一种环境胁迫下植物中酶活性的原位可视化测定方法与流程
本发明涉及可视化植物毒性评价
技术领域:
,尤其涉及一种原位可视化污染物胁迫下植物叶片c、p代谢有关酶活性空间分布的表征技术,更具体地涉及邻苯二甲酸酯胁迫下,叶片中葡萄糖苷酶、木聚糖酶和酸性磷酸酶活性的空间分布的原位可视化表征方法。
背景技术:
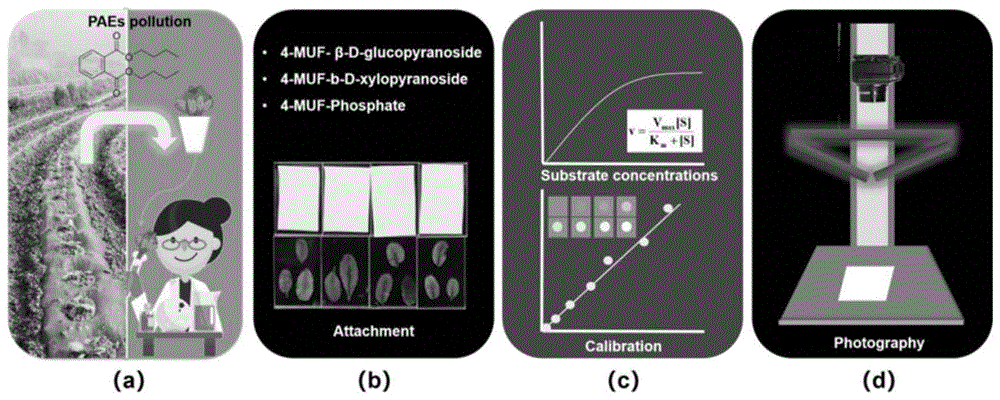
:邻苯二甲酸酯(paes)是一类重要的人工合成有机化合物,广泛应用于塑料增塑剂(俗称塑化剂),以及汽车、服装、化妆品、润滑剂和农药等行业,大量进入环境中,并具有半挥发性和远距离迁移性,成为全球性环境污染物。paes具有致癌、致畸、致突变等毒性,尤以造成人体生殖功能异常最受关注,是典型的内分泌干扰物,被人们称为“第二个全球性pcb污染物”。目前,包括我国在内的许多国家和环保组织将其列入优先控制污染物,并采取相应措施防止paes对人类健康的危害。迄今为止,植物酶活性已被广泛用作评价重金属和有机污染物等环境胁迫的生物指标。例如,超氧化物歧化酶(sod)、过氧化氢酶(cat)、过氧化物酶(pod)等几种抗氧化酶被用来评价在胁迫条件下(包括盐度、辐照、低温、有机污染物和重金属等)活性氧(ros)的生成。与蔗糖磷酸合成酶(sps)、转化酶(inv)和蔗糖合成酶(ss)一样,ros通常用于评估环境条件对植物碳代谢的影响。酶活性对污染物的响应不均匀,酶的选择因研究的不同而不同。然而,以往的研究仅在生化水平上进行;很少涉及到在环境样品中,特别是在植物中。事实上,酶活性测定已有60多年的历史,常用的方法有分光光度法、荧光法、滴定法和放射性同位素法,而且大多数利用荧光成像方法的研究都是定性的,而关于植物中酶活性对应激条件的反应的可视化目前还未见报道。酶谱是通过底物转换(本质上是酶成像)来显示酶活性。利用荧光成像来测量酶活性是一种强大的方法,因为它具有灵敏度高、图像采集相对容易、获得的环境样品中酶活性空间异质性的大量精细尺度信息。目前关于酶谱的研究主要集中在土壤酶谱技术,可用于作用于任何生物底物上的水解酶或氧化酶,包括蛋白质和多肽、低聚糖和多糖、脂质和糖。然而,关于叶片酶谱的研究至今未有报道。技术实现要素:本发明的目的在于克服现有技术中存在的上述缺陷和不足,提供一种污染物胁迫下植物中酶活性的原位可视化测定方法。本发明的第二个目的在于提供上述原位可视化测定方法的应用。本发明的上述目的是通过以下技术方案给予实现的:一种环境胁迫下植物中酶活性的原位可视化测定方法,包括如下步骤:s1.将植物在环境胁迫下进行预处理;s2.配置酶活性荧光标记底物溶液,将聚酰胺膜浸泡在所述酶活性荧光标记底物溶液中,待膜被底物溶液均匀饱和后取出,风干后将其与s1预处理后的植物叶片表面紧密贴合,固定,叶片两侧施加压力,于暗室中反应;反应结束后,将聚酰胺膜从叶片表面剥离,于激发波长为355nm,发射波长为460nm的紫外灯下,对聚酰胺膜进行拍摄,获得叶片酶谱图像;s3.配置系列浓度的标准荧光产物溶液,以未添加荧光产物的缓冲液为空白对照,将聚酰胺膜于缓冲液中浸泡;再将上述系列浓度的标准荧光产物溶液滴加到风干的聚酰胺膜上,使用与s2酶谱成像相同的拍摄条件进行拍摄,获得标曲图像;s4.将s3获得的标曲图像,原始图像转化为8-bit灰阶图,以σg=0.5的高斯滤波核进行降噪,再以灰度值为98作为全局阈值对所有降噪后的梯度浓度标准荧光灰阶图进行图像分割并计算灰度均值,最终得到扣除背景的梯度浓度标准荧光均值,然后根据不同浓度的标准荧光产物与其图像灰度的变化拟合出分段校正曲线:其中为施加于膜上的荧光底物浓度;a1,a2,b1,b2是线性回归拟合的参数;g*为分断点处的灰度值;是经过分割的图像的平均灰度值;分段校正曲线的第一部分用于将酶谱中的像素灰度值转化为酶活性,并将校正曲线中的灰度值赋予成比例关系的rgb颜色,获得的rgb颜色条将用于创建查找表lut,用于生成伪彩色叶片酶谱图;将s2获得的叶片酶谱图像转化为8-bit灰阶图,以σg=0.5的高斯滤波核进行降噪,记录各图像四角5×5范围内的灰度均值,以此作为图像的背景灰度并在全图扣除得到最终的灰度图像,然后将lut应用于叶片图像,从而将叶片酶活性的空间分布可视化。本发明是利用高淬灭荧光底物法来进行植物酶活的原位可视化;底物一端连接一分子的荧光色素结合,另一端与荧光淬灭剂结合,正常情况下荧光信号无法释放。当被该底物饱和的膜与预处理后的植物相接触后,底物被植物叶片中的酶降解,淬灭剂脱落,作用消失,在紫外的激发光下发射出荧光,此时通过检测荧光信号强度在膜上的分布便可获得植物酶活性在叶片上的分布情况。在上述步骤中,其中,叶片图像处理是一个重要的问题,它会对所有后续统计分析的结果产生深远影响。但是,先前的研究缺乏有关如何重建高质量图像从而提高其定性准确性的有用信息。本发明通过使用σg=0.5的高斯滤波器对进行降噪增强处理。同时为了获得荧光底物的真实荧光强度,本发明在计算标曲图像的灰度平均值之前进行图像分割,使用了全局阈值的方法来分割背景和前景;最终通过图像去噪和分割对图像进行处理以重建高质量图像从而提高其定性准确性的有用信息。优选地,还包括以叶片酶谱中灰度值超过校正曲线第一部分75%的区域为热区,通过计算叶片酶谱中热区面积的变化来研究酶活性较高区域在环境胁迫下空间分布的变化,用于表示酶活性从叶基到叶尖的分布趋势。从而为了更好地说明沿主叶脉酶活性的空间分布具体地,所述植物为菜心。具体地,所述环境胁迫为污染物胁迫。具体地,所述污染物为邻苯二甲酸酯。具体地,所述污染物为邻苯二甲酸二丁酯。具体地,所述酶为β-葡萄糖苷酶、酸性磷酸酶或木聚糖酶中的至少一种。优选地,所述荧光标记为4-甲基伞形酮;即所述酶活性荧光标记底物为4-甲基伞形酮-β-d-葡糖苷(muf-g)、4-甲基伞形酮-β-d-磷酸酯(muf-p)或4-甲基伞形酮-β-d-木糖苷(muf-x)中的至少一种,其浓度为80~120μm。优选地,所述聚酰胺膜孔径为0.45μm,大小为3×5cm2。具体地,本发明通过上述酶谱技术来量化植物叶片中dbp胁迫下的酶活性。,研究葡萄糖苷酶、木聚糖酶和酸性磷酸酶在植物叶子的二维分布,为污染物胁迫下的植物毒性评价和生理反应提供了新的思路。作为一种优选地可实施方式,所述dbp胁迫下植物中葡萄糖苷酶、木聚糖酶和酸性磷酸酶的原位可视化测定方法包括如下步骤:(1)植物种植和暴露:种植无污染的植物幼苗,例如菜心;配置一系列dbp溶液,将幼苗置于dbp环境中暴露,进行污染物胁迫预处理;(2)酶谱成像:幼苗完成暴露后,将4-甲基伞形酮-β-d-葡糖苷(muf-g)、4-甲基伞形酮-β-d-磷酸酯(muf-p)和4-甲基伞形酮-β-d-木糖苷(muf-x)配置成浓度为100μm的酶底物缓冲液;将孔径为0.45μm的聚酰胺膜裁剪成3×5cm2大小,在配置的酶底物缓冲液中浸泡8~12min,风干2~4min后将其与不同浓度dbp暴露的幼苗叶片表面紧密贴合;使用亚克力板固定并在两面各施予1.0kg的压力,于暗室中反应30~40min。反应结束后,将聚酰胺膜小心的从叶片表面剥离并置于激发波长为355nm,发射波长为460nm的紫外灯下,对聚酰胺膜进行拍摄,获得叶片酶谱图像;(3)标曲制作:配置一系列浓度(0,4,8,10,15,20,40,80,100,200,400μm)的4-muf溶液,以未添加4-muf的缓冲液为空白对照,将聚酰胺膜于缓冲液中浸泡8~12min;再将上述系列浓度的4-muf溶液滴加到风干的聚酰胺膜上,使用与s2酶谱成像相同的拍摄条件进行拍摄,获得标曲图像;(4)图像处理:将s3获得的标曲图像,原始图像转化为8-bit灰阶图,以σg=0.5的高斯滤波核进行降噪,再以灰度值为98作为全局阈值对所有降噪后的梯度浓度标准荧光灰阶图进行图像分割并计算灰度均值,最终得到扣除背景的梯度浓度标准荧光均值,然后根据不同浓度的标准荧光产物与其图像灰度的变化拟合出分段校正曲线:其中为施加于膜上的muf浓度;a1,a2,b1,b2是线性回归拟合的参数;g*为分断点处的灰度值;是经过分割的图像的平均灰度值;分段校正曲线的第一部分用于将酶谱中的像素灰度值转化为酶活性,并将校正曲线中的灰度值赋予成比例关系的rgb颜色,获得的rgb颜色条将用于创建查找表lut,用于生成伪彩色叶片酶谱图;将s2获得的叶片酶谱图像转化为8-bit灰阶图,以σg=0.5的高斯滤波核进行降噪,记录各图像四角5×5范围内的灰度均值,以此作为图像的背景灰度并在全图扣除得到最终的灰度图像,然后将lut应用于叶片图像,从而将叶片酶活性的空间分布可视化。本发明还保护上述任一所述方法在污染物胁迫下的植物毒性评价和生理反应检测中的应用。与现有技术相比,本发明具有以下有益效果:本发明提供了一种污染物胁迫下植物中酶活性的原位可视化测定方法,弥补了现有技术中植物叶片酶谱的技术空白。同时,本发明针对叶片酶谱,通过提供一系列的图像处理手段,重建叶片酶谱高质量图像从而提高其定性准确性的有用信息,具有较大的应用前景。附图说明图1为本发明开发的植物酶谱原位可视化方法的技术流程示意图。图2为本发明植物酶谱原位可视化方法中饱和底物膜孵育及反应示意图。图3为本发明植物酶谱原位可视化方法中图像处理流程示意图。图4为本发明植物酶谱原位可视化方法中标准曲线绘制。图5为本发明植物酶谱原位可视化方法中图像降噪结果;(a)fft图像的灰度径向分布图;(b)不同浓度muf下预处理对图像分析的影响。图6为实施例1中dbp胁迫下植物叶片酶谱图及热区分析:(a)木聚糖酶;(b)磷酸酶;(c)β-葡萄糖苷酶。图7为实施例1中dbp胁迫下叶片酶活性沿主叶脉的变化。图8为实施例1中dbp胁迫下叶片总酶活性的定量分析:(a)叶片酶谱法;(b)生化分析法。图9为不同浓度muf的标曲图像:(a)空域图;(b)频域图。图10为降噪处理后标曲图像的灰度直方图。具体实施方式以下结合说明书附图和具体实施例来进一步说明本发明,但实施例并不对本发明做任何形式的限定。除非特别说明,本发明采用的试剂、方法和设备为本
技术领域:
常规试剂、方法和设备。除非特别说明,以下实施例所用试剂和材料均为市购。本发明通过下述dbp胁迫下植物中葡萄糖苷酶、木聚糖酶和酸性磷酸酶的原位可视化测定来说明本发明的意图。实际研究时,根据研究目标选择环境胁迫类型、植物种类、酶种类等试验对象。实施例1dbp胁迫下植物中葡萄糖苷酶、木聚糖酶和酸性磷酸酶的原位可视化测定一、方法使用本发明开发的原位酶谱可视化方法表征dbp胁迫下植物叶片中葡萄糖苷酶、木聚糖酶和酸性磷酸酶的酶活性,方法流程如图1所示,具体包括如下步骤:1、植物种植和暴露以课题组模式植物绿宝菜心为研究对象,在人工气候箱中进行沙培(恒温25℃,相对湿度60%),每天浇灌改良的8692营养液(配方见表1)。在生长18天后,选择3个大小相似的预处理幼苗,并将其置于50mldbp(0、10.0、50.0和100.0mg/l)的离心玻璃管中。这些dbp溶液的浓度是根据三种不同的有效浓度(ec20、ec50和ec75)确定的,这三种效应浓度分别对应菜心幼苗的20%、50%和75%的相对生长抑制率。每个处理有四次生物重复。本研究使用了保证试剂邻苯二甲酸二丁酯(dbp,cas:84-74-2)。为了最大限度地减少水分蒸发,防止藻类生长,离心玻璃管覆盖了铝箔。所有幼苗均置于人工气候箱中暴露3天。表1no.reagentconc.no.reagentconc.1kno357.04g/l8mncl20.83g/l2mgcl2·6h2o12.00g/l9fe-edta4.38g/l3cacl218.00g/l10namoo435.00mg/l4mgso415.00g/l11cuso462.50mg/l5kh2po433.46g/l12znso414.95mg/l6nahco330.00g/l13cocl215.00mg/l7h3bo30.371g/l2、酶谱成像幼苗完成暴露后,将4-甲基伞形酮-β-d-葡糖苷(muf-g)、4-甲基伞形酮-β-d-磷酸酯(muf-p)和4-甲基伞形酮-β-d-木糖苷(muf-x)溶于100μl的二甲基亚砜并震荡至完全溶解后,加入0.1m的2-(n-吗啉)乙磺酸溶液(mes缓冲液)配置成浓度为100μm的酶底物缓冲液。上述3种底物可以分别与叶片中的β-葡萄糖苷酶、酸性磷酸酶和木聚糖酶发生反应,使底物中的猝灭剂脱落失效。将孔径为0.45μm的聚酰胺膜裁剪成3×5cm2大小,在配置的酶底物缓冲液中浸泡10min,风干2min后将其与不同浓度dbp暴露的幼苗叶片表面紧密贴合(每组设置4个生物重复)。使用亚克力板固定并在两面各施予1.0kg的压力(图2),于暗室中反应30min。反应结束后,将聚酰胺膜小心的从叶片表面剥离并置于遮光暗箱的黑色吸光布上进行拍摄。由于叶片上的酶促反应产物能够扩散到聚酰胺膜的对应位置并在特定紫外激发波长下发出与酶活性呈正相关的淡蓝色荧光,因此在暗箱中加装了两条激发波长为355nm,发射波长为460nm的紫外灯管用于拍摄过程中激发酶谱的荧光。紫外灯管的放置需要保证黑色背景的平场化。为保证图像分析的条件相同,酶谱拍摄过程中应使用相同的拍摄条件(fujifilmx-t20,xf18-55mm,f/2.8,1/125s,iso-12800)。得到的图像使用图像分析软件image-j进行分析。3、标曲制作为了能够定量分析酶谱图像表现出的叶片酶活性,本研究建立了一个以系列muf浓度酶谱标准图像获得定量标准曲线的方法。将4-甲基伞形酮(4-muf)溶解于100μl的二甲基亚砜并震荡至完全溶解后,加入0.1m的mes缓冲液并梯度稀释以获得0,4,8,10,15,20,40,80,100,200,400μm的系列标准荧光产物,未添加4-muf的mes缓冲液作为空白对照。将2×2cm2的聚酰胺膜在0.1m的mes缓冲液中浸泡10min并风干2min后取出。用移液枪吸取15μl各浓度的标准荧光产物滴加到风干的聚酰胺膜(n=3)的中央后,使用与酶谱成像相同的拍摄条件进行摄影。4、图像处理图像均使用image-j软件进行处理,流程如图3所示。对于获得的标曲图像,将每个校准图像裁剪为具有72900像素的正方形,并转换为8-bit图像,通过获得8-bit灰阶图中各像素点的灰度值来进行统计分析。由于摄影机的晃动以及取景器被动产生的噪点会影响图像分析的准确性,因此首先需要对灰阶图进行降噪处理。然而传统的利用黑色背景修正或直方图偏差降噪的方法无法去除随机出现的内部干扰噪点,会对标准曲线的拟合造成较大的误差。我们通过对原始图像进行观察,发现图像中出现的噪点较为均匀,因此考虑使用高斯滤波器应用于原始的8位标曲图像进行降噪增强处理:其中g(x,y)是二维卷积计算使用的高斯卷积核,σg是卷积矩阵中心衰减到e-0.5的矩阵半径。为兼顾对图像进行的高斯模糊修饰达到较好的降噪效果并避免边缘细节损伤,使用系列高斯卷积核(σg=0-9)对原始8-bit标曲图像进行卷积运算以优化降噪过程。同时,在频域内利用快速傅里叶变换(fft)得到与降噪后8-bit标曲图像相对应的32-bit频域幅值图像:其中对n×n像素图像内的灰度值进行离散傅立叶变换的结果,(kx,ky)是频域中的位置坐标;(x,y)是空域中的位置坐标。在频域中,变化快的噪声对应高频分量,平缓的背景对应低频分量,因此可以利用灰度在频谱图中的径向分布来评估不同σg的降噪效果。通过使用radialprofile工具比较不同σg降噪后图像的低频保留和高频消除情况,选择出标曲图像降噪的最佳σg值并对所有标曲图像进行降噪处理。随后,从没有添加4-muf的空白图像的灰度直方图中计算出空白图像灰度值的平均值(μ)和标准偏差(σ)。由于在最后荧光强度的计算过程中只需要膜中滴加有muf的部分,因此需要选择合适的阈值将标曲图像中muf和膜背景分割。而标曲图像在经过高斯滤波器降噪后,其灰度直方图表现出明显的正态分布,因此我们选择μ+σ处的灰度值作为图像分割的全局阈值以去除图像中至少84%的膜背景。所有标曲图像的分割过程都以空白图像μ+σ处的灰度值为参考,并计算分割后图像的平均灰度值通过对razavi等提出的方法进行修改,通过不同的muf浓度与图像灰度的变化拟合出分段校正曲线(图4),以分段线性回归的两个部分来描述灰度值和浓度之间的关系:其中为施加于膜上的muf浓度;a1,a2,b1,b2是线性回归拟合的参数;g*为分断点处的灰度值;是经过分割的图像的平均灰度值。分段校正曲线的第一部分可以将酶谱中的像素灰度值较为准确的转化为酶活性,并将校正曲线中的灰度值赋予成比例关系的rgb颜色,获得的rgb颜色条将用于创建查找表(lut),以生成伪彩色叶片酶谱图。对于叶片图像,在转换为8-bit灰度图像后,使用σg=0.5的高斯滤波器对叶片图像进行处理,在保留映射在膜上的叶片边缘的同时进行简单的平滑处理。通过分别在4个角处测量每个5×5像素的平均灰度值来获得每个叶片酶谱图的背景灰度值(gb)。为了标准化模糊的叶片酶谱图,引入了一个运算符δ来将其背景灰度值调整为校准线的空白灰度值b1:δ=gb-b1分别将所有叶片酶谱图中的像素灰度减去该运算符δ以获取最终的灰度图像。随后,将lut应用于叶片图像将叶片酶活性的空间分布可视化。5、数据建模分析为了计算酶活性从叶基到叶尖的分布趋势,我们使用plotprofile工具选取了沿叶脉1个像素宽的线段,并记录了线段上的灰度变化。我们将叶片酶谱中灰度值超过校正曲线第一部分75%的区域称为“热区”,通过计算叶片酶谱中热区面积的变化来研究酶活性较高区域在paes胁迫下空间分布的变化。此外,我们还使用传统的紫外分光光度法对paes胁迫下叶片中β-葡萄糖苷酶、酸性磷酸酶和木聚糖酶的活性进行分析,并对叶片酶谱结果进行验证。本发明的数据分析使用spss20.0(spss,chicago,illinoisusa)进行双向方差分析,多重比较采用邓肯后检验(p<0.05)。最终数据以均值±标准差来表示。所有柱状图都是使用originpro9.0绘制的。在图像处理方法的步骤中均使用imagej工具。二、结果1、图像前处理叶酶谱分析技术的关键步骤包括:1)化学暴露,2)饱和荧光底物膜的孵育,3)固定的相机设置和摄影条件,4)校准研究(图1)。其中,图像处理是一个重要的问题,它会对所有后续统计分析的结果产生深远影响。但是,先前的研究缺乏有关如何重建高质量图像从而提高其定性准确性的有用信息。我们将10个不同半径的高斯滤波器内核应用于原始的8-bit标曲图像以选择合适的σg进行降噪,并获得了由不同σg的高斯滤波器滤波后的fft图像的灰度径向分布图(图5a)。结果表明,频域中的幅度通常随着半径的增加而减小,这是由于高斯滤波核在空域中的卷积会在fft图像中表现出低通滤波器的特点。低通滤波器可以衰减高频信号,因此可以减小噪声对图像平均灰度的影响。当σg<3时,大量包含噪声的高灰度值像素以及细节信息仍保留在空域图像中(σ=19.4~3.8)。对应的fft图像显示高频信号逐渐减小。空域中的噪声表示为频域中的高频信号,该信号可以通过傅立叶频谱进行定性分析。从σg=3~5,滤波效果进一步增强,图像在空域中的标准偏差减小(σ=2.8~1.9)。对应的fft图像中的高频信号被进一步衰减,并且在σg=5时实现了最强的降噪效果。当σg=6~9时,图像在空域中变得更平滑,标准偏差的变化已经很小(σ=1.9~1.4)(图5a,图9)。为了实现强力的降噪效果,对象和背景之间一定程度的边缘平滑是可以接受的。综上所述,高斯滤波器核σg的半径固定为5,并应用于所有标曲图像。为了获得muf的真实荧光强度,在计算标曲图像的灰度平均值之前必须进行图像分割。这里我们使用了全局阈值的方法来分割背景和前景。灰度直方图显示,降噪后标曲图像的直方图由一个主峰和不同数量的副峰组成(图10)。参考空白图像,可以得知直方图中的主峰代表了背景的荧光强度。因此,降噪后的空白图像的平均灰度值为95±3。考虑到灰度低于该平均值的像素点不包含任何校准研究信息,而使用高斯滤波器后灰度直方图的背景强度呈正态分布,因此将阈值选择为98(95+3)以除去至少84%的背景像素。图像获取和预处理过程中的缺陷可能会导致图像信息的误读。然而,在先前关于荧光成像的研究中并没有或仅仅简要提及图像预处理过程的质量控制。在本发明中,与没有分割的图像相比,去噪后的标准偏差值正在减小(图5b)。同时,由于高斯核卷积给出的结果是基于邻域的线性输出,这使得降噪前后的平均灰度值变化很小。但是该区域中包含了很多噪声,这些噪声没有被分割,为灰度的平均计算带入了较大的误差。如果对图像进行分割而不进行降噪,由于直方图中没有明显的聚类,直接进行图像分割导致阈值附近像素的失真将不容忽视。因此,图像去噪和分割对于酶谱图像的分析都是必不可少的。2、叶片酶活性的空间分布在本发明中,选择木聚糖酶,β-葡萄糖苷酶和磷酸酶作为生物指示剂,以确定dbp对绿宝菜心幼苗的毒性。酶谱图显示,酶的最高活性主要位于主叶脉,并逐渐衰减至边缘(图6)。木聚糖酶和β-葡萄糖苷酶的活性主要分布在叶片主叶脉中,而磷酸酶活性的空间分布似乎分布在整叶中。显然,dbp暴露(3天)可以改变植物叶片中木聚糖酶,β-葡萄糖苷酶和磷酸酶活性的空间分布。然而,大多数利用荧光成像方法的研究都是定性的。因此,应该建立一种合适的方法,将像素灰度值转换为酶活性,以收集和定量分析此类图像。图像分割后计算出校准图像的灰度平均值。我们使用分段线性回归来描述灰度值与muf浓度之间的关系,结果如下:y1=2.07x+95.17(r2=0.997)和y2=0.59x+124.48(r2=0.635)。该结果表明高浓度的muf(≥100mm)可能导致膜过饱和,构成“不确定区域”。因此,我们选择分段曲线的第一部分作为“安全区”以进一步定量叶片中的酶活性。为了更好地说明沿主叶脉酶活性的空间分布,在每个图像中沿主叶脉取一个像素宽的线段,以计算木聚糖酶,β-葡萄糖苷酶和磷酸酶的活性(图7)。叶片酶谱分析揭示了三种酶分布的特定模式,是从叶基到叶尖距离的函数。木聚糖酶沿叶脉的分布呈下降趋势。而β-葡萄糖苷酶和磷酸酶沿叶脉的分布呈反u形曲线。这些结果表明,叶基中木聚糖酶,β-葡萄糖苷酶和磷酸酶的活性高于叶尖。为了量化酶活性的敏感性水平,可以将拟合曲线积分的面积用作评估dbp毒性作用的敏感生物指标(表2)。表2热区分析结果表明,叶基中木聚糖酶,β-葡萄糖苷酶和磷酸酶的敏感性值均高于叶尖,而叶基中β-葡萄糖苷酶活性的敏感性值最高。暗示可以选择叶基中的β-葡萄糖苷酶活性作为dbp胁迫的敏感生物指示剂。主要的生物地球化学过程发生的微型地点,被称为“热区”。在这项研究中,木聚糖酶,β-葡萄糖苷酶和磷酸酶的四种区域类别(极低,低,中和高热区)对酶谱区域总活性的贡献如图6和表3所示:表3敏感性分析(y=a0+a1*x+a2*x^2+a3*x^3+a4*x^4)值得注意的是,β-葡萄糖苷酶和磷酸酶的热区比木聚糖酶的热区显着(p<0.05)。在0、10、50和100mgdbp/l处理下,木聚糖酶的热区分别占总木聚糖酶活性面积的4.23%,1.44%,2.66%和1.33%。在不同的dbp处理下,β-葡萄糖苷酶的热区分别占总β-葡萄糖苷酶活性区域的13.44%,17.34%,30.52%和27.64%。磷酸酶热区的定位与β-葡萄糖苷酶类似。总体而言,与对照相比,dbp处理显着增加了叶片中β-葡萄糖苷酶和磷酸酶的热区区域。而木聚糖酶的趋势相反,这表明dbp处理会导致植物叶片中的β-葡萄糖苷酶和磷酸酶的活性升高,但抑制木聚糖酶的活性。这种分布模式主要归因于dbp的植物毒性和基因表达的响应。这些结果表明,在dbp压力下,使用热区作为生物指标具有很大的潜力。3、叶片总酶活性先前的研究表明,木聚糖酶的活性对小麦根部和枝条中的as(iii)胁迫都非常敏感。β-葡萄糖苷酶可以调节脱落酸(aba)的含量,从而减轻拟南芥的干旱胁迫。而在大麦根部,在铝处理期间可以观察到酸性磷酸酶活性显着增加。然而,前人很少关注dbp胁迫下植物中木聚糖酶,β-葡萄糖苷酶和磷酸酶活性的变化。在本发明中,叶片总酶活性被定义为叶片每个像素点的酶活性总和除以叶片的面积(图8a)。我们发现,随着paes浓度的增加,在叶片酶谱中观察到磷酸酶和β-葡萄糖苷酶总酶活性的增加,而木聚糖酶则明显下降。为了验证全叶中的酶活性,在这项研究中对木聚糖酶,磷酸酶和β-葡萄糖苷酶进行了生化分析(图8b)。正如预期的那样,木聚糖酶,磷酸酶和β-葡萄糖苷酶活性的变化趋势与叶片酶谱分析结果相似,表明叶酶谱分析技术是可靠的分析方法。我们基于叶片酶谱技术对木聚糖酶,β-葡萄糖苷酶和磷酸酶的活性进行了定量分析,表现了在环境胁迫下叶片酶谱在酶活性趋势分析中的优势。以上所述,仅是本发明的较佳实施例而已,并非对本发明作任何形式上的限制,依据本发明的技术实质对以上实施例所作的任何简单修改、等同变化与修饰,均仍属于本发明技术方案的范围内。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1