平均10%更高的产量(p=0.003)。图7显示,当将ee-gm5渗入(bc2f3)易感scn的原种mgi(成熟组i)系中后,在2016年在明尼苏达州的一个位置和北达科他州的一个位置(每个位置具有高scn侵染水平)测试的ee-gm5的籽粒产量。在2017年,在相同的两个位置(每个再次都具有高scn侵染)和在威斯康星州的另一个位置(此后者具有中等的scn压力),对相同的mgi系进行了测试,并且与缺少ee-gm5的无效分离子相比,包含ee-gm5的植物的籽粒产量一致地更高。最后,在2017年在巴西在晚种试验中在具有中度至高度scn压力的三个位置的初步研究显示,在原种易感系中,具有ee-gm5的植物与无效分离子(缺少ee-gm5)相比,31%的显著平均增加(p=0.01)。由于种植日期晚,这些初步巴西试验的总产量倾向于较低,一次试验中的变异性相当高,这可能影响产量增加的幅度,但对于具有ee-gm5的植物发现明显显著且视觉上可观察到的产量增加。因此,事件ee-gm5赋予scn侵染的田地中大豆植物显著的产量增加。
在评估与天然scn抗性结合时事件ee-gm5对产量影响的研究中,从ee-gm5与原种mgiii常规系(携带来自pi88788的rhg1抗性基因)的单杂交中,开发了一系列f3群体。在这些f3群体中,将对于事件ee-gm5和rhg1等位基因二者均纯合的一个“堆积”(stacked)群体,与仅仅rhg1等位基因纯合的群体(缺少ee-gm5)进行比较。用这些群体,在2016年在具有scn中度至高度侵染的三个位置以及在2017年在具有低到高的scn压力的七个位置,建立了产量试验。结果示出于图8中。2017年的所有试验包括三种不同的种子处理。在单独的任何这些种子处理中均没有观察到显著的产量差异或相互作用,因此跨种子处理将数据汇集以提供纯合(hh)事件和无效分离子之间产量差异的最佳统计估计。如图8所示,在2016年在所有三个位置中,“堆积”群体(ee-gm5事件和rhg1等位基因均纯合的植物)比仅携带rhg1等位基因的群体产生高8%的产量(p=0.08);并且2017年的试验中,与仅携带rhg1等位基因的群体相比,ee-gm5事件和rhg1等位基因均纯合的植物提供11%的平均产量增加(p=1.24-11)。作为参考,仅使用基础种子处理(
由于许多因素对结果产生影响,因此在中度至高度scn侵染下进行产量试验具有挑战性。田间scn群体密度可能有实质性变异,并因此scn对产量的总体影响也可能因试验小区不同而异(参见,例如,www.plantmanagementnetwork.org/pub/php/review/2009/sce08/)。有利的土壤类型、良好的肥力和充足的降雨可以减轻scn侵染对大豆植物的影响,并且即使在高scn群体下也可能使产量影响最小化。许多具有非常高scn群体的田地往往土壤贫瘠,因此产量潜力较低,因此难以辨别出对产量的统计上显著的影响。因此,来自scn田间试验的产量数据可能变化很大,并且预期不会在具有高scn群体的每个试验中均看到产量的显著改善。试验的总体趋势是用于判断事件性能的最相关标准。
用含有ee-gm5的植物进行的scn田间试验在具有天然scn侵染的田间建立。实验单元由包括2至4行(间隔0.76m,并且长度为从3.8至9.1m)的田间小区构成。行数/小区和小区长度基于田地大小和仪器配置在不同位置有变化。将小区按26粒种子/米播种,并因此每个实验单元含有200至960粒种子。使用裂区或复式裂区(split-splitplot)设计在田间将小区随机化。裂区设计非常适合于使得土壤类型或scn群体中高变异性的影响最小化,所述变异性在scn侵染的田地中很常见。在scn田间试验中,将包含ee-gm5的植物种植在子小区中,该子小区靠近或非常接近于含有无效分离子植物(不含ee-gm5)的相伴子小区。两个小区的紧密接近有助于最小化(scn)田地变异性对具有和不具有事件ee-gm5的植物之间差异估计的影响。大多数试验被重复四次,但少数重复三次,少数重复五或六次。
2016年,在两个位置(印第安纳州和爱荷华州),观察到猝死综合征的中度至重度侵染。针对sds症状的发生率和严重程度将这两个位置处的小区分级,并使用“sds评分的siuc方法”(www.scnresearch.info/462.pdf)计算sds疾病指数(dx)。与易感无效分离子(缺少ee-gm5)上的dx分级相比,ee-gm5纯合植物的dx分级在印第安纳州降低61%,并且在爱荷华州降低55%,表明该事件提供了抵抗sds侵染的保护。sds和scn常在田间密切相关,并在植物中显示一些相互作用(参见,例如,
www.soybeanresearchinfo.com/pdf_docs/sdsupdate.pdf,和www.apsnet.org/edcenter/intropp/lessons/fungi/ascomycetes/pages/suddendeath.aspx)。
在2017年,在观察到idc症状的美国的一个试验位置(具有高scn侵染),在具有ee-gm5的植物(和其无效分离子)上收集了缺铁性萎黄病(idc)得分。该试验是一个裂区设计,研究三种不同背景中的事件的影响。如cianzio等人.(1979)cropscience[作物科学]19:644-646所述进行idc分级。图11显示了,在三个遗传背景(1个scn-抗性(pi88788抗性),1个scn易感性、和scn易感性thorne背景)中,具有事件ee-gm5的植物和相应无效分离子(缺少ee-gm5)的idc得分平均值。与其无效分离子相比,发现包含ee-gm5的植物的idc得分显著更低。因此,在大豆植物同时受到scn和idc攻击的田间试验中,ee-gm5显著降低了idc的叶面严重性。这种减少发生在所有三个大豆系中,其中一个包括pi88788型天然scn抗性。
此外,使非转化的thorne和ee-gm5种子萌发并种植在温室中以检查对腐线虫,最短尾短体线虫,的控制。在2周龄时,将最短尾短体线虫(#1500/植物,不同的发育阶段)施用于植物。施用后30天,从根中提取短体线虫并计数。将含有ee-gm5的植物根部中发现的线虫的平均数量与在野生型thorne植物根中发现的短体线的平均数量进行比较。与thorne对照根相比,在含有ee-gm5的植物的根中发现少平均约80%至90%的短体线虫,表明通过大豆事件ee-gm5显著控制腐线虫。
图9显示了来自美国最短尾短体线虫温室试验的结果,该试验中比较了在5个原种大豆系(一个scn易感性(mg1)、一个scn抗性(pi88788,mg3)、一个scn易感性(mg6.2)、一个scn抗性(peking,mg6.2)、和一个scn易感性(mg9))中具有ee-gm5的原种系与scn易感性和scn抗性美国大豆系。使这些大豆植物在小锥形盆中生长并保持在温度25℃-32℃之间变化的温室中。从南卡罗来纳州获得并在温室中增加的最短尾短体线虫,用于接种处于v2-v3发育阶段的植物。将每株植物接种约1500个卵+成虫,并且每个条目具有5株植物。侵染后30天,从根中提取线虫和卵并计数。每个条目都在两个独立的实验中运行。虽然scn-易感性和scn抗性美国大豆系没有显示出对短体线虫的控制,但具有ee-gm5的植物显示出对短体线虫的约90%控制。
图10显示了来自巴西的最短尾短体线虫温室试验的结果,将具有ee-gm5的大豆植物与不具有抗性的巴西大豆系、和1个低rf系、和scn易感性和抗性植物进行比较。使这些大豆系在小锥形盆中生长并保持在温度25℃-32℃之间变化的温室中。从巴西田地获得并在温室中增加的最短尾短体线虫,用于接种处于v2-v3发育阶段的植物。将每株植物接种约1000个卵+成虫,并且每个条目具有5株植物。侵染后30天,从根中提取线虫和卵并计数。显示的结果来自单个实验。一个巴西大豆系(brs7380),标记为对于短体线虫具有低繁殖因子,显示出约89%的短体线虫减少。具有ee-gm5的植物给出对短体线虫的约99%控制。携带scn天然抗性(rhg1+rhg4)的大豆系不控制最短尾短体线虫。
此外,包含ee-gm5的植物还可以用于控制根结线虫(rkn)(例如,南方根结线虫)。虽然南方根结线虫群体不能很好地侵染thorne野生型大豆,但与未转化的thorne植物相比,具有ee-gm5的thorne植物显示出平均rkn卵数量/根质量的进一步减少。
1.2.2原种事件ee-gm5的侧翼区和插入t-dna的鉴定
包含在ee-gm5原种事件中的、插入t-dna的侧翼区序列和与其邻接的t-dna的序列,显示在随附的序列表中。
1.2.2.15’t-dna侧翼区
对鉴定为包含ee-gm5的5’t-dna侧翼区的片段测序,其核苷酸序列示于seqidno.5中,核苷酸1-166。该5’t-dna侧翼区由对应于插入前基因座序列(seqidno.5,核苷酸1-166)的大豆基因组序列组成。包含插入t-dna序列的部分和与其邻接的t-dna5’侧翼序列的部分的5’连接区,显示在seqidno.1和3中。
1.2.2.23’t-dna侧翼区
对鉴定为包含ee-gm5的3’t-dna侧翼区的片段测序,其核苷酸序列示于seqidno.6中,核苷酸359-691。该3’t-dna侧翼区由以下构成:39个核苷酸的填充dna序列(在seqidno.6中从位置359至位置397),接着是对应于插入前基因座序列的大豆基因组序列(从seqidno.6的位置398至位置691)。包含插入t-dna序列的部分和与其邻接的t-dna3’侧翼序列的部分的3’连接区,显示在seqidno.2和4中。
1.2.2.3ee-gm5的插入t-dna
对邻接以上5’t-dna侧翼序列的插入t-dna进行测序,其核苷酸序列表示在seqidno.5中,核苷酸167-353。此外,将邻接以上3’t-dna侧翼序列的插入t-dna进行测序,其核苷酸序列表示在seqidno.6中,核苷酸1-358。在转化过程中,在插入前基因座序列中缺失了63bp的基因组dna,并且其被插入的dna(由t-dna和填充dna(fillerdna)的一小部分构成)代替。
对转化质粒psz8832中的t-dna区域(t-dna边界之间的部分)的测序,产生seqidno.11中报道的序列。嵌合cry14ab-1.b基因序列(包含ubi10启动子和35s3’非翻译区)显示在seqidno.11核苷酸131-5276中(逆时针)。在seqidno.5(核苷酸167-353)中5’侧翼区的插入t-dna序列与seqidno.11的核苷酸1至核苷酸187的核苷酸序列相同,并且在seqidno.6(核苷酸1-358)中3’侧翼区的插入t-dna序列与seqidno.11的核苷酸7102至核苷酸7459的核苷酸序列相同。因此,ee-gm5中的插入t-dna的5’末端对应于seqidno.11的转化质粒序列中的核苷酸1,并且ee-gm5中的插入t-dna的3’末端对应于seqidno.11的转化质粒序列中的核苷酸7459。在seqidno.5的序列和seqidno.6的序列之间的ee-gm5中的插入t-dna包含在atcc保藏号pta-123625下保藏的种子中,并且具有与seqidno.11的核苷酸188至核苷酸7101的序列基本上相似或相同的序列。
基于本文提供的5’和3’t-dna侧翼序列(seqidno.5的nt1至nt166和seqidno.6的nt359至nt691),通过本领域已知的方法,可以从野生型大豆品种thorne确定事件ee-gm5的插入基因座。大豆基因组中的插入前基因座序列按顺序对应于以下序列:seqidno.5中的核苷酸位置1至核苷酸位置166,63nt缺失,和seqidno.6中的核苷酸位置398至核苷酸位置691。完整的插入前基因座序列给出在seqidno.33中,其中nt1-1000是5’侧翼基因组序列,nt1001-1063是靶位点缺失,并且1064-2063是3’侧翼基因组序列。
1.2.3原种事件ee-gm5的侧翼区和插入t-dna的确认
使用靶向插入t-dna的上游和下游的植物dna的引物,以及靶向ee-gm5中的插入t-dna的引物,pcr扩增确认并延伸了ee-gm5的5’和3’侧翼序列。
1.2.3.1.5’连接序列ee-gm5-特异性反应
设计两个引物,glpa210和glpb167以扩增大约5118bp的扩增子,该扩增子跨越事件ee-gm5的5’t-dna侧翼序列与t-dna插入片段的连接区。引物glpa210的序列源自威廉大豆(glycinemaxwilliams)82.a2.v1的大豆参考序列。
靶向ee-gm5t-dna5’侧翼序列的正向引物:
glpa2105’-ctctcacccagatttcac-3’(seqidno.26)
靶向ee-gm5插入t-dna序列的反向引物:
glpb1675’-tacaacgtgctcgctattcc-3’(seqidno.28)
用于5’连接序列反应的反应混合物的组成:
水补足至50μl
5’连接序列反应的热循环条件:
获得的延伸的t-dna5’侧翼序列的序列(如seqidno.5所示,该序列邻接并位于插入t-dna的部分的上游)显示在seqidno.24中。1.2.3.2.3’连接序列ee-gm5-特异性反应
设计两个引物,glpb170和glpa212以扩增大约4982bp的扩增子,该扩增子跨越事件ee-gm5的t-dna插入片段与3’t-dna侧翼序列的连接区。引物glpa212的序列源自威廉大豆(glycinemaxwilliams)82.a2.v1的参考序列。
靶向ee-gm5插入t-dna序列的正向引物:
glpb1705’-tctcggtatcagcgttcttg-3’(seqidno.29)
靶向ee-gm5t-dna3’侧翼序列的反向引物:
glpa2125’-cccatgcggtattatgtg-3’(seqidno.27)
用于3’连接序列反应的反应混合物的组成:
水补足至50μl
3’连接序列反应的热循环条件:
获得的延伸的t-dna3’侧翼序列的序列(如seqidno.6所示,该序列邻接于插入t-dna的部分并在其下游)显示在seqidno.25中。
由于上述2个反应中所得的扩增子重叠,这允许重建ee-gm5插入t-dna序列和延伸的5'和3'侧翼序列,如seqidno.23所示。seqidno.23中5’t-dna侧翼序列是从核苷酸位置1至核苷酸位置1113(对应于插入前基因座基因组序列);插入t-dna序列是从核苷酸位置1114至核苷酸位置8572;并且seqidno.23中3’t-dna侧翼序列是从核苷酸位置8573至核苷酸位置9663(对应于39nt填充dna(在seqidno.23中nt8573-8611);以及插入前基因座基因组序列(在seqidno.23中nt8612-9663))。
2.针对ee-gm5的鉴定方案的开发
2.1.ee-gm5身份分析的终点方法
该方法描述了聚合酶链式反应检测方法,使用标准dna提取程序以分析从生物样品(例如,植物材料(例如,叶和种子))中获得的dna样品中事件ee-gm5-特异性dna序列的存在。
该方法描述概述了方法设计,包括寡核苷酸引物和探针序列,反应混合物的组成,进行反应所需的热循环条件,以及发现适合于扩增子检测的荧光读数器设置。还提供了关于对照样品的性质和用途的一般性建议。此外,还提供了用于数据分析和解释的指南,包括在考虑对照材料的使用建议和数据分析指南的情况下的方法结果的实例。
2.1.1.方法设计
该方法使用taqman化学来扩增和检测两个靶序列:ee-gm5特异性反应确定事件的存在,分类单元特异性反应验证事件特异性反应的阴性结果。
2.1.1.1.ee-gm5-特异性反应
设计两个引物,prim1038和prim1039以扩增85bp的扩增子,该扩增子跨越事件ee-gm5的3’侧翼序列与t-dna插入片段的连接区。
使用fam作为荧光标记并且使用bhq1作为猝灭剂的探针tm1788被设计用于检测扩增的序列。
靶向ee-gm5t-dna序列的正向引物:
prim10385’-gagccaccttccttttccacta-3’(seqidno.12)
靶向ee-gm5t-dna3’侧翼序列的反向引物:
prim10395’-atagggttactgcttcgtaaaataagca-3’(seqidno.13)
靶向ee-gm5t-dna和其’3’侧翼序列的连接序列的探针:
tm1788fam5’-cgcgtccatgatgctgcgactatg-3’bhq1(seqidno.14)
2.1.1.2.分类单元特异性反应
设计两个引物,kvm164和kvm165以扩增大豆内源性植物凝集素1基因序列的102bp的扩增子。
使用joe作为荧光标记并且使用bhq1作为猝灭剂的探针tm1242被设计用于检测扩增的序列。
靶向内源性植物凝集素1基因序列的正向引物:
kvm1645’-ctttctcgcaccaattgaca-3’(seqidno.15)
靶向内源性植物凝集素1基因序列的反向引物:
kvm1655’-tcaaactcaacagcgacgac-3’(seqidno.16)
靶向内源性植物凝集素1基因序列的探针:
tm1242joe5’-ccacaaacacatgcaggttatcttgg-3’bhq1(seqidno.17)
2.1.2.反应混合物的组成
水补足至10μl
注意:
·2xperfectaqpcrfastmixii,rox由量子生物科学公司(quantabioscience)提供。可以使用其他酶缓冲液,但应验证其性能。
·向整合dna技术公司(integrateddnatechnologies)定购引物和标记探针
·*每次反应中模板dna的量可能会变化,但应予以验证2.1.3.热循环条件
注意:
·该热循环条件被验证用于在bioradc1000热循环仪上使用。可以使用其他仪器,但应验证其性能
2.1.4.波长和带宽设置
注意:
·该波长和带宽设置被验证用于在tecanm1000读板仪上使用。可以使用其他仪器和设置,但应验证其性能
2.1.5.对照样品
以下对照样品应包含在实验中以验证测试样品的结果:
·阳性对照:包含靶和内源序列的dna样品
·阴性对照:仅包含内源序列的dna样品
·无模板对照:水样品(无dna)
2.1.6.数据分析
·对于所有样品,针对靶和内源性反应两者计算荧光信号与背景的比率(s/b)。
·对照样品应给出预期结果,即:
o阳性对照应评分为“检测到”
o阴性对照应评分为“未检测到”
o无模板对照应仅显示荧光背景水平
·将样品评分如下:
o检测到:靶s/b和内源性s/b超过可接受的阈值比率(例如,2)
o未检测到:靶s/b低于可接受的阈值比率(例如,1)并且另外,内源性s/b超过可接受的阈值比率(例如,2)
o不确定:靶和内源性s/b低于可接受的阈值(例如,1)
图2显示了针对含有ee-gm5的一系列大豆样品和常规大豆样品的方法结果实例。对于每个样品,显示了针对ee-gm5特异性反应和内源性反应的s/b比率。
2.2.用于ee-gm5身份和合子型分析的终点方法
该方法描述了聚合酶链式反应检测方法,使用标准dna提取程序以分析从生物样品(例如,植物材料(例如,叶和种子))中获得的dna样品中事件ee-gm5-特异性dna序列的存在和合子型状态。
该方法描述概述了反应试剂、寡核苷酸引物和探针序列,进行反应所需的热循环条件,以及发现适合于扩增子检测的荧光读数器设置。还提供了关于对照样品的性质和用途的一般性建议。此外,还提供了用于数据分析和解释的指南,包括考虑到对照材料使用建议和数据分析指南的方法结果实例。
应注意,由于插入前基因座序列的性质,(以下选项1的)合子型分析的方法性能可能是品种依赖性的。因此,对于渗入事件的每个品种,都需要性能验证。对于性能不足的情况,可以使用针对合子型的可替代的终点方法设计(如以下的选项2),或可以将基于拷贝数分析的可替代的实时pcr方法用于合子型测定,例如以下在2.3节中所述。
2.2.1.方法设计
该方法使用taqman化学来扩增和检测两个靶序列:ee-gm5特异性反应确定事件的存在,插入前基因座-特异性反应确定事件的插入前基因座的存在。
仅检测到ee-gm5特异性序列表明纯合子型状态的事件ee-gm5的存在。
检测到ee-gm5特异性和插入前基因座特异性序列表明半合子型状态的事件ee-gm5的存在。
仅检测到插入前基因座特异性序列表明不存在事件ee-gm5。
2.2.1.1.ee-gm5-特异性反应
a.选项1
设计两个引物,prim1040和prim1041,以扩增84bp的扩增子,该扩增子跨越事件ee-gm5的t-dna5’侧翼序列与t-dna插入片段的连接区。
使用fam作为荧光标记并且使用bhq1作为猝灭剂的探针tm1789被设计用于检测扩增的序列。
靶向ee-gm5t-dna序列的正向引物:
prim10415’-cattgtgctgaataggtttatagctatgat-3’(seqidno.18)
靶向ee-gm5t-dna5’侧翼序列的反向引物:
prim10405’-tcaaatcaacatgggtgactagaaa-3’(seqidno.19)
靶向ee-gm5t-dna和其5’侧翼序列的连接序列的探针:
tm1789fam5’-cagtactgggcccttgtggcgct-3’bhq-1(seqidno.20)
b.选项2
设计两个引物,prim2123和prim1041,以扩增134bp的扩增子,该扩增子跨越事件ee-gm5的t-dna5’侧翼序列与t-dna插入片段的连接区。
使用fam作为荧光标记并且使用bhq1作为猝灭剂的探针tm1789被设计用于检测扩增的序列。
靶向ee-gm5t-dna序列的正向引物:
prim10415’-cattgtgctgaataggtttatagctatgat-3’(seqidno.18)
靶向ee-gm5t-dna5’侧翼序列的反向引物:
prim21235’-gcactgtttaactttaaataactcatttgag-3’(seqidno.30)
靶向ee-gm5t-dna和其5’侧翼序列的连接序列的探针:
tm1789fam5’-cagtactgggcccttgtggcgct-3’bhq-1(seqidno.20)
2.2.1.1.插入前基因座特异性反应
a.选项1
设计两个引物,prim1629和prim1040,以扩增72bp的扩增子,该扩增子跨越插入前基因座和ee-gm5插入前基因座的5′侧翼序列的连接区。
使用vic作为荧光标记并且使用mgb-nfq作为猝灭剂的mgb探针tm2083被设计用于检测扩增的序列
靶向ee-gm5插入前基因座序列的正向引物:
prim16295’-ttggtgaaaaacaatttggtgtaca-3’(seqidno.21)
靶向ee-gm5t-dna5′侧翼序列的反向引物:
prim10405’-tcaaatcaacatgggtgactagaaa-3’(seqidno.19)
靶向插入前基因座和5’侧翼序列的连接序列的野生型探针:
tm2083vic5’-aatcaaatcgacatcaatgt-3’mgb-nfq(seqidno.22)
b.选项2
设计两个引物,prim2122和prim2123,以扩增193bp的扩增子,该扩增子跨越插入前基因座和ee-gm5插入前基因座的5′侧翼序列的连接区。
使用vic作为荧光标记并且使用mgb-nfq作为猝灭剂的mgb探针tm2327被设计用于检测扩增的序列。
靶向ee-gm5插入前基因座序列的正向引物:
prim21225’caagcaaaataagcaactagatctattgg-3’(seqidno.31)
靶向ee-gm5t-dna5’侧翼序列的反向引物:
prim21235’-gcactgtttaactttaaataactcatttgag-3’(seqidno.30)
靶向插入前基因座和5’侧翼序列的连接序列的野生型探针:
tm2327vic5’-tttggtgaaaaacaatttggtgt-3’mgb-nfq(seqidno.32)
2.2.2.反应混合物的组成
a.选项1
水补足至10μl
b.选项2
水补足至10μl
注意:
·2xperfectaqpcrfastmixii,rox由量子生物科学公司(quantabioscience)提供。可以使用其他酶缓冲液,但应验证其性能。
·向整合dna技术公司(integrateddnatechnologies)定购引物和标记探针
·*每次反应中模板dna的量可能会变化,但应予以验证2.2.3.热循环条件(对于选项1和2是相同的)
注意:
·该热循环条件被验证用于在bioradc1000热循环仪上使用。可以使用其他仪器,但应验证其性能
2.2.4.波长和带宽设置(对于选项1和2是相同的)
注意:
·波长和带宽设置被验证用于在tecanm1000读板仪上使用。可以使用其他仪器和设置,但应验证其性能
2.2.5.对照样品(对于选项1和2是相同的)
以下对照样品应包含在实验中以验证测试样品的结果:
·纯合子对照:包含处于纯合状态的靶序列的dna样品
·半合子对照:包含处于半合子状态的靶序列的dna样品
·野生型对照:不包含靶序列的dna样品
·无模板对照:水样品
2.2.6.数据分析(对于选项1和2是相同的)
·对于所有样品,针对靶和插入前基因座反应,计算荧光信号与背景的比率(s/b)。
·对照样品应给出预期结果,即:
o纯合子对照应评分为“纯合子”
o半合子对照应评分为“半合子”
o野生型对照应评分为“野生型”
o无模板对照应仅显示荧光背景水平
·将样品评分如下:
o纯合子:靶s/b超过可接受的阈值比率(例如,2),并且插入前基因座s/b低于可接受的阈值比率(例如,1)
o半合子:靶和插入前基因座s/b超过可接受的阈值比率(例如,2)
o野生型:靶s/b低于可接受的阈值比率(例如,1),并且插入前基因座s/b超过可接受的阈值比率(例如,2)
o不确定:靶和插入前基因座s/b低于可接受的阈值(例如,1)
图3显示了对于包含纯合状态的ee-gm5的大豆样品、包含半合子状态的ee-gm5的大豆样品和常规大豆样品的(以上选项1的)方法结果的实例。
2.3.用于ee-gm5身份和合子型分析的实时pcr方法
该方法描述了定量实时聚合酶链式反应检测方法,使用标准dna提取程序以分析从生物样品(例如,植物材料(例如,叶或种子))中获得的dna样品中事件ee-gm5dna序列的合子型状态。
该方法描述概述了反应试剂、寡核苷酸引物和探针序列,进行反应所需的热循环条件,以及发现适合于扩增子检测的荧光读数器设置。还提供了关于对照样品的性质和用途的一般性建议。此外,还提供了用于数据分析和解释的指南。
该方法不依赖于品种,并且可以用作以上第2.2节中描述的方法的替代性的合子型分析方法。
2.3.1.方法设计
该方法使用taqman化学和实时pcr原理来量化ee-gm5特异性序列的相对拷贝数。
该方法包括ee-gm5特异性反应以量化ee-gm5拷贝数,以及分类单元特异性反应以标准化ee-gm5拷贝数。
包含处于纯合状态的ee-gm5插入序列的样品具有比半合子样品高两倍的相对拷贝数。无合子的样品不扩增ee-gm5序列。
2.3.2.ee-gm5-特异性反应
设计两个引物,prim1038和prim1039以扩增85bp的扩增子,该扩增子跨越事件ee-gm5的3’侧翼序列与t-dna插入片段的连接区。
使用fam作为荧光标记并且使用bhq1作为猝灭剂的探针tm1788被设计用于量化扩增的序列。
靶向ee-gm5t-dna序列的正向引物:
prim10385’-gagccaccttccttttccacta-3’(seqidno.12)
靶向ee-gm53’侧翼序列的反向引物:
prim10395’-atagggttactgcttcgtaaaataagca-3’(seqidno.12)
靶向ee-gm5t-dna和其3’侧翼序列的连接序列的探针:
tm1788fam5’-cgcgtccatgatgctgcgactatg-3’bhq-1(seqidno.14)
2.3.3.分类单元特异性反应
设计两个引物,kvm164和kvm165以扩增大豆内源性植物凝集素1基因序列的102bp的扩增子。
使用joe作为荧光标记并且使用bhq1作为猝灭剂的探针tm1242被设计用于量化扩增的序列
靶向内源性植物凝集素1基因序列的正向引物:
kvm1645’-ctttctcgcaccaattgaca-3’(seqidno.15)
靶向内源性植物凝集素1基因序列的反向引物:
kvm1655’-tcaaactcaacagcgacgac-3’(seqidno.16)
靶向内源性植物凝集素1基因序列的探针:
tm1242joe5’-ccacaaacacatgcaggttatcttgg-3’bhq1(seqidno.17)
2.3.4.反应混合物的组成
水补足至10μl
注意:
·2xperfectaqpcrfastmixii,lowrox由量子生物科学公司(quantabioscience)提供。可以使用其他酶缓冲液,但应验证其性能。
·向整合dna技术公司(integrateddnatechnologies)定购引物和标记探针
·*每次反应中模板dna的量可能会变化,但应予以验证
2.3.5.热循环条件
注意:
·**在每个循环中,在60℃下完成引物延伸步骤后,进行荧光读数
·该热循环条件被验证用于在viia7和quantstudio7实时pcr装置上使用。可以使用其他仪器,但应验证其性能
2.3.6.对照样品
以下对照样品应包含在实验中以验证测试样品的结果
·纯合子对照:包含处于纯合状态的靶序列的dna样品
·半合子对照:包含处于半合子状态的靶序列的dna样品
·野生型对照:不包含靶序列的dna样品
·无模板对照:水样品
2.3.7.数据分析
·使用ddct方法进行数据分析。在该方法中,相对于所选的参考样品,针对所有样品计算拷贝数。建议使用半合子对照作为参考样品。
·对照样品应给出预期结果,即:
o纯合子对照应评分为“纯合子”
o半合子对照应评分为“半合子”
o野生型对照应评分为“野生型”
o无模板对照应仅显示荧光背景水平
·将样品评分如下:
o纯合子:相对拷贝数是2+/-可接受的阈值,例如0.5
o半合子:相对拷贝数是1+/-可接受的阈值,例如0.25
o野生型:相对拷贝数是0+可接受的阈值,例如0.1
o不确定:相对拷贝数超出针对纯合子、半合子和野生型样品的可接受范围
2.4.用于ee-gm5低水平存在分析的实时pcr方法
该方法描述了检测方法,其中使用标准dna提取程序分析从大块的植物材料(例如,叶或种子)或加工材料(例如,包含经加工的大豆籽粒的食品或饲料)中获得的事件ee-gm5dna序列的低水平存在。
该方法描述概述了反应试剂、寡核苷酸引物和探针序列,以及进行反应所需的热循环条件。还提供了关于同时使用分类单元特异性方法的一般建议来支持数据分析和结果解释。另外,提供了关于对照样品的性质和用途的建议。
应注意,替代性方法可用于预期目的,包括但不限于数字液滴pcr方法。数字液滴pcr方法使用1.1节所述的用于事件身份分析的终点方法,并结合对提取的dna样品进行二次取样(subsample)的原理。在该方法中,基于针对事件序列发现的阳性和阴性dna子样品的比率来确定事件的低水平存在。
2.4.1方法设计
该方法使用taqman化学和实时pcr原理来检测或量化dna样品中低水平的ee-gm5。
设计两个引物,prim1040和prim1041以扩增84bp的扩增子,该扩增子跨越事件ee-gm5的5’侧翼序列与t-dna插入片段的连接区。
使用fam作为荧光标记并且使用bhq1作为猝灭剂的探针tm1789被设计用于量化扩增的序列。
靶向ee-gm5t-dna序列的正向引物:
prim10415’-cattgtgctgaataggtttatagctatgat-3’(seqidno.18)
靶向ee-gm5t-dna5’侧翼序列的反向引物:
prim10405’-tcaaatcaacatgggtgactagaaa-3’(seqidno.19)
靶向ee-gm5t-dna和其5’侧翼序列的连接序列的探针:
tm1789fam5’-cagtactgggcccttgtggcgct-3’bhq-1(seqidno.20)
2.4.2反应混合物的组成
水补足至20μl
注意:
·2xperfectaqpcrfastmixii,lowrox由量子生物科学公司(quantabioscience)提供。可以使用其他酶缓冲液,但应验证其性能。
·向整合dna技术公司(integrateddnatechnologies)定购引物和标记探针
·*每次反应中模板dna的量可能会变化,但应予以验证2.4.3热循环条件
注意:
·**在60℃下完成引物延伸步骤后,在每个循环中进行荧光读数
·该热循环条件被验证用于在viia7和quantstudio7实时pcr装置上使用。可以使用其他仪器,但应验证其性能
2.4.4分类单元特异性方法
靶向内源序列的实时pcr检测方法应当在与靶特异性实时pcr方法中使用的相同量的模板dna上同时进行。应使用分类单元特异性方法的结果以支持数据分析和解释,即标准化输入dna的量,并验证靶特异性反应的任何阴性结果。
2.4.5测试样品、校准样品和对照样品
·建议对所有测试样品进行一式两份的分析。
·实验中包括一组校准样品,以生成针对靶和分类单元特异性方法的标准曲线。
·此外,还包括以下对照样品:
o阳性对照:包含处于检测限水平的靶序列的dna样品,
o阴性对照:仅包含内源序列的dna样品
o无模板对照:水样品
2.4.6数据分析
·对于所有样品,针对靶和分类单元特异性方法,测定阈值循环值(即,ct值)。阈值循环被定义为给定样品的扩增曲线达到所定义的信号阈值的循环数(参见图4)
·使用校准样品的ct值和基因组拷贝数,对靶和分类单元特异性方法,计算标准曲线公式
·标准曲线参数应满足斜率和线性(r2)的可接受标准,例如
o-3.2<斜率<-3.6
or2>0.98
·对于所有样品,使用线性回归分析,计算靶和内源性方法的基因组拷贝数。
·通过计算靶和分类单元特异性方法的基因组拷贝数的%比率来测定相对于分类单元特异性dna总量的低水平存在量。
·对照样品应给出预期结果,即:
o阳性对照应评分为“检测到”
o阴性对照应评分为“未检测到”
o无模板对照应仅显示荧光背景水平
·将样品评分如下:
o检测到:考虑到该方法的测量误差,低水平存在为对于所有重复而言均高于检测限
o未检测到:考虑到该方法的测量误差,低水平存在为对于所有重复而言均低于检测限
o不确定:重复样品给出不一致的得分
图4显示了在校准样品上进行的该方法的结果实例。
3.ee-gm5渗入到优选的栽培品种
通过反复回交到六个不同的原种大豆系而引入原种事件ee-gm5。选择这些系以代表一系列的成熟系:来自mgi的两个系、来自mgiii的一个系、来自mgvi的两个系、和来自mgix的一个系。mgi系的一个和mgiii系包含来自pi88788的rhg1天然抗性等位基因,并且mgvi系的一个携带来自pi437654的rhg1和rhg4天然抗性等位基因。其他三个系对scn是易感的。
此外,在初步测试中,在若干个实验中,当事件ee-gm5从thorne背景渗入到其他大豆种质背景(不同的成熟度,在基因渗入的不同阶段)时,与针对thorne背景中的ee-gm5发现的结果相比,在温室中生长的植物的叶中测量(如用elisa或蛋白质印迹法测定(仅观察到正常的测定变异)),对于cry14ab-1未观察到hppd-4蛋白质的表达水平方面的生物学显著的差异;并且在标准的温室scn测定结果(测量scn胞囊相比于thorne对照的%减少)中未观察到显著的差异。
原种事件ee-gm5向其他大豆栽培品种中的渗入不会显著影响这些栽培品种的任何期望表型或农艺学特征(无连锁累赘(linkagedrag)),而转基因的表达满足商业上可接受的水平。这证实了事件ee-gm5作为原种事件的状态。
此外,原种事件ee-gm5可以有利地与其他大豆原种转化事件组合。根据本发明特别有用的植物是包含ee-gm5与另一大豆转化事件的组合、或与一个以上的其他大豆转化事件的组合的植物,这些转化事件例如是在不同国家或地区管理机构的数据库中列出的那些,包括但不限于,事件mon87751(描述于wo2014201235和usda-aphispetition13-337-01p中)、事件pdab8264.42.32.1(描述于wo2013010094)、事件das-81419-2(也称作conkestatm大豆,描述于wo2013016527和usda-aphispetition12-272-01p中)、事件ee-gm3(也称作fg-072,mst-fg072-3,描述于wo2011063411,usda-aphispetition09-328-01p中)、事件syht0h2(也称作0h2,syn-
如以下权利要求中所使用的,除非另有明确说明,术语“植物”旨在包括植物组织(在成熟的任何阶段),以及取自或衍生自任何这种植物的任何细胞、组织或器官,包括但不限于任何种子、叶、茎、花、根、单细胞、配子、细胞培养物、组织培养物或原生质体。
包含原种事件ee-gm5的参考种子于2016年11月9日在atcc(10801universityblvd.,马纳萨斯,va20110-2209)以atcc保藏号pta-123625保藏,并且确认了其存活力。ee-gm5的替代名称是事件gmb151或bcs-gm151-6。
本发明的以上描述旨在是举例说明性的而非限制性的。
本领域技术人员可以想到所述实施方案中的各种变化或修饰。这些可以在不脱离本发明的精神或范围的情况下进行。
- 还没有人留言评论。精彩留言会获得点赞!