用于富集循环肿瘤DNA的方法与流程
发明领域
本发明涉及一种方法,其用于从血液、血清或血浆中纯化或富集肿瘤来源的循环无细胞核小体及相关的循环肿瘤dna。
发明背景
细胞dna以称为染色质的蛋白质-核酸复合物形式存在。核小体为染色质结构的基本单元,且由缠绕在蛋白质复合物周围的双链dna(dsdna)而组成。所述dna以常常称为类似“串珠”的结构缠绕在连续的核小体周围,这形成开放染色质或常染色质的基本结构。在压缩染色质或异染色质中,所述绳串以紧密且复杂的结构为螺旋的和超螺旋的。
染色质中的各个核小体由八个高度保守的核心组蛋白(包含各一对组蛋白h2a、h2b、h3和h4)的蛋白质复合物组成。围绕该复合物缠绕着约146个碱基对(bp)的dna。据报道无细胞核小体包含160-200bpdna,其可能是因为存在另外的接头dna。另一种组蛋白h1或h5,充当接头并涉及染色质压缩。
成年人中的正常细胞更新包括通过细胞分裂每天产生约1011个细胞并主要通过凋亡使类似数量死亡。在凋亡的过程中,染色质分解成单核小体和寡核小体,其中的某些可存在于循环中。在正常条件下,在健康受试者中发现的循环核小体的水平报道为较低的。在具有多种病况的受试者中发现升高的水平,所述病况包括许多癌症、自身免疫性疾病、炎性病况、中风和心肌梗死(holdenreider&stieber,2009)。
dna异常为所有癌症疾病的特征。癌细胞的dna在许多方面都与健康细胞的dna不同,包括但不限于点突变、易位、基因拷贝数、微卫星异常、dna链完整性和核苷酸修饰(例如第5位胞嘧啶的甲基化)。在dna结构或序列上的这些肿瘤相关的改变,通常在活检或手术时移除的癌细胞或组织中进行研究,以用于临床诊断、预后和治疗目的。肿瘤遗传特征和表观遗传特征,在不同的肿瘤类型之间以及在患有相同肿瘤疾病的不同患者之间均不同。此外,随着所述疾病进展以及在形成对药物或其他疗法的获得性耐药期间,在相同患者的相同癌症中,这些特征亦随时间改变。因此,对手术或活检时移除的细胞中的肿瘤dna的连续研究,可帮助临床医师监测疾病进展并在早期(可能比放射学检测早许多个月)检出任何复发或获得性治疗耐药性,并允许疗程中有可能的成功的变化。
然而,组织dna检测具有局限性,因为不能对患者反复进行侵入性活检程序来用于监测目的。对于一些患者而言,根本不能使用活检。进行活检很昂贵,对患者而言为不适的,造成患者风险,且可能导致手术并发症。此外,患者中的肿瘤可由位于相同肿瘤的不同区域内或不同转移内(在转移癌中)的多个肿瘤样克隆组成,其并非全部都可在活检上取样。因此,在处于时间上的特定时刻的位于肿瘤的不同区域内的不同肿瘤克隆之中,组织活检dna研究同时在时间上和空间上提供肿瘤的快照(snap-shot)。
癌症患者的血液含有循环肿瘤dna(ctdna),认为其来源于从濒死或死亡的癌细胞释放到循环中核小体或染色质片段。来自癌症患者的匹配血液和组织样品的研究显示,存在于患者肿瘤中(但不存在于他/她的健康细胞中)的癌症相关的突变,亦存在于获自相同患者的血样的ctdna中(newman等,2014)。类似地,在癌细胞中差异甲基化(通过胞嘧啶残基的甲基化在表观遗传上改变)的dna序列,在循环的ctdna中亦可检测为甲基化的序列。此外,由ctdna组成的无细胞循环dna(cfdna)的比例与肿瘤负荷相关,因此可通过ctdna存在的比例定量监测同时通过其遗传和/或表观遗传组成定性监测疾病进展。ctdna的分析可产生关于起源于肿瘤内的所有或许多不同克隆的dna的非常有用且临床上精确的数据,并其因此在空间上整合所述肿瘤克隆。此外,随时间的重复取样为远更实际且经济的选项。(ctdna)的分析具有变革肿瘤的检测和监测的潜力,以及在早期检出复发和获得性耐药,用以通过不使用侵入性组织活检程序的肿瘤dna研究来选择用于肿瘤的治疗。所述ctdna检验可用于研究所有类型的癌症相关的dna异常(例如点突变、核苷酸修饰情况、易位、基因拷贝数、微卫星异常和dna链完整性),且对于常规癌症筛查、定期和更频繁的监测以及定期检查最佳治疗方法而言应具有适用性(zhou等,2012)。
血液、血浆或血清可用作ctdna测定的基质,且可使用任何dna分析法,包括但不限于dna测序、表观遗传dna测序分析(例如对于含5-甲基胞嘧啶的序列)、pcr、beaming、ngs(靶定基因组或全基因组)、数字pcr、冷pcr(较低变性温度下的共扩增-pcr)、map(midi-激活的焦磷酸解)、pare(重排末端的个性化分析)和质谱法。
因为dna异常为所有癌症疾病的特征,且对于其中已进行研究的所有癌症疾病中均观察到ctdna,所以ctdna检验在所有癌症疾病中均具有适用性。研究的癌症包括但不限于膀胱癌、乳腺癌、结肠直肠癌、黑素瘤、卵巢癌、前列腺癌、肺癌、子宫内膜癌、卵巢癌、淋巴瘤、口腔癌、白血病、头颈癌和骨肉瘤(crowley等,2013;zhou等,2012;jung等,2010)。现在将通过概述三个(非限制性的)示例方法来阐述ctdna检验的特性。
第一个实例包括检测ctdna中的一种癌症相关的基因序列突变。包括检测ctdna中的单基因突变的血液检验,一般具有低临床灵敏性。对此有两个原因。首先,尽管所有癌症均具有突变,但特定癌症疾病中任何特定突变的频率通常很低。例如,尽管k-ras和p53突变被认为是更高频癌症突变中的两个,且已在包括膀胱癌、乳腺癌、结肠癌、肺癌、肝癌、胰腺癌、子宫内膜癌和卵巢癌在内的大范围的癌症中进行研究,但其分别在23%-64%和17%-54%的癌组织样品中检出。其次,即便患者的癌组织确实含有所述突变,但存在于所述患者血液中的突变ctdna的水平或浓度可能很低且难以检出。例如,k-ras和p53突变可在0%-75%的k-ras和p53组织阳性患者的ctdna中检出。这两个结果的总和,意味着k-ras或p53突变在小于40%的癌症患者的血液中检出(jung等,2010)。
第二个实例包括检测ctdna中的多个癌症相关的基因序列突变。尽管诸如k-ras或p53等任何特定基因的突变可能仅存在于少数癌症中,但所有癌症均含有突变,所以研究足够大量的突变在原则上应利于检出大多数或甚至所有肿瘤。因此,增加所述检验的临床灵敏性的一个方式为检测许多基因中的大范围的突变。newman等采用该方法用于非小细胞肺癌(nsclc)并研究了来自139个周期性发生的突变基因的521个外显子和13个内含子序列。研究的突变包括多个种类的癌症相关的遗传改变,包括单核苷酸变异(snv)和融合基因。以此方式,作者报道检出大于95%的ii-iv期肿瘤和大于50%的i期肿瘤,在ctdna血液检验中以96%特异性(newman等,2014)。
第三个实例包括检测对于ctdna中的特定基因序列的癌症相关的表观遗传改变。此方法可应用于任何dna或核苷酸修饰。此方法的主要实例为检测在某些癌症中其在胞嘧啶残基上为差异甲基化的基因。对此目的,在多种癌症中研究了大量基因。这些基因中的一些为在膀胱癌、乳腺癌、结肠直肠癌、黑素瘤、卵巢癌和前列腺癌中研究的septin-9、apc、dapk、gstp1、mgmt、p16、rassf1a、t1g1、brca1、erα、prb、tms1、mlh1、hltf、cdkn2a、socs1、socs2、pax5、pgr、ptgs2和rarβ2。此方法的一个示例性实例为检测ctdna中的甲基化septin-9以检测结肠直肠癌(crc),其报道为检出68%的crc病例,以89%的临床特异性(grutzmann等,2008)。
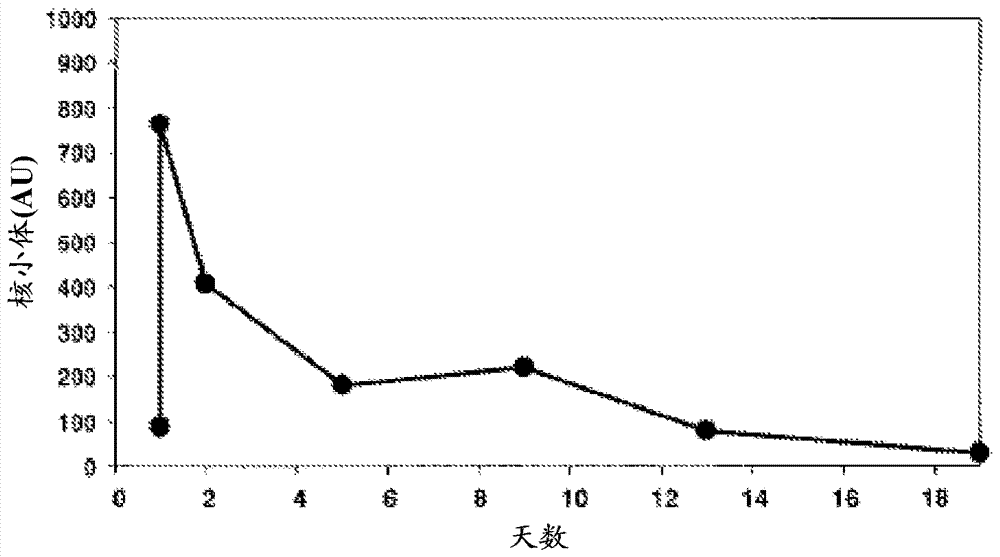
所述cfdna的肿瘤来源的ctdna流分,作为中值长度为约180个碱基对(bp)的小dna片段循环,所述中值长度为对于以单核小体的形式循环的dna片段所预期的大小(newman等,2014)。认为这些180bpdna片段包括核小体dna加上某些接头dna。据报道,癌症患者比健康受试者具有更高的cfdna水平。本领域工作人员曾报道对于健康受试者0-100ng/ml(平均30ng/ml)cfdna的范围以及对于患有癌症的受试者0-1000ng/ml(平均180ng/ml)cfdna的范围(schwarzenbach等,2011)。循环cfdna由长度高达20,000个碱基对的不同大小的dna分子组成(zhou等,2012)。与ctdna主要作为单核小体循环的假说一致,在癌症患者中测得的循环中的无细胞核小体的水平,如dna水平,高于健康受试者中的水平(holdenrieder等,2001)。然而,升高的自身循环核小体水平并未在临床上用作癌症的生物标志物,因为核小体为细胞死亡的非特异性产物,且对于涉及增加的细胞死亡的许多情况,包括急性创伤,均观察到升高的水平(holdenrieder和stieber,2009)。作为细胞死亡的产物,在使用细胞毒性药物或放疗治疗之后,循环核小体水平可显著升高。然而,核小体亦被从循环中清除,所以水平可随治疗达到峰值并随后下降,如在从holdenrieder等,2001重现的图1和2中所示。
尽管自身循环无细胞核小体的水平并未在临床实践上用作癌症中的基于血液的生物标志物,但是就其组蛋白修饰、组蛋白变体、dna修饰和加合物含量方面而言循环无细胞核小体的表观遗传组成,已作为癌症中的基于血液的生物标志物进行研究(wo2005/019826;wo2013/030577;wo2013/030579;wo2013/084002)。
cfdna的生物学来源并未充分了解。产生单核小体和寡核小体的染色质的片段化,为凋亡性细胞死亡的特征。虽然认为坏死细胞产生长度为数千个碱基对的较大dna分子,但dna片段化亦可在一些坏死的情况下发生。另外,常见的dna重复序列(例如alu或line1序列)可作为200-400碱基对dna片段从正在经历非凋亡性或坏死性细胞死亡的细胞中释放(schwarzenbach等,2011)。dna片段亦可作为胞间交流的形式通过细胞分泌。认为ctdna的来源与癌细胞的死亡相关。dna片段可作为核小体从坏死和/或凋亡的肿瘤细胞中释放。然而,坏死和凋亡的细胞通常被巨噬细胞或其他清除细胞吞噬,且dna可通过吞入了坏死或凋亡细胞的巨噬细胞释放(schwarzenbach等,2011)。
存在可用于从血液、血清或血浆中提取cfdna的多种方法,且已针对提取dna的产率和其提取不同长度的dna片段的效率进行比较。酚-氯仿和碘化钠提取法提供最高产率且提取长度小于200bp的小dna片段。测试的其他方法(包括市购可得的方法)报道为具有较低的dna提取率且未能提取长度小于200bp的小dna片段(fong等,2009)。
从血液、血清或血浆中提取cfdna用于ctdna的分析,通常使用市购可得的dna提取产品进行。所述提取法声称具有较高的循环dna回收率(>50%),且某些产品(例如qiagen生产的qiaamp循环核酸试剂盒)声称提取小尺寸的dna片段。使用的典型样品体积范围为1-5ml的血清或血浆。
因为许多限制,目前没有用于临床肿瘤学目的的常规使用的基于ctdna的检验。主要的方法学限制为需要高质量dna。因为样品的特性,当前的ctdna取样法得到低质量的ctdna样品。主要困难在于在循环中存在大量的非肿瘤cfdna,其使ctdna的任何分析变复杂。尽管来自不同工作者的评价不同,但存在于循环中的ctdna的流分可能太低而无法检出或者高于50%的cfdna。然而,对于大多数癌症患者而言,所述ctdna流分为cfdna的一小部分。例如,近期研究报道,在预治疗肺癌患者中,ctdna流分随肿瘤大小增加。虽然在具有较大肿瘤负荷的患者中发现的最高水平为3.2%,但发现大多数患者具有低于0.1%的ctdna流分(newman等,2014)。这意味着对于许多患者样品而言,必须在存在远远更高水平的非肿瘤来源dna的情况下分析非常低水平的ctdna。此外,该dna来自相同受试者并因此就有类似序列,且在用于定量或分析ctdna的任何方法中均会产生干扰。
对于测量作为癌症的生物标志物的循环无细胞核小体和/或循环核小体的表观遗传学组成而言存在类似的问题,这是因为自身核小体为细胞死亡的非特异性指示物,且作为身体的正常细胞更新过程的部分而释放,以及在与升高的细胞死亡水平相关的情况下释放,所述情况例如自身免疫性疾病、中风、脓毒症、创伤后、烧伤、心肌梗死、脑卒中,器官移植之后的移植物排斥期间和剧烈锻炼之后的病况。因此,肿瘤来源的核小体与各种细胞和组织来源的其他非肿瘤核小体一起循环。这些非肿瘤核小体将在用于肿瘤来源的核小体的定量或表观遗传分析的任何方法中产生干扰。
因此,对用于从血液、血清或血浆样品中富集肿瘤来源的循环核小体和ctdna的方法,存在极大需求。
发明概述
根据本发明的第一方面,提供组蛋白h3.1和/或h3.2和/或h3t结合剂用于从生物样品中检测、分离和/或纯化肿瘤来源的无细胞核小体或循环肿瘤dna(ctdna)的用途。
根据本发明的又一方面,提供通过亲和纯化从生物样品中分离肿瘤来源的循环无细胞核小体的方法,其中所述方法包括步骤:
(i)使所述样品与组蛋白h3.1和/或h3.2和/或h3t结合剂接触;
(ii)从所述样品中分离结合的核小体;和
(iii)分析所述分离的核小体和/或相关dna。
根据本发明的又一方面,提供用于从生物样品中分离纯化的循环肿瘤dna(ctdna)的方法,其中所述方法包括步骤:
(i)分离含组蛋白3.1和/或h3.2和/或h3t的循环无细胞核小体;
(ii)从在步骤(i)中产生的核小体样品提取dna;和
(iii)分析所述提取的dna。
根据本发明的又一方面,提供用于在生物样品中检测肿瘤来源的循环核小体的表观遗传表位的免疫测定法,其中所述方法包括步骤:
(i)使所述样品与组蛋白h3.1和/或h3.2和/或h3t结合剂接触;
(ii)使所述核小体或样品与第二结合剂接触,所述第二结合剂与所述表位结合;
(iii)检测和/或定量所述第二结合剂与所述表位的结合;和
(iv)使用所述结合的存在或程度作为所述样品中的肿瘤来源核小体的特定表位的存在的测量结果。
根据本发明的又一方面,提供用于在生物样品中检测肿瘤来源的循环核小体的表观遗传表位的免疫测定法,其中所述方法包括步骤:
(i)使所述样品与第一结合剂接触,所述第一结合剂与所述表位结合;
(ii)使所述核小体或样品与组蛋白h3.1和/或h3.2和/或h3t结合剂接触;
(iii)检测和/或定量所述组蛋白h3.1和/或h3.2和/或h3t结合剂与所述样品中的核小体的结合;和
(iv)使用所述结合的存在或程度作为所述样品中的肿瘤来源核小体的特定表位的存在的测量结果。
根据本发明的又一方面,提供诊断癌症的方法,其包括在获自人类或动物受试者的生物样品中检测循环无细胞核小体相关的组蛋白变体h3.1和/或h3.2和/或h3t的步骤。
附图简述
图1:在患有头颈癌、用16次日剂量放疗的患者中的自身无细胞核小体的循环水平。使用市购的核小体elisa,用涉及与核心核小体表位结合的固定化抗体来测量水平(复制自holdenrieder等,2001)
图2:在患有小细胞肺癌、用细胞毒性化疗第1天的患者中的自身无细胞核小体的循环水平。使用利用涉及与核心核小体表位结合的固定化抗体的市购的核小体elisa,在大于42天时间内测量水平(复制自holdenrieder等,2001)
图3:在术前以及术后6、24、48、72和96小时时6个患有crc的患者中的自身无细胞核小体的循环水平以及含组蛋白变体h3.1和/或h3.2和/或h3t的无细胞核小体的循环水平。使用利用涉及与核心核小体表位结合的固定化抗体的市购的核小体elisa来测量自身无细胞核小体的水平。使用类似的但使用涉及与组蛋白变体h3.1、h3.2或h3t结合的固定化抗体测定法测量含组蛋白变体的循环无细胞核小体的水平。
图4:在术前以及术后6、24、48、72和96小时时6个患有crc的患者中的自身无细胞核小体的循环水平和含有组蛋白变体h3.1、h3.2或h3t和5-甲基胞嘧啶(5mc)的无细胞核小体的循环水平。使用利用涉及与核心核小体表位结合的固定化抗体的市购的核小体elisa来测量自身无细胞核小体的水平。使用采用涉及与组蛋白变体h3.1、h3.2或h3t结合的固定化抗体和涉及与5mc结合的标记抗体的测定来测量含组蛋白h3变体和5mc的循环无细胞核小体的水平。
图5:健康患者和患有crc的患者中的含(i)组蛋白变体h3.1、h3.2或h3t和组蛋白修饰h3k27ac二者或(ii)组蛋白变体h3.1、h3.2或h3t和5-甲基胞嘧啶(5mc)二者的无细胞核小体的循环水平。将含组蛋白h3变体以及组蛋白修饰h3k27ac的循环无细胞核小体的水平相对于含5mc的水平按一定比率表达,得到在获自健康受试者和患有crc的受试者的血样之间改进的鉴别力(iii)。
图6:新诊断为前列腺癌的患者和类似年龄的健康对照受试者中的含(i)组蛋白变体h3.1、h3.2或h3t和5-甲基胞嘧啶二者或(ii)含5-甲基胞嘧啶(5mc)的自身核小体的无细胞核小体的循环水平。对于含(iii)a,组蛋白变体h3.1、h3.2或h3t和5-甲基胞嘧啶二者或(iii)b,含5-甲基胞嘧啶(5mc)的自身核小体的患者,亦将数据显示为箱线图(boxplot)。
发明详述
核小体的结构,就其表观遗传信号组成而言,在癌细胞中可不同。涉及与表观遗传信号结合的抗体或其他结合剂的使用,对于结合获自受试者的生物样品中的肿瘤来源的无细胞核小体,可具有选择性,所述表观遗传信号在癌细胞中比在其他细胞中更常见,所述生物样品含有伴随细胞来源的混合物的无细胞核小体。
根据本发明的第一方面,提供组蛋白h3.1和/或h3.2和/或h3t结合剂用于从生物样品中检测、分离和/或纯化循环肿瘤dna(ctdna)的用途。
在一个实施方案中,肿瘤来源的无细胞核小体为分离的和/或纯化的。
患者样品中的ctdna常常以非常低或不可检测的浓度存在,并且此外仅包含存在的cfdna的一小部分。如果能得到质量更好的样品,则基于ctdna分析的癌症检验的临床性能和实用性将得到改进。类似地,肿瘤来源的循环无细胞核小体作为具有各种来源的核小体的混合物的一部分进行循环且仅包含存在的无细胞核小体的一部分。出人意料的是,我们在此显示,从血液、血清或血浆样品中富集或分离肿瘤来源的核小体,连同cfdna的核小体相关ctdna流分,可使用亲和纯化分离法进行。
holdenrieder已报道的是,升高的循环无细胞核小体水平为恶性和良性肿瘤疾病的特征(holdenrieder等,2001)。然而,这对于癌症检测并没有用,因为循环核小体水平为细胞死亡的非特异性标志物且会在多种病况下升高,包括自身免疫性疾病、中风、脓毒症、创伤后、烧伤、心肌梗死、脑卒中、器官移植之后的移植物排斥期间和剧烈锻炼之后的病况(holdenrider&stieber,2009)。holdenrieder使用elisa技术测量了循环核小体,其中使用的第一固定化抗体涉及与常见核小体核心表位结合,而第二标记抗体涉及结合双链dna。该elisa法设计成检测所有核小体或核小体自身,连同相关的dsdna而不论表观遗传结构。
循环核小体水平可在由任何数目的不同原因包括创伤、中风或使用细胞毒性药物或放疗的治疗造成的细胞死亡中突然增加之后的2-5天明显达到峰值。然后水平在2-3天的时间内下降,如在图1和2中所示(重现自holdenrieder等,2001)。此结果是因为诱导细胞死亡接着将其从循环中清除(holdenrider&stieber,2009)。
使用利用涉及与常见核小体核心表位结合的相同固定化抗体以及涉及与双链dna结合的相同的标记抗体的相同的市购可得的核小体elisa(holdenrieder等使用的),我们进行了类似试验来检测所有核小体而不论表观遗传情况。在这些试验中,我们在术前以及术后6、24、48、72和96小时时在6个患有crc患者中测量了循环无细胞核小体水平。与holdenrieder的研究结果一致,在各例中均观察到在核小体水平中的术后升高,如在图3中所示。所述在核小体水平上的升高的详细时间,以及任何后续下降的时间,在各患者之间不同。
清楚的是,释放到未患肿瘤疾病的患者的循环中的核小体(包括例如因手术创伤所致),不会具有肿瘤来源。这些核小体将有助于(contribute)cfdna但不含ctdna。本领域技术人员亦应清楚的是,在我们自己的试验中术后释放的核小体可能具有肿瘤来源,但亦可能是因为手术的创伤而具有非肿瘤来源。具有肿瘤来源的所述cfdna的流分(即ctdna流分)可确定为所述cfdna的等位基因流分,其含有肿瘤相关的突变。我们开发了一种方法,藉此可从其他核小体和cfdna中富集或分离含ctdna的肿瘤来源的循环核小体。
我们使用第二种elisa法重新测定了获自6个crc患者术前和术后的样品。所述第二种elisa法使用与holdenrieder所用的市购可得的方法相同的涉及与双链dna结合的标记抗体,但使用涉及与组蛋白变体h3.1、h3.2或h3t结合的固定化抗体。结果显示,在含h3.1、h3.2或h3t的核小体水平上对手术的响应,与使用采用涉及与常见核小体核心表位结合的抗体的elisa对核小体自身观察到的响应不类似。含这些h3变体的循环核小体的水平与(总)循环核小体自身的水平具有不同的响应特征,且受手术影响较小(图3)。
多种表观遗传修饰的核苷酸已描述于文献中,且已知dna和/或dna核苷酸残基中的表观遗传修饰模式在癌症中改变。这些表观遗传修饰中阐述最充分的包括第5位胞嘧啶的甲基化。含5-甲基胞嘧啶的dna常常称为甲基化dna。我们开发了第三种elisa法并对相同的6个crc患者实施所述方法。该elisa为用于测量含有掺入5-甲基胞嘧啶残基的dna的核小体的方法,如同描述于wo2013/030577的用于含表观遗传修饰的核苷酸的dna测定法的实例,但使用固定化抗-h3.1、h3.2或h3t抗体作为捕获抗体。因此,所述第三种elisa法检测含组蛋白变体h3.1、h3.2或h3t以及5-甲基胞嘧啶残基二者的无细胞核小体,且因此类似于上述第二种elisa法,但使用不同的标记抗体(涉及与5-甲基胞嘧啶结合,而不是与dsdna结合)。对于所述6个crc患者的结果显示,与(总)循环核小体自身的水平相比,h3变体核小体相关的5-甲基胞嘧啶的水平较低且受手术影响较小(图4)。
此外,我们使用此测定法在健康受试者和患有crc的受试者中测量了循环核小体中共存的组蛋白变体h3.1、h3.2或h3t与5-甲基胞嘧啶甲基化dna水平。癌症受试者中甲基化dna的水平低于健康受试者中的水平。此研究结果与发现dna在癌细胞中为总体低甲基化的出版文献一致。与健康细胞的dna相比,预计癌细胞中dna的甲基化降低约50%(guerrero-preston等,2007;soares等,1999)。然而,在循环核小体的水平上的癌症相关的增加报道为平均970%(holdenrieder等,2001),和在cfdna上的增加为约600%(schwarzenbach等,2011)。尽管在含甲基化dna的循环无细胞核小体的比例上有(小量)下降,但预期在总cfdna上的大量增加可能会导致在循环核小体相关的甲基化dna的绝对水平上的总体上升。然而,使用针对组蛋白变体h3.1、h3.2或h3t的捕获抗体的本测定的结果,显示在癌症患者中在含甲基化dna(和组蛋白h3变体)的循环核小体的绝对水平上的降低,表明在(总)循环核小体自身测定中观察到的大部分癌症相关的上升,并非为肿瘤细胞来源的。此外,我们已显示所述h3变体核小体相关的5-甲基胞嘧啶测定,对于在血液检验中检测癌症而言为有效的,这表明通过其鉴定的全部或大部分无细胞核小体为肿瘤来源的。显示对癌症患者和健康患者的所述测定的结果的箱线图在图5中显示。
我们已开发和进行了第四种elisa法,其用于测量含组蛋白变体h3.1、h3.2或h3t和组蛋白修饰h3k27ac(组蛋白h3的乙酰化赖氨酸27)的循环无细胞核小体。这与类似于文献(wo2005/019826)中描述的用于含表观遗传修饰的组蛋白的核小体的测定法,但使用固定化抗-h3.1、h3.2或h3t抗体作为捕获抗体。所述第四种elisa与上述第三种elisa法相同,只是其使用涉及与h3k27ac(而不是5-甲基胞嘧啶)结合的不同的标记抗体。我们类似地使用所述第四种测定法在健康受试者和患有crc的受试者中测量相同的无细胞循环核小体中共存的h3.1、h3.2或h3t和h3k27ac水平。癌症受试者中的h3k27ac水平高于健康受试者中的水平。此研究结果与发现h3k27的乙酰化在癌细胞的染色质中升高的出版文献一致(karczarski等,2014)。此外,我们已显示此测定法对于在血液检验中检测癌症而言为有效的,这表明通过其鉴定的全部或大部分无细胞核小体为肿瘤来源的。显示对癌症患者和健康患者的所述测定的结果的箱线图在图5中显示。所述测定结果在获自健康受试者和患有癌症的受试者的血样之间存在差别。当本文所述的第三种和第四种elisa测定法的结果分别用于含组蛋白h3变体以及5-甲基胞嘧啶(5mc)或h3k27ac的表观遗传修饰的循环无细胞核小体时,在患有癌症的受试者和健康受试者之间的鉴别力得到进一步增强。这些结果显示,用于含组蛋白修饰h3k27ac以及组蛋白变体h3.1、h3.2或h3t二者的循环无细胞核小体的测定法,在临床肿瘤学上为有用的,包括但不限于用于检测癌症,以及用于预后预报、疗法选择、患者监测和复发监测/检测。用于核小体相关h3k27ac的测定法可独立进行或者作为包含表观遗传检验和/或其他检验的测定组的一部分。
为了进一步证实所述含组蛋白变体h3.1、h3.2或h3t的循环核小体流分富含肿瘤来源的核小体且对于肿瘤学血液检验而言为有用的,对于获自新诊断为前列腺癌的9个男性和类似年龄的10个健康男性的样品,我们使用固定化抗h3.1、h3.2或h3t抗体作为捕获抗体和标记的抗5-甲基胞嘧啶抗体进行elisa法(即上述第三种elisa法),用于含组蛋白变体h3.1、h3.2或h3t以及掺入5-甲基胞嘧啶残基的dna二者的核小体。与健康男性相比,发现患有前列腺癌的男性具有较低的循环核小体相关的5-甲基胞嘧啶水平,因此该测定法可单独使用或作为诊断组的一部分使用,作为检测前列腺癌的方法。然后重复该elisa,但使用涉及与常见核小体核心表位结合的固定化抗体(第五种elisa设计)。所述第五种测定法显示对前列腺癌更低的鉴别力。所述结果在图6中显示。
我们得出结论,所述含组蛋白变体h3.1、h3.2或h3t的循环无细胞核小体亦可含有其他表观遗传信号。我们亦得出结论,含组蛋白h3变体和其他特定表观遗传信号二者的循环无细胞核小体的水平,在癌症受试者和健康受试者中可以是不同的,且这可与在癌细胞中观察的水平一致地发生。我们进一步得出结论,即便所述特定表观遗传信号的水平在癌细胞的染色质中可能升高或受到阻抑,亦仍是如此。捕获完全相同的循环核小体流分且仅在标记抗体靶定的表观遗传信号转导结构上不同的其他相同的elisa法,可产生与在癌细胞的染色质中发现的相同表观遗传信号的表达相一致的升高或下降(对于相同的含组蛋白h3变体的循环核小体流分而言)的结果的事实,表明此核小体流分的肿瘤来源。
我们进一步得出结论,含组蛋白h3变体的循环核小体亦具有与其他循环核小体不类似的细胞释放特征。所述核小体具有不同的来源且更少因创伤诱导的细胞死亡所致。因为含h3.1、h3.2或h3t的核小体不会很大程度地由创伤诱导的细胞死亡产生且可用作癌症的生物标志物,所以清楚的是,其为癌症所特有且主要为肿瘤来源。含组蛋白h3.1、h3.2或h3t的核小体的分开或分离,产生从诸如血液、血清或血浆等体液样品中分离或富集含ctdna的肿瘤来源的循环核小体。
本领域技术人员应清楚的是,作为存在核小体的比例的含组蛋白h3变体的核小体的水平,可用作样品中包含ctdna的cfdna的比例的测量结果。所述测量结果类似于ctdna中癌症相关突变的等位基因频率测量结果,且可用作肿瘤负荷测量结果和对疗法的响应。
癌症特有的其他表观遗传标志可类似地用于本发明的方法,只要与其他循环核小体或cfdna相比,其在肿瘤来源的循环核小体或ctdna中更频繁地出现便可。虽然所述表观遗传信号对于肿瘤来源的核小体或dna而言并不是独特的,但增加的频率应足以富集肿瘤来源的核小体和dna。所述表观遗传学标志可包括组蛋白变体(或同种型)、组蛋白修饰、dna修饰和核小体加合物。
在本发明的第一个实施方案中,从生物流体的样品中分离完全或部分纯化的肿瘤核小体制品。所述纯化方法包括使用与所述表观遗传表位结合的结合剂,对含癌症特有的组蛋白或dna表观遗传信号表位的无细胞核小体进行亲和分离。然后可对所述肿瘤核小体制备物和/或其相关ctdna进行分析。在一个优选的实施方案中,通过使用针对组蛋白h3.1或h3.2或h3t的结合剂的免疫亲和纯化法进行肿瘤核小体和ctdna分离。本领域技术人员应清楚的是,能够与组蛋白h3变体(或癌症特有的其他适当的核小体表位)特异性结合的任何结合剂,均可用于本发明的亲和纯化法。所述结合剂可包括但不限于抗体、适体或结合蛋白(例如核小体结合蛋白)。
抗体可通过本领域已知的多种方法产生,包括免疫法和文库法,例如噬菌体展示。可针对关注的部分或抗原诱导免疫反应,或针对与关注部分或抗原的结合来选择文库。涉及与组蛋白h3变体结合的抗体可针对多种所述部分来产生,包括完整的组蛋白h3同种型蛋白氨基酸序列,并且可任选包含翻译后组蛋白修饰。所述蛋白质可从活细胞中纯化,或者以合成法产生。或者可使用代表h3同种型氨基酸序列的一部分的肽序列,且其亦可任选包含翻译后组蛋白修饰。亦可使用含组蛋白h3同种型的核小体或其他染色质流分。
在本研究中,使用了与肽序列atkaarksapatggvkkph结合的抗体。所述氨基酸序列存在于组蛋白变体h3.1、h3.2和h3t的序列中,但不存在于其他组蛋白h3变体中。本领域技术人员应清楚的是,在本发明的方法中可使用涉及与组蛋白变体h3.1和/或h3.2和/或h3t和/或h3.2和/或h3t的任何或全部结合的结合剂。虽然我们在本文中主要提及组蛋白h3.1,但意图此符号(notation)始终包括组蛋白h3.1和/或h3.2和/或h3t。
根据本发明的又一方面,提供通过亲和纯化从生物样品中分离肿瘤来源的循环无细胞核小体的方法,其中所述方法包括步骤:
(i)使所述样品与组蛋白h3.1和/或h3.2和/或h3t结合剂接触;
(ii)从所述样品中分离结合的核小体;和
(iii)分析所述分离的核小体和/或相关dna。
所述分离的肿瘤来源的核小体的分析可包括任何合适的分析方法,其中许多为本领域已知的。这些方法包括但不限于通过使用针对常见核小体表位(例如dsdna)或针对关注的表观遗传结构(包括组蛋白修饰、组蛋白变体、dna修饰或与核小体加合的另一个分子)的第二抗体或其他结合剂的elisa的分析。这些方法通过引用包含在本文中,所有方法描述于wo2005/019826、wo2013/030577、wo2013/030579和wo2013/084002中,其中使用组蛋白h3变体结合剂替换常见的抗核小体表位结合剂,由此对肿瘤来源的循环核小体(而不是自身核小体)分析表观遗传组成。这些方法亦包括用于分析存在于肿瘤来源的循环核小体中的多种表位的多种方法。
根据该方面的一个实施方案,提供用于从任何特定的核小体相关的修饰组蛋白、组蛋白变体、修饰核苷酸方面或者从存在与核小体加合的任何其他分子方面来分析肿瘤来源循环核小体的特定表观遗传表位的免疫测定法,其包括步骤:
(i)使所述样品与组蛋白h3.1和/或h3.2和/或h3t结合剂接触;
(ii)使所述核小体或样品与第二结合剂接触,所述第二结合剂与所述表位结合;
(iii)检测和/或定量所述第二结合剂与所述表位的结合;和
(iv)使用所述结合的存在或程度作为所述样品中肿瘤来源核小体的特定表位的存在的测量结果。
根据一个备选的实施方案,提供用于从任何特定的核小体相关的修饰组蛋白、组蛋白变体、修饰核苷酸方面或者从存在与核小体加合的任何其他分子方面检测和测量含特定表观遗传表位的无细胞核小体或肿瘤来源循环核小体的组成的方法,其包括步骤:
(i)使所述样品与第一结合剂接触,所述第一结合剂与所述表位结合;
(ii)使所述核小体或样品与组蛋白h3.1和/或h3.2和/或h3t结合剂接触;
(iii)检测和/或定量所述组蛋白h3.1和/或h3.2和/或h3t结合剂与所述样品中的核小体的结合;和
(iv)使用所述结合的存在或程度作为所述样品中肿瘤来源核小体的特定表位的存在的测量结果。
通过本发明的方法分离的肿瘤来源的核小体的分析亦可包括本领域已知的任何蛋白组学方法,包括但不限于电泳法、色谱法和涉及质谱法的任何方法,其包括涉及色谱和质谱法和/或稳定同位素标记质谱法的方法和/或涉及蛋白质消化以产生用于通过质谱法或与任何其他方法任意组合的质谱法鉴定和/或定量的肽的方法。在本发明的一个优选的实施方案中,通过在获自癌症患者的血液、血清或血浆样品中进行循环核小体的亲和纯化来制备富集了肿瘤来源的核小体的循环核小体制品,且通过涉及质谱法的方法研究所述核小体制备物的表观遗传组成。在一个实施方案中,所述方法包括步骤:
(i)使所述样品与组蛋白变体h3.1和/或h3.2和/或h3t结合剂接触;
(ii)从所述样品中分离结合的核小体;和
(iii)使用包括质谱法的方法分析在步骤(ii)中分离的核小体。
在一个实施方案中,通过免疫亲和纯化法进行含ctdna的核小体的分离,所述方法包括步骤:
(i)使所述样品与涉及与表观遗传表位结合的结合剂接触,与非肿瘤来源的核小体相比,所述表观遗传表位在肿瘤来源的核小体中更常存在;
(ii)从所述样品中分离结合的核小体;和
(iii)任选从在步骤(ii)中分离的核小体中提取dna。
在又一个实施方案中,涉及与肿瘤来源的核小体特有的表观遗传表位结合的结合剂,在免疫亲和纯化法中涉及与组蛋白变体h3.1和/或h3.2和/或h3t结合,所述方法包括步骤:
(i)使所述样品与组蛋白变体h3.1和/或h3.2和/或h3t结合剂接触;
(ii)从所述样品中分离结合的核小体;和
(iii)任选从在步骤(ii)中分离的核小体中提取dna。
纯化或分离的ctdna的研究可包括分析任何或所有类型的癌症相关的dna异常,包括但不限于表观遗传学分析,其包括dna序列的甲基化、点突变、易位、基因拷贝数、微卫星异常和dna链完整性。此外,可使用任何dna分析法,包括但不限于dna测序、甲基化dna测序分析、pcr、beaming、ngs(靶定基因组或全基因组)、数字pcr、冷pcr(较低变性温度下的共扩增-pcr)、map(midi-激活的焦磷酸解)、pare(重排末端的个性化分析)和质谱法。
在一个实施方案中,所述生物样品包括血液、血清或血浆样品。
根据本发明的又一个方面,提供诊断癌症的方法,其包括在获自人类或动物受试者的生物样品中检测循环无细胞核小体相关的组蛋白变体h3.1和/或h3.2和/或h3t的步骤。
在一个实施方案中,所述诊断方法另外包括检测一种或多种组蛋白修饰、修饰核苷酸、组蛋白变体或同种型或核小体加合物。
在又一个实施方案中,所述组蛋白修饰包括h3k27ac和/或5-甲基胞嘧啶。
根据本发明的又一方面,提供包含组蛋白h3.1和/或h3.2和/或h3t结合剂的试剂盒在本文所述的任何方法中的用途。
现在将参考下列非限制性实施例来阐述本发明。
实施例1
涉及与组蛋白同种型h3.1、h3.2或h3t特异性结合的抗体,通过制造商推荐的方法进行生物素化并固定在链霉抗生物素包被的磁珠(dynal)上。使用磁性分离系统,用上样缓冲液将所述珠粒清洗数次。用上样缓冲液将获自癌症患者的血清或血浆稀释,并加到所述珠粒上。使含组蛋白h3变体的核小体吸附到所述珠粒上。其他血清/血浆组分留在溶液中并借助磁性分离来去除。用缓冲液清洗珠粒。此时所述含组蛋白h3变体的核小体被分离到珠粒上。通过酚/氯仿法或其他标准提取法提取与所述分离的核小体相关的ctdna。可对所述提取的dna分析癌症的遗传特征或表观遗传特征。
实施例2
涉及与组蛋白同种型h3.1、h3.2或h3t特异性结合的抗体,通过制造商推荐的方法进行生物素化并固定在链霉抗生物素包被的磁珠(dynal)上。使用磁性分离系统,用上样缓冲液将所述珠粒清洗数次。用上样缓冲液将获自癌症患者的血清或血浆稀释,并加到所述珠粒上。使含组蛋白h3变体的核小体吸附到所述珠粒上。其他血清/血浆组分留在溶液中并通过磁性分离来去除。用缓冲液清洗珠粒。此时所述含组蛋白h3变体的核小体被分离到珠粒上。使用洗脱缓冲液从所述磁珠去除分离的核小体,并通过包括质谱法在内的蛋白组学法对核小体进行分析。
实施例3
对获自术前以及术后6、24、48、72和96小时时6个患有crc的患者的血清样品,使用roche生产的商品化(总)核小体elisa测定循环无细胞核小体水平,其使用常见的抗核小体核心表位和标记的抗dsdna抗体。然后对所述样品进行再次测定,但使用抗组蛋白变体h3.1、h3.2或h3t捕获抗体。使用商品化elisa,测得的(总)核小体水平在手术创伤之后增加,但对于使用采用抗组蛋白h3.1、h3.2或h3t抗体的测定法测量的含组蛋白h3变体的核小体,对手术的响应得到改变和消减。结果在图3中显示。
实施例4
使用采用了抗组蛋白变体h3.1、h3.2或h3t捕获抗体和标记的抗组蛋白修饰h3k27ac抗体或标记的抗5-甲基胞嘧啶抗体的elisa,测定了获自患有crc的患者和健康对照受试者的血清样品。当使用标记的抗组蛋白修饰h3k27ac抗体时,结果显示,在癌症患者的循环中存在比健康对照的循环中更高水平的核小体相关h3k27ac。相反地,当使用标记的抗5-甲基胞嘧啶抗体时,结果显示,在癌症患者的循环中存在比健康对照的循环中更低水平的核小体相关5-甲基胞嘧啶(图5)。这两个研究结果均与对癌细胞和组织的染色质报道的表观遗传改变相一致(减少的总体dna甲基化和增加的总体h3k27乙酰化)。此外,当结果为表达为比率的两个表观遗传修饰的核小体结果时,所述组合的数据以高准确度区分健康受试者与crc受试者,如在图5中所示。这些结果表明,这些elisa法可用于检测癌症,以及与所述固相抗组蛋白h3变体抗体结合的核小体主要为肿瘤来源的。
实施例5
使用采用固定化抗h3.1、h3.2或h3t抗体作为捕获抗体和用于含有掺入5-甲基胞嘧啶残基的dna的核小体的标记抗5-甲基胞嘧啶抗体的elisa法,测定获自新诊断为前列腺癌的9个男性和类似年龄的10个健康男性的血清样品。发现患有前列腺癌的男性比健康男性具有更低的循环核小体相关5-甲基胞嘧啶水平,且因此该测定法可单独使用或作为诊断组的一部分使用,作为检测前列腺癌的方法。然后重复所述elisa,但使用涉及与常见核小体核心表位结合的固定化抗体。该测定法显示对前列腺癌更低的鉴别力。结果在图6中显示。
参考文献
crowley等,naturereviewsclinicaloncology(自然综述临床肿瘤学),10,472-484,2013。
fong等,clinicalchemistry(临床化学)55(3),587-589,2009。
grutzmann等,plosone3(11):e3759.doi:10.1371/journal.pone.0003759,2008。
guerrero-preston等,epigenetics(实验胚胎学)2(4),223-226,2007。
holdenrieder等,intjcancer(癌症)95,114-120,2001。
holdenrieder和stieber,criticalreviewsinclinicallaboratorysciences(临床实验室科学中的重要综述);46(1):1–24,2009。
jung等,clinicachimicaacta(临床化学学报),411,1611–1624,2010。
karczarski等,clinicalproteomics(临床蛋白组学),11:24,2014。
newman等,naturemedicine(自然医学)20(5),548-554,2014。
schwarzenbach等,naturereviewscancer(自然综述癌症),11(6),426-437,2011。
soares等,cancer(癌症)85(1),112-118,1999。
zhou等,seminarsinoncology(肿瘤学研讨会),39(4),440-448,2012。
- 还没有人留言评论。精彩留言会获得点赞!
- 一种检测哺乳动物血液中微量循环肿瘤细胞的特异性方法与制造工艺
- 一种循环肿瘤细胞阴性富集方法与制造工艺
- 高通量测序检测人循环肿瘤DNA EGFR基因的捕获探针及试剂盒的制造方法与工艺
- 用于肿瘤早期筛查的肿瘤血小板RNA定量检测模型及方法与制造工艺
- 肿瘤循环DNA技术检测‑癌症早期易发风险评估数据法的制造方法与工艺
- 一种偶联抗体的仿生免疫磁球及其制备方法与制造工艺
- 一种循环肿瘤细胞快速检测试剂盒及其制备和应用方法与制造工艺
- 循环肿瘤细胞表面标志分子磷脂酰肌醇蛋白聚糖‑3的检测方法与制造工艺
- 使用组合的核酸酶、连接酶、聚合酶和测序反应识别和枚举核酸序列、表达、拷贝或DNA甲基化变化的方法与制造工艺
- 一种循环肿瘤细胞分析仪的制造方法与工艺