增强抗肿瘤免疫反应的新型免疫策略和免疫组合物的制作方法
本发明属于免疫学领域;更具体地,涉及增强抗肿瘤免疫反应的新型免疫策略和免疫组合物。
背景技术:
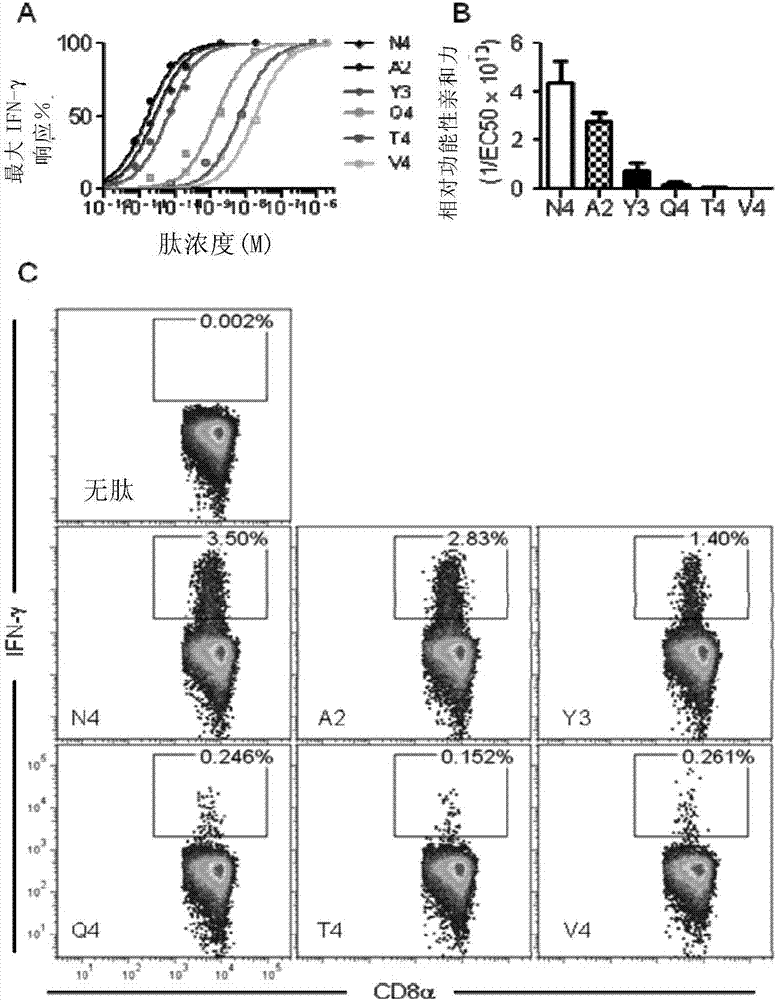
:肿瘤是当前严重威胁人类健康和导致人类死亡的主要疾病。研发有效的癌症治疗药物是势在必行。治疗性肿瘤疫苗可以通过激活人体免疫系统对肿瘤抗原产生免疫应答,特异性清除固体肿瘤组织并形成免疫记忆对患者提供终身保护。和传统的癌症治疗方式(手术切除、放射治疗和化学治疗)以及其他免疫治疗方式相比,治疗性肿瘤疫苗作为一种主动的治疗方法,更易在患者身上使用,且不会引起强烈的副作用。尽管治疗性肿瘤疫苗的研究已经取得了一定进步,然而总体看来,得到综合临床实验结果数据仍然并不理想。低临床疗效是疫苗研发中面临的主要问题。在2003年以前,至少有200种疫苗进行了二期或三期临床实验,而其中的大部分疫苗在临床实验中被证明不能有效的激活患者体内的肿瘤抗原特异性t细胞产生免疫反应抑制肿瘤生长,仅有2.6%的患者在疫苗作用下产生了抗肿瘤免疫肿瘤抗原作为自身抗原一直存在着免疫原性差的问题,影响了治疗性肿瘤疫苗在临床上的应用和发展。影响治疗性肿瘤疫苗有效性的一个重要原因在于中枢耐受机制对抗肿瘤免疫反应的抑制作用。中枢耐受机制是指高亲和力的t细胞在胸腺发育过程中因接触其特异性识别的自身抗原而被阴性选择机制清除而导致的免疫耐受。虽然现有技术中开发了多种技术,如使用佐剂、使用突变的肿瘤抗原等,以期望增强抗肿瘤免疫反应水平,但是在临床实验中的治疗效果并不明显。综上所述,本领域尚缺乏令人满意的、具有高效的抗肿瘤效果的肿瘤疫苗。因此,本领域迫切需要开发新的能够有效增强抗肿瘤免疫反应的方法和组合物,以便获得更好的抗肿瘤预防或治疗效果。技术实现要素:本发明的目的就是提供一种增强抗肿瘤免疫反应的方法和组合物及其应用。在本发明的第一方面,提供了一种非治疗性或者治疗性的增强抗肿瘤免疫反应 的方法,包括步骤:(a)提供第一肿瘤抗原和第二肿瘤抗原,其中所述的增强肿瘤免疫反应是增强的、针对第一肿瘤抗原的肿瘤免疫应答;所述的第一肿瘤抗原为来源于一哺乳动物的低免疫原性的天然多肽或其免疫原性片段,并且当所述第一肿瘤抗原被免疫施用于所述的哺乳动物时,无法有效产生有效的针对所述第一肿瘤抗原的抗肿瘤免疫应答;以及所述的第二肿瘤抗原是与所述第一肿瘤免疫抗原同源且交叉的多肽或其免疫原性片段;并且所述第一肿瘤抗原与来自所述哺乳动物的抗原特异性cd8+t细胞的亲和力为q1,所述第二肿瘤抗原与来自所述哺乳动物的抗原特异性cd8+t细胞的亲和力为q2,并且所述q2/q1的比值r1为0.02-0.80;(b)将所述的第二肿瘤抗原免疫施用于所述的哺乳动物的对象,激发针对所述第二肿瘤抗原的免疫应答,从而获得经初次免疫激发的哺乳动物对象;(c)在上一步骤的免疫施用之后t天后,其中t为3-60,将所述的第一肿瘤抗原免疫施用于所述的经初次免疫激发的哺乳动物对象,激发针对所述第一肿瘤抗原的免疫应答,从而获得经再次免疫激发的哺乳动物对象,其中在所述再次免疫激发的哺乳动物对象中,产生了增强的针对第一肿瘤抗原的肿瘤免疫应答。在另一优选例中,所述的“增强的针对第一肿瘤抗原的肿瘤免疫应答”指与用所述第一肿瘤抗原进行初次免疫施用并用所述第一肿瘤抗原再次免疫施用的对照中的针对第一肿瘤抗原的肿瘤免疫应答水平yc相比,步骤(c)中用所述第二肿瘤抗原进行初次免疫施用并用所述第一肿瘤抗原再次免疫施用的哺乳动物对象中的针对第一肿瘤抗原的肿瘤免疫应答水平yv显著高于yc。在另一优选例中,所述的“yv显著高于yc”指yv/yc的比值r2≥1.5,较佳地≥2,更佳地≥5,更佳地≥10,最佳地≥20。在另一优选例中,所述的肿瘤免疫应答水平为免疫的哺乳动物t细胞中抗原特异性cd8+t细胞的数量和/或比例。更佳地,所述的肿瘤免疫应答水平包括:用第一种抗原体外刺激哺乳动物t细胞后检测细胞因子(如伽玛干扰素)的分泌水平,或者用主要组织相容性复合体(mhc)-抗原表位多肽的四聚体或者多聚体经流式细胞仪分析抗原特异性cd8+t细胞的数量和比例。在另一优选例中,所述的t为4-45,较佳地t为5-30,更佳地t为6-25,最佳地t为7-21。在另一优选例中,所述的q2/q1的比值r1为0.04-0.60,较佳地为0.05-0.50,更佳地为0.06-0.40。在另一优选例中,第一肿瘤抗原与t细胞的亲和力q1在9-100纳摩尔(um)之 间,和/或第二肿瘤抗原与t细胞的亲和力q2在0.01-10纳摩尔之间。在另一优选例中,所述的亲和力的定义如下:mhc-抗原表位多肽与t细胞受体之间结合的强度。在另一优选例中,所述的亲和力为功能性亲和力或ic50的倒数。在另一优选例中,所述的功能性亲和力指,抗原表位多肽体外刺激t细胞分泌细胞因子(如伽玛干扰素)达到最高水平的50%所需要的多肽浓度。在另一优选例中,所述的“无法有效产生有效的抗肿瘤免疫应答”指在所述的被免疫施用于所述哺乳动物时,产生针对所述第一肿瘤抗原的抗肿瘤免疫应答率≤25%,较佳地≤15%,更佳地≤10%,更佳地≤5%,最佳地≤3%。在另一优选例中,所述的第一肿瘤抗原是来自所述哺乳动物的野生型肿瘤相关抗原(蛋白)的表位多肽(即多肽片段)。在另一优选例中,所述的“同源的且交叉的”指所述的第二肿瘤抗原和所述的第一肿瘤抗原在每10个氨基酸长度上具有平均1-2个氨基酸突变,因此不仅能够引起针对第二肿瘤抗原的免疫应答,而且能够引起针对所述第一肿瘤抗原的交叉的免疫应答反应。在另一优选例中,所述的第一肿瘤抗原和第二肿瘤抗原来自或衍生自同一致肿瘤相关抗原。在另一优选例中,所述的第一肿瘤抗原和第二肿瘤抗原来自或衍生自同一致肿瘤相关抗原的同一区段。在另一优选例中,所述的第一肿瘤抗原和第二肿瘤抗原为长度分别为n1和n2个氨基酸的表位多肽,其中n1和n2分别为6-200,较佳地7-100,更佳地8-50,最佳地8-25或8-12的正整数。在另一优选例中,所述的哺乳动物包括非人哺乳动物和人。在另一优选例中,所述的哺乳动物包括灵长动物和啮齿动物(如小鼠、大鼠)。在另一优选例中,所述的第一肿瘤抗原为中枢耐受的肿瘤特异性表位。在另一优选例中,所述的第一肿瘤抗原来源于选自下组的肿瘤相关抗原:ny-eso-1、her2、egfr、cea、gpc3、afp、pap、psa、psma、psca、或其组合。在另一优选例中,所述的“免疫施用”指将所述的肿瘤抗原或被所述肿瘤抗原致敏的树突状细胞(dc)直接或间接地施用于所述对象,从而引发免疫应答反应。在另一优选例中,所述的“免疫施用”包括以疫苗组合物的形式进行施用。在另一优选例中,所述的疫苗组合物含有:(i)所述的肿瘤抗原(如第一肿瘤抗原或第二肿瘤抗原)和/或被所述肿瘤抗原致敏的树突状细胞;(ii)任选的佐剂;和(iii)药学上或免疫学上可接受的载体。在另一优选例中,所述的树突状细胞与所述的第一肿瘤抗原来源于同一种哺乳动物。在本发明的第二方面,提供了一种增强抗肿瘤免疫反应的组合物产品,所述产品包括:(1)第一组合物,所述的第一组合物包括(i)第一肿瘤抗原和/或被所述第一肿瘤抗原致敏的树突状细胞;(ii)任选的佐剂;和(iii)药学上或免疫学上可接受的载体;(ii)第二组合物,所述的第二组合物包括(i)第二肿瘤抗原和/或被所述第二肿瘤抗原致敏的树突状细胞;(ii)任选的佐剂;和(iii)药学上或免疫学上可接受的载体;并且,所述的第一组合物和第二组合物是独立的,其中,所述的第一肿瘤抗原为来源于一哺乳动物的低免疫原性的天然多肽或其免疫原性片段,并且当所述第一肿瘤抗原被免疫施用于所述的哺乳动物时,无法有效产生有效的针对所述第一肿瘤抗原的抗肿瘤免疫应答;以及所述的第二肿瘤抗原是与所述第一肿瘤免疫抗原同源且交叉的多肽或其免疫原性片段;并且所述第一肿瘤抗原与来自所述哺乳动物的抗原特异性cd8+t细胞的亲和力为q1,所述第二肿瘤抗原与来自所述哺乳动物的抗原特异性cd8+t细胞的亲和力为q2,并且所述q2/q1的比值r1为0.02-0.80,并且所述的增强肿瘤免疫反应是增强的、针对第一肿瘤抗原的肿瘤免疫应答。在另一优选例中,所述的组合物产品是用于治疗或预防肿瘤的瘤苗或疫苗组合物。在另一优选例中,所述的疫苗组合物还含有佐剂。在另一优选例中,所述疫苗组合物为核酸疫苗组合物。在另一优选例中,所述的佐剂包括氧化铝、皂苷、胞壁酰二肽、矿物油或植物油、基于囊泡的佐剂、非离子嵌段共聚物或deae葡聚糖、细胞因子(包括il-1、il-2、ifn-r、gm-csf、il-6、il-12、和cpg)。在本发明的第三方面,提供了本发明第二方面所述的组合物产品的用途,它们被用于制备增强抗肿瘤免疫反应的药物。在另一优选例中,所述药物用于治疗选自下组的肿瘤:肝癌、肺癌、胃癌、乳腺癌、卵巢癌、前列腺癌、皮肤癌、黑色素瘤、宫颈癌、脑癌、甲状腺癌和胆管癌、膀胱癌和胰腺癌。在本发明的第四方面,提供了一种(非诊断性和非治疗性的)挑选具有增强针对低免疫原性的肿瘤抗原的抗肿瘤免疫反应的候选多肽免疫原的方法,包括步骤:(a)提供一测试组,所述测试组包括i个待选择的多肽免疫原,其中所述的各多肽免疫原是同源的且交叉的,其中i为≥1的正整数,并且,所述的“同源的且交叉的”指一个多肽免疫原(a抗原或天然抗原)与所述测试组中另一多肽免疫原(b抗原或相近抗原)在每10个氨基酸长度上具有平均1-2个氨基酸突变,因此不仅在哺乳动物中能够引起针对自抗原的免疫应答,而且能够引起针对所述相近抗原的交叉的免疫应答反应;其中,所述的多肽免疫原中至少1个为突变型多肽免疫原;(b)测定所述各多肽免疫原与各自所引发的抗原特异性cd8+t细胞的反应亲和力,分别记为qj,其中j=1-i中任一正整数;(c)将所述各qj进行排序,选择排序位于中间的多肽免疫原,作为具有增强针对低免疫原性的肿瘤抗原的抗肿瘤免疫反应的候选多肽免疫原;和/或将各qj与qz进行比较,选择所述qj/qz的比值r为0.02-0.80的多肽免疫原,作为具有增强针对低免疫原性的肿瘤抗原的抗肿瘤免疫反应的候选多肽免疫原,其中,qz是所述第一肿瘤抗原与来自所述哺乳动物的抗原特异性cd8+t细胞的亲和力,而所述的第一肿瘤抗原为来源于哺乳动物的低免疫原性的天然肿瘤抗原或其免疫原性片段,并且当所述第一肿瘤抗原被免疫施用于所述的哺乳动物时,无法有效产生有效的针对所述第一肿瘤抗原的抗肿瘤免疫应答。在另一优选例中,在步骤(c)中,包括:将所述各qj进行排序,将qj中的最大值定义为qmax,并选择qj/qmax的比值r为0.02-0.80的多肽免疫原,作为具有增强针对低免疫原性的肿瘤抗原的抗肿瘤免疫反应的候选多肽免疫原。在另一优选例中,在步骤(c)中,包括:将所述各qj进行排序,并选择与q的算术平均值qaverage或中位数qmean最接近(即|qaverage-qj|或|qmean-qj|最小)的s个多肽免疫原,且s为<i的正整数,i为≥3正整数,且i-s≥2。在另一优选例中,s/i为1/3至3/5。在另一优选例中,当i≥3时,被挑选出的多肽免疫原不包括qj最小的以及qj最大的多肽免疫原。在另一优选例中,所述的qj/qz或qj/qmax的比值r为0.04-0.60,较佳地为0.05-0.50,更佳地为0.06-0.40。在另一优选例中,所述的方法还包括步骤:(d)对于上一步骤所挑选出的候选多肽免疫原,测试其增强针对低免疫原性的肿瘤抗原的抗肿瘤免疫反应的能力。在另一优选例中,所述的步骤(d)中,包括:用上一步骤挑选出的候选多肽免疫原hx,对所述哺乳动物进行初次免疫施用,从而激发针对所述候选多肽免疫原hx的免疫应答,从而获得经初次免疫激发的哺乳动物对象;以及在所述的初次免疫施用操作之后t天后,其中t为3-60,将所述的第一肿瘤抗原免疫施用于所述的经初次免疫激发的哺乳动物对象,激发针对所述第一肿 瘤抗原的免疫应答,并测定经再次免疫激发的哺乳动物对象中针对第一肿瘤抗原的肿瘤免疫应答水平,从而判断所述候选多肽免疫原hx是否具有增强针对低免疫原性的肿瘤抗原的抗肿瘤免疫反应的能力和/或增强的程度。在本发明的第五方面,提供了一种第二肿瘤抗原的用途,所述第二肿瘤抗原是与第一肿瘤免疫抗原同源且交叉的多肽或其免疫原性片段,其中所述的第二肿瘤抗原被用于制备一药物组合物,所述药物组合物(a)用于增强哺乳动物产生针对所述第一肿瘤免疫抗原的抗肿瘤免疫反应;和/或(b)用于突破哺乳动物机体对自身抗原的免疫耐受并增强抗肿瘤免疫反应。在另一优选例中,所述的“同源的且交叉的”指第一肿瘤抗原(a抗原)与第二肿瘤抗原(b抗原)在匹配区,在每10个氨基酸长度上具有平均1-2个氨基酸突变。在另一优选例中,所述第二肿瘤抗原是长度为n2个氨基酸的表位多肽,其中n2为6-200,较佳地7-100,更佳地8-50,最佳地8-25或8-12的正整数。在本发明的第六方面,提供了一种增强抗肿瘤免疫反应的治疗方法,包括步骤:(a)提供一来自来自患者的分离的淋巴细胞,所述淋巴细胞选自下组:肿瘤侵润的淋巴细胞(til)、外周血淋巴细胞(pbmc)、或其组合;以及提供提供第一肿瘤抗原和第二肿瘤抗原;其中所述的第一肿瘤抗原为来源于一哺乳动物的低免疫原性的天然多肽或其免疫原性片段,并且当所述第一肿瘤抗原被免疫施用于所述的哺乳动物时,无法有效产生有效的针对所述第一肿瘤抗原的抗肿瘤免疫应答;以及所述的第二肿瘤抗原是与所述第一肿瘤免疫抗原同源且交叉的多肽或其免疫原性片段;(b)在第二肿瘤抗原存在下,对所述的til淋巴细胞和/或pbmc淋巴细胞进行培养;然后在第一肿瘤抗原存在下,再次进行培养,从而获得经致敏的t细胞;(c)任选地对所述的致敏的淋巴细胞进行传代,从而获得经传代的、经致敏的t细胞;(d)将上一步骤获得的经致敏的t细胞,回输给患者。在另一优选例中,在步骤(b)中,所述的培养还包括在含在树突状细胞、细胞因子或其组合的培养体系中,进行体外t细胞培养。在另一优选例中,在步骤(b)之中和之后、步骤(c)之中和之后、或步骤(d)之前和之中,所述方法还包括:检测所述经致敏的t细胞与第一肿瘤抗原和/或第二肿瘤抗原的亲和力。在另一优选例中,所述第一肿瘤抗原与来自所述哺乳动物的抗原特异性cd8+t细胞的亲和力为q1,所述第二肿瘤抗原与来自所述哺乳动物的抗原特异性cd8+t 细胞的亲和力为q2,并且所述q2/q1的比值r1为0.02-0.80(即第二肿瘤抗原的亲和力低于第一肿瘤抗原的亲和力)。在本发明的第七方面,提供了一种增强抗肿瘤免疫反应的治疗方法,包括步骤:(a)提供第一肿瘤抗原和第二肿瘤抗原,其中所述的增强肿瘤免疫反应是增强的、针对第一肿瘤抗原的肿瘤免疫应答;所述的第一肿瘤抗原为来源于一哺乳动物的低免疫原性的天然多肽或其免疫原性片段,并且当所述第一肿瘤抗原被免疫施用于所述的哺乳动物时,无法有效产生有效的针对所述第一肿瘤抗原的抗肿瘤免疫应答;以及所述的第二肿瘤抗原是与所述第一肿瘤免疫抗原同源且交叉的多肽或其免疫原性片段;并且所述第一肿瘤抗原与来自所述哺乳动物的抗原特异性cd8+t细胞的亲和力为q1,所述第二肿瘤抗原与来自所述哺乳动物的抗原特异性cd8+t细胞的亲和力为q2,并且所述q2/q1的比值r1为0.02-0.80;(b)将所述的第二肿瘤抗原免疫施用于所述的哺乳动物的对象,激发针对所述第二肿瘤抗原的免疫应答,从而获得经初次免疫激发的哺乳动物对象;(c)在上一步骤的免疫施用操作之后t天后,其中t为3-60,将所述的第一肿瘤抗原免疫施用于所述的经初次免疫激发的哺乳动物对象,激发针对所述第一肿瘤抗原的免疫应答,从而获得经再次免疫激发的哺乳动物对象,其中在所述再次免疫激发的哺乳动物对象中,产生了增强的针对第一肿瘤抗原的肿瘤免疫应答。应理解,在本发明范围内中,本发明的上述各技术特征和在下文(如实施例)中具体描述的各技术特征之间都可以互相组合,从而构成新的或优选的技术方案。限于篇幅,在此不再一一累述。附图说明图1显示了ova天然抗原表位n4及突变抗原的结构。图2显示了天然抗原n4及突变抗原对内源性多克隆t细胞的功能性亲和力。其中,(a)wt小鼠尾静脉注射感染lm-n45000cfu/只后第7天,取脾脏细胞体外用梯度浓度的天然抗原和突变抗原刺激(10-12m-10-6m),检测ifn-γ+cd8+t细胞比例。根据其数值与对应的刺激浓度拟合剂量-效应曲线(n≥4);(b)计算每组剂量-效应曲线对应的ec50,取倒数(1/ec50)代表相对功能性亲和力(relativeavidity);(c)wt小鼠尾静脉注射感染lm-n45000cfu/只后第7天,取脾脏细胞体外用天然抗原或突变抗原2μg/ml刺激培养,检测ifn-γ+cd8+t细胞比例。图3显示了中枢耐受机制使t细胞库更倾向于对中等亲和力突变抗原反应。其 中,(a)天然抗原和突变抗原感染后识别抗原本身的ifn-γ+cd8+t细胞比例的统计平均值(n≥4)。(b)天然抗原和突变抗原感染后识别n4的ifn-γ+cd8+t细胞比例的统计平均值(n≥4)。(c)wt小鼠和rip-mova小鼠用lm-n4或突变株尾静脉注射感染后,其对天然抗原n4反应的ifn-γ+cd8+t细胞比例与对相应抗原反应的ifn-γ+cd8+t细胞比例间的比值(n≥4)。图4显示了基于ova抗原耐受的rip-mova小鼠黑色素瘤模型的构建。其中,(a)如图示,2×104,10×104,50×104细胞数的mo5分别皮下接种wt小鼠,每3天测量肿瘤大小并记录。当肿瘤长径和短径长度的平均值超过20mm时,处死小鼠,视为小鼠死亡。(b)50×104细胞数的mo5皮下接种rip-mova小鼠,每3天测量肿瘤大小并记录。当肿瘤长径和短径长度的平均值超过20mm时,处死小鼠,视为小鼠死亡。(c)50×104细胞数接种后,wt荷瘤小鼠和rip-mova荷瘤小鼠的存活曲线(n≥7)。(d)50×104细胞数接种后20天,rip-mova荷瘤小鼠脾脏和腹股沟引流淋巴结中n4-tetramer+cd44+的cd8+t细胞比例。(e)50×104细胞数接种后20天,rip-mova荷瘤小鼠脾脏和腹股沟引流淋巴结细胞体外用天然抗原n4或突变抗原y3刺激,检测其cd8+t细胞中细胞因子ifn-γ和tnf-α的表达。图5:中枢耐受机制使得低亲和力的自身反应性t细胞不能诱导有效的抗肿瘤免疫反应。其中,(a)wt小鼠尾静脉感染lm-n4或lm-y3后7天,皮下接种5×105/只mo5黑色素瘤细胞。mo5接种后第6天开始,每隔2天测量肿瘤尺寸,计算肿瘤体积。(b)rip-mova小鼠尾静脉感染lm-n4或lm-y3后7天,皮下接种5×105/只mo5黑色素瘤细胞。mo5接种后第6天开始,每隔2天测量肿瘤尺寸,计算肿瘤体积。(c)wt小鼠和rip-mova小鼠尾静脉感染lm-n4或lm-y3后7天,取脾脏细胞体外用梯度浓度的n4刺激(10-12m-10-6m),检测ifn-γ+cd8+t细胞比例。根据其数值与对应的刺激浓度拟合剂量-效应曲线。wtsp.t细胞:lm-n4感染wt小鼠;wtcr.t细胞:lm-y3感染wt小鼠;rocr.t细胞,lm-y3感染rip-mova小鼠。(d)计算每组剂量-效应曲线对应的ec50,取倒数(1/ec50)代表相对功能性亲和力。图6显示了中枢耐受下的自身抗原反应性t细胞库vβ使用多样性。其中,(a)wt小鼠和rip-mova小鼠尾静脉注射感染lm-y3后第7天,n4-tetramer抗体和y3-tetramer抗体双染色,检测识别不同tetramer的cd44+cd8+t细胞比例。(b)wt小鼠和rip-mova小鼠尾静脉感染lm-n4或lm-y3后7天,取脾脏细胞体外用n4刺激培养,检测ifn-γ+cd8+t细胞使用不同vβ的比例。wtsp.t细胞:lm-n4感染wt小鼠;wtcr.t细胞:lm-y3感染wt小鼠;rocr.t细胞,lm-y3感染rip-mova小鼠。(c)rip-mova小鼠尾静脉感染lm-y3后7天,取脾脏细胞体外用y3刺激培养,检测ifn-γ+cd8+t细胞使用不同vβ的比例示意图。(d)rip-mova小鼠尾静脉感染lm-y3后7天,取脾脏细胞体外用y3刺激培养,检测ifn-γ+cd8+t细胞使用不同vβ的比例(n≥8)。(e)wtsp.t细胞和rocr.t细胞使用不同vβ比例的比较。(f)wt cr.t细胞和rocr.t细胞使用不同vβ比例的比较。图7显示了同种抗原疫苗的重复免疫不能诱导有效的抗肿瘤免疫反应。其中,(a)wt小鼠接种黑色素瘤细胞mo5后第7天开始,用表达对应抗原的lm菌株腹腔注射进行治疗,剂量为104cfu/只。治疗共重复3次,每次间隔一周。mo5接种6第6天开始,每隔2天测量肿瘤大小并计算体积。(b)rip-ova小鼠接种黑色素瘤细胞mo5后第7天开始,用表达对应抗原的lm菌株腹腔注射进行治疗。治疗共重复3次,每次间隔一周。(c)wt小鼠接种黑色素瘤细胞mo5后第7天开始用lm-n4或lm-y3治疗,治疗共重复3次,每次间隔一周。第30天的时候处死小鼠,取脾脏、腹股沟引流淋巴结、肠系膜淋巴结细胞,体外用天然抗原n4刺激,检测ifn-γ+cd8+t细胞比例。(d)rip-ova小鼠接种黑色素瘤细胞mo5后第7天开始,用cpg活化的dc负载突变抗原y3,足垫免疫进行治疗,剂量为106细胞/只。治疗共重复3次,每次间隔一周。(e)rip-ova小鼠接种黑色素瘤细胞mo5后第7天开始,用cpg活化的dc负载突变抗原y3,足垫免疫进行第一次治疗,剂量为106细胞/只;一周后,用lm-y3腹腔注射进行第二次治疗,剂量为104cfu/只。图8显示了优化的双重异源免疫策略增强自身反应性t细胞的功能性和亲和力。其中,(a)rip-ova小鼠用lm-y3或dc-y3进行第一次免疫;根据实验描述一周后用表达同种突变抗原y3再次免疫。4天后取小鼠脾脏体外用n4刺激,检测ifn-γ+cd8+t细胞、tnf-α+cd8+t细胞和ifn-γ+tnf-α+cd8+t细胞的比例(n≥4)。(b)根据实验描述,rip-ova小鼠采用双重异源免疫策略,间隔一周,分两次免疫。第2次免疫4天后取脾脏体外用n4刺激,检测ifn-γ+cd8+t细胞、tnf-α+cd8+t细胞和ifn-γ+tnf-α+cd8+t细胞的比例。(c)rip-ova小鼠如图示描述分别用lm-y3免疫一次,lm-y3免疫两次,dc-y3-lm-y3免疫,dc-y3-lm-n4免疫后,脾脏细胞体外用梯度浓度n4刺激,检测对应刺激浓度的ifn-γ+cd8+t细胞比例,拟合剂量效应曲线。(d)计算图6c中每组剂量-效应曲线对应的ec50,取倒数(1/ec50)代表相对功能性亲和力。(e)rip-ova小鼠用dc-疫苗起始免疫后,用改变抗原的lm-疫苗免疫,检测cd8+t细胞表达细胞因子ifn-γ和tnf-α的比例。(f)rip-ova小鼠用dc-y3-lm-n4免疫后,脾脏细胞体外用梯度浓度n4刺激,检测ifn-γ+cd8+t细胞中各vβ的使用比例。(g)rip-ova小鼠用lm-y3免疫一次后产生的rocr.t细胞中倾向使用的vβ比例(图4b)和用dc-y3-lm-n4免疫后识别n4的ifn-γ+cd8+t细胞中的比较。图9显示了双重异源免疫策略激活的自身反应性t细胞具有抗肿瘤作用。其中,(a)rip-ova小鼠上第-14天用lm-y35000cfu/只或dc-y3106细胞/只预免,7后用lm-y35000cfu/只或lm-n45000cfu/小鼠再次免疫增强反应。第0天时接种黑色素瘤细胞mo55×105细胞/只。自mo5接种后第6天开始,每隔2天测量肿瘤大小并计算体积。(n=3或4)。(b)rip-ova小鼠各免疫组接种mo5后,在各时间点 每组小鼠的平均肿瘤体积大小。(c)rip-ova小鼠dc-y3-lm-n4免疫组,lm-y3-lm-y3和未免疫组小鼠接种mo5后的存活率。图10显示了双重异源免疫能作为治疗性疫苗在荷瘤小鼠上抑制肿瘤生长。其中,(a)rip-ova小鼠上接种黑色素瘤mo55×105/只,第7天根据描述给予第一次免疫治疗,第14天第二次免疫治疗。dc疫苗足垫免疫治疗,剂量1×106/只;lm疫苗腹腔注射免疫,剂量10000cfu/只。(b)根据图10a步骤,双重异源免疫治疗荷瘤rip-ova小鼠。mo5接种第6天开始,每隔2天测量肿瘤大小并计算体积,测定小鼠血糖浓度。(c)荷瘤rip-ova小鼠不治疗组,dc-y3-lm-y3治疗组,dc-y3-lm-n4治疗组在各时间点每组小鼠的平均肿瘤体积大小(n≥5)。(d)荷瘤rip-ova小鼠不治疗组,dc-y3-lm-y3组,dc-y3-lm-n4组的小鼠存活曲线。图11显示了基于佐剂的双重异源免疫策略可提高抗肿瘤特异性t细胞免疫反应,并显著性地抑制肿瘤生长。具体实施方式本发明人经过广泛而深入的研究,通过大量筛选和实验,首次开发了一种增强抗肿瘤免疫反应的方法以及用于该方法的组合物。本发明方法通过双重异源免疫策略,可以在中枢耐受环境下有效激活识别天然肿瘤抗原的t细胞,并有效提高其亲和力及多功能性,从而达到抑制肿瘤生长,延长存活期等效果。在此基础上完成了本发明。具体地,具体地,本发明人利用表达模式抗原ova作为自身抗原的rip-mova小鼠构建了ova(n4)特异性t细胞中枢耐受模型,并以n4及其5种突变抗原的重组李斯特菌株作为模式疫苗感染该转基因小鼠,研究了中枢耐受机制作用下,机体t细胞库对天然(自身)抗原和突变抗原的识别和反应。本发明人发现,中枢耐受不仅完全抑制免疫系统对自身抗原n4的反应,也部分抑制了对突变抗原的反应。而中等亲和力的突变抗原y3能诱导最多的交叉识别天然抗原n4的t细胞,即自身反应性t细胞。该免疫反应机制说明,因肿瘤抗原为自身抗原,在中枢耐受环境下,直接用肿瘤抗原不能诱导有效的抗肿瘤免疫反应。因为胸腺发育过程中选择性的清除了高亲和力的自身反应性t细胞,所以直接识别肿瘤抗原的t细胞库很小,而大部分识别突变抗原的t细胞库受到中枢耐受机制的影响有限,其中依然存在足够的对肿瘤抗原具有低亲和力的自身反应性t细胞,这群t细胞更依赖于某些特定vβ的表达,可能是因为使用该vβ的tcr亲和力低,所以更容易逃离阴性选择。但是这也影响了自身反应性t细胞的功能性,使其不具有有效的抗肿瘤作用。因此,在利用突变抗原诱导交叉识别自身抗原的t细胞库的基础上,选择性增强这群自身反应性t细胞的亲和力和功能性,对抗肿瘤免疫反应来说尤为重要。本发明人在实验中发现,增强自身反应性t细胞的功能性和亲和力受两种不同的机制调控。免疫载体的改变可以影响自身反应性t细胞的亲和力,而抗原的改变能促进自身反应性t细胞的功能性细胞因子表达。以cpg活化成熟后dc作为载体进行第一次免疫,以lm作为载体进行第二次免疫,可以有效的增强自身反应性t细胞的亲和力。而两次都使用dc或者两次都使用lm,或者颠倒dc和lm作为载体的使用顺序则没有同样的效果。通过对自身反应性t细胞库vβ的研究,本发明人发现,中枢耐受机制不仅减少了自身反应性t细胞的绝对数量,在存留下来的低亲和力的识别自身抗原的t细胞库,其多样性也降低了。在本发明人的系统里,低亲和力的自身反应性t细胞中vβ3,vβ7,vβ10b,和vβ13的使用比例高于识别同一抗原的高亲和力的t细胞,而使用其他(未检测的vβ)的比例则明显下降,这大概是因为在胸腺阴性选择的过程中会特异性清除了高亲和力t细胞克隆,进而导致了对某些低亲和力vβ的使用偏好和总体上细胞库多样性的降低。而在优化的双重异源免疫策略dc-y3-lm-n4免疫后,识别自身抗原的t细胞库中仅仅vβ3的比例显著减少,而vβ4的使用比例明显增加,即有效提高了自身反应性t细胞的功能性亲和力。有趣的是,在高亲和力t细胞库中vβ4的使用比例却并不高。本发明人推测经过优化的双重异源免疫后自身反应性t细胞的亲和力和功能性增加,主要是通过特异性的不扩增某些低亲和力和功能性的t细胞克隆,而增加了其他t细胞克隆被选择的机会。本发明人在rip-mova小鼠上接种表达ova的黑色素瘤细胞mo5,模拟人自身肿瘤发生情况,然后利用优化的双重异源免疫策略dc-y3-lm-n4作为疫苗治疗荷瘤小鼠,发现相比对照组,优化后的免疫策略可以抑制肿瘤生长,有效延长小鼠的存活期。说明抗肿瘤反应的效率主要还是依赖于自身反应性t细胞功能性和亲和力的提高。术语如本文所用,术语“本发明肿瘤抗原”指第二肿瘤抗原,即与所述第一肿瘤免疫抗原同源且交叉的多肽或其免疫原性片段。例如,当trp2sd肽(seqidno.:7)为第一肿瘤抗原时,一种第二肿瘤抗原是trp2sk(seqidno.:8)。如本文所用,术语“抗原表位(肽)”指拟诱导动物产生免疫反应的其它蛋白的一段肽。通常,抗原表位指免疫反应拟靶向的肽段,较佳地来源于哺乳动物(如人)蛋白的一段肽。中枢耐受模型在本发明中,针对中枢耐受模型的实验证明,本发明方法可有效激活识别天然低免疫性抗原(包括肿瘤抗原或自身肿瘤抗原)的t细胞,从而有效增强针对低免疫 性性抗原的免疫反应。在本发明中,一种代表性的中枢耐受模型是ova的转基因小鼠(rip-mova),该转基因小鼠是人工构建的针对天然抗原n4特异性t细胞的中枢耐受模型。该小鼠在髓质胸腺上皮细胞、胰腺β细胞和肾脏近端小管中表达ova作为自身抗原,因此在免疫系统发育过程中中枢耐受机制已经选择性清除了小鼠体内n4特异性t细胞。本发明人的实验表明,该中枢耐受模型小鼠体内仍存在有低亲和力的n4自身反应性t细胞。本发明人利用表达天然ova抗原的李斯特菌株(lm-n4)或表达突变ova抗原的李斯特菌株(lm-a2~lm-v4)感染rip-mova和野生型(wt)小鼠,结果发现,中枢耐受机制作用下,小鼠的t细胞库对自身抗原,亦即天然抗原不反应,而更容易被中等亲和力的突变抗原所激活。卵清蛋白天然抗原及其同源且交叉的突变抗原模式抗原卵清蛋白(ova)的天然抗原和系列突变抗原一直应用于t细胞反应的研究。ova257-264位的氨基酸序列siinfekl(n4)能被i型mhc分子kb识别并递呈给t细胞,从而引起强烈的抗原特异性cd8+t细胞反应。在天然抗原n4序列基础上根据氨基酸对应位置进行单个氨基酸突变,获得一系列突变抗原。如图1所述,n4及其代表性的突变抗原包括:天然抗原n4:siinfekl(seqidno.:1)突变抗原a2:sainfekl(seqidno.:2)突变抗原y3:siynfekl(seqidno.:2)突变抗原q4:siiqfekl(seqidno.:4)突变抗原t4:siitfekl(seqidno.:5)突变抗原v4:siivfekl(seqidno.:6)这组突变抗原和天然抗原对小鼠抗原递呈细胞mhc分子kb具有相似的亲和力,而对cd8+t细胞受体的功能性亲和力则不同。n4用于模拟第一(肿瘤)抗原,而y3是优选的同源且交叉的第二(肿瘤)抗原。组合物和施用方法本发明还提供了一种组合物(或组合物产品),它含有:(1)第一组合物,所述的第一组合物包括(i)第一肿瘤抗原和/或被所述第一肿瘤抗原致敏的树突状细胞;(ii)任选的佐剂;和(iii)药学上或免疫学上可接受的载体;(ii)第二组合物,所述的第二组合物包括(i)第二肿瘤抗原和/或被所述第二肿瘤抗原致敏的树突状细胞;(ii)任选的佐剂;和(iii)药学上或免疫学上可接受的载体;并且,所述的第一组合物和第二组合物是独立的。本发明中,术语“含有”表示各种成分可一起应用于或存在于本发明的组合物中。因此,术语“主要由...组成”和“由...组成”包含在术语“含有”中。本发明的组合物包括药物组合物和疫苗组合物。本发明的组合物可以是单价的(仅含有一种重组蛋白或多核苷酸),也可以是多价的(含有多种重组蛋白或多核苷酸)。本发明的药物组合物或疫苗组合物可制备成各种常规剂型,其中包括(但并不限于):注射剂、粒剂、片剂、丸剂、栓剂、胶囊、悬浮液、喷雾剂等。(1)药物组合物本发明的药物组合物包含(或含有)治疗有效量的第一肿瘤抗原和/或被所述第一肿瘤抗原致敏的树突状细胞;以及第二肿瘤抗原和/或被所述第二肿瘤抗原致敏的树突状细胞。本文所用的术语“治疗有效量”指治疗剂治疗、缓解或预防目标疾病或状况的量,或是表现出可检测的治疗或预防效果的量。该效果可通过例如抗原水平来检测。治疗效果也包括生理性症状的减少。对于某一对象的精确有效量取决于该对象的体型和健康状况、病症的性质和程度、以及选择给予的治疗剂和/或治疗剂的组合。因此,预先指定准确的有效量是没用的。然而,对于某给定的状况而言,可以用常规实验来确定该有效量。为了本发明的目的,有效的剂量为给予个体各自约0.001毫克/千克至1000毫克/千克,较佳地约0.01毫克/千克至100毫克/千克体重的所述的第二肿瘤抗原以及第一肿瘤抗原。药物组合物还可含有药学上可接受的载体。术语“药学上可接受的载体”指用于治疗剂(例如本发明的重组蛋白)给药的载体。该术语指这样一些药剂载体:它们本身不诱导产生对接受该组合物的个体有害的抗体,且给药后没有过分的毒性。合适的载体可以是大的、代谢缓慢的大分子,如蛋白质、多糖、聚乳酸(polylacticacid)、聚乙醇酸等。这些载体是本领域普通技术人员所熟知的。在remington'spharmaceuticalsciences(mackpub.co.,n.j.1991)中可找到关于药学上可接受的载体或赋形剂的充分讨论。组合物中药学上可接受的载体可包括液体,如水、盐水、甘油和乙醇。另外,这些载体中还可能存在辅助性的物质,如润湿剂或乳化剂、ph缓冲物质等。通常,可将组合物制成可注射剂,例如液体溶液或悬液;还可制成在注射前适合配入溶液或悬液、液体赋形剂的的固体形式。脂质体也包括在药学上可接受的载体的定义中。(ii)疫苗组合物本发明的疫苗(组合物)可以是预防性的(即预防疾病)或治疗性的(即在患病后 治疗疾病)。这些疫苗包含免疫性抗原(即本发明的第二肿瘤抗原和第一肿瘤抗原),并且通常与“药学上可接受的载体”组合,这些载体包括本身不诱导产生对接受该组合物的个体有害的抗体的任何载体。合适的载体通常是大的、代谢缓慢的大分子,如蛋白质、多糖、聚乳酸、聚乙醇酸、氨基酸聚合物、氨基酸共聚物、脂质凝集物(如油滴或脂质体)等。这些载体是本领域普通技术人员所熟知的。另外,这些载体可起免疫刺激剂(“佐剂”)作用。另外,抗原也可以和细菌类毒素(如白喉、破伤风、霍乱、幽门螺杆菌等病原体的类毒素)偶联。增强免疫组合物效果的优选佐剂包括但不限于:(1)铝盐(alum),如氢氧化铝、磷酸铝、硫酸铝等;(2)水包油型乳剂配方,例如,(a)mf59(参见wo90/14837),(b)saf,和(c)ribitm佐剂系统(ras)(ribiimmunochem,hamilton,mt),(3)皂素佐剂;(4)freund完全佐剂(cfa)和freund不完全佐剂(ifa);(5)细胞因子,如白介素(如il-1、il-2、il-4、il-5、il-6、il-7、il-12等)、干扰素(如γ干扰素)、巨噬细胞集落刺激因子(m-cfs)、肿瘤坏死因子(tnf)等;(6)细菌adp-核糖基化毒素(如大肠杆菌热不稳定毒素lt)的脱毒变异体;以及(7)作为免疫刺激剂来增强组合物效果的其它物质。包括免疫原性组合物在内的疫苗组合物(例如,可包括抗原、药学上可接受的载体以及佐剂),通常含有稀释剂,如水,盐水,甘油,乙醇等。另外,辅助性物质,如润湿剂或乳化剂、ph缓冲物质等可存在于这类运载体中。更具体地,包括免疫原性组合物在内的疫苗,包含免疫学有效量的免疫原性多肽,以及上述其它所需的组分。“免疫学有效量”指以单剂或连续剂一部分给予个体的量对治疗或预防是有效的。该用量可根据所治疗个体的健康状况和生理状况、所治疗个体的类别(如人)、个体免疫系统合成抗体的能力、所需的保护程度、疫苗的配制、治疗医师对医疗状况的评估、及其它的相关因素而定。预计该用量将在相对较宽的范围内,可通过常规实验来确定。通常,可将疫苗组合物或免疫原性组合物制成可注射剂,例如液体溶液或悬液;还可制成在注射前适合配入溶液或悬液、液体赋形剂的固体形式。该制剂还可乳化或包封在脂质体中,以增强佐剂效果。此外,本发明的疫苗组合物可以是单价的或多价疫苗。(iii)给药途径和剂量一旦配成本发明的组合物,可将其直接给予对象。待治疗的对象可以是哺乳动物,尤其是人。当用作疫苗时,可用已知的方法将本发明的重组蛋白直接施用于个体。通常采用与常规疫苗相同的施用途径和/或模拟病原体感染路径施用这些疫苗。给予本发明药物组合物或疫苗组合物的途径包括(但并不限于):肌内、皮下、皮内、肺内、静脉内、经鼻、经口服或其它肠胃外给药途径。如果需要,可以组合给药途径,或根据疾病情况进行调节。疫苗组合物可以单剂量或多剂量给予,且可以包括给予加强剂量以引发和/或维持免疫力。应以“有效量”给予重组蛋白疫苗,即重组蛋白的量在所选用的给药路径中足以引发免疫应答,能有效促使保护宿主抵抗相关的疾病。代表性的疾病包括(但并不限于):肿瘤等,例如肝癌、肺癌、胃癌、乳腺癌、卵巢癌、前列腺癌、皮肤癌、黑色素瘤、宫颈癌、脑癌、甲状腺癌和胆管癌、膀胱癌和胰腺癌。在各疫苗剂份中所选用的重组蛋白的量,是按可引发免疫保护性应答而无明显的副作用的量而定。通常,在感染宿主细胞后,各剂的疫苗足以含有约1μg-1000mg,较佳地为1μg-100mg,更佳地10μg-50mg蛋白质或多肽(包括第二肿瘤抗原或第一肿瘤抗原)。可用包括观察对象中的抗体滴定度和其它反应的标准研究方法来确定具体疫苗的最佳用量。可通过监控疫苗提供的免疫力水平来确定是否需要增强剂量。在评估了血清中的抗体滴定度后,可能需要选用增强剂量免疫接种。施用佐剂和/或免疫刺激剂就可提高对本发明的蛋白质的免疫应答。优选方法是从肠胃外(皮下或肌内)途径通过注射给予免疫原性组合物。此外,本发明的疫苗可以结合其它免疫调节剂一起给予,或者与其他治疗剂一起给予。本发明的主要优点包括:(a)本发明的免疫策略和组合物可以有效突破机体对自身抗原的免疫耐受。(b)本发明的免疫策略和组合物可以有效激发机体针对肿瘤抗原的免疫反应。(c)本发明有助于针对性地解决当前治疗性肿瘤疫苗普遍低效的问题,对加快肿瘤疫苗的研发具有重要的推动作用。(d)本发明可应用于肿瘤免疫治疗、疫苗开发等方面。下面结合具体实施例,进一步阐述本发明。应理解,这些实施例仅用于说明本发明而不用于限制本发明的范围。下列实施例中未注明具体条件的实验方法,通常按照常规条件,例如sambrook等人,分子克隆:实验室手册(newyork:coldspringharborlaboratorypress,1989)中所述的条件,或按照制造厂商所建议的条件。除非另外说明,否则百分比和份数是重量百分比和重量份数。所有抗体均为市售抗体,购自ebioscience或bdbioscience公司。例如,anti-mousevβ2tcr(b20.6,ebioscience);anti-mousevβ3tcr(kj25,bd bioscience)。所有试剂除非另外说明,否则均为市售。实验动物:6到8周野生型c57bl/6小鼠购于中国科学院上海实验动物中心slac公司;rip-mova小鼠获自华盛顿大学霍华德休斯医学研究所。通用方法(a)李斯特菌的培养和感染1.-80℃冻存的李斯特菌划线涂tsb培养基固体平板,37℃培养箱过夜后挑取单克隆菌落,30℃,200rpm/min,摇床培养过夜14小时后,按照1:20比例转接菌液150μl至3ml新鲜tsb液体培养基中,37℃,200rpm/min,摇床培养2.5~3小时后到对数生长期,测od600值,此时od600值在0.6~1间。2.取1ml菌液,12000rpm/min离心10min,重复用1ml磷酸盐缓冲溶液清洗离心两遍,根据测得od600值用磷酸盐缓冲溶液重悬菌体沉淀,od600与菌液浓度间的对应关系为当od600值为1时,菌液浓度为109cfu/ml。3.按照1:10比例依次取100μl菌液稀释至1ml磷酸盐缓冲溶液,直至菌液浓度稀释为105cfu/ml,继续按照1:4比例稀释菌液浓度为2.5×104cfu/ml用于小鼠的尾静脉注射。(b)小鼠骨髓来源树突状细胞的培养、活化和免疫1.第1天:完整取出年龄6周至8周c57/bl6雌性小鼠下肢的股骨和胫骨,去除附着的肌肉组织,放置预冷磷酸盐缓冲溶液中,转移置细胞工作台。用无菌pbs冲洗骨头3次。无菌剪刀剪去骨头两端,1ml注射器冲洗骨髓腔,直至骨变白。重悬于磷酸盐缓冲溶液,尼龙膜过滤为单细胞悬液。300g,10min,4℃,离心去上清后,加入1ml红细胞裂解液,冰上裂解5分钟后加入10ml磷酸盐缓冲溶液终止。300g,10min,4℃离心去上清,细胞计数后重悬于kmedium铺24孔板,细胞浓度为1×106细胞/ml,1ml/孔。移入细胞培养箱,37℃,5%co2条件下培养。2.第3天:24孔板小角度倾斜10分钟后,移液枪吸取移走孔内上清500μl。补充新鲜培养基500μl,添加生长因子gm-csf和il-4,37℃,5%co2培养。3.第5天:吸取24孔板内所有培养基及细胞,4℃,300g,10min离心去培养基上清,补充新鲜培养基1ml/孔,添加生长因子gm-csf和il-4,移入细胞培养箱。4.第7天:吹打收集24孔板内所有的悬浮细胞,300g,10min离心后去上清,重悬于新鲜培养基,细胞浓度为2×106细胞/ml,加入cpg800ng/ml,移入细胞培养箱,37℃刺激活化16小时。5.cpg刺激过夜后的小鼠树突细胞300g,10min离心后去上清,重悬于新鲜培养基,细胞浓度为1×106细胞/ml,加入测试多肽2μg/ml负载,37℃,5%co2培 养2.5小时。6.负载多肽后的小鼠树突细胞300g,10min,4℃离心后去上清,10ml磷酸盐缓冲溶液清洗3次后,重悬于无血清磷酸盐缓冲溶液,细胞浓度为20×106细胞/ml,用于小鼠免疫。(c)黑色素瘤细胞的培养和黑色素瘤模型的建立黑色素瘤细胞系mo5细胞(获自中国科学院)从液氮冻存中复苏培养(第一代),第二代培养开始加入浓度为1mg/ml抗生素g418筛选特异性表达ova抗原的mo5细胞。筛选传代3-4次,第四代或第五代细胞处于指数生长期时收集,重悬于磷酸盐缓冲溶液或无血清dmem培养基中。小鼠用2.5%的avertin麻醉,后背去毛,在后腰正中皮下接种mo5细胞,5×105细胞/小鼠。从接种后第6天开始,每隔2天用游标卡尺测量肿瘤长径和短径的长度,肿瘤体积依照公式【长径×(短径)2×π/6】计算42。当长径和短径的平均长度超过2厘米后,出于依据动物实验伦理准则将小鼠断颈处死减轻痛苦。(d)小鼠肿瘤的免疫预防和免疫治疗小鼠肿瘤的预防:小鼠根据实验方案预免表达抗原李斯特菌和/或树突细胞疫苗后第7天,皮下接种黑色素瘤细胞系mo5细胞5×105细胞/小鼠,观察并记录肿瘤生长情况。小鼠肿瘤的治疗:小鼠皮下接种黑色素瘤细胞系mo5后,根据设计的治疗方案,给予李斯特菌和树突细胞疫苗治疗,观察并记录肿瘤生长情况。(e)骨髓来源的树突细胞培养1.第1天:完整取出年龄6周至8周c57/bl6雌性小鼠下肢的股骨和胫骨,去除附着的肌肉组织,放置预冷磷酸盐缓冲溶液中,转移置细胞工作台。用无菌pbs冲洗骨头3次。无菌剪刀剪去骨头两端,1ml注射器冲洗骨髓腔,直至骨变白。重悬于磷酸盐缓冲溶液,尼龙膜过滤为单细胞悬液。300g,10min,4℃,离心去上清后,加入1ml红细胞裂解液,冰上裂解5分钟后加入10ml磷酸盐缓冲溶液终止。300g,10min,4℃离心去上清,细胞计数后重悬于kmedium铺24孔板,添加生长因子gm-csf和il-450ng/ml,细胞浓度为1×106细胞/ml,1ml/孔。移入细胞培养箱,37℃,5%co2条件下培养。2.第3天:24孔板小角度倾斜10分钟后,移液枪吸取移走孔内上清500μl。补充新鲜培养基500μl,添加生长因子gm-csf和il-4,37℃,5%co2培养。3.第5天:吸取24孔板内所有培养基及细胞,4℃,300g,10min离心去培养基上清,补充新鲜培养基1ml/孔,添加生长因子gm-csf和il-4,移入细胞培养箱。4.第7天:吹打收集24孔板内所有的悬浮细胞,300g,10min离心后去上清,重悬于新鲜培养基,细胞浓度为2×106细胞/ml,加入cpg800ng/ml,移入细胞培养箱,37℃刺激活化16小时。5.cpg刺激过夜后的小鼠树突细胞300g,10min离心后去上清,重悬于新鲜培养基,细胞浓度为1×106细胞/ml,加入测试多肽(如trp2sd或trp2sk)2μg/ml负载,37℃,5%co2培养2.5小时。6.负载多肽后的小鼠树突细胞300g,10min,4℃离心后去上清,10ml磷酸盐缓冲溶液清洗3次后,重悬于无血清磷酸盐缓冲溶液,细胞浓度为20×106细胞/ml,用于小鼠免疫。(f)统计学分析除了特殊说明的地方,独立实验至少重复2次,每组样品总数量n≥5。原始数据经核对后用prism5.0(graphpad,美国)软件进行统计学分析。实验数据用平均值±标准误差来表示。当p值小于0.05(信赖区间为50%)被认为具有统计学差异。实施例1天然抗原和突变抗原对内源性多克隆t细胞功能性亲和力的鉴定本发明人用表达天然抗原的李斯特菌lm-n4尾静脉注射感染小鼠,7天后取脾脏细胞体外用呈梯度浓度的天然抗原n4或突变抗原a2,y3,q4,t4,v4等分别刺激培养,检测ifn-γ+cd8+t细胞的比例,做剂量反应关系曲线(图2a)。结果如图2所示。本发明人发现,相对于cd8+t细胞对天然抗原n4的剂量反应曲线,对于其他突变抗原的剂量反应曲线都发生了右移。本发明人用半最大效应浓度(ec50)的倒数(1/ec50)表示天然抗原和修饰抗原的相对功能性亲和力,发现尽管天然抗原和突变抗原对内源性多克隆t细胞体系的功能性亲和力相比对ot-it细胞降低,但是其亲和力高低顺序并没有改变。天然抗原n4对内源性多克隆ova特异性t细胞的功能性亲和力最高(ec50=27.58pm),a2为仅次于n4的高功能性亲和力抗原(35.4pm),y3和q4为中等亲和力抗原(253.51pm和689.17pm),t4和v4为低亲和力抗原(3221pm和20930pm)(图2b)。功能性亲和力的差异可能会影响特异性识别n4的内源性t细胞库对突变抗原的交叉反应性,在常规的抗原刺激浓度2μg/ml条件下,识别n4的内源性多克隆t细胞库对突变抗原的交叉反应水平总体上和突变抗原的功能性亲和力正相关。高亲和力的抗原与n4特异性t细胞库的交叉反应性高于低亲和力的抗原(图2c)。实施例2中枢耐受使小鼠免疫系统更倾向于对中等亲和力的突变抗原反应鉴定了天然抗原和突变抗原对ova抗原特异的内源性多克隆t细胞体系的功能性亲和力后,本发明人进一步研究小鼠内源性多克隆t细胞库对天然抗原和突变抗原的反应规律及中枢耐受机制对该规律的影响。用表达天然抗原n4或突变抗原的李斯特菌株分别感染wt小鼠和rip-mova小鼠,7天后取脾脏细胞体外用各自感染株对应的抗原分别刺激,检测ifn-γ+cd8+t细胞的比例。结果如图3所示。wt小鼠体内的t细胞库对天然抗原和突变抗原都能产生免疫反应;而在rip-mova小鼠中,因为中枢耐受机制的存在,对天然抗原n4和高亲和力抗原a2的免疫反应几乎被完全抑制,对其他中等亲和力和低亲和力突变抗原的免疫反应也有不同程度的降低,而中枢耐受对y3、q4、v4三个突变抗原诱导免疫反应水平的影响较低。该结果说明在中枢耐受机制的存在下,机体的t细胞库对自身抗原和高亲和力突变抗原不能反应,而对突变抗原的免疫反应水平被抑制,但是一些中等亲和力和低亲和力的突变抗原仍可以诱导激活产生一定数量的抗原特异性t细胞克隆。由于在抗肿瘤免疫反应中,起主要作用的是突变抗原诱导的免疫反应中识别天然抗原的那一群t细胞,因此在本实施例中,进一步检测突变抗原免疫后,t细胞库对天然抗原的交叉反应。用携带突变抗原的李斯特菌株分别免疫wt小鼠和rip-mova小鼠后,取脾脏细胞体外用天然抗原n4刺激培养,检测ifn-γ+cd8+t细胞的比例。在wt小鼠中,对天然抗原反应的t细胞的比例都低于对突变抗原本身反应的t细胞的比例,突变抗原诱导的对天然抗原的交叉反应和突变抗原的功能性亲和力总体上呈正相关关系,高亲和力的突变抗原能诱导更高比例的天然抗原反应性t细胞,低亲和力的突变抗原倾向于诱导较低比例的天然抗原反应性t细胞。而在rip-mova小鼠中,由于天然抗原为自身抗原,其他突变抗原的免疫仅产生极少能对天然抗原反应的t细胞,只有突变抗原y3可以诱导明显的、天然抗原交叉反应性t细胞(平均比例为0.9%),亦即自身反应性t细胞。然而有趣的是,高亲和力的突变抗原a2免疫后产生的天然抗原交叉反应性t细胞占所有抗原反应性t细胞的比例在wt小鼠中接近100%,说明几乎所有a2特异性的t细胞都能对天然抗原反应,这个比例在rip-mova小鼠中为55%,降低了接近一半。而y3免疫后的交叉反应率从wt小鼠中的29%降低至rip-mova小鼠中的19%,仅降低了约30%(图3)。这些实验结果说明,当天然抗原为外源性抗原时,机体t细胞库对突变抗原的免疫反应水平主要与其功能性亲和力相关;而当天然抗原为自身抗原时,中枢耐受机制清除了大部分自身反应性t细胞和对高功能性亲和力突变抗原反应的t细胞,使得t细胞库更倾向于对中等亲和力的突变抗原反应。而用lm-y3免疫在rip-mova小鼠中检测到了明显的对天然抗原反应的t细胞,说明在中枢耐受机制对自身反应 性t细胞的清除并不彻底,t细胞库内仍存在足够数量的克隆可以识别应答自身抗原。而在中枢耐受机制下,识别自身抗原的t细胞反应水平与交叉反应率的关系也并不明显,说明决定抗肿瘤反应有效性的因素仍然在于体内存在的低亲和力克隆的绝对数量,高的交叉反应率并不表明能激活更多的自身反应性t细胞。实施例3在wt小鼠和rip-mova小鼠上构建黑色素瘤模型鉴定出在中枢耐受机制作用下,中等亲和力的突变抗原y3可以最好的诱导自身反应性t细胞,因此具有潜在的抗肿瘤作用。为了后续的实验,在本实施例中,用不同剂量的表达ova的黑色素瘤细胞mo5皮下接种wt小鼠,建立肿瘤抗原为外源性抗原的黑色素瘤非耐受模型(图4a),确定了在5×105细胞/只的接种量时,可以确保每只小鼠形成生长均一的黑色素肿瘤。用5×105细胞/只剂量接种rip-mova小鼠,模拟人肿瘤发生过程中肿瘤抗原为自身抗原的情况,黑色素瘤mo5可以在小鼠上正常生长,因此建立对了ova抗原自身耐受的黑色素瘤模型(图4b),其存活曲线略高于wt小鼠(图4c)。进一步观察了rip-mova小鼠接种肿瘤后是否会产生肿瘤抗原特异性的t细胞(图4d和e),用n4特异性的tetramer检测发现rip-mova小鼠脾脏和引流淋巴结内都没有识别自身抗原n4的体细胞(图4d),体外用n4刺激也几乎不产生ifn-γ+cd8+t细胞。有趣的是,用y3在体外刺激脾脏细胞和引流淋巴结细胞,产生了一定数量的表达ifn-γ和tnf-α的cd8+t细胞比例(图4e)。这提示,体内的t细胞库更容易对突变抗原y3反应。实施例4中等亲和力突变抗原诱导的自身反应性t细胞不能有效抑制肿瘤生长中枢耐受机制存在下,中等亲和力突变抗原y3能诱导t细胞库中产生最多的自身抗原反应性t细胞,而在ova抗原中枢耐受小鼠荷瘤模型的检测中也发现,小鼠体内存在对突变抗原y3反应的t细胞。然而,进一步的研究表明lm-y3的感染免疫对对表达天然ova抗原的肿瘤并没有明显的抑制作用,抗肿瘤反应效率低下。在本实施例中,用lm-n4和lm-y3分别预免wt小鼠和rip-mova小鼠,然后接种表达ova抗原的黑色素瘤细胞系mo5。结果如图5所示。尽管在rip-mova小鼠上lm-y3可以比lm-n4更好抑制黑色素瘤生长(图5b),但仍然明显低于在wt小鼠上两种抗原的抑制效果(图5a)。这暗示,在抗肿瘤过程中,起主要作用的仍然是针对天然抗原n4的自身反应性t细胞,而不是只对突变抗原y3反应的那群细胞。接着,本发明人用天然抗原lm-n4和突变抗原lm-y3分别感染wt小鼠和 rip-mova小鼠,绘制wt小鼠的n4-特异性t细胞(wtsp.t细胞)和n4-交叉反应性t细胞(wtcr.t细胞),以及rip-mova小鼠的n4-交叉反应性t细胞(rocr.t细胞。rosp.t细胞因存在比例过低无法计算)剂量反应曲线(图5c),并计算相对功能性亲和力。结果表明,与wtsp.t细胞相比,wtcr.t细胞的相对功能性亲和力下降了约109倍,而rocr.t细胞的亲和力下降了490倍(图5d)。说明在中枢耐受不存在的情况下,突变抗原诱导的识别天然抗原的t细胞库只具有较低的亲和力;而中枢耐受机制存在时,突变抗原激活的识别天然抗原的t细胞库为自身反应性t细胞,其亲和力范围进一步降低。以上结果说明,中枢耐受环境下突变抗原激活的低亲和力自身反应性t细胞不能对肿瘤抗原进行有效的免疫反应,抑制其生长。实施例5识别同一抗原的自身反应性t细胞库和高亲和力t细胞库偏向使用不同的vβ在内源性多克隆免疫体系中,抗原与t细胞间的功能性亲和力,实际为t细胞库中识别应答同一抗原的t细胞克隆的平均功能性亲和力。因为单个t细胞的受体vβ基因通过片段重排,可以产生不同的vβ链进而组合为不同的tcr形成巨大的由不同克隆组成的t细胞库。在本实施例中,进一步研究突变抗原激活的识别自身抗原的t细胞库和识别同一抗原的高亲和力t细胞库间的差异。用突变抗原y3分别免疫wt小鼠和rip-mova小鼠,用n4-tetramer抗体和y3-tetramer抗体共染色检测其对y3和n4分别识别的t细胞库。结果如图6所示。wt小鼠中交叉识别天然抗原n4的t细胞库与识别y3的t细胞库有部分重叠,n4-tet+y3-tet+cd8+t细胞比例为3.20%,仍有1.05%的cd8+t细胞仅识别n4和15.1%的cd8+t细胞仅识别y3;而在rip-mova小鼠中,识别n4的t细胞库中84%都来自于识别突变抗原y3的t细胞库(图6a)。通过进一步对wtsp.t细胞、wtcr.t细胞和rocr.t细胞中vβ表达情况的检测发现,尽管wtcr.t细胞的功能性亲和力显著低于wtsp.t细胞,两者的vβ使用倾向性和t细胞库多样性却基本一致,相反地,尽管亲和力只相差大约4倍,rocr.t细胞的t细胞库多样性和另两者相比却明显不同(图6b),而rip-mova小鼠中识别y3的t细胞库却基本上不受中枢耐受影响(图6c)。这说明,中枢耐受后,剩下的t细胞库主要针对突变抗原,而激活的自身反应性t细胞大部分来自突变抗原特异性的t细胞库。还发现自身反应性t细胞库的多样性相比另两者降低,更倾向于使用某些特定的vβ。通过对vβ使用率的比较,结果发现,相较于wt小鼠,rocr.t细胞中 对vβ3,vβ7,vβ10b和vβ13的使用率显著上升,未检测的vβ(others)比例显著下降(图6d和e)。该结果暗示,由于中枢耐受选择性的清除高亲和力的自身反应性t细胞克隆,存留下来的识别自身抗原的t细胞更依赖于使用突变抗原t细胞库中某些低亲和力的特定vβ。实施例6中枢耐受下同种突变抗原的重复免疫治疗不能有效抑制肿瘤生长由于中枢耐受作用对自身抗原反应性t细胞库的抑制作用,突变抗原免疫后产生的自身反应性t细胞只具有极低的亲和力。因为突变抗原诱导的自身抗原t细胞反应是有效的抗肿瘤免疫的基础,本发明人尝试用重复免疫突变抗原作为治疗性肿瘤疫苗,看是否能够增强抗肿瘤免疫反应,抑制肿瘤生长。本发明人在rip-ova小鼠上接种表达ova的黑色素瘤细胞mo5,建立天然肿瘤抗原n4的中枢耐受模型;并在wt小鼠上接种同样的肿瘤,作为天然肿瘤抗原的非中枢耐受模型。接种肿瘤后第7天开始,本发明人用表达天然抗原的lm-n4或突变抗原的李斯特菌株治疗各组小鼠,治疗一共进行三次,每次时间间隔一周。结果如图7所示。在非中枢耐受的wt小鼠荷瘤治疗模型中本发明人发现,天然抗原n4和高亲和力突变抗原a2的重复治疗可以有效抑制肿瘤生长,其他中等亲和力和低亲和力的突变抗原则没有治疗效果(图7a)。通过对lm-n4和lm-a2治疗后的荷瘤wt小鼠进行检测,本发明人发现小鼠的脾脏、肠系膜淋巴结、腹股沟引流淋巴结,都产生了一定数量的对天然抗原n4反应的t细胞(图7c)。而在基于rip-ova小鼠构建的中枢耐受模型中,包括能在rip-ova小鼠中最好诱导自身反应性t细胞的突变抗原y3在内,所有的抗原治疗都不能产生有效的抗肿瘤作用,抑制肿瘤生长(图7b)。考虑到也许在rip-ova小鼠中存在着对ova抗原的外周耐受机制,其中体内未成熟的dc递呈抗原或许会诱导对自身反应性t细胞的抑制作用,为此,本发明人构建了新的免疫策略:利用cpg刺激成熟的骨髓来源树突细胞负载y3(dc-y3),作为疫苗治疗荷瘤rip-ova小鼠。本发明人首先用不同浓度的cpg刺激骨髓来源培养的dc细胞,检测其活化分子cd80和cd86的表达。结果发现,在2μg/ml的浓度刺激下,dc细胞可以最好的被活化。但是,无论是dc-y3的重复免疫治疗(图7d),还是dc-y3结合lm-y3的双重免疫治疗策略,都不能有效抑制肿瘤(图7e)。这些实验结果表明,基于突变抗原y3,传统的同种抗原重复免疫治疗不足以产生足够的抗肿瘤免疫反应,而需要设计新的免疫策略以提高中心耐受条件下的自 身抗原反应性t细胞反应,才可能对肿瘤治疗有效。实施例7双重异源免疫能有效提高中枢耐受下的免疫反应在本实施例中,本发明人进一步利用成熟的树突细胞负载不同抗原,构建树突细胞疫苗,和表达不同抗原的李斯特菌组合对rip-ova小鼠进行免疫,检测其激活的识别自身抗原的t细胞免疫反应。本发明人首先检测在免疫抗原不改变的情况下,仅改变两次免疫中递呈抗原的载体(李斯特菌或成熟的树突细胞),对中枢耐受环境下的自身抗原特异性t细胞反应是否有影响。本发明人用负载突变抗原y3的树突状细胞疫苗(dc-y3)或lm-y3分别免疫rip-ova小鼠,7天后,再根据实验设计用dc-y3或lm-y3再次免疫各组小鼠,4天后在体外用天然抗原n4刺激,检测cd8+t细胞中细胞因子ifn-γ和tnf-α的表达情况。结果如图8所示。在不改变免疫抗原的情况下,两次免疫后各组小鼠产生的ifn-γ+cd8+t细胞比例相比仅用lm-y3免疫一次后的比例在数量上并没有增加,反而降低了大约50%,而tnf-α+cd8+t细胞和ifn-γ+tnf-α+cd8+t的比例没有明显改变(图8a)。但是,通过将免疫后小鼠的脾脏细胞体外用梯度浓度的n4刺激绘制剂量效应曲线(图8c)并计算比较相对功能性亲和力(图8d)后本发明人发现,和仅用突变抗原y3免疫一次后产生的自身反应性t细胞相比,两次lm-y3免疫和dc-y3-lm-y3综合免疫后产生的自身反应性t细胞的相对功能性亲和力均显著增加,分别为2.6倍和15倍。有趣的是,后者的相对性功能性亲和力,甚至超过了wtcr.t细胞的相对性亲和力。结果表明,在免疫抗原不变情况下,自身反应性t细胞亲和力的提高,依赖于疫苗载体的影响,而dc-lm这样的组合能有效提高自身反应性t细胞的亲和力。进一步地,本发明人在两次使用不同载体组合免疫的基础上,改变免疫抗原,检测t细胞的功能性细胞因子的表达和功能性亲和力是否受到影响。根据实验设计和图8b所示,本发明人用两种载体dc和lm,与天然抗原n4和突变抗原y3的不同组合,间隔7天两次免疫rip-ova小鼠,第二次免疫4天后检测识别天然抗原n4的自身反应性cd8+t细胞的细胞因子表达和功能性亲和力(图8b,c和d)。本发明人发现,只有dc-y3-lm-n4这样的免疫组合能够产生明显的ifn-γ+cd8+t细胞,尽管其比例0.68%,仍然略低于lm-y3免疫一次后产生的ifn-γ+cd8+t细胞的比例0.93%;但是,其产生的tnf-α+cd8+t细胞比例明显高于其他的免疫组合,且超过了lm-y3免疫一次后的比例大约3倍;而其产生的ifn-γ+tnf-α+cd8+t细胞的比例是lm-y3免疫一次后的大约4倍,超过了其中水平最低的免疫组合lm-n4-dc-y3产生的ifn-γ+tnf-α+cd8+t细胞比例13倍(图8b);此外,其自身 反应性t细胞的相对功能性亲和力尽管相比lm-y3一次免疫后的亲和力显著增加了10.5倍,但是和dc-y3-lm-y3组合免疫后自身反应性t细胞的功能性亲和力相比,没有明显的差异(图8c和d)。这些结果说明,在确定了载体的基础上,自身反应性t细胞的功能性受到免疫抗原的影响,先突变抗原再天然抗原的组合可以促进自身反应性t细胞表达多种细胞因子。利用载体与抗原相结合的双重异源免疫策略,先使用突变抗原再使用天然抗原免疫,dc-y3-lm-n4这种组合能够同时增强自身反应性t细胞的多功能性和功能性亲和力。本发明人进一步检测这群活化的自身反应性t细胞的vβ使用情况,以确定t细胞库的多样性是否有改变。通过对ifn-γ+cd8+t细胞的vβ用特定抗体染色后发现,和lm-y3免疫一次后的rocr.t细胞的各vβ比例相比(图4b),双重异源免疫后在rip-ova小鼠中尽管其他的vβ使用比例仅略微增加,t细胞库多样性缩小(图8f),自身反应性t细胞表达vβ3的比例却相较于只用前者显著下降了大约80%,其比例近似于高亲和力的wtsp.t细胞中vβ3的比例,此外,表达vβ4的自身反应性t细胞的比例也显著增加了大约3.4倍(图8f和g)。该结果说明,第二次用天然抗原免疫,对突变抗原免疫激活的自身反应性t细胞库进行了筛选。综上,本发明人发现使用特定的双重异源免疫策略,可以在中枢耐受抑制下增强自身反应性t细胞的免疫反应,先使用突变抗原再使用天然抗原能促进多功能性t细胞的产生,而改变免疫抗原递呈载体有效能够提高t细胞的功能性亲和力。通过对自身反应性t细胞库做vβ使用分析发现,突变抗原免疫后产生的低亲和力自身反应性t细胞可能倾向于使用某些特定的vβ,而优化免疫策略后产生的自身反应性t细胞库中,对使用这些特定vβ的t细胞克隆的不选择,可能是自身反应性t细胞功能性和亲和力增加的原因。实施例8优化的双重异源免疫能在中枢耐受机制下有效激活肿瘤特异性免疫反应,抑制肿瘤生长基于突变抗原y3,本发明人确定了在中枢耐受机制作用下下,优化的双重异源免疫组合dc-y3-lm-n4能最有效激活rip-ova小鼠体内的自身抗原反应性t细胞,进而可能产生潜在的抗肿瘤免疫反应。本发明人进一步检测当肿瘤抗原为自身抗原时,双重异源免疫策略诱导的自身反应性t细胞是否具有抗肿瘤作用,并是否能有效抑制肿瘤生长。用dc-y3或lm-y3分别免疫rip-ova小鼠,7天后用lm-y3或lm-n4再次免疫。第二次免疫7天后接种mo5黑色素瘤细胞,并观察肿瘤生长。结果如图9所示。双重异源组合dc-y3-lm-n4免疫后的rip-ova小鼠上,肿瘤 的生长速度明显慢于同种组合lm-y3-lm-y3和仅改变载体的组合dc-y3-lm-y3免疫后rip-ova小鼠上的肿瘤生长速度(图9a)。通过比较平均肿瘤大小,本发明人发现dc-y3-lm-n4的组合免疫诱导的免疫反应可以有效抑制黑色素瘤的生长(图9b),在同时间段内,相比没有免疫过的小鼠,其存活率增加了50%(图9c)。以上结果说明,双重异源免疫策略激活的自身反应性t细胞能够特异性识别并杀伤肿瘤细胞,具有显著的抗肿瘤作用。实施例9优化的双重异源免疫能作为治疗性肿瘤疫苗在荷瘤小鼠上抑制肿瘤生长本发明人已经验证了双重异源免疫组合dc-y3-lm-n4激活的自身反应性t细胞可以杀死肿瘤细胞,在rip-ova小鼠上抑制肿瘤生长。因此,在本实施例中,进一步检测在构建的中枢耐受荷瘤小鼠模型上,双重异源免疫治疗策略能否作为治疗性疫苗,抑制肿瘤并延长小鼠的存活期。用表达ova的黑色素瘤细胞mo5皮下接种rip-ova小鼠,第7天进行第一次治疗,第14天进行第二次治疗(图10a)。结果表明,相比不治疗的对照组,和其他双重异源免疫治疗组,dc-y3-lm-n4明显能更有效抑制肿瘤生长(图10b)。和改变载体的同种抗原免疫治疗组dc-y3-lm-y3相比(图5d),双重异源免疫组合也能更有效的抑制肿瘤生长(图10c),并提高小鼠的存活率(图10d)。不治疗组的小鼠平均存活期为26.25天,dc-y3-lm-y3治疗组的平均存活期为26.4天,而dc-y3-lm-n4治疗的平均存活期可达到32天,相比另两组延长小鼠存活期近1周。实施例10基于佐剂的双重异源免疫提高抗肿瘤特异性t细胞免疫反应并抑制肿瘤生长(a)小鼠免疫1.第1天,取50μl(1×106细胞)负载trp2sd或trp2sk的树突细胞,脚掌注射免疫年龄6周至8周c57/bl6雌性小鼠,左右脚掌各25μl。2.第5天,使用trp2sd或trp2sk(50μg/只)腹腔注射免疫小鼠,以lps(5μg/只)和cpg(10μg/只)作为佐剂。3.第12天,重复步骤2。4.第19天,处死小鼠,取小鼠脾脏细胞,流式染色分析。其中,trp2sd:svydffvwl(native)(seqidno.:7);trp2sk:svykffvwl(mutated)(seqidno.:8)(b)流式染色1.取小鼠脾脏置于预冷磷酸盐缓冲溶液中研磨为单细胞悬液,尼龙膜过滤后4℃,320g,10min离心后去上清,加入1ml红细胞裂解液,冰上裂解5分钟后加入10ml磷酸盐缓冲溶液终止,4℃,300g,10min离心去上清,重悬于1ml1640培养基中。2.脾脏细胞悬液铺入96孔板,4×106细胞/well。添加多肽刺激(trp2sd),多肽浓度为2μg/ml。37℃,5%co2培养30分钟后加入bfa,浓度为2.5μg/ml。继续刺激培养5小时。3.24孔板内细胞转移至流式离心管,加入预冷的1ml流式染色液清洗,4℃,320g,5min离心去上清。4.细胞置于冰上,每管加入1μl的fcblockeranti-cd16/cd32及50μl流式染色液,轻柔震荡混匀,冰上孵育10min。5.每管加入流式抗体pe-cy7-cd8,percp-cy5.5-tcrβ,pe-cd44及50μl流式染色液,细胞震荡混匀,避光,冰上孵育30min。6.每管加入1ml流式染色液清洗两次,4℃,320g,10min离心去上清。7.每管加入250μl试剂盒中的细胞固定液,细胞震荡混匀,避光,冰上孵育50min。8.每管加入1ml试剂盒中的细胞破膜液清洗,震荡混匀,4℃,320g,10min离心去上清。重复清洗2遍。9.细胞置于冰上,每管加入1μl的fcblockeranti-cd16/cd32及50μl细胞破膜液,震荡混匀,避光冰上孵育10min。10.每管加入流式抗体fitc-ifnγ,apc-tnfα及50μl细胞破膜液,细胞震荡混匀,避光,冰上孵育30min。11.每管加入1ml细胞破膜液清洗,震荡混匀,4℃,320g,10min离心去上清。最后重悬于300μl的流式染色液,经流式细胞仪上样分析。实验结果:初次免疫使用负载突变肽trp2sk的树突细胞,再次免疫使用trp2sd及佐剂的小鼠记为sk-sd小鼠,初次免疫使用负载突变肽trp2sk的树突细胞免疫,再次免疫使用trp2sk及佐剂的小鼠记为sk-sk小鼠,初次免疫使用负载突变肽trp2sd的树突细胞免疫,再次免疫使用trp2sd及佐剂的小鼠记为sk-sd小鼠。sd-sd小鼠的cd8t细胞免疫反应较低,sk-sk小鼠的免疫反应也较低,与sd-sd小鼠无显著差异,sk-sd小鼠的免疫反应显著高于sd-sd小鼠和sk-sk小鼠(图11)。以上结果再次证明,只使用天然肽(trp2sd)或只使用突变肽(trp2sk)免疫均不能高效诱导cd8t细胞的免疫反应,先使用突变肽扩增t细胞库,然后使用天然肽诱导交叉反应的免疫策略是行之有效的。讨论肿瘤抗原作为自身抗原一直存在着免疫原性差的问题,影响了治疗性肿瘤疫苗在临床上的应用和发展。对肿瘤抗原上的t细胞表位进行氨基酸修饰可以提高其免疫原性,增强抗肿瘤免疫反应水平,但是在临床实验中的治疗效果并不明显。过去的研究表明,中枢耐受环境下修饰抗原引起免疫反应对天然肿瘤抗原的交叉反应性低是目前基于修饰抗原的治疗性肿瘤疫苗效果差的原因之一,而对造成这一问题分子机制目前仍不十分清楚。本发明人利用表达卵清蛋白模式抗原天然t细胞表位或修饰表位的李斯特菌株感染野生型小鼠及卵清蛋白模式抗原转基因rip-mova小鼠发现,中枢耐受机制作用下免疫系统对天然抗原不反应,只有中等亲和力的修饰抗原在免疫后能诱导机体免疫系统产生交叉识别天然抗原的低亲和力t细胞,并且t细胞库多样性显著降低。通过双重异源免疫策略,可以在中枢耐受环境下激活识别天然肿瘤抗原的t细胞,有效提高其亲和力及多功能性,抑制肿瘤生长,延长小鼠存活期。本发明为治疗性肿瘤疫苗的抗原优化和筛选,以及免疫策略的设计提供了理论基础和指导意义。根据上述实验结果,总结出治疗性肿瘤疫苗的构建宜基于以下几点原则:①合适的突变抗原的选择,使其作为初次免疫抗原能够激活t细胞库中数量足够的自身抗原反应性t细胞;②初次免疫载体和再次免疫载体的组合与改变,以提高自身反应性t细胞亲和力为主要目的和衡量标准;③再次免疫中天然抗原对t细胞库筛选的效率,使筛选后的自身反应性t细胞更好的表达功能性细胞因子。在本发明提及的所有文献都在本申请中引用作为参考,就如同每一篇文献被单独引用作为参考那样。此外应理解,在阅读了本发明的上述讲授内容之后,本领域技术人员可以对本发明作各种改动或修改,这些等价形式同样落于本申请所附权利要求书所限定的范围。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1